Els factors ambientals que actuen per sota el nivell de la mar són essencialment diferents als que regeixen les comunitats emergides. En un món constantment submergit, la disponibilitat hídrica deixa de ser un problema: els organismes, que no estan preparats per a sobreviure als períodes d’emersió, moren quan queden accidentalment en sec a causa d’unes fortes minves.
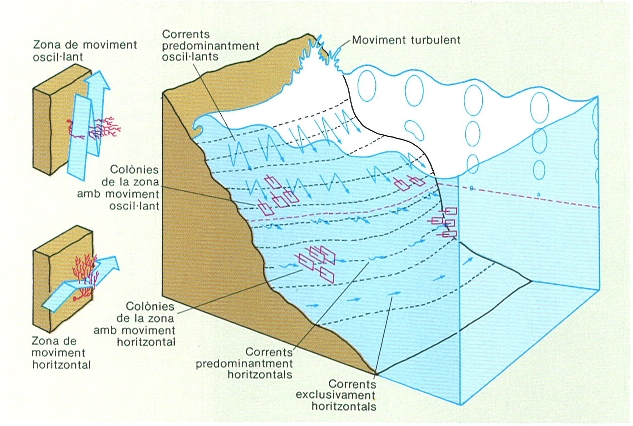
L’hidrodinamisme varia qualitativament i quantitativament segons la fondària. És turbulent i fort a la zona litoral, té un moviment de vaivé en els primers metres (zona d’oscil·lació) i minva i es torna unidireccional i horitzontal a més fondària. La disposició de les gorgònies, orientades transversalment o longitudinalment al pla vertical segons la fondària, és un bon indicador d’aquest canvi.
Jordi Corbera, original de R. Riedl, modificat
L’hidrodinamisme és un factor important, no com a fornidor d’humectació, sinó com a element renovador de l’aigua que envolta plantes i animals bentònics, ja que posa a disposició de les plantes els compostos químics i dels animals les partícules alimentàries que necessiten per a viure. També té, però, un efecte desestabilitzador o disruptor a causa del seu poder destructor quan és massa intens. Cal també tenir en compte que l’hidrodinamisme té un component direccional diferent segons la fondària que considerem. A la zona estrictament litoral, l’hidrodinamisme és totalment turbulent, i no hi ha cap direcció predominant de circulació. Una mica més avall, la turbulència generada pel trencament de les onades és substituïda per un moviment de vaivé. Queda així delimitada una zona on domina el moviment bidireccional de l’aigua segons un eix vertical (zona d’oscil·lació). Aquest moviment oscil·lant decreix a mesura que augmenta la fondària, i els moviments unidireccionals de l’aigua segons un pla horitzontal passen a ser els dominants. Aquesta zona, amb un hidrodinamisme regit pels corrents dominants, és anomenada zona de flux. La intensitat d’aquest flux és vital per a la supervivència de molts organismes suspensívors, l’orientació dels quals reflecteix la direcció dominant del flux.
La llum és un altre factor important, sens dubte el principal, ja que d’ell depèn la producció fotosintètica, per a comprendre la distribució dels organismes segons la fondària. L’atenuació de la llum és molt gran ja en el primer metre a causa dels processos de reflexió que pateix en travessar la inferíase aire/aigua; a partir d’aquí, segueix una corba exponencial amb un coeficient d’extinció inversament proporcional a la transparència de l’aigua. La intensitat de llum que arriba a una fondària determinada depèn, doncs, de la matèria en suspensió, inorgànica i orgànica, incloent dins d’aquesta darrera el fitoplàncton; per tant, varia segons la situació geogràfica i les característiques hidrogràfiques i productives de la zona. En termes generals, la transmitància és menor a les costes continentals que a les insulars, i és molt afectada per les aportacions de les desembocadures dels rius, les clavegueres i els emissaris.
La composició espectral de la llum també varia segons la fondària. L’absorció és màxima en la zona de les longituds d’ona del vermell, i mínima en les del blau. Les partícules, i d’una manera especial el fitoplàncton, varien la composició espectral: la transmitància pel blau i pel violeta és més baixa en les aigües riques en fitoplàncton. Això és important, perquè en aigües relativament productives la màxima transmitància es produeix a la zona del verd, precisament la zona de l’espectre en què l’absorció dels fotons per part de la clorofil·la és mínima. En aigües molt transparents, com les que hi ha a les Balears, la transmitància a la zona del blau és elevada i la clorofil·la absorbeix la llum que arriba al fons amb més eficiència. La proliferació i conservació, al llarg de l’evolució, de pigments accessoris en les algues (ficobilines, clorofil·la c, fucoxantines, sifonoxantines, etc.) s’ha interpretat sovint com un mecanisme d’adaptació a un medi on l’absorbància de les diferents longituds d’ona és selectiva. Això no obstant, i segons els estudis més recents, sembla que la fotosíntesi depèn més de la quantitat de llum que no pas de la qualitat, la qual cosa invalida (almenys parcialment) la teoria de l’adaptació cromàtica, segons la qual les algues verdes estarien més adaptades als nivells superficials, les roges als profunds (on rebrien, respectivament, llum de les longituds d’ona complementàries) i les brunes a profunditats intermèdies.
La zona ben il·luminada és ocupada per extenses poblacions d’algues anomenades fotòfiles, això és, amants de la llum. En aquesta zona, els herbívors són els principals organismes animals, i els suspensívors, en clar desavantatge respecte de les algues, es reclouen principalment als racons més ombrívols. Les poblacions d’algues fotòfiles típiques desapareixen aproximadament allà on arriba només el 10-15% de la irradiància superficial, i a partir d’aquesta fondària dominen les algues esciòfiles, és a dir, amants de l’ombra. El límit inferior de les comunitats dominades per algues esciòfiles és variable i depèn també de l’hidrodinamisme i la concentració de matèria orgànica particulada, factors que afavoreixen els organismes suspensívors. En termes generals, però, aquest límit pot situar-se vers un 10 MJ/m2·any, que correspon aproximadament al 0,5% de la irradiància superficial. Les algues, però, com a components minoritaris de les comunitats, són capaces de créixer a irradiàncies molt inferiors. En particular, hom ha detectat el creixement d’algues bentòniques tropicals a irradiàncies al voltant de 0,007 MJ/m2·any (és a dir, de 2 a 3 milionèsimes de la irradiància superficial!). A les nostres costes, hom observa coral·linàcies incrustants fins a irradiàncies inferiors al 0,05% de la irradiància superficial, sense arribar, però, a les cotes observades a les mars tropicals. Per a valors inferiors al 0,5% de la irradiància superficial els animals acostumen a predominar sobre les algues i, malgrat estar encara dins la zona fital, la dominància animal és manifesta. La principal entrada d’energia al sistema és proporcionada pels organismes suspensívors, els quals prenen el relleu de les algues en el paper de principals estructuradors de les comunitats.
El tipus de substrat és un factor bàsic a l’hora d’avaluar l’existència d’unes determinades comunitats. El substrat rocós és el més apropiat per a mantenir una densa cobertura d’organismes. Així i tot, sobre substrat sorrenc o de graves poden establir-se fanerògames marines que estabilitzen el substrat i afavoreixen el desenvolupament d’algues i animals, bé sobre les seves fulles, o bé sobre els seus rizomes. Pel que fa a la naturalesa química de la roca, l’efecte sobre les comunitats és pràcticament nul, llevat de la fauna i flora endolítiques, molt més abundants en roques carbonatades, més fàcilment perforables. La taxa de sedimentació i el tipus de sediment són també importants; taxes de sedimentació molt elevades són desfavorables a la implantació de les algues i de qualsevol organisme sèssil en general. La temperatura és també un factor important, ja que probablement és la principal responsable de gran part de les diferències de distribució geogràfica dels organismes. La diferència de 2 a 3°C en la temperatura mitjana de l’aigua superficial a la Costa Brava respecte a la de les illes Balears o del País Valencià són suficients perquè hom observi una tropicalització, no solament en les flores i faunes respectives, sinó també en l’estructura i la dinàmica de les comunitats.
La disponibilitat de nutrients i matèria orgànica particulada són factors determinants de les comunitats que s’estableixen en una zona. La Mediterrània és una mar caracteritzada per les baixes concentracions de nutrients i partícules en suspensió, i qualsevol increment en els valors naturals comporta una simplificació de les comunitats i una banalització de les espècies que hi viuen. A les zones pol·luïdes químicament (ports, litoral d’àrees industrials), la reducció de la riquesa i la diversitat del bentos és encara més manifesta, fins al punt que en situacions extremes només sobreviuen organismes inferiors (bacteris, cianòfits i fongs).
La freqüència dels que podem anomenar factors catastròfics és molt important en les comunitats submergides. L’esdeveniment d’algunes d’aquestes catàstrofes té un origen climàtic i les diferents espècies i comunitats han acoblat el seu cicle preveient aquestes situacions. Les minves i els grans temporals poden ser catalogats com a factors catastròfics de periodicitat anual en moltes comunitats de dominància algal. Això no obstant, hi ha catàstrofes (generalment d’origen humà) que arriben a destruir totalment les comunitats. D’aquests casos, en tenim exemples clars en l’abocament indiscriminat de fangs procedents de depuradores i que estan carregats de metalls pesants i altres contaminants; l’extracció de sorres destinades a «regenerar» platges; o la pesca a ròssec.
Llum, hidrodinamisme, temperatura, tipus de substrat, taxes de sedimentació, concentració de nutrients, pol·lució química, esdeveniments catastròfics: tots són factors importants en el món submergit. Cal tenir en compte, però, que els factors no actuen sols, sinó que els organismes estan sotmesos al seu conjunt de manera que és la resultant de la seva interacció la que té una influència decisiva per a cada organisme. D’altra banda, cada organisme és afectat de forma diferent i això condiciona la seva capacitat de resposta davant de l’ambient que l’envolta. Amb això volem dir que no són els factors en ells mateixos que interactuen amb cada organisme, sinó que són els factors com a modificadors de les potencialitats de cada organisme per separat els que determinen la composició específica, la dominància, l’estructura i, parcialment, la dinàmica de cada comunitat. La competència per l’espai, la depredació i, en definitiva, els factors biòtics són cabdals, i ho són més com més constants són les condicions ambientals.
Els fenòmens que intervenen en la composició de les comunitats són, com hem pogut veure, força més complexos sota l’aigua que als estatges supralitoral i mediolitoral i, a més, tenen un mecanisme d’actuació que podríem qualificar de més subtil. I aquesta subtilesa augmenta amb la fondària: els gradients físics s’esmorteeixen i els factors biòtics tenen cada cop més rellevància. El resultat és que la zonació en el món submergit és més desdibuixada que en l’emergit. Les comunitats són distingibles per elles mateixes però, llevat dels primers metres, les discontinuïtats són difícils d’observar en indrets on les característiques del substrat (consistència, inclinació, orientació) es mantenen constants. El substrat és generalment molt irregular i per això, l’estatge infralitoral apareix com un món extraordinàriament heterogeni; però hi ha una direcció de màxima variació ambiental, difuminada en un ample mosaic de taques corresponents a comunitats o fàcies diferents. Potser un símil ajudarà a fer més entenedora aquesta idea.
Podem imaginar el bentos marí com un món en miniatura on totes les coses són molt més petites i van més ràpides que a les comunitats terrestres. Els arbres de 15 o 20 m d’alçada són substituïts per algues que rarament sobrepassen els 15 o 20 cm. El temps que triga a reconstituir-se, després d’una maltempsada, un «bosc» d’aquestes característiques és, com a màxim, de 2 a 3 anys, en lloc dels 40 anys o més que triga un bosc mediterrani. La mida relativa dels herbívors i dels animals dels altres nivells de la cadena tròfica és, en canvi, molt gran. Pensem en un conill i un bosc i comparem-lo amb una garota i un prat d’algues. Observem també l’extraordinària densitat de garotes i altres herbívors que hi ha a la majoria de comunitats de dominància algal. La contínua explotació de les algues per part d’aquestes poblacions d’herbívors crea tota una sèrie d’estadis de regeneració de la comunitat clímax amb un efecte i una estructura semblants als que provoca l’home sobre la vegetació en una zona d’elevada densitat de població. Si unim això a l’heterogeneïtat creada pels mateixos gradients físics que es troben a l’eix vertical (tant o més forta a 30 m de desnivell sota l’aigua com a 2000 m en un ambient terrestre) i a l’eix horitzontal, comprendrem el perquè de la immensa diversitat dels fons marins. La nostra enorme mida relativa i la nostra situació quan nedem per sobre del fons ens fan veure les comunitats bentòniques mediterrànies de la mateixa manera que un gegant de 100 m d’alçada veuria qualsevol paisatge humanitzat en una zona molt muntanyosa.
Les comunitats de dominància algal
Les comunitats de dominància algal són pròpies de la zona infralitoral però també es desenvolupen a la part superior de la zona circalitoral. Per l’estructuració de la vegetació en estrats, les comunitats algals bentòniques recorden comunitats terrestres en miniatura. Hi trobem des de comunitats molt senzilles, equivalents als prats de conreu gairebé monoespecífics, fins a comunitats amb diversos estrats de vegetació, equivalents, en certa manera, a les selves tropicals. Ja que són la llum i la disponibilitat de nutrients els factors que acostumen a limitar el creixement de les algues, no és estrany que en el curs de la successió es tendeixi a comunitats amb una màxima estratificació, dominades per espècies erectes, les quals competeixen favorablement en la recepció de la llum i en l’explotació del medi aquàtic com a. font de nutrients. Hi ha diversos factors que impedeixen la consecució d’aquesta màxima estructuració. El més important és, sens dubte, l’efecte dels herbívors, els quals, com ja hem comentat, exploten les comunitats algals i les mantenen en estats variables de regeneració. Els factors físics, com l’hidrodinamisme i la disponibilitat de nutrients, poden també (freqüentment a causa de la interacció amb els factors biòtics) impedir o potenciar la constitució de comunitats ben estructurades.
Les comunitats de dominància algal: composició i estructura
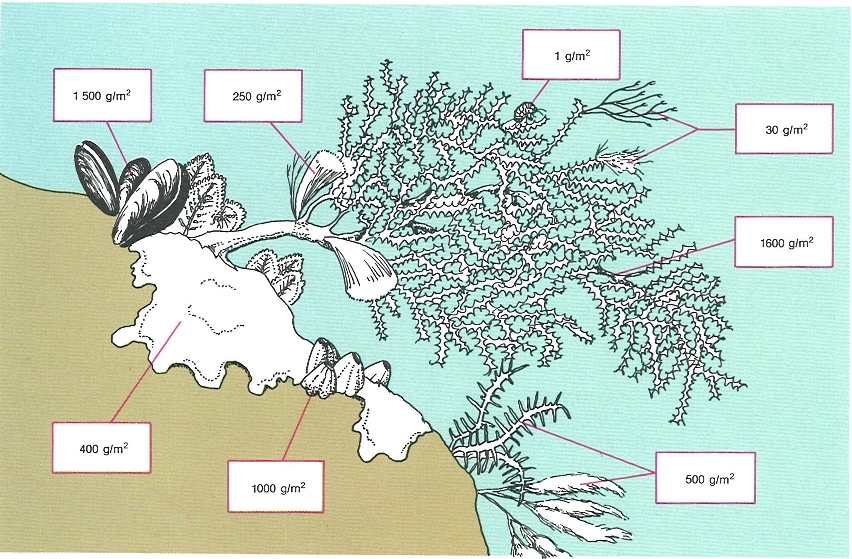
Estructura d’una comunitat de Cystoseira mediterranea de la Costa Brava i distribució de la biomassa (en g de pes sec/m2). Els estrats «arbustiu» i «herbaci» són poc importants en comparació d’altres comunitats d’algues fotòfiles a causa del gran recobriment de l’espècie dominant i de l’intens hidrodinamisme a què és sotmesa. La turbulènaa afavoreix, però, la presència d’organismes filtradors (Mytilus galloprovincialis i Balanus perforatus).
Jordi Corbera, amb dades d’E. Ballesteros.
En el cas de màxima complexitat, una comunitat d’algues mediterrànies pot arribar a tenir cinc estrats de vegetació diferenciats. L’estrat erecte, més elevat (15-40 cm), acostuma a estar constituït per poblaments d’una o diverses espècies, sovint del gènere Cystoseira. Per sota seu apareixen una sèrie d’espècies fotòfiles de mida considerable, de 5 a 15 cm (Halopteris, Padina, Dilophus), que equivaldrien a un estrat arbustiu elevat. En un nivell inferior, hi ha nombroses espècies que poden formar gespes o petits manyocs que recobreixen els espais que deixen les altres algues: es tracta d’espècies de requeriments ecològics molt variats, on trobem tant formes fotòfiles (Acetabularia, Dasycladus, Anadyomene), com esciòfiles (Udotea, Rhodymenia, Peyssonnelia) o polivalents (Coral·lina, Gelidium). Totes aquestes espècies rara vegada s’implanten sobre la roca, sinó que creixen sobre algues incrustants, sovint calcificades, que recobreixen totalment, o gairebé, el substrat (estrat incrustant: Lithophyllum, Neogoniolithon, Polystrata), substrat que també pot ser una bioconstrucció (per exemple, conquilles anastomitzades de Dendropoma petraeum).
Finalment, hi ha un estrat epifític que creix, principalment, sobre les algues dels estrats més elevats. Les algues epífites són molt nombroses i constitueixen l’estrat més ric en espècies; quantitativament poden arribar a tenir tanta biomassa com les algues de l’estrat erecte. Cal tenir en compte també algunes espècies animals epífites, bé que són poc nombroses i poc importants quant a biomassa.
La simplificació de les comunitats es realitza per la pèrdua progressiva dels diferents estrats en l’ordre següent: estrat elevat, estrat arbustiu alt, estrat arbustiu inferior. Les zones més afectades per la depredació o per qualsevol altre factor desestabilitzador arriben a estar constituïdes només per un estrat d’algues incrustants sobre el qual es desenvolupen algues microscòpiques i petites algues filamentoses o laminars de morfologia molt senzilla i de creixement ràpid. En general, les comunitats ben estructurades es caracteritzen per espècies pròpies de medis estables, que no viuen als llocs on hi ha un estrès físic o biòtic important; en aquests darrers indrets dominen les algues oportunistes, les quals, malgrat ser presents a les comunitats ben estructurades, mai no hi tenen un desenvolupament important.
Hom pot ordenar les diferents espècies d’algues al llarg d’un eix hipotètic els extrems del qual fossin l’estratègia de la r (algues oportunistes) i l’estratègia de la K. Una ordenació de les algues segons aquestes estratègies ecològiques es relaciona íntimament amb una classificació segons una sèrie de grups funcionals, que se situarien al llarg de l’eix esmentat anteriorment. L’augment de la mida i el gruix del tal·lus són factors evolutivament lligats, els quals afavoreixen la persistència en el temps, perquè en dificulten el consum per part dels animals herbívors. La calcificació dels tal·lus és un pas més en aquest sentit, però només ha estat aconseguit per alguns grups vegetals (coral·linàcies, peissonneliàcies, algunes caulerpals i dictiotals). Finalment, la síntesi de productes que actuen com a repel·lents enfront dels herbívors, encara que no és una característica funcional, és un mecanisme bàsic per a augmentar la perdurabilitat de les algues. En general, hom observa un augment de les formes carbonatades i de les algues més perdurables en augmentar l’estabilitat i la complexitat del medi, tant des d’un punt de vista geogràfic (tropicalització) com general (regularitat en els paràmetres físics i químics). En aquest sentit, hom observa un augment de la importància de les algues incrustants i carbonatades a les comunitats de les illes Balears i el País Valencià respecte de la Costa Brava o la Catalunya del Nord, com també un augment del nombre d’espècies calcificades en augmentar la fondària. El quocient P/B (producció/biomassa), indicador de la taxa de renovació, disminueix en el mateix sentit.
Els organismes animals són presents en les comunitats de dominància algal, però rara vegada són prou rellevants perquè hi prenguin part com a definidors d’una comunitat determinada. En són excepcions les poblacions denses de garotes, cirrípedes i mol·luscs: hom ha descrit dues associacions en les quals els vogamarins Paracentrotus lividus i Arbacia lixula tenen una significació important, tant en termes de biomassa global com per llur acció rosegadora, que impedeix l’evolució d’aquestes comunitats cap a poblaments més estructurats; els musclos (Mytilus galloprovincialis) i les glans de mar (Balanus perforatus), com també el gasteròpode vermiforme Dendropoma petraeum (= Vermetus [Spiroglyphus] cristatus), poden constituir poblacions importants en indrets amb un elevat hidrodinamisme. Llevat d’aquests casos, els animals són components minoritaris en les comunitats d’algues fotòfiles. Així i tot, el bentos fotòfil no és comparable a les comunitats terrestres, ja que molts animals treuen el seu aliment de les partícules en suspensió o dels detrits. Això fa que la biomassa animal pugui ser, en termes comparatius, molt més alta en una comunitat bentònica que en una comunitat terrestre. Quan la fondària augmenta, els organismes animals desplacen progressivament les algues com a elements principals en les comunitats.
La fauna que habita les comunitats dominades per algues no pot qualificar-se de fotòfila. La llum és un factor d’importància secundària per als animals. La presència d’animals en una comunitat d’algues pot tenir diverses raons. La principal és que siguin organismes fitòfags i, en aquest cas, es tracta d’espècies vàgils que recorren el substrat mentre s’alimenten de les algues. El creixement dels organismes suspensívors solament és possible si resulten prou competitius. L’epifitisme, amb totes les característiques pròpies d’espècies estrategues de la r que porta associades, és una solució. Una altra possibilitat és deixar-se recobrir per les algues, la qual cosa pot tenir una funció críptica enfront els depredadors habituals; ho fan filtradors que bomben l’aigua amb una certa potència (Microcosmus, Ostraea). Finalment, també és possible entrar en competència directa amb les algues i impedir o dificultar el seu creixement mitjançant procediments bioquímics o d’altres que n’impedeixin la fixació o la germinació.
En un intent d’agrupar els diferents animals que viuen en les comunitats de dominància algal, hom pot establir les categories que esmentem a continuació. En primer lloc, els organismes sèssils incrustants que recobreixen el substrat d’una manera semblant a la de les algues incrustants; en són exemples diverses esponges (Crambe crambe, Hymeniacidon sanguinea), poliquets serpúlids, mol·luscs (Dendropoma petraeum) i alguns briozous i tunicats. En segon lloc, els organismes sèssils no incrustants que competeixen fortament amb les algues en l’ocupació de l’espai; aquest grup inclou la majoria de suspensívors que viuen a les comunitats d’algues fotòfiles: esponges (diverses Ircinia, Spongia virgultosa, Adocia simulans), cnidaris (Anemonia sulcata, Aiptasia diaphana, Balanophyllia regia, molts hidraris), mol·luscs (Ostraea edulis, Arca noae, Spondylus gaederopus), tunicats (Microcosmus sabatieri, Aplidium pseudolobatum, Polycarpa pomaria), poliquets tubícoles (Spirographis spallanzani) i briozous (Myriapora truncata, Crisia occidentalis). Les espècies sedentàries, fortament fixades al substrat, de les quals són exemples Mytilus galloprovincialis i Balanus perforatus, formen una tercera categoria. Una altra la constitueixen les espècies epibiòtiques d’algues i animals dels grups anteriors, grup que inclou moltes espècies d’animals suspensívors (hidraris, esponges, briozous, poliquets i tunicats, entre d’altres). En cinquè lloc, les espècies endobiòtiques (sobretot d’esponges i tunicats) i endolítiques (especialment les esponges del gènere Cliona i el dàtil de mar Lithophaga lithophaga). En sisè lloc, els organismes criptòfils, que viuen en els intersticis i les microcavitats naturals o produïdes per les algues i animals estructuradors de l’espai; la llista d’espècies és molt llarga i inclou poliquets (Nereis zonata, Lepidonotus clava), mol·luscs (Jujubinus gravinae, Haliotis tuberculata), ofiures (Ophiothrix fragilis, Amphipholis squamata), estrelles de mar (Asterina gibbosa) i sipuncúlids (Phascolosoma granulatum). En setè lloc, les espècies vàgils pertanyents a la microfauna, que es mouen entre les algues i que sovint se n’alimenten, o bé dels organismes colonials epífits; aquest grup inclou un gran nombre de crustacis (amfípodes i isòpodes, principalment), mol·luscs (prosobranquis i opistobranquis), picnogònides i poliquets errants. I finalment, les espècies vàgils de la macrofauna, com alguns crancs (Pilumnus hirtellus, Alpheus dentipes), molts gasteròpodes, pops (Octopus vulgaris), garotes (Paracentrotus lividus, Arbacia lixula), estrelles de mar (Echinaster sepositus) i molts peixos, sobretot gòbids i blènnids.
Ja hem comentat l’extraordinària heterogeneïtat del món submergit, principalment en els primers metres. És lògic, doncs, que l’estudiós dels fons marins es capfiqui a esbrinar quines són les diferències entre les taques que observa. Això ha dut a definir una sèrie d’associacions, sobretot vegetals, representatives de diferents combinacions en els factors ambientals i biòtics, com també de les diferents zones geogràfiques. La llista de comunitats i associacions que hom pot trobar als fons dels Països Catalans és massa llarga per a comentar-la aquí, però podem fer-ne una síntesi i remarcar-ne els aspectes funcionals.
Característiques de les estratègies r i K en tres grups d’organismes bentònics
| ESTRATEGS DE LA r | ESTRATEGS DE LA K |
|---|---|
| ALGUES | |
| Grups representatius: ulvàcies, ectocarpàcies, ceramiàcies | Grups representatius: udoteàcies, cistoseiràcies, peissonneliàcies |
| Bons colonitzadors | Mals colonitzadors |
| Durada de la vida efímera o anual; perennes, amb grans alts i baixos al llarg de l’any | Durada de la vida llarga, amb cicles vitals complexos |
| Tal·lus senzill; quocient àrea/volum elevat | Tal·lus ben diferenciat estructuralment; quocient àrea/volum baix |
| Creixement ràpid i discontinu; teixit totalment fotosintetitzador | Creixement estacional o continu; amb teixit no fotosintetitzador |
| Capacitat reproductora molt elevada i activa durant tot l’any | Capacitat reproductora confinada a un teixit reproductor especialitzat; reproducció estacional |
| Contingut calòric elevat i uniforme | Contingut calòric distribuït diferentment segons les parts del tal·lus; capacitat d’emmagatzemar substàncies de reserva |
| Tal·lus juvenils amb la mateixa estratègia que els adults | Tal·lus juvenils amb característiques i estratègies pròpies de les formes oportunistes |
| Mecanismes de defensa: poden fugir dels herbívors gràcies a la seva impredicibilitat temporal i espacial o al seu creixement ràpid | Mecanismes de defensa: redueixen l’apetència dels depredadors amb mecanismes químics i estructurals de defensa |
| CNIDARIS | |
| Grup representatiu: hidrozous | Grup representatiu: antozous |
| Dimensions dels individus petites (de mm) | Dimensions dels individus mitjanes (de cm) |
| Dimensions de les colònies molt variables, segons el substrat, però predominantment petites | Dimensions de les colònies generalment grans |
| Colònies estolonials | Colònies reptants, erectes o estolonials |
| Esquelet present o no, sempre corni | Esquelet rarament absent, calcari o corni |
| Alimentació suspensívora, micròfaga | Alimentació suspensívora, micròfaga o macròfaga |
| Reproducció asexual per estolons i gemmació | Reproducció asexual per gemmació i escisió |
| Reproducció sexual amb formació de larves i meduses; maduració ràpida, proporcional a les dimensions de la colònia | Reproducció sexual amb formació de larves, que poden incubar-se; maduració lenta i irregular |
| Iteroparitat, amb producció de pocs ous en cadascun dels múltiples períodes reproductors | Semelparitat, amb producció de molts ous en l’únic (generalment) període reproductor |
| Més d’una generació anual, generalment | Menys d’una generació anual, generalment |
| Durada de la vida curta (formes fugaces), de pocs dies a un any | Durada de la vida llarga (formes perennes), fins a 50 anys |
| Taxa de renovació alta | Taxa de renovació baixa |
| Formes de creixement lineal, per estolons | Formes de creixement tridimensional, per gemmació |
| Tipus de creixement indeterminat, exponencial al llarg de tota la colònia (estoló) | Tipus de creixement més ràpid en colònies joves, més lent en les adultes i amb esquelet calcari |
| Epibiosi molt comuna | Epibiosi només en algunes espècies |
| Mecanismes de competència: nematocists, toxines, recobriment del contrari, formes de resistència (estolons) | Mecanismes de competència: nematocists, toxines, recobriment i depredació del contrari |
| Ambients sobretot inestables | Ambients especialment estables |
| MOL·LUSCS OPISTOBRANQUIS | |
| Grups representatius: tectibranquis (i eolidacis) | Grups representatius: nudibranquis (i doridacis) |
| Dimensions variables, de petits a grans (de cm) | Dimensions variables, de petits a grans (de cm) |
| Hàbits actius | Hàbits sedentaris |
| Alimentació micròfaga, eurífaga, aliment transitori | Alimentació micròfaga o macròfaga, estenòfaga, aliment perdurable |
| Ambient inestable | Ambient estable |
| Substrat tou | Substrat dur |
| Diversitat poblacional baixa | Diversitat poblacional alta |
| Mecanismes de defensa senzills o nuls | Mecanismes de defensa complexos |
| Reproducció per producció contínua d’ous; diverses generacions a l’any i desenvolupament planctotròfic | Reproducció per producció discontínua d’ous; una generació a l’any i desenvolupament planctotròfic, lecitotròfic i directe |
| Creixement asincrònic, molt ràpid, subanual | Creixement sincrònic, lent, anual |
| Estadis inicials de la successió | Estadis finals de la successió |
| Distribució geogràfica àmplia | Distribució geogràfica restringida |
| Dades dels autors | |
Les comunitats més superficials
En la zona més propera a la superfície, en llocs ben il·luminats i batuts per les ones, d’una certa estabilitat i d’aigües netes, es desenvolupa arreu de les costes dels Països Catalans la comunitat de pèl salper (Cystoseira mediterranea). Aquesta comunitat és molt sensible a qualsevol tipus de pertorbació, i la seva presència o absència és indicadora de la qualitat de les aigües i de l’estabilitat del medi. La seva desaparició comporta la substitució per una comunitat en forma de tapís ras, dominada per Corallina elongata. En ambdues associacions, a part un elevat nombre d’algues, hom hi troba abundants musclos (Mytilus galloprovincialis) i glans de mar (Balanus perforatus). A les illes Balears i al País Valencià hom observa en determinats indrets una formació que recorda estructuralment les tenasses de Lithophyllum tortuosum, però que són construïdes per les closques calcàries del gasteròpode vermiforme Dendropoma petraeum (= Vermetus [Spiroglyphus] cristatus). Aquestes voravies són totalment infralitorals, menys potents i molt més dures que la tenassa típica, i poden ser recobertes per algues fotòfiles.
L’hidrodinamisme minva en llocs arrecerats o en augmentar la fondària. Cystoseira mediterranea perd vitalitat i al seu lloc apareixen altres espècies dominants. A les zones arrecerades trobem sovint poblacions d’altres espècies de Cystoseira (C. compressa, C. caespitosa, C. crinita, C. balearica), però més freqüentment hom troba comunitats algals dominades per diferents espècies de feòfits: Padina pavonica, Halopteris scoparia, Dictyota dichotoma, Cladostephus hirsutus, Dilophus fasciola, etc. Si la pressió dels animals fitòfags (especialment garotes) és molt intensa, la cobertura vegetal desapareix i les algues coral·linàcies incrustants (Lithophyllum incrustans, Neogoniolithon notarisii) són les dominants, juntament amb les nombroses garotes. Si l’hidrodinamisme oscil·lant és intens apareix una comunitat dominada per Corallina elongata i altres petites algues. Totes aquestes comunitats són considerades d’algues fotòfiles i només apareixen en aquells indrets on la llum és abundant (igual o superior al 10% de la irradiància superficial).
En les baumes i zones més fosques (esquerdes, coves) l’aspecte canvia totalment com a conseqüència de l’aparició d’unes espècies d’algues diferents i del considerable augment dels organismes animals. Els feòfits desapareixen i els rodòfits i alguns cloròfits són les algues dominants. Hom distingeix a les zones batudes dues associacions; una és pròpia de les aigües fredes del golf del Lleó, i l’altra és present a la resta dels Països Catalans. A les zones encalmades, l’aspecte de les comunitats recorda molt el que trobarem a més fondària, ja que les espècies vegetals i animals són pràcticament les mateixes.
A les zones pertorbades, les comunitats fins ara esmentades hi són absents i la diversitat decreix molt: tot és més homogeni. L’enciam de mar (Ulva) pot assolir creixements puntuals extraordinaris, sobretot als llocs encalmats. Als indrets més batuts domina Corallina elongata. Altres algues que podem trobar en aquests ambients són els fideus (Codium fragile), Taonia atomaria, Dictyota dichotoma i un elevat nombre de petites algues filamentoses. Els organismes suspensívors poden ser localment abundants: musclos (Mytilus), ostres (Ostraea edulis), glans de mar (Balanus), esponges (diverses Ironia, Verongia aerophoba) i ascidis (Diplosoma spongiforme, Clavellina lepadiformis), com també alguns depredadors sèssils: les anèmones de mar Anemonia sulcata i Aiptasia diaphana.
Les comunitats profundes de substrat dur
En endinsar-nos per sota de la fondària on arriba el 10% de la llum superficial (8-10 m a la Costa Brava; 20 m a les Balears) les comunitats canvien gradualment d’aspecte. Al principi, les algues són encara les espècies dominants i l’estructura de les comunitats és idèntica a la descrita per a les algues fotòfiles. Els animals suspensívors són, però, més abundants: esponges (Ircinia variabitis, I. fasciculata, Spongia virgultosa), cnidaris (Eudendrium racemosum i altres hidraris, Eunicella singularis, Alcyonium acaule, les manetes de mort), briozous (diverses Crisia i Scrupocellaria, Myriapora truncata), tunicats (el rovell de mar Microcosmus sabatieri, l’ascidi roig Halocynthia papillosa, Aplidium pseudolobatum, Polysyncraton lacazei). Les algues dominants són feòfits (Cystoseira spinosa, Dictyopteris membranacea, Dictyota dichotoma) o cloròfits (Codium vermilara). A mesura que la fondària augmenta, l’estrat d’algues arborescents perd densitat. Les algues incrustants són ben visibles i tenen un desenvolupament força important (Mesophyllum lichenoides, Lithophyllum expansum, Peyssonnelia rosamarina, Neogoniolithon mamillosum). Si les condicions de llum són prou atenuades, el seu creixement en potència crea l’anomenat coral·ligen, comunitat de dominància animal on les algues es localitzen exclusivament en la part superior. Comentarem àmpliament aquesta comunitat en un capítol a part, però aquí farem referència a la seva cobertura algal, ja que és independent que el substrat sigui o no el resultat de la concreció produïda per les algues calcàries.
En una primera situació, no gaire esciòfila, la cobertura d’algues erectes pot ser molt important. Hom hi troba els cloròfits Udotea petiolata i Halimeda tuna i un nombre variat de rodòfits (Rhodymenia ardissonei, Vidalia volubilis, diverses Kallymenia i Peyssonnelia, etc.). Els feòfits són abundants en llocs amb corrents intensos on Cystoseira zosteroides, juntament amb Arthrocladia villosa i Sporochnus pedunculatus, constitueix una comunitat molt característica. Els fons de Laminaria rodriguezii apareixen també en llocs amb corrents intensos, però sempre per sota els 60 m, principalment a les Balears. En les zones més insolades dels Països Catalans, és a dir, les costes alacantines i balears, es dona una certa «pujada», encara no ben explicada, d’espècies d’algues que més amunt hem qualificat d’esciòfiles. Udotea petiolata i, sobretot, Halimeda tuna, poden trobar-se gairebé a flor d’aigua, situació insòlita a les costes septentrionals de la Mediterrània occidental. Potser cal cercar-ne la raó en l’escassa competència que hi tenen, tant amb altres algues com amb animals bentònics, ja que sembla que aquest factor és el que pot haver arraconat aquestes espècies en fons més pregons a la resta del litoral català. Totes les comunitats comentades fins ara creixen sobre substrat dur, on la majoria d’algues, proveïdes només de discs fixadors, poden instal·lar-se i desenvolupar-se.
Les comunitats profundes de substrat tou
Els fons tous, és a dir, de sorra o fang, estan sovint desproveïts d’una cobertura algal densa. En constitueixen una excepció importantíssima els herbeis de fanerògames marines, als quals ens referirem en un capítol a part. Hi ha, però, dues comunitats de dominància algal característiques dels fons tous.
La comunitat de coletes (Caulerpa prolifera) és pròpia de les zones sorrenques, ben il·luminades i d’aigües relativament càlides i encalmades (i de vegades un xic dessalades o fins i tot hiperhalines de les llacunes litorals); als Països Catalans és abundant a totes les costes insulars, al País Valencià i al delta de l’Ebre. La coleta és una alga estolonífera que colonitza les zones sorrenques gràcies a una sèrie de rizoides, petites rels que la mantenen fixada. Els fons de coletes són exclusius de la zona infralitoral i mai no sobrepassen els 20 m de fondària. Sovint, la mateixa presència d’aquesta alga contribueix a fer que els fons, sorrencs en principi, esdevinguin fangosos, de vegades amb un fang anòxic. Hi contribueix tant la captura de sediment per les algues com, sobretot, l’acumulació entre els rizoides, un cop l’any, de la matèria orgànica de les seves frondes que pràcticament cap herbívor consumeix (hi ha substàncies, com la caulerpina, que impedeixen aquest consum per part d’animals, a excepció d’alguna espècie d’opistobranqui).
L’altra comunitat de dominància algal que es desenvolupa sobre fons tous és pròpia dels indrets sorrencs i de graves de la zona circalitoral i és al costat del coral·ligen. És constituïda, bàsicament, per algues carbonatades de vida lliure, de forma arbuscular (Lithothamnium corallioides, L. valens, Phymatolithon calcareum, Lithophyllum racemus) o laminar (Peyssonnelia rosa-marina, P. crispata, Lithophyllum expansum), sobre les quals es desenvolupen una flora i una fauna molt característiques. A la Costa Brava els pescadors anomenen aquesta comunitat grapissar o glapissar, i a les Balears rep el nom d’avellanó, que recorda el terme francès de «praliné» (ametlla garapinyada) donat als mateixos fons (també anomenats fons de «maërl»). Apareix entre els 30 i els 90 m, i els nòduls formats pel creixement de les algues calcàries són sovint el substrat adient per a moltes altres algues (Vidalia volubilis, Laminaria rodriguezii i altres) i animals (gorgònies, esponges, tunicats, etc.) que necessiten un substrat dur mínim sobre el qual instal·lar-se.
Les comunitats de dominància algal: dinàmica i funcionalitat
Tal com hem vist fins ara, hom observa molts canvis en la constitució específica de les comunitats a mesura que augmenta la fondària. Però a més de les espècies, també canvia l’estructura de la comunitat. En termes generals, si ens fixem en les comunitats que podem considerar climàciques de cada nivell, veiem uns models de variació. La biomassa algal, per exemple, assoleix un màxim a les comunitats de Cystoseira superficials i decreix cap avall. La disminució de la biomassa és, però, molt lenta, ja que les algues carbonatades adquireixen una importància més gran amb la fondària. Això significa que les comunitats, a mesura que la fondària augmenta, sacrifiquen un creixement ràpid per aconseguir la formació d’estructures perdurables (carbonatades), que són menys atacables pels depredadors. El recobriment i la matèria orgànica tenen un comportament molt semblant al de la biomassa, però la seva disminució amb la fondària és molt més manifesta; així i tot, aquesta disminució és molt inferior a la de la llum. Es pot afirmar, per tant, que les comunitats d’algues de fondària són molt més eficients que les superficials, ja que amb una mateixa quantitat de llum són capaces de mantenir una quantitat de matèria viva (o d’estructura) força superior.
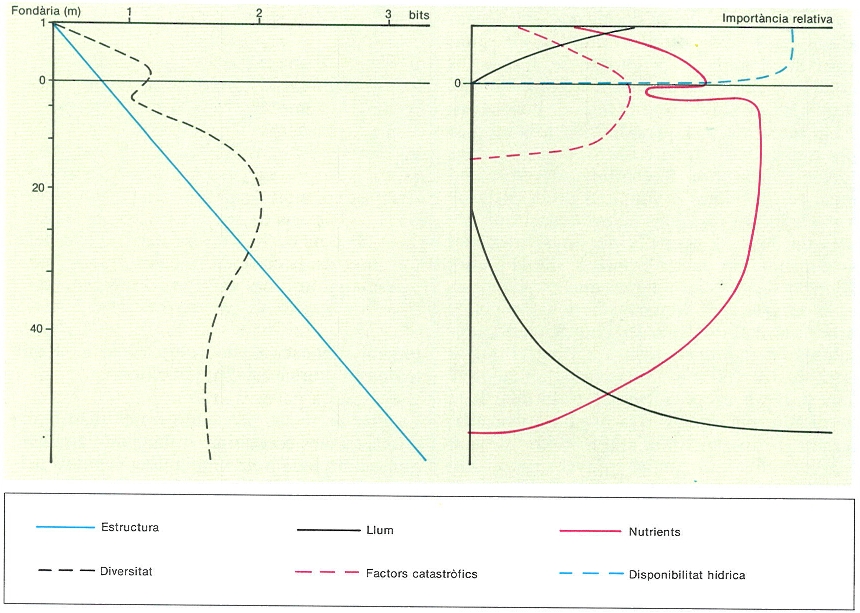
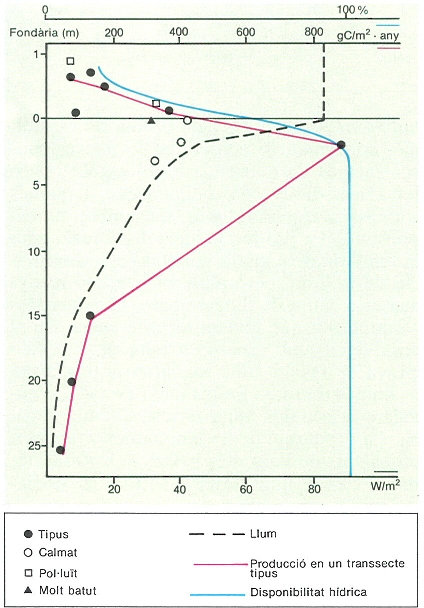
Variació de la diversitat específica i l’estructura (diversitat beta) de les comunitats fitobentòniques al llarg d’un transsecte tipus a Tossa de Mar (Selva). L’augment de l’estructuració indica graus més elevats de compartimentació i una repartició més laxa de les espècies en augmentar la fondària. La màxima diversitat s’obté, però, a fondàries intermèdies (10-20 m), on les condicions són òptimes per al desenvolupament d’un gran nombre d’espècies algals. Disponibilitat hídrica, nutrients i llum són, batimètricament, per aquest ordre, els factors principals que regeixen l’estructura i la dinàmica de les comunitats fitobentòniques.
Eduard Clavem, original d’E. Ballesteros.
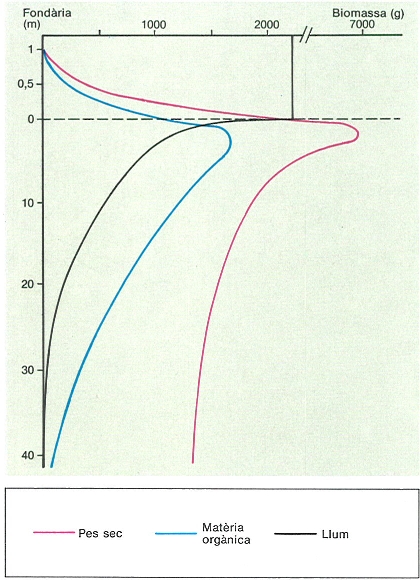
Variació de la biomassa algal al llarg d’un transsecte tipus a Tossa de Mar (Selva). A la zona mediolitoral (representada a escala augmentada en el dibuix) la biomassa augmenta d’una manera proporcional a la disponibilitat hídrica. En la zona submergida també augmenta al principi a causa de l’elevat hidrodinamisme i la intensitat de llum propis dels primers metres. La disminució posterior no segueix, però, l’extinció de la llum. La raó és principalment l’adquisició de les algues d’estructures carbonatades, però també una eficiència fotosintètica més gran, com ho demostra el comportament de les corbes de pes sec i de matèria orgànica.
Eduard Clavero, amb dades d’E. Ballesteros.
L’estructura pròpiament dita, és a dir, la distribució de les espècies dins la comunitat, també varia d’una manera regular amb la fondària. Això és lògic si pensem que la llum disminueix i la competència per l’espai augmenta. En aquest sentit, les comunitats d’algues dels fons coral·lígens són molt més estructurades, ja que les espècies que les formen es distribueixen molt laxament en l’espai. Ara bé, les condicions de vida per a les algues en aquests indrets on hi ha poca llum no són del tot favorables: la competència per l’espai amb els animals filtradors és gran i les algues estan en una posició no gens avantatjosa; a més, com que el creixement és lent, l’acció dels herbívors es fa sentir molt més. El resultat és que hom observa un empobriment del nombre d’algues a partir d’una certa fondària.
Biomassa (quadrats i rectangles) i producció (cercles) dels diferents compartiments algals en un coral·ligen d’Halimeda tuna (en verd) i Mesophyllum lichenoides (en blanc); s’han indicat en rosat els epífits i en blau les restants algues erectes.
Eduard Clavero, original d’E. Ballesteros.
En general, les comunitats amb més espècies d’algues són les situades a la part inferior de la zona infralitoral, lloc on les que són fotòfiles encara tenen prou llum per a viure i moltes de les esciòfiles es troben immerses en la intensitat de llum adequada per al seu desenvolupament. La riquesa d’aquestes comunitats és extraordinària: hom troba sovint més d’un centenar d’espècies en un espai de dos o tres pams quadrats. Les comunitats d’algues fotòfiles són també molt riques en espècies, principalment a la Costa Brava, on la gran proliferació de l’estrat erecte crea unes condicions de llum amortida en el subestrat, que afavoreix el desenvolupament de les algues esciòfiles. Els factors que provoquen estrès, com l’hidrodinamisme, la manca de llum o la pol, lució, disminueixen notablement aquesta riquesa. Així, per exemple, les comunitats de Cystoseira mediterranea, d’Schottera nicaeensis o de Corallina elongata, pròpies de llocs batuts, són molt més pobres que les comunitats de Cystoseira caespitosa o de Padina pavonica, típiques d’indrets encalmats. La diversitat, mesura que ens dona una idea de la riquesa específica juntament amb l’equitabilitat de les espècies, té un comportament semblant a la riquesa específica. En el coral·ligen, però, la diversitat algal és relativament elevada; encara que hi ha menys espècies, les que hi creixen estan representades d’una manera més equitativa.
La disponibilitat hídrica era el factor limitant de la producció a la zona mediolitoral. Quan la humectació és suficient, com passa a les comunitats submergides, hi ha dos factors que limiten la producció: la llum i la disponibilitat de nutrients. A poca fondària la irradiància és elevada i els nutrients són llavors els limitants. L’hidrodinamisme té aquí un paper molt important ja que, en renovar l’aigua que envolta les plantes, potencia la captació de nutrients: amb una mateixa concentració de nutrients, la disponibilitat és més gran en els llocs amb un hidrodinamisme intens. A les comunitats més fondes la llum disminueix i limita el creixement algal; la disponibilitat de nutrients és llavors un factor secundari. Les algues mediterrànies, sobretot les estrategues de la K, estan molt adaptades al seu medi i tenen diversos mecanismes per a superar, almenys en part, aquests problemes. Un d’aquests mecanismes és la gran eficiència en la captura dels compostos inorgànics de nitrogen i fòsfor. Per altra banda, moltes algues són capaces d’acumular substàncies de reserva o compostos rics en nitrogen i fòsfor, que utilitzen quan els convé. Finalment, les algues de fondària aprofiten al màxim la radiació que els arriba, ja sigui gràcies a la seva morfologia, com a una elevada eficiència en el mecanisme captador de fotons. Les ficobilines (ficoeritrina i ficocianina) possibiliten que les algues vermelles capturin la llum de tot l’espectre visible amb una eficàcia màxima, la qual cosa els permet créixer fins a irradiàncies molt més baixes que qualsevol altra planta.
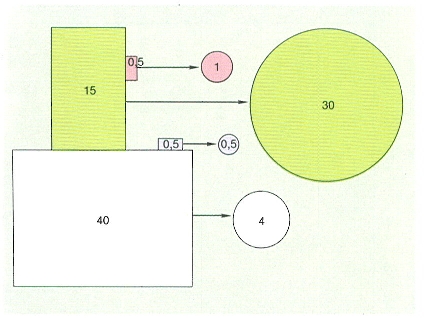
La producció de les comunitats fltobentòniques depèn de la disponibilitat hídrica (estatge mediolitoral) i de la llum (estatges infralitoral i circalitoral). L’hidrodinamisme moderat potencia la producció ja que afavoreix la captura de nutrients. La limitació de la producció pels nutrients és òbvia a les comunitats superficials de llocs encalmats.
Eduard Clavero, original d’E. Ballesteros.

Biomassa (quadrats i rectangles) i producció (cercles) dels diferents compartiments algals en una comunitat de Cystoseira mediterranea. S’indica en color verd l’espècie dominant (separada en ràmuls, tronc i base), en rosat els epífits, en blanc les espècies incrustants i en blau l’estrat «arbustiu».
Eduard Clavero, original d’E. Ballesteros.
Els rangs de producció primària de les comunitats d’algues bentòniques de la Mediterrània són molt variables. La comunitat més productiva és la de Cystoseira mediterranea, la qual produeix al voltant de 1000 g C/m2·any, equiparable al que produeixen les comunitats naturals més productives de la biosfera (cinyells de vegetació helofítica, selva tropical). En el món marí només trobem valors de producció lleugerament superiors en les comunitats oceàniques dominades per laminaríais i en els alguers de la fanerògama Thalassia testudinum. Aquesta elevada producció és molt més sorprenent si pensem en les baixes concentracions de nutrients de les aigües mediterrànies; el fort hidrodinamisme a què és sotmesa la comunitat n’és, sens dubte, el responsable. La producció algal disminueix amb la fondària de manera semblant a la disminució de la llum, però amb un pendent menys acusat a causa de l’increment d’eficiència fotosintètica a què ja hem fet referència. En el coral·ligen amb Halimeda tuna de la Costa Brava, a uns 25 m de fondària, la producció és només d’uns 35 g C/m2·any, és a dir, equivalent a la d’una zona terrestre semidesèrtica. La producció de les comunitats situades a intensitats de llum més baixes és encara més petita i en el límit de la zona fital és, òbviament, propera a zero.
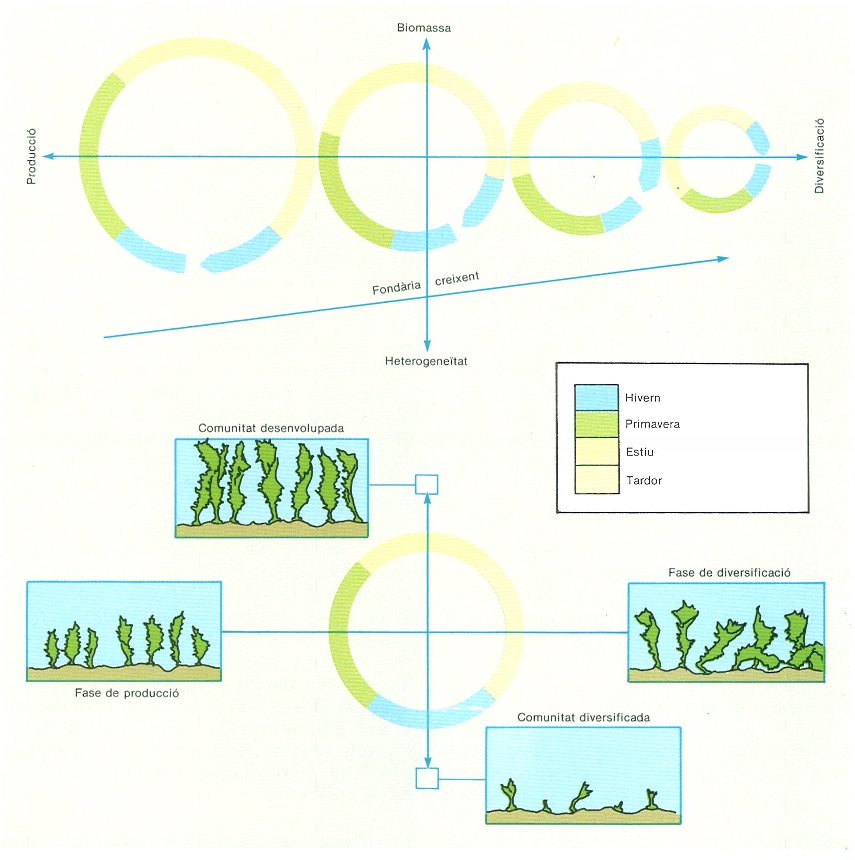
Representació de les comunitats fitobentòniques mediterrànies i de llur cicle anual segons els eixos producció/diversificació i biomassa/heterogeneïtat, i exemple en la comunitat de Rissoella verruculosa (estatge mediolitoral). El diàmetre dels cercles és proporcional a la variació estacional. Les comunitats superficials són les més productives, i les comunitats circalitorals, les més estructurades. Hi ha un desplaçament de primavera a estiu del moment de màxima producció als llocs profunds.
Jordi Corbera, original d’E. Ballesteros, modificat.
Aquestes dades ens fan veure l’escassa importància del bentos en la contribució a la producció primària global de la mar. Per donar xifres més comparables podem dir que, aproximadament, la producció del fitobentos en una costa rocosa equival a la producció del fitoplàncton situat entre la línia de costa i uns 2 km mar enfora. La producció fitobentònica per metre de línia de costa i any seria d’uns 125 kg de carboni. Si estimem, aproximadament, en 1.000 km la longitud de la costa de tots els Països Catalans i considerem que només un 50% és ocupada per roca, hom pot avaluar una producció primària global d’unes 62.500 t de carboni a l’any, que es poden arribar a duplicar si hom considera els valors de producció habituals en les comunitats de posidònia (Posidonia oceanica). Malgrat que la xifra de 125.000 tones de carboni a l’any que les caracteritzen pugui semblar elevada, hem de pensar que equival a la producció calculada per al fitoplàncton d’uns 1.600 km2 de mar Mediterrània, que també equival a quatre vegades la producció normal d’un alzinar mediterrani d’una extensió comparable a la que ocupa tota la ciutat de Barcelona.
La producció de les algues no es distribueix d’una manera uniforme al llarg de l’any sinó que, com passa a les comunitats vegetals dels països temperats, hi ha uns períodes favorables durant els quals les algues creixen molt més que en la resta de l’any: llevat d’algunes algues efemerofícies que aprofiten qualsevol moment favorable per a desenvolupar-se, la gran majoria de comunitats bentòniques concentren la producció en una època de l’any. Aquesta fase de producció coincideix amb el període en què hi ha un creixement net de l’espècie o espècies dominants i això està lligat al cicle anual dels dos factors que controlen el creixement algal: la irradiància i la disponibilitat de nutrients. Atès que els màxims d’irradiància es presenten a l’estiu (coincidint amb els mínims de nutrients) i els mínims a l’hivern (coincidint amb els màxims de nutrients), les algues dels diferents nivells han acoblat llurs cicles vitals al cicle anual del factor que els és més limitant. Per això el creixement màxim de les algues de les zones superficials té lloc a l’hivern i la primavera, mentre que els màxims de producció en les zones profundes es desplacen cap a l’estiu. Hom anomena fase de diversificació el període en què no hi ha un guany net de biomassa per a l’espècie o espècies dominants. En aquesta situació els processos dominants són els de consum i descomposició de la matèria orgànica produïda, la qual es recicla en part dins del mateix sistema local i en part s’exporta cap al sistema pelàgic, com també al sistema bentònic profund.
L’alteració de la dinàmica de les comunitats va associada a canvis d’estructura, i per això hom pot distingir, a manera d’abstraccions dins d’un continu, dos estadis successius: l’estadi de comunitat desenvolupada i l’estadi de comunitat diversificada. L’estadi de comunitat desenvolupada es dona al final del període de producció i es caracteritza pels valors elevats de biomassa i recobriment i per la gran estructuració espacial. L’estadi de comunitat diversificada es presenta al principi de la fase de producció; és caracteritzada per la seva baixa biomassa, el seu recobriment escàs i per una estructuració espacial mínima, ja que sovint hi manquen els estrats de vegetació més elevats o hi són pobrament representats. Com que moltes comunitats mediterrànies són dominades per una sola espècie, l’estadi de comunitat desenvolupada coincideix amb una diversitat baixa i una gran homogeneïtat, mentre que en l’estadi de comunitat diversificada la diversitat és alta i hi ha una marcada heterogeneïtat. Aquests estadis i fases són també ben distingibles en les comunitats d’algues mediolitorals.
Principals característiques adaptatives de les algues de superfície i de fondària per a maximitzar-ne la producció
| CARACTERÍSTIQUES | ALGA DE SUPERFÍCIE | ALGA DE FONDÀRIA | |
|---|---|---|---|
| Afinitat dels sistemes per a capturar nitrats, amoni, fosfats | + + + | + + | |
| Capacitat d’emmagatzemar midó | + | + | |
| Capacitat d’emmagatzemar nitrogen i fòsfor | + + | + | |
| Capacitat de translocar productes | + | + + | |
| Pigments especials que absorbeixen a la banda del verd | – | + + | |
| Concentració de clorofil·la a | + + | + + + | |
| Morfologia interceptora de la llum | – | + + + | |
| Estructuració de la comunitat amb una eficiència màxima per a la captació de la llum | – | ++ + | |
| FACTOR LIMITANT | NUTRIENTS | LLUM | |
| Dades dels autors | |||
Els alguers de fanerògames

L’alguer de posidònia (Posidonia oceanica) és una de les comunitats bentòniques mediterrànies més riques i més esteses. El dibuix correspon a l’herbei de les illes Medes (Baix Empordà), on distingim en un primer pla nombrosos epífits foliars com Fosliella (1), Gifforfia mitchelliae (2) i Castagnea irregularis (3); els hidraris Obelia dichotoma (4) i Sertularia perpusilla (5); els briozous Fenestrulina joannae (6), Disporella hispida (7), Lichenopora radiata (8) i Electra posidoniae (9); el poliquet Spirorbis (10); sobre els rizomes apareixen els briozous Platones stoechas (11) i Schizobrachiella sanguinea (12); en el substrat hi ha les garotes Sphaerechinus granularis (13) i Paracentrotus lividus (14), la pixota Holothuria forskali (15), l’estrella Echinaster sepositus (16), la nacra Pinna nobilis (17), el cloròfit Codium bursa (18) i conquilles de Lima lima (19). Les brànquies del poliquet Spirographis spallanzani (20) sobresurten entre les fulles de Posidonia; les xucles (Spicara maena) (21) i les julioles (Coris julis) (22) evolucionen per l’herbei.
J.Corbera / M.Zabala, original d’A.A. Ramos.
La vida en la mar comporta una adaptació al medi aquàtic d’algunes plantes amb progenitors terrestres. Les fanerògames presenten una sèrie de característiques que les fan més competitives que les algues. El fet de tenir rels els permet d’obtenir nutrients del lloc on estan arrelades, de manera que disposen de dues fonts potencials de substàncies minerals, l’aigua i el sediment. Les rels també els permeten de fixar-se en indrets on molt poques algues han reeixit a viure: els fons tous. La diferenciació morfològica i funcional dels diferents òrgans i teixits de les fanerògames és d’una complexitat impensable en una planta inferior, i això en facilita i perfecciona el metabolisme. Actualment, les fanerògames marines formen praderies submergides, anomenades herbeis o alguers arreu del món (llevat de les regions àrtiques), sobre substrat sedimentari i en zones ben il·luminades.
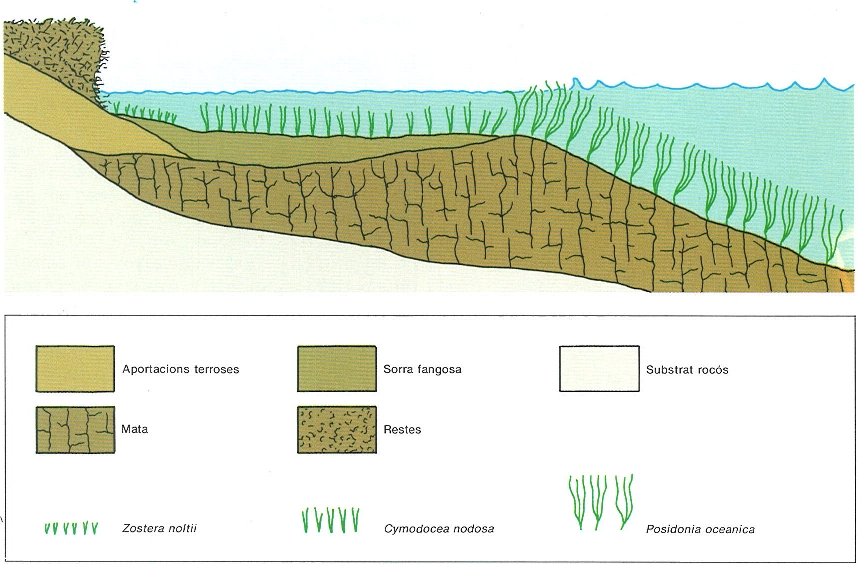
Als Països Catalans només viuen quatre espècies de fanerògames marines, de les quals Posidonia oceanica és, de molt, la més abundant i coneguda; espècie endèmica de la Mediterrània, forma extensos alguers a la zona infralitoral, principalment sobre substrat sorrenc, encara que també creix esporàdicament sobre roques; se la coneix també amb els noms equívocs d’alga o altina. Cymodocea nodosa és una espècie comuna que forma petits alguers al llarg de la costa; a Catalunya viu sovint en una certa fondària (15-25 m), encara que a les Balears i al País Valencià apareix principalment a les badies d’aigües somes i tranquil·les. Zostera noltii, molt més rara que les dues espècies anteriors, viu exclusivament sobre substrat fangós o llimós, sempre a poca fondària (0-5 m). Finalment, Zostera marina és una espècie raríssima localitzada només a Banyuls de la Marenda (Vallespir), Portlligat (Alt Empordà) i la badia dels Alfacs (delta de l’Ebre).
Els alguers de fanerògames marines tenen una gran importància en l’àmbit dels ecosistemes bentònics mediterranis. Constitueixen un deis sistemes estructuralment més complexos, sobretot si es té en compte que es desenvolupen sobre un sediment que, si no tingués aquest recobriment vegetal, estaria desproveït de qualsevol comunitat ben estructurada.
Els alguers de posidònia (Posidonia oceanica) són els més característics de la Mediterrània, els més estesos i ben estructurats, i també els més exhaustivament estudiats; per això, en aquest text hi farem referència d’una manera gairebé exclusiva. Tenen una acció molt important en la protecció de la zona costanera, ja que els seus rizomes fixen el sediment i formen una estructura que amorteix l’acció dels corrents i les onades. En aquest sentit, són el mitjà més segur i natural per a estabilitzar el sediment i impedir l’erosió de la línia de costa. Així mateix, els alguers són zones de refugi per a moltíssimes espècies de peixos, que a més hi troben aliment, hi dipositen llurs postes i utilitzen la zona com a «nursery». La fauna i flora associades a aquests fons són molt més riques que les dels fons sorrencs desproveïts d’alguers. I, sobretot, a causa de l’escassetat d’herbívors adaptats al consum d’aquesta planta d’origen terrestre, una part molt important de la producció de posidònia s’exporta en forma de detrits, en bona part cap a nivells més profunds i, per tant, deficitaris de producció primària autòctona. Tot això, el paper de protecció del litoral, la riquesa del seu poblament i l’exportació de la producció excedent fa dels alguers de posidònia una de les comunitats bentòniques mediterrànies més interessants i, alhora, la més necessitada de protecció, atesa la seva situació superficial i les múltiples agressions que rep (pol·lució, pesca a ròssec massa superficial, ancoratge, dragatge de fons, etc.).
Posidonia oceanica és una espècie constituïda per uns rizomes ajaguts o erectes, sovint totalment o parcialment recoberts pel sediment, proveïts d’una sèrie de rels que els fixen al substrat. Els rizomes tenen el gruix d’un dit i són recoberts d’esquames provinents de les restes dels pecíols d’antigues fulles. Aquestes fulles apareixen a les parts terminals dels rizomes i s’agrupen en feixos. Les fulles són en forma de cinta, d’aproximadament 1 cm d’amplada i d’una llargada variable, que oscil·la entre els 20 i els 100 cm. El seu creixement s’inicia a partir d’un meristema basal, de tal manera que les zones més joves són les més properes al rizoma. L’aparició de fulles noves es produeix a la part interna dels feixos, i així les fulles externes corresponen a les de més edat. Una de les característiques principals dels alguers és la creació d’un substrat amb un cert grau de persistència que possibilita la fixació i el creixement de molts organismes. La forta diferenciació morfològica de la posidònia ha contribuït a la constitució de dos poblaments ben diferents, adaptats a viure sobre les fulles o sobre els rizomes.
Les fulles són ocupades per espècies petites, de vida molt curta, estrategues de la r, especialment adaptades al ràpid i particular creixement que presenta el substrat orgànic. Les parts més joves de les fulles són ocupades per les espècies pioneres; les colònies són petites i l’important és la rapidesa en la fixació i el posterior desenvolupament. A les parts terminals el recobriment pot superar el 100% i es produeixen fenòmens típics de competència per l’espai entre els principals grups d’organismes. En termes generals, hom distingeix una sèrie de tipus morfològics bàsics: les espècies incrustants, les pulvinulars i les erectes. Les espècies incrustants s’adhereixen fortament a les fulles i la seva estratègia consisteix a ocupar ben aviat tot l’espai disponible; són les primeres a colonitzar les fulles. Algunes algues (Myrionema magnusii, Fosliella lejolisii, Dermatolithon litorale) i briozous (Electra posidoniae, Chorizopora brogniartii, Fenestrulina joannae) són clars exponents d’aquesta estratègia. Les espècies pulvinulars creixen directament sobre les fulles, com és el cas dels briozous Lichenopora radiata i Disporella hispida i del poliquet Spirorbis corrugatus, o bé sobre les espècies incrustants, com ho fan la majoria d’algues (Giraudia sphacelarioides, Myriactula gracilis). Les espècies erectes apareixen tant a la part basal de les fulles com a les parts senescents. Els hidraris tenen una sèrie de representants especialment adaptats a viure sobre les posidònies, amb una gran varietat de preferències (fulles joves/fulles velles, base/àpex, cara interna/cara externa, marges/cares). Destaquen Sertularella ellisii, Sertularia perpusilla, Campanularia assymetrica i Plumularia obliqua forma posidoniae. Les algues erectes (diverses Castagnea i Audouinella, per exemple) es desenvolupen a la part terminal de les fulles i poden recobrir totalment els àpexs de posidònia durant la primavera.
Els rizomes són ocupats per una gran diversitat d’organismes provinents de les comunitats dels fons rocosos. Quan l’alguer és dens, penetra poca llum i les espècies que hi creixen són preferentment esciòfiles. Hi dominen les algues (Peyssonnelia, Udotea, ceramiàcies), els briozous (Schizobrachiella sanguinea, Schizoporella dunkeri, Platonea stoechas), les esponges (diverses Dysidea i Ircinia) i el foraminífer Miniacina miniacea. Si l’alguer és esclarissat, les algues fotòfiles són els principals colonitzadors dels rizomes i la comunitat és molt semblant als poblaments d’algues de substrat rocós de la zona infralitoral, amb Padina, Halopteris, Dasycladus, Udotea i un llarg etcètera. La part enterrada dels rizomes és ocupada per una fauna pròpia dels fons tous, rica en bivalves, poliquets, nematodes, copèpodes i petits decàpodes.
L’alguer manté també una rica fauna vàgil. Hi destaca el principal consumidor de posidònia, el vogamarí vermell (Paracentrotus lividus). Aquesta espècie s’alimenta de nit i al capvespre, quan s’enfila a les fulles per rosegar-les; durant el dia s’amaga entre els rizomes. Pot assolir concentracions de 40 individus per metre quadrat i, en cas de superpoblació, pot arribar a devastar extenses zones d’alguer. Un altre vogamarí, Sphaerechinus granularis, s’alimenta bàsicament de les algues epífites dels rizomes. Diversos cargols herbívors recorren les fulles (espècies de Rissoa, Rissoina bruguierei, Bittium reticulatum). En el grup dels crustacis també hi ha consumidors de posidònia (Idothea hectica, Achaeus cranchii, Pisa nodipes). Finalment, les salpes (Sarpa salpa), peixos de la família dels espàrids, són també consumidors importants de les altines: la visió de grans bancs de salpes, pasturant en una mena de dansa que omple la mar de reflexos argentats, s’associa als herbeis de posidònia. La posidònia és, però, una planta provinent del medi terrestre, i els herbívors marins no tenen molts dels enzims necessaris per a digerir-la bé. Les excrecions d’aquests herbívors, riques en fragments de posidònia parcialment digerits, són atacades per microorganismes descomponedors. Aquests detrits són llavors ingerits per les holotúries (Holothuria tubulosa), organismes força abundants en els alguers.
L’estructura de l’alguer de posidònia (Posidonia oceanica) és regida per la particular morfologia de l’espècie. La posidònia ocupa l’espai mitjançant el creixement del rizoma en sentit horitzontal i això li permet colonitzar el substrat del voltant. Els rizomes també creixen en alçada i, si la població és molt densa, es produeix un aixecament generalitzat del fons. Hi ha un creixement continu dels rizomes per la seva part apical mentre que les parts inferiors moren progressivament. Es crea, doncs, una estructura, la mata, constituïda per rizomes inactius entrelligats amb sediment i material detrític provinent dels organismes de l’alguer, principalment de la mateixa posidònia. El creixement dels rizomes cap amunt s’ha interpretat com una estratègia per a evitar de quedar colgats pel sediment i per a deslliurar-se de la competència per la llum dels feixos adjacents. Una acumulació massa ràpida de sediment provoca la mort de la part apical dels rizomes; si l’acumulació, en canvi, no és suficient, o bé si hi ha erosió, pot produir-se el trencament dels rizomes, i l’alguer es destrueix. Quan hi ha un equilibri creixement-sedimentació, les fulles i la mata subjacent s’aixequen lentament cap a la superfície. Aquest aixecament és molt lent a escala humana però considerable a una escala més gran. Ha estat estimat en una mitjana d’1 cm/any.
Secció esquemàtica d’un escull barrera de posidònia (Posidonia oceanica). El creixement de la mata provoca l’arribada de les fulles a la superfície (altina). L’espai entre la costa i l’altina, d’aigües encalmades, és generalment ocupat per Cymodocea nodosa i Zostera noltii.
Jordi Corbera, original de J.M. Pérès i J. Picard, modificat.
En els llocs més arrecerats de les badies l’aixecament progressiu de l’alguer pot fer-lo arribar a la superfície. En un primer estadi, són els feixos més propers a la costa els que l’assoleixen; més endavant, però, els feixos de les zones més fondes també hi arriben, i això provoca un estancament de l’aigua vora la costa, que causa la mort dels feixos més interns. A la llarga es crea un escull barrera constituït per feixos vius de posidònia que separa la mar lliure d’una zona d’aigües estancades, desproveïda de posidònia, on s’instal·len comunitats d’algues fotòfiles o bé alguers de cimodocea (Cymodocea nodosa). En l’actualitat, els esculls barrera de gran part de la Mediterrània han desaparegut a causa de l’activitat humana ja que l’home hi ha construït ports i/o ha modificat llurs condicions originals perquè són indrets molt acollidors. Als Països Catalans no tenim referència de cap escull barrera ben constituït, però en trobem restes o indicis en alguns indrets de l’Alt Empordà i de la costa mallorquina.
Mapa de distribució de l’alguer de posidònia (Posidonia oceanica) en un fons de l’illa de Tabarca (Alacantí), on domina àmpliament.
J.Corbera / M.Zabala, original d’A.A. Ramos
L’alguer de posidònia té una gran heterogeneïtat espacial. La distribució dels feixos és de contagi, de tal manera que hi ha parts molt denses i d’altres que estan desproveïdes de posidònia. Aquestes clarianes sense posidònia, anomenades calbes, tenen l’origen en zones concretes on la mata s’enfonsa. Les onades i els corrents poden ampliar aquest forat i deixar amples taques de sorra entremig de l’alguer, sovint conformades en canals més o menys perpendiculars a la línia de la costa.
Els alguers de posidònia tenen un límit batimètric inferior determinat per la irradiància. Aquest límit se situa prop dels 25 m a les costes peninsulars i dels 35 m a les insulars. Hom ha observat, però, feixos isolats a fondàries superiors. L’estructura de l’alguer varia segons la fondària; en general, s’observa una disminució de la biomassa amb la profunditat, a causa d’una abundància més gran de calbes i d’una densitat més petita de feixos dins de la mata. A l’alguer de les illes Medes, per exemple, a 5 m de fondària, el 80% de la superfície és cobert de posidònia amb una densitat de 600 feixos/m2; en canvi, a 13 m de fondària, el percentatge de superfície ocupada és només del 40%, i la densitat, de solament 350 feixos/m2.
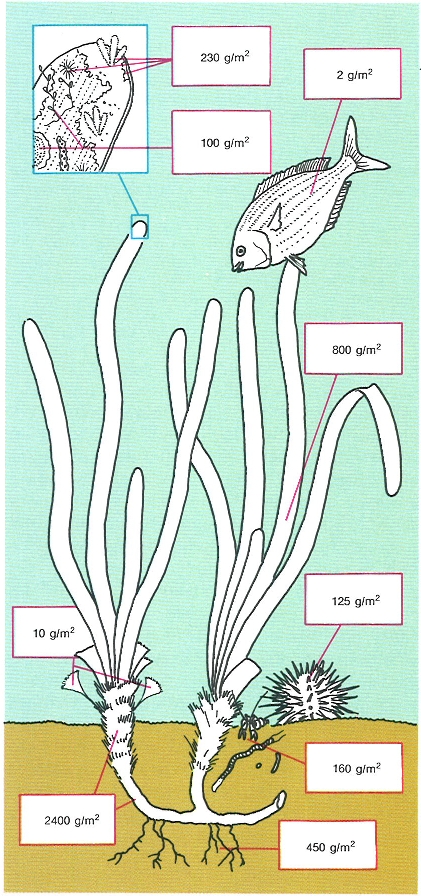
Estructura i distribució de la biomassa (g pes sec/m2) en un alguer de posidònia (Posidonia oceanica) durant la fase de maduresa. Les variacions estacionals afecten principalment les fulles i els epífits, compartiments que redueixen la seva biomassa fins a una cinquena part, aproximadament.
Jordi Corbera, amb dades de C.F. Boudouresque i A. Meinesz i de J. Romero i E. Ballesteros.
En conjunt, l’alguer de posidònia és la comunitat bentònica mediterrània que manté una biomassa viva més gran. En un alguer tipus, situat a 5-10 m de fondària, la biomassa total és d’uns 4,3 kg (pes sec)/m2, equivalent a 43 t/ha, de la qual un 90% és biomassa vegetal. L’alguer de posidònia, com les comunitats algals, té un cicle anual molt característic. La fase de producció s’inicia a l’octubre i, en un primer període, els feixos són constituïts per fulles curtes, d’aparició recent, i poc epifitades. El creixement augmenta gradualment fins al mes de febrer, en què s’observa un decreixement momentani de la producció. En aquest primer període, anomenat fase juvenil, part del creixement sembla realitzar-se gràcies a la mobilització del midó acumulat en els rizomes com a reserva durant l’estiu. La petita baixada de producció del mes de febrer pot atribuir-se a l’esgotament del midó. A partir d’aquest moment, però, el creixement continua augmentant i tot el material produït en la fotosíntesi s’inverteix en creixement foliar. No hi ha, però, producció de noves fulles, sinó creixement de les ja formades. Els epífits tenen també un desenvolupament important a causa de l’augment de la superfície foliar i de l’edat creixent de les fulles, la qual cosa augmenta les possibilitats de colonització. Als alguers d’aigües somes es produeix un creixement molt ràpid d’algunes espècies de petits feòfits que donen als àpexs de les fulles un color bru i un aspecte pilós. Durant el mes de juny el creixement de les fulles és màxim; ens trobem dins l’estadi de comunitat desenvolupada o fase de maduresa. La fase de diversificació o senectut s’inicia al juliol, amb una baixada molt important del creixement. Els epífits tenen la biomassa màxima, almenys en els alguers no situats en aigües somes, on la persistència de les fulles és més gran. El material foliar envelleix i, encara que la biomassa foliar és màxima, una gran part és metabòlicament poc activa. La fotosíntesi es manté, però, relativament elevada, i els productes fotosintetitzats s’inverteixen en la producció de noves fulles (primordis) i, sobretot, en la síntesi de midó, que s’acumula en els rizomes. La caiguda de les fulles comença durant el mes d’agost i és màxima durant el de setembre.
És interessant de constatar que el cicle de creixement és semblant a qualsevol alguer, independentment de la fondària on es trobi. Hi ha, però, una petita diferència: els alguers profunds tenen un retard temporal en diversos aspectes del cicle (creixement de les fulles i dels epífits, caiguda de les fulles), com passa a les comunitats algals. La producció és també més petita als alguers profunds a causa, bàsicament, de la menor densitat de l’alguer i no d’una menor producció per feix; això suggereix que hi ha una acomodació del sistema fotosintetitzador a condicions de baixa irradiància.
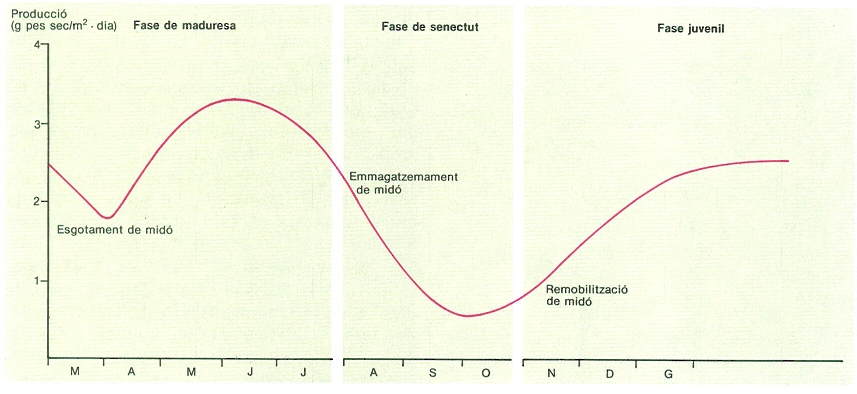
Cicle anual de producció de fulles de posidònia (Posidonia oceanica) a l’alguer de les illes Medes (Baix Empordà). Si bé encara com a hipòtesi, l’emmagatzemament i la remobilització de midó permetrien explicar el creixement tardorenc i la minva de creixement durant l’estiu, tot i que la planta té, en aquest darrer període, una elevada taxa de fixació de carboni.
Eduard Clavero, amb dades de J. Romero.
El cicle de la posidònia està internitzat, és a dir, es comporta d’una manera semblant, en condicions ambientals constants, any rere any. El creixement de fulles no es relaciona, però, de forma clara amb cap dels factors ambientals que, versemblantment, podrien afectar-lo. Això significa que hi ha hagut una adaptació integrada als diferents factors biòtics i abiòtics que marquen el ritme estacional. El creixement hivernal, sustentat en part per les reserves de midó, permetria a la planta crear nou teixit fotosintetitzador en un període en què els fitoepífits no tenen condicions favorables per a créixer, a causa de les baixes irradiàncies. El creixement de les fulles durant la primavera i l’estiu, quan no hi ha nutrients dissolts a l’aigua, podria sustentar-se per l’absorció radicular o per un hipotètic magatzem de compostos nitrogenats en els rizomes. Durant l’estiu, el més «econòmic» seria emmagatzemar hidrats de carboni per a utilitzar-los durant la tardor següent.
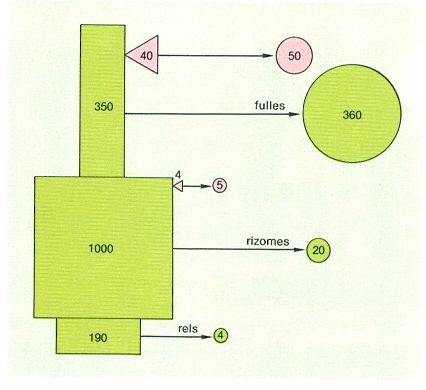
Biomassa (quadrats i rectangles) i producció (cercles) dels diferents compartiments vegetals en l’alguer de posidònia (Posidonia oceanica), representats en verd, i dels seus epífits, en rosat.
Eduard Clavero, amb dades de C.F. Boudouresque i A. Meinesz i de J. Romero i E. Ballesteros.
La producció primària d’un alguer tipus de posidònia es pot estimar entre els 400 i els 500 g C/m2·any, de la qual més d’un 80% correspon a producció foliar, un 5% a producció dels rizomes i la resta a producció de les algues epífites. L’alguer de posidònia ocupa, doncs, la segona posició en el barem de producció de les comunitats fitobentòniques mediterrànies, només superada per la comunitat de Cystoseira mediterranea. En valors absoluts, i ateses les grans superfícies que ocupen, els alguers de posidònia són probablement els principals productors bentònics de la Mediterrània. Una gran part de la producció es queda dins l’alguer en part com a material edificador de la mata i en part com a aliment per als herbívors; almenys un 30%, però, és exportada a d’altres sistemes: una petita part és acumulada a les platges, on serveix d’aliment a la fauna de les zones supralitoral i mediolitoral; els corrents s’emporten la resta, sovint acomboiats pels canals del mateix alguer, arreu de la plataforma continental i, també, als fons de l’estatge batial. Un cop dipositades, les fulles senceres o fragmentades de posidònia són atacades pels descomponedors (principalment bacteris). Els consumidors d’aquestes restes només aprofiten els bacteris i eliminen les restes de les fulles, que són novament atacades pels descomponedors. La importància de la posidònia com a material primari en les cadenes alimentàries de les poblacions profundes i de les que hi ha en les coves submarines és desconeguda, però ben segur que no és negligible, almenys localment: el sòl d’alguns trams ben foscos de coves de Mallorca i de les illes Medes es troba completament cobert de detrits de posidònia, la qual cosa comporta, d’altra banda, una entrada considerable de matèria orgànica en un ambient on és molt escassa.
El coral·ligen
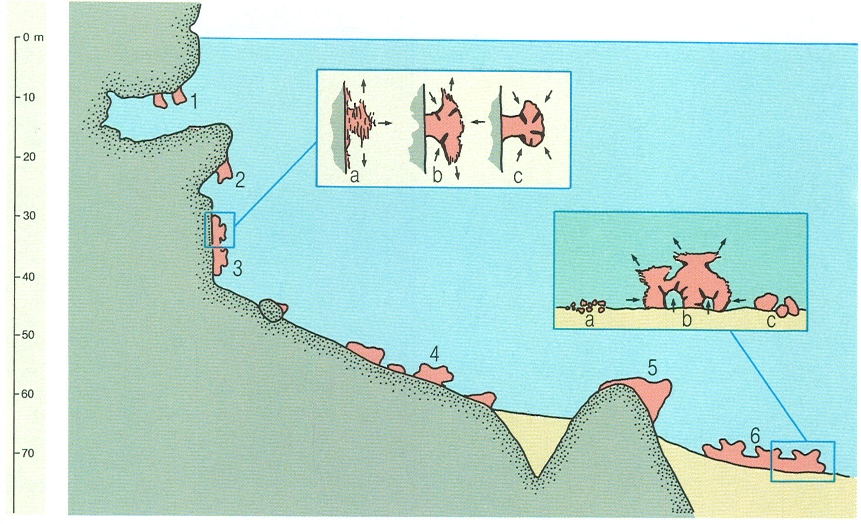
Diferents tipus de concreció coral·lígena segons la seva situació i el substrat sobre el qual es desenvolupa. Dins del coral·ligen de roca costanera hom distingeix el d’entrades de coves (1), el de cornises (2), el de parets verticals (3), el de relleixos i esllavissades del talús (4) i el de recobriment sobre sòcols rocosos (5). Aquest darrer és bionòmicament molt semblant al coral·ligen de plataforma (6), originat sobre substrats tous. En el dibuix s’han representat, dins dels requadres, els processos de formació (a) i de degradació (b, c) de les concrecions coral·lígenes.
Jordi Corbera, original de J. Laborel, modificat.
Sobre substrat dur, les algues constitueixen el grup dominant al bentos marí mediterrani, si la llum és prou elevada. El descens de la intensitat lumínica associat a la fondària o a les particularitats del relleu submarí (extraploms, entrada de coves) comporta una minva important del creixement algal. Les primeres algues afectades pertanyen a l’estrat erecte. Les comunitats esdevenen menys atapeïdes, la qual cosa dona cabuda a algunes espècies d’animals suspensívors. L’estrat incrustant d’algues calcàries, en veure’s desproveït d’un estrat d’algues erectes que en situacions d’elevada irradiància competeixen favorablement amb les calcàries en la captació de llum i nutrients, en resulta afavorit. La calcificació dels tal·lus n’assegura la perdurabilitat perquè, tot i tenir un creixement lent, són difícilment atacades pels herbívors. El resultat és un desenvolupament força important de les algues carbonatades, els tal·lus de les quals competeixen per la llum: es donen fenòmens de superposició i recobriment d’uns tal·lus per uns altres. El creixement de les algues no és, tampoc, homogeni. Determinats suspensívors creixen en alçada i formen un estrat erecte que els permet un filtratge més eficient de l’aigua i fa disminuir la llum que arriba als tal·lus algals. D’altres suspensívors recobreixen les algues, afavorits per les baixes intensitats de llum. Les cares inferiors dels tal·lus i els espais buits deixats per les algues en el seu creixement són ocupats també per una multitud d’organismes suspensívors, carnívors i detritívors. Els esquelets d’algues i animals morts reomplen els buits creats. Aquests materials s’acumulen per sedimentació i es compacten i se cimenten per processos químics. A 20 cm de fondària l’estructura ja és totalment cimentada i les restes dels organismes que l’han originat són completament indistingibles. Els organismes perforants obren noves galeries i en provoquen l’esfondrament de petites parts. La destrucció pot ser causada també per processos físics (hidrodinamisme) o químics (redissolució dels carbonats). D’aquesta manera, a poc a poc, la complicada estructura formada sobre la base de les algues carbonatades i ocupada per una munió d’organismes animals va creixent en potència en un procés continu de construcció i destrucció. Estem assistint als processos de formació d’una estructura molt heterogènia, pròpia del bentos mediterrani i amb una complexitat estructural i una bellesa molt superiors a les comunitats que hem comentat fins ara: el coral·ligen.

Elements principals de les comunitats de les illes Medes. Representació idealitzada del coral·ligen corresponent a uns 35 m de fondària, com a exemple d’aquest tipus de comunitat, amb una dominància gairebé exclusiva de suspensívors. S’hi han indicat: l’alga coral·linàcia Lithophyllum expansum (1), les esponges Cliona viridis (2), Acanthella acuta (3), Agelas oroides (4), Spirastrella cunctatrix (5), Chondrosia reniformis (6), Petrosia ficiformis (7), Clathrina clathrus (8) i Dysidea avara (9); els hidraris Halecium halecinum (10), Aglaophenia septifera (11), els antozous Synthecium evansi (12), Paramuricea clavata (13), Eunicella singularis (14), Alcyonium acaule (15), Caryophyllia inornata (16), Parazoanthus axinellae (17) i Corallium rubrum (18), el poliquet Serpula vermicularis (19), els mol·luscs Pteria hirundo (20) i Peltodoris atromaculata (21); crustacis com el sastre (Galathea strigosa) (22) i la llagosta (Palinurus elephas) (23); el briozou Porella cervicornis (24) i l’ascidi Halocynthia papillosa (25); i diversos peixos: cabuts (Anthias anthias) (26), variades (Diplodus vulgaris) (27), la castanyola (Chromis chromis) (28) i el nero (Epinephelus guaza) (29).
Jordi Corbera / Mikel Zabala
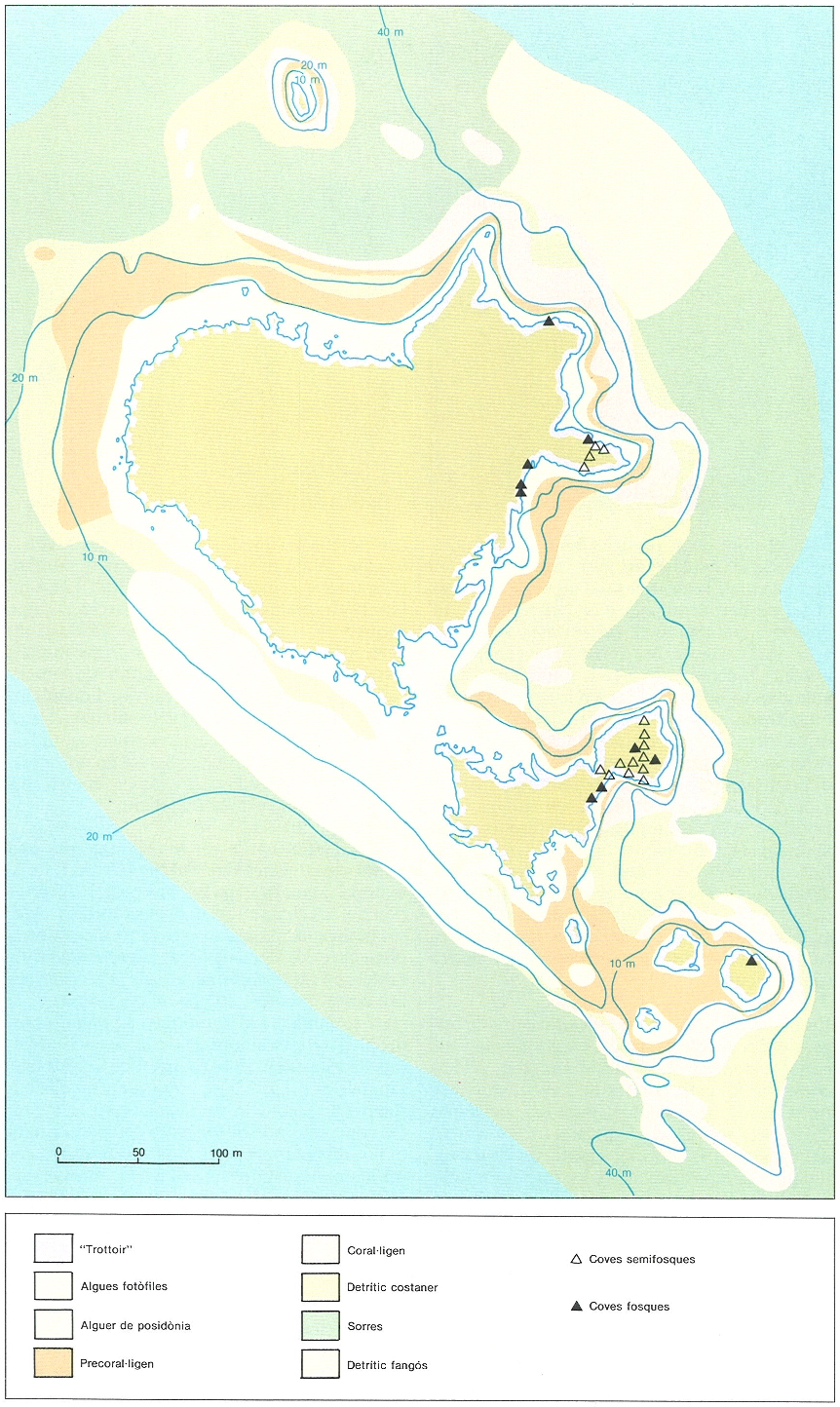
Extensió del coral·ligen i el precoral·lígen als vessants N i E de les illes Medes, de fons abruptes sotmesos a forts corrents. S’hi fa palesa la gran extensió de la comunitat i l’asimetria amb el vessant S, de fons sorrencs, amb un reduït alguer de posidònia.
Jordi Corbera / Mikel Zabala
Així, el coral·ligen pot ser interpretat com una estructura organògena, això és, construïda pels organismes, pròpia dels fons circalitorals de la Mediterrània. Els principals organismes edificadors d’aquesta estructura són les algues carbonatades pertanyents bàsicament a la família de les coral·linàcies. Això no obstant, les algues no acostumen a ser el grup dominant en termes de nombre d’espècies o de biomassa, privilegi que correspon als animals suspensívors. La seva distribució batimètrica varia segons la intersecció de diversos factors ambientals, entre els quals hi ha la llum, l’hidrodinamisme, la concentració de matèria orgànica particulada, la disponibilitat de nutrients i els factors biòtics. Aquests paràmetres afecten diferentment les algues i els animals suspensívors. Quan les condicions globals impedeixen el creixement esponerós de les algues toves, afavoreixen el desenvolupament de l’estrat incrustant i permeten l’ocupació de l’espai pels suspensívors; llavors el coral·ligen s’estableix. Aquesta situació es produeix des dels 25-30 m fins als 60-70 m de fondària a la Costa Brava, i dels 40-45 m als 110-150 m a les costes insulars. En aquestes fondàries, el coral·ligen esdevé la comunitat climàcica. Això no obstant, pot desenvolupar-se a fondàries inferiors, en parets fortament inclinades o verticals, orientades al N, que reben irradiàncies molt inferiors a les que els pertocaria per la fondària.
Sota el nom de coral·ligen hom agrupa tota una sèrie de poblaments molt diversos que tenen en comú l’estrat basal d’algues calcàries i el fet de pertànyer a l’estatge circalitoral. El desenvolupament dels organismes suspensívors varia segons les zones. En general és màxim en els indrets menys il·luminats, amb abundància de matèria orgànica particulada i un hídrodinamisme accentuat, causat per corrents intensos que sovintegen a la zona. Els diversos grups de suspensívors estan també diferentment representats segons aquestes característiques. Les esponges, els ascidis i els briozous, per exemple, són presents amb abundància a qualsevol tipus de coral·ligen. Les gorgònies, en canvi, tenen uns requeriments més estrictes pel que fa a la mida i la densitat de les partícules en suspensió, i això és la causa, per exemple, de la seva rarefacció a les costes insulars, més pobres en material particulat.
En el cas de màxima complexitat hom distingeix els següents estrats. Les gorgònies i algunes esponges de forma arborescent poden constituir un estrat elevat de més de mig metre d’alçada, generalment molt lax, però que en indrets sotmesos a corrents forts i a irradiàncies molt baixes pot assolir un desenvolupament espectacular. L’estrat intermedi, constituït per diverses colònies massives de briozous, algunes esponges, ascidis, hidraris i poliquets, és de densitat variable, molt condicionada també per la disponibilitat d’aliment. L’estrat incrustant és format per les algues calcàries i per diverses espècies de madreporaris, esponges i briozous que competeixen amb les algues per l’ocupació de l’espai. Al seu dessota, els forats de la concreció coral·lígena mantenen un poblament d’espècies adaptades a la foscor, idèntiques sovint a les que viuen a les coves, i un cert nombre d’esponges i mol·luscs endolítics. Els epibionts de mida petita creixen sobre els que formen els substrats erectes més elevats i inclouen representants de tots els grups animals i també algues no carbonatades de desenvolupament primaveral o estival. Finalment, hi ha una rica fauna vàgil (poliquets, altres verms, mol·luscs, crustacis, picnogònides, equinoderms, peixos) que habita en els intersticis de l’estructura coral·lígena i recorre tots els racons a la recerca d’aliment.
El coral·ligen ha de ser interpretat com una unitat paisatgística dins del bentos marí, de la mateixa manera com ho és l’alguer de posidònia o les comunitats d’algues fotòfiles. Ateses la seva localització i la seva formació, hom distingeix dos tipus de coral·ligen: el que s’estableix en els nivells inferiors de la zonació sobre costes rocoses, anomenat coral·ligen de la roca costanera, i el coral·ligen de plataforma, propi d’indrets allunyats de la costa i envoltats de fons tous. El coral·ligen de la roca costanera pot presentar-se a diferents nivells batimètrics, segons la irradiància que hi arribi. La concreció no sol ser gaire gruixuda i hom hi pot distingir aspectes molt diferents. El coral·ligen de plataforma, en canvi, forma masses força potents, solcades de canals i túnels, la mida i consistència de les quals depenen de l’activitat constructiva de les espècies concrecionants; una mida tipus, però, pot ser la de blocs de 4-6 m de diàmetre per 2-3 m d’alçada. La situació aïllada d’aquests blocs fa pensar que han estat originats sobre substrats tous. És possible, però, que la seva instal·lació només sigui possible si hi ha substrats durs que, encara que siguin de mida molt petita, tinguin prou estabilitat per a permetre l’inici de la concreció. Les cornises i les platges fòssils que voregen les nostres costes com a testimoni de les variacions del nivell de la mar durant el Miocè afavoreixen, sens dubte, la instal·lació del coral·ligen de plataforma. Els fons de grapissar i cascall són també àrees favorables per a la formació d’aquest coral·ligen. Bé que alguns fons de grapissar són rics en espècies d’algues laminars, la seva contínua mobilitat fa difícil la formació d’un nucli estable. L’eventual presència de petits sortints rocosos facilita enormement la concreció. El fet que els fons tous de la Mediterrània estiguin literalment sembrats de restes de naufragis, àncores i altres restes de l’antiguitat ha afavorit, sens dubte, la formació de molts blocs de coral·ligen de plataforma.
Independentment de la seva localització, el coral·ligen ofereix diferents aspectes segons els organismes que el constitueixen. La seva tipificació és, però, més difícil que en cap altra comunitat bentònica, ja que la seva complexitat intrínseca permet la coexistència en un espai molt reduït d’espècies amb requeriments ecològics molt diferents. Essent conscients d’aquestes limitacions, podem comentar alguns dels aspectes que ofereix el coral·ligen a les costes dels Països Catalans.
En l’aspecte més fotòfil, el coral·ligen es desenvolupa sobre substrats horitzontals o poc inclinats i la seva concreció és escassa. Les algues erectes hi poden ser ocasionalment abundants i constitueixen la comunitat de Cystoseira zosteroides, ja comentada. Entre els organismes suspensívors podem remarcar algunes gorgònies (Eunicella singularis) i alcionaris (Alcyonium acaule), esponges (Petrosia ficiformis, Ircinia dendroides, Spongia virgultosa, S. agaricina, S. officinalis), briozous (Myriapora truncata, Turbicellepora avicularis, Porella cervicornis, Pentapora fascialis) i tunicats (Aplidium tabarquensis, Pseudodistoma crucigaster, Microcosmus sabatieri, M. polymorphus, M. nudistigma, Pyura durà). És abundant a totes les costes rocoses dels Països Catalans, a fondàries discretes (15-20 m). A fondàries superiors el recobriment algal disminueix i, en canvi, el coral·ligen s’enriqueix amb nous filtradors, sobretot la gorgònia Eunicella cavolinii.
A les parets molt inclinades de les roques costaneres pot haver-hi una forta concreció causada per les algues coral·linàcies Mesophyllum lichenoides i Lithophyllum expansum. Si la irradiància és suficient, les algues erectes poden ser abundants, i hi dominen Halimeda tuna, Udotea petiolata i diverses Peyssonnelia. Els organismes suspensívors recobreixen totalment les anfractuositats i les bases de les algues incrustants. Hom hi distingeix, com a elements principals, les esponges Faciospongia cavernosa, Anchinoe tenacior, Axinella damicornis, A. verrucosa, Oscarella lobularis, Chondrosia reniformis i Petrosia ficiformis; els cnidaris Eudendrium racemosum i Clavularia ochracea; els bivalves Arca noae i Spondylus gaederopus; els briozous Myriapora truncata, Sertella septentrionalis i Hincksina flustroides; i els tunicats Halocynthia papillosa i Microcosmus sabatieri. Els llocs sotmesos a forts corrents s’enriqueixen d’organismes filtradors que ocupen les superfícies externes del coral·ligen: esponges (Hippospongia communis, Spongia officinalis), cnidaris (Alcyonium acaule), grans briozous (Porella cervicornis, Pentapora fascialis, Hornera frondiculata) i tunicats (Aplidium conicum, A. tabarquensis, Eudistoma banyulensis).
La constitució específica del coral·ligen de plataforma és molt semblant a la descrita aquí. Entre les diferències més remarcables hi ha la substitució de Mesophyllum lichenoides per d’altres espècies d’algues inscrustants (principalment Neogoniolithon mamillosum i Peyssonnelia rosa-marina), espècies que donen lloc a un coral·ligen amb poques microcavítats però, en canvi, amb molts petits túnels i anfractuositats més grans.
El coral·ligen amb Paramuricea clavata, propi de llocs molt poc il·luminats sotmesos a corrents intensos, constitueix, probablement, el paisatge submarí més bonic i divers de tota la Mediterrània. Sota un estrat elevat d’aquesta gorgònia multicolor, els suspensívors assoleixen un desenvolupament extraordinari. La llista seria gairebé interminable, però entre les espècies més abundants podem destacar un elevat nombre d’esponges (Chondrosia reniformis, Petrosia ficiformis, Axinella verrucosa, A. polypoides, Acanthella acuta, Ircinia fasciculata, I. oros, Dictyonella obtusa, Faciospongia cavernosa), diversos antozous (Caryophyllia smithi, Hoplangia durotrix, Parazoanthus axinellae, Leptopsammia pruvoti), poliquets (Filograna implexa), briozous (diverses Schizomavella, Pentapora fascialis, Porella cervicornis, Adeonella calveti, Cellaria fistulosa), el cirrípede Balanus perforatus i alguns ascidis (Halocynthia papillosa, Eudistoma planum, Cystodytes dellechiajei). Totes aquestes espècies i moltes d’altres competeixen activament per l’espai i es recobreixen les unes a les altres, sense deixar un sol buit. L’espectador d’una paret vertical amb aquests poblaments queda admirablement sorprès; la llum d’un focus potent sobre aquests fons produeix un esclat de grocs, vermells, taronges, blaus, verds, blancs i roses, que es combinen en una simfonia de colors inimaginable. Després d’aquesta visió hom s’adona de la pobresa de colors del món emergit, dominat pel color de les roques, el blau de la mar i el verd de les plantes. Aquesta comunitat, probablement el representant més genuí dels fons coral·lígens de la Mediterrània occidental, és particularment abundant a la zona del golf del Lleó, des de Portvendres fins a Palamós. Apareix també als Columbrets, la costa alacantina i en algun indret molt concret de la costa balear.
A intensitats lluminoses inferiors a les dels aspectes comentats fins ara i, per tant, en llocs situats a més fondària o en zones extraplomades i entrades de coves i túnels, la cobertura algal disminueix, la concreció és molt inferior i l’estrat elevat de gorgònies i grans esponges té una densitat més petita o és inexistent. Els poblaments són dominats per les esponges (Axinella vaceleti, A. damicornis, Agelas oroides i les esmentades a l’aspecte anterior) i els antozous Parazoanthus axinellae i Leptopsammia pruvoti. Si la sedimentació és escassa i la disponibilitat de matèria particulada ho permet, apareix el corall vermell (Corallium rubrum), malauradament molt malmès a causa de la seva intensa explotació. Aquest poblament és pràcticament indistingible del de les coves semifosques, del qual parlarem més endavant.
Els aspectes que hem comentat del coral·ligen no són els únics que hom pot trobar. De fet, encara que les situacions tipus són freqüents, el coral·ligen és extraordinàriament divers i molt sovint el que trobem són situacions mixtes o, fins i tot, molt diferents dels aspectes que hem descrit. La seva gran heterogeneïtat a petita i gran escala i la fondària considerable on es troben la majoria de poblaments en dificulta moltíssim l’estudi, i l’explicació dels processos que intervenen en la seva constitució no deixa d’estar en el terreny de les hipòtesis.
A part els organismes a què hem fet referència, sèssils i de mida gran, hi ha una enorme diversitat d’animals vàgils o sèssils dels mateixos grups o bé d’altres, que contribueixen a la diversitat del coral·ligen. Una llista mitjanament exhaustiva seria del tot improcedent aquí; en farem, doncs, una síntesi sumària, no sense remarcar que moltes espècies viuen en aquesta comunitat en les seves primeres etapes larvals o juvenils.
Els organismes epibiòtics són molt nombrosos i pertanyen al grup dels hidraris (Nemertesia antennina, Aglaophenia kirchenpaueri), els antozous (Parerythropodium coralloides), els poliquets (Filograna implexa), els briozous (diverses espècies de Bugula, Scrupocellaria i Aetea) o els mol·luscs (Pieria hirundo). La fauna vàgil és representada per diversos poliquets (Eunice, Nereis, Pontogenia), l’equiürid Bonellia viridis, diversos amfípodes (Lilljeborgia brevicornis, Maera inaequipes), grans decàpodes (Palinurus elephas, la llagosta, i Hommarus gammarus, el llobregant, en són els més coneguts), gasteròpodes prosobranquis (Acmaea virginea, Fusinus rostratus, Muricopsis cristatus), opistobranquis (Peltodoris atromaculata, Hypselodoris luteorosea, Flabellina affinis), bivalves (Anomia ephippium, Chlamys multistriatus, Chama gryphoides), ofiures (Ophioíhrix fragilis, Amphipholis squamata), estrelles de mar (Echinaster sepositus, Ophidiaster ophidianus) i garotes (Sphaerechinus granularis, Echinus acutus). Totes aquestes espècies i d’altres recorren la superfície del coral·ligen o bé viuen dins les nombroses cavitats, i sovint es comporten més com a organismes fixos que no pas vàgils (els bivalves, Bonellia, etc.). Els peixos són també força abundants, principalment els cabuts (Anthias anthias), peixos suspensívors d’un llampant color vermell i groc; els xuclans (Labrus bimaculatus), cap-roigs (Scorpaena scrofa i altres escórpores), congres (Conger conger) i corballs (Sciaena umbra). El coral·ligen és també un dels indrets on l’escafandrer pot ser obsequiat amb la visió cada cop més rara d’un nero (Epinephelus guaza).
Hi ha molts pocs estudis experimentals que versin sobre la dinàmica del coral·ligen. La producció primària és molt baixa a causa de l’escassa importància que tenen les algues. Hom ha avaluat la producció d’algues erectes en 68 g C/m2·any per una comunitat de Cystoseira zosteroides, i de 35 g C/m2·any per una comunitat d’Halimeda tuna, a la Costa Brava. La producció algal al coral·ligen de Paramuricea és, sens dubte, molt inferior: ateses les dades de biomassa algal, no sobrepassa els 10 g C/m2·any. En el coral·ligen de corall vermell (Corallium rubrum) la producció primària és totalment negligible: és inferior a 5 g C/m2·any). El creixement de les espècies incrustants no augmenta gaire aquestes xifres. En el millor dels casos hom pot avaluar la fixació de carboni de Mesophyllum lichenoides i Lithophyllum expansum En uns 12 g C/m2·any dels quals només 5 formen part de matèria orgànica. El creixement del coral·ligen en potència és, doncs, molt lent. Mitjançant la mesura de l’alçada del coral·ligen de plataforma edificat sobre àmfores de l’època romana hom ha avaluat un creixement d’uns 0,8 mm anuals per a formacions d’aquest tipus localitzades en una fondària de 42 m i amb una irradiància corresponent al 0,3% de la superficial.
Les dades sobre el creixement dels organismes suspensívors són molt escasses, però suficients per a revelar la lenta velocitat de canvi de les poblacions. Paramuricea clavata, per exemple, creix al voltant de 15 mm l’any en una població de les illes Medes. El creixement d’altres gorgònies (Eunicella cavolinii, E. singularis, Lophogorgia ceratophyta) és semblant. L’edat d’una gorgònia adulta és, doncs, d’uns 20 anys. Pel que fa al corall vermell, el creixement és també molt lent; hom ha avaluat un creixement en alçada situat al voltant dels 5 mm anuals i un creixement en gruix inferior a 1 mm a l’any.
El coral·ligen és, per tot el que acabem de veure, una màquina complexa, que funciona molt a poc a poc. Els processos són lents i cada organisme, cada estructura, és una cosa molt treballada, que s’ha construït no per atzar, sinó per la intervenció conjunta d’una sèrie de factors, entre els quals hi ha la competència i l’optimització en l’ocupació de l’espai. La impressió que fan els fons coral·lígens de cosa ben feta, de cosa acabada, es pot atribuir, precisament, a aquesta lentitud d’edificació d’una estructura per una sèrie d’organismes molt diferents, que interactuen fortament entre ells.
Ja hem comentat que el component algal té un desenvolupament màxim al final de la primavera i durant l’estiu, quan la irradiància (factor limitant del creixement) és màxima. Es desconeix, però, si dins l’activitat dels animals que formen part del coral·ligen hi ha cicles anuals. La descoberta d’anells de creixement a les gorgònies i al corall vermell fa pensar en una resposta positiva, encara que no és evident que es formin sota la influència de les variacions estacionals. La seva reproducció acostuma a ser estacional i a Paramuricea clavata, per exemple, l’expulsió dels ous es produeix durant l’estiu. El ritme de creixement dels organismes suspensívors podria anar lligat a les diferències estacionals de disponibilitat d’aliment, encara que la irradiància podria tenir, indirectament, alguna repercussió sobre el metabolisme dels suspensívors. Cal també tenir en compte la temperatura, a la qual s’han atribuït moltes de les diferències de caràcter estacional trobades en el creixement de molts organismes, tant terrestres com aquàtics. Però l’escassa variació d’aquest paràmetre en les comunitats d’una certa fondària fa pensar que hi té una importància secundària.
De fet, però, i encara que s’ha demostrat l’existència de ritmes endògens en determinats organismes suspensívors, la impressió de l’escafandrer que visita assíduament un mateix fons coral·ligen és que la variació estacional es manifesta només en petits detalls: una activitat més gran de la fauna vàgil durant l’estiu, la successió de diferents espècies de peixos al llarg de l’any i, en els poblaments ben il·luminats, una cobertura algal més reeixida al final de la primavera. La comunitat per ella mateixa es manté, tanmateix, invariable.
En bona part això és degut, com ja s’ha dit, a les característiques de llarga vida de les espècies animals. Moltes d’aquestes espècies, siguin vàgils o sèssils, suspensívores o carnívores, són bons exemples d’organismes estrategs de la K: constitueixen poblacions no gaire abundants, la mida dels individus sol ser considerable, inverteixen més en estructura que en biomassa fàcilment utilitzable pels altres nivells tròfics, dediquen una part important del metabolisme a produir descendents ben protegits (que en gran proporció tenen desenvolupament directe, sense passar per una fase planctònica) i a ocupar l’espai, estratègia en la qual, com ja hem explicat, han excel·lit al llarg de l’evolució.
Les coves submergides
Les coves i els túnels submarins exerceixen una atracció fascinadora sobre l’escafandrer esportiu. El descobriment inesperat de qualsevol cavitat submergida provoca un sentiment mixt d’alegria i angoixa que l’impulsa a endinsar-se en un món fosc i desconegut. Aquesta curiositat es veu, gairebé sempre, àmpliament recompensada. Ja a l’entrada, sota la llum del focus, apareix una decoració bigarrada, dominada pels colors clars i pastels, però amb precioses pinzellades d’un vermell intens corresponents a les colònies del preuat corall ver. A mesura que ens aventurem cap als indrets més foscos, la decoració s’empobreix estèticament i la roca queda parcialment al descobert. Sovint, del sostre pengen estalactites, reveladores de l’origen aeri de moltes d’aquestes cavitats, que formen autèntiques columnes que hi donen una vistositat escenogràfica extraordinària. Els eixams de petits misidacis semblants a minúscules gambetes ens envolten, però fugen espaordits dels raigs de llum del nostre focus. És un ambient totalment inhòspit però increïblement bell, on el temps sembla haver-se aturat per sempre i on l’únic soroll és el de la nostra respiració i la del nostre company. Si apaguem el focus, el sentiment de soledat, d’abandó, és absolut. Podem estar-nos-hi quinze segons, potser trenta, però no més: ens supera l’angoixa de trobar-nos envoltats de roca per tot arreu, aïllats de la reconfortant visió de la superfície blava i segura de la mar oberta. Encenem de nou el focus i desfem el camí que havíem recorregut. Primer les esponges, després el corall i una altra vegada la llum i els fons oberts: les gorgònies, les algues, els peixos. En definitiva, un món diferent, el nostre món submarí.
Malgrat que constitueixen un espai inapreciable en el conjunt del bentos marí, les coves submarines són d’un interès extraordinari pel que fa als seus pobladors i als processos que s’hi desenvolupen. Tenim al nostre abast un exemple d’un món submergit sense llum, generalment inaccessible per a l’home si no és mitjançant la utilització de costosos enginys. L’ambient tan peculiar de les coves, la seva reclusió i la presència gairebé sempre d’una única entrada i, alhora, sortida, donen un interès ecològic addicional a aquests sistemes. En conjunt, actuen com una capsa fosca consumidora de matèria i energia que arriba tota de l’exterior, atès que els vegetals hi són exclosos per la manca de llum. Dins d’aquesta capsa, una maquinària complicada (encara no totalment entesa pels científics i formada amb peces de museu, interconnectades de vegades de manera maldestra o insòlita) hi funciona lentament, especialment adaptada a un indret on no es pot anar de pressa.
La característica ambiental comuna a totes les coves és la manca de llum. Al llarg d’una cova hi ha un gradient de llum molt accentuat de tal manera que en un recorregut horitzontal de pocs metres l’atenuació de la llum és molt superior a la que es produeix en l’eix vertical en la mar lliure. La màxima variació es dona, doncs, en l’eix horitzontal, la qual cosa és una excepció al que passa regularment en les comunitats aquàtiques. En les coves més profundes l’extinció de la llum arriba a ser total. L’altre factor d’importància és la circulació de l’aigua a través de la cova. La seva renovació és molt important, ja que tota la vida de la cova depèn de les entrades de material extern. L’aigua aporta als organismes de la cova l’oxigen i l’aliment necessaris per a subsistir, i n’elimina el diòxid de carboni i els productes excretats. També serveix com a mitjà dispersador de les diàspores emeses pels seus habitants. En general, la circulació de l’aigua és més reduïda com a més fondària es trobi la cova i més extensa i complicada sigui. Òbviament, també hi ha un gradient en la intensitat de circulació de l’aigua, que decreix de les zones externes a les internes. Aquesta disminució porta associats un augment de la sedimentació i una disponibilitat d’oxigen i aliment més petita.
La temperatura també en resulta afectada. A les zones més internes acostuma a ser inferior a la dels trams més externs, encara que durant els mesos d’hivern pugui ser una mica més alta. L’estabilitat tèrmica és, en tot cas, superior a la dels fons oberts. Finalment, la salinitat és un nou factor que cal tenir en compte. Íntimament associades als massissos càrstics, les coves solen tenir filtracions d’aigua dolça a les parts terminals, per la qual cosa es produeix un gradient de salinitat creixent de l’interior a l’exterior. La gran estabilitat de l’aigua impedeix la barreja i es formen dues capes d’aigua ben diferenciades, la superficial totalment dolça i la més profunda de característiques marines. En algunes coves, com la del Pont d’en Gil, a Menorca, la picnoclina és molt ben delimitada i les masses d’aigua allotgen poblaments ben diferents. No són rars els petits llacs d’aigua dolça en contacte amb cambres d’aire. En alguna d’aquestes coves, especialment a les de les Balears, hi trobà encara refugi la foca mediterrània o vell marí (Monachus monachus) quan la pressió de l’home la va foragitar dels seus llocs habituals de descans i cria, les cales solitàries. Ara no trobarem foques mediterrànies enlloc dels Països Catalans.
Esquema d’una secció longitudinal del túnel llarg de la Meda petita (Baix Empordà), amb la distribució a les parets i al sostre del recobriment (en percentatge) d’esponges, cnidaris i altres grups (briozous, poliquets, etc.). Observeu la dominància de les esponges a les parets i dels cnidaris als sostres.
Jordi Corbera, original de M.A. Bibiloni i altres, modificat.
Tots aquests gradients, llevat del de la llum, que és bidireccional, no existeixen en els túnels. Els túnels submarins, com el Túnel Llarg de la Meda Xica (Costa Brava), són recorreguts per forts corrents i les poblacions que apareixen en les seves parets reflecteixen aquesta peculiaritat.
La distribució dels organismes cavernícoles és un fidel reflex d’aquests gradients físics a què hem al·ludit. Hom observa un accentuat gradient biòtic des de les parets externes fins a les internes. Les espècies canvien, i també ho fan, en sentit decreixent, els paràmetres reveladors de l’estructura de les comunitats, com la biomassa, el recobriment o el nombre d’espècies. La fauna s’emprobreix qualitativament i quantitativament. Els organismes adaptats a l’ambient de les coves més profundes estan altament especialitzats i de vegades són veritables relíquies de grups taxonòmics antics, pràcticament extingits, que han quedat reclosos en aquests ambients perquè són els únics llocs on la poca competència els ha permès de sobreviure.
El poblament de les coves prové bàsicament de certs organismes de la fauna coral·lígena que han aconseguit superar el sever filtre ecològic que comporten les particulars condicions ambientals; els organismes preadaptats a aquesta situació són els criptòfils, és a dir, els que viuen als forats, intersticis i anfractuositats dels blocs coral·lígens, sotmesos a una irradiància i una circulació de l’aigua molt reduïdes. Un segon component, molt menys important, prové de la fauna batial; aquestes espècies, adaptades a una vida a les fosques i a un aliment escàs, troben dins les coves un ambient propici per al seu desenvolupament. Finalment, ja hem comentat la importància de les coves com a indrets especialment favorables per a la conservació d’espècies relictes. La manca de productors primaris, l’absència de llum i l’estrès alimentari apropa les comunitats cavernícoles a les de l’estatge batial. Però el fort lligam entre la seva constitució faunística i la del coral·ligen fa que, bionòmicament, siguin molt més properes a aquest tipus de formacions. De fet, les coves són un indret especial, un ambient pont que fa de connexió entre els poblaments coral·lígens i les comunitats de la zona afital, i de vegades els poblaments d’aigua dolça de les coves continentals (especialment dels massissos càrstics).
Cada cova té unes peculiaritats que la individualitzen: no hi ha dues coves de formes i condicions ambientals idèntiques. Això fa que el poblament de cada cova sigui gairebé propi, i això, a més, s’accentua perquè, per a alguns grups animals (però no tots), les coves són veritables illes biològiques pel que fa a llur colonització i a l’establiment de les faunes. El seu interès faunístic és, per això, considerable.
Valors de biomassa (a dalt), en g de pes sec/m2, i grams de matèria orgànica (a l’esquerra) dels principals grups d’animals bentònics en les parets d’una cova, i aspecte dels diferents poblaments que caracteritzen cada transsecte. Els organismes dominants són, de l’exterior a l’interior, cnidaris, esponges i poliquets. Noteu, també, l’empobriment qualitatiu i quantitatiu dels poblaments, causat per l’estrès tròfic i la manca de fixació larval (el nombre d’espècies de cada grup s’assenyala dins dels histogrames).
Jordi Corbera, original de J.M. Gili i altres, i de J.G. Harmelin i altres.
Deixant de banda les característiques de cada cova, hom observa una sèrie de particularitats generals en el seu poblament. La manca de llum provoca l’absència d’algues, i aquesta la d’herbívors. La densitat de depredadors macròfags és relativament baixa a causa de l’escassetat d’aliment. Els detritívors són abundants i els suspensívors resulten especialment afavorits en els túnels o els llocs amb un cert intercanvi d’aigua. Les esponges són el grup dominant a la fauna cavernícola, llevat de les zones més recòndites, on els poliquets tubícoles constitueixen, en termes de biomassa i recobriment, els organismes més abundants. Antozous i briozous hi són també ben representats, i, juntament amb els depredadors de tots aquests grups sèssils (crustacis, mol·luscs i verms brostejadors i peixos), completen la llista dels principals pobladors de les coves.
Les diferències faunístiques varien també segons la posició del substrat: El sòl de les coves és un ambient poc propici per a la vida dels suspensívors, ja que la sedimentació interfereix en els seus sistemes d’alimentació. Només alguns antozous i poliquets hi estan adaptats; els sedimentívors hi constitueixen la fauna dominant. Les parets i el sostre de les coves no tenen, generalment, una sedimentació excessiva, i l’estratègia suspensívora hi és molt ben representada. Les esponges són el grup dominant a les parets, i els cnidaris, principalment els madreporaris, són especialment abundants en els sostres.
La variació específica i estructural de les comunitats de les coves en l’eix horitzontal és contínua, però es diferencien clarament els poblaments de l’entrada dels del fons. Hom parla de dos tipus bàsics de comunitats: el de les coves semifosques i el de les coves fosques.
La comunitat de les coves semifosques
La comunitat de coves semifosques es presenta, en algun dels seus aspectes, en gairebé qualsevol zona costanera rocosa. Apareix no solament a l’entrada de les coves i els túnels sinó també a les cavitats, a les escletxes i els forats prou grans, als extraploms i a les baumes. La seva estructuració espacial és semblant a la del coral·ligen, però se’n diferencia per la manca d’un estrat elevat de gorgònies i grans esponges i per una concreció reduïda a causa de la manca del component algal. La fauna intersticial és, per això, molt més pobra. Els estrats intermedi i incrustant estan ben desenvolupats i hi pertanyen les espècies principals. Les esponges constitueixen aproximadament el 50% de la biomassa i el recobriment total. Les espècies típiques del coral·ligen en l’aspecte més esciòfil són aquí dominants: Oscarella lobularis, Petrosia ficiformis, Chondrosia reniformis, Agelas oroides, Axinella vaceleti, A. damicornis. Spongia virgultosa, Clathrina clathrus; a les quals s’afegeixen Spirastrella cunctatrix, Erylus euastrum, Remera fulva, Haliclona mediterranea i Verongia cavernicola, entre altres. La fauna d’antozous és especialment rica i hi destaquen Leptopsammia pruvoti, Parazoanthus axinellae, Corallium rubrum, Hoplangia durotrix, Rolandia rosea i Caryophyllia inornata. Els hidraris són representats bàsicament per les grans colònies d’Eudendrium racemosum i Halecium beani. Els briozous són també abundants (diverses Sertella, Adeonella calveti, Crassimarginatella maderensis, Prenantia inerma, Schizotheca serratimargo, Porella cervicornis, Cribrilaria radiata, etc.), i els tunicats tenen alguna espècie comuna a les coves (Pyura dura, Cystodytes dellechiajei, Didemnum maculosum). Diversos poliquets (Serpula vermicularis, Pomatoceros triqueter, Spirobranchus polytrema), mol·luscs (Barbatia barbata, Àcar pulchella, Rocellaria dubia i altres bivalves), el cirrípede Balanus perforatus i el foraminífer Miniacina miniacea contribueixen també al poblament sèssil de les coves semifosques. Les espècies endolítiques són representades pel mol·lusc Lithophaga lithophaga i l’esponja Cliona smithii. La fauna vàgil és constituïda per diversos poliquets (espècies de Ceratonereis, Glycera, Lumbrinereis); els mol·luscs Calliostoma zizyphinus, Luria lurida i diversos nudibranquis; crustacis com l’esclop (Scyllarus arctus), la cabrella (Maja verrucosa), el cranc pelfut (Dromia vulgaris), etc.; i estrelles de mar (Marthasterias glacialis, Coscinasteriastenuispina) i ofiures (Ophiocomina nigra, Ophioderma longicauda). Algunes espècies de peixos es refugien a les coves tot i que també s’estenen per altres indrets. El moll reial (Apogon imberbis) i Thorogobius ephippiatus són, però, força exclusius d’aquest ambient.
La comunitat de les coves fosques
A les coves fosques la fauna està molt empobrida. El recobriment no és mai total i el color fosc de les parets, incrustades d’òxids metàl·lics, és la nota dominant. La simplificació estructural és enorme, amb un sol estrat d’organismes irregularment dispersos per les parets. Els tubs blancs dels serpúlids (Omphalopoma aculeata, diversos Vermiliopsis) constitueixen sovint la major part de la fauna, sobretot en els llocs més inaccessibles. Les esponges de les coves semifosques creixen també en aquesta comunitat, però ho fan anormalment, amb formes de creixement estranyes i despigmentades. Algunes espècies són, però, pròpies d’aquests indrets, com ara Rhabderemia minutula, Dendroxea lenis, Diplastrella bistellata i els fòssils vivents Merlia normani, Pectoroninia hindei i Petrobiona massiliana. Els cnidaris i els briozous són representats per espècies amb pòlips i colònies petites, com ara Guynia annulata, Polycyathus muellerae i Madracis pharensis entre els antozous, i Celleporina lucida, Setosellina cavernicola i Crassimarginatella crassimarginata entre els briozous. De l’escassa fauna vàgil podem destacar els crustacis Stenopus scaber i Herbstia condylata, els opistobranquis Berthella aurantiaca i Susania testudinaria, l’ofiura Ophiopsila aranea i un peix cec, Grammonus ater. Autèntics núvols de misidacis (diferents espècies d’Hemimysis, cadascuna pròpia d’una cova o sistema de coves diferent) són moradors constants del fons de les coves durant el dia; a la nit surten de la cova per alimentar-se a les comunitats bentòniques exteriors, molt més productives. L’entrada diària de material extern que comporta la migració nictemeral d’aquests misidacis és, sens dubte, molt important en el balanç tròfic de la cova.
Les comunitats de les coves es mantenen constants durant tot l’any: l’existència de cicles anuals passa desapercebuda als poblaments cavernícoles. D’altra banda, hom no disposa d’estudis sobre el creixement de les espècies en aquests indrets, però tot fa pensar que és extraordinàriament lent. La introducció experimental de substrats verges a les coves ha demostrat que la colonització és pràcticament inexistent, per la qual cosa hom pensa que la pobresa faunística de la comunitat de les coves fosques és causada en part per la manca de fixació larval. És probable que la responsable d’aquesta impossibilitat de les larves per a fixar-s’hi sigui la manca de llum. Cal pensar que les coves són una situació atípica en el bentos marí: les baixes irradiàncies, en general, van associades a grans fondàries i, per tant, a una baixa disponibilitat d’aliment. És possible, doncs, que al llarg de l’evolució el comportament larval hagi escollit la llum com a indicadora de la riquesa tròfica d’un sistema. Aquesta, i no la mateixa escassetat alimentària, seria la causa de l’empobriment faunístic fins i tot a les coves on la velocitat de renovació de l’aigua és elevada. D’altra banda, l’estrès tròfic sembla també un element selectiu d’importància que provoca un creixement anòmal o la mort de la majoria d’organismes no adaptats especialment a la vida de les coves, la qual cosa n’explicaria la composició faunística insòlita.
Els substrats durs profunds
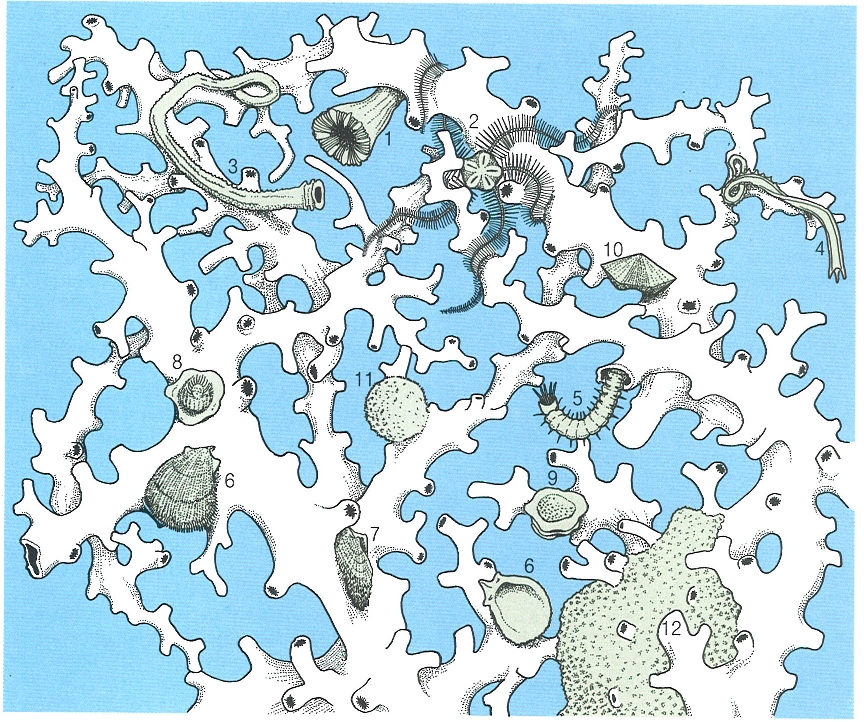
Sobre el corall blanc Madrepora oculata, que creix sobre els escassos substrats rocosos de l’estatge batial, hi viu una munió de petits organismes, filtradors i carnívors, dels quals hem representat en el dibuix l’antozou Caryophyllia smithii (1), l’ofiura Ophiothrix lutkeni (2), els poliquets Vermiliopsis (3), Placostegus tridentatus (4) i Eunice floridana (5), els bivalves Spondylus gussoni (6) i Barbatia scabra (7), els briozous Disporella hispida (8) i Plagioecia inoedificata (9), el braquiòpode Crania anomala (10) i les esponges Hamacantha johnsoni (11) i Tylodesma inornata (12).
Jordi Corbera.
La plataforma continental de les costes que voregen la Mediterrània no és gaire ampla, i generalment acaba vers els 120-150 m de fondària, en una zona de pendent més o menys acusat, travessada per grans canyons submarins o recs, com els de la costa catalana. L’inici del talús continental no sempre coincideix batimètricament amb el final de la zona fital, a causa de les grans diferències de transparència de l’aigua als diferents indrets dels Països Catalans. D’altra banda, el límit inferior de les algues no és una frontera clara i ben definida; les crostes d’algunes algues calcàries escassament desenvolupades, com també algunes diatomees adaptades a baixes irradiàncies o els filaments endobiòtics de la clorofícia Ostreobium, arriben a trobar-se a fondàries considerables. En aquestes condicions la concreció algal és nul·la i, com passava a les coves, el poblament animal és relativament abundant. A mesura que va augmentant la profunditat, els fons durs són més escassos, per la qual cosa on hi ha un aflorament rocós enmig dels fons sedimentaris s’hi amuntega una gran varietat d’espècies animals, en epibiosis múltiples, resultat d’utilitzar com a substrat qualsevol suport, inorgànic o orgànic. Com a conseqüència, els poblaments centrats en aquests afloraments són veritables oasis d’una certa riquesa, en comparació del «desert» que formen els fons tous.
La fauna de les roques situades a la part inferior de la plataforma continental i la fauna de les coves semifosques tenen una semblança inequívoca, com ja hem comentat. El component algal incrustant és minoritari o totalment absent i hi ha un nucli important d’espècies comunes, com el corall vermell o moltes esponges (diverses Axinella, Petrosia ficiformis, Ircinia muscarum, I. oros, Acanthella acuta, etc.). Tot i això, la mida dels individus és molt superior: Axinella polypoides, per exemple, pot assolir un metre d’alçada, i les colònies de corall vermell (que en aquestes grans profunditats creixen erectes sobre el fons i no a redós dels extraploms foscos, com passa a poca fondària) tenen unes dimensions insòlites. D’altra banda, hi apareixen nombrosos organismes que no trobàvem a les coves i que aquí atenyen un desenvolupament espectacular; es tracta, en concret, dels coralls negres (Antipathes subpinnata i Parantipathes larix), que formen grans colònies arborescents, i d’un madreporari colonial d’enormes pòlips grocs, Dendrophyllia cornigera. Diverses esponges grosses i de colors clars són també pròpies d’aquests poblaments (Phakellia ventilabrum, P. robusta, Polymastia mamillaris, Raspailia viminalis, Reniera mucosa, Stelletta grubei, S. hispida, Mycale syrinx, M. truncata). Diversos briozous que ja trobàvem al coral·ligen dels gèneres Hornera, Sertella, Porella) i els braquiòpodes Terebratulina caputserpentis i Gryphus vitreus són components habituals d’aquests poblaments. Els poliquets també hi són ben representats (Serpula vermicularis, Omphalopomopsis fimbriata).
La fauna vàgil hi és força abundant. Hi destaquen la tabaquera (Cidaris cidaris), una garota (Echinus melo) l’euriàlid Astrospartus mediterraneus i l’ofiura Ophiacantha setosa, entre els equinoderms; diversos crustacis, com la llagosta salamenya (Palinurus mauritanicus) i el cabrot de fonera (Paromola cuvieri); i, entre els peixos, els serrans penegals (Helicolenus dactylopterus). La dominància de l’estratègia suspensívora en aquests poblaments i el gran desenvolupament que assoleixen les esponges i alguns cnidaris posen de manifest una gran riquesa en matèria orgànica particulada i una notable circulació de l’aigua. La proximitat de les costes i, per tant, de les àrees de producció afavoreixen, sens dubte, aquestes espècies. Moltes les podem trobar a l’Atlàntic, a fondàries menors; això, i el fet que no es trobin a les coves o els túnels situats a l’abast de l’escafandrer, fa pensar que la temperatura i l’aliment són factors determinants de la seva exclusivitat a fondàries situades per sota del límit estival de la termoclina.
A partir dels 250 m de fondària la fauna s’empobreix; el corall vermell, el corall negre i les grans esponges desapareixen progressivament a causa, probablement, de la manca d’aliment i de la reducció creixent de substrats durs. La minva de l’hidrodinamisme i l’augment de la distància a la costa fan que la disponibilitat tròfica hi sigui molt disminuïda. Els poblaments, que ja són exclusivament animals, canvien, i els rars substrats rocosos estan recoberts per dos madreporaris colonials, els coralls blancs Madrepora oculata i Lophelia pertusa. Associats amb ells hi ha altres madreporaris solitaris (Desmophyllum cristagalli, Caryophyllia smithii), gorgònies (Isidella elongata) i corall negre (Antipathes fragilis). Les esponges (Amacantha, Medesmia i Microciona) incrusten les colònies de madrèpora, i els poliquets, força comuns, hi són representats per espècies d’àmplia distribució, com ara Eunice floridana, juntament amb tàxons característics de la fauna abissal (Placostegus tridentatus, Lagischa drachi, per exemple). Alguns bivalves, com la caixeta (Arca nodulosa) i l’ostra vermella (Spondylus gussoni), crustacis (el cirrípede Verruca i la gamba Pandalina) i equínids (Cidaris) constitueixen la resta de la fauna que hom hi pot trobar. L’espècie Coralliophaga lithophagella és un bivalve molt comú que viu dins les colònies de Madrepora.
El límit inferior d’aquesta comunitat se situa cap als 1000 m de fondària, sempre que el pendent sigui prou inclinat per a permetre l’aflorament de promontoris rocosos. Actualment, en molts indrets, les colònies de corall blanc (Madrepora i Lophelia) són mortes i només en sobreviuen les parts apicals; la resta és recoberta per incrustacions negroses de limonita i òxids de ferro i manganès. Les bases del corall i la roca acostumen a estar enterrades en el fang, i més avall hom observa un sediment cimentat d’origen organogen, ric en espècies subfòssils, pròpies d’aigües més càlides i actualment extingides a la Mediterrània. Llur desaparició s’hauria esdevingut ara fa uns 40 000 anys; això dona una idea de l’activitat bioconstructora extraordinàriament lenta d’aquests coralls ahermatípics. Aquesta activitat és, doncs, molt inferior a la del coral·ligen o les cornises de Lithophyllum tortuosum i, naturalment, encara més que la dels seus parents hermatípics, amb zooxantel·les simbiòtiques, formadores dels esculls coral·lins.
Els fons tous
Caladors de la zona de costa compresa entre Calella (Maresme) i Tossa de Mar (Selva), on feinegen, principalment, els bous del port de Blanes. És remarcable la gran extensió dels fons tous per dessota els 50 m. Alguns d’aquests caladors reben el nom de l’espècie o espècies que hi són més abundants: sot de la Gamba (per aquest decàpode), les Garotes (per Spatangus purpureus), la Forbiana (per Leptometra phalangium), els Capets (pels peixos de la família dels tríglids) i les Bruixes (pels peixos de la família dels bòtids).
Carto-Tec / Institut de Ciències del Mar, Barcelona.
La diversitat d’espècies, de formes, de colors, de moviment, és molt inferior en un fons tou que en un de dur. Però els fons tous són molt productius per a l’home; efectivament, les pesqueries de ròssec actuen sobre aquests fons (les xarxes es trencarien si treballaven sobre fons rocosos) i n’extreuen moltes espècies de peixos demersals i bentònics, com també de crustacis, mol·luscs i altres espècies comercials. En la plataforma continental dels Països Catalans no hi ha pràcticament cap àrea de sorra o fang que no constitueixi un calador, més o menys productiu, conegut des d’antic i explotat per les flotes pesqueres dels ports més propers.
Els fons sorrencs i fangosos estan desproveïts, però, de l’animació frenètica que caracteritza les zones rocoses. Sovint, sobretot a poca fondària, semblen immensos deserts a l’observador submarí. Aquesta primera impressió és, però, errònia. La vida en aquests ambients és intensa encara que no tingui l’espectacularitat i la diversitat de la que es desenvolupa sobre substrat dur. La mobilitat del substrat condiciona l’absència d’organismes sèssils i redueix enormement les possibilitats de crear estructures complexes. Però la mateixa naturalesa tova del substrat permet l’existència d’organismes que s’hi enterren, bo i excavant-hi galeries (l’estratègia perforadora és seguida per poques espècies en els substrats durs), o que viuen entre els grans de sorra, constituint el que podríem anomenar un «plàncton» intersticial (estratègia desconeguda en el fons durs).
Algunes espècies disposen de mecanismes per a fixar-se en aquests ambients i estabilitzar el substrat, amb la qual cosa s’obté un tipus de fons equiparable als durs. Ja hem parlat extensament de les dues situacions més típiques: l’alguer de posidònia i el coral·ligen de plataforma. Ací tractarem d’aquells indrets que no tenen un substrat estable biogènic.
Els factors que determinen el poblament
Els factors que determinen les condicions ambientals i els organismes que viuen en un fons sedimentari són diversos.

Característiques granulomètriques del sediment marí, importants per als organismes.
Dades dels autors
Potser el més important és la mida de les partícules que constitueixen el sediment. La porositat d’un substrat tou depèn de la mida mitjana dels grans de sediment que el constitueixen; aquesta és una característica d’una gran importància perquè determina el contingut i la renovació de l’aigua (i dels materials o gasos que porta) dintre del sediment, com també l’espai disponible per als organismes que hi viuen. La mida de les partícules és determinada bàsicament per l’hidrodinamisme i per la distància del lloc a la costa i als al·luvions fluvials. Els indrets amb un moviment intens de l’aigua no permeten la sedimentació de les partícules fines, i el sediment és gruixut; en canvi, si l’hidrodinamisme és escàs, les partícules fines poden sedimentar-se i el gra és petit. Hi ha una correlació molt forta entre la mida del gra i el contingut de matèria orgànica i l’oxigenació del sediment, factors d’una importància capital perquè determinen la capacitat productiva d’aquests tipus de fons. Els sediments de gra gruixut estan, en general, ben oxigenats i són pobres de matèria orgànica i bacteris, i rics en fauna intersticial (i en flora, a profunditats adients); mentre que els sediments de gra fi, al contrari, són rics en matèria orgànica, sovint anòxics i mancats de fauna intersticial.
L’oxigenació està també molt lligada a l’època de l’any i a la fondària: la concentració d’oxigen disminueix a l’estiu i a mesura que ens endinsem en el sediment. El quimisme de l’aigua intersticial es modifica, d’aquesta manera, en augmentar la fondària. La anòxia fa que hi hagi un potencial d’oxidoreducció negatiu i provoca la reducció de determinats elements, com ara el ferro, i també dels nitrats o els sulfats. A més, no permet la vida d’organismes amb respiració aeròbia, de tal manera que només poden viure-hi els organismes de respiració anaeròbia, com diversos ciliats i bacteris. L’hidrodinamisme és també molt important, ja que posa a disposició dels organismes suspensívors la matèria orgànica particulada en suspensió i determina, com hem vist, la mida de gra del sediment. Els factors catastròfics d’origen natural, com són les tempestes, provoquen modificacions importants en l’estructura interna del sediment i afecten, directament o indirectament, el component viu.
Distribució amb la fondària d’algunes característiques químiques d’un sediment fi amb una capa externa oxidada i una capa interna reduïda.
Carto-Tec, original de T. Fenchel i R. Riedl.
Altres factors que determinen poc o molt les característiques de les comunitats de fons tou són la llum (necessària per a l’existència de productors primaris), les característiques de l’aigua situada per sobre del sediment (el contingut de nutrients, la temperatura i la salinitat, aquest darrer factor en zones estuarianes) i, finalment, els factors biòtics i antròpics. La competència i la depredació en determinen, com passa en les comunitats bentòniques de substrat dur, la composició faunística i florística.
La incidència de l’acció de l’home sobre els fons tous no és gaire diferent de la que es dona en els fons durs. Com passa també en d’altres situacions al bentos marí, aquesta acció pot afectar molt l’estructura i la composició de les comunitats de fons tous. Però hi ha una activitat humana que se centra precisament sobre aquests fons. Ja hem parlat de la pesca a ròssec; l’acció continuada de les arts sobre el fons equival a llaurar la terra en els ecosistemes continentals: les comunitats s’empobreixen (perden estructura i espècies), però es fan més productives i explotables, perquè les espècies més fràgils, típiques estrategues de la K, són substituïdes per d’altres, estrategues de la r, més robustes, prolífiques i oportunistes. Igual que amb els conreus a terra, on les plantes que l’home explota actualment són el resultat de la selecció agrícola sobre les males herbes dels camps primitius, les espècies de peixos amb què les flotes pesqueres assorteixen les llotges de peix del litoral dels Països Catalans són, cada cop més, les més resistents a la pesca, en detriment d’altres espècies més preuades, però més escasses. En d’altres situacions, l’acumulació de fangs i llots orgànics (procedents del clavegueram de les grans ciutats i de les depuradores litorals, per exemple) sobre un substrat de gra gros, o bé sobre un substrat dur, també empobreix el poblament; fins i tot, en casos extrems, crea extenses àrees azoiques o bé poblades només per una o dues espècies tolerants (com ara el poliquet Capitella capitata).
Les estratègies i les comunitats
Organismes característics de la meiofauna intersticial dels fons sorrencs submarins: 1 rotífers, 2 hidrozous, 3 turbel·laris, 4 gastròtrics, 5 cinorincs, 6, 7, 8 nematodes, 9 poliquets, 10 copèpodes harpacticoides, 11 ostràcodes, 12 tardígrades.
Jordi Corbera, original d’Schlieper i Fincham
D’un primer cop d’ull hom observa que els fons sorrencs i fangosos són molt més homogenis que no els rocosos. Els organismes dominants hi acostumen a ser diferents: les algues dominants són formes unicel·lulars, bàsicament diatomees (si hom exceptua els fons de coletes i els de grapissar), i sovint són els únics productors primaris que hi ha. Cal remarcar la seva baixa biomassa i la seva elevada taxa de renovació, la qual cosa apropa els fons tous al món pelàgic pel que fa a l’estratègia dels organismes fotosintetitzadors. Com que no és possible la creació d’estructures perdurables, la colonització d’aquests fons la fan espècies típicament oportunistes, estrategues de la r, que, així com en els fons durs representen els estadis inicials de la successió vers la reconstitució de les comunitats, sobre els fons tous poden esdevenir una comunitat permanent.
La fauna és més diversificada, però només se’n veu una petita part (epifauna): la majoria d’organismes viuen amagats dins del sediment (endofauna o infauna). Els habitants d’aquests fons també es poden classificar per les seves dimensions: microfauna, meiofauna i macrofauna. La meiofauna, constituïda per organismes que fan aproximadament entre 0,05 i 2 mm, és potser el grup en què hom troba els representants més genuïns dels fons tous: turbelaris, nematodes, arquianèl·lids, quinorrincs, gastrotrics, tardígrades, rotífers, ostràcodes, copèpodes harpacticoides, com també formes aberrants de grups més ubics, com cnidaris i mol·luscs. La microfauna ha estat molt poc estudiada, malgrat la seva importància ponderal i metabòlica; hi dominen sobretot els protozous, especialment els ciliats. La macrofauna, per la seva part, és constituïda per mol·luscs, poliquets, crustacis, equinoderms, algunes esponges i cnidaris i diversos peixos.
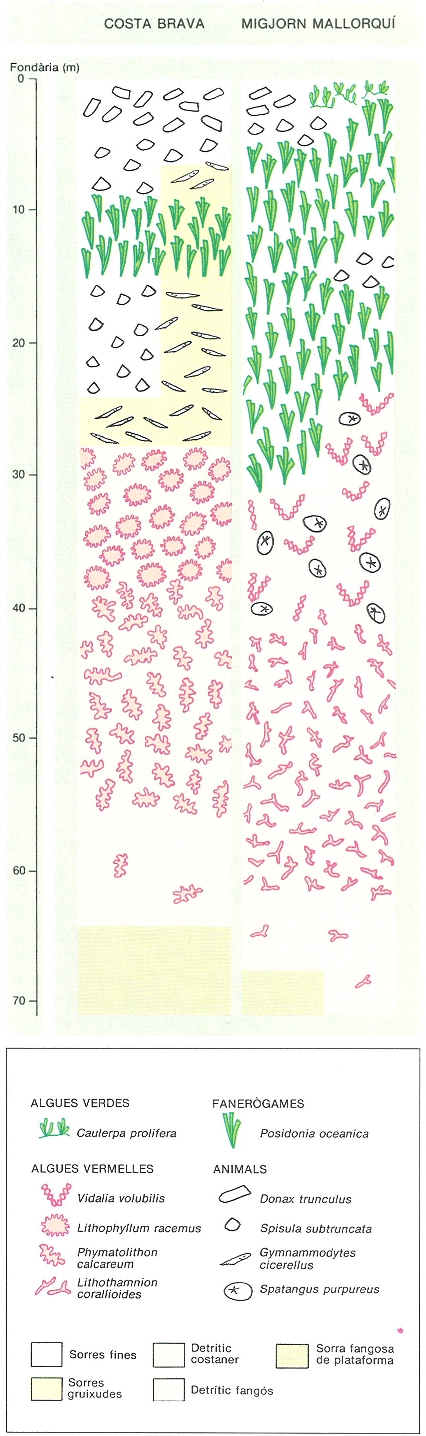
Models de zonació sobre fons tous en dos indrets diferents dels Països Catalans. Aquests models s’alteren quan predomina el fang sobre la sorra, cosa que impedeix l’aparició de fons de grapissar ben constituïts; la pesca al ròssec, tot i estar prohibida per sobre dels 50 m, es practica en aquests fons i malmet extraordinàriament les comunitats. A Mallorca són remarcables la raresa de sorres gruixudes, l’amplitud batimètrica dels alguers de posidònia (Posidonia oceanica), la presència de fons detrítics dominats per l’herba torta (Vidalia volubilis) i la constitució una mica diferent dels fons de grapissar.
Jordi Corbera, amb dades dels autors
Són dues les estratègies bàsiques que segueixen els animals en aquests tipus de fons: la dels suspensívors i la dels sedimentívors. Aquests darrers són característics dels fons tous i poden ser catalogats com a detritívors especialment adaptats a aquests ambients. Els sedimentívors mengen la matèria orgànica particulada que hi ha en el sediment i, principalment, digereixen els bacteris que la descomponen. Diversos poliquets, les holotúries, les garotes irregulars i alguns bivalves són representatius d’aquesta categoria. Els suspensívors són representats per alguns poliquets tubícoles, un gran nombre de bivalves i algun gasteròpode i, amb menys importància, per cnidaris i esponges. Els pocs herbívors que hi ha s’alimenten del microfitobentos i, generalment, es tracta d’organismes pertanyents a la microfauna i a la meiofauna. Els carnívors són relativament poc abundants, i exploten la resta de grups; hi ha mol·luscs, crustacis, equinoderms asteroïdeus i peixos.
La dominància de sedimentívors o suspensívors en un indret determinat és indicadora de les característiques del sediment. Els sediments fins afavoreixen els sedimentívors ja que el seu contingut de matèria orgànica és alt i, per tant, les poblacions bacterianes de les quals s’alimenten són més nombroses. L’activitat dels sedimentívors inestabilitza el sediment, la qual cosa no afavoreix la fixació de les larves dels organismes suspensívors i n’obtura els mecanismes de filtració. Els suspensívors, en canvi, dominen en els sediments de gra més gruixut, situats en àrees de corrents més forts. En aquests llocs, la quantitat de matèria orgànica en el sediment és més petita i, per tant, els sedimentívors hi tenen menys menjar. Pel fet que el substrat és més estable, els suspensívors no tenen problemes per a establir-s’hi, i mitjançant el filtratge continu d’aigua impedeixen la introducció de larves de sedimentívors. Aquestes són, naturalment, dues situacions extremes; en la realitat es donen situacions intermèdies, i una determinada extensió de fons és ocupada per clapes on dominen, respectivament, suspensívors o sedimentívors, la qual cosa introdueix una certa heterogeneïtat en uns fons bastant homogenis.
Tipus de fons que hom pot distingir a la costa del Principat segons la macrofauna que acompanya les captures de les embarcacions de pesca al ròssec (allò que els pescadors anomenen «brutícia»). Noteu la dominància del fang per sota la isòbata dels 100 m.
Carto-Tec, original de J.M. Gili i altres
Els organismes que viuen dins un mateix sediment competeixen per l’espai. Els suspensívors han de tenir els seus sistemes de captació a la columna d’aigua i, si l’aliment és suficient, la manera de poder mantenir poblacions més grans d’individus és distribuir-los verticalment dins del substrat. Paral·lelament, els organismes sedimentívors poden menjar a qualsevol nivell del sediment sempre que hi hagi prou matèria orgànica i oxigen per a satisfer les seves necessitats. L’acomodació de les espècies en els diferents nivells maximitza l’eficàcia en l’explotació dels recursos. El resultat és una estructuració vertical en la distribució de les espècies dins del substrat. Un segon eix de variació és la fondària, a la qual estan íntimament lligats diversos factors d’influència directa per a la vida dels organismes (llum, temperatura, disponibilitat d’aliment) o indirecta, com a determinants de les característiques granulomètriques del sediment (hidrodinamisme). Finalment, hi ha una heterogeneïtat horitzontal deguda a les característiques particulars de cada indret, que es reflecteixen bàsicament en el tipus de sediment que hi ha.
Les comunitats que trobem a les costes dels Països Catalans evidencien les diferents condicions ambientals, i podem distingir-les segons la mida de gra del sediment i l’hidrodinamisme (factors que es presenten intrínsecament lligats), i segons la fondària. En la relació que exposem a continuació hem obviat una descripció minuciosa de les comunitats i, com hem fet abans amb els fons durs, hem preferit una descripció més global.
Les comunitats de les platges infralitorals
A les zones més superficials de la zona infralitoral, a les platges de còdols de mida mitjana moguts constantment per l’onatge, i on la matèria orgànica retinguda pel pedruscall no és gaire important, hi trobem amfípodes (Allorchestes aquilinus, Melitta hergensis), turbel·laris, nemertins i peixos que característicament viuen enganxats a la cara inferior dels còdols mitjançant ventoses ventrals (són els aixafa-roques, Gouania wildenowii i Lepadogaster gouanii), com també algun decàpode molt actiu (Porcellana bluteli). Quan els còdols són substituïts per graves de gra gros, netes, els pobladors típics són encara més escassos; hi destaquen un nemertí (Lineus lacteus) i un arquianèlid (Saccocirrus papillocercus).
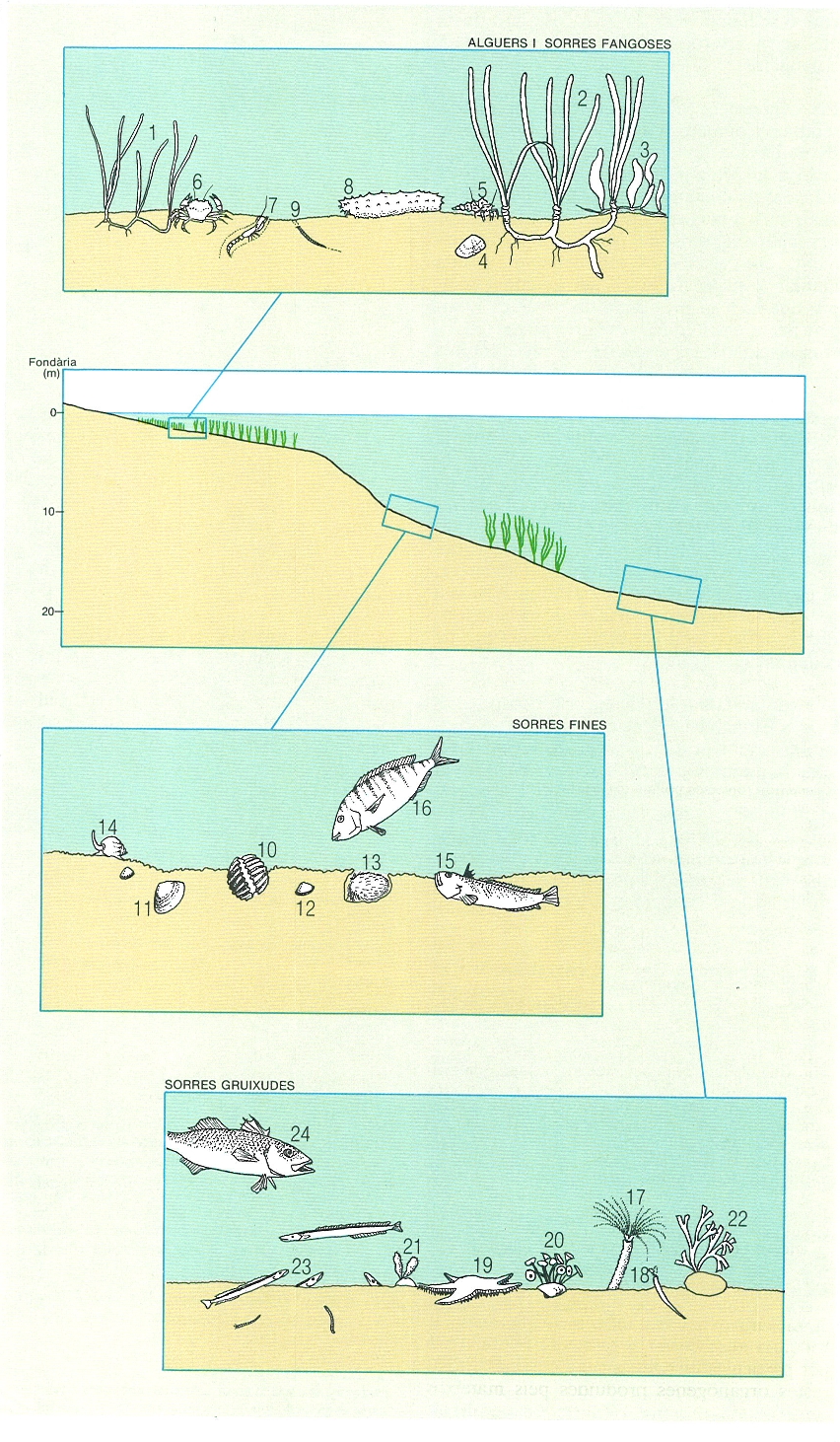
Zonació i aspecte de les comunitats sobre fons tou de l’estatge infralitoral. A les parts més encalmades i menys fondes es desenvolupen alguers de Zostera noltii (1). En llocs amb un hidrodinamisme més intens i sorres de gra més gruixut hi creix Cymodocea nodosa (2), acompanyada sovint de Caulerpa prolifera (3). Comuns en aquests fons hi ha els crancs de sorra (Carcinus mediterraneus, 6), el decàpode Upogebia pusilla (7), pixotes (Holothuria tubulosa, 8), cloïsses (Tapes decussatus, 4) i poliquets (9). Les conquilles de gasteròpodes morts (Gourmya) són ocupades pel bernat ermità Clibanarius erythropus (5). A una fondària superior, enterrades a la sorra fina, hom hi troba escopinyes (Cardium tuberculatum, 10), tellerines (Donax, 11), Spisula subtruncata (12) i l’eriçó de sorra Echinocardium cordatum (13). El gasteròpode Sphaeronassa mutabilis (14), les aranyes (Trachinus vipera, 15) i les mabres (Pagellus mormyrus, 16) són comuns en aquests fons. En llocs de sorra gruixuda apareixen el poliquet Sabella penicillum (17), l’escafòpode Antalis tarentina (18) i l’estrella Astropecten aurantiacus (19). Les conquilles mortes estan ocupades per algues (Acetabularia acetabulum, Dasycladus vermicularis, Dictyota dichotoma, 20, 21, 22). Característics d’aquests fons són els sonsos (Gymnammodytes cicerellus, 23), peixets molt preuats per depredadors com el llobarro (Dicentrarchus labrax, 24).
Jordi Corbera, amb dades facilitades pels autors.
A les platges de sorra relativament gruixuda, si l’hidrodinamisme és moderat o elevat, es desenvolupa un poblament dominat per diverses tellerines (Donax trunculus, D. multistriatus, Macoma tenuis, Lentidium mediterraneum), l’equínid Echinocardium mediterraneum i diverses espècies d’isòpodes i poliquets. Si l’hidrodinamisme és escàs, s’acumulen sorres fangoses favorables al desenvolupament d’alguers de Caulerpa prolifera, Cymodocea nodosa i Zostera noltii. La presència d’una cobertura vegetal més o menys desenvolupada no afecta gaire el poblament animal, el qual és constituït per algunes holotúries (Holothuria tubulosa, H. polii) i mol·luscs del gènere Gourmya (G. vulgata, G. rupestris). Les closques buides d’aquests i d’altres gasteròpodes són ocupades per un bernat ermità de petites dimensions (Clibanarius erythropus). També hi és present el cranc de sorra (Portumnus latipes). Les cloïsses (Tapes decussatus) són habitants típics d’aquests ambients. En aigües salabroses s’hi afegeixen, encara, molts altres bivalves, entre els quals hi ha les escopinyes (Cardium lamarckii) i altres espècies de cloïsses (Abra ovata, Scrobicularia plana). A les zones fortament pol·luïdes la fauna és molt poc diversificada i hi dominen denses poblacions dels poliquets Capitella capitata, Magelona papillicornis, Scolelepis fuliginosa i algun altre, tots especialment resistents a la contaminació orgànica.
A fondàries superiors, entre 3 i 30 m aproximadament, la composició específica de les zones sorrenques varia segons la dimensió dels grans de sorra. La comunitat més estesa correspon a la dominada per diverses espècies de bivalves: les escopinyes (Cardium tuberculatum), les xirles (Mactra corallina), les rossellones (Chamaelea gallina), diverses tellerines (Donax, Tellina) i, sobretot, Spisula subtruncata, que caracteritza la comunitat. Els gasteròpodes hi són també presents (diverses espècies de Nassa, Neverita josephiniae, Murex trunculus), i també uns quants poliquets i crustacis, i l’equínid Echinocardium cordatum.
Si la sorra és gruixuda o es tracta d’un fons de graves de gra petit, les espècies que hi viuen són unes altres. Aquest sediment acostuma a tenir un origen organogen, resultat del trencament i la degradació dels esquelets calcaris dels organismes que viuen sobre substrat dur o a l’alguer de posidònia. També trobem aquest tipus de sediment en les costes granítiques amb una aportació terrígena important. Els bivalves són representats pel petxinot (Glycimeris glycimeris) i diverses tellerines (Donax variegatus, Tellina crassa). Són també característics d’aquest problament l’ofiura Ophiopsila annulosa, l’escafòpode Antalis tarentinum, diversos poliquets i crustacis, el cefalocordat Branchiostoma lanceolatum (que ha donat nom a aquest sediment: sorres d’Amphioxus) i els sonsos (Gymnammodytes cicerellus). A partir d’uns 20 m de fondària, en llocs amb un cert corrent, es desenvolupa l’alga coral·linàcia lliure Lithophyllum racemus, la qual, juntament amb les restes dels esquelets carbonatats, serveix de fixació per a nombroses espècies d’algues, principalment Arthrocladia villosa, Sporochnus pedunculatus i Vidalia volubilis. L’equínid Spatangus purpureus i l’escopinya Circomphalus casinus poden ser localment abundants. D’altres espècies que podem trobar són la gran estrella de mar Astropecten aurantiacus i la bonica actínia Condylactis aurantiaca.
Aquests fons tous infralitorals constitueixen l’hàbitat d’un gran nombre de peixos, que s’alimenten principalment dels bivalves i dels poliquets, molt abundants en aquest tipus de fons. Com a espècies més comunes, encara que no puguin ser considerades com a característiques, poden esmentar les lluernes (Trigla), alguns gòbits (Gobius), dragons (Callionymus), aranyes (Trachinus), rogers (Mullus surmuletus), rajades (Raja clavata), calets (Pagellus acarne), pagells (P. erythrinus), mabres (P. mormyrus), pagres (Pagrus pagrus) i unes quantes espècies de peixos plans (bòtids i solèids). Gran part de la pesca artesanal de palangre i xarxa es fa en aquest tipus de fons, i moltes d’aquestes espècies són habituals en les subhastes dels pobles costaners.
A mesura que ens allunyem de la costa, la fondària augmenta i el sediment és constituït per partícules terrígenes cada cop més fines i restes organògenes produïdes pels mateixos organismes bentònics. En termes generals, els elements terrígens se sedimenten més lluny de la costa com més fins són. La mida mitjana de les partícules hauria de decréixer, doncs, en anar cap enfora. No obstant això, aquesta condició no es compleix a causa de la irregular distribució de les entrades d’aigua continental provinents dels rius. A més, cal pensar que des del Pliocè fins ara hi ha hagut nombrosos canvis eustàtics, segons els períodes de transgressió i regressió del nivell de la mar. (Hom creu que aquests canvis del nivell de la mar poden haver estat els causants de les extincions de bona part de les espècies que avui dia trobem només com a restes subfòssils al llarg de la plataforma continental mediterrània.) Per això, un fons situat actualment a 100 m de fondària pot haver estat no fa gaire temps (en termes geològics) a només 20 m i haver rebut sediments de gra gruixut. Amb les transgressions posteriors, aquest sediment pot haver estat exposat a condicions molt variables, atesa la intensitat dels dipòsits terrígens provinents dels rius i la intensitat dels corrents de la zona. Segons aquests tres factors, el factor històric, la deposició actual de sediments i l’hidrodinamisme, hom troba diversos tipus de sediments caracteritzats per poblaments força diferents. A les zones amb entrada de dipòsits terrígens considerables (delta de l’Ebre, per exemple) els fons són constituïts per sorres fines i fangs, mentre que a llocs sotmesos a forts corrents (Balears, Costa Brava), els fons tous són formats principalment per material detrític o sediment de gra gruixut. En una primera aproximació, hom pot diferenciar quatre grans tipus de comunitats que sovint se situen en bandes paral·leles al llarg de la costa; de les més costaneres a les més allunyades, són: la del substrat detrític costaner, la del detrític fangós, la de la sorra fangosa i la del detrític profund de final de plataforma.
Les comunitats dels fons detrítics costaners
Mapa bionòmic detallat de la plataforma continental situada entre el cap de Creus i el cap de Begur. Les comunitats instal·lades sobre sorres es limiten a la zona més propera a la costa, sense ultrapassar gairebé els 50 m de fondària. El detrític apareix a continuació (llevat de davant de la badia de Roses). Els fons fangosos (sota la forma de diferents fàcies o variants) ocupen la resta del fons, excepte una ampla taca de detrític situada entre els 100 i els 200 m de fondària davant el golf de Roses. L’explicació d’aquesta distribució una mica aleatòria cal cercar-la en l’existència de diferències en l’hidrodínamisme, l’aportació actual de partícules exògenes, els organismes vivents actualment i els factors històrics.
Jordi Corbera, original de D. Desbruyères i altres.
Les comunitats dels fons detrítics costaners sovint anomenades genèricament detrític costaner, substitueixen la comunitat d’Spisula subtruncata o la comunitat de sorres gruixudes amb Spatangus purpureus a partir dels 35-40 m de fondària, i, a les zones amb escassa deposició de material fi, s’estenen fins al límit de la plataforma continental, on es confonen amb el detrític profund; si el dipòsit de material fi és important, és substituït pel detrític fangós i les sorres fangoses.
El sediment detrític costaner consisteix en una barreja de graves organògenes dels poblaments actuals o subrecents, juntament amb una quantitat més o menys elevada de sorra. Aquests fons acostumen a ser dominats per algues carbonatades de vida lliure, a les quals ja hem fet referència en parlar de les comunitats algals de fons tous. La formació de sediments organògens per aquestes algues és important i, si l’estabilitat és prou gran, el fons detrític pot esdevenir un coral·ligen de plataforma. A causa de l’escassa entrada de sediments fins i de la relativa estabilitat del fons, el recobriment de l’epibentos animal i vegetal és elevat. Entre les espècies epibentòniques més destacables hi ha un elevat nombre de rodofícies carbonatades (dels gèneres Phymatolithon, Peyssonnelia i Lithothamnium), algues toves (Dictyota dichotoma, Cryptonemia tunaeformis, Vidalia volubilis, Phyllophora crispa, etc.), algunes esponges (Actinoe fictitius, Raspaciona aculeata, Eurypon clavatum, Dysidea fragilis), cnidaris (Lophogorgia ceratophyta, Eunicella singularis, Veretillum cynomorium, Epizoanthus arenaceus), poliquets (Hyalinoecia fauveli), bivalves (Chlamys varia, Pecten jacobaeus, Lima loscombei), gasteròpodes (Turritella communis, Murex trunculus), equínids (Sphaerechinus granularis), estrelles de mar (Astropecten irregularis, Luidia ciliaris) i ascidis (Phallusia mammillata, Diazona violacea, Rhopalaea neapolitana). Diverses espècies de bernat ermità (Paguristes oculatus, Dardanus arrosor, Pagurus) ocupen les closques buides dels mol·luscs, recobertes per anèmones (Calliactis parasitica, Adamsia palliata) i esponges (Suberites domuncula). L’endofauna és també abundant i és representada per diversos bivalves (Laevicardium oblongum, Moerella donacina, Circomphalus casinus), petites holotúries (Cucumaria), ofiures (Ophiopsila aranea, Ophiocomina nigra) i garotes de sorra (Spatangus purpureus, Echinocyamus pusillus).

Aspecte d’un fons de sorra fangosa situat a la plataforma continental, a uns 125 m de fondària. Hom hi distingeix una galera (Squilla mantis, 1), el cranc Liocarcinus depurator (2), la ploma de mar (Pennatula phosphorea, 3), l’ofiura Ophiura texturata (4) i el gasteròpode Lunatia fusca (5). En segon terme hi ha mans de mort (Alcyonium palmatum, 6), la garota Echinus melo (7), l’esponja Axinella verrucosa (8) i l’estrella Astropecten irregularis (9).
Jordi Corbera, cedit pel Servei de Medi Ambient, Diputació de Barcelona.
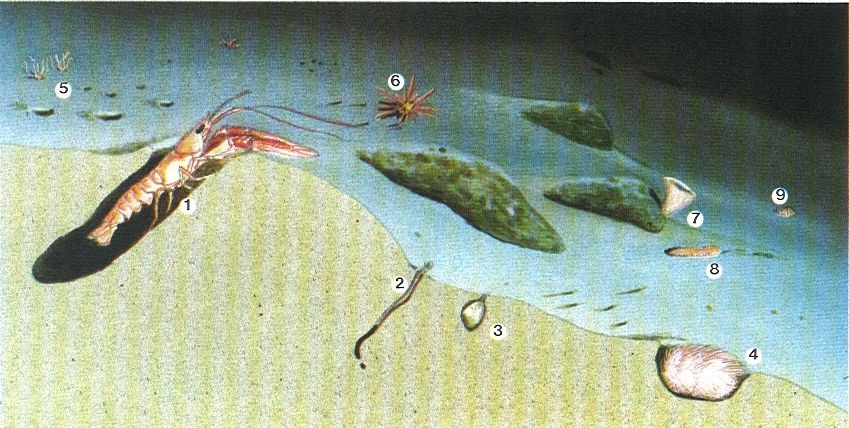
Fons de fang de talús continental, a uns 200 m de fondària. Enterrats al fang hi ha un escamarlà (Nephrops norvegicus, 1), el cuc Hyalinoecia fauveli (2), el bivalve Nucula sulcata (3) i l’equínid Brissopsis lyrifera (4). En segon terme apareixen Amphiura filiformis (5), una tabaquera, Cidaris cidaris (6), l’esponja Poecillastra compressa (7), l’espardenya Stichopus regalis (8) i el cargol Cassidaria echinophora (9).
Jordi Corbera, cedit pel Servei de Medi Ambient, Diputació de Barcelona.
Si el dipòsit de material fi és considerable, el sediment és constituït per una barreja de partícules gruixudes i fines, amb una proporció variable; el resultat és una sorra fangosa amb trossets de petxines i grava. Els organismes sèssils de l’epifauna no assoleixen el desenvolupament dels dels fons detrítics costaners. Els organismes sedimentívors són més abundants, però encara hi ha força filtradors, com ara l’esponja Raspailia viminalis, l’antozou Alcyonium palmatum i diversos ascidis (Phallusia mammillata, Microcosmus vulgaris, M. savignyi, Polycarpa mamillaris); les ofiures hi són abundants: Ophiothrix quinquemaculata, Amphiura chiajei i O. filiformis, com també els poliquets Aphrodite aculeata, Nephtys hombergi, Euphantalis kinbergi i altres.
En determinats indrets on el fons és substituït totalment per fang o sorra fangosa, sense material detrític apreciable, la vida dels petits suspensívors no està gens afavorida, i hi dominen els sedimentívors, principalment diverses espècies de poliquets i les holotúries Oerstergrenia digitata i Stichopus regalis. Les plomes de mar (Virgularia mirabilis, Pteroides griseum i Pennatula phosphorea) són característiques d’aquests fons, on es fixen mitjançant el seu llarg peu; hi són també abundants els gasteròpodes Scaphander lignarius i Lunatia fusca, les estrelles Astropecten irregularis i Anseropoda placenta, la galera (Squilla mantis) i l’ofiura Ophiura texturata.
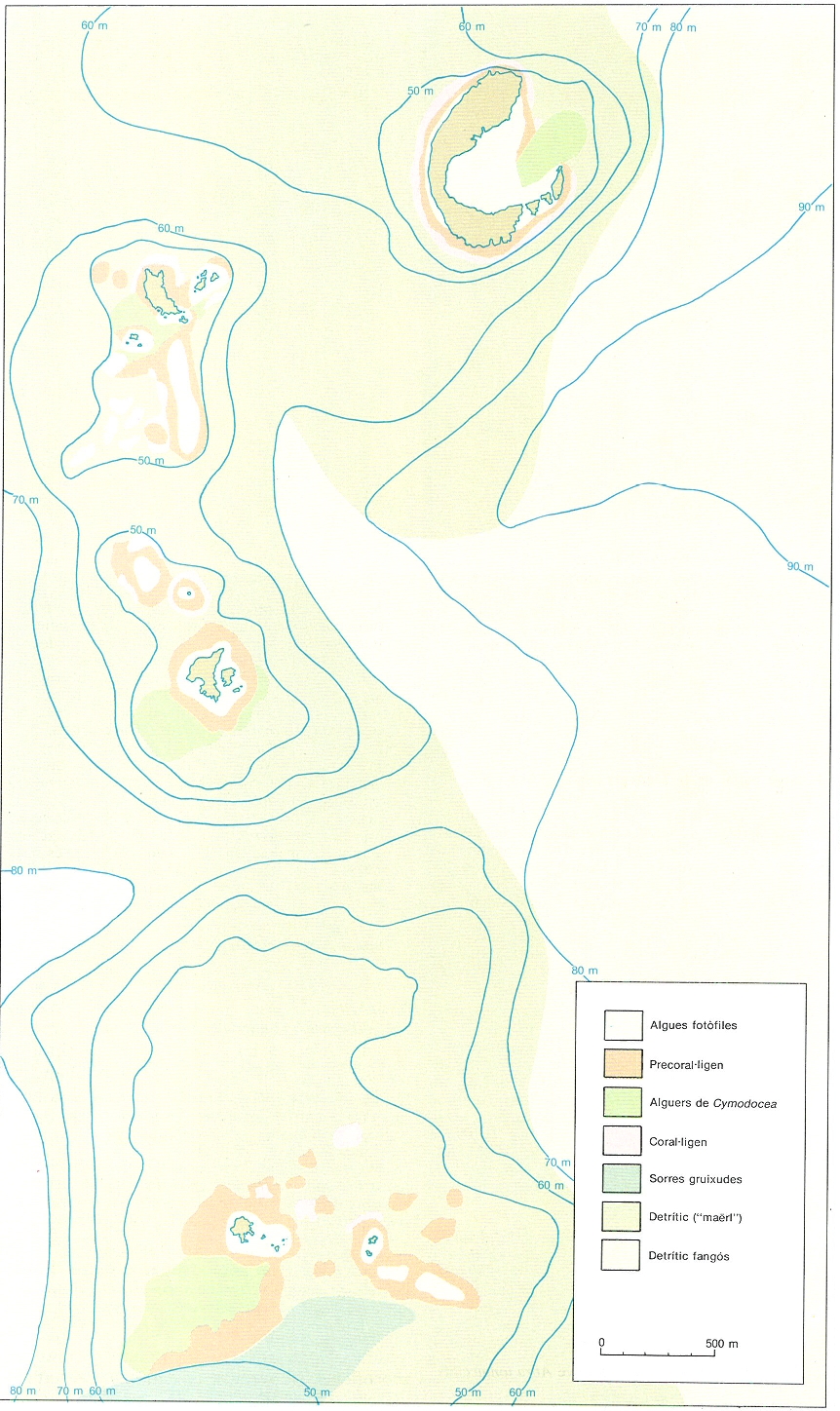
Els Columbrets, arxipèlag d’origen volcànic situat unes 35 milles mar enfora de la costa castellonenca, s’aixequen sobre un fons de naturalesa detrítica, ocupat principalment per bancs de grapissar («maërl»). L’absència de posidònia és un fenomen totalment atípic, si tenim en compte que es tracta d’unes illes mediterrànies; la raó cal cercar-la en la reduïda extensió dels fons sorrencs infralitorals i en la manca històrica de colonització per aquesta espècie.
Jordi Corbera, original d’AM. Garcia Carrascosa.
A les àrees amb un dipòsit de material terrigen molt escàs i allunyades de la costa, ja vora el límit de la plataforma continental, el sediment és constituït bàsicament per graves amb sorra gruixuda i restes organògenes, majoritàriament fòssils o subfòssils. A les fondàries on es desenvolupen aquests fons detrítics (90-150 m), les algues ja són poc abundants o inexistents. L’espècie dominant és la forbiana (Leptometra phalangium), crinoïdeu que aprofita els forts corrents per a obtenir la matèria orgànica particulada de l’aigua mitjançant els seus llargs braços. Altres espècies força abundants són els hidraris Lytocarpia myriophyllum i Nemertesia antennina, els bivalves Circomphalus casinus, Chlamys clavata i Astarte sulcala, l’escafòpode Dentalium panormum, els equínids Echinus acutus, Neolampas rostellata i Spatangus purpureus, les holotúries Thyone gadeana i Neocucumis marioni i diverses ofiures.
Els peixos de la plataforma i el talús continental, encara que menys abundants que a l’Atlàntic, han de ser tinguts en compte, a causa també de la seva importància econòmica. Les principals espècies comercials, pescades amb barques de ròssec que operen aproximadament entre els 50 i els 500 m de profunditat, són el lluç (Merluccius merluccius), la mòllera pigada (Phycis blennoides), la maire (Micromesistius poutassou) i la mòllera (Trisopterus minutus capelanus). Altres espècies menys importants, però molt apreciades, són els rogers o molls (Mullus), els llenguados (Solea), els raps (Lophius), el gall (Zeus faber) i diversos espàrids (Pagellus, Pagrus).
Les comunitats de l’estatge batial

Aspecte d’un fons batial, a uns 400 m de fondària. Sobre el fang, hom hi observa el pennatulaci Virgularia mirabilis (1), la gorgònia Isidella elongata (2), la gamba Aristeus antennatus (3) i el poliquet Aphrodite pallida (4). Sobre la roca apareixen l’antozou Dendrophyllia cornigera (5), el braquiòpode Gryphus vitreus (6) i el crustaci Munida perarmata (7).
Jordi Corbera, cedit pel Servei de Medi Ambient, Diputació de Barcelona.
L’estatge batial de la Mediterrània occidental és constituït, llevat de determinats afloraments rocosos amb poblaments de madrèpora (Madrepora oculata), per un sediment fi, fangós, bastant compacte. El poblament d’aquest fons és relativament pobre si el comparem amb els que trobem a l’Atlàntic, i hi ha diverses raons que ho justifiquen. La primera és la dificultat que tenen les espècies batials atlàntiques de penetrar a la Mediterrània, tant en el passat com en el present, a causa de l’escassa fondària de l’estret de Gibraltar i dels forts corrents d’aigua mediterrània que flueix cap a l’Atlàntic pel fons; no ha estat possible, doncs, un repoblament total després que la dessecació de la Mediterrània, a la fi del Miocè, eliminés gairebé del tot la fauna profunda. En segon lloc, la temperatura de l’estatge batial de la Mediterrània es manté en uns 13°C, temperatura molt elevada per a les espècies batíais atlàntiques. Finalment, l’oligotròfia dificulta enormement la subsistència de diverses espècies a causa de l’escassetat d’aliment.
La llista d’espècies que viuen a l’estatge batial és llarga; hom hi pot diferenciar les espècies característiques d’aquests poblaments de les que tenen una repartició més àmplia. Entre les primeres cal esmentar diverses esponges, com ara Thenea muricata, Rhizaxinella pyrifera, Poecillastra compressa i Craniella cranium, les quals es presenten a fondàries superiors als 200 m, o Trichostemma sarsi i Hyalonema thomsoni, confinades a fondàries encara més grans; els cnidaris Funiculina quadrangularis, Virgularia mirabilis i Isidella elongata; diversos poliquets (Aphrodite pallida, Harmothoe impar, per exemple); un elevat nombre de crustacis, entre els quals hi ha els explotats comercialment, com les gambes blanca (Parapenaeus longirostris), rosada (Aristeus antennatus) i vermella (Aristeomorpha foliacea); el bivalve Abra longicallus; dues espècies d’escafòpodes, Dentalium agile i Siphonodentalium quinquangulare; el pogonòfor Siboglinum carpinei; el braquiòpode Gryphus vitreus; diversos cefalòpodes (Sepietta oweniana, Rossia caroli) i estrelles de mar (Odontaster mediterraneus, Brissingella coronata).
Entre les espècies presents a l’estatge batial que també es trobaven als fons fangosos de l’estatge circalitoral podem destacar, per la seva abundància, les plomes de mar (Veretillum cynomorium i Pennatula phosphorea); els escamarlans (Nephrops norvegicus); els bernats ermitans Eupagurus prideauxi i Dardanus arrosor; l’ofiura Amphiura chiajei; les estrelles de mar Astropecten irregularis i Anseropoda placenta; els equínids Echinus acutus, Brissopsis lyrifera i Cidaris cidaris; l’holotúria Oerstergrenia digitata, l’espardenya (Stichopus regalis), molt preuada pels «gourmets», el gasteròpode Cassidaria echinophora i l’ascidi Diazona violacea. La fauna de peixos d’aquests fons és escassa; hom pot esmentar Chimaera monstrosa, Bathypterois mediterraneus i dues espècies del gènere Chalinura. La meiofauna, molt poc estudiada, és molt abundant i, a fondàries superiors als 2000 m, en zones gairebé desproveïdes de macrofauna, hom n’ha comptabilitzat una densitat mitjana de 5 individus/cm2 de fons.
Dinàmica i funcionalitat dels fons tous
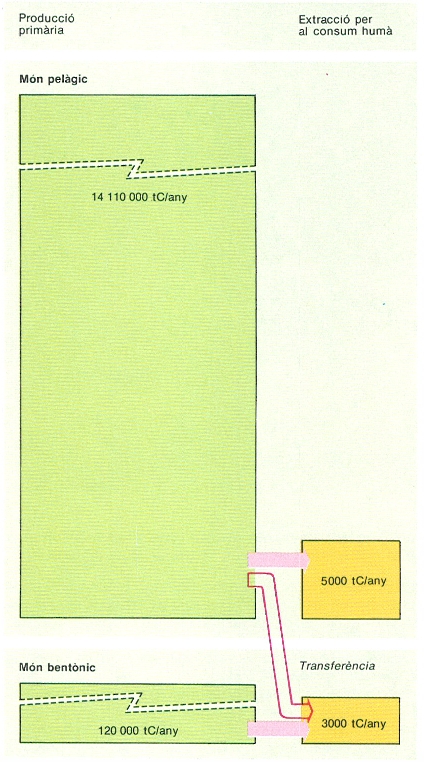
A la mediterrània nord-occidental adjacent als Països Catalans, l’extracció d’organismes del bentos per al consum humà és, amb relació a la producció primària, setanta vegades superior a la d’organismes pelàgics. Una causa és l’eficàcia superior de les pesqueries de peixos bentònics (conseqüència simplement de la seva concentració), i una altra, ecològicament més important, és la transferència d’energia del món pelàgic al món bentònic, que fa augmentar extraordinàriament la matèria aprofitable per les darreres (i explotables) baules de la xarxa tròfica.
Román Montull, amb dades dels autors.
Des d’un punt de vista quantitatiu, els fons tous han estat escassament estudiats a la Mediterrània. Se sap, però, que la biomassa vegetal és molt baixa a la zona infralitoral, llevat dels casos en què hi ha poblaments d’algues o de fanerògames marines. Els herbeis de coleta (Caulerpa prolifera) de la badia de Pollença, a Mallorca, tenen una fitomassa d’uns 300 g (pes sec)/m2, mentre que els de cimodocea (Cymodocea nodosa) i zostera (Zostera noltii) del delta de l’Ebre tenen una fitomassa que oscil·la entre 150 i 550 g/m2. La producció d’aquestes comunitats no és gaire elevada, i es pot estimar en uns 100 g C/m2, ·any per als herbeis de coleta i en uns 200 g/m2·any per als de cimodocea. La producció de les diatomees bentòniques que es desenvolupen en fons oberts desproveïts de clorofícies o fanerògames marines ha estat estimada en uns 20 g C/m2·any a les badies del delta de l’Ebre. Els grans valors de producció obtinguts a irradiàncies de gairebé l’1% de la irradiància superficial fan pensar que la contribució del microfitobentos a la producció de les zones sorrenques i fangoses pot ser considerable, fins i tot a la part superior de la zona circalitoral. D’altra banda, la producció de les comunitats algals amb Lithophyllum racemus, pròpies de les sorres gruixudes de la part inferior de l’estatge infralitoral, i la producció del grapissar («maërl»), ja en ple estatge circalitoral, han de ser també d’un ordre de magnitud semblant a l’obtingut a les comunitats coral·lígenes; valors de 10 a 15 g C/m2·any han de ser habituals en poblaments vegetals d’aquestes característiques.
La biomassa de la fauna oscil·la generalment entre 2 i 15 g (pes sec descalcificat)/m2, amb poques variacions entre la zona infralitoral i la circalitoral. Els principals contribuents a aquesta xifra són els poliquets, els mol·luscs, els equinoderms, els crustacis i els ascidis, per aquest ordre. Hom observa una abundància relativa més gran de mol·luscs a poca fondària i un predomini de poliquets i equinoderms a les zones més profundes. No hi ha, pràcticament, estimacions de producció de la fauna, encara que hom observa grans oscil·lacions dels valors de biomassa i del nombre d’individus de les poblacions al llarg de l’any; així, per exemple, en un poblament d’Spisula subtruncata de la zona infralitoral, la biomassa oscil·la entre 0,4 (novembre) i 7,5 (març) g (pes sec descalcificat)/m2, per la qual cosa pot estimar-se una producció mínima d’uns 3 g C/m2·any.
D’altra banda, l’extracció dels peixos bentònics o lligats alimentàriament al bentos (demersals) pels pescadors mediterranis equival, aproximadament, a uns 10 mg C/m2·any, és a dir, poc més del 30% del total de la producció pesquera. Així, doncs, i malgrat que la producció primària del bentos sigui només de gairebé una centèsima part de la producció planctònica, la producció bentònica utilitzada pel consum humà és d’un terç del total. Això ens recorda que el bentos, tot i ser un sistema amb una explotació més eficient des del punt de vista pesquer, és altament deficitari, que explota el sistema pelàgic, tant d’una manera activa com passiva, capturant les partícules en suspensió. El paper dels organismes suspensívors i sedimentívors com a introductors d’energia en el sistema bentònic és, doncs, importantíssim, ja que a través d’ells entra una quantitat d’energia molt més gran que a través dels productors primaris pròpiament bentònics. El bentos depèn absolutament del món pelàgic des del punt de vista tròfic (recordem també aquí que la majoria d’animals bentònics tenen larves pelàgiques, que passen com a components del plàncton períodes més o menys llargs, alimentant-se d’altres plànctons), la qual cosa fa que aquests dos sistemes siguin totalment indissociables per a aconseguir una idea global del funcionament de l’ecosistema marí.