Concepte de rèptil

Els rèptils són un grup de vertebrats molt més ben adaptats a la vida a la terra que no els amfibis. Contribueix a aquesta capacitat una de les seves característiques més sobresortints: la possessió d’escates còrnies que recobreixen tot el cos i formen una mena d’esquelet extern. Una altra de les seves característiques és l’estructura de l’esquelet, que permet un tipus de moviment especial, la reptació, i dóna fins i tot una gran capacitat grimpadora a algunes espècies, com la serp blanca (Elaphe scalaris), que veiem a la fotografia.
Xavier Parellada.
Els rèptils són vertebrats tetràpodes, en els quals la pell és revestida d’una capa gruixuda d’epiteli ceratinitzat, completament seca i pràcticament desproveïda de glàndules. Les escates epidèrmiques còrnies són, en certs casos, sostingudes per plaques òssies dèrmiques subjacents i cauen periòdicament. La muda pot ser de tota la capa còrnia sencera o saltant a trossos. El crani s’articula a la primera vèrtebra, l’atlas, mitjançant un sol còndil occipital. La vida i la reproducció d’aquests animals tenen lloc a terra; en tot cas, secundàriament, tornen a l’aigua els adults, però la reproducció i el desenvolupament són exclusivament terrestres, gràcies a l’aparició d’una nova mena d’ou, l’ou amniota, que presenta vesícula vitel·lina amb reserves nutritives suficients per a un desenvolupament directe i també noves capes o membranes extraembrionàries que eviten la dessecació de l’embrió.
Els rèptils, si hom considera llurs formes fòssils, a més de les actuals, resulten certament difícils de caracteritzar, perquè dos dels caràcters fonamentals que s’observen en els vivents i que representen l’avenç més important a partir del tronc amfibi (el tegument i l’ou amniota), no poden ser verificats en estat fòssil. De les moltes característiques anatòmiques i fisiològiques que diferencien els rèptils de la resta de vertebrats, les dues més notables són precisament el tipus de tegument i el tipus de reproducció, especialment l’ou amniota. En altres aspectes, la progressió respecte dels antecessors amfíbics és més petita, com ocorre amb l’ectotèrmia (encara que la regulació tèrmica no sigui idèntica) o amb el tipus de cor, en què el ventricle és encara únic, si bé comença ja a marcar-se la divisió entre dos ventricles i arriba a ser quasi completa en alguns grups; o en la homodòntia, encara existent en els rèptils, la persistència, de la cloaca, etc.
Les escates epidèrmiques, renovades periòdicament, impermeabilitzen totalment la pell i no permeten ni l’intercanvi de gasos ni l’intercanvi hídric. El rèptil adult esdevé, així, completament independent de l’aigua i, fins i tot, del medi humit, per la qual cosa és adaptat a biòtops notablement àrids i, fins i tot, plenament desèrtics. Evidentment, la impermeabilització tegumentària requereix adaptacions anatomofisiològiques importants: el pulmó ha esdevingut perfectament funcional i imprescindible i cobreix completamente les necessitats d’intercanvi de gasos dels rèptils; al mateix temps, la regulació hídrica efectuada pel ronyó, orientada a retenir un màxim d’aigua (amb l’eliminació d’un tipus d’orina semisòlida, amb àcid úric), és igualment adaptada a hàbitats secs. Però aquestes modificacions no són suficients per a la independència completa de l’aigua per part dels rèptils. El problema de la reproducció, que obligava els amfibis a la reproducció aquàtica o, excepcionalment, en un medi humit específic, ja és, en aquest nou grup, completament resolt. La fecundació ha esdevingut interna en tots els casos i l’òvul presenta gran quantitat de vitel·lus nutritiu, que permet un desenvolupament embrionari directe, dins del propi ou, sense passar per una fase larval de vida lliure; aquest desenvolupament embrionari, normalment efectuat sense atencions dels progenitors, llevat excepcions, pot produir-se a terra ferma, en qualsevol lloc protegit que reuneixi unes condicions de temperatura, sobretot, i d’hurnitat dins uns límits precisos. Això és possible gràcies a l’aparició d’una sèrie de membranes que protegeixen l’embrió en creixement: l’amni, dins el qual s’acumula el líquid àmnic (que és el que protegeix l’embrió i evita la seva dessecació, ja que actua com un medi aquàtic independent), l’al·lantoide, un diverticle de l’intestí posterior, que acumula els residus tòxics de les cèl·lules embrionàries, en forma d’urea dissolta en aigua i, finalment, el còrion envolupant i protector. Els ous amb les reserves nutritives són dipositats encara recoberts d’una closca, coriàcia o ja calcària, que permet l’intercanvi de gasos però alhora contribueix a evitar la pèrdua d’aigua.
Els fòssils que hom considera corresponents als primers rèptils autèntics són difícils de determinar. Ultra assemblar-se als amfibis fòssils contemporanis en l’aspecte morfològic i anatòmic, hom desconeix el moment de l’aparició de l’ou amniota en el curs de la història del grup. De tota manera, és evident que els rèptils s’originen a partir dels amfibis i comencen llur camí al final del Primari, esdevenen dominants durant el Secundari i minven fortament en el pas al Terciari, en què comencen a dominar els mamífers. El grup no era biològicament prou eficaç, en les formes aleshores presents, o no conservava prou plasticitat per a evolucionar en el sentit que demanaven els canvis climàtics produïts, i, en part, fou afectat per l’evolució dels grups endotèrmics que descendien d’ell: els ocells i els mamífers.
Els grups actuals de rèptils
Els rèptils actuals són, com els amfibis, una minsa representació del que foren en el passat. Hom troba representants vivents de cinc ordres, tres dels quals tenen representants als Països Catalans.
El primer ordre és el dels quelonis o testudins, les tortugues, grup molt antic, amb representants terrestres, d’aigua dolça i marins, que reuneix unes 230 espècies. Els quelonis presenten plaques dèrmiques òssies i un recobriment de plaques epidèrmiques còrnies; ambdós tipus d’escates constitueixen una closca dividida en dues parts, dorsal i ventral, articulades o soldades pels flancs i obertes pel davant i pel darrere, per a permetre, respectivament, la sortida del cap i les potes anteriors i la de la cua i les potes posteriors. Encara que rares, hom troba als Països Catalans dues formes terrestres, dues d’aigua dolça i dades escasses de tres espècies marines.
L’ordre dels crocodilians és representat per rèptils lacertiformes amb plaques òssies i escates còrnies, força aquàtics, amb potes curtes i cua musculosa i potent. Anatòmicament i fisiològicament són molt evolucionats, amb paladar secundari, musell allargat i dentició poderosa. L’ordre inclou 21 espècies, exclusivament de climes tropicals i subtropicals.
Els rincocèfals són representats avui només per una espècie, un dels anomenats fòssils vivents, endèmic de certs illots de la costa N de Nova Zelanda, d’aspecte lacertiforme i, des del punt de vista anatòmic, molt primitiu.
Els escatosos inclouen els rèptils d’escata petita i es divideixen en dos ordres, ben representats a les nostres terres. Són els saures, i els ofidis. Els primers són, normalment, lacertiformes, però també, en alguns casos, àpodes, amb obertures auditives externes i parpelles mòbils, símfisi mandibular forta i llengua ben desenvolupada. Inclouen llangardaixos, sargantanes i dragons, i del total de 3000 espècies conegudes unes divuit són autòctones dels Països Catalans, on també n’hi ha algunes d’introduïdes. Els ofidis o serps són un grup d’escatosos completament àpodes, sense estèrnum ni orifici auditiu extern, amb parpelles foses i transparents, símfisi mandibular lligamentosa i llengua fina, bífida i protràctil. Tota llur morfologia i anatomia és relacionada amb l’allargament extremat del cos. Comprenen unes 2700 espècies, dotze de les quals són regularment presents a les terres catalanes, valencianes i balears.
Els models morfològics
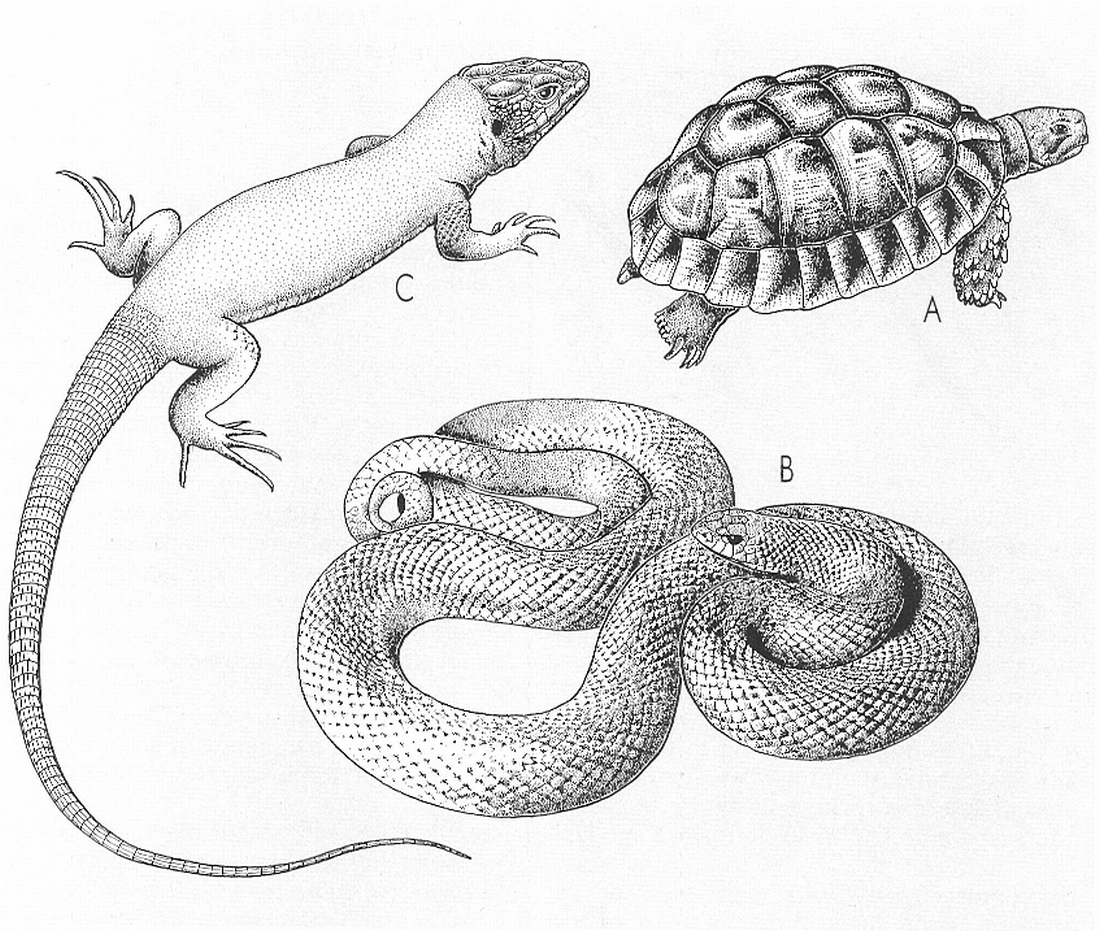
La morfologia dels rèptils respon a tres models bàsics: A tipus queloniforme, amb una peça òssia dorsal i una altra de ventral que constitueixen la closca; B tipus serpentiforme, de cos allargat i àpode, que comparteixen els ofidis i alguns saures; i C tipus lacertiforme, amb el cap, el tronc i la cua ben diferenciats, i les potes més o menys ben desenvolupades, que mostren la majoria dels saures, els rincocèfals i els corcodilians.
Gustavo Hormiga, a partir de fonts diverses.
Els tipus morfològics dels rèptils vivents corresponen als models següents: testudiniforme, lacertiforme i serpentiforme.
El tipus testudiniforme, que presenta exclusivament els quelonis o tortugues, té el cos recobert d’una closca rígida, de forma més o menys ovalada; la peça dorsal (espatllera) és bombada i la peça ventral (plastró) pràcticament plana; ambdues són formades per plaques òssies dèrmiques, recobertes per grosses escates còrnies, unides lateralment per una soldadura rígida o bé mitjançant un lligament. Aquestes dues peces poden, en algunes espècies, arribar a tancar-se com una caixa, que inclou tot l’animal dintre seu. Normalment, però, encara que el cap i les potes anteriors (que surten per la part anterior de la closca) i la cua i les potes posteriors (que surten per la vora posterior) poden retraure’s dins el refugi ofert per la pròpia closca, aquesta no es tanca completament.
El tipus lacertiforme el presenten els saures, els crocodilians i els rincocèfals. El cap i el tronc són ben individualitzats, les quatre potes presenten una llargada pràcticament equivalent i la cua és ben desenvolupada. Les proporcions corporals relatives varien d’un ordre a l’altre, d’una família a una altra i, fins i tot, en espècies del mateix gènere. També apareixen altres modificacions en la forma dels dits, en la folidosi i tipus d’escates, etc. Pot presentar-se una forta reducció de les potes, que acaba amb l’aparició de formes àpodes.
Els rèptils serpentiformes pertanyen, en general, a l’ordre dels ofidis, però es troben també, alguns representants d’aquest tipus a l’ordre dels saures. El seu tronc és molt llarg i generalment esvelt i el cap no és sempre ben individualitzat, la qual cosa vol dir que el coll no és sempre patent. La seva cua és més o menys afilada i mai no presenten potes, encara que en algunes famílies es conserven restes de cintura pelviana. Hi ha patents difèrencies en caràcters tan importants com la presència de parpelles o la disposició de les escates segons si es tracta d’ofidis o de saures àpodes.
Les característiques i les adaptacions anatòmiques dels rèptils
El tegument
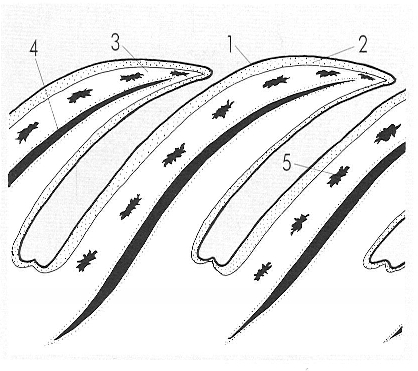
El tegument dels rèptils es caracteritza per la presència d’escates. Normalment es tracta d’escates epidèrmiques juxtaposades o imbricades, però, a més d’aquestes escates còrnies, poden haver-hi escates òssies subjacents. 1 capa còrnia, 2 epidermis, 3 dermis, 4 escata òssia (present només en alguns casos), 5 melanina.
Ferran Rodríguez, a partir de fonts diverses.
La característica principal del tegument dels rèptils és la seva sequedat i impermeabilitat. L’epidermis presenta un estrat corni de cèl·lules ceratinitzades, que es renova periòdicament, a trossos o en una sola peça (muda). La dermis és ben desenvolupada i conté nombrosos cromatòfors. En resulta un cuir excel·lent, que és explotat en la indústria en alguns casos, la qual cosa posa en greu perill determinades poblacions d’ofidis i crocodilians. Les escates epidèrmiques es formen en àrees engruixides del tegument, creixen cap amunt i cap enrera i esdevenen després ceratinitzades; les bases d’aquestes àrees són contínues. Les noves escates es formen sota les antigues, que cauen amb la muda: aquest model de formació correspon als escatosos. En el cas dels crocodilians i dels quelonis, les escates es desenvolupen independentment l’una de l’altra i es renoven gradualment, a conseqüència de llur desgast progressiu.
Les glàndules són pràcticament absents i les que hi ha actuen, normalment, com a factors de protecció (com les glàndules anals de les serps) o tenen secrecions amb funció sexual, com les glàndules femorals dels saures. La pèrdua d’aigua és quasi nul·la.
L’esquelet i la musculatura
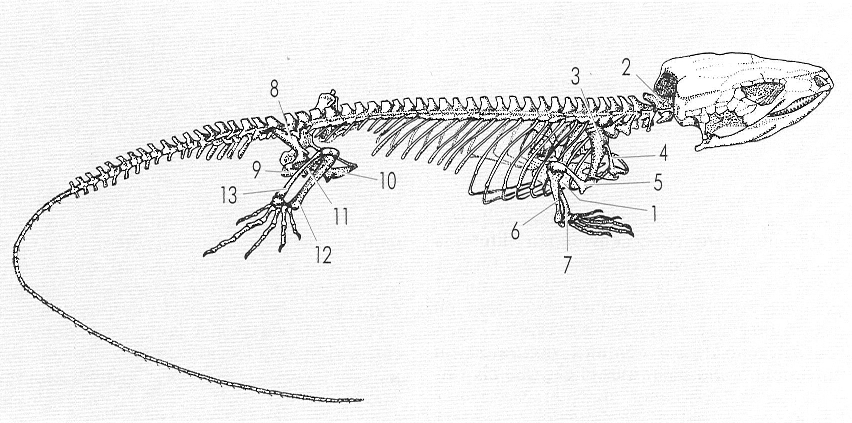
Esquema de l’esquelet reptilià típic, que posa de manifest les modificacions característiques del grup, que concerneixen especialment les cintures i les potes. Cal destacar la presència de costelles ben desenvolupades i de l’estèrnum, que tanca la caixa toràcica per sota. 1 estèrnum, 2 axis, 3 suprascàpula i escàpula, 4 clavícula, 5 húmer, 6 ulna, 7 radi, 8 il·li, 9 isqui, 10 pubis, 11 fèmur, 12 tíbia, 13 fíbula.
Ferran Rodríguez.
L’esquelet dels rèptils ha sofert les modificacions necessàries per a la adaptació a terra ferma, modificacions que ja havien aparegut en els amfibis, però que experimenten en els rèptils una sensible progressió en la direcció d’una major eficàcia locomotora.
El crani no és aplatat, sinó més alt que en els amfibis, amb ossos més densos i, en conjunt, més ossificat. Té només un còndil occipital, cosa que dóna molta mobilitat al cap, i, en els crocodilians, apareix un paladar secundari ossi, que, en separar millor les vies respiratòries de les digestives, permet una eficàcia més gran en el funcionament dels processos respiratori i digestiu.

Les potes dels rèptils representen notables variacions sobre el model bàsic de cinc dits i arriben fins a l’absència completa. En el grup dels saures aquestes modificacions són encara més paleses, com mostren els exemplars il·lustrats: potes reduïdes, pròpies d’animals amb costums excavadors, com la bívia tridàctila (Chalcides chalcides, a dalt), dits amples amb làmines adhesives subdigitals, propis d’animals lapidícoles, com el dragó comú (Tarentola mauritanica, al mig) i potes ben desenvolupades i dits fins, com en la sargantana cuaroja (Acanthodactylus erythrurus, a baix), que es desplaça per sòls sorrencs.
Ramon Torres i Javier Andrada.
La columna vertebral és constituïda per vèrtebres en forma de rodet, amb un arc neural superior que envolta la medul·la nerviosa. Les apòfisis vertebrals són variables, segons la posició de les vèrtebres i els grups de rèptils. Són diferenciades la vèrtebra atlas, que s’articula amb el crani, l’axis, que permet de girarlo, i les vèrtebres cervicals, toràciques, lumbars, sacres i caudals. Les vèrtebres toràciques s’articulen amb les costelles. Els rèptils escatosos àpodes presenten només dues regions vertebrals: una anterior i l’altra posterior a la cloaca; la primera presenta costelles lliures. La columna dels crocodilians és ja molt evolucionada i les seves costelles són molt variades en nombre, estructura i articulació, amb una o dues apòfisis articulables al cos vertebral (diferentment dels amfibis, en què sempre hi ha dues apòfisis costals). En el cas dels quelonis, les vèrtebres es troben fusionades a la closca. L’estèrnum és absent en els quelonis i els ofidis, però es troba en els crocodilians i els saures.
La seva cintura pectoral és ben desenvolupada i és constituïda pel coracoide, el precoracoide i l’escàpula; de vegades, no presenten ni clavícula ni interclavícula. La cintura pelviana és sòlida, i és formada pels ossos ili, isqui i pubis, units mitjançant una símfisi púbica i una símfisi isquiàtica als ossos respectius de cada banda.
L’apèndix locomotor és típicament pentadàctil, amb l’estructura bàsica descrita en parlar dels amfibis: avantbraç, braç i mà de cinc dits, i cuixa, cama i peu de cinc dits. Existeix, però, en alguns grups, una forta tendència a la reducció apendicular, que pot arribar a desaparèixer completament, no sols pel que fa al membre, sinó també a les cintures. La locomoció terrestre és més eficaç i sovint més ràpida que en els amfibis, però encara es realitza per ondulació de la columna vertebral; en els rèptils àpodes també s’efectua aquesta ondulació, però es desenvolupen diferents models peculiars de locomoció reptant.
En els rèptils, la musculatura evoluciona en relació amb l’aparició de les costelles i d’un coll diferenciat. En conjunt, presenten un moviment mandibular més eficaç que els amfibis i també, en comparació, les potes més ben desenvolupades i més actives.
Els sistemes de relació i control
El sistema nerviós
Els hemisferis cerebrals dels rèptils són més ben desenvolupats que els dels amfibis. Apareix, per primera vegada en els vertebrats, el neopal·li, constituït per grups de cèl·lules neuronals agrupades per funcions específiques, de manera que, de fet, és un primer pas envers l’escorça o còrtex cerebral dels vertebrats superiors, que hom pot considerar ja present en els crocodilians.
La medul·la espinal segueix tota la llargada de la columna vertebral, amb eixamplaments cervical i lumbar, que es corresponen als centres d’innervació de les potes, motiu pel qual són absents en els rèptils àpodes, que presenten un plexe lumbosacre, ja que deriven d’animals amb membres desenvolupats. Hi ha dotze parells de nervis cranials, per cefalització dels primers parells de nervis espinals, a partir de l’estructura que és present en els amfibis.
Els òrgans sensorials
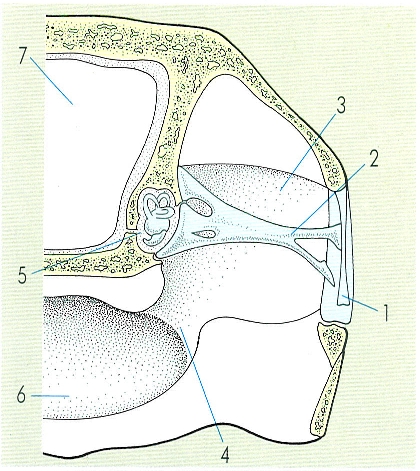
Esquema de l’oïda d’un rèptil. Hom hi ha indicat: 1 timpà, 2 estrep, 3 oïda mitjana, 4 trompa d’Eustaqui, 5 oïda interna, 6 gola, 7 cavitat cranial.
Amadeu Blasco, segons Romer.
Els òrgans sensorials són desenvolupats a partir del model amfíbic i no presenten excessives modificacions, ja que els amfibis ja havien resolt els problemes bàsics plantejats pel fet d’adquirir vida terrestre.
L’ull presenta dues parpelles mòbils i una altra, la membrana nictitant, llevat dels ofidis, que tenen parpelles soldades i transparents. L’òrgan auditiu presenta oïda interna ben desenvolupada, amb membrana timpànica ben visible, i oïda mitjana amb columel·la. És excepció el cas de les serps, on no s’observa ni timpà ni oïda mitjana, de manera que llur percepció de les ones de pressió sembla produir-se pel contacte entre el seu cos i el substrat i capten les vibracions transmeses per aquest darrer mitjançant la mandibula inferior. L’òrgan de Jacobson és un òrgan vòmeronasal que és petit en els crocodilians i els quelonis i esdevé més important en els escatosos, en què s’obre directament a la cavitat bucal; rep la informació química mitjançant les puntes de la llengua, que és molt activa en aquests animals. L’olfacte és molt important en el reconeixement de les preses, però també en cas de presència d’enemics o en la identificació de la parella reproductora.
Entre les peculiaritats dels rèptils en l’àmbit sensorial figuren l’ull pineal, que és quasi un ull normal en alguns grups, com els rinocèfals, però que pot faltar en molts altres; o les fossetes facials d’algunes serps, els crotàlids, amb funció termoreceptora, que permeten realitzar la localització de preses homeotermes.
Les glàndules endocrines
Són, en els rèptils, ben desenvolupades i sense cap peculiaritat específica. Inclouen la glàndula tiroide, les paratiroides, les suprarenals, els illots de Langerhans al pàncreas, i la hipòfisi.
L’aparell digestiu

La llengua dels rèptils té una gran mobilitat i sovint capta estímuls químics del medi, com en el cas del cròtal de la fotografia. Els extrems de la llengua bífida, característica dels rèptils escatosos, recullen les partícules químiques i les porten fins a l’òrgan de Jacobson, situat al sostre de la boca.
Aisa/J. Novak.
La llengua dels quelonis i els crocodilians no és retràctil i es troba situada al terra de la boca. Els escatosos, en canvi, presenten llengües ben desenvolupades, retràctils i, en el cas dels ofidis, bífides, que poden sortir per una escotadura especial mentre la boca roman tancada i que tenen una funció sensorial. També són ben desenvolupades les glàndules salivals. En ofidis, com els vipèrids, i alguns saures (els helodermàtids), existeixen glàndules verinoses, que deriven de les glàndules labials del maxil·lar superior, convenientment modificades i connectades, en general, a un aparell inoculador, que és una dent acanalada o tubular. Algunes serps no el tenen (són àglifes) i són inofensives; altres presenten dents verinoses en posició anterior (proteròglifes) o posterior (opistòglifes). La composició i toxicitat del verí són molt variables. Fonamentalment n’hi ha de dos tipus: un neurotòxic i l’altre hemolític.
Les dents són més localitzades que en els amfibis i només es troben en els maxil·lars, els palatins, els pterigoides i els vòmers. Les tortugues no tenen dents i la seva boca és recoberta d’un bec corni molt dur que supleix funcionalment aquelles; els crocodilians només presenten dents als maxil·lars i a les mandíbules. La dentició dels rèptils és polifiodonta (renovable diverses vegades) i homodonta (totes les dents iguals), encara que hi ha algunes modificacions, com les ja esmentades dents verinoses dels ofidis, en forma d’ullal, corbades, acanalades o tubulars. La implantació és acrodonta, a la part superior de la geniva, o pleurodonta, és a dir, en posició lateral, encara que els crocodilians, que també en això mostren llur estadi molt més evolucionat, presenten dents tecodontes, implantades dins d’alvèols dentaris.
El tub digestiu presenta un esòfag llarg, amb plecs longitudinals que permeten una distensió i l’engolida de preses grosses, un estòmac ben diferenciat de l’esòfag, llarg i fusiforme en els escatosos, i amb una regió muscular ben diferenciada en els crocodilians. L’intestí prim és llarg i ben enrotllat; el segueix l’intestí gros, amb cec, que desemboca, finalment, en una cloaca, com en els amfibis.
L’aparell respiratori
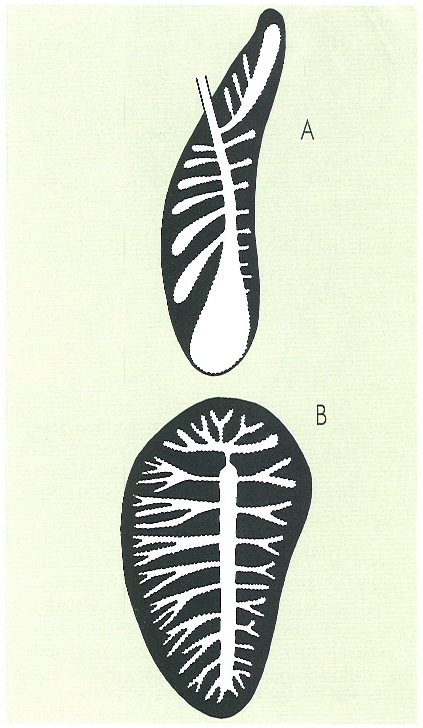
Els pulmons dels rèptils tenen les ramificacions dels sistemes bronquials molt profundes i això els permet acomplir íntegrament la funció d’intercanvi gasós. En el dibuix s’han representat els pulmons d’un queloni, concretament de la tortuga babaua (Caretta caretta, A) i d’un saure, del gènere Varanus (B).
Gustavo Hormiga, a partir de Bellairs i Attridge.
Els rèptils respiren exclusivament pels pulmons. La impermeabilitat de la pell no els permet una respiració cutània i només algunes tortugues i serps aquàtiques sembla que presenten respiració faríngia i en alguns casos cloacal. Els pulmons han evolucionat per a cobrir totes les necessitats de l’intercanvi de gasos, encara que són relativament simples i poc lobulats. En els rèptils, s’inicia la formació d’un paladar ossi que tendeix a independitzar les vies respiratòries nasal i bucal. Les coanes, en conseqüència, es desplacen cap enrere i es produeix un allargament conseqüent de la distància entre els orificis nasals externs i les coanes. La laringe és poc desenvolupada i no hi ha cordes vocals, de manera que els rèptils són normalment muts, amb molt poques excepcions, com els gecònids, alguns dels quals emeten alguns crits, espontàniament o, de vegades, en ser manipulats.
L’aparell circulatori
No hi ha grans modificacions respecte dels amfibis. La circulació és doble i completa, amb un circuit pulmonar i un circuit sistèmic general. El cor, però, amb tres cavitats, presenta les aurícules completament separades, mentre que el ventricle presenta una paret incompleta que separa els dos corrents sanguinis, l’oxigenat, procedent dels pulmons, i el no oxigenat, que prové de la resta del cos. En els crocodilians, aquesta paret és pràcticament completa i augmenta l’eficàcia del funcionament cardíac. Es mantenen el tercer, quart i sisè arcs aòrtics. El tronc arterial es divideix en tres branques: l’aorta pulmonar i les aortes sistèmiques dreta i esquerra; la dreta porta la sang oxigenada i d’ella parteixen les caròtides cap al cap, i l’esquerra porta sang parcialment oxigenada. Ambdós conductes s’uneixen després del cor i constitueixen l’aorta dorsal. No hi ha modificacions importants en el sistema venós respecte del cas dels amfibis.
L’aparell excretor
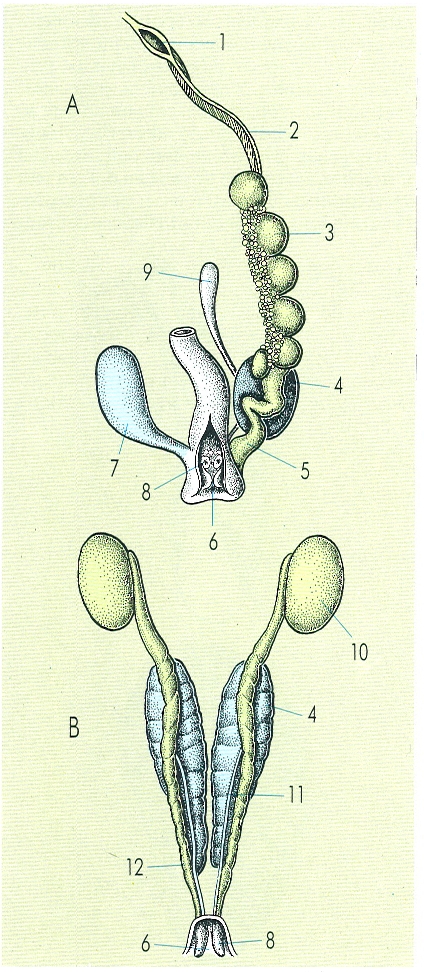
Esquema de l’aparell urogenital d’una femella (A) i d’un mascle (B) de rèptil. Hom hi ha indicat: 1 pavelló, 2 oviducte, 3 ovari, 4 ronyó, 5 úter, 6 cloaca, 7 bufeta urinària, 8 papil·la urogenital, 9 epidídim vestigial, 10 testicle, 11 urèter, 12 conducte deferent.
Gustavo Hormiga, segons Romer 1955.
Els rèptils són els primers vertebrats que presenten un ronyó de tipus metanèfric, el més evolucionat, encara que en fase embrionària és encara un mesonefre. És un ronyó més curt i compacte que el dels amfibis. La part posterior del conducte de Wolff esdevé l’urèter i es fon amb els túbuls col·lectors del ronyó metanèfric. La resta del conducte de Wolff només porta productes reproductors i esdevé, en el mascle, l’epidídim, els conductes deferents i les vesícules seminals. En la femella, la independència entre l’aparell excretor i l’aparell reproductor és total. La bufeta urinària és bilobulada i es desenvolupa com una evaginació de la cloaca en els saures i en els quelonis.
L’aparell reproductor
En la reproducció dels rèptils, l’avenç més important respecte dels amfibis és l’ou amniota. La resta de l’aparell reproductor, en el sentit anatòmic, i del procés reproductiu presenta modificacions menys notables. La reproducció fora de l’aigua exigeix forçosament una fecundació interna, per la qual cosa el mascle desenvolupa un aparell copulador.
En la femella, els ovaris són, en els escatosos, saculars i allargats; a la resta de rèptils, són de forma compacta, com en els tetràpodes superiors. Els òvuls formats en aquests ovaris són telolecítics, és a dir, amb abundant vitel·lus nutritiu, la qual cosa permet que les cries neixin en fase avançada, amb capacitat per valer-se totalment per elles mateixes. Són grossos i, per això, llur nombre és petit: les postes dels rèptils presenten xifres discretes d’ous, compreses entre dos i trenta o quaranta ous, per bé que les tortugues marines en dipositen sovint més de cent. Els òvuls s’alliberen pel trencament de la paret ovàrica van a parar als oviductes, en els quals desemboquen les glàndules encarregades de la secreció de l’albumen i la closca, coriàcia o calcària. Aquestes capes es van dipositant progressivament a mesura que l’òvul descendeix per l’oviducte i la fecundació té lloc a la part distal d’aquest. Els dos oviductes desemboquen independentment a la cloaca.

Hemipenis estès d’un mascle d’escurçó (Vipera). Hom aprecia a la fotografia el caràcter doble de l’òrgan i les espines característiques.
Antonio Bea.
En el mascle, hi ha un parell de testicles suspesos dins la cavitat corporal, de forma i mida variables, esfèrics, ovals o en forma de pera; els túbuls seminífers són llargs i enrotllats. El conducte de Wolff ha esdevingut exclusivament dedicat a la sortida dels espermatozoides a l’exterior i les vies renals s’han independitzat de les genitals. L’òrgan copulador, imprescindible per a traspassar els espermatozoides a la cavitat femenina, pot ser un penis simple o bilobulat; el p rimer cas es dóna en els quelonis i els crocodilians i el segon, en els escatosos, en els quals es constitueixen dos hemipenis. En tots els casos, l’òrgan copulador és protrusible per l’orifici cloacal, però no sempre pel mateix mecanisme: si és unic, és el cos cavernós el que permet l’extensió del penis, en omplir-se de sang, mentre que en els hemipenis, l’extensió i la retracció es fan per un mecanisme muscular.
Les característiques i les adaptacions fisiològiques dels rèptils
La regulació tèrmica

La regulació de la temperatura del cos en els rèptils, animals poiquiloterms, és ecoetològica. La fotografia mostra una sargantana (Podarcis) captant la calor directa emesa per un focus lluminós (heliotèrmia) i recollint calor del substrat mitjançant l’eixamplament del cos per extensió lateral de les costelles (tigmotèrmia).
Antonio Bea.
Els rèptils, com els amfibis, són ectotèrmics. Tenen, per això mateix, temperatures crítiques, aproximadament situades entre -3° C i 50° C, encara que sovint l’interval de temperatures que suporta l’individu és molt més estret. Així com els amfibis no tenen una excessiva delimitació de temperatures òptimes, els rèptils mostren clares preferències per un marge de temperatures estret, lleugerament diferent d’unes espècies a les altres o d’uns grups als altres i, en general, sensiblement superior a les temperatures de funcionament òptim dels amfibis.
En els rèptils, els mecanismes fisiològics d’ajustament de temperatures per al manteniment d’una temperatura corporal idònia no són òptims; llur regulació esdevé marcadament etològica. La temperatura corporal d’un rèptil es manté constant durant tot el període en què és actiu mitjançant aquests mecanismes, contràriament als amfibis, en què la temperatura corporal oscil·lava més directament lligada a la del medi. En cert sentit, els rèptils efectuen una termoregulació; l’òptim tèrmic oscil·la, per a molts d’ells, entre 28° i 40° C, llevat del cas dels rincocèfals, en què les temperatures òptimes són extremament baixes (10° a 12° C).
En els saures i en els ofidis és fàcil d’observar aquest procés. Quan surten del refugi, la seva temperatura corporal és baixa i no permet una activitat intensa. Cerquen un lloc adient, normalment sempre el mateix, prop del refugi, i es col·loquen apropiadament. En els saures, el fet que les costelles s’estenguin lateralment (l’animal esdevé més ample i aplanat) fa que rebin calor per insolació directa (heliotèrmia) o per contacte amb la superfície del substrat (tigmotèrmia), fins arribar a la temperatura òptima de funcionament. A partir d’aquest moment, l’animal pot esdevenir vertaderament actiu i partir a la caça d’aliment, en cerca de parella o per qualsevol altra activitat. Com sigui que la temperatura de l’ambient pot continuar augmentant i l’individu no necessita o no suporta temperatures superiors, regula activament la seva, fugint dels llocs massa oberts i desplaçant-se a indrets més obacs del seu territori, sota la vegetació; si la temperatura corporal disminueix, torna a la insolació directa i, així, pot mantenir constant la temperatura del seu cos durant tot el període actiu del dia.
El temps d’activitat dels rèptils és variable; en part, depèn de l’estació i, per tant, del temps que triguen en adquirir l’òptim tèrmic. És evident que del principi al final de l’estació activa hi ha diferències; a la primavera, en sortir de la hivernació, surten a assolellar-se relativament tard, quan els substrats són ja una mica tebis, i són actius al pic del migdia; a mesura que avança l’estació, surten cada cop més d’hora i a l’estiu eviten les hores massa càlides del migdia, en no poder compensar l’excès tèrmic, i, en tot cas, tornen a sortir a la tarda, quan baixa el sol. A la tardor s’inverteix el procés i tornen a centrar llur activitat enmig del dia, fins arribar novament a la hivernació.
L’intercanvi de gasos
La pell dels rèptils és totalment impermeable als gasos i tot l’intercanvi entre el medi intern i el medi extern es fa per la via pulmonar. Això comporta concentracions altes de CO2 als teixits, en forma de CO3 H2 convertit en ió bicarbonat CO3 H-, que els rèptils han de tolerar a llur sang.
Tot l’aportament gasós fet pels pulmons s’aconsegueix per ventilació pulmonar, bombeig bucofaringi i panteix. El mecanisme és més evolucionat que en els amfibis; l’aire entra per expansió muscular de la caixa toràcica i de la paret del cos, de manera que es produeix una depressió dintre del pulmó, que xucla l’aire (excepte en les tortugues, a les quals les costelles soldades a la closca, no permeten aquest moviment i introdueixen l’aire en els pulmons per contraccions alternes de muscles antagònics dels flancs i de les espatlles). D’aquesta manera, els rèptils aconsegueixen quasi tot l’oxigen necessari; l’eficàcia del mecanisme és en funció de la temperatura.
El bombeig bucofaringi sembla tenir, només, una funció olfactiva, durant les pauses respiratòries, encara que algunes tortugues aquàtiques pot ser que l’usin per l’obtenció d’O2, almenys en condicions de repòs de l’animal, la qual cosa els permet una immersió indefinida, afavorida per mecanismes complementaris, com la inhibició del batec cardíac, la glicòlisi anaeròbia, l’augment de l’ió CO3 H- al plasma, etc. De tota manera, només algunes espècies poden funcionar així; la resta de tortugues aquàtiques respira similarment als altres grups de rèptils, fonamentalment per ventilació pulmonar.
La regulació hídrica
Els problemes principals de la regulació osmòtica en els rèptils són, com en tots els casos, el control de l’aigua i els ions minerals i l’eliminació de residus nitrogenats. Els rèptils han aconseguit una independència molt gran de l’aigua i això significa que han resolt la regulació hídrica millor que els amfibis.
Els residus nitrogenats són emmagatzemats en forma d’àcid úric, que no és tòxic i que es pot conservar durant un cert temps sense dissoldre en aigua. Així se soluciona el problema en els adults i també en els embrions, que poden acumular-lo a la cavitat de l’al·lantoide sense problemes i, per tant, completar el seu desenvolupament dintre de l’ou amniota. Els adults poden també eliminar-lo en forma d’orina semisòlida, la qual cosa representa un estalvi hídric important que els permet de sobreviure en medis molt àrids. Com sigui que la pèrdua d’aigua a través de la pell, encara que existent perquè la impermeabilització cutània no és total, és molt més petita que en els amfibis, els rèptils necessiten quantitats molt més petites d’aigua per a sobreviure, la qual cosa els permet colonitzar medis desèrtics i, fins i tot, les aigües marines, com en el cas de les tortuges i d’algunes serps.
L’aigua obtinguda procedeix de l’aliment, i és, per tant, d’origen metabòlic, o pot ser adquirida prenent-la directament del medi —com fan les sargantanes, que aprofiten gotes de pluja o de rosada— o absorbida per canals especials, com en el cas d’animals desertícoles. La regulació de la pèrdua d’aigua és un dels resultats de l’adaptació evolutiva de cada espècie al seu medi. Les formes terrestres perden aigua més lentament que no les d’aigua dolça i, dins les terrestres, les que habiten a climes humits la perden més ràpidament que les que viuen en zones desèrtiques. Els rèptils marins i alguns saures desertícoles tenen glàndules de sal que eliminen activament ions i ajuden, així, a la funció renal en un medi pobre en aigua o excessiu en sal; l’excreció es produeix en forma de sals deshidratades de sodi i potassi.
L’estratègia biològica
La reproducció i el desenvolupament
El pas de l’aigua a terra modifica profundament la reproducció dels rèptils en relació amb la dels amfibis i soluciona prou correctament el problema de la dependència de l’aigua: es produeix íntegrament a terra, fins i tot en animals de residència permanentment aquàtica, com les tortugues marines. La fecundació és interna; apareix, per aquesta raó, un òrgan copulador i la posta es fa en un sòl tou, sota pedres o en qualsevol indret on els ous puguin rebre suficient aportació de calor durant tot el desenvolupament, ja que les capes protectores que recobreixen els ous eviten la dessecació. Rarament la posta i les cries reben atencions dels seus genitors. Encara que, fonamentalment, els rèptils són ovípars, hi ha espècies ovovivípares o pràcticament vivípares.
Encara que no han estat observats casos espectaculars d’hibridació, s’ha comprovat que es pot produir en captivitat entre espècies emparentades. Les observacions de camp són escasses, però no és improbable que formes molt properes s’aparellin i tinguin descendència, fèrtil o no. El camp més estudiat experimentalment afecta els lacèrtids, sobretot els llangardaixos verds.
L’ou amniota
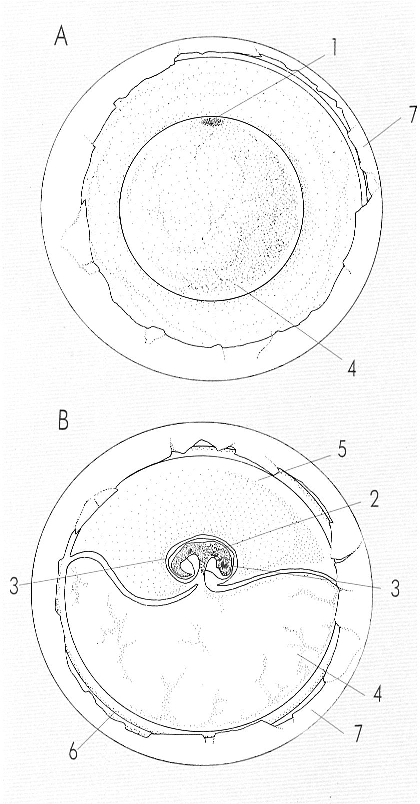
La formació d’un ou amniota, és a dir, proveït de determinades membranes embolcallants, és una característica que en l’escala evolutiva s’inicia amb els rèptils i permet que el procés de la reproducció s’esdevingui íntegrament en el medi estrictament terrestre. El dibuix mostra dos estadis del desenvolupament d’aquest ou: A posició del disc embrional, i B un estadi de desenvolupament més avançat. Hom hi ha indicat: 1 disc embrional, 2 embrió, 3 amni, 4 vitel·lus, 5 al·lantoide, 6 còrion, 7 closca.
Ferran Rodríguez, a partir de fonts diverses.
El desenvolupament directe i terrestre exigeix ous telolecítics, amb quantitat suficient de reserves nutritives. La femella produeix una quantitat d’ous inferior a la dels amfibis. L’òvul fecundat és protegit per membranes constituïdes a partir de teixits del propi embrió (amni, al·lantoide i còrion) i per membranes procedents dels teixits materns (les de la closca). A part això, hi ha la pròpia closca, segregada per les glàndules corresponents de l’oviducte que és present a totes les espècies ovípares i ovovivípares, amb o sense impregnacions de CO3Ca damunt una sèrie de capes fibroses coriàcies. L’oviducte matern segrega també l’albumen, el paper del qual és fer de reserva d’aigua.
El vitel·lus conté proteïnes, lípids i elements minerals diversos; és utilitzat per al desenvolupament embrionari, però una petita part pot conservar-se per a l’alimentació del nadís, almenys en algunes espècies. Part del calci necessari prové, en certes formes, de la closca i l’aigua procedeix de l’albumen o clara en altres casos, tot el calci és d’origen vitel·lí. La segmentació de l’ou dóna un disc embrionari petit (segmentació discoïdal), perquè la major part de l’ou és vitel·lus que no se segmenta. L’embrió va desenvolupant-se al mateix temps que el vitel·lus disminueix, en ser utilitzat per la construcció del nou rèptil.
Les membranes extraembrionàries apareixen molt aviat; l’amni és una capa ectodèrmica recoberta pel mesoderma, que embolcalla l’embrió a manera de sac i delimita una cavitat plena de líquid àmnic, que presenta una composició semblant a la del sèrum i proporciona protecció. Aquest líquid té una funció paral·lela a la del medi aquàtic en la reproducció de les espècies aquàtiques. L’al·lantoide és format per endoderma i cobert de mesoderma, i procedeix de la part posterior del tub digestiu embrionari, que acaba creixent enormement; actua com un dipòsit de residus nitrogenats de les cèl·lules embrionàries i, allà on connecta amb el còrion, estableix una mena de pulmó embrionari, altament vascularitzat, que intercanvia gasos a través de la closca, amb la qual té un contacte directe. El còrion, ectoderma recobert de mesoderma, se situa sota la closca i les membranes subjacents a aquesta, i embolcalla integralment l’embrió i el sac vitel·lí. Totes aquestes capes són abandonades després de descloure’s l’ou, juntament amb la closca.
L’aparellament i la posta

Durant el període de reproducció és corrent en els rèptils que es produeixin baralles entre els mascles, que són seguides, en l’espècie de la fotografia, el dragó comú (Tarentola mauritanica), de la precòpula, que igualment és molt agressiva.
Javier Andrada.
En els rèptils no es produeix el gregarisme reproductor dels amfibis, sinó que la reproducció la realitzen parelles isolades, poc conspícues. Encara que els animals presenten modificacions en el moment de la reproducció, aquestes es limiten a variacions, no gaire espectaculars, de la coloració, com la gola blava del mascle del lluert (Lacerta viridis); en formes tropicals, hi ha, amb tot, casos forja evidents de dimorfisme sexual, però el més freqüent són les variacions moderades en la mida i en els proporcions corporals o en la coloració i les ornamentacions, en general poc exagerades.
El ritual previ a la còpula tampoc és gaire complex, però segueix patrons ben definits per a cada família i, fins i tot, per a cada espècie. El mascle persegueix la femella i, després d’identificar-la per l’olfacte, la vista o el comportament, s’entortolliga a ella i la mossega fins immobilitzar-la i aconseguir completar la còpula. En els ofidis, les danses poden resultar espectaculars, i encara més els combats entre mascles, previs a l’època d’aparellament, igualment ritualitzats i que rara vegada acaben amb lesions greus o mortals, com passa amb freqüència en els vipèrids i en alguns altres grups. Les amenaces tracten sempre d’intimidar l’adversari donant la sensació que l’animal és d’una mida més grossa que la real. Els mascles, dilaten el seu cos, estenen prolongacions diverses i s’ataquen amb la boca molt oberta, o adopten qualsevol altra actitud en aquest sentit. No tots els rèptils disposen d’estructures específiques, com collars o espines, però la majoria de mascles defensen asprament la seva femella. Un cop allunyats els competidors i aconseguida la femella, la còpula es verifica per aposició de cloaques i entortolligament de cues, en els escatosos, o muntant el mascle al dors de la femella, en els quelonis.

L’ovoviviparisme, present en algunes espècies de rèptils, es considera una adaptació a climes freds. En són exemple els vipèrids; a les fotografies hom observa dos estadis successius, corresponents a un escurçó (Vipera). Els ous són envoltats d’una membrana fina i transparent que es trenca just feta la posta i allibera un individu preparat per a valer-se tot sol.
Antonio Bea.
La reproducció terrestre per parelles, aïllades, té l’avantatge d’afavorir la dispersió dels ous i la capacitat de supervivència de les postes i les cries. Però també té el problema de la localització de mascles i femelles —cal tenir en compte la manca de comunicació auditiva—, encara que en molts casos la hivernació conjunta d’individus d’ambdós sexes facilita que, a la primavera, quan comença el període actiu, mascles i femelles es trobin ja junts per a una reproducció pràcticament immediata.
La fecundació pot ser, en certs casos, retardada, i això augmenta l’eficàcia reproductora, especialment si es tenen compte les dificultats existents per a trobar parella en el moment propici. Els receptacles seminals conserven l’esperma, que serà utilitzable en temporades posteriors. Els rèptils són majoritàriament ovípars, però també en aquesta situació els ous són dipositats amb l’embrió en ple desenvolupament; la retenció dintre de l’oviducte permet de mantenir un correcte nivell hídric i també una temperatura constant.
Per això, especialment si les condicions ecològiques són desfavorables, hi ha casos d’ovoviviparisme, com el del vidriol (Anguis fragilis) o la sargantana vivípara (Lacerta vivipara), en què la posta coincideix amb el moment de la desclosa dels ous, o, fins i tot, un cert grau de viviparisme, amb una mena de placenta en la qual es produeix un intercanvi sanguini entre la mare i l’embrió. Aquests casos afecten especialment formes d’alta muntanya i de climes freds, amb un cicle molt curt i en un medi altament desfavorable, en què el desenvolupament embrionari pot, fins i tot, estendre’s per més d’un any. En algunes espècies hi ha poblacions ovovivípares en llocs desfavorables, mentre que la majoria són ovípares en localitats més temperades.

La posta, en els rèptils, es fa sempre a terra, àdhuc en el cas d’espècies que menen una vida aquàtica. Hom mostra a les fotografies alguns exemples d’aquestes postes. A dalt, moment de la posta d’una tortuga mediterrània (Testudo hermanni), els ous de la qual són calcaris; al mig, posta, en el moment de la desclosa, del dragó comú (Tarentola mauritanica), d’ous també amb closca calcària; a baix, posta de colobra (Natrix), amb closca còrnia, com una gran majoria dels rèptils.
Fèlix de Pablo, Javier Andrada i Ramon Torres.
La posta és dipositada unes setmanes o mesos després de l’aparellament. Els ous són quasi esfèrics o bé ovoides, molt allargats, i d’una mida relacionada amb la de l’animal adult, però acostumen a ésser relativament grossos en proporció a la mida de la femella; el color és blanc pur o brut, amb closca calcària, en els gecònids i en els quelonis, i coriàcia, en altres saures i en els ofidis. Normalment, els ous romanen a terra, en llocs on hi ha suficient humitat i calor per a la incubació, a la sorra, sota la fullaraca, en troncs podrits, enterrats al sòl, sota pedres, etc. Les postes, per les raons ja esmentades, són més petites que en els amfibis i consten de dos a 150 ous. El nombre de postes per any varia des d’una sola, en climes temperats, o diverses, en clima càlid, fins a una cada dos anys, en clima fred. La durada de la incubació és variable i depèn del període previ a la posta i de l’espècie.
Rares vegades hi ha atencions dels progenitors, com ocorre en els aligàtors i en les pitons. El normal és que, en descloure’s l’ou, la cria s’hagi de valer per ella mateixa i ho faci amb extrema competència. La cria té, damunt el musell, una dent de desclosa que li facilita el trencament de la closca, que cau un o dos dies després. Les cries són molt similars als adults, però poden presentar diferències de dibuix i coloració, que o bé pot ser més viva o bé, simplement, presentar un altre disseny; el seu cap sol ser més curt i arrodonit, proporcionalment, que el de l’adult i presentar ulls força més grossos en relació a la mida del cos. La cria creix inicialment de manera molt ràpida; després continua creixent cada vegada més lentament.
La maduresa sexual pot trigar anys en aparèixer i l’animal l’assoleix quan ja té una mida respectable. No hi ha gaires dades segures respecte de la longevitat dels rèptils en condicions naturals, però, encara que és variable d’una espècie a l’altra i és probablement relacionada amb la mida dels animals, sembla ser llarga i, en alguns casos, com en molts quelonis, arribar a ésser reconegudament excepcional.
Els cicles d’activitat
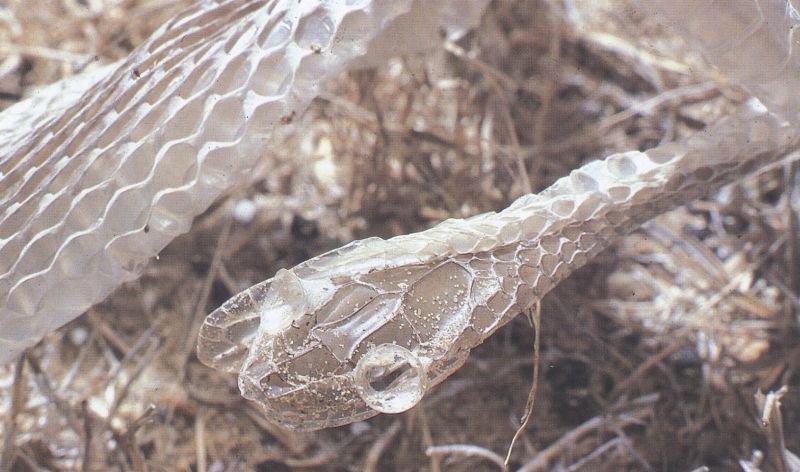
La renovació periòdica de la capa còrnia que recobreix el tegument dels rèptils és imprescindible per a permetre el creixement de l’animal. No obstant això, la manera com es desprèn aquesta muda difereix segons els grups: ho fa en escates soltes en els quelonis, a trossos en els saures i en una sola peça en els escatosos àpodes, tant en els ofidis com en els saures. És un exemple d’aquest darrer cas, la «camisa» de serp verda (Malpolon monspessulanus) que ens mostra la fotografia, en la qual hom identifica perfectament les escates del cap que fan inconfusible l’espècie.
Javier Andrada.
Com a animals ectotèrmics, encara que llur capacitat de regulació tèrmica sembla superior a la dels amfibis, també els rèptils presenten una diapausa hivernal, almenys en climes freds i temperats. La durada de la hivernació depèn de la latitud i l’altitud i passa de ser pràcticament inexistent a allargar-se fins a deu mesos sencers. Un cop l’animal surt de la hivernació, passa a seguir el cicle d’activitat normal; els rèptils són més termòfils que els amfibis i de costums més diürns, sobretot al principi de l’estació activa, en què les temperatures són més baixes. La fase reproductora es produeix molt poc després d’entrar en activitat (entrada que a la mateixa latitud i altitud és, sempre, més tardana que en els amfibis), encara que la posta és decalada i la desclosa dels ous se separa bastant de l’aparellament i té lloc al pic de l’estiu. Els animals adults segueixen primer un període d’insolació en començar el dia, quan la temperatura arriba a un mínim determinat, característic de cada espècie, que puja en arribar els mesos de calor i torna a disminuir a finals de l’estiu, de manera que els animals són actius a temperatures més baixes a la primavera i a la tardor que a l’estiu; un cop assolellats, passen a la caça i a períodes de repòs, al sol directe o a l’ombra, segons llurs necessitats tèrmiques. Algunes espècies, les menys termòfiles, són francament crepusculars o nocturnes, com els escurçons o els vidriols.
Dels rellotges interns que controlen l’entrada en el període actiu o la sortida a l’activitat de cada dia no se’n coneix gaire cosa. Com hem exposat en parlar dels amfibis, aquests mecanismes semblen estar més relacionats amb el calendari que amb les condicions concretes d’un any determinat, però, de tota manera, aquestes condicions tenen alguna moderada influència. Els animals s’agrupen per parelles en les etapes reproductores; fora d’aquestes etapes, són solitaris. Moltes espècies són territorials, sobretot en el cas dels mascles, fins i tot fora de l’estació reproductora.
L’adequació a l’ambient
Factors físics
Els rèptils són animals d’exigències força termòfiles i amb una higrofília limitada, però disposen d’una regulació tèrmica superior a la dels amfibis i, de fet, són animals estesos per tots els continents i totes les latituds i, fins i tot, arriben a notables altituds.
Les necessitats d’assolellament per a adquirir la temperatura òptima porten els rèptils a preferir àrees solellades i a defugir, en general, les obagues. Igualment, eviten, en la majoria de casos, les formacions vegetals massa denses. Prefereixen, doncs, les localitats amb una bona exposició solar. L’aridesa és també un factor ben suportable per moltes espècies de rèptils, que no exigeixen gaire humitat, en general, aigua líquida, i habiten, fins i tot, a indrets desèrtics.
Hi ha, però, excepcions notables a aquestes exigències bàsiques d’insolació. Algunes formes poc termòfiles, com el vidriol (Anguis fragilis) o els escurçons (Vipera), a les nostres terres poden esdevenir crepusculars o, fins i tot, nocturnes, i prefereixen àrees obagues, ben humides i amb força coberta vegetal.
Un altre factor important el constitueix el substrat. Normalment els rèptils no són exigents amb la qualitat química del substrat, però de la naturalesa de les roques depenen aspectes tan importants com l’estructura del sòl o la possibilitat de formació dels imprescindibles refugis. Així, algunes formes excavadores exigeixen sòls tous, esponjats; el vidriol, que els prefereix, pot viure també en altres localitats; els gecònids europeus, molt antropòfils, es comporten originàriament com lapidícoles i exigeixen afloraments rocosos amb esquerdes adients; no es troba cap espècie a localitats nues, sense pedres ni vegetació on refugiar-se.
Algunes formes exigeixen medi líquid; són les colobres d’aigua (com les nostres Natrix), que toleren, però, moltes menes d’aigües quietes, les tortugues d’aigua dolça, que volen aigües tranquil·les amb força vegetació, i les tortugues marines, de vida exclusivament aquàtica, tret del període de la reproducció. L’exigència d’aigua líquida pot ser independent de les condicions requerides fora de l’aigua, ja que un rèptil de costums amfibis pot suportar perfectament climes molt càlids, que aprofita per assolellar-se apropiadament, regulant la temperatura corporal amb estades alternatives a la vora i dins de l’aigua.
En definitiva, com s’observa en molts grups zoològics, el més important en la distribució dels rèptils, des d’un punt de vista abiòtic, és l’estructura i l’orientació dels substrats, que condicionen els factors restants d’insolació, temperatura, humitat i vegetació.
La vegetació
Els rèptils no mostren preferència per una vegetació determinada, si no és a nivell de grans formacions; és a dir, no depenen, per exemple, d’un tipus de bosc, sinó, en tot cas, del bosc com a estructura. La termofília els obliga, per altra banda, a escollir formacions vegetals que ofereixin superfícies d’assolellament —àrees, per tant, més o menys obertes— i, al mateix temps, possibilitat de refugiar-se dels excessos tèrmics a l’ombra de les plantes, les arrels de les quals, a més, poden servir com a refugis a moltes espècies. Encara que hom troba rèptils a tots els indrets i a llocs amb tota mena de masses vegetals, són més escassos en zones molt obagues i denses; són també poc diversos, encara que no rars, en localitats de molt escassa coberta vegetal i, evidentment, són quasi inexistents en substrat nu. Esdevenen especialment abundants, almenys en terres temperades com les nostres, en els límits entre dos medis, com poden ser les vorades de bosc, les bardisses, sobretot enmig de camps o ran de bosquines i a altres localitats similars.
Les relacions tròfiques
L’alimentació

El règim alimentari carni és el predominant en els rèptils, amb poques excepcions, com la de molts quelonis i algunes espècies que tenen hàbitats peculiars. La classe i les dimensions de llurs preses depenen de les dimensions del mateix rèptil i de les disponibilitats del medi. En els saures, com el llangardaix ocel·lat (Lacerta lepida) que veiem a la fotografia de l’esquerra, predominen les espècies insectívores; en els ofidis, gràcies a la peculiar disposició de la mandíbula, es pot observar el consum de preses de diàmetre força superior al del predador, com veiem a la fotografia de la dreta, on la serp d’aigua (Natrix maura) consumeix un gripau del gènere Bufo.
Javier Andrada i Xavier Parellada.
Els rèptils són fonamentalment carnívors i les excepcions herbívores són poques: algunes tortugues presenten un règim predominantment, però no exclusivament, vegetarià i alguns saures, en medis pobres, com les sargantanes als illots balears, també desvien llurs fonts alimentàries cap al regne vegetal, per manca d’un aliment animal suficient. Els tipus i mides de les preses, en el cas dels carnívors, semblen dependre de la fauna del medi on viu l’animal i de la mida d’aquest propi animal. La majoria no semblen tenir hàbits selectius en llur alimentació, com tampoc els tenien els amfibis.
Però la mida de cada espècie i el medi condicionen fins un cert punt l’alimentació. Les sargantanes mostren preferència pels insectes, mentre que els llangardaixos, més grossos, també en consumeixen, però poden igualment capturar petits vertebrats, fins i tot, de llur propi grup o les pròpies cries, que no són ni poc ni molt protegides davant d’aquesta situació. Les serps poden consumir insectes i invertebrats diversos quan són petites, però en esdevenir adultes llurs preses preferides són els petits vertebrats i, fins i tot, arriben a especialitzar-se: les colobres d’aigua consumeixen moltes larves d’amfibi, amfibis adults i peixos, principalment; algunes serps africanes consumeixen quasi exclusivament ous, altres mengen amb preferència saures. Els crocodilians mengen tota mena de vertebrats.
La captura de l’aliment es fa a l’aguait i a la carrera, principalment gràcies a la vista, però en el cas dels herbívors, potser, l’olfacte hi té un paper fonamental. Un cas especial el constitueixen les serps verinoses (vegeu la pàgina següent, sobre recursos defensius), que ataquen la presa des del punt on eren a l’aguait i que, un cop la tenen, poden retenir-la fins que mori o deixar-la fugir, com fan els escurçons, i seguir-ne després el rastre olfactivament, traient les puntes de la llengua per tal que les partícules olfactives siguin portades per aquesta fins a l’òrgan de Jacobson. Els cròtals disposen d’una fosseta tèrmica facial especial que els permet de localitzar animals homeoterms per la diferència de temperatura que presenten respecte de l’entorn.
La competència
La competència interspecífica i intraspecífica és relacionada, sobretot, amb l’aliment disponible.
Dins d’una mateixa espècie, les diferències en la mida dels individus diversifiquen llurs exigències i els adults més grossos acostumen a explotar preses més grosses que els subadults; els joves sovint tenen un espectre alimentari completament diferent, per raó de la seva petitesa. En cas de pobresa extrema de recursos, com, per exemple, en el de les sargantanes balears, la competència pels pocs insectes o per les escasses fonts vegetals disponibles esdevé aferrissada i només els animals més sans arriben a l’estat adult, de manera que la població es manté dins dels nombres, limitats, que els recursos alimentaris toleren.
En la competència interspecífica el problema no es planteja, en teoria, perquè cada espècie ocupa en exclusiva un nínxol ecològic i explota uns recursos alimentaris determinats. En la realitat, però, hi ha una coincidència almenys parcial de recursos alimentaris, i algunes espècies són desplaçades per altres de semblants i equivalents que els exploten millor. Aquest podria ser el cas, en les regions dels NE ibèric, del llangardaix ocel·lat (Lacerta lepida) i el lluert (Lacerta viridis); dels dos llangardaixos, el primer, més gros, i també més termòfil, limita clarament el segon; el lluert podria viure, probablement, en localitats més baixes i més càlides que les que ocupa, però hom interpreta que el llangardaix ocel·lat, que el supera en mida, és més capacitat per a la captura de preses i aconsegueix de desplaçar-lo i substituir-lo. El lluert només apareix en aquelles localitats més altes, humides i fredes on el llangardaix ocel·lat no pot viure, perquè la seva termofília no li ho permet. Tal interpretació és documentada pel fet que, en localitats d’idèntica altitud i latitud, hi trobem el llangardaix ocel·lat si són prou assolellades i seques, encara que siguin dels Prepirineus o dels Pirineus, on predomina el lluert. En altres àrees, però, aquesta competència no existeix i, fins i tot, a Catalunya, és discutible.
Els recursos defensius
La pressió de predació
Els enemics dels rèptils són múltiples. Normalment es tracta de vertebrats i, de vegades, poden ser de llur mateixa classe. Molts mamífers petits són especialitzats en la captura de sargantanes o almenys en capturen sovint. Alguns rapinyaires diürns, com l’àguila marcenca (Circaetus gallicus), capturen predominantment serps. Les serps, per la seva part, com les del gènere Coronella, consumeixen sargantanes. Algunes serps adultes poden consumir-ne de més petites i ser ulteriorment consumides per mamífers carnívors grossos.
Els efectes de la predació són, però, selectius. Dels predadors naturals, les poblacions de rèptils se’n ressenten poc. Només en el cas de cries, les captures poden esdevenir importants, però la quantitat d’ous dipositada i amb possibilitats d’èxit és suficient per a contrarestar aquest efecte de destrucció en massa dels joves, més maldestres. Normalment, només en el cas de l’home i també en el dels seus animals doméstics, es porta a terme una predació en massa de les poblacions, que no són en condicions de compensar-la, sobretot si aquesta ataca els ous, com en el cas de les tortugues marines, o si es tracta de captures abusives amb alguna finalitat industrial (per exemple, l’explotació de la pell en crocodilians i serps o l’obtenció d’exemplars comestibles de certes tortugues).
Els quelonis i els crocodilians no corren perill de caure en mans de predadors quan són adults, perquè la closca dels primers i la mida i la dentició dels segons normalment són suficients per a defensar-los. Les postes i els joves, més febles, solen ser víctimes d’alguns carnívors de mida més respectable o de l’home. Tanmateix, hi ha rapinyaires, i també còrvids, especialitzats en la captura i depredació de quelonis, la closca dels quals trenquen llençant-los des de gran altura.
L’autotomia

La facultat d’autotomitzar la cua, és a dir, la capacitat de seccionar-la pel mig d’una vèrtebra caudal proveïda d’un pla de fractura especial, és propietat de molts saures. Després de desprendre’s, la cua és regenerada, en un termini breu, encara que sense un esquelet normal. A les fotografies podem veure (a l’esquerra) una regeneració normal, en el dragó comú (Tarentola mauritanica), en què el fragment de cua regenerat es diferencia bé de la base, i un fenomen gens rar (a la dreta), pel qual la cua original no acaba de seccionar-se i, en produirse igualment el procés de regeneració, dóna lloc a la curiosa imatge de la fotografia: un exemplar de Podarcis, en què coexisteixen dues cues.
Fèlix de Pablo i Francesc Junyent.
Un mecanisme característic que concerneix la defensa de l’animal és l’autotomia caudal. Encara que altres grups, com certs crustacis, presenten autotomia en certes parts del cos, el cas dels rèptils és únic entre els vertebrats. Consisteix en una automutilació amb desprendiment de la cua. L’òrgan eliminat, que no és vital, és regenerable i es pot compensar la seva pèrdua. Només els rincocèfals, els saures i algunes serps presenten autotomia, que és exclusivament caudal.
L’autotomia es produeix per partició d’alguna de les vèrtebres caudals segons un pla predeterminat i amb una pèrdua mínima de sang. El tros de cua segueix bellugant-se després de despendre’s, davant del predador, durant bastants minuts o fins a mitja hora, cosa que permet la fugida del seu antic propietari. És necessari que la cua sigui manipulada perquè l’animal se’n desprengui, per un procés de contracció muscular que acompanya la fractura del pla vertebral predeterminat. Normalment, els animals actius es desprenen de la cua més fàcilment quan són mig letàrgics i sembla que els joves la trenquen més fàcilment que els adults, o, si es vol, sembla que els adults que l’han conservada sencera, que en algunes poblacions són ben escassos, ja no la deixen anar amb facilitat, encara que se’ls manipuli.
Un cop perduda la cua, la ferida cicatritza molt aviat i comença immediatament la regeneració. La nova cua no té esquelet ossi, sinó que moltes vegades té una mena d’eix cartilaginós i és sensiblement diferent de la cua original, de manera que es pot veure, a ull nu, la patent diferència en coloració i en folidosi entre la cua original i la regenerada. La nova cua creix prou ràpidament i acostuma a assolir una mida semblant a la de l’original. De vegades, la cua queda només mig autotomitzada, però comença igualment la regeneració de la cua nova; no és pas rar observar sargantanes amb dues cues. En algunes espècies, la regeneració és escassa o quasi nul·la.
La cripsi

Una reacció de defensa que empren alguns rèptils és l’il·lustrat a la fotografia, d’un exemplar de serp d’aigua (Natrix natrix), que consisteix a fer-se el mort, deixant-se caure panxa enlaire.
Antonio Bea.
Els mitjans de defensa dels rèptils són similars als emprats per altres grups zoològics, encara que, entre ells, es troben espècies verinoses, que no és un cas tan general.
La primera defensa és, per a la majoria de rèptils, la cripsi, afavorida per llur immobilitat durant hores. Es dissimulen tan bé en el medi, sobretot els escatosos, que només un ull avesat els distingeix i sovint només es capta llur moviment de fugida si hom se’ls apropa massa. La majoria de rèptils, si corren perill i tenen la possibilitat de fer-ho, s’escapen ràpidament; és el seu medi de defensa més usual. Les coloracions aposemàtiques no serveixen de defensa, en el sentit del terme que usem aquí, sinó que més aviat constitueixen, en el cas d’animals territorials, una advertència. Només algunes serps tòxiques americanes, les corals, i els saures helodermàtids presenten coloració d’advertència de llur toxicitat i determinen l’aparició de mecanismes de mimetisme (són imitades per formes inofensives que així queden també protegides).
Algunes serps també fan el mort, recurs molt socorregut en el regne animal, i es posen panxa enlaire, absolutament immòbils, fins i tot quan se les manipula. Alguns saures poden adoptar postures d’amenaça, com, per exemple, en cas de perill, presentar la boca ben oberta al contrari o l’enemic per tal d’intimidar-lo, tot aparentant una mida superior a la real.
La toxicitat
Una cas peculiar és el de la possessió de verí. Hi ha una família de saures verinosa, els helodermàtids centramericans, però els rèptils verinosos típics són els ofidis: de cròtals, cobres, mambes, escurçons, etc., n’hi ha moltes espècies que posseeixen glàndules tòxiques. Algunes les tenen connectades amb dents verinoses acanalades o tubulars, i altres no les tenen connectades amb cap dent. En certs casos, són en posició posterior (opistòglifes) i es claven en la presa quan l’engoleixen, per immobilitzar-la; en altres, les dents verinoses són en posición anterior (proteròglifes), plegades contra el sostre de la boca en repòs, i inoculen el verí a la primera mossegada.
Cal tenir en compte que el verí és, primordialment, un recurs per a l’alimentació, més que un medi de defensa, i que la serp triga un cert temps en elaborar-ne suficient per a inocular-lo en una presa o en un enemic. Inocular-lo injustificadament és malbaratar-lo. Per això, les serps verinoses tenen recursos per a evitar usar el verí fora de llur aplicació més immediata, pràctica i rendible, que és obtenir el menjar. Els cròtals fan sonar el cròtal (estructura d’anells ossis de la cua) en advertència, les cobres despleguen llur coll en aixecarse per intimidar i advertir l’enemic, que les identifica i fuig, o s’espanta en veure aquell simulacre de cap amb dos ulls grossos; els nostres escurçons prefereixen fugir, si poden, abans que mossegar gratuïtament, i bufen en clara advertència de perill. L’atac d’aquests animals és, però, quan es desencadena, rapidíssim.