L’ecofisiologia del desert
Per tal de comprendre per què una àrea determinada de la superfície de la Terra és un desert cal, en primer lloc, esbrinar les causes que impedeixen que hi prosperi la vegetació. A la majoria dels deserts la causa primera sol ser un abastament d’aigua insuficient. Per això convé saber diferenciar entre aridesa, de sequera i de desertització. El grau d’aridesa d’un clima és una característica permanent i mesurable, a despit de fluctuacions. La sequera, al contrari, és sempre de natura relativa i temporal, difícil de definir de manera precisa, si no és amb relació a una pluviositat respecte a la qual un any o una estació es desvia en el sentit d’accentuar el dèficit hídric. La desertització, a diferència de la sequera, té conseqüències perdurables, de vegades permanents i sovint irreversibles; més endavant es tractarà extensament. (Vegeu també Els tipus de clima de la Terra)
Els constrenyiments hídrics de la vida vegetal
Les plantes terrestres contenen una elevada proporció d’aigua, en la majoria de casos superior al 50%, i fins i tot, de vegades, al 95% o més, però el més important és que l’aigua està íntimament lligada a la seva activitat. L’intercanvi de diòxid de carboni amb l’atmosfera en la respiració i la fotosíntesi implica la presència d’uns canals oberts a l’intercanvi de vapor d’aigua. El citoplasma de les cèl·lules fotosintetitzadores —amb un contingut d’aigua del 75% com a mínim — ha d’estar exposat a l’atmosfera per tal de poder capturar i assimilar el diòxid de carboni i, per exemple, a una temperatura de 20°C i una pressió de vapor de 15 mbar, ha d’experimentar una pèrdua neta d’aigua de fins a uns 10 mbar de mitjana.
El risc de perdre l’aigua indispensable per a l’activitat protoplasmàtica ha estat una conseqüència inevitable de la migració de les plantes des del seu ambient aquàtic original cap a les terres emergides ara fa uns 500 milions d’anys. Les plantes terrestres han desenvolupat mecanismes per a restringir les pèrdues d’aigua. Generalment consisteixen a envoltar la membrana citoplasmàtica amb una cutícula impermeable a través de la qual el bescanvi gasós queda limitat a uns porus que es poden obrir i tancar, els estomes. En algunes espècies, els estomes es tanquen durant el dia i s’obren de nit, quan la humitat atmosfèrica exterior és relativament elevada. D’aquesta manera la pèrdua d’aigua queda molt limitada mentre que l’intercanvi de diòxid de carboni no es veu obstaculitzat.
A més del paper que juga en mantenir el protoplasma actiu, l’aigua també és fonamental per a transportar els nutrients des del sòl cap a l’interior de la planta i fins als centres metabòlics. El motor d’aquest procés és la transpiració, i perquè sigui possible, l’aigua s’ha de trobar en forma líquida al sòl, i ser captada a través d’òrgans especialment adaptats per a aquesta funció, les arrels. Els nutrients són arrossegats juntament amb l’aigua gràcies al continuum sòl-planta-atmosfera que s’estableix. Les plantes terrestres han conservat, doncs, com a memòria de l’ambient aquàtic ancestral, una dependència de la disponibilitat d’aigua al medi tant per al transport de nutrients com per a reemplaçar l’aigua perduda a causa de la necessària exposició de les superfícies fotosintètiques a una atmosfera no saturada. Per això mateix, la colonització de la superfície terrestre començà en zones on l’atmosfera es trobava en un estat pròxim a la saturació i on el sòl proveïa d’aigua de manera constant i abundosa. La penetració de les plantes superiors a zones amb una menor disponibilitat d’aigua i amb l’atmosfera en condicions molt més allunyades de la saturació, no fou possible fins molt més tard i després d’un seguit de cicles adaptatius.
Tot això és aplicable a les plantes superiors, proveïdes de fulles, estomes i sistemes radiculars diferenciats, com també de processos reproductius independents de l’alliberament en el medi aquàtic dels elements reproductors. Probablement, però, altres organismes fotosintètics colonitzaren la superfície de la Terra molt abans. Eren organismes capaços de sobreviure en períodes eixuts en un estat de dessecació, per reviscolar i tornar a ser actius tan bon punt hi hagués disponibilitat d’aigua. Aquest és el cas de determinades algues, líquens i molts briòfits, que no són dependents d’una disponibilitat constant d’aigua, i que aconsegueixen limitar la seva activitat a les oportunitats esporàdiques ofertes per les pluges, durant les quals la superfície del sòl és humida i la pel·lícula d’aigua conté tant diòxid de carboni com nutrients dissolts, dels quals s’alimentaran durant el breu període que la superfície triga a assecar-se.
Els marges tèrmics de la supervivència de les plantes
A més de l’aigua, les plantes tenen certes exigències pel que fa a la temperatura. El rang de temperatures que permeten la seva supervivència és bastant estret, i aquell que els permet mantenir-se actives encara ho és més (per a la majoria d’espècies queda limitat entre 0 i 40°C aproximadament). L’oceà primigeni era, hipotèticament, líquid, de manera que la seva temperatura havia de situar-se entre 0 i 100°C i, atès que l’aigua té la calor específica més elevada que es coneix, les variacions que podien resultar de la influència del bescanvi energètic eren molt lleus i la seva manifestació, molt lenta. Així doncs, les plantes evolucionaren inicialment en un ambient amb un rang de temperatures molt limitat, per trobar-se, en colonitzar la superfície terrestre, amb un marge molt més ampli en el qual temperatures inferiors a 0°C poden ser freqüents durant els mesos d’hivern mentre les d’alguns microhàbitats poden ser molt superiors a les dels oceans.
Les condicions extremes de temperatura, anàlogament a la manca d’aigua, haurien estat menys severes en els ambients terrestres més humits —aiguamolls i altres zones humides— que foren, sens dubte, els primers espais colonitzats i a partir dels quals s’inicià la migració cap a ambients terrestres més extremats. Les darreres zones colonitzades foren les menys hospitalàries a causa de la seva aridesa i temperatures extremes, fins a arribar als deserts, on la petjada de la vida vegetal és encara molt lleu.
Les vies metabòliques privilegiades
La llum i la radiació no constitueixen generalment un factor limitador de la producció vegetal als deserts i subdeserts, ja que el flux energètic és generalment superior a 140 000 cal/cm2 i any. Només localment, a les zones amb boires de les riberes oceàniques i d’algunes muntanyes, la lluminiositat i el flux energètic poden ser localment o temporal un factor limitador. Tanmateix, la disponibilitat d’aigua i l’estat dinàmic dels ecosistemes de deserts i subdeserts condicionen poderosament la producció primària, altrament dit, la productivitat és limitada per l’aridesa, per la migradesa de la vegetació i la condició del sòl i no totes les vies fotosintètiques resulten igual d’eficaces en les condicions d’aridesa i de disponibilitat d’aigua al sòl dels deserts i subdeserts.
La majoria de les plantes vasculars apliquen en la fotosíntesi la via metabòlica anomenada C3. Les altres dues vies fotosintètiques que es coneixen a les plantes vasculars, la C4 i la CAM, són adaptacions fisiològiques a temperatures elevades i humitat escassa. Els termes C3 i C4 es refereixen al diferent processament dels productes interns de la fotosíntesi. En les plantes que segueixen la via C3 es forma l’àcid 3-fosfoglicèric, un àcid de tres carbonis i en les plantes C4, es produeix l’àcid oxalacètic, de quatre carbonis. Tanmateix, la via CAM deu el seu nom a les sigles angleses de “Crassulacean Acid Metabolism” (‘metabolisme àcid de les crassulàcies’), la família de plantes on aquest procés es va detectar per primera vegada. Les plantes CAM fixen el diòxid de carboni en forma de malat durant la nit i el descarboxilen durant el dia assimilant el diòxid de carboni que s’allibera. D’aquesta manera, aquestes plantes tenen els estomes tancats durant el dia, quan hi ha més perill de perdre aigua a causa de les temperatures elevades, cosa que els permet una gran eficiència en l’ús de l’aigua (de vegades en tenen prou amb uns 50 o 100 kg d’aigua per produir 1 kg de matèria seca, encara que generalment necessiten de 200 a 300 kg).
Les espècies CAM, com les C4, estan molt esteses a la natura. A més de donar-se entre les crassulàcies, aquesta via metabòlica s’ha trobat en espècies de les famílies de les agavàcies, les aizoàcies, les asclepiadàcies, les bromeliàcies, les cactàcies, les compostes, les cucurbitàcies, les didieràcies, les euforbiàcies, les liliàcies, les orquidàcies, les polipodiàcies, les portulacàcies i les vitàcies. Molt possiblement es dóna també en les geraniàcies, les labiades, les oxalidàcies i les piperàcies, mentre que en algunes espècies de lesfamíles de les butàcies, les cariofil·làcies, les convolvulàcies, les plantaginàcies i les quenopodiàcies ja no és tan clar; en canvi, sembla segur que no es dóna en cap gramínia. La majoria són espècies tropicals suculentes de famílies com agavàcies, bromeliàcies, cactàcies o euforbiàcies, encara que en una mateixa família es poden trobar tant espècies CAM com C4; així, les espècies d’Euphorbia de l’Àfrica tropical presenten la via CAM, mentre que les espècies subtropicals nord-americanes del mateix gènere presenten la via C4.
Tot i que la via CAM sol anar associada amb la suculència, hi ha plantes suculentes, per exemple els halòfits, que no la presenten, i algunes espècies CAM es poden descriure més correctament com a semisuculentes. Algunes espècies, com per exemple les aïzoàcies Mesembryanthemum crystallinum, M. forsskalii i M. nodiflorum, utilitzen la via CAM quan es veuen sotmeses a condicions extremes de sequera o salinitat, però es converteixen en plantes C3 quan les condicions tornen a la normalitat. Encara que les plantes CAM utilitzin l’aigua d’una manera molt més eficient que les C3 i les C4, la seva productivitat és més baixa. També s’ha associat a un creixement lent però, tot i que moltes plantes CAM, com per exemple els cactus baixets i diverses epífites, són efectivament de creixement lent, l’energètica de la fixació del diòxid de carboni demostra que això no ha de ser necessàriament així en totes les plantes que segueixen la via CAM. De fet, les espècies CAM caracteritzen principalment medis poc contrastats amb temperatures nocturnes moderades i humitats relatives nocturnes una mica elevades, tals com els deserts i els subdeserts litorals o les zones de condensació de núvols d’altitud.
Plantes C3 i plantes C4
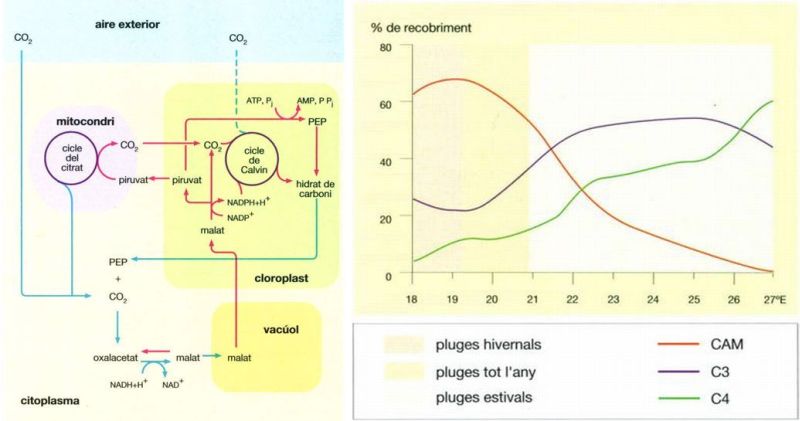
Jordi Corbera, a partir de fonts diverses
Les diferents estratègies vegetals per a fixar el carboni a partir del diòxid de carboni atmosfèric estan adaptades a ambients també diferents. Hi ha plantes que el fixen inicialment, gràcies a la RuDP (ribulosa 1,5 difosfat), en sucres de tres carbonis: són les anomenades plantes C3. Altres, les C4, fabriquen inicialment sucres de quatre carbonis mitjançant l’acceptor PEP i unes darreres, les CAM (a dalt), fixen el CO2 durant la nit (fletxes blaves) i l’emmagatzemen temporalment en forma d’àcids orgànics, principalment malat; durant el dia (fletxes vermelles) continuen la formació de sucres seguint el cicle de Calvin. La via que utilitza cada espècie vegetal depèn en gran part del clima local. L’estratègia CAM té l’avantatge que l’absorció del CO’2 durant la nit, amb els estomes oberts, limita considerablement les pèrdues d’aigua, i és per aquesta raó que és molt comuna entre les plantes dels deserts. Però si bé l’estratègia CAM és perfecta per a ambients on no plou gaire, no tolera temperatures excessivament altes o molt baixes. Per això les plantes CAM sovint es restringeixen a les regions costaneres dels deserts, de clima suavitzat per la presència de la mar. Això es posa de manifest si es representa el percentatge de recobriment vegetal segons cadascuna d’aquestes estratègies (a baix) al llarg d’un transsecte pel sud de l’Àfrica, al nivell dels 30°30’ S, des de la costa occidental fins a una regió interior del continent, a 27°E. Mentre les plantes C3 i C4 constitueixen el recobriment vegetal principal a les regions orientals interiors de pluges d’estiu relativament abundoses, les espècies CAM són molt més abundants a prop de la costa on les pluges són escasses o concentrades a l’hivern.
Les disfuncions del medi físic
Fins aquí s’ha tractat el paper del clima i de la resposta de les plantes en la formació dels deserts. Hom es pot preguntar, però, quin és el paper que hi té el substrat. En realitat, un de ben petit: el substrat contribueix a determinar quin tipus de ecosistema desèrtic es desenvoluparà, però no si en resultarà un desert o algun altre tipus de bioma. La geologia també té una clara influència, però només a escala regional. Com ja s’ha dit, els moviments de l’escorça causants de la formació de les muntanyes han tingut un paper molt important en la creació de molts deserts a les zones d’ombra pluviomètrica.
Les desfavorables condicions del substrat
Tot i que la roca mare no determina si es desenvoluparà un desert, pot modificar la vegetació i determinar quin tipus de desert es formarà. En aquest punt, hi ha una important interacció entre les plantes i el substrat. En altres condicions climàtiques es forma un sòl, de manera que el substrat on les plantes creixen és molt diferent de la roca o el tipus de sediment donats pels processos geològics. El flux de l’aigua i l’activitat animal incorporen residus vegetals al sòl, els quals són modificats per altres animals i microorganismes; les arrels de les plantes i les seves secrecions contribueixen a descompondre les partícules minerals, i el resultat final després de mil·lennis pot ser un gruix d’uns quants metres de sòl sobre de la roca mare, molt diferent d’aquesta tant pel que fa a composició com a estructura. En els deserts, el procés de formació del sòl està molt restringit per l’escàs creixement vegetal; a més, l’escassa biomassa vegetal es tradueix en una també baixa biomassa d’invertebrats i de microorganismes, de gran importància en la formació del sòl i en el desenvolupament de la seva estructura. Així, l’estructura i les propietats dels sòls dels deserts sovint no difereixen gaire dels dels materials parentals subjacents. El contingut de matèria orgànica acostuma a ser inferior a 1%, fins i tot a les capes més superficials, i la poca matèria orgànica present és subjecta a una ràpida descomposició mentre les temperatures són altes. El desenvolupament del sòl es veu frenat fins a nivells imperceptibles, de manera que l’expressió ‘sòl desèrtic’ és gairebé una fal·làcia.
La salinitat és una característica dels substrats desèrtics que sol ser molt limitadora per a l’establiment i el creixement de les plantes. Molts sòls desèrtics, especialment a les depressions, contenen elevades concentracions de sal comuna o de guix provinents de les roques a través de les quals o per sobre de les quals ha passat l’aigua de la pluja, seguida d’una evaporació in situ. Això fa dels deserts, fins i tot sense considerar les adverses condicions climàtiques, uns hàbitats molt inhòspits per a les plantes.
A causa de la seva immaduresa, els sòls dels deserts solen ser inestables i mòbils; són prou conegudes, per exemple, les tempestes de sorra del desert. El vent, sovint fort, aixeca les partícules que encara no s’han consolidat ni han format un sòl veritable, i pot transportar-les a molts quilòmetres. El vent també garbella les partícules, així, la sorra més gruixuda es diposita abans que la més fina a les velocitats del vent més elevades. Una consolidació molt limitada de la superfície del desert, per exemple per una crosta d’algues i líquens, pot ser suficient per a aturar aquesta erosió provocada pel vent i preservar l’incipient sòl. Tanmateix, un cop trencada, per exemple per acció de les peülles d’animals, aquesta escorça esdevé altament vulnerable a l’erosió.
La facilitat de l’erosió geològica
Per altra banda, existeix als deserts, com a tot arreu, una erosió geològica normal, l’ordre de magnitud de la qual és d’entre 0,1 a 1 t/ha i any. Aquest índex pot ser considerablement accelerat per l’acció humana, especialment per una mala gestió del territori i el seu consegüent impacte sobre la vegetació i els sòls. Aquesta acceleració arriba a índexs mitjans anuals de 10 a 700 t/ha i aquests índexs són proporcionals als índexs d’escolament. L’índex d’escolament geològic normal és de l’ordre de l’1%, però pot ser accelerat fins a valors d’entre el 20 i el 80% per les causes vinculades a la gestió del territori, de la vegetació i dels sòls suara evocades. És clar que l’erosió geològica pot ser molt més activa localment, per raons diverses: plegament orogènic, ciclons, solifluxió, colades de lava, colades de fang, pluges torrencials prolongades. Un exemple en zona àrida seria el d’una àrea experimental de la Tunísia meridional, l’escolament mitjà anual de la qual és del 14 al 25% i les pèrdues anuals de sòl de 8,2 t/ha per a una precipitació mitjana anual de 150 mm; s’hi ha mesurat, sobre sòl llimós amb un pendent del 5%, arran d’una pluja de 250 mm en 26 hores (de probabilitat centenària, és a dir que se’n pot produir una per segle), un escolament del 80% i una erosió de 39 t/ha en un dia.
La coberta vegetal contra l’acció erosiva de les pluges
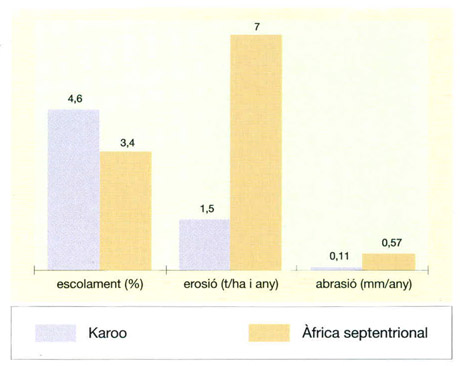
IDEM, a partir de dades proporcionades per l’autor
Els estudis comparatius sobre l’erosió i l’abrasió en dues zones àrides africanes de condicions climàtiques força similars demostren que regions de característiques ecològiques força similars poden patir efectes erosius molt diferents segons com estiguin gestionades. Les dues zones són el Karoo, a l’Àfrica meridional, i les estepes semidesèrtiques de la regió septentrional del continent. L’índex d’escolament és un 35% més elevat al Karoo que a les estepes nord-africanes, però l’erosió hídrica és 4,6 vegades més elevada en aquesta darrera zona. Al Karoo pràcticament no hi ha desertització, mentre que a les estepes de l’Àfrica septentrional, l’erosió progressa prop del 0,7% anual. Com que ambdues zones reben un volum de precipitacions molt semblant, entre 100 i 400 mm anuals, aquest fet es pot explicar per les diferències en la coberta vegetal, que protegeix de l’acció erosiva de les pluges. Al Karoo, la coberta vegetal perenne cobreix entre el 20 i el 30% del territori i la biomassa perenne és de 1 000-1 500 kg de matèria seca/ha. A més a més, al Karoo no hi ha pràcticament conreus de secà (que permeten una infiltració molt més gran), ja que han estat prohibits per sota la isohieta dels 400 mm, excepte sobre sòls de més d’1,2 m de profunditat. A les estepes nord-africanes, en canvi, el 50% del territori ha estat artigat pels conreus de cereals, que, d’altra banda, donen un volum de collites poc predicible. El recobriment i la biomassa perennes, inicialment comparables als del Karoo, arriben només al 5% i a 250-300 kg matèria seca/ha, respectivament.
La indefensió davant de les erosions hídrica i eòlica
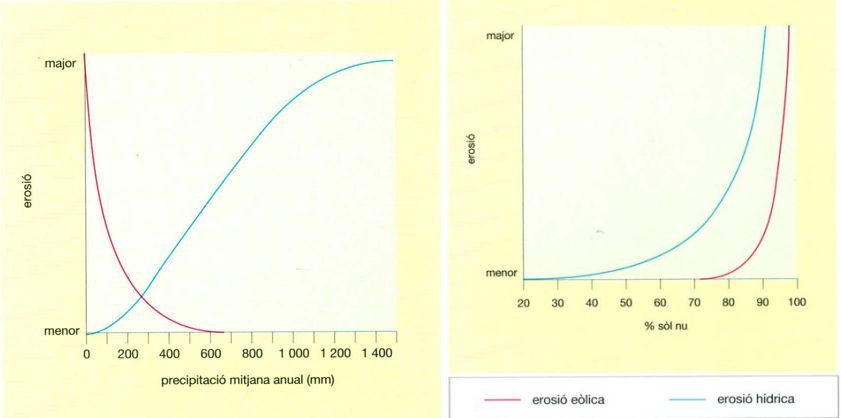
El risc d’erosió, tant eòlica com hídrica, depèn de la precipitació mitjana anual (a dalt) i del grau de cobertura vegetal (a baix). Com és lògic, l’erosió hídrica és directament proporcional a les precipitacions i, per tant, als deserts, on la pluja és tan escassa, semblaria poc important, bé que la natura del substrat i la topografia també hi tenen molt a veure. Si plou poc, gairebé tota l’erosió és eòlica. Als deserts, aquest tipus d’erosió té un paper essencial en la conformació de la geomorfologia i el paisatge. Però així que comença a ploure, l’erosió eòlica disminueix ràpidament, ja que la terra humida no és tan fàcilment enduta pel vent. A més a més, en zones on les pre- cipitacions comencen a ser significatives, creix ràpidament una coberta vegetal que protegeix el substrat del vent i, en menor grau, de l’impacte de les gotes de pluja, l’aixaragallament i l’escolament. Quan les precipitacions són molt elevades, és possible l’existència d’un mantell vegetal important i, per tant, l’erosió eòlica es redueix de manera considerable.
Jordi Corbera, a partir de dades proporcionades per l’autor
L’erosió hídrica és, certament, proporcional a les precipitacions. Així, serà més feble en zones àrides; tanmateix, determinades zones àrides amb formacions geològiques argil·lo-guixenques presenten uns índexs d’abrasió anual de 200-300 t/ha, que combinen l’aixaragallament i l’erosió per escolament superficial concentrat o difús, amb l’erosió per galeries resultat de l’escolament subterrani.
L’erosió hídrica als deserts i subdeserts prové, en gran part, de pluges torrencials. S’ha d’apreciar en dues escales espacials diferents: l’escala concreta de la parcel·la de terreny (d’unes dimensions que poden anar d’algunes àrees a algunes hectàrees) i la de les conques (d’extensions que van d’algunes centenes a uns quants milers de quilòmetres quadrats). Els índexs són inversament proporcionals a les dimensions de les superfícies. Hi ha prop d’un ordre de magnitud de diferència entre els valors obtinguts al nivell de la parcel·la i els obtinguts al nivell de la conca vessant. Aquesta erosió al nivell de la parcel·la pot ser avaluada per l’equació universal de pèrdua de sòls per erosió, de Wischmeier (USLE) que és el producte de 5 factors, en principi independents: el clima, el tipus de sòl, la topografia, la coberta vegetal i les pràctiques de conservació. Es poden definir altres índexs, com l’erosió climàtica, que correspondria a l’escolament anual, és a dir el quocient entre l’escolament mitjà anual (E) i la pluviositat mitjana anual (P). Pel que fa a la conca l’índex de referència és el de Fournier: p2/P on p és la pluviositat del mes més plujós, i P la precipitació mitjana anual. A les zones àrides del Sàhara septentrional els índexs d’erosió hídrica de 10 a 20 t/ha i any (cosa que equivaldria a l’abrasió anual d’un gruix imaginari de 0,7 a 1,4 mm) són habituals.
L’erosió eòlica caracteritza els subdeserts i sobretot els deserts; és habitualment negligible quan la pluviositat ultrapassa els 600 mm, mentre als veritables deserts la seva influència és decisiva en la geomorfologia. Hi ha poca informació, i només puntual, sobre l’erosió eòlica. Han estat ressenyats índexs de 150 a 300 t/ha i any a la Tunísia meridional (i també a Mongòlia, en deserts freds), a conseqüència de l’artigament d’estepes subdesèrtiques sorrenques. Com l’erosió hídrica, l’erosió eòlica resulta de la denudació de la superfície del sòl. Existeix un valor crític, el 25% de recobriment permanent del cobricel pel qual el dipòsit eòlic compensa la deflació.