La llum motriu que ve del Sol

Algunes unitats bàsiques i les seves respectives dimensions (M, masca; L, longitud; T, temps). Una de les unitats més intuïtives és la caloria.
Dades elaborades per l’autor
Hi ha un equivalent mecànic de la calor i de la radiació electromagnètica. Tot treball mecànic comporta una certa producció de calor que es difon de mil maneres; una part d’aquesta calor mai no és totalment recuperable. Aquesta partida perduda ha donat lloc al concepte d’entropia, que és una manera de comptabilitzar aquella mena d’impost que es paga en tots els bescanvis energètics, i que depèn de la temperatura a la qual es fa el bescanvi.
La vida comporta canvis materials continus que requereixen energia. La biosfera s’organitza per transportar energia allí on faci falta. Recordem la circulació en les plantes i els moviments dels animals. La guerra del Golf Pèrsic de 1990-91 proporciona un exemple extraordinari del que significa poder concentrar materials i recursos en un lloc precís. Aquest seria un exemple del control ràpid de dalt a baix que exerceixen els carnívors o els organitzadors en els ecosistemes, i que contrasta amb el control de baix a dalt, que parteix dels que ingressen l’energia, dels productors primaris. És com si tant els arbres com els hostes dels paràsits tinguessin la benevolència de cedir nodriment a altres éssers. La falta de recursos bàsics des del principi, com en un desert, seria un altre exemple de control de baix a dalt.
Algunes de les especulacions que s’han fet sobre l’origen de la vida accepten la disponibilitat inicial i local de molècules orgàniques de gran energia, com les del petroli, les quals haurien estat produïdes per síntesi abiòtica o prebiòtica. Ara, però, i molt probablement també des del principi, la vida depèn de l’ús d’energia electromagnètica radiant, procedent del Sol. És una energia prou concentrada i que viatja per l’espai sense pèrdues, atesa la naturalesa immaterial dels fotons.
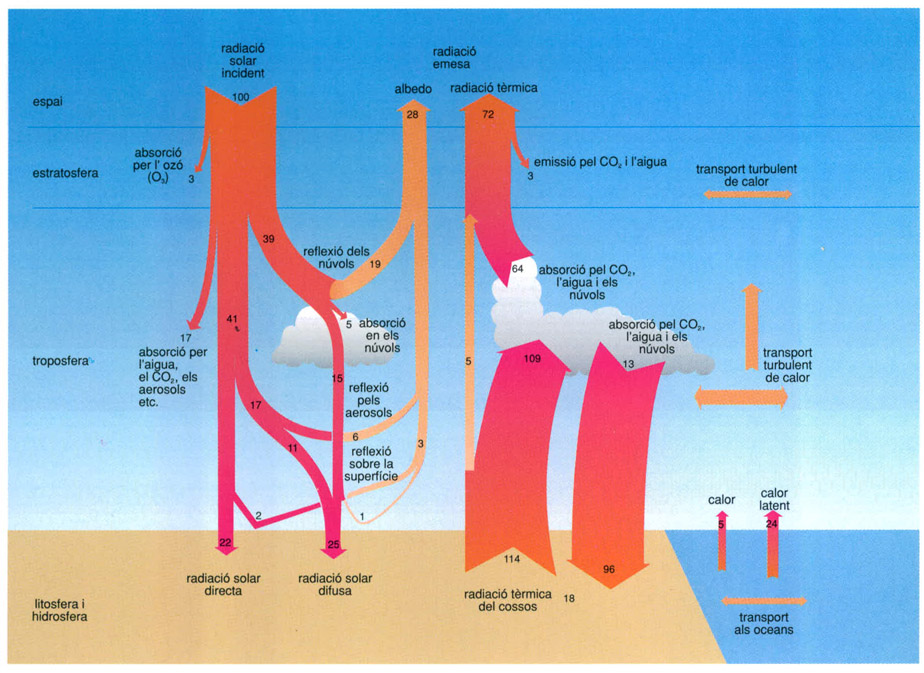
Radiació rebuda del Sol comparada amb radiació emesa per la Terra.
Biopunt, a partir de Raschke, 1989
Hi ha una relació entre la qualitat de l’energia i la mida d’una màquina que pot obtenir-ne treball. Així, l’energia de poca qualitat, com la hidràulica, demana instal·lacions molt grans. Els organismes, en canvi, són miniaturitzats, organitzats a una escala molt fina, i llur funcionament necessita energies d’alta qualitat, com l’electromagnètica i la dels enllaços químics. L’energia química té limitacions perquè ha de circular damunt d’un suport material. Així, hom s’explica l’adopció primària de la radiació electromagnètica, un fet que lliga la biosfera a la superfície de la Terra, que és exposada d’una manera natural a aquella radiació. El fet que la disponibilitat de radiació sigui intermitent com a conseqüència de la rotació de la Terra té una importància indubtable, encara que secundària. Altres organismes depenen dels productors primaris típics, i poden situar-se en llocs on no arriba directament la radiació solar, però on condueix una via material, viva o no viva.
L’energia radiant es converteix en altres formes d’energia que són les que identifiquem en el funcionament de la biosfera. En aquesta conversió es passa dels fotons a les probabilitats de desplaçaments d’electrons, els quals, al seu torn, fan més o menys probable que certes molècules intervinguin en determinades reaccions químiques. L’energia química dels components dels ecosistemes se sol mesurar cremant completament els materials en presència d’oxigen en un aparell anomenat calorímetre, i mesurant la calor produïda, en calories, per l’augment de la temperatura de l’aigua que envolta la cambra de combustió. Aquest és l’origen de diverses xifres que hom emprarà. Quan es tracta d’avaluar transferències o resultats de processos, hom parla d’eficiència, entesa com el quocient entre el resultat obtingut i el resultat esperat; l’eficiència és calculable com un quocient energètic i dóna, per tant, una relació senzilla i sense dimensions.
El verd vegetal de la vida autònoma
La captació de l’energia
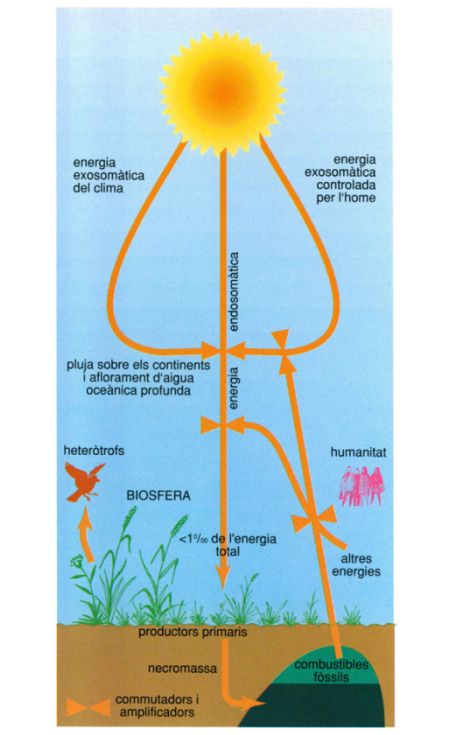
En el funcionament de la biosfera no solament intervé l’energia lluminosa aprofitada inicialment pels productors primaris i més tard, amb l’aliment, pels consumidors; energia que circula pels ecosistemes en forma de llum i per vies químiques d’un organisme a un altre i que s’anomena endosomàtica. També hi ha una part no negligible de l’energia solar que opera en vies exteriors als organismes, però que té un paper força important en el funcionalisme ecològic: és l’energia que anomenem exosomàtica. La circulació de corrents marins i de pertorbacions atmosfèriques, la pluja o els afloraments, són alguns dels fenòmens que fan possible aquesta energia. Una part d’ella, canalitzada pels humans, actualment té un paper prou important.
Biopunt, a partir d’un original de Ramon Margalef
El receptor-detector vivent de l’energia solar consisteix en una mena d’“antenes” els elements essencials de les quals són molècules que absorbeixen radiacions d’una longitud d’ona definida; tenen color, són pigments. Els fotons absorbits determinen canvis que afecten els electrons en el receptor, amb la qual cosa l’energia associada amb certs enllaços varia. Seguidament es produeix una migració d’electrons vers altres molècules, on poden ocórrer canvis químics, i finalment els pigments tornen a quedar en condició de rebre nous fotons. Quan els organismes no cerquen energia sinó orientació respecte de la llum, els detectors no són fets per treballar amb gran intensitat sinó amb la màxima delicadesa (òrgans visuals).
En la captació de l’energia del Sol per les plantes es repeteix a diverses escales una modalitat de construcció laminar. A l’escala màxima, la coberta vegetal s’estén com un estrat relativament prim (uns 100 m de gruix màxim) que, comparat amb el radi de la Terra, ve a ser com un tel finíssim (100 m: 6 380 000 m = 1 / 63 800), seu de l’activitat bàsica que fa rutllar la biosfera. A l’escala següent, la de les fulles (1-10 cm), tenim estructures també òbviament laminars. Dins les cèl·lules foliars especialitzades hi ha els cloroplasts,en forma de discos de pocs micròmetres de diàmetre. En el seu interior es reconeixen sovint unitats menors o grànuls (grana), de 0,3 a 0,5 µm de diàmetre. I a un nivell entre les molècules i els cloroplasts hom troba uns orgànuls laminars, els tilacoides, en forma de bosses aplanades, la membrana contínua dels quals té menys de 6 nm de gruix (0,006 µm) i dóna suport a estructures moleculars força complexes i engruixides. Aquestes estructures moleculars no són uniformes; hom distingeix, almenys, entre les que corresponen als dos sistemes fotosintetitzadors, el fotosistemaI i el fotosistemaII. Cada unitat conté un gran nombre de proteïnes i els pigments que capten la radiació del Sol.
Els pigments
L’estudi de l’aparell fotosintetitzador progressa ràpidament, i aquí sols hi ha lloc per a recordar, de manera molt simplificada, algunes característiques d’un interès funcional i evolutiu especial. Els pigments, les molècules que absorbeixen la llum, pertanyen a tres grups diferents.
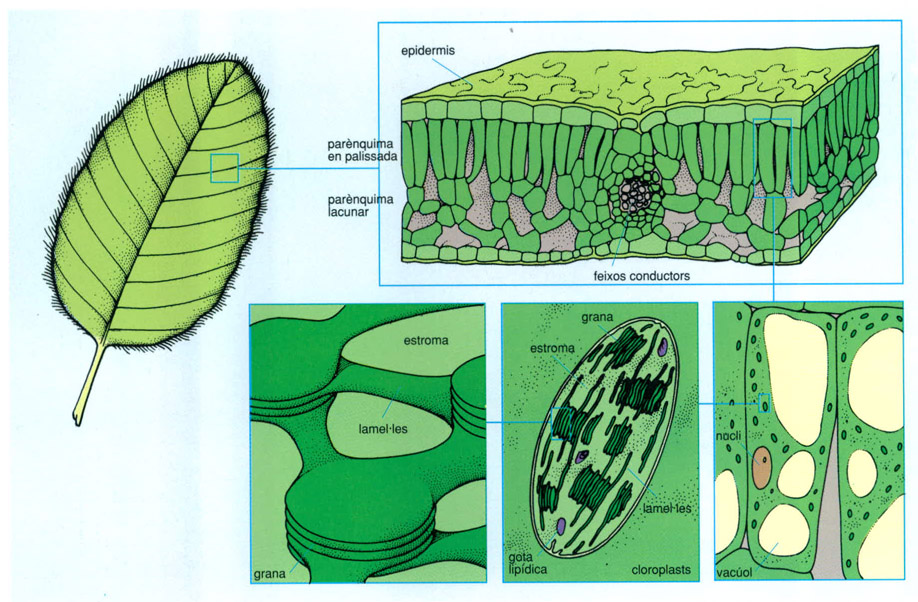
Els sistemes de captació de l’energia solar pels productors primaris repeteixen a diferents escales una estructura laminar. A escala macroscòpica la fulla té forma de làmina. A sota l’epidermis de l’anvers de la fulla es troba el parènquima en palissada, en forma de làmina, que conté les cèl·lules més actives en la fotosíntesi. A escala cel·lular, els cloroplasts solen tenir la forma de petits discos d’uns pocs micròmetres de diàmetre. A escala ultrastructural, els tilacoides formen bosses planes que donen suport a les estructures moleculars actives en la fotosíntesi.
Biopunt, a partir de fonts diverses
En el primer s’inclouen les clorofil·les,la molècula de les quals consta de quatre grups pirròlics (amb nitrogen) disposats en forma d’anell entorn d’un àtom de magnesi, i amb una cua constituïda per un alcohol; les clorofil·les tenen dos màxims d’absorció: un entorn dels 430 nm, i l’altre vers els 665 nm, i això fa que es vegin de color verd (el color corresponent a la llum que no absorbeixen, de longituds d’ona intermèdies). Hi ha diverses modalitats de clorofil·les, a, b i c, i encara d’altres.
En segon lloc, les ficobilines,les quals es podrien comparar amb molècules de clorofil·la obertes, més lineals que circulars i associades amb proteïnes. Les ficobilines absorbeixen precisament les radiacions de longitud d’ona compresa entre els màxims d’absorció de les clorofil·les, és a dir, cap a la zona dels 470-520 nm; per això el seu color aparent és blavós o bé porpra.
En tercer lloc tenim els carotenoides,derivats d’hidrocarburs de cadena llarga, els quals absorbeixen radiacions d’ona relativament curta (blava), vers els 400-480 nm, cosa que fa que el seu color aparent sigui el complementari del blau, és a dir, entre el groc i el taronja.
L’absorció de la llum
La llum travessa l’atmosfera amb poca pèrdua relativa, i les absorcions més importants són en el segment ultraviolat (UV) de l’espectre (segment fortament absorbit per l’oxigen i per l’ozó atmosfèrics) i en el segment de l’infraroig (IR), que és absorbit pels gasos d’efecte hivernacle, entre ells el CO2 i el vapor d’aigua. Altres fenòmens contribueixen a modificar la qualitat de la llum: una fracció creixent de la llum esdevé difusa (deixa de ser de trajectòries aproximadament paral·leles) i una part important es polaritza, la qual cosa vol dir que els plans de vibració dels diversos raigs adquireixen alguna orientació preferent. Però alguns d’aquests canvis no són intranscendents en absolut; per exemple, els cloroplasts estan fets per utilitzar llum difusa, i si haguessin evolucionat per emprar llum de trajectòries paral·leles, possiblement llur eficiènca hauria pogut esdevenir considerablement més alta. Molts animals poden percebre el pla de polarització de la llum, aptitud que utilitzen per a orientar-se amb relació ala llum del cel, fins i tot quan no veuen directament el Sol.
La radiació electromagnètica és fortament absorbida per l’aigua, la qual té la màxima transparència cap a la banda que correspon a la llum verdosa blavosa (entorn dels 510 nm). En canvi, en l’aire fa poca diferència la posició d’una fulla, però unes fulles fan ombra a les altres. Les plantes tenen tendència a créixer cap amunt i la selecció resultant determina la progressiva evolució en alçada dels arbres.
És més rigorós el condicionament que deriva de la forta absorció de la llum per l’aigua. Hi ha un mètode molt simple de mesurar aquesta absorció, basat en la fondària fins a la qual es veu un disc blanc que hom baixa lligat a l’extrem d’una corda (disc de Secchi). Multiplicant per 2,2 la distància fins a la qual es veu aquest disc, hom obté el nivell aproximat en el qual la llum representa l’1% de la llum a la superfície, i en la pràctica hom té definit així el límit inferior de la vida vegetal, encara que algunes algues de fons aconsegueixen viure a més de 200 m de fondària en aigües particularment transparents. El rècord de la profunditat de visió del disc de Secchi en la Mediterrània (oriental) és 53 m; les mars més fèrtils tenen una fondària de visió sovint entre 5 i 20 m.
L’aprofitament òptim de la llum en el medi aquàtic demanaria concentrar la clorofil·la (uns 450 mg m-2) just en la superfície del líquid. Així s’esdevé en els tels verds que es veuen de vegades en bassals, i també en la vegetació de llentilles d’aigua (Lemna) i de jacint d’aigua (Eichhornia). Aquestes catifes vegetals esgoten ràpidament el nodriment local i solament es poden mantenir en aigües fertilitzants just en la superfície i amb poca agitació, condicions que són pròpies de les llacunes associades amb rius. Són típiques d’aquestes situacions les plantes de fulles flotants més impressionants (Victoria, a l’Amèrica del Sud).
Els mecanismes de la fotosíntesi
La unitat de cada “antena” prové del fet de tenir un mecanisme comú de transducció d’energia. El conjunt es podria comparar amb un embut que tingués una sortida limitada, representada pel que hom anomena centre de reacció, en el qual es manifesta l’energia captada. D’una manera simplificada hom pot dir que aquesta manifestació consisteix a donar poder reductor a la cèl·lula, és a dir, capacitat per a moure electrons i, precisament, per a afegir electrons a alguns elements químics. El mecanisme passa a través dels fosfats de dinucleòtids (nicotinamida + adenina) de la piridina (NADP + NADPH2), però és més complicat que això. El resultat final és la reducció d’elements químics que, en l’entorn, hom acostuma a trobar en forma oxidada, com el carboni, el nitrogen i el sofre. Aquests elements químics en forma reduïda s’integren en el substrat de la vida, i, més tard i a la fi, tornaran a oxidar-se i passaran de nou a l’entorn. Aquesta via de retorn ara és causa de preocupació, ja que s’hi afegeixen les oxidacions suplementàries (combustions) degudes a la nostra civilització, que donen tant CO2, amplificador de l’efecte hivernacle, com compostos de sofre i de nitrogen, responsables de les pluges àcides.
L’energia i els fotons
Cada “antena” comporta entre 200 i 900 molècules de clorofil·la, i qualsevol fotó que hi incideixi es mou pràcticament sense pèrdua d’energia fins anar a raure al centre de reacció. Però les molècules de clorofil·la no són equivalents. La probabilitat esbiaixada de transport es deu a la tendència general a canviar radiació d’ona curta (equivalent a fotons més petits i que porten més energia) per radiació d’ona més llarga (corresponent a fotons més grossos i que porten menys energia).
L’energia E per quant la radiació és igual a la constant de Planck (h = 4,14 × 10-15 eV · S) multiplicada per la freqüència (igual a la velocitat de la llum dividida per la longitud d’ona). Així, per a la radiació roja de 668 nm l’energia d’un quant de radiació, d’un fotó, en electrovolts, és de 4,14 × 10-15 × 299 792 458 m s-1 / 668 × 10-9 = 1,858 eV (= 2,97 × 10-12 ergs). Si hom suposa una radiació d’ona més curta, vers el blau violat, per exemple de 445 nm, l’energia per fotó serà de 2,787 eV.
Hom disposa actualment d’aparells capaços de comptar els quants de radiació, i és molt freqüent expressar la radiació en einsteins (E). Un einstein de fotons és un mol de fotons, és a dir, el nombre d’Avogadro de fotons, igual a 6,0220943 ×1023 fotons. L’energia d’un einstein de fotons de longitud d’ona 1 és igual a 119 610 083 J dividits per la longitud d’ona en nanòmetres (nm). Multiplicant el nombre d’einsteins per segon, per l’energia que els correspon segons la longitud d’ona, obtenim l’energia en vats (W). Així, un E (einstein) per segon de radiació de 500 nm és igual a 239 220 W. Es comprèn, per tant, que la unitat pràctica serà el microeinstein, µE. Per a llum de 500 nm, 1 W = 4,18 µEs-1. Tot això pot resultar feixuc, però cal deixar anotats, per una vegada, valors precisos d’algunes relacions que són importants en ecologia planetària i de les quals hom haurà de fer ús repetidament.
La fluorescència
Si la disminució de l’energia associada a un fotó correspon a l’allargament de la longitud d’ona, es comprèn que la tendència entròpica natural sigui que àtoms i molècules retornin, en general, radiació de longitud d’ona més llarga de la que reben. Això és la fluorescència. Els fotons han de rodar camí de les molècules que absorbeixen a longitud d’ona més llarga. I això passa en cada “antena” receptora, de manera que els centres de reacció són funcionalment més propers a molècules de clorofil·la que absorbeixen fotons associats a una longitud d’ona més llarga. Per això, a l’hora d’avaluar l’energia en la fotosíntesi, resulta acceptable comptar els fotons en lloc de mesurar exactament l’energia que porten. Actualment s’usen molt els comptadors de fotons, que donen el resultat en einsteins i no en energia. Hom limita la finestra de la seva sensibilitat, mitjançant filtres adients, a la faixa de la radiació fotosintèticament efectiva (PAR o “photosynthetically active radiation”), aproximadament entre 380 i 750 nm.
Si hi ha massa llum, part de la radiació electromagnètica no s’usa, ja que els processos que ocorren en el centre de reacció limiten el flux. Cada unitat es comporta com un embut que recull pluja que cau i que, si és excessiva, vessa per damunt la vora, de manera que no tota l’aigua que ha plogut se’n va pel broc. L’excés de llum escalfa els teixits i els pot danyar, o produeix un augment excessiu de la respiració, o l’energia és emesa en forma de radiació d’ona més llarga, en un efecte de fluorescència.
Les manifestacions de la fluorescència són importants en la biologia de la producció: els líquens contenen substàncies fortament fluorescents que absorbeixen radiació de longitud d’ona molt curta i emeten radiació d’ona més llarga, la qual tal vegada s’aprofita per a la fotosíntesi de les algues simbionts. Una cosa semblant ocorre en els coralls, en els quals la transmissió d’energia útil de la part animal a les algues simbionts és realment significativa. La fluorescència és un mitjà de transferir energia entre els diferents sistemes fotosintetitzadors dins d’una planta, i concretament del sistema I al sistema II.
Els fotosistemes
Els diferents sistemes fotosintetitzadors d’una planta reben, en efecte, el nom de fotosistemes. L’anomenat fotosistema II ha adquirit una funció preeminent i nodreix amb un sobrant de fotons el fotosistema I, el qual en general absorbeix a longitud d’ona més llarga. El fotosistema I conté principalment clorofil·la a; el fotosistema II també en conté com a pigment majoritari, peró té proporcions més altes d’altres clorofil·les i de carotinoides. L’absorció efectiva de la radiació i la seva reemissió en forma de fluorescència són, com és natural, complementàries. Si s’obstaculitza la fotosíntesi a escala bioquímica per mitjà d’algun recurs adient (per exemple, amb una substància, DCMU, que és també o principalment apreciada com a herbicida) l’emissió fluorescent resulta intensificada, i aquest és un fenomen que s’ha intentat fer servir per a mesurar la capacitat de fer fotosíntesi.
Qualsevol sistema fotosintetitzador genera una diferència de potencial que farà moure els electrons entre uns valors determinats de reduccióoxidació en acoblar-se a la màquina bioquímica. El fotosistema I funciona entre -0,6 i +0,4 volts i, en la cèl·lula vegetal, la seva funció quantitativament més important és la reducció del CO2. El fotosistema II actua entre -0,2 i +0,8 V i usa l’energia aportada pels fotons per a descompondre l’aigua, l’hidrogen de la qual intervé en les reaccions alimentades pel fotosistema I, mentre que l’oxigen queda lliure. Si l’entorn és relativament reductor, n’hi ha prou amb el fotosistema I, que és l’únic existent en organismes que viuen en ambients amb sulfhídric o amb molta matèria orgànica, com ara els bacteris fotosintetitzadors. És versemblant que aquest fotosistema fos l’únic existent en uns temps en què bona part de l’entorn era reductor, abans que es formés una atmosfera oxidant, atmosfera que es produí per la mateixa acció dels organismes. En el curs de l’evolució s’hi afegí el fotosistema II, i ambdós, funcionalment enllaçats com en una cursa de relleus, poden moure electrons sobre una banda més ampla, que abasta pràcticament un volt i mig. La tensió de les piles seques usuals coincideix amb el voltatge a què treballa la vida.
L’acoblament entre tots dos fotosistemes encara té altres aspectes d’interès. L’“antena” del fotosistema II és entre 2 i 4,5 vegades més gran que en el fotosistema I: és natural que aquell capti fotons en excés i pugui transferir energia dintre la cèl·lula al fotosistema I per fluorescència. Això s’entén i s’adiu amb la diferent composició dels pigments. El fotosistema II comporta una fracció més gran de carotinoides i, en general, de pigments que absorbeixen a una longitud d’ona més curta, i, per tant, de fotons amb més energia, part de la qual es transfereix fàcilment per fluorescència a l’altre fotosistema. El resultat és que el fotosistema II sembla com si estigués adaptat a emprar una lluminositat més baixa (“antenes” més extenses), i alhora adquireix una posició preeminent, ja que ell sol és suficient per a subministrar energia a allò que queda del fotosistema I.
Cadascun dels fotosistemes mou electrons sobre una diferència de potencial d’un volt aproximadament, però els dos fotosistemes combinats solament cobreixen un àmbit d’aproximadament un volt i mig. Hi ha una superposició en aquest joc de relleus. Això, d’una banda es podria veure com un mecanisme de seguretat, però a més l’excés d’energia troba una altra aplicació en la màquina bioquímica de la cèl·lula: en efecte, té a veure amb l’emmagatzematge d’energia i l’assimilació de fòsfor (pas d’ADP a ATP).
Plantes C3 i plantes C4/
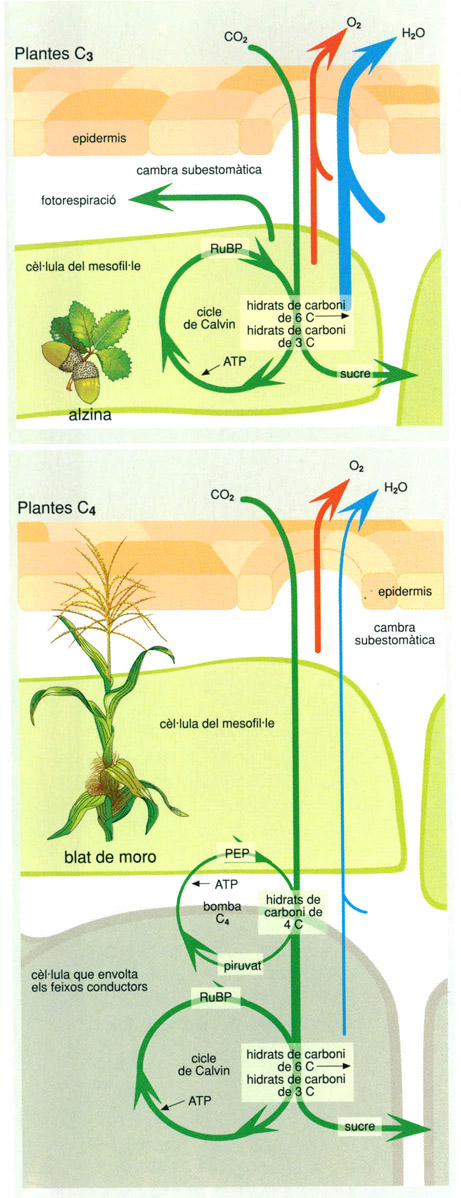
Hi ha diferents maneres de fixar el CO2 atmosfèric: la de les anomenades plantes C3, que són la majoria de les plantes forestals de la zona temperada; i la de les anomenades plantes C4, que són moltes fanerògames d’origen tropical.
La fotosíntesi de les plantes C3 condueix a la formació inicial de molècules de carbohidrats de sis àtoms de carboni, les quals s’escindeixen aviat en dues molècules d’àcid 3-fosfoglicèric, de tres carbonis cada una; per això aquest sistema de fixar del CO2, incorporat per la ribulosa 1,5 difosfat (RuDP), s’anomena C3.
Les plantes C4, entre les quals hi ha el blat de moro i la canya de sucre, tenen un rendiment fotosintètic extraordinari fins i tot quan el CO2 escasseja. En aquestes plantes l’acceptor del CO2 és el fosfoenolpiruvat (PEP), i els carbohidrats que es formen tenen quatre àtoms de carboni en comptes de tres; primer es forma àcid oxalacètic i després àcid màlic; es regenera el piruvat i queda lliure, altre cop, el CO2, el qual inicia un cicle similar al de Calvin. Aquestes plantes, que reben el nom de C4, solen presentar diferències anatòmiques, ja que tenen els feixos vasculars de les fulles envoltats d’una capa de cèllules on es concentren els cloroplasts. La bomba C4 fa que es concentrin molècules de CO2 prop del centre actiu d’un enzim crucial per a la fotosíntesi, la ribulosa difostat-carboxilasa, amb la qual cosa s’impedeix que s’hi fixi l’oxigen i, per tant, s’eviten pèrdues d’energia per fotorespiració.
L’esquema és fet per Editrònica, a partir de Bazzaz i Fajer, 1992.
La fotosíntesi biosfèrica
No és objecte d’aquesta obra presentar la complicada estructura i la funció de la maquinària bioquímica de la cèl·lula. Però podem entendre que l’entrada d’energia que fa funcionar tota la màquina de la biosfera és fonamental.
La fotosíntesi es limita a les parts de la biosfera situades a una altura relativa més gran, que contrasta amb la distribució, menys localitzada, o més generalitzada, de la respiració. Com a resultat d’aquest contrast es genera un gradient vertical en el valor del potencial d’oxidació-reducció. La llum dóna capacitat de reducció a la part alta de la biosfera; més avall predomina de manera general la respiració sobre la fotosíntesi, i la respiració normalment implica els d’oxigen. Així, hi ha un flux d’electrons per dintre de la biosfera que va de dalt a baix, com un corrent elèctric, entre un pol superior, negatiu, i l’inferior, positiu. Aquest corrent queda compensat per fora dels organismes, excepte quan s’acumula molt material químicament reduït en el sòl o en el sediment. Un altre cas de gradients elèctrics verticals, molt més intensos i més erràtics, es dóna en els núvols i entre ells i la terra, i es fan prou evidents en les tempestes i en els llampecs.
El rendiment fotosintètic
La constant solar en el límit de l’atmosfera és de 139 mW cm-2 (Vegeu també L’atmosfera). La part aprofitable per a les plantes (PAR) s’apropa a la meitat d’aquella energia i la superfície de la Terra és quatre vegades l’extensió del seu cercle màxim, de manera que la vida vegetal pot disposar, de mitjana, d’uns 172 W m-2. Hom hauria d’ajustar, localment i per a cada cas, aquest valor segons la latitud i l’estació, sense comptar altres factors més imprevisibles i inconstants, com ara l’absorció i la reflexió que té lloc als núvols.
En fer extraccions dels pigments de les fulles o del plàncton per mitjà de dissolvents orgànics (ordinàriament metanol o acetona) es veu que una dissolució que conté 450 mg de clorofil·la estesa en un estrat uniforme sobre una superfície d’1 m2, absorbeix pràcticament el 99% de la radiació visible incident. Si la clorofil·la i els altres pigments no estan dissolts, sinó més compactats en estructures petites, com s’esdevé en les fulles, hi pot haver més pigment amb el mateix efecte global.
El plàncton vegetal no arriba gairebé mai a posar més de mig gram de clorofil·la per metre quadrat d’extensió. Aquesta és també la concentració referida a l’extensió de les fulles, i per damunt d’aquesta concentració ja no augmenta el rendiment. Potser en són una excepció les fulles que funcionen per les dues cares, com per exemple les dels arbres molt verds de l’àrea mediterrània, com l’alzina i altres acompanyants, que tenen concentracions de clorofil·la més elevades. Com que en la vegetació terrestre l’extensió total de les fulles és unes 4 vegades més gran que l’extensió del sòl sobre el qual es projecten, hi ha més clorofil·la de la que semblaria estrictament necessària.
La producció primària global
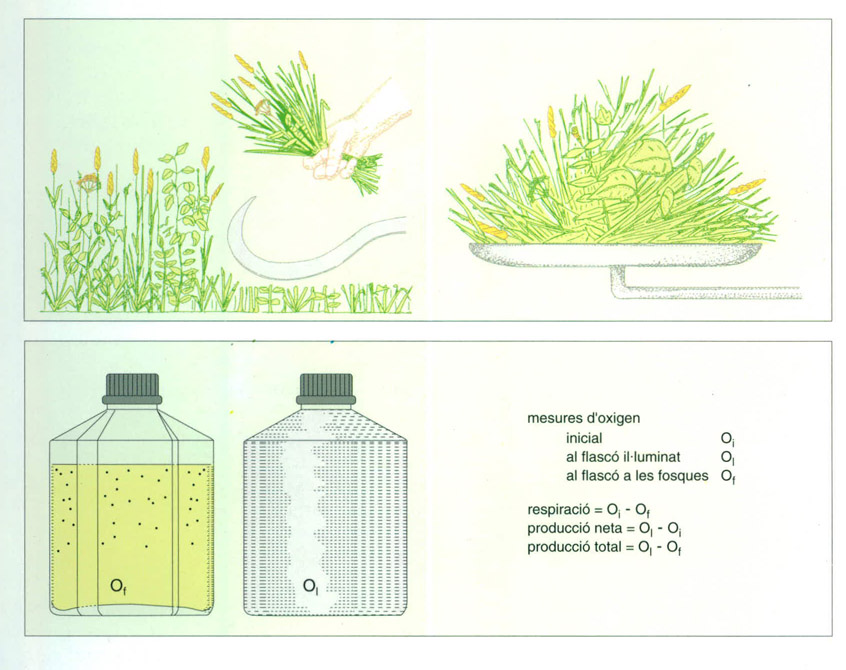
La producció primària real de la vegetació s’estima de maneres molt diverses: la més senzilla és segar i pesar la collita i avaluar-ne el contingut energètic en el calorímetre. Hom també pot avaluar la producció per l’oxigen produït en el bescanvi, o seguint la velocitat d’assimilació de carboni usant l’isòtop radioactiu 14C. Es mesura la respiració de les plantes i la intensificació de la respiració que pot acompanyar la fotosíntesi.
Biopunt, a partir de dades elaborades per l’autor
Com a reacció fotoquímica que és, la fotosíntesi resulta relativament poc accelerada davant de qualsevol augment de temperatura; però una temperatura més alta, que sempre intensifica la respiració, facilita sovint un aprofitament més ràpid dels nutrients. Un augment de 10°C duplica aproximadament la velocitat d’una reacció o redueix a la meitat el temps necessari per a acomplir un procés. Una temperatura fluctuant té un efecte accelerador més gran que el que correspondria a la seva mitjana aritmètica.
La mesura de la producció primària més simple possible de la vegetacició terrestre és la que consisteix a segar i pesar la collita d’un prat o d’un conreu herbaci; té l’inconvenient que no es coneix la tracció de la producció consumida en la respiració. Amb el plàncton, mitjançant mesures de l’oxigen, es poden obtenir dades més precises. N’hi ha prou d’omplir dos flascons idèntics, l’un transparent i l’altre opac, amb la mateixa suspensió de plàncton en aigua; mesurar-ne el contingut d’oxigen inicial (Oi) i al cap d’un temps el contingut d’oxigen del tascó transparent, que ha rebut llum solar (Ol) i del flascó opac, que ha romàs a les fosques (Of). Unes senzilles restes permeten estimar la producció total (Ol - Of), la respiració (Oi - Of) i la producció neta (Ol - Oi).
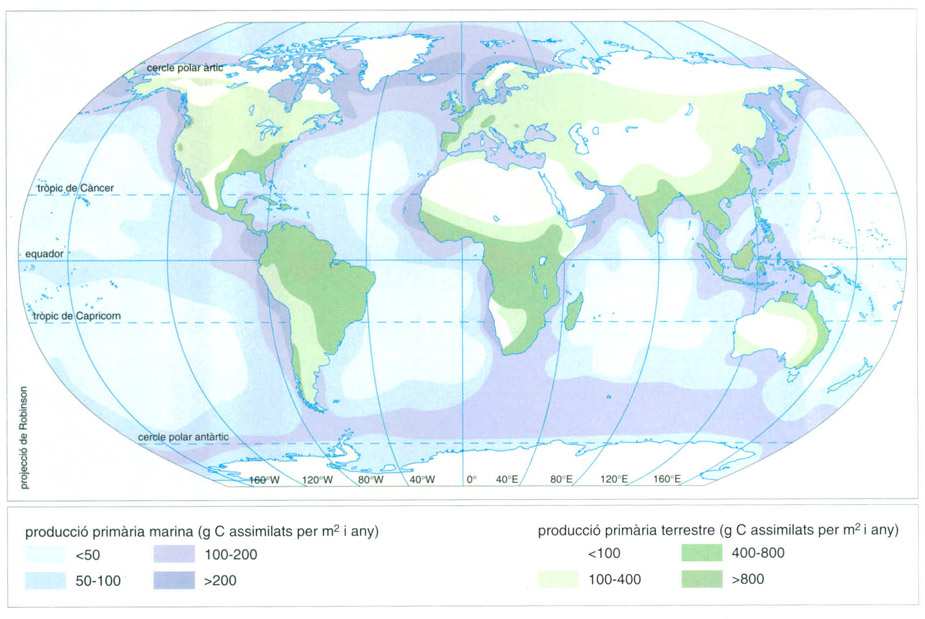
Distribució de la producció primària, expressada en grams de carboni assimilats per metre quadrat i any, als oceans i als continents.
Editrònica, a partir de fonts diverses
Una fracció de l’energia absorbida en la fotosíntesi s’usa per a reduir compostos de nitrogen i de sofre, i una altra fracció dóna lloc a l’energització de compostos de fòsfor. Però el canvi que correspon a l’assimilació del carboni és el més important, i sovint en parlar de producció es fa referència exclusiva o més directa al carboni. Així, les produccions primàries mitjanes aproximades i integrades en un període anual són de prop de 300 g de carboni per metre quadrat en terra i de prop de 100 en els oceans. La producció global de la biosfera seria entorn de 78 × 109 tones de carboni per any, que equivalen a 114 × 1012 W.
Les xifres que hom repeteix en els llibres es basen en mesures en bona part bastant antigues, massa poc nombroses, i sovint interpretables de maneres no sempre coincidents. Considerant l’interès que té saber realment per on anem, seria hora de reprendre amb més entusiasme la tasca d’obtenir xifres noves, emprant procediments que corresponguin a l’estat actual dels coneixements i dels recursos tècnics. La visió remota des de l’espai ens pot il·lustrar sobre la distribució dels pigments, però no ens dóna molta informació sobre la seva activitat.
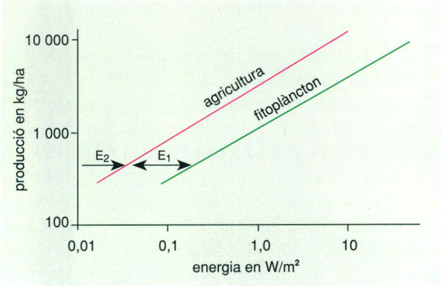
L’energia exosomàtica intervé en la producció, com l’endosomàtica. Això és particularment palès en el cas dels sistemes en l’explotació dels quals els humans intervenen activament, com són els conreus on s’esmerça energia en llaurar, adobar, regar, collir i totes les activitats involucrades en l’agricultura, però succeeix també en el cas del plàncton, que rep energia exosomàtica en forma d’onades, corrents, afloraments, etc., que faciliten la barreja. En el cas de la producció agrícola no s’ha comptabilitzat, per manca de dades, l’energia de l’evapo-transpiració de les plantes; si s’hagués comptabilitzat, com caldria, resultaria més gran (superior a l’esmerçada pel fitoplàncton) l’energia total esmerçada per a una producció donada.
Editrònica, a partir de fonts diverses
Si l’energia total que arriba del Sol és de 178 × 1015 W, i si fem cas de les xifres de producció esmentades unes línies enrere, és clar que solament el 0,6‰ aproximadament d’aquella energia reapareix en la producció primària. Aquesta és una xifra més propera a la producció primària neta, avaluada al nivell de les plantes, de manera que hom ha d’afegir-hi una estimació de la respiració de les mateixes plantes. No cal anar massa enllà per a concloure que amb prop de l’1‰ de l’energia que ens arriba del Sol n’hi ha prou per a fer funcionar la biosfera. Aquesta energia es qualifica d’endosomàtica, ja que circula per dins els organismes. Tota l’energia restant, tant la no visible com la de longitud d’ona adient, però no aprofitada, no es pot donar per perduda, des del punt de vista de la biosfera, ja que configura el clima i, per tant, fa habitable la Terra. Serà considerada com energia externa o exosomàtica per als organismes.
El canvi energètic que es pot associar a la reducció del carboni (l’energia que el carboni o els seus compostos donen en cremar-se) és de 116-144 kcal (entre 485 668 i 602 900 J) per mol (12 g de C); 8 einsteins de fotons d’una longitud d’ona entorn de la màxima absorció de la clorofil·la (668 nm) representen una energia de 8 × 42,6 kcal (= 14 268 614 J). La conversió de llum en energia química es fa amb una eficiència de menys del 50%. Energèticament, tres fotons serien suficients per a reduir un àtom de carboni i produir una molècula d’oxigen. En realitat, en calen vuit (hi ha valors experimentals entre sis i deu). Hom no pot dir que aquesta relativa ineficiència sigui important, tal com funciona la vida. La selecció natural ha donat preferència a l’acumulació d’estructura sobre un aprofitament de l’energia més eficient.
Dins dels organismes, la maquinària química de la vida va mobilitzant i degradant a poc a poc l’energia assimilada a través de cicles bioquímics més estesos, que afavoreixen l’estabilitat i l’organització més que no pas una mobilització ràpida de l’energia. La vida esmerça prudentment l’energia acumulada en els enllaços químics: s’usa en la síntesi de molècules més complexes que formaran part de la mateixa matèria viva, sota la guia dels mecanismes cel·lulars de transmissió d’informació (DNA → RNA → enzims → substrats). La via de degradació i de cessió d’energia és inversa. Característicament, la mobilització d’energia passa per introduir molècules de sucre en un cicle regular de canvis en el qual intervenen compostos fosforats (el fosfat és sempre un bon vehicle d’energia) i així l’energia es fa utilitzable d’una manera gradual i relativament lenta.
La producció i la productivitat
La síntesi de nova matèria orgànica, i fins i tot de matèria orgànica viva, és comparable al rèdit que dóna un capital. El capital es podria comparar amb la biomassa i el rèdit amb la producció primària. Hom fa una distinció entre producció primària bruta i producció primària neta. Ambdós conceptes queden clars si es fan experiments per mesurar la producció a partir del bescanvi d’oxigen.
La manera més senzilla de mesurar la producció és a partir d’una suspensió d’algues unicel·lulars, sigui de plàncton natural, sigui d’un cultiu de laboratori. L’experiment és més favorable si es tracta d’un cultiu axènic o pur d’alguna espècie d’alga, ja que el plàncton natural conté molts bacteris que respiren i, per tant, usen part de la matèria orgànica produïda i “embruten” l’experiment: si a l’ampolla tenim heteròtrofs, és a dir, bacteris o animals, el resultat serà falsejat, ja que el que tindrem serà el bescanvi d’un segment més ampli d’ecosistema, i no solament el corresponent als productors primaris.
Si, en un altre tipus d’experiment, seguim l’assimilació amb carboni radioactiu (14C), resulta encara més difícil treure’n l’entrellat, ja que sempre una fracció del 14C (per exemple, del respirat) va al líquid i en part es recicla. L’altra manera, més òbvia, i per on hauríem d’haver començat, és estimar la producció primària a partir de l’augment net de massa, és a dir, de la collita, que és com es fa amb el producte dels conreus; si seguíssim aquest procediment en el cas de les algues, segurament negligiríem una petita part del material produït, que les algues cedeixen a l’entorn en forma de material orgànic soluble. Hom no pot atendre mai tanta complexitat.
S’ha dit que la producció primària (P) és com el rèdit que dóna una biomassa (B), la qual és comparable a un capital. És natural, doncs, que en ecologia s’hagi usat molt el quocient P/B emprant unitats adients (pes total, pes sec, carboni; o bé energia, com per exemple calories o joules i aleshores la productivitat té la forma de watts dividits per joules); el quocient P/B té la dimensió d’un invers del temps, una velocitat. El seu invers, B/P, és el temps mitjà que triga a renovar-se la biomassa (“turnover time”). És clar que els valors derivats de la biomassa total o calculats amb referència a l’energia no tenen per què coincidir: pensem què passa si en el desenvolupament de la població augmenta el valor energètic per unitat de massa del material acumulat. En català i en llengües de construcció equivalent, productivitat s’aplica a la producció dependent d’algun dels factors de producció; un factor molt important, potser el més important de tots, és la biomassa. P/B és una productivitat.
Hom pot parlar també de productivitat amb relació a altres factors externs de producció com la intensitat de la llum, la temperatura, etc., i interns, com ara la concentració de clorofil·la. Ja que la clorofil·la i els pigments fotosintetitzadors en general són bons d’avaluar, la relació producció/pigments és una de les més emprades, generalment en la forma de grams de carboni orgànic, que són assimilats en una hora per cada gram de clorofil·la. Aquesta xifra té un valor teòric màxim de 25, un valor pràctic màxim d’11, i els valors alts, sovint observats, queden entre 3 i 4; aquests es donen en condicions favorables de llum (més de 70 µE m-2 s-1). Sota una llum menys intensa, aquell factor es fa aproximadament proporcional al logaritme de la il·luminació.
La supeditació a la llum i als nutrients
La disponibilitat i la utilització de la llum influeix molt sobre la disposició de les fulles i l’organització de les plantes terrestres. És significatiu que la concentració de la clorofil·la a les fulles correspongui a la quantitat necessària per a una utilització plena, i també que la vegetació terrestre tendeixi a tenir un índex foliar (quocient entre l’expansió horitzontal de les fulles i la superfície del sòl sobre la qual es projecta la vegetació) proper a 4, encara que sovint hom troba valors més alts, principalment en arbres corpulents. Evidentment hi ha molt més pigment del necessari, cosa que no ocorre en la vegetació de molt poca alçada, com ara els líquens, les algues, les hepàtiques i algunes molses. Això fa creure que l’estructura esponjada d’un bosc permet una optimació de l’ús de la llum, ja que solament una fracció de la llum incident (potser 1/3) és absorbida per la fulla mentre que la resta és reflectida, i una petita part, la travessa; bona part d’aquesta llum seria finalment absorbida per altres fulles. A més sempre queden entre el fullam escletxes a través de les quals es projecta una imatge circular del Sol, i els moviments de branques i fulles sota el vent, no solament acceleren el bescanvi gasós, sinó que també tenen alguna incidència en l’ús de la llum. Un àlber (Populusalba)és una imatge suggeridora, però fins ara no ha produït més que algunes consideracions sobre la mecànica del moviment de les fulles. Ja que 4 és la superfície d’una esfera de cercle màxim 1, hom podria veure en aquella relació una expressió estadística més general de condicions d’optimació en la interacció de la radiació, i podria ser una interpretació acceptable de l’índex foliar més comú, que és proper a 4.
L’extensió i la disposició de les fulles crea un clima determinat de llum dins el bosc i unes espècies s’ajusten o se subordinen a les altres en haver de compartir la llum disponible. Entre els vegetals no falta la competència, que es manifesta especialment en la tendència a créixer d’alçada de les plantes que acaben essent les dominants. Tot això es combina amb el condicionament de la disponibilitat i el transport vertical de l’aigua procedent del sòl pel tronc, a través de les arrels. Els botànics han interpretat que tota l’organització de les plantes superiors, dels macròfits, és orientada principalment cap al transport i l’economia de l’aigua, cosa que és certa solament fins a un cert punt.
La productivitat també depèn, com és obvi, de la concentració de nutrients necessaris (carboni, nitrogen, fòsfor, sofre, etc.). Si falta algun nutrient, o es troba a una concentració molt baixa, sovint sembla que hi hagi una assimilació excessiva de carboni, la qual es reconeix en les estructures vegetatives i, en el cas de les algues, sovint en l’excreció de mucílags. És habitual relacionar la velocitat d’assimilació amb la concentració d’algun nutrient limitant [Cs].
És natural que totes aquestes complicacions de la fisiologia s’hagin de tenir en consideració en ecologia. Però no ens han de desviar excessivament dels objectius més generals, els quals sovint toleren aproximacions força grolleres. Tot i així, no hem d’oblidar la necessitat de millorar-les. Recordem que l’alternació entre períodes de llum i períodes de foscor sotmet la fisiologia de l’assimilació vegetal a un ritme molt marcat, l’estudi precís del qual té un gran interès.
L’índex foliar
Editrònica, a partir d’un original de Ramon Margalef
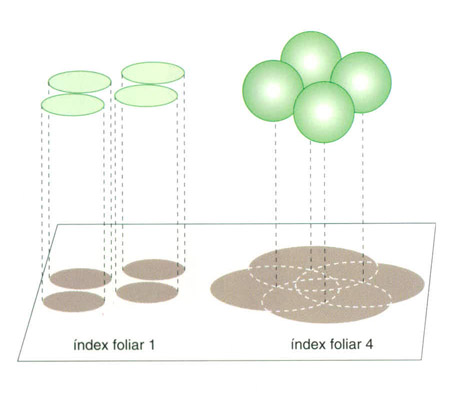
A les plantes terrestres l’índex foliar, és a dir, la relació entre l’extensió total de les fulles i l’extensió del sòl sobre el qual es projecten, té generalment un valor proper a 4. Això equivaldria a una superfície esfèrica el radi de la qual fos igual al d’un cercle que tingués la mateixa extensió que la projecció de les capçades.
A la figura adjunta s’esquematitzen geomètricament les situacions cor-responents a uns índexs fo-liars 1 i 4 respectivament. Amb un índex foliar 1, que seria la situació d’una capçada en la qual no hi hagués cap mena de superposició ni encavallament de les fulles, l’extensió dels cercles que representen la superfície de la capçada i de la seva projecció és idèntica. Amb un índex foliar 4, tot i que la projecció seria la mateixa (la dels cercles màxims, que tenen el mateix radi que els cercles del primer cas), el desenvolupament de cada superfície esfèrica té una extensió 4 vegades més gran (E = 4π2).
L’estimació de la producció i la productivitat
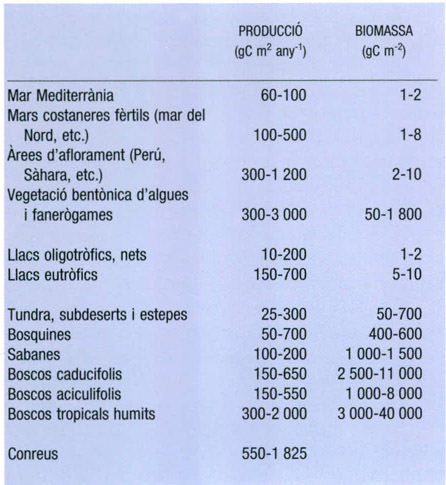
Estimacions de la producció primària de diferents ecosistemes del món expressades en grams de carboni assimilat per metre quadrat i any. A partir del Programa Biològic Internacional es començaren a aplegar dades a escala global sobre la producció primària de diferents biomes. Cal advertir que, tot i que algunes d’aquestes xifres, si no la majoria, són força criticables, sí que donen una idea comparativa dels ordres de valors en diferents àrees del món. En els casos en què és prudent de fer-ho, s’indica també la biomassa mitjana dels productors primers considerats, en grams de carboni per metre quadrat.
Dades elaborades per l’autor
Actualment hom està fent un esforç per deduir la productivitat i interpretar les condicions de producció a través d’imatges obtingudes des de l’espai. Hom ha d’interpretar el significat de la reflexió en longituds d’ona que es poden atribuir a la clorofil·la i a d’altres pigments. Però en la mar la concentració màxima de clorofil·la sovint es troba entre 60 i 100 m de fondària i no és visible. En ecologia terrestre es prescindeix massa sovint de combinar la interpretació del color amb detalls texturals que podrien indicar, per exemple, l’alçada i la mida de gra de la vegetació. Val a dir, però, que la informació és cada vegada més abundant i més correcta, de manera que hom pot creure que ens anem aproximant a un coneixement millor de la distribució de la biomassa vegetal i de la seva producció primària.
És d’actualitat considerar canvis recents i possibles dels valors de la biomassa i la necromassa amb relació a acceleracions o desacceleracions de la productivitat i amb transferències de material (principalment de carboni) cap a d’altres compartiments de la Terra (per exemple, el CO2 de l’atmosfera), i amb possibles modificacions del cicle d’altres elements, com el nitrogen. El nitrogen sol desvetllar més interès que el fòsfor, tot i que aquest és un candidat més raonable al paper de limitant definitiu de la producció planetària.
Si hom porta els diversos valors de producció sobre un mapa, tant per als sistemes terrestres com per als oceànics, veu fàcilment que hi ha trets comuns en les distribucions respectives: generalment hi ha regions, més aviat poc extenses, on la producció primària és molt alta, i àrees d’extensió creixent que corresponen a valors cada vegada més baixos de la producció. Aquesta ordenació pot ser més regular en els oceans que damunt la terra. Per al cas dels oceans, hom ja ha comentat el mecanisme bàsic amb relació a les disponibilitats dels elements limitants: si el funcionament dels ecosistemes marins porta a acumular el nodriment cap al fons, llavors el que controla la producció és el fet de portar de nou vers la superfície, vers la llum, aquestes masses d’aigua; per tant, la producció primària en els oceans apareix fortament correlacionada amb l’energia localment esmerçada en promoure el moviment vertical de l’aigua. En els sistemes terrestres, les relacions, al cap i a la fi, són equivalents: hom sap des de sempre que la productivitat depèn en aquests sistemes de la disponibilitat d’aigua líquida i, per tant, també de ta temperatura; la fracció de l’any durant la qual l’aigua és sòlida constitueix la limitació més seriosa, encara que no ho és tant com la falta o l’escassetat total d’aigua. S’han proposat expressions, derivades empíricament, que donen la producció primària en funció d’aquells descriptors; n’és un exemple l’expressió que apareix en els mapes confegits per Lieth.
Els pagesos saben que calen entre 200 i 500 l d’aigua per a produir 1 kg de gra. Naturalment, hom pot expressar el cost de l’aigua per l’energia implicada en l’evaporació, la pluja i l’evapo-transpiració. Igual que en el cas de la mar, concloem que la producció primària depèn d’una energia externa o exosomàtica que decau o es degrada en l’espai perifèric de la Terra. De manera global, la producció primària depèn d’una potència fraccionària (entorn de 0,6) de l’energia exosomàtica que es degrada per unitat de superfície.
Hom ha donat per als oceans una producció mitjana de 100 g C m-2 any-1, xifra que correspon al plàncton. La vegetació litoral és més productiva per unitat de superfície, però ocupa una extensió molt més petita, i la seva contribució a la producció marina total probablement és inferior al 2%. Caldria fer un lloc a part per als baixos de corall, on els principals productors primaris són les algues que viuen en simbiosi dintre dels animals colonials i són explotades per aquests, de manera que l’eventual producció neta sobrant és extraordinàriament petita amb relació a la producció primària neta de les algues.
En els sistemes terrestres, la producció primària mitjana és unes tres vegades més elevada que en els marins, entorn de 300 g C m-2 any-1. Si hom té en compte l’extensió relativa de la terra i de la mar, es podria dir —i fóra correcte, i també bo de recordar— que la producció total de la Terra es reparteix gairebé equitativament entre els continents i els oceans. El camí mitjà que ha de seguir el nodriment en el seu retorn és molt més llarg en els oceans, que tenen 4 km de fondària mitjana, que no en terra, on la capa de sòl és retativament prima i el transport horitzontal tampoc no és excessiu. A més, la vegetació terrestre (especialment els arbres) controla de manera molt efectiva el camí de l’aigua cap amunt a través d’un sistema ben organitzat de canals. En la regió marina pelàgica els fluxos cap amunt es produeixen en un sistema fluid, no organitzat, de manera que no és estrany que siguin molt menys efectius si hom els compara amb l’energia del clima que hi intervé.
La biosfera deu haver funcionat segons aquestes línies des del seu començament, ja que no sembla que els mecanismes bàsics hagin variat gaire en el curs de l’evolució. Sembla talment com si la natura, si se’ns permet personalitzar-la, hagués intensificat més la riquesa de formes i la diversitat que no pas la simple quantitat i la capacitat de desenvolupar treball físic. De tota manera, hom no pot excloure la possibilitat que hagin ocorregut variacions importants. Hi ha indicis que durant les èpoques glacials la producció primària de tota la biosfera fou més baixa, potser entre 3/4 i 2/3 de l’actual, però no sembla que hi hagi hagut canvis gaire més grans que aquest.
El valor dels valors
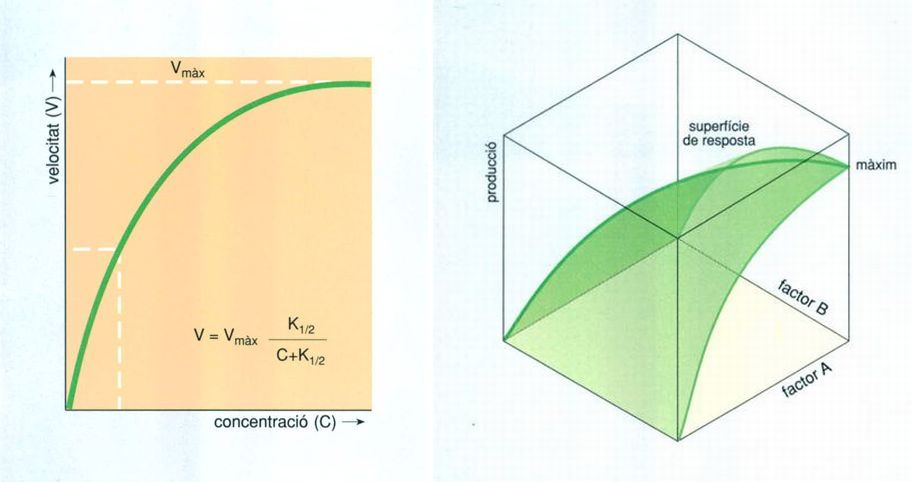
Biopunt, amb dades elaborades per l’autor
En ecologia, els processos sovint s’han referit als valors màxims permesos (velocitat màxima, població màxima, etc.). Lògicament, és un procediment dubtós, però, a la pràctica es continua emprant amb un èxit relatiu. En el gràfic de l’esquerra veiem una aplicació de l’equació de Michaelis-Menten, que relaciona la velocitat d’un procés (V) i la concentració del factor que el limita, per exemple la concentració d’un nutrient (C). K1/2 és la concentració a la qual la velocitat és la meitat de la màxima. Es poden construir models on s’interfereixen diversos factors limitants, tal com es mostra en el gràfic de la dreta.
Menjar: servitud animal de la vida subsidiària
La vida heterotròfica
No cal recórrer a hipòtesis complicades per entendre com es pogué originar la vida dependent o heterotròfica: n’hi ha prou amb pensar que en un règim en què alternen el dia i la nit, la continuïtat de la vida durant la nit s’havia de basar en materials sintetitzats durant el dia, i si aquests materials permetien la continuïtat de la vida a qui els sintetitzava i després els utilitzava, també eren una constant temptació a emprendre l’evolució del règim de vida heterotròfic pur en organismes que perderen o renunciaren a la capacitat de captar, emmagatzemar i utilitzar l’energia solar.
L’osmotròfia i la fagotròfia
Hi ha dues maneres diferents d’exercir la vida heterotròfica. Una, l’osmotròfia, és usar molècules presents en l’entorn, per exemple dissoltes en l’aigua i que poden travessar la membrana que envolta la superfície de la cèl·lula; ho fan els heteròtrofs osmòtrofs, representats principalment pels bacteris i els fongs. L’altra, la fagotròfia, té a veure amb el fet que la concentració de matèria orgànica assequible es manté alta si es colonitza l’interior d’altres organismes (paràsits), o bé si es confinen temporalment dins d’un sistema digestiu pertanyent a organismes o parts d’organismes aliens, de manera que llur descomposició dóna una solució altament concentrada en tota mena de molècules adients; això és menjar, tal com ho inventaren els heteròtrofs fagòtrofs, que són pròpiament els animals.
Els avantatges de la fagotròfia
El règim de vida fagòtrof permet especialitzar-se en diverses direccions. La varietat de conformacions funcionals es manifesta en l’evolució de dos sistemes que van associats, encara que menys estretament del que es podria sospitar: l’evolució dels dispositius de detecció, persecució i captura, i l’evolució paral·lela, menys espectacular o si més no menys visible i potser no tan extremada, dels mecanismes químics d’atac, descomposició i utilització dels materials ingerits. El desenvolupament de pinsos molt complets i equilibrats, especialment per a la cria de l’aviram, ha permès utilitzar-los per mantenir vius, en parcs zoològics, animals tan exòtics com la gallina (originària del SW d’Àsia); això prova que sovint són els mètodes d’obtenir aliment més que no pas els de digerir-lo el que es troba a la base del problema de la inserció d’una espècie animal en el seu entorn.
Des del punt de vista de l’evolució, és evident que el joc que dóna l’alimentació fagotròfica permet seguir i acumular informació per moltes vies divergents: l’enorme varietat de la vida animal, enfront dels vegetals, n’és testimoni. S’han donat noms a un nombre d’espècies de plantes, animals i microorganismes que, en conjunt, excedeix potser d’1,4 milions i se suposa que n’hi ha per descobrir un nombre igual o més elevat. Però la suma de les plantes fanerògames (220 000 espècies) i altres vegetals autòtrofs (algues) no són més que el 25% del total, de manera que la diversificació de la vida ha fet més camí entre els heteròtrofs que no entre els autòtrofs, malgrat que els darrers representen la forma inicial i autònoma de la vida. Les espècies de fongs i de liquens són legió. Es pot creure que entre els animals hi ha hagut més estímul, i que s’ha fet més camí en les múltiples possibles direccions d’evolució. Ara es fa molt ús d’aquesta mena d’estadístiques i de consideracions, especialment per a comentar, amb més o menys compunció, la pérdua de la diversitat biòtica en el nostre món.
La naturalesa dels aliments
Els aliments subvenen tant a les necessitats plàstiques com energètiques. Comporten un ingrés d’energia i de matèria a l’organisme, però també un ingrés d’informació: la funció de les vitamines i els casos de simbiosi serien exemples d’aquest valor afegit dels aliments ingerits.
L’energia alimentària
En els nivells de producció més enllà de la producció primària, la vida usa energia emmagatzemada en enllaços químics relativament làbils; no serveixen, per exemple, els enllaços dels metalls o els de la matèria cristal·lina. L’energia que pot proporcionar l’oxidació total d’un mol de matèria orgànica corrent és de l’ordre de 2 a 8 × 105 J, que són 0,5 - 2 × 102 kcal. Un gram de carboni representa al voltant de 10 kcal, xifra que resulta fàcil de recordar a l’hora de fer estimacions aproximades. Compostos rics d’energia, com l’ATP (trifosfat d’adenosina), i que són moneda corrent en el metabolisme dels organismes no donen més de 40 o 50 kcal/mol, en condicions fisiològiques normals.
El carboni comprèn aproximadament el 10% del pes de la matèria viva, de manera que 100 g de pes fresc total contenen entre 40 i 160 kcal utilitzables, com algú podrà recordar de les taules presentades sovint amb finalitats dietètiques. Materials poc hidratats (fusta, nèctar, pol·len, cera, resina, llavors, pèls i plomes) donen valors fins a 240-600 kcal per 100 g de pes fresc; resulten de mal digerir, però aquells que aconsegueixen digerir-los en tenen el monopoli.
Solament en la seva forma reduïda, el carboni de les molècules orgàniques pot adoptar les infinites variants dels esquelets moleculars. Les substàncies en aquesta forma reduïda es poden oxidar combinant-se amb l’oxigen de l’aire. Per respiració o per combustió carreguem l’atmosfera amb el compost més oxidat del carboni (CO2). Paral·lelament es formen compostos oxidats de nitrogen i de sofre.
La incorporació d’informació biològica viva: simbiosi
En casos singularitzats es pot acceptar informació biològica viva, quan és suficientment distinta perquè sigui inofensiva. A vegades, com en els virus, solament aconsegueix fer-se copiar. O bé resulta tolerable el seu funcionament continuat de manera paral·lela, i això ha estat l’origen de moltes simbiosis, com algues o bacteris que viuen dins de cèl·lules d’altres organismes (coralls i ciliats amb algues endosimbiòtiques).
Ja des del principi de segle, alguns investigadors pensen que l’adopció i assimilació d’organismes aliens o d’algunes de les seves parts, ha estat un procés repetit en el curs de l’evolució, que ha permès compondre la cèl·lula eucariòtica i accelerar l’evolució biològica. La major part dels organismes es podrien considerar monstres, a l’estil del mític monstre del doctor Frankenstein, és a dir, fets de peces creades per la vida segons línies independents i que els atzars de les trobades i l’interès mutu han permès de combinar. També es poden assimilar òrgans complexos i pràcticament desproveïts d’informació biològica, i per tant no reproduïbles, com les càpsules urticants dels celenterats, que algunes espècies de mol·luscs marins nus agafen impunement sense descarregarles i les utilitzen de nou en defensa pròpia.
El cas de les vitamines
El tema del nivell màxim de complicació i de pes molecular que poden atènyer les molècules que s’assimilen va lligat a la qüestió de les vitamines. Es donà el nom de vitamina a substàncies de mida molecular mitjana, ni massa senzilles ni tampoc excessivament complicades, que són produïdes per diversos organismes i que altres continuen necessitant, després d’haver perdut la capacitat de sintetizar-les. El resultat és que els segons organismes arriben a ser dependents d’aquells primers, capaços de sintetitzar-les. La interpretació biològica i ecològica és senzilla. En bacteriologia i en micologia experimental, quan s’afegeixen molècules necessàries als cultius, s’aconsegueix fàcilment de fer sobreviure mutacions gèniques defectuosos, que feien impossible la síntesi d’aquelles molècules. Les nissages amb aquesta mancança poden continuar propagant-se en medis enriquits amb les substàncies necessitades.
Hi ha ecosistemes molt complexos que contenen en la seva matriu o entorn físic, d’una manera habitual, una munió de molècules relativament complicades, la presència de les quals permet la supervivència de castes d’altres espècies que les necessiten, però que havien perdut la capacitat de produir-les. És clar que alguns canvis importants que tinguin com a resultat un subministrament defectuós de vitamines poden determinar que les espècies que necessiten vitamines es trobin en inferioritat i fins puguin desaparèixer. Es natural trobar més espècies amb necessitats vitamíniques en els ecosistemes que les poden oferir, és a dir, rics d’espècies i que no han experimentat pertorbacions gaire freqüents. Sembla obvi que, per a l’home, el neolític, caracteritzat segurament per l’adopció d’una vida més sedentària i una alimentació més rutinària, basada en un espectre sobtadament limitat de productes del conreu, anà acompanyada d’una epidèmia d’avitaminosis variades. Ara, al contrari, vivim l’aprofitament de vies metabòliques naturals per contaminants que no fan gens de falta per al funcionament del cos, sinó tot al contrari; un exemple clar són molts productes i derivats de la nostra indústria, o bé metalls que se solubilitzen i mobilitzen a través dels canvis incessants (acidificació, per exemple) a què tenim sotmès l’entorn.
El processament alimentari
En l’evolució de la vida es manté la identitat de l’espècie. Els organismes disposen d’una jerarquia precisa de portadors d’informació, la qual, en darrera instància, depèn del DNA nuclear. No admet pertorbacions importants, però conserva prou flexibilitat per manejar reconeixement, acceptació i reaccions, com veiem que passa contínuament en el sistema immunitari. Els aliments es degraden per acció enzimàtica en peces prou petites, portadores d’energia o aprofitables com a peces de construcció, però que ja no presenten el risc d’introduir una informació aliena en els sistemes genètics.
El valor alimentari dels glúcids i dels lípids
Els glúcids, hidrats de carboni o sucres són aliments típicament energètics. Es tracta de molècules relativament poc específiques, que contenen àtoms de carboni, sovint disposats en anell, i àtoms d’oxigen i hidrogen en les proporcions de l’aigua. Poden combinar-se molècules elementals per formar-ne d’altres de pes molecular més elevat. Els lípids o greixos contenen els mateixos elements químics, bé que mostren una proporció menor dels components de l’aigua i formen molècules més llargues. Un gram de glúcids dóna, en el calorímetre, 4 100 calories, en canvi usat dins un organisme en dóna una mica menys, 3 850, ja que no s’oxida del tot en una atmosfera d’oxigen, cosa que passa en el calorímetre. Per als lípids hom té, també en el calorímetre i en l’organisme respectivament, 9 300 i 8 800 calories per gram.
La degradació d’aquestes molècules mai no és sobtada, com ho seria si es cremessin en una flama, sinó que es fa progressivament, per passos, i mai no s’arriba al final de tot. La tendència en la vida és disminuir el flux d’energia per unitat de massa. Això es materialitza en els cicles metabòlics, com el de Krebs, que separa l’energia dels sucres seguint una successió de passos químics en què intervenen fosfats; així, es pot passar de l’ADP a l’ATP, que és un dels acumuladors d’energia d’ús més comú en el metabolisme intermediari. Les cadenes de greixos es consumeixen com una candela, des d’una punta (o des de les dues) i amb la mateixa parsimònia. No hi ha perill que la vida es cremi a si mateixa, mentre sigui activa, la qual cosa vol dir que la concentració d’aigua és prou elevada com per a evitar-ho. Pensem només que l’evaporació de l’aigua demana una quantitat de calories que, a temperatures normals, és d’unes 600 calories gram (o aproximadament 2 500 J) per gram d’aigua i que l’aigua representa entre el 70 i el 80% del pes dels organismes. Però la fusta i els teixits vegetals secs contenen menys aigua, cosa que fa que els boscos puguin cremar-se.
El valor alimentari de les proteïnes
Les proteïnes també donen energia, però són més importants com a font de compostos nitrogenats. Així, s’incorporen aminoàcids com a tals, i n’hi ha alguns que molts organismes no sintetitzen i que, per tant, han de ser presos amb els aliments. Després tots ells s’empalmen per formar proteïnes específiques. Ens trobem amb una situació que recorda la de les vitamines. És comprensible aquesta estratègia de la vida: resulta prudent usar com a vehicles de l’energia enllaços que pertanyin a molècules trivials, sense recórrer, amb aquesta finalitat, als enllaços adjacents a àtoms menys banals que els de carboni, oxigen i nitrogen.
Les xarxes tròfiques
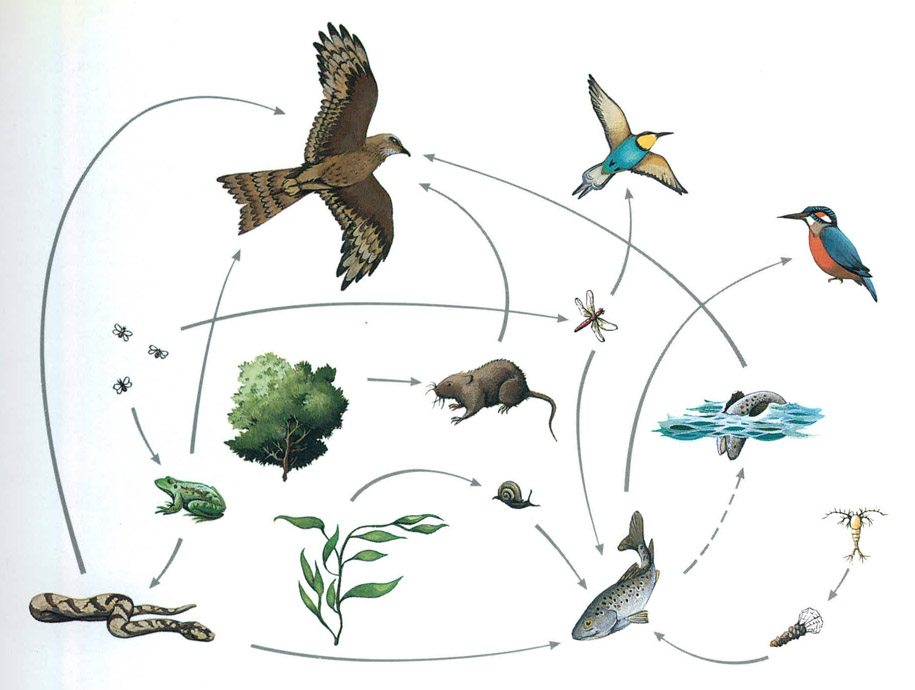
Les relacions tròfiques entre els organismes rarament poden expressar-se com a cadenes simples en les quals successivament cada espècie se’n menja una altra i és menjada per una tercera. Diferents espècies depredadores poden alimentar-se d’una mateixa espècie de presa, o bé poden ser diverses les preses susceptibles de sentir d’aliment a un determinat depredador. Per això, fins i tot en presentacions simplificades com la d’aquesta làmina, que podria correspondre a un rierol de l’Europa mitjana o de la muntanya mediterrània i a les seves ribes, és preferible tractar-la com una xarxa, la xarxa tròfica.
Biopunt
El conjunt de les relacions alimentàries, com la popularment expressada amb la frase “el peix gran es menja el peix xic”, científicament es dignifica, o es fa més pedant, sota el nom de la cadenatròfica o xarxa tròfica, expressió del fet que uns animals es puguin menjar altres animals, en una seqüència que pot incloure diverses etapes, fins a cinc o més. Es poden comptar molts més nivells, especialment si fem un lloc als paràsits i hiperparàsits (o paràsits de paràsits) els quals poden afegir fins a cinc graons més. És natural que els individus d’una espècie depredadora siguin més grossos (i visquin més temps, la qual cosa és tan important o més en biologia) que els individus de les espècies que li serveixen de presa.
Mai no es pot parlar de cadenes tròfiques senzilles, ja que generalment hi ha connexions laterals, en el sentit que dues espècies depredadores poden perseguir una mateixa espècie de presa. O, ben sovint a la inversa: un depredador s’alimentarà de preses distintes. Per aquesta raó, és més apropiat parlar de xarxes tròfiques que de cadenes tròfiques, tot i reconèixer el nombre limitat de nivells. I l’estructura és, com si diguéssim, fibrosa, per una certa tendència a eliminar, com a resultat de la competició, un nombre excessiu de connexions horitzontals a una mateixa alçada. Relacions de canibalisme, o relacions en les quals el depredador i la presa canvien de paper són molt poc eficients i, per tant, escasses o excepcionals.
Els nivells tròfics i el parasitisme
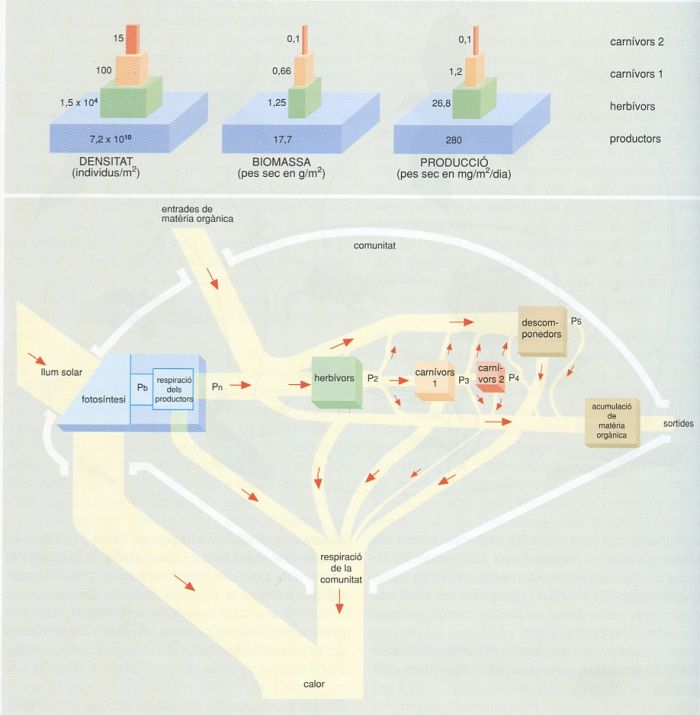
L’ecòleg anglès Charles Elton ideà una representació dels successius nivells de les cadenes tròfiques emprant diferents descriptors (nombre, biomassa, productivitat), en forma de gràfics comparables a piràmides escalonades, com les del gràfic superior. Aquestes representacions, anomenades precisament piràmidesd’Elton, són molt usuals en ecologia. Altres representacions que inclouen els fluxos d’energia, com les proposades per Howard T. Odum, de les quals el gràfic inferior és un exemple, poden ser alhora més subtils i realistes.
Editrònica, a partir de fonts diverses
Ha estat tradició, almenys des dels temps de l’ecòleg Elton, a la dècada dels anys 20, representar propietats quantitatives selectes dels successius nivells tròfics en un ecosistema (vegetarians, carnívors de primer grau, de segon grau, etc.) en forma d’un gràfic que recorda les piràmides de pisos de maies i asteques. En sentit vertical cada pis depèn del pis inferior, i sigui quina sigui la magnitud emprada per a la caracterització, la seva expressió numèrica generalment va disminuint cap al cim de la piràmide. I disminueix més ràpidament la producció o el flux d’energia que no la biomassa, perquè s’escau que els carnívors de nivell més alt tenen unes poblacions que es renoven molt lentament. És a dir, la duració mitjana de la vida dels carnívors acostuma a ser més llarga que la dels vegetarians. I ho és en part perquè al damunt no tenen ningú que els pugui exigir (la qual cosa s’aplica especialment als depredadors del cim); unes poques espècies de vegetarians han seguit la via d’espolsar-se els enemics augmentant enormement de mida, com els elefants, els rinoceronts, els hipopòtams i, indubtablement, diverses castes de dinosaures. La relació segons la qual els subsistemes portadors de més informació són també els que es renoven més lentament, es retroba des dels sistemes interns de la cèl·lula (substrats-enzims-RNA-DNA). Emprant un símil mecànic, cada ecosistema es podria comparar a un joc reductor d’engranatges, en el qual els pinyons giren més de pressa que les respectives rodes dentades de diàmetre més gran, amb les quals s’engranen.
Els segments de les cadenes tròfiques que corresponen als paràsits són una altra cosa, perquè s’hi inverteix la regularitat precedent, ja que els paràsits són menys corpulents i de vida més curta que els seus hostes respectius. És com si la vida parasitària abandonés el control real als hostes. D’altra banda, la condició parasitària és molt diversa. Quan un paràsit intestinal metabolitza o escindeix molècules llargues i en treu prou energia, i l’hoste acaba de degradar els segments mo-leculars que queden, el paràsit queda com integrat en la maquinària metabòlica de l’hoste i pot esdevenir necessari. Aleshores podríem dir que es comporta com un simbiont. Però aquestes qualificacions (paràsit, simbiont) són massa acolorides d’un maniqueisme molt humà que seria bo de bandejar.
Els fluxos energètics
D’una manera general, s’accepta que el flux d’energia que corre a través de nivells tròfics successius queda dividit per un valor pròxim a 10 a cada pas, valor que és comparable a la quantitat de carn de porc obtinguda segons el pes del pinso subministrat. En principi aquesta convergència entorn d’un valor comú és sorprenent, atesa la gran varietat d’organismes implicats en relacions de la mena de les que s’estableixen entre menjador i menjat.
De fet, però, resulta raonable, perquè els animals que mengen de manera automàtica un aliment que pot ser poc nutritiu (els que mengen terra, o fang, o filtren material en suspensió en l’aigua, animals que podríem qualificar com “animals de cullera”) solen treballar de manera molt seguida i usen un material que sovint és poc alimentari. Aquests animals tenen, però, un metabolisme baix, són poc actius, i inverteixen en la reproducció una notable fracció d’allò que han assimilat. Contràriament, els carnívors (animals que podríem definir com “de forquilla”), que poden ser molt actius, digereixen a fons peces de carn força nutritiva, però tenen un metabolisme elevat i es mouen molt, mentre que una part relativament més petita del que assimilen es destina a produir una nova generació. En aquest darrer cas, naturalment, una part més alta de la dieta pot consistir en sucres o greixos, simples subministradors d’energia; animals molt actius, com els borinots o els colibrís, tenen una dieta amb elevada proporció de sucres.
En l’evolució hom troba exemples de nissagues que han tornat enrere en l’evolució tròfica, animals que han passat de ser “de forquilla” a tornar a fer-se “de cullera”, cosa que sovint ha demanat profundes reconstruccions orgàniques, com les que s’intueixen en el cas del bec i la llengua dels flamencs quan es comparen amb els òrgans homòlegs d’altres ocells. En el flamenc, el bec i la llengua són eines que trien molt bé el fang i en separen els bulbil·les, les llavors i els petits crustacis. Una transformació evolutiva semblant s’observa en els cetacis misticets, les balenes veres, que es nodreixen de crustacis, mol·luscs i peixos relativament petits i que han reorganitzat profundament l’aparell bucal, amb l’adquisició dels filtres palatals de lamines còrnies esfilagarsades que constitueixen les anomenades barbes.
Si el flux d’energia es divideix aproximadament per deu a cada nivell, al cinquè pis només quedaria el 10-5 de la producció primària, i aquesta seria una raó, potser la principal encara que no l’única, de la relativa limitació de la llargada de les cadenes tròfiques. La conclusió més immediata és l’exclusió de grans carnívors de les àrees d’extensió limitada: un tigre no podria sobreviure en una illa gaire petita.
Les plantes ofereixen possibilitats molt diverses de vida als organismes fitòfags. Considerem, simplement, la gran quantitat d’insectes que podem trobar adaptats a viure sobre qualsevol espècie de planta. La vida d’un arbre, que pot arribar a durar milers d’anys, amb un grau elevat de conservatisme en el genoma, contrasta amb els milers de generacions d’alguns insectes que habiten damunt d’ell, i que evolucionen a més velocitat. Heus aquí un condicionament a què generalment es fa referència breu en parlar del conservatisme ben justificat dels vegetals.
La pols dels morts
Hom ha dit que moltes de les insuficiències de l’ecologia actual es deuen a la poca afecció que tenen els ecòlegs a fer observacions de nit, sota terra, o en els excrements. El llarg cicle del retorn implica una quantitat de matèria que ha viscut i ja no viu, la necromassa, que excedeix en quantitat la mateixa biomassa. Entretant continua essent portadora d’una quantitat d’energia utilitzable i utilitzada per organismes molt diversos, particularment abundants i diversificats en els sòls i en els sediments. Ja s’ha assenyalat que hi ha dominis crepusculars en els quals la mort no és total, com és ara el representat per la fusta.
El reciclatge total del material mort no va tan ràpid com es podria suposar, ja que en la naturalesa hi ha hagut la pressió d’una selecció que ha afavorit l’aparició de molècules resistents a la descomposició, tal com s’ha esmentat amb relació al sistema de suport i transport dels vegetals superiors. També són força resistents a la descomposició els materials de reserva; són un tresor del qual és prudent guardar la clau (els enzims propis, que permeten utilitzar aquests materials en exclusiva). La cansalada i el llegum en són un bon exemple, i també n’és la manera com l’home els aprolita. També són molècules molt resistents, recalcitrants a la descomposició, les dels missatgers químics, hormones externes o feromones.
Hom es meravella de la distància a la qual es poden comunicar químicament insectes de la mateixa espècie de diferent sexe, encara que molt més rarament es fa notar que, per tal que les molècules actives funcionin, cal que siguin molècules de llarga durada. És exactament el mateix que fa l’home amb els seus antibiòtics i biocides en general; per exemple, és millor que el DDT costi de destruir; i igualment, no convé que una ampolla destinada a contenir líquid sigui biodegradable.
El material mort
La presència i l’eventual acumulació de material mort, o més ben dit, de compostos de carboni químicament reduïts (com correspon als que han estat sintetitzats per la vida) té un origen mixt. D’una banda es deu a la resistència particular dels enllaços químics, sigui en els materials originaris, o bé en les molècules més petites o modificades d’alguna manera, que queden després que s’hagin desfet els enllaços més atacables. D’altra banda, molècules que serien oxidables no s’oxiden perquè van a parar o s’acumulen en llocs on l’oxigen és escàs, com pot ser en els fons anòxics de mars i llacs i en les torberes.
Aquesta necromassa no sols queda implicada en els cicles ecològics, amb algun significat positiu o utilitari en el cas de la fusta, sinó també en altres situacions. Sovint fa de suport a moltes reaccions mentre es descompon lentament, com ara l’anomenat humus del sòl i el dissolt en les aigües naturals. L’humus és un dels grans desconeguts pel que fa a la seva composició química precisa. Amb el nom de virosta, i amb altres denominacions, es fa referència tradicionalment a material mort que conserva una estructura tal que permet reconèixer-ne la procedència. L’humus ja és un pas més enllà, ja que és amorf i no deixa veure, ni al microscopi, el seu origen: la seva composició molecular encara no és prou coneguda.
Una gran part d’aquest material és emprat pels heteròtrofs del sòl i torna a ingressar en l’ecosistema reciclant-s’hi. Podria dir-se el mateix dels components minerals, per exemple el potassi o el fòsfor. El seu reciclatge té lloc en els arbres vius. La pluja renta el potassi acumulat en la superfície de les fulles, però sovint s’assimila, poc després, en les arrels. Un cicle complet i local de la major part dels elements necessaris es dóna més freqüentment en els boscos tropicals humits que en comunitats vegetals situades en pendent o en la vegetació de temporada, ja que aquestes perden elements contínuament i en guanyen d’altres per erosió de les roques immediates o de les situades a nivells més elevats.
La interacció entre la biosfera i la litosfera
L’intercanvi continuat entre la biosfera i la litosfera ha determinat que en la litosfera es produïssin canvis molt més importants dels que s’hi haurien produït si no hi hagués hagut vida; els canvis soferts per les roques carbonatades i la descomposició dels silicats queden sota el control de la biosfera: hom ha remarcat sovint que l’alteració de les roques de la perifèria de la Terra ha anat almenys dues vegades més de pressa (és difícil fer-ne estimacions amb pretensions quantitatives) de com hauria anat si no hi hagués existit vida. Fet i fet, l’accelerada transformació selectiva dels materials sòlids superficials de la litosfera es pot atribuir al manteniment, gràcies a la vida, del gradient vertical d’oxidació-reducció. Les fases finals tant de la descomposició dels materials húmics com de l’alteració de les argiles es fan en condicions de poca energia, la qual cosa permet trobar una gran variabilitat en els productes finals. Humus i argiles estan condemnats a adaptar-se mútuament com a components essencials dels sòls, especialment dels més madurs, als quals donen llurs propietats més interessants.
El destí de l’humus que es troba dissolt en l’aigua podria ser més conegut del que és. Un exemple n’és la matèria que dóna el color bru als rius anomenats “d’aigües negres” al subcontinent sudamericà, i en aigües torboses de tot el món. Hom sap de fa temps que en els oceans hi ha una considerable quantitat de carboni orgànic en solució (sovint entorn d’un mil·ligram de carboni per litre o encara més), intensament fluorescent i esmentat, a vegades, com a Gelbstoff o substància groga. Certament, el seu contingut de carboni 14 ha estat força estudiat amb l’intent de conèixer el temps transcorregut des del moment en què aquests materials foren assimilats per organismes que estaven en equilibri amb la composició isotòpica de l’atmosfera d’aquell temps. Hom ha obtingut xifres sorprenents, de 6 000 anys o més per a nivells profunds d’aigua del Pacífic, i de mig mil·lenni o més per a aigües superficials de l’Atlàntic, el qual sembla molt més jove o biològicament més actiu.
En cada ecosistema, la major part d’aquestes molècules són les que es descontrolen temporalment en els cicles ordinaris i locals dels materials orgànics. Però a més hi ha la sospita que en els oceans hi ha una fracció relativament excessiva de material procedent dels continents, la síntesi del qual depèn molt de la fusta, dels fongs i d’altres materials, que han passat per l’estadi d’humus. Heus aquí un tema de recerca poc fressat i que, certament, té un gran interès amb relació als cicles globals de materials, fonamentalment del carboni.
La destrucció dels sòls comporta l’oxidació d’una part del material húmic que contenen, i en els incendis forestals actuen en el mateix sentit. Hom ha fet avaluacions d’aquests efectes i ha trobat que la destrucció d’humus per desertització, incendis i treball agrícola, pot representar una contribució molt important a l’efecte hivernacle global; és a dir, l’oxidació de tot aquest material fa que augmenti la concentració de CO2 en l’atmosfera. Estimacions relativament recents avaluen aquesta contribució entre 2 i 3 milers de milions de tones de carboni per any, per a la superfície dels continents, la qual cosa representa la meitat dels combustibles fòssils que crema la humanitat.
A tota vela o a tota màquina
L’energia endosomàtica i l’energia exosomàtica
El manteniment de la biosfera depèn de l’u per mil de la radiació solar que arriba al nostre planeta i s’aconsegueix amb concentracions de clorofil·la, que són poc més altes d’un gram per metre quadrat en els sistemes terrestres i en els d’aigua dolça, i que mai no arriben a aquest valor en els oceans. Però aquesta no és tota l’energia de què acabem servint-nos.
En quantificar el funcionament dels ecosistemes, no és correcte comptabilitzar solament l’energia que ingressa a través de la clorofil·la, és a dir, la que segueix una via metabòlica interna: aquesta energia és la que qualifiquem d’energia endosomàtica perquè circula per dins dels organismes, primer de les plantes, i després al llarg de la sèrie d’heteròtrofs. La resta de l’energia que ve del Sol també és important: és l’energia del clima, que fa la Terra habitable i que en defineix les condicions desiguals en les diverses contrades (les quals es poden interpretar com més o menys favorables a la vida en general, o favorables per a uns organismes i menys favorables per a d’altres en particular), de manera que esdevé un factor important a l’hora de definir les diferències regionals; diem que aquesta fracció rellevant de l’energia és energia exosomàtica amb relació als organismes, perquè circula o fa treball per fora del cos.
També és exosomàtica l’energia de les marees, que no és electromagnètica en el seu origen sinó gravitacional. És exosomàtica l’energia que barreja verticalment l’aigua marina i fa pujar cap a la superfície elements (fòsfor, nitrogen, silici) que estaven més concentrats en aigües profundes, cosa que assegura la continuïtat de la producció i de la vida pelàgica. També ho és l’energia associada a la pluja i a l’evapo-transpiració que es produeix en les fulles, la qual té una funció anàloga, en el sentit de fer arribar elements necessaris (fòsfor, nitrogen, potassi, etc.) al nivell il·luminat on té lloc la fotosíntesi.Tant en un exemple com en l’altre, l’energia exosomàtica és quantificable i ve a ser entre 20 i 50 vegades més gran que l’energia endosomàtica associada. Com en qualsevol màquina, la temperatura local condiciona l’aprofitament de l’energia externa i el grau d’irreversibilitat del treball realitzat.
Les relacions entre l’energia exosomàtica i l’endosomàtica passen sovint per l’organització d’un entorn físic o per la construcció d’artefactes específics. Hi ha espècies que han aconseguit controlar l’energia a través d’estructures materials; el resultat és que hem augmentat o regulat la producció biològica, amb les conseqüències esperables en la competició. Hom ja ha esmentat que en la història de la vida es poden reconèixer tres cims on l’evolució ha obtingut un èxit considerable: el conjunt de les plantes superiors, amb tiges, troncs i arrels; el conjunt dels coralls formadors de baixos i esculls; i algunes nissagues d’animals superiors: els insectes socials en un grau relativament modest i, superlativament, la nostra espècie.
Hom ha comparat també el bosc amb el plàncton: la secció del xilema o vasos ascendents de la saba bruta, multiplicada per la velocitat de flux, resulta comparable a l’efecte d’ascensió de nutrients que s’aconsegueix en la barreja d’aigües oceàniques. En les plantes, la pujada de materials és limitada també per la quantitat d’aigua a disposició.
El cas de les formigues Atta
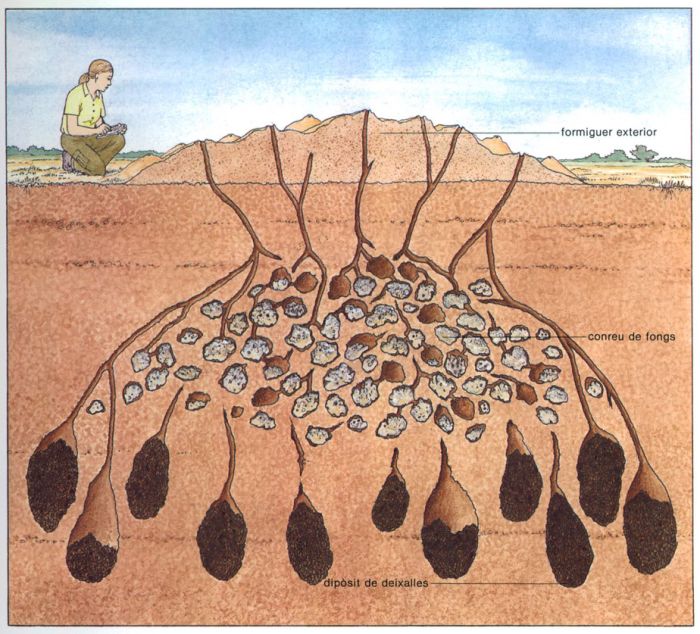
Aina Bonner
Les formigues Atta són insectes socials de comportament relativament complex, per tal com obtenen l’aliment gràcies a una mena d’agricultura subterrània: els fragments de fulles que tallen als arbres serveixen només com a substrat, un cop les han triturat convenientment amb les mandíbules i les han regat amb el seu líquid fecal. El que veritablement mengen les formigues Atta són els micelis d’uns fongs que elles mateixes implanten sobre la pasta de fulles obtinguda per masticació en cambres subterrànies dels seus nius, que funcionen com a autèntics horts. Alguns d’aquests fongs encara no han estat identificats, i molts no es troben fora d’aquests “horts” de les Atta. El dibuix mostra un tall vertical d’un niu del qual veiem al capdamunt el caramull superior, que pot arribar a ocupar una superfície de 250 m2 i en les galeries del qual poden arribar a viure centenars de milers de formigues; més avall, les cavitats o horts on els fongs són cultivats en pasta de fulles i, més avall encara, les grans cavitats que utilitzen les formigues com a dipòsits de deixalles, als quals van a parar les restes de pasta de fulles ja no aprofitables pels fongs.
Els efectes de l’energia exosomàtica sobre la vida
Les desigualtats que hi ha de distribució, de característiques i de possibilitats d’aprofitament de l’energia exosomàtica expliquen el repartiment heterogeni de les espècies: el treball realitzat per a portar aigua pot ser més important que el subministrament de llum a l’hora de configurar la distribució geogràfica de la vegetació. Les fluctuacions de la quantitat d’energia exosomàtica expliquen els canvis ocorreguts en el desenvolupament de la vida i de les seves activitats. Aquestes van associades al dia i la nit, al pas de les estacions i a d’altres fluctuacions més irregulars que guarden un considerable misteri; per exemple, la successió d’anys humits i anys secs, amb una possible distribució irregular o de contagi (almenys per parelles), que porta inacabablement a especular sobre la possible realitat de cicles de periode llarg o indeterminat, 4, 7 o 11 anys. A una escala més llarga de temps hi hagué les glaciacions, i un llarg etcètera que enllaça amb els canvis relacionats amb els moviments de les plaques continentals, els quals al seu torn depenen d’energia interna de la Terra, però que és energia externa amb relació a la vida i l’evolució dels organismes.
L’energia externa és important també perquè introdueix un element d’aleatorietat. Fluctuacions, intermitències i irregularitats contribueixen a l’eficàcia dels mecanismes ecològics. Pensem, per exemple, en les successions interrompudes per pertorbacions. Fluctuacions irregulars han afavorit el desenvolupament d’estructures destinades a persistir durant l’època de menor activitat vegetativa (troncs, arrels), i han facilitat l’evolució de plantes explotables i de sòls apropiats per a retenir nutrients, cosa que els ha fet idonis per a l’agricultura. Les zones temperades i la zona nòrdica particularment, n’han estat beneficiades, ja que és allà on el pas dels fronts atmosfèrics és prou freqüent i on els mateixos fronts expressen unes discontinuïtats més marcades.
Els coralls construeixen edificis ingents de carbonat càlcic en forma d’ilies o esculls, capaços de transformar l’energia dels corrents marins en energia turbulenta, la qual, en part, fa pujar aigua de capes més fondes i, en general, torna més fèrtils les aigües superficials, cosa que aprofiten més directament les algues simbionts. Recentment s’han trobat arguments per creure que els edificis coral·lins poden fer pujar nodriment del fons a través de l’estructura porosa de la massa calcària.
Els animals socials construeixen pistes i nius que configuren l’entorn de manera diversa i alhora els ajuden a aconseguir aliment. Això és ben clar en molts insectes, en els quals l’emmagatzemament i la conservació de recursos equival a obtenir-ne més i amb més regularitat. La vida social de les formigues i d’altres animals comporta una estructuració de residències i camins que en els tèrmits i les formigues és essencial per a l’establiment de cultius intensius; s’hi associa el maneig i la transformació de la fusta pels tèrmits.
L’espècie humana i el consum d’energia
Funcions i capacitats comparables a les esmentades esdevenen encara més evidents en l’espècie humana. L’energia exosomàtica en aquest cas és mesurable pel consum de fusta, petroli, electricitat, etc., i va des d’uns valors comparables als de l’energia endosomàtica (les 3 000 kcal diàries, o una mica menys, 120 watts, acceptats com a valors normals) que corresponen a una vida senzilla (és a dir, l’ús d’una mica de llenya per cuinar i escalfar-se) fins a un valor tan gran com es vulgui. En una etapa intermèdia s’hi afegeix l’energia associada amb l’emmagatzemament, empaquetament, etc., dels aliments, que evita moltes pèrdues (pèrdues per a la nostra espècie, peró no per a l’ecosistema), entre altres les causades per bacteris, fongs i diversos animals comensals, des dels corcs fins a les rates. La il·luminació, el transport, el condicionament climàtic i la indústria, incrementen enormement l’ús d’energia.
Avui dia la mitjana mundial de despesa d’energia exosomàtica deu quedar entre 15 i 20 vegades la mitjana referent a l’energia endosomàtica. No hi ha un límit superior individual: una persona pot usar, en forma d’energia exosomàtica, 100, 1 000 o més vegades els 120 watts de referència bàsica. El càlcul pot ser difícil perquè l’ús d’energia exosomàtica és, en bona part, col·lectiu. L’energia exosomàtica té propensió a multiplicar-se ella mateixa, de manera que qui en gasta molta esta en condicions d’augmentar-ne el consum encara més.
L’energia endosomàtica segueix una distribució gairebé normal en la humanitat, ja que els valors extrems, tant per fam com per excés de menjar, en limiten la distribució i la mantenen acotada. El repartiment de l’energia exosomàtica és ben diferent, i són les freqüències dels logaritmes del consum allò que s’aproxima a la distribució normal. Queda, doncs, prop d’una distribució aleatòria de tipus brownià, on cada trajecte a l’atzar no parteix de zero sinó del punt al qual s’havia arribat, i on la variància és proporcional al quadrat de la mitjana. En augmentar el consum mitjà d’energia exosomàtica, la seva variància augmenta, i aquesta característica es pot veure com a negativa i injusta. El funcionament de la nostra societat genera aquesta distribució estadística, i sembla que hi ha poques esperances de sortir-ne, per més que hom pugui creure indesitjable la situació present.
Consum energètic al llarg de la història dels humans
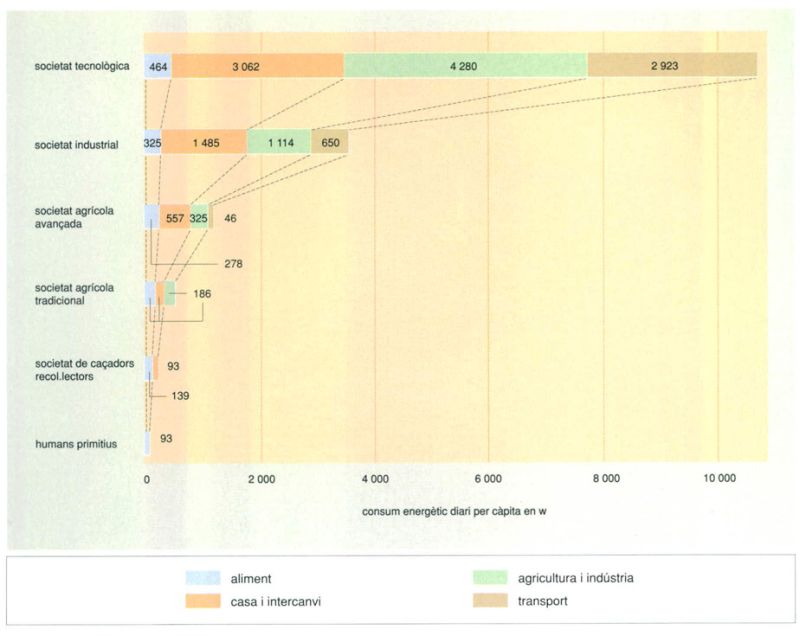
Editrònica, a partir de fonts diverses
El consum d’energia pels humans ha variat enormement d’uns pobles a uns altres i al llarg de la història segons el desenvolupament tecnològic. Els humans primitius es limitaven a consumir l’energia necessària per a llur supervivència i la prenien dels aliments. A les societats de caçadors-recol·lectors, el consum d’energia exosomàtica era nul o insignificant, però l’ús del foc per a la cocció d’aliments i per a l’escalfament dels llocs d’habitatge, de vegades construïts expressament, representà un primer recurs voluntari de fonts d’energia exosomàtica. L’aparició de l’agricuitura, amb la subsegüent sedentarització, que implicava la creació d’excedents de producció i l’acumulació de béns, significà la possibilitat d’aparició de consum. En societats agrícoles més avançades, les necessitats del transport de mercaderies a distàncies cada cop més grans es feren paleses. Amb l’aparició de la societat industrial, i la societat tecnològica posterior, el consum d’energia exosomàtica va créixer de manera espectacular. Cal tenir present que l’increment de l’energia vinculada al consum d’aliments no representa un increment paral·lel de l’energia ingerida amb els propis aliments sinó que és l’energia d’allò associat als aliments que no s’aprofita i que va a les deixalles. El consum energètic d’un ésser humà, avui com a les societats primitives, no arriba als 100 W diaris.