Els primers humans pròpiament dits: Homo erectus
Els canvis que se succeeixen en el transcurs del temps en Homo habilis, amb un conjunt de formes de transició a l’est africà, donarà lloc a H. erectus, un homínid amb una grandària corporal superior, una bipèdia més eficient, un cervell molt més gran, una cultura més complexa i interpretable com a adaptació, amb llocs d’habitatge més o menys permanents, i una dependència alimentària més centrada en la carn, probablement obtinguda en caceres.
La difícil delimitació de l’espècie
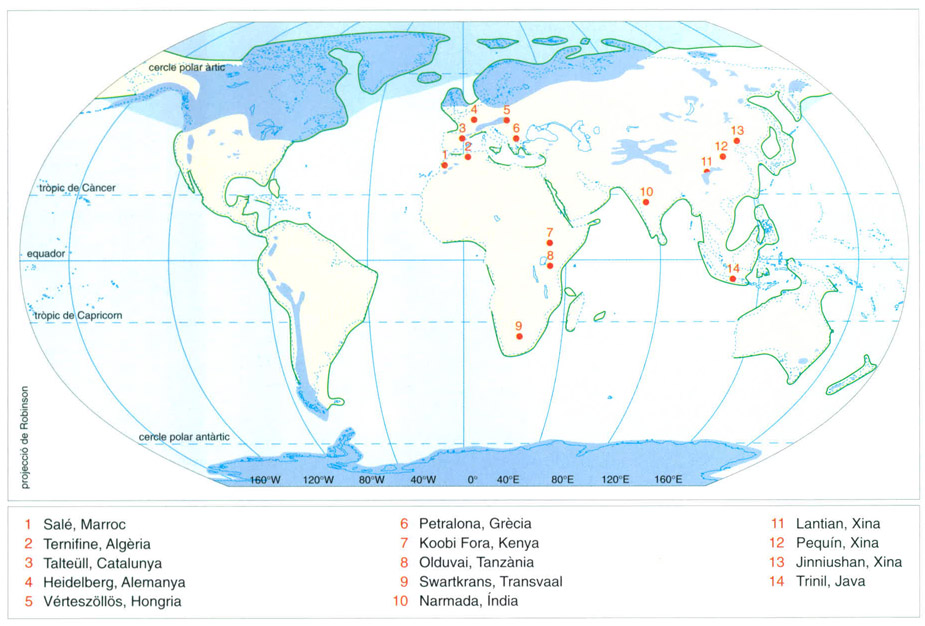
Les troballes corresponents a Homo erectus es localitzen en indrets geogràfics ben diversos; la diàspora d’aquesta cronospècie el portà a regions del sud-est asiàtic, a l’Àsia oriental, al continent indi i, segons alguns autors, també a Europa (les línies de costa són les estimades per a aquell temps). Als Homo erectus se’ls deu la conquesta de les zones temperades del planeta, com també altres invents importants en la història de la humanitat: la domesticació del foc, el descobriment de la simetria (amb l’aparició dels primers bifaços), l’ús de colorants, la invenció de la tècnica elaborada per a la talla de pedra i les primeres activitats rituals.
Editrònica, a partir de fonts diverses
Hem d’entendre Homo erectus com una cronospècie. És a dir, un conjunt de formes més aviat homogènies que viu en un període de temps, sense la pretensió de donar un significat biològic més profund al concepte en termes de reproducció recíproca. De tota manera, la imatge detallada no és senzilla i algunes restes són difícils d’encaixar dins l’esquema general. Molt probablement és en H. erectus en qui trobem els canvis culturals més marcats, lligats a la conquesta de nous territoris, amb noves estratègies evolutives que mostren el camí de la complexificació cultural en estreta relació amb l’adaptació biològica. Hi ha, en l’estratègia evolutiva, una dependència de l’adaptació cultural. Es reconeixen activitats complexes, com la fabricació d’instruments elaborats, la cacera major, l’ús del foc o la construcció de cabanes.
A grans trets comencem a reconèixer les formes d’Homo erectus en restes d’l,6 milions d’anys enrere a l’est àfrica, s’expandeixen fora d’Àfrica fa més d’un milió d’anys, en una data encara discutida, i dins d’una remarcable continuïtat tant morfològica com cultural els trobem per Àfrica i per Euràsia fins fa uns 300 000 anys, en què ja podem parlar de les formes arcaiques d’Homo sapiens. La transició, però, no es presenta amb la mateixa claredat ni amb la mateixa datació en els diversos llocs. H. erectus degué ocupar bona part d’Àfrica, amb restes a l’est, però també al Magrib i, expandint-se fora d’Àfrica, el trobem al SE i a l’E asiàtics, a l’Índia i a Europa. Aquí el coneixement és molt més gran per les nombroses restes fòssils i jaciments arqueològics que, de fet, ens permeten una anàlisi no únicament del grup com a tal, sinó de la variació en l’espai i el temps.
Malgrat tot, moltes vegades es fa difícil una delimitació clara de l’espècie, fet que porta a llargues i sovint inútils discussions. Per exemple, alguns autors, tot acceptant el tàxon Homo erectus, defensen que les formes europees s’haurien d’assignar exclusivament a Homo sapiens: però tot i que certes proves poden refermar-ho, tal com veurem seguidament, el panorama conjunt és molt més complicat. Es complica molt especialment per l’exigència de delimitació que imposa el fet de parlar d’espècies diferents (Homo habilis, H. erectus i H. sapiens), entitats discretes no sempre delimitables.
Els trets dels primers humans
Les característiques generals de l’Homoerectus el mostren poc diferent de les formes posteriors i, fins i tot, dels humans moderns pel que fa a l’esquelet postcranial. D’estatura alta i en algun cas fins i tot extraordinària amb relació a la dels humans actuals, devia caminar de manera molt semblant a la nostra. Les grans diferències respecte als humans moderns es troben en el crani, on sovint hi ha una evolució en mosaic. És a dir, hi ha alhora caràcters primitius, clarament típics de formes anteriors, i altres de derivats, que seran típics de formes posteriors. Tots plegats donen una morfologia característica que trobem en restes d’àrees geogràfiques ben diferents i que fan acceptable posar-les sota una denominació única.

L’estructura somàtica dels Homo erectus era molt semblant a la dels homes actuals excepte en el crani, on era prou diferent. El crani d’Homo erectus tenia una forma pentagonal —en norma posterior— i un important tor nucal per a la inserció de la musculatura del coll; també tenia, generalment, una aresta sobre la línia sagital. La cara mantenia encara trets arcaics, com el prognatisme —cara projectada cap endavant—; una arcada supraorbitària força marcada, amb engruiximent ossi; un front escàs tirat cap enrere; i una mandíbula robusta però sense barbeta, La dentició d’Homo erectus era diferent de la dels homes moderns en certs trets i similar en d’altres: les dents del davant eren voluminoses, els pre-molars, bicuspidals, i els molars morfològicament molt similars als dels homes actuals. L’esquelet de les extremitats d’Home erectus s’assemblava molt al de l’home actual.
Biopunt, a partir de fonts diverses
El crani d’Homo erectus és gros: la capacitat craniana té una mitjana d’uns 1 000 cm3 (oscil·la entre els 800 i els 1 200) amb un augment manifest en el temps. El volum gros va acompanyat d’una forma caracteritzada per un engruiximent de l’os sobre les òrbites, que fa una mena de visera o tor supraorbital; el front no és vertical, sinó que està tirat enrere i dóna un aspecte aplanat a la volta del crani. La part del darrere és sortint, i en comptes de ser arrodonida com en els humans moderns, és angulada i forma un tor nucal, en el qual, per la part inferior, s’insereix la musculatura del coll. Vista de perfil, la cara es mostra amb una projecció cap endavant, tret anomenat prognatisme: la mandíbula està mancada de barbeta i té una branca molt gruixuda, tret indicador d’una gran zona d’inserció d’una potent musculatura mandibular.
La dentició és similar a la de les poblacions modernes, però més grossa i amb característiques típiques. Per exemple, les incisives tenen la forma anomenada de pala, caracteritzada per un engruiximent de l’esmalt dels dos costats del darrere de cada dent; sembla que sigui un caràcter relacionat amb un fort ús de la dentició del davant, que dóna suport i més resistència al desgast.
Les restes descobertes i l’embolic taxonòmic
La millor aproximació a Homo erectus s’aconsegueix, sens dubte, resseguint les restes dels jaciments més importants on se n’han trobat. En força casos, i especialment en els més antics, es donaren a aquestes restes d’H.erectus noms científics propis que contribuiren a crear confusió, però d’ençà de la unificació del grup per Franz Weidenreich l’any 1940, aquests noms només formen part de l’anecdotari, tot i que es fan servir informalment per a designar restes concretes. Per raons expositives el recorregut anirà d’Àsia (Java, Xina i l’Índia) a l’Àfrica (E i N) i finalment a Europa. No farem referència, ni de lluny, a totes les restes o jaciments: són massa nombrosos fins i tot per esmentar-los tots.
L’home de Java
Java va ser, de manera casual i espectacular alhora, un dels primers llocs on es recuperaren restes dels nostres avantpassats. El 1887, Eugene Dubois, un metge holandès, decidí deixar la seva feina a la càtedra d’anatomia de la Universiteit Amsterdam per anar al que llavors eren les Índies Holandeses a buscar aquella espècie hipotètica que Ernst Haeckel havia batejat com a Pithecanthropus alalus,l’humà pòngid sense parla que, segons Haeckel, havia de ser “la” baula entre els altres primats i els humans. Per concepcions equivocades sobre l’origen de primats del SE asiàtic, Dubois va decidir anar a Indonèsia, primer a Sumatra i després a Java. I no tardà a fer grans descobertes.
L’any 1891, prop de la població de Trinil, als marges del riu Solo, a Java oriental, Dubois descobrí la part superior d’un crani amb una forma desconeguda llavors i que inicialment cregué que era d’un ximpanzé (espècie que actualment sabem que mai no havia viscut en aquella regió). L’any següent descobrí un fèmur en el mateix nivell, amb una clara aparença humana; convençut que les restes corresponien veritablement a la baula perduda que cercava, i notant que la posició erecta era la característica més destacable d’aquest homínid, les anuncià com a forma de transició cap als humans amb el nom de Pithecanthropus erectus:l’humà pòngid de posició erecta. Com altres vegades, la reacció no es féu esperar, tant per part d’antievolucionistes com de científics que creien que el fèmur podia ser d’un individu actual i el crani d’algun gibó gegant. Dubois, ofès i desconfiat, amagà les restes durant molts anys, fins que altres troballes aplanaren el camí de la interpretació de les restes de l’“home de Java”.
En conjunt, el panorama de les troballes javaneses resulta complicat per l’extraordinària complexitat de l’estratigrafia, que en molts casos no ha permès datacions acurades i en altres només n’ha consentit de molt dubtoses. En general, les edats s’han estimat a partir de la fauna associada al jaciment. Se sap molt poca cosa del tipus de vida, ja que són molt escasses les restes de cultura material i en cap cas no s’ha excavat un sòl d’habitació. Algunes restes (com per exemple algunes de Sangiran) podrien ser molt antigues, de més d’un milió d’anys, però no hi ha evidència suficient per acceptar les datacions que donen més edat. Amb un augment de capacitat craniana continuat en el temps, s’arriba a formes molt més recents, encara atribuïbles a Homoerectus, i amb una datació de 300 000 anys o possiblement menor en el cas de les restes de Solo, de les quals tornarem a parlar. Trobar restes tan antigues en un lloc tan allunyat de l’Àfrica, i a més en una illa, no deixa de sorprendre. Cal tenir molt present, però, que Java no era una illa quan hi vivia H. erectus, sinó que llavors formava part de les terres altes de Sunda, una península que abastava gran part de les illes actuals del SE asiàtic.
L’home de Pequín
La història de les troballes més importants de la Xina, les de Pequín, no fou tan tumultuosa com la de les de Java, però en canvi les restes trobades tingueren un final molt pitjor: desaparegueren definitivament durant la Segona Guerra Mundial. El que passà exactament encara avui és un misteri, però sortosament Franz Weidenreich, abans d’abandonar Pequín, on havia dirigit del 1935 al 1941 el laboratori de recerques sobre el Terciari del Servei de Geologia de la Xina, va poder treure del país, malgrat la situació de guerra, una bona col·lecció de reproduccions, fotografies i dibuixos que encara avui són la base per als estudis de les restes.
Les restes inicials es trobaren en la cova superior de Zhoukoudian (Chou-kou-tien, en transliteració Wade) i s’assignaren a un nou tàxon, Sinanthropus pekinensis, però la semblança amb les formes javaneses portaren a incloure-les en el mateix grup. A partir del 1949 es reprenen les excavacions i, sortosament, es troba molt més material, especialment eines de pedra (més de 100 000). En total hi ha 6 cranis gairebé complets i altres restes postcranials que representen un total d’uns 40 individus.
Gràcies a les monografies d’autors xinesos es coneix bé la datació dels diferents nivells de la cova, entre 460 000 i 230 000 anys, i s’ha pogut estudiar bé el tipus de vida. Així, s’ha dit que “l’home de Pequín era un habitant de coves, feia servir el foc, caçava cérvols, recollia llavors i feia eines especialitzades”. Una descripció que, pet que havíem vist fins ara, dóna una perspectiva completament nova i molt més complexa i coneguda del tipus de vida.
A més de les extraordinàries restes de Pequín, altres jaciments xinesos han donat importants fòssils d’Homoerecrus. A Lantian, prop de Xi’an, a Shaanxi, hi ha la resta més antiga, un crani datat en uns 750 000 o 800 000 anys, amb una capacitat craniana de 780 cm3. La resta més recent, que alguns autors situen ja dins d’Homosapiens, és la de Jinniushan, a Hubei, d’uns 280 000 anys.
El crani de Narmada
Durant molt de temps, les restes de Java i la Xina han estat les úniques conegudes a l’Àsia. Però la descoberta, l’any 1982, d’un crani a les terrasses del riu Narmada, prop de la localitat d’Hathnora, a l’estat de Madhya Pradesh, al centre de l’Índia, situava la primera passera entre les poblacions d’Àsia i Àfrica. Es tracta del crani de Narmada, de datació desconeguda i d’una morfologia que s’ha descrit com d’Homo erectus evolucionada.
Les restes africanes
Molt més tard que les restes asiàtiques, s’han descrit restes d’Homo erectus de l’est africà al barranc d’Olduvai i a les ribes del llac Turkana. Del barranc d’Olduvai Louis Leakey va descriure diverses restes (anomenades pel número de catàleg OH9 i OH12) que clarament són d’H. erectus amb una edat de prop del milió d’anys el primer i dels 700 000 anys el segon. Les restes més interessants, però, provenen de les ribes del llac Turkana. L’any 1975 es trobaren a FE del liac, en la formació de Koobi Fora, les restes més antigues, d’1,5 milions d’anys: dos cranis complets (anomenats KNM-ER 3733 i 3883, d’acord amb el número de catàleg de les restes del Kenya National Museum) amb una capacitat craniana de 800 i 850 cm3 respectivament, que morfològicament entroncaven amb Homo habilis. Més tard (1984) en la part W del llac es descobriren les restes més sorprenents: l’esquelet gairebé complet d’un noi d’uns 12 anys, d’elevada estatura (165 cm, que potser hagués augmentat en fer-se adult) i capacitat cranial elevada (900 cm3). La datació, també sorprenent, el situa a 1,6 milions d’anys: el més antic conegut i amb unes característiques ja a allunyades de les d’H. habilis. Aquesta resta ha fet proposar una data més antiga a la transició i una velocitat més alta de canvi.
A Sud-àfrica, al jaciment de Swartkrans, es trobà un conjunt de fòssils que s’assignen també a aquest tàxon; la seva datació, com calia esperar, és poc precisa.
Finalment hem de fer referència a les troballes de restes d’homínids, especialment mandíbules, al N d’Àfrica, amb una datació d’ entre 700 000 anys (les de Ternifine, al sud d’Orà, a Algèria) i 350 000 anys (les de Salé, al Marroc).
Les restes europees
Els primers assentaments humans a Europa daten, segurament, d’entorn d’un milió d’anys, tot i que hi ha algunes propostes d’endarrerir aquesta data. La resta humana més antiga és la mandíbula de Mauer, trobada prop de Heidelberg, a Alemanya. Tot i que hi ha jaciments arqueològics anteriors que no han donat restes humanes, aquesta mandíbula és probablement la resta humana més antiga i la que presenta una morfologia més semblant a les formes no europees d’Homo erectus. Altres restes (Vértesszöllös, a Hongria; Petralona, a Grècia; Bilzingsleben, a Turíngia, Alemanya; Aragó, a Talteüll, prop de Perpinyà, a la Catalunya Nord) solen situar-se dins del mateix tàxon però amb certa polèmica. De fet són poblacions contemporànies de les d’Homo erectus a l’Àsia i a l’Àfrica. Però et fet que posseïssin algunes característiques que fan pensar ja en els Homo sapiens més arcaics i la manca d’una discontinuïtat entre uns i altres fa que alguns autors s’estimin més no parlar d’H. erectus a Europa i posar totes les restes en un ampli calaix d’H. sapiens arcaic.
Diverses evidències en molts jaciments, no únicament europeus, fan pensar que l’Homo erectus tenia un tipus de vida de gran caçador. Així ho mostraria l’acumulació d’ossos de grans animals, alguns dels quals presenten marques d’haver estat esquarterats amb eines tallants de pedra. Per algunes de les caceres observades s’ha deduït que havien estat fetes en grup, fet indicatiu d’una complexitat social. Les eines són molt més complexes; destaquen especialment els bifaços, característics de la cultura anomenada acheuliana (pel jaciment francès de Saint-Acheut, a la Picardia). En alguns casos, com a Terra Amata, a Niça, s’han descobert sòls d’habitació en els quals sembla que hi havia cabanes temporals que serviren d’habitatge i taller per tallar pedra.
La ruta més probable d’expansió fora d’Àfrica és a través de ta península Aràbiga, llavors encara unida a l’Àfrica, i amb rutes que degueren arribar per un cantó a l’Àsia i per l’altre a Europa. No es descarta un pas des del nord d’Àfrica a Itàlia i es discuteix ta possibilitat del pas a través de l’estret de Gibraltar.
El jaciment de Terra Amata

Biopunt, a partir de fonts diverses
El jaciment de Terra Amata (Niça, Costa Blava) ha permès conèixer cents aspectes de la vida dels habitants d’Europa durant el Plistocè mitjà. Hom ha trobat evidència de cabanes ovals, de 7 a 15 m de llarg per 4 o 6 m d’amplada, prou grans per a haver pogut refugiar uns deu individus. L’estacionalitat anual influïa enormement en els assentaments humans; després d’estar un període de temps a Terra Amata, els caçadors desmuntaven el campament i es dirigirien cap a altres zones; en tornar a la mateixa zona l’any després, devien construir noves cabanes sobre l’assentament anterior, ja que les antigues restarien recobertes per sorra. Dins les cabanes, els humans que hi vivien fabricaven eines i excavaven petits togons on encendre foc. Hi ha proves suficients per a pensar que els homes d’aquella època eren capaços d’explotar amb habilitat els recursos de l’entorn, caçaven elefants, conills i urs, i pescaven mamífers aquàtics, peixos i marisc.
L’emergència de la nostra espècie: Homo sapiens primitius
Les restes més arcaiques
La base del problema que posen alguns autors d’acceptar com a Homo erectus els primers humans a Europa (ells proposarien incloure’ls com a Homo sapiens) és l’evidència d’un canvi gradual cap a aquesta darrera espècie. Així usualment es considera que les formes de fa uns 400 000 anys presenten unes característiques cranials que ja no són tes típiques d’H. erectus que hem vist i que fan que ja es parti d’H. sapiens.
Les restes europees
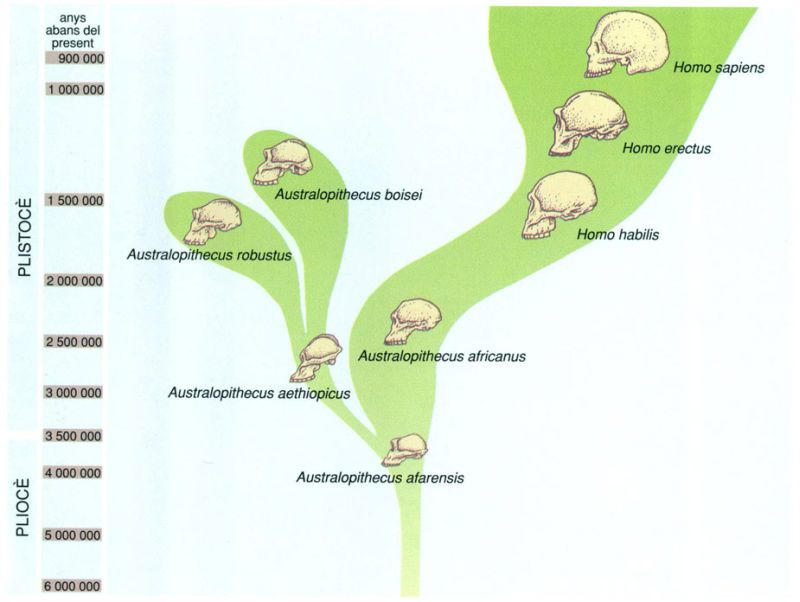
Esquema evolutiu dels homínids del Quaternari inferior elaborat amb les dades de que es disposa en l’actuahtat. La nissaga dels homínids se separa dels primats a l’Àfrica tropical i subtropical ara fa uns 5 milions d’anys, quan aparegueren els primers prehomínids (dels quals es te-nen restes de fa uns 4 milions d’anys); per tant, es pot dir que, com que el bressol de la humanitat ha estat Àfrica, en aquest continent han existit humans, com a mínim, 2,5 milions d’anys abans que en qualsevol altre. Se suposa que hi ha hagut dues dispersions d’humans per Euràsia, la segona de les quals va arribar fins a Amèrica passant per l’estret de Bering. La primera (Homo erectus) fou fa prop d’un milió d’anys i la segona (Homo sapiens) probablement fa entre 60 000 140 000 anys.
Biopunt, a partir de fonts diverses
És a Europa on aquesta transició s’observa millor, potser perquè la quantitat de restes és més gran. Seria una discussió sense fi intentar esbrinar de manera taxativa si restes com tes d’Aragó, Petralona, Bilzingsleben o altres estan ben classificades com a Homo erectus i no s’haurien de classificar com a Homo sapiens. Les restes europees un xic més modernes, cap als 250 000 anys d’antiguitat, comencen a presentar característiques que fan pensar en les poblacions humanes que, ben delimitades per la morfologia i per la distribució en l’espai i temps, viuran a Europa entre fa 70 000 i 30 000 anys, les de neandertal; ens podem referir a aquest conjunt de poblacions com a una raça de la nostra espècie. Les característiques morfològiques que els vinculen a la nostra espècie fan referència sobretot al crani. La capacitat craniana és alta, amb una mitjana de 1 350 cm3, molt semblant ja a la de les poblacions actuals; el crani en general globulós i gràcil; el front vertical i amb poques marques d’inserció muscular, especialment a la regió nucal; no hi ha el tor superciliar continu, sinó arcs superciliars damunt de cada òrbita; la mandíbula és gràcil, amb barbeta i una dentició de mida reduïda.
Com cal anomenar els humans que reconeixem que pertanyen a la nostra espècie, però que són anteriors als ben delimitats neandertals? Les propostes s’han succeït amb més o menys fortuna. No s’accepten denominacions com “pre-sapiens”, per raons òbvies, ni tampoc “pre-neandertals” o “anteneandertals”, ja que la referència a les formes posteriors els dóna poca identitat i sembla que indiqui que són els avantpassats directes dels neandertals; aquest fet, tot i que es pot acceptar, no té per què reflectir-se en el nom. Altres denominacions són més emprades, com els primers Homo sapiens oles formes de transició o les formes arcaiques d’H. sapiens. A vegades també s’anomenen formes pre-würmianes, referint-se al fet que visqueren abans de la gran glaciació de Worm, viscuda directament pels neandertals.
Dues restes es consideren com a prototípiques d’aquests primers Homo sapiens, la de Swanscombe, a Anglaterra, a l’estuari del Tàmesi i la de Steinheim, prop de Stuttgart, a Alemanya. Solen posar-se juntes per la proximitat de la datació, entre 200 000 i 250 000 anys. Però no són pas les úniques, ni de bon tros; també en són representants els fòssils de Biache-Saint-Baast, al Flandes francès, diverses troballes al conjunt de La Chaise, a la Charente, a l’W de França, i els més tardans, ja propers a l’última glaciació, d’Ehringsdorf, a Turíngia (Alemanya), i de Fontechevade, a la Charente (França). La mandíbula de Banyoles, a Catalunya, durant molt de temps melosa en aquest grup per afinitats morfològiques, una datació recent la situa dins de l’època del neandertal.
Les restes de Swanscombe comprenen dos parietals i l’occipital de la volta del crani. Es tracta d’una troballa afortunada, ja que els tres fragments del mateix individu es trobaren amb molts anys de diferència. La capacitat craniana estimada és de 1 325 cm3, molt per sobre de la d’Homo erectus i té un occipital arrodonit. Pet que fa al crani de Stcinheim, és més petit (1 100-1 200 cm3) i és parcialment deformat per la fossilització. Una característica, però, ha provocat grans discussions: la base del crani està trencada però d’una manera que suggereix que fou tallada per extreure’n el cervell. Això ha donat base a la hipòtesi del canibalisme, ja que sembla clar que la part trencada ho fou intencionadament abans d’iniciar-se la fossilització. De fet no es pot demostrar que la mutilació fos per menjar-se el cervell i no per a una activitat ritual o màgica, però ambdós fets podrien estar estretament relacionats. La possibilitat del canibalisme o almenys la modificació intencionada de la part inferior del crani es manifesta també en altres jaciments.
El jaciment que està donant més material de tot Europa és el d’Atapuerca, a Castella. Cada any hi ha noves restes, especialment mandíbules i peces dentàries, algunes d’extraordinària qualitat. Les més interessants s’han trobat en el fons d’una cova, amb associació a restes d’Ursusdeningeri, l’ós avantpassat del més conegut ós de les cavernes (U. spelaeus). La datació, temptativa, se situa entre 200 000 i 350 000 anys. Morfològicament, tot i ser unes restes molt robustes que recorden de prop la morfologia d’Homo erectus, ja s’hi troben alguns caràcters que fan pensar en els futurs neandertals.
Fora d’Europa diverses restes també mostren la transició cap a Homo sapiens, tot i que són molt més escasses. En cap cas, però, no hi ha una derivació cap a les formes neandertals, típiques d’Europa i, en canvi, algunes formes sud-africanes de més de 100 000 anys enrere seran uns bons candidats com a avantpassats dels humans anatòmicament moderns que trobarem més endavant.
A l’Àfrica hi ha encara les restes de Broken Hill (actualment Kabwe), al nord de Lusaka, a Zàmbia: un crani robust amb 1 280 cm3 de capacitat; les de Bodo, a la vall d’Awash, a l’E d’Addis Abeba, a Etiòpia, amb clares marques d’incisions sobre l’os que indiquen que l’individu fou descarnat amb un instrument tallant de pedra, i algunes altres, amb un mosaic de característiques entre Homo erectus i Homo sapiens.
A l’Àsia oriental també hi ha una transició que ha estat interpretada per alguns autors com a signe d’una evolució, independent de la d’Europa i Àfrica, des d’Homo erectus fins als humans moderns. És a dir, fins a les poblacions aborígens actuals. Aquesta teoria, que tal com veurem és rebutjada per molts altres que defensen l’origen africà dels humans moderns, postula una continuïtat morfològica, i per tant d’origen i descendència, des de les restes d’H. erectus de Zhoukoudian fins als humans anatòmicament moderns passant pels primers Homo sapiens (restes de Dali, prop de Jiefangeun, 120 km al NE de Xi’an, a Shaanxi, i de Maba, prop de Shaoguan, al nord de la província de Guangdong). El que sí que s’accepta, però, és una persistència més llarga de les característiques d’H. erectus a la Xina i al SE asiàtic que no pas a altres llocs, on la transició a H. sapiens ja s’hauria produït.
L’instrumental i l’habitatge
Des d’una perspectiva estrictament arqueològica, hi ha poques diferències entre l’instrumental d’Homo erectus i el dels primers Homo sapiens. Continua la tradició acheuliana, però els jaciments tenen escassos bifaços, especialment a Europa, i en canvi un gran nombre d’ascles, que són les peces de pedra, usualment sílex, que se separen d’un nucli en ser colpejat; posteriorment es poden retocar per aconseguir una forma més idònia. La diferència bàsica és que en els bifaços l’eina s’aconsegueix traient peces, i les ascles, en canvi, són les peces que es desprenen d’un bloc en ser colpejat.
Els primers Homo sapiens habitaven tant coves com llocs a l’aire lliure, amb un ús cada vegada més freqüent de les coves i hi ha evidències de l’ús del foc. El jaciment que millor ens mostra el tipus de vida és el de la cova de Lo Lazaret, a Niça, a la Costa Blava. S’hi ha trobat l’evidència d’una cabana que recolzava sobre la paret de la cova, delimitant un espai d’11 m per 3,5, en el qual es trobaren restes de dos focs i acumulacions de petites petxines, potser portades amb les algues que s’empraren per a fer els jaços. A l’interior, no hi ha acumulacions de restes de talla de pedres, però sí a l’exterior, cosa que mostraria una delimitació de l’espai. La presència d’un crani de llop a l’entrada de restes de cabanes que s’han trobat fora de la cova té un clar significat simbòlic.
S’hi pot veure que els recursos eren variats segons els hàbitats, amb molta diversitat d’aliments. Hi ha una utilització de recursos marins i també, en alguns casos, de pesca d’aigua dolça. S’ha suggerit que entre els primers Homo sapiens podria haver-hi signes de violència interpersonal que podrien haver provocat, per exemple, la fractura del crani d’Aragó; l’evidència, però, no és clara tot i que cal acceptar que hi hagué fracturacions rituals de les restes humanes.
L’esclat neandertalià
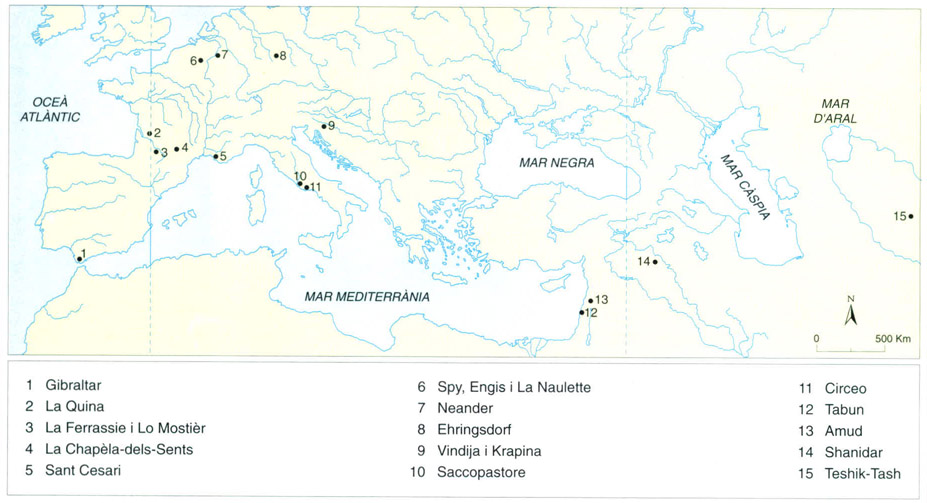
Distribució de les restes de neandertals. Els neandertals són els fòssils humans no moderns més famosos i ben coneguts, s’han trobat des de la part meridional i occidental d’Àsia fins a l’extrem més occidental d ’Europa, on visqueren en iniciar-se la glaciació Würm, fa entre 75 000 135 000 anys. No sembla difícil d’esbrinar per què els neandertals no sobrevisqueren en aparèixer sobre la Terra humans amb pautes de comportament més modernes. Les dades arqueològiques indiquen que, en qualsevol aspecte estudiat (instruments, objectes, construcció dels assentaments i altres), les pautes de comportament dels neandertals eren força inferiors a les de llurs successors, més moderns. No se sap gaire bé com es van extingir els neandertals ni per què, però es pensa que, a més de la desaparició per compètencia, es produí un fenomen de desplaçament i/o un fenomen de captació genètica per part de les poblacions humanes més modernes.
Editrònica, a partir de fonts diverses
Els neandertals representen un grup homogeni que potser es correspondria amb el concepte de raça tal com el coneixem actualment, amb la complexitat que implica el fet de tractar-se de poblacions pretèrites. De vegades se’ls dóna un rang de subspècie i s’anomenen Homo sapiens neanderthalensis. Es tracta, a grans trets, de les poblacions humanes que visqueren a Europa i al SW d’Àsia durant l’inici de la glaciació de Würm, entre 75 000 i 35 000 anys enrere, originades a partir de les poblacions que habitaven les mateixes regions i que, d’una manera certament sobtada, deixaren pas, a la fi d’aquest període, a les poblacions del que anomenem humans anatòmicament moderns, que trobem associades amb les cultures del Paleolític superior i que són indistingibles morfològicament de les poblacions actuals.
L’aspecte de l’home de Neandertal
El crani neandertalià era de grans dimensions, ample i llarg, amb un occipital arrodonit, sortint i sense angulació. La capacitat craniana és gran, amb una mitjana de 1 550 cm3, per sobre de la de moltes poblacions actuals, i amb alguns exemplars amb valors per sobre de 1 700 cm3. El front no era tan vertical com en l’home modern i sobre les òrbites els arcs superciliars són pronunciats. La cara era grossa i d’aspecte massís, projectada cap endavant. El nas devia ser gros, angulós i prominent i s’ha interpretat funcionalment per escalfar l’aire abans que arribés als pulmons. La dentició era més grossa que en les poblacions actuals, especialment les peces anteriors; les posteriors, en canvi, ja eren semblants a les de les poblacions modernes. La mandíbula usualment era sense barbeta.
Pel que fa a l’esquelet postcranial, les diferències amb els humans moderns consisteixen en una extraordinaria robustesa. Una característica ha cridat especialment l’atenció: el pubis, en la part antero-inferior de la pelvis, era llarg i prim, sense la robustesa que es troba en altres parts del cos, i deixava un canal del part ample. Aquesta característica s’ha relacionat per una banda amb la mida del cap, força gran: per això el canal del part havia de ser ample; però per una altra banda l’obertura més grossa es pot relacionar també amb un temps de gestació més llarg. Tot i que aquesta segona hipòtesi és especulativa, si fos certa tindria interessants implicacions evolutives. En conjunt en resulta una morfologia que esdevé familiar només d’haver vist uns quants fòssils, i que és característica exclusiva d’aquest grup.
Les restes neandertalianes

Representació figurativa d’un ritus d’enterrament neandertalià de la cova de Shanidar, Iraq. Hom creu que els homes de neandertal foren els primers humans a enterrar els seus morts amb algun tipus de cerimònia ritual, si no sempre, sí d’una manera ocasional. De totes les restes neandertalianes trobades a la cova de Shanidar, una d’elles va ser enterrada al principi del mes de juny, ara fa uns 60 000 anys. Les restes de pol·len indicaren, a més dels tipus de vegetals que hi havia en el moment de l’enterrament, que hi hagué intervenció humana: el vent no hauria pogut dur núvols de pol·len tan enormes ni deixar-lo en una disposició tan concreta. La investigació palinològica indica que només podia haver estat que les despulles estiguessin envoltades per flors completes.
Biopunt
Un dels exemplars més famosos és el de la Chapèla-dels-Secnts (la Chapelle-aux-Saints), al Llmosí, al centre de l’estat francès, que presenta les característiques esmentades molt exagerades, i que moltes vegades s’ha pres com a prototip del grup. De fet, quan Marcellin Boule l’estudià de primer a la década del 1910, en féu una descripció deformada i irreal, com si hagués estat un pòngid, ferotge i poc intel·ligent. L’error es pot relacionar amb clares marques d’artritis, que no interpreta correctament, o amb la poca convicció que sembla que tenia en l’evolució humana.
Les restes del centre d’Europa (jaciments de Krapina i Vindija, tots dos a Croàcia) tenen un interès especial. D’una banda apunten la possibilitat de canibalisme o, almenys, de descarnament dels cadàvers per un enterrament secundari, pràctica que ha perdurat en moltes poblacions durant mil·lennis. D’altra banda la morfologia no segueix tan clarament el patró d’Europa occidental i algunes característiques han estat interpretades com a indicadors d’una possible continuïtat amb les formes del paleolític superior. Aquesta interpretació té una clara repercussió evolutiva en entendre els canvis en la transició de neandertal a modern, tal com veurem.
Cap a l’Àsia trobem jaciments a Israel (Tahun, Amud). Iraq (Shanidar) i el ja esmentat a l’Uzhekistan (Teshik-Tash). La morfologia és semblant a la dels europeus, amb una menor robustesa i algunes característiques que fan pensar en restes més modernes. Aquí també és evident el problema de la substitució dels neandertals perquè en altres jaciments molt propers s’hi han trobat restes molt antigues que presenten una morfologia relacionable amb les formes modernes d’Homosapiens.
El jaciment de Shanidar, amb nou esquelets excavats, s’ha fet famós per un conjunt d’evidències indirectes de gran importància per a entendre la manera de viure i de pensar d’aquests humans. Les restes d’un dels individus mostren clarament signes de supervivència després d’un accident que li enfonsà l’òrbita esquerra. També presenta degeneració de l’extremitat superior dreta i cama dreta, probablement com a conseqüència de la ferida cranial que li pogué afectar el cervell. És dubtós si tenia el braç amputat; si es confirmés seria l’operació quirúrgica més antiga coneguda. Altres individus també han deixat signes de ferides: un d’ells, amb una forta artritis, probablement morí d’una ferida a l’alçada dels pulmons feta amb un instrument penetrant que ha deixat una clara marca en una costella, i un altre presenta una ferida cicatritzada en el costat dret del front. Aquests són alguns exemples de ferides i malalties que mostren la supervivència d’alguns individus que no es podien valer per ells sols.
Fora de l’àrea descrita, les restes de l’època dels neandertals no presenten les mateixes característiques morfològiques, i per tant no es consideren dins del mateix grup o raça. Els humans habitants de l’Àfrica i l’Àsia oriental i sud-oriental (no estava habitada l’àrea de Sibèria ni s’havia iniciat la conquesta d’Oceania ni la d’Amèrica) no són inclosos en una denominació única per manca de prou restes que en puguin donar una imatge de conjunt, i els antics noms de neardertals no-europeus o neandertaloides no són acceptables. Es parla, simplement, de contemporanis dels neandertals. A l’Àfrica hi ha una gran heterogeneïtat i, al costat de restes amb morfologia arcaica, relacionada amb les primeres formes d’Homosapiens,en trobem altres que, sorprenentment, mostren característiques modernes en el sentit que fan pensar ja en els humans anatòmicament moderns que veurem més endavant.
La cultura neandertaliana

Les eines que els neandertalians fabricaren a Europa i Àsia occidental s’assignen de manera universal a la cultura mosteriana i corresponen al Paleolític mitjà. La indústria mosteriana era tallada principalment en nuclis de sílex i, menys sovint, en calcàries o quars. La tècnica emprada per a aconseguir aquests instruments s’anomenà “tècnica Levallois”, i amb ella aconseguien un seguit d’eines. En aquesta indústria lítica mosteriana hi ha una remarcable uniformitat en el temps i l’espai, si bé es pot distingir una certa varietat regional. A més dels estris de pedra hom creu que els neandertalians també fabricaven habitualment instruments amb materials no perdurables, ja que en alguns jaciments mosterians s’han trobat restes d’objectes de fusta, com ara bastons.
Biopunt, a partir de fonts diverses
La cultura de l’home de Neandertal s’associa, a grans trets, amb la indústria mosteriana, el nom de la qual es deriva del nom del jaciment occità de Lo Mostièr (en francès Le Moustier), al Perigord, ben caractcritzada per la producció de nombroses ascles a partir d’un nucli i un posterior retoc per aconseguir la peça pretesa (un rascador, un gratador, una punta, un ganivet) amb tècniques molt més refinades i eficients que en èpoques anteriors. Probablement també empraren l’os i la banya, que no es troben de manera generalitzada, però, fins al següent període cultural. La tipificació de l’utillatge és molt complexa i ha ajudat els arqueolòlegs a definir i delimitar arces i períodes amb un denominador cultural comú.
La cova és el lloc d’habitació per excel·lència, tot i que hi ha indicis de vida en balmes i cabanes. Cal tenir present que visqueren en el temps i en els llocs més freds de tota la història humana fins llavors. Els jaciments mostren una ocupació temporal, d’acord amb un tipus de vida nòmada. L’alimentació estava repartida entre els vegetals recollits, amb evidència de diferents fruites, i la carn procedent de caceres que, per l’abundància de restes, degueren ser molt productives.
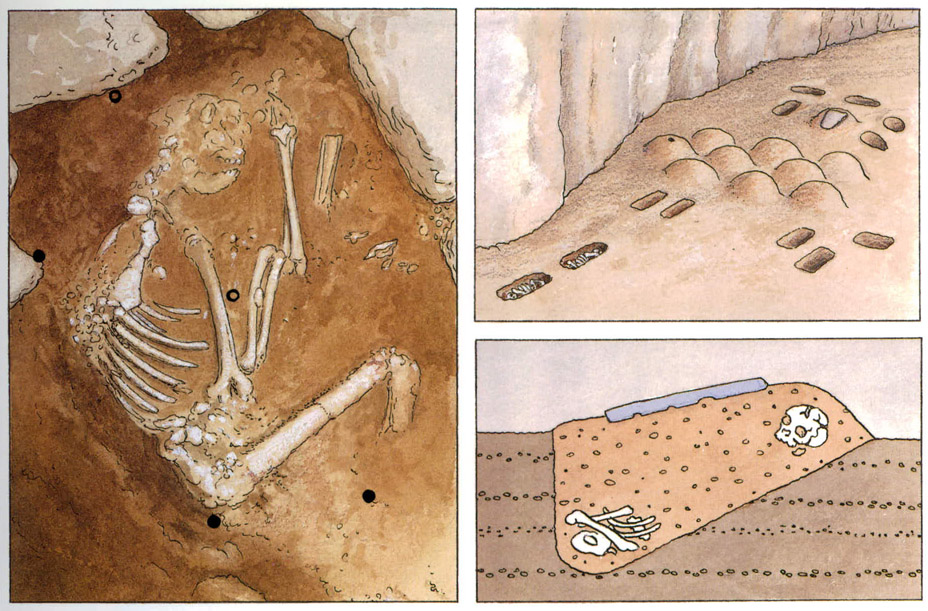
Les troballes d’enterraments neandertalians indiquen que foren els humans més antics a enterrar llurs morts. A l’esquerra, representació de com va ser trobat l’esquelet neandertalià Shanidar IV (Iraq) Els punts negres són mostres amb pol·len de flors; els punts buits, mostres sense pol·len. A la dreta, representacions del descobriment de l’enterrament, també neandertalià, de l’abric de La Ferrassie (Dordonya) on es trobà un cementiri familiar.
Aina Banner
Però l’evidència d’activitat més clara del grup es troba en els nombrosos enterraments fets de manera intencionada i ritual. Entre els neandertals s’aprecien, doncs, per primera vegada, signes d’espiritualitat o religió: el cadàver es dipositava en un forat fet expressament, en una posició específica, normalment fetal i acompanyat d’ofrenes que podien ser utensilis tallats (per fer servir en una altra vida?), restes d’animals com banyes de cérvols o espècies semblants. En diversos casos hi ha signes de descarnament i en alguns s’observa el trencament de la base del crani. Les restes de Monte Circeo, a Itàlia, mostren aquest trencament intencional i el crani fa pensar que l’individu morí d’un cop en el costat dret del cap i fou dipositat al mig d’un cercle de pedres; alguns autors han interpretat que hi hagué un sacrifici ritual. A l’entorn d’un esquelet de Shanidar, l’anàlisi del pol·len mostrà unes acumulacions que resultaren ser de flors de florida primaveral, i que sembla que haurien estat dipositades en la tomba durant l’enterrament.
A l’enterrament de l’esquelet neandertalià Shanidar IV (Iraq), els arqueòlegs prengueren mostres de la terra que envoltava el cadàver per tal de poder determinar-ne la composició. Gràcies a aquestes mostres es pogueren determinar les espècies de les flors que havien servit per al ritual de l’enterrament, algunes de les quals eren medicinals. A l’abric de La Ferrassie (Dordonya) es trobà un cementiri neandertalià familiar. Sota l’abric rocallós foren enterrats, cap amb cap, i separats uns 50 cm de distància, un home d’uns 45 anys i una dona d’uns 25; juntament amb ells hi havia les despulles de quatre nens i dos fetus. Hom pensa que els neandertals coneixien la noció de sepultura perquè hi ha restes d’utensilis i monticles de terra.
La distribució i la datació dels jaciments
La història de les descobertes de neandertals començà molt aviat, amb troballes a la cova d’Engis, prop de Lieja, a Bèlgica, el 1828, i a Gibraltar el 1848, però no foren reconegudes com a restes de poblacions pretèrites. De fet, el primer descobriment amb impacte en l’àmbit científic fou el d’una volta craniana en una pedrera a la vall (“tal” en alemany) de Neander, prop de Düsseldorf, a Alemanya, el 1856. Inicialment es valora com a representant d’una població antiga des del punt de vista prehistòric, sense pensar, però, en termes evolutius. Se sap que Darwin en coneixia l’existència quan publica “The origin of species”el 1859, però no va fer-hi referència. La primera discussió evolutiva relacionada amb el crani de Neandertal fou feta per Thomas Huxley, el 1863, i li donà un estatut semblant al que té actualment; un humà primitiu però molt allunyat dels pòngids. El nombre de restes ha anat creixent amb el pas del temps fins a arribar a centenars, la qual cosa ha permès estudis poblacionals acurats.
La delimitació en l’espai i el temps és complexa, ja que depèn de la definició que es faci del grup. Així, l’edat dels neandertals més antics pot ser donada per restes que comencen a presentar-ne característiques o bé que ja les presenten plenament. Atès que hi ha una bona quantitat de formes de transició, la delimitació resta flexible i alguns autors arriben a incloure-hi les restes d’Ehringsdorf, ja esmentades, i les de la pedrera de Saccopastore, a pocs quilòmetres de Roma, que usualment es consideren Homo sapiens arcaic. La delimitació a partir de 75 000 anys considera les formes ja morfològicament típiques i és el moment d’inici de la glaciació de Würm, que reforça l’elecció. Les dates més tardanes, en canvi, són més clares. La resta de Sant Cesari (Saint-Césaire), prop de Grassa, a Provença, descoberta el 1979, però datada el 1991, marca, de moment, l’aparició de la morfologia neandertal de datació més propera en el temps (uns 36 300 anys enrere).
La delimitació en l’espai és clara en la part occidental: tot Europa, incloent-hi Gran Bretanya (si més no, el sud d’Anglaterra), amb Alemanya i Polònia com a extrem nord. Cap a l’E, però, l’àrea de distribució s’adapta a les escasses i esparses troballes, fins abastar el llunyà jaciment de Teshik-Tash, a l’Uzbekistan. Segons l’àrea de distribució solen distingir-se el ric grup de l’Europa occidental, el de l’Europa central i les restes asiàtiques, especialment concentrades a Israel.
L’expansió de la humanitat: Homo sapiens actuals
L’origen dels humans moderns
La humanitat experimenta un esclat important a partir del Paleolític superior. Aleshores els humans començaren a ensenyorir-se de la Terra.
De manera global, pot dir-se que els humans del Paleolític superior foren grans caçadors. La cacera, especialment a causa de la fosa parcial del gel, era molt abundosa en tota la tundra i l’estepa eurasiàtiques. La tecnologia de la cacera era molt elaborada, amb innovacions especialment en el darrer període del Paleolític superior, el Magdalenià, que inclouen el propulsor per a tirar llances amb gran força, arpons per a la pesca o l’arc i la fletxa, de gran potència, precisió i llargada. Els instruments de pedra eren molt variats, amb una talla de gran precisió i bellesa.
El Paleolític superior també és el gran període de l’art. Les coves pintades, especialment de la regió cantàbrica i del vessant nord dels Pirineus, i les escultures humanes, anomenades “venus”, en són els exemples més clars, amb una gran perfecció tècnica, un sentit estètic i una gran càrrega simbòlica que, en ser interpretada, parla de creences, de mites, de religió.
Aquests humans paleolítics ja són plenament Homo sapiens sapiens, és clar. El seu origen exacte i el seu parentiu amb l’home de Neandertal o altres formes anteriors no és gaire clar. Hom ha formulat diverses hipòtesis pel que fa al cas.
La hipòtesi de l’origen africà
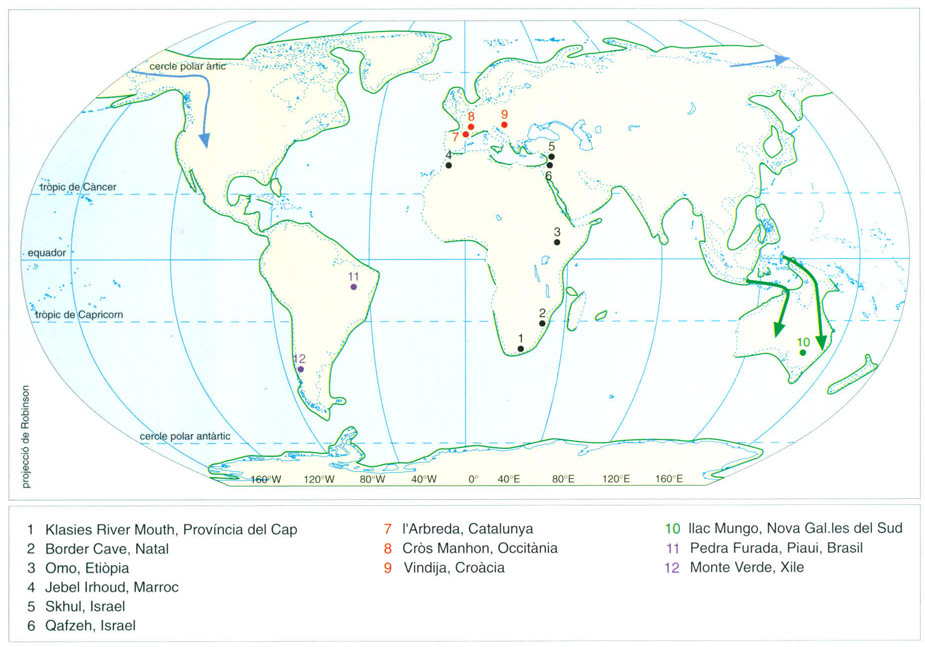
Lorigen africà dels humans moderns és refermat per les proves paleontològiques: els cranis trobats al continent africà que insinuen característiques noves (punts negres) són els més antics. En el dibuix podem veure les localitats més significativos en les quals han estat trobades restes d’Homo sapiens moderns esmentades al text (en vermell els jaciments europeus). Les línies de costa són les que hom pensa que hi havia entorn del màxim glacial de 20 000 anys enrere, si bé el pas d’Àsia a Amèrica només fou possible abans o després d’aquell màxim, per un suposat passadís lliure de gel que resseguia el peu oriental de les Rocalloses (sageta blava); els jaciments sud-americans estan indicats amb punts liles. La colonització d’Austràlia es féu per mar (sagetes verdes).
Editrònica, a partir de fonts diverses
Les restes més antigues que presenten característiques modernes es troben a l’Àfrica i l’acceptació d’aquest fet ha donat hoc a la hipótesi, dins del model de substitució, de l’origen africà dels humans moderns (anomenada “out of Africa”). Els antropòlegs tenen la mirada posada en els jaciments de Sud-àfrica: la cova de Klasies River Mouth, a la costa meridional de la província del Cap, i l’anomenada Border Cave, al NE de Natal, prop de la frontera amb Ngwame (Swazilàndia). Tots dos amb datacions superiors als 100 000 anys i morfologies modernes, especialment pet que fa a l’absència de tor supraorbital, a la forma general del crani, a la presència de barbeta, etc. De tota manera els crítics addueixen que el primer ha donat poques restes per confirmar la seva assignació i en el segon, indiscutible a nivell morfològic, la datació no és definitiva. Són, però, peces clau per a una explicació global, i la posició com a primeres formes modernes s’accepta per gran part dels estudiosos.
Fòssils d’altres parts d’Àfrica han estat estudiats de nou recentment i reinterpretats com a portadors de característiques morfològiques modernes. Així, un crani d’Omo, a Etiòpia, i les restes de Jebel Irhoud, al Marroc, sembla que morfològicament són en la mateixa línia. Si aquesta interpretació és correcta, les restes més antigues d’humans moderns serien africanes i podrien ser la base o l’origen de totes les poblacions modernes: de l’Àfrica hi hauria hagut una expansió a l’Orient mitjà, des d’on s’haurien estès cap a l’Àsia i Europa. De fet, les restes més antigues no africanes amb caràcters moderns es troben a l’Orient mitjà i són més modernes que les africanes que hem vist.
Dos jaciments, coneguts des de ja fa temps, han proporcionat les restes més importants: Jebel Qafzeh (normalment referit com a Qafzeh) i Mugharet es-Skhul (referit com a Skhul: el crani més conegut és el Skhul V), tots dos a Israel. La morfologia dels diversos individus d’aquests jaciments mostren, dins d’una considerable robustesa, caràcters indiscutiblement moderns. De fet durant un temps es postulà que aquestes formes podien ser els avantpassats del humans moderns europeus i que podrien haver evolucionat a partir dels neandertals de la mateixa regió. Però revisions posteriors de les datacions els han situat en dates anteriors a alguns dels neandertals: entre 90 000 i 100 000. No hi ha, doncs, evolució dins del mateix lloc i, en canvi, s’obre la possibilitat que siguin formes modernes emigrants de l’Àfrica abans de l’expansió cap a la resta del món.
Darrerament un nou tipus d’evidència ha proporcionat noves dades: l’anomenada antropologia molecular. Amb diversos tipus de dades moleculars (especialment freqüències gèniques i seqüències del DNA mitocondrial) s’ha afavorit de manera clara la hipòtesi de l’origen africà. Les diferències gcnètiques entre les poblacions humanes actuals refermen la idea d’un temps relativament curt de diversificació, d’acord amb la proposta d’origen africà de les poblacions modernes i l’expansió a tot el món, amb substitució de les poblacions prèvies; no dóna suport, en canvi, a un origen molt antic de les poblacions actuals.
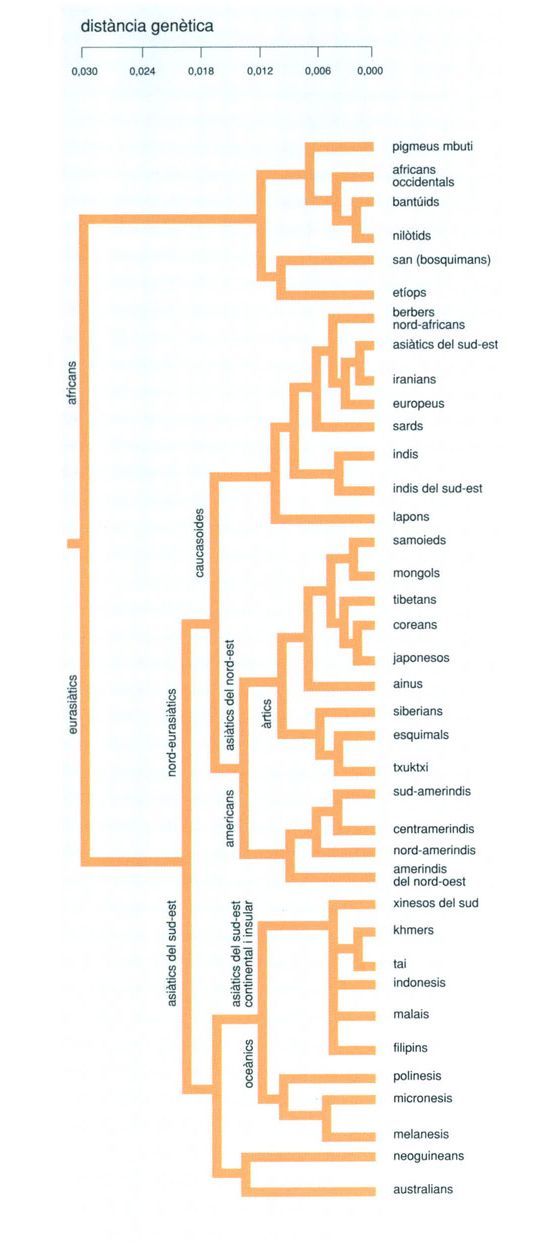
Les distàncies genètiques entre les diverses poblacions humanes són un fidel reflex del temps passat des que se separaren, i n’indiquen el parentiu. Estudiant llur distribució hom pot elaborar una mena d’arbre genealògic de l’espècie que il·lustra la dispersió humana pel planeta. No és gens sorprenent que es trobin força similituds entre la distribució dels gens i la distribució geogràfica de les llengües.
Biopunt, de Cavalli-Sforza, 1992
L’aparent discontinuïtat, o canvi sobtat, que es manifesta entre el Paleolític mitjà i el Paleolític superior a Europa, en particular a la seva part occidental, acaba de refermar la hipòtesi d’una substitució relativament ràpida dels neandertals pels humans moderns. El canvi es manifesta tant en l’aspecte físic com en el cultural. Es fa palès un canvi sobtat entre les tecnologies del Paleolític mitjà (Mosterià, associades a les neandertals) i les primeres del Paleolític superior (Aurinyacià, normalment associades a humans moderns). A l’Arbreda, un jaciment de la localitat catalana de Serinyà, prop de Girona, per exemple, una anàlisi detallada del jaciment ha permès detectar una ràpida transició datada en uns 40 000 anys, data molt antiga i que en altres llocs encara es troba en ple Paleolític mitjà i amb restes clarament neandertalianes.
Les restes d’humans moderns del Paleolític superior a l’Europa occidental són molt nombroses. Descobertes d’antic, donaren lloc a la descripció de l’anomenada raça de cromanyó, denominació que cal evitar ja que no correspon a res en especial. De fet, les restes del jaciment del Cròs Manhon (més divulgat amb el nom francés de CroMagnon), situat prop de la localitat occitana de Las Eisiàs de Taiac, a la Dordonya, al SW de l’estat francès, s’han mantingut durant temps com a prototip d’humà modern a Europa, però de fet hi ha una gran diversitat dins de les característiques inconfusiblement modernes; d’altra banda, són restes poc antigues, d’uns 25 000 anys, i per tant de molt temps després de la transició.
A l’Europa central i de l’est les restes són més discutides ja que, en alguns jaciments com Vindija i Velika Pecina a Croàcia, hi ha individus amb caràcters moderns i alhora una robustesa i algunes altres característiques que poden recordar el neandertal. Per això alguns autors donen suport a una evolució in situ en comptes de la substitució dels neandertals pels humans moderns.
La hipotesi multicèntrica o de continuïtat regional
L’aparició de les formes modernes s’hauria produït independentment en diversos llocs, segons la d’hipòtesi multicèntrica o de continuïtat regional, que manté que des de la fase d’Homo erectus hi ha hagut una evolució continuada almenys a l’Àfrica, Europa i a l’Àsia. Promotors d’aquesta teoria defensen que, després de la diversificació creada en l’expansió d’H. erectus per la selecció natural i la deriva genètica deguda a poblacions reduïdes, hi hauria hagut una evolució paral·lela per selecció i flux gènic entre poblacions gràcies a les migracions. És en el continent asiàtic (especialment al SE) i a Austràlia on es pot parlar d’una certa continuïtat morfològica des de formes antigues a modernes, amb exemples de transició.
De fet, però, hi ha molts arguments contraris, com per exemple la gran diversitat morfològica dels primers australians, els quals presenten característiques que no estan presents en les formes arcaiques de Java i en canvi sí que hi són en les formes modernes africanes. D’altra banda, no hi ha enlloc fòssils que indiquin clarament una transició i, en canvi, sí que es pot parlar en alguns llocs de substitució. Finalment, si la hipòtesi multicèntrica fos correcta, hi hauria d’haver una velocitat d’evolució constant en diversos llocs que en realitat no es produeix i, en canvi, s’observa una aparició escalada en el temps de les formes modernes a mesura que ens allunyem de l’Àfrica.
L’expansió i la colonització de noves terres
Tots aquests nuclis d’humans actuals, sorgits o implantats a l’Àfrica i a Euràsia, han conegut una expansió formidable en els darrers mil·lennis. La Terra sencera ha estat ocupada pels humans, però cal no oblidar que molts territoris no van ser petjats per l’espècie humana fins a temps realment molt recents. És el cas d’Austràlia i les illes oceàniques del nord d’Euràsia mateix, o singularment de tot el continent americà.
L’ocupació completa d’Euràsia
La completa ocupació d’Euràsia s’aconseguí amb l’establiment humà en les inhòspites terres de Sibèria, que no sembla haver-se produït abans de fa uns 35 000 anys. Hi ha diversos jaciments en la vall del riu Lena i una prova indirecta n’és el poblament del Japó, que es féu fa uns 30 000 anys o més, probablement a través de les terres que unien les illes més importants amb el SE de Sibèria durant l’última glaciació.
La colonització d’Austràlia
El poblament d’Austràlia s’assolí fa, com a mínim, 33 000 anys, probablement 40 000. Això mostra que ja existia la tecnologia per viatjar per mar en temps molt reculats: tot i que les illes d’Indonèsia estaven unides a l’Àsia durant les glaciacions. Austràlia estava separada per mar. Tot i aprofitant les illes intermèdies, calia fer navegant a mar obert diversos trajectes d’uns 30 km i com a mínim un de l’ordre d’uns 90, distàncies que requereixen un bon nivell tecnològic. L’entrada dels humans al continent australià probablement es féu pel NE més proper a Nova Guinea, i primer se’n degueren ocupar les costes. La totalitat del territori ja sembla ocupat fa 20 000 o 25 000 anys.
Les restes humanes més antigues conegudes d’Austràlia són les del llac Mungo, al SE de l’illacontinent, datades de fa entre 26 000 i 30 000 anys. La seva morfologia s’assembla a la de restes anteriors del SE asiàtic, però és diferent de la coneguda dels aborígens australians de temps recents, que sí que es reconeix en restes fòssils més recents. Probablement la colonització d’Austràlia fou un complex fenomen de diferents arribades i extincions de grups. Per exemple, l’arribada del dingo, un gos mig domesticat i estretament lligat als ahorígens, es va produir fa només entre 3 000 i 4 000 anys. I les nombroses extincions de mamífers marsupials semblen estretament lligades a l’activitat depredadora dels humans.
El procés de poblament d’Amèrica
El poblament d’América és un dels grans temes de discussió. Si bé es reconeix que es féu a través del que actualment és l’estret de Bering i, per tant, a partir de poblacions asiàtiques, la datació de la primera entrada és controvertida. L’ocupació generalitzada del continent americà fa uns 12 000 anys s’accepta plenament, però el dubte rau en datacions anteriors, algunes situades al voltant dels 40 000 anys. Durant l’última glaciació un pont de terra ferma de més de 1 000 km d’amplada unia Sibèria i Alaska, i pogué facilitar el pas; el problema, però, és l’ocupació de terres més cap al S, ja que una gran capa de gel cobria el que actualment és el Canadà. S’ha postulat que si la penetració fou fa 25 000 anys o més, es pogué seguir un estret corredor sense gel que separava les dues grans plaques de gel a l’E del Canadà; el pas havia de restar pràcticament impracticable en el pic de la glaciació, entre 25 000 i 14 000 anys enrere.
Entre les dues propostes hi ha una gran separació en el temps i, de moment, no hi ha consens a l’hora d’acceptar o rebutjar els casos pretesament més antics, com per exemple alguns jaciments del Brasil (abric de Pedra Furada, al S de l’estat de Piauí) o de Xile (Monte Verde, a la valla del riu Maullin, a la província de Los Lagos). Si la data tardana és correcta, l’expansió es degué produir molt de pressa, ja que jaciments molt australs tenen datacions semblants als del nord. De tota manera, la velocitat d’expansió sembla compatible amb la mobilitat associada a grups de grans caçadors i possiblement pogueren recórrer de cap a cap el continent en pocs milers d’anys.
L’impacte de l’arribada de l’home a Amèrica, o almenys de la clara expansió que s’observa fa entre 12 000 i 10 000 anys, és manifest en la massiva extinció de grans mamífers. Tot i que coincideix amb el canvi climàtic del final de la darrera glaciació, tot fa pensar que l’acció humana directa en forma de cacera fou molt important. De fet, la capacitat caçadora i, en general, d’adaptació de les poblacions del Paleolític superior feren sentir poderosament la seva acció en termes d’extinció d’espècies a tot arreu, però Amèrica sembla presentar, per grans àrees, l’exemple més clar.
Gens, pedres i llengües per reseguir la colonització humana de la Terra
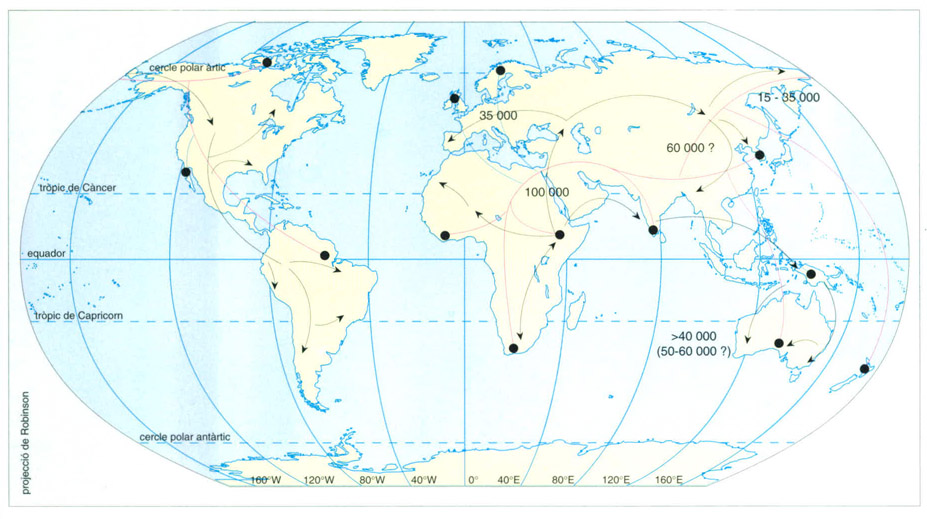
Editrònica, a partir de Cavall-Sforza, 1992
Gens, pedres i llengües ens expliquen una història semblant. L’enorme diversitat de poblacions humanes que hi ha sobre la superfície del planeta és un reflex del que succeí en temps passats quan els humans moderns s’assentaren als diversos indrets que havien colonitzat i començaren a diferenciar-se els uns dels altres. Els humans, però, no sols portaven amb ells llur dotació genètica, sinó que també duien una enorme empremta cultural. El fet cultural, transmès de manera extrasomàtica, pot do-nar força informació sobre l’origen de cada persona. Un dels fenòmens no genètics més característics de l’espècie humana és el llenguatge; aquests estudis mostren que la distribució de les llengües és totalment equiparable a la distribució genètica (excepte pocs casos en què hi ha hagut penetració d’una llengua sense penetració genètica o viceversa), ja que, quan Homo sapiens colonitzà el planeta dugué amb ell el seu llenguatge que, en romandre aïllat, es diferencià molt més que la genètica. El primer arbre genètic (vermell) s’ha projectat sobre un mapamundi i els seus extrems acaben sobre poblacions actuals (punts negres). Treballs més recents semblen assenyalar que hi hagueren dues vies migratòries d’Àfrica a Àsia (negre); si bé no tenim proves detallades de les rutes seguides, les aproximacions s’han fet amb estimacions arqueològiques.
La diversificació de l’espècie humana actual
Així com l’inici del camí evolutiu humà presentava certes dificultats i una necessitat de justificació, el final també és difícil de delimitar o precisar. Després d’haver vist l’aparició de les formes modernes, que relacionem directament amb les poblacions humanes actuals, i llur expansió per tota la Terra podríem donar per acabada la descripció. Però podem no donar-nos per satisfets. Podem, per exemple, resseguir més finament no sols l’evolució a l’escala de l’espècie, sinó dels grups humans, tant si els anomenem races com si els anomenem simplement poblacions.
El problema és que, com si fos una fractal la complexitat de la qual no disminueix quan s’observen detalls en comptes de conjunts, la història dels grups humans ateny una complexitat immensa si pretenem reconèixer detalladament el procés de formació, la relació amb altres grups, les migracions i barreges, les adaptacions. Altrament, a les consideracions estrictament antropològiques se sumen aleshores molts factors culturals i el fenomen —al capdavall també cultural— de les continues mescles entre les races o poblacions, sobretot d’uns segles, o fins i tot només decennis, ençà. Davant d’això, l’antropologia física fa servir altres eines, com la genètica de poblacions, per a aconseguir marcs més precisos, però l’aproximació global necessita encara altres instruments, com la prehistòria i història o la lingüística històrica per tal de donar imatges coherents i precises.
En tot cas, l’antropologia física, avui, fonamentant-se en la constatació de fenòmens genètics i bioquímics, tendeix a relativitzar la importància de certes manifestacions anatòmiques o morfològiques de les poblacions, tals com la pigmentació o els trets facials, per exemple. Això equival a dir que minimitza els elements que, tradicionalment, eren presos com a determinants a l’hora d’establir les preteses races humanes. Tant és així, que el mateix concepte de raça ha fet crisi entre els antropòlegs físics fins a esdevenir obsolet atesa la impossibilitat objectiva d’aplicar-la a les poblacions d’una espècie com la nostra en la qual és excepcional l’endogàmia de cap població i en canvi són fenòmens corrents les migracions i els intercanvis gènics d’un cap a l’altre del planeta. Com ja escrivia Johann Gottfried von Herder el 1784: “El terme raça fa referència a una diferència originària inexistent [...]; en una paraula, les races humanes no són quatre, ni cinc, ni varietats exclusives [...]; els tipus físics s’interpenetren i segueixen els trets hereditaris i no són finalment més que les ombres d’un gran miratge que s’estén sobre totes les edats i tots els continents; pertanyen menys a un sistema d’història natural que a una història física i geogràfica del gènere humà.” Avui només les estretes percepcions de les ideologies racistes donen a fenòmens com el color de la pell una rellevància, totalment injustificada, com a element caracteritzador de grups humans substancialment diferents. És cert que històricament, des de la interpretació esclavista que “els negres no tenen ànima” (és a dir, que no pertanyen a l’espècie humana) o de les jerarquitzacions racials fetes pels nazis, fins a la pràctica desestimació del concepte de raça postulada pels antropòlegs moderns, passant per les classificacions elementals de races blanca, negra, groga i roja o altres de més elaborades (i/o interferides amb criteris etnològics), els humans hem establert tota mena de judicis i de prejudicis a propòsit de la seva mateixa realitat física. Tanmateix, dels avenços en el coneixement científic d’aquesta realitat en resta una sola evidència incontestable: hi ha hagut diverses espècies d’humans en un passat remot, però actualment (en una actualitat que dura pel cap baix 40 000 anys) només n’hi ha una, la qual es manifesta sota morfologies molt semblants, encara que no pas idèntiques, atès que les datacions genètiques de les poblacions varien geogràficament.