Concepte d’amfibi

Els amfibis són els primers vertebrats que en l’escala evolutiva han aconseguit colonitzar el medi terrestre. La vida a terra ferma requereix una llarga sèrie d’estructures especialitzades que inclouen diverses modificacions de l’esquelet per a suportar el pes del cos, com l’aparició dels membres quiridis, articulats i aptes per a la locomoció i, fins i tot, en algunes espècies, per al salt, com en la reineta comuna (Hyla meridionalis) de la fotografia.
Javier Andrada
Els amfibis són els primers vertebrats que han reeixit a sortir de l’aigua, almenys parcialment. En algun moment de la història de la Terra, algun grup de peixos es veié forçat a ocupar el medi terrestre, que llavors era disponible i lliure de competència per a aquests animals, per bé que oferia la presència de plantes i invertebrats de diversos grups, que ja havien dut a terme el mateix pas. Tanmateix, el procés evolutiu de l’aigua a la terra, que degué ser prou gradual, però no lent, ha deixat pocs rastres, i això és el que ha dificultat la individualització del grup, del qual encara es discuteix l’origen monofilètic o polifilètic, és a dir, si el poblament de la terra ferma per part dels vertebrats fou iniciat en un punt o en més d’un punt i a partir d’un o de més d’un grup de peixos.
El terme amfibi deriva del grec i significa, lliurement traduït, "doble vida", la qual cosa es refereix a les fases aquàtica i terrestre presents al cicle biològic de la majoria d’amfibis actuals. Fou enunciat per Linné, que hi agrupà una barreja heterogènia de vertebrats més o menys parcialment aquàtics i amb certes similituds morfològiques. Més tard, Latreille restringí l’ús del terme a les granotes, els gripaus, les salamandres i els tritons, als quals foren afegides, més tard, les cecílies; així quedaren constituïts els amfibis vivents actuals, que són relativament homogenis. El terme batraci, que deriva igualment del grec i equival a "granota", els fou aplicat també durant un temps, però ha anat perdent-se a poc a poc perquè era inadequat.
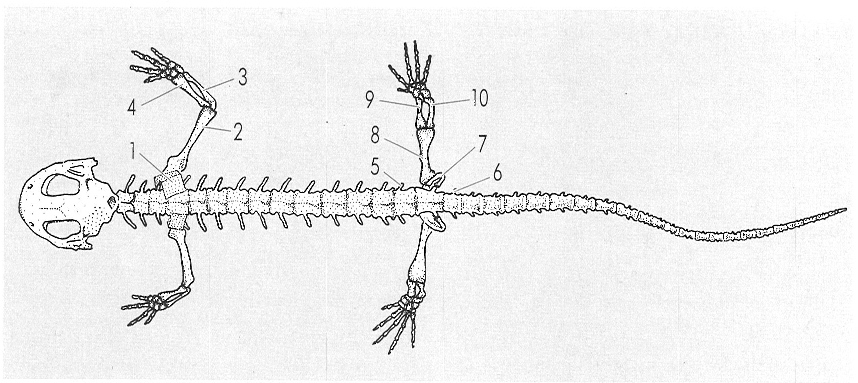
Entre les modificacions aparegudes en el pas a la vida terrestre destaquen les que afecten l’esquelet (de salamandra, en el dibuix), com les de les cintures i les potes, les de l’estructura de la columna vertebral i de cadascuna de les vèrtebres, i l’escurçament del crani. Les potes mostren els tres segments característics, amb la mà de quatre dits i el peu de cinc dits. 1 Omòplat, 2 húmer, 3 cúbit, 4 radi, 5 costella sacra, 6 primera vèrtebra caudal, 7 ili, 8 fèmur, 9 tíbia, 10 peroné.
Ferran Rodríguez, original de Schaeffer.
La sortida de l’aigua exigeix solucions a una sèrie de problemes plantejats per la baixa densitat de l’aire, com ara el desenvolupament de membres que suportin el pes corporal, la constitució d’un tegument amb glàndules que impedeixin la seva dessecació i d’un aparell respiratori intern, pulmonar, solucions per a la reproducció i el desenvolupament, i encara d’altres. Com sigui que en les formes fòssils la informació disponible concerneix especialment les restes òssies, hom coneix prou bé l’evolució esquelètica dels amfibis per a adaptar-se a la terra ferma però desconeix quasi completament el seu procés d’evolució fisiològica per a la vida aèria.
Els amfibis vivents mostren que el grup ha resolt prou aspectes d’aquesta sèrie de problemes, però no ha aconseguit una independència total de l’aigua. La reproducció els exigeix tornar a l’aigua. La fecundació és externa en molts casos i quasi sempre té lloc en medi aquàtic, i els ous, amb poques reserves nutritives, es desclouen en una larva estrictament aquàtica i pisciforme, que sofrirà una profunda metamorfosi que afectarà la morfologia, l’anatomia i la fisiologia de l’animal fins a esdevenir adult. Aquest adult tampoc no aconsegueix una total independència de l’aigua: té la pell nua i en molt pocs casos hi presenta algunes escates; compta, però, amb unes glàndules que ajuden a mantenir-la humida. L’ull s’acompanya de parpelles que el protegeixen de la dessecació. La respiració és pulmonar, però, pel fet que els pulmons són de reduïda eficàcia, es complementa amb una respiració cutània importantíssima, que és possible gràcies a la nuesa de la pell, la qual, en algun cas com el de certs urodels cavernícoles, és l’únic òrgan respiratori. La circulació segueix un circuit doble, però el cor té dues aurícules i un sol ventricle, per la qual cosa hi ha un cert grau de mescla de sang oxigenada i sang carbònica. Els amfibis són ectotèrmics i no aconsegueixen regular fisiològicament llur temperatura corporal, sinó que ho fan mitjançant mecanismes ecoetològics, i sovint necessiten hivernar o estivar.
Els amfibis són vertebrats tetràpodes, proveïts de quatre membres locomotors, fonamentalment pentadàctils, destinats al desplaçament sobre terra ferma durant llur vida adulta. El seu crani s’articula a la vèrtebra atlas, mitjançant dos còndils occipitals. Presenten una important reducció de peces esquelètiques i, al mateix temps, una forta unió entre la cintura pelviana i les vèrtebres sacres. Si bé les formes actuals poden ser caracteritzades per certes particularitats anatòmiques i biològiques i el grup sembla ben definit, no ocorre el mateix si hom considera simultàniament les formes fòssils.
Els amfibis actuals són només una minsa representació del que foren des de llur aparició al final de l’Era Primària fins al final de la Secundària. D’aquests primers representants del grup no en queden gaires restes completes, però les que posseïm són suficients per a demostrar l’abundància i la diversitat dels amfibis en el passat. Molt sovint és difícil separar-los dels seus antecessors, els peixos, o dels seus descendents, els rèptils, que aparegueren igualment a finals del Primari, no gaire més tard. Es produí una radiació evolutiva amplíssima que donà similituds notables entre els diversos ordres i classes, i, per això, la separació d’amfibis i rèptils fòssils és molt difícil, perquè no poden observar-se, en general, detalls relacionats amb la biologia reproductiva, el fet diferencial més important. Els grups actuals, en canvi, residuals en ambdós casos, són ben caracteritzats i prou homogenis. És evident que els amfibis d’avui no són antecessors dels rèptils; aquests procedeixen d’algun d’aquells grups primitius que cal considerar avantpassats comuns dels representants actuals de tots els vertebrats.
Hom classifica els amfibis vivents en tres grups: cecílies o àpodes (gimnofions, Gymnophiona, amb 150 espècies, de distribució tropical i subtropical), sireniformes, tritons i salamandres (urodels, Urodela, amb unes 310 espècies arreu del món) i granotes i gripaus (anurs, Anura o Salientia, amb 2510 espècies conegudes). La seva mida oscil·la entre 1 cm i 1 m de llargada, i han aconseguit d’adaptarse a força medis; es troben repartits quasi arreu del món i, en alguns casos, arriben a ésser bastant independents de l’aigua. Es troben pertot arreu tret de les regions més fredes i les més àrides.
Els models morfològics
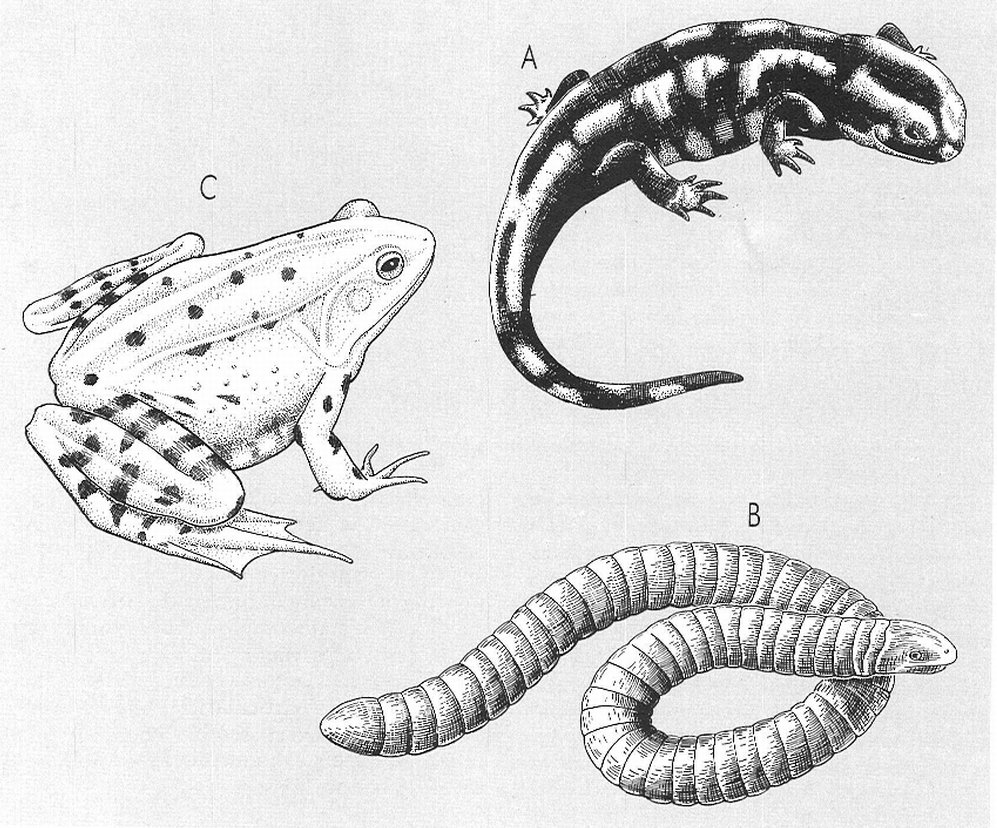
Models morfològics que trobem en els amfibis actuals: A salamandriforme, que correspon als urodels; B serpentiforme o vermiforme, que correspon als gimnofiònids (exòtics); i C raniforme, que correspon als anurs.
Amadeu Blasco, original de Canka i Vit.
Cadascun dels grups actuals d’amfibis presenta una morfologia bàsica diferent, a més d’una gran variabilitat dins de cada patró, per les adaptacions corresponents a les diverses maneres de viure. Els tipus principals vénen determinats per l’estructura lacertiforme (urodels), la raniforme (anurs) i la vermiforme (gimnofions).
Els urodels lacertiformes
Els urodels lacertiformes presenten un cap ample, un tronc diferenciat, quatre potes de llargada equivalent —que en algun grup poden ser molt reduïdes— amb quatre dits als membres anteriors i cinc als posteriors, i una cua de secció circular, en els animals de costums més terrestres (de cos més robust) o comprimida, en els d’hàbit aquàtic més marcat (que són, al mateix temps, animals de cos més esvelt), que persisteix durant tota la vida de l’animal. Els mascles d’algunes espècies desenvolupen, durant el període de zel, crestes caudals i dorsals de coloracions característiques.
Els anurs raniformes
Els anurs raniformes presenten, en general, cap i cos sense solució de continuïtat, més aviat robustos, i no tenen cua en estat adult; les potes posteriors, amb cinc dits, són —com a conseqüència de l’adaptació al salt— molt més llargues que les anteriors, amb quatre dits. El tipus raniforme presenta dues varietats ben definides: la granota pròpiament dita i el gripau, identificables, fins i tot, pel coneixement popular. La granota és més esvelta, de potes més llargues i més aquàtica; el gripau és més rabassut, amb potes posteriors proporcionalment més curtes i d’hàbits més terrestres. Aquestes dues formes són els extrems d’un grup on apareixen moltes espècies que presenten caràcters intermedis.
Els àpodes
Els àpodes són amfibis excavadors completament desproveïts de membres, amb més similitud, des del punt de vista morfològic, a un oligoquet que no a un amfibi anur o urodel—a causa de llur pell plegada—; s’acantonen a les zones càlides de la terra, presenten una coloració molt vistent i no arriben als 50 cm de llargada.
Les característiques i les adaptacions anatòmiques dels amfibis
El tegument
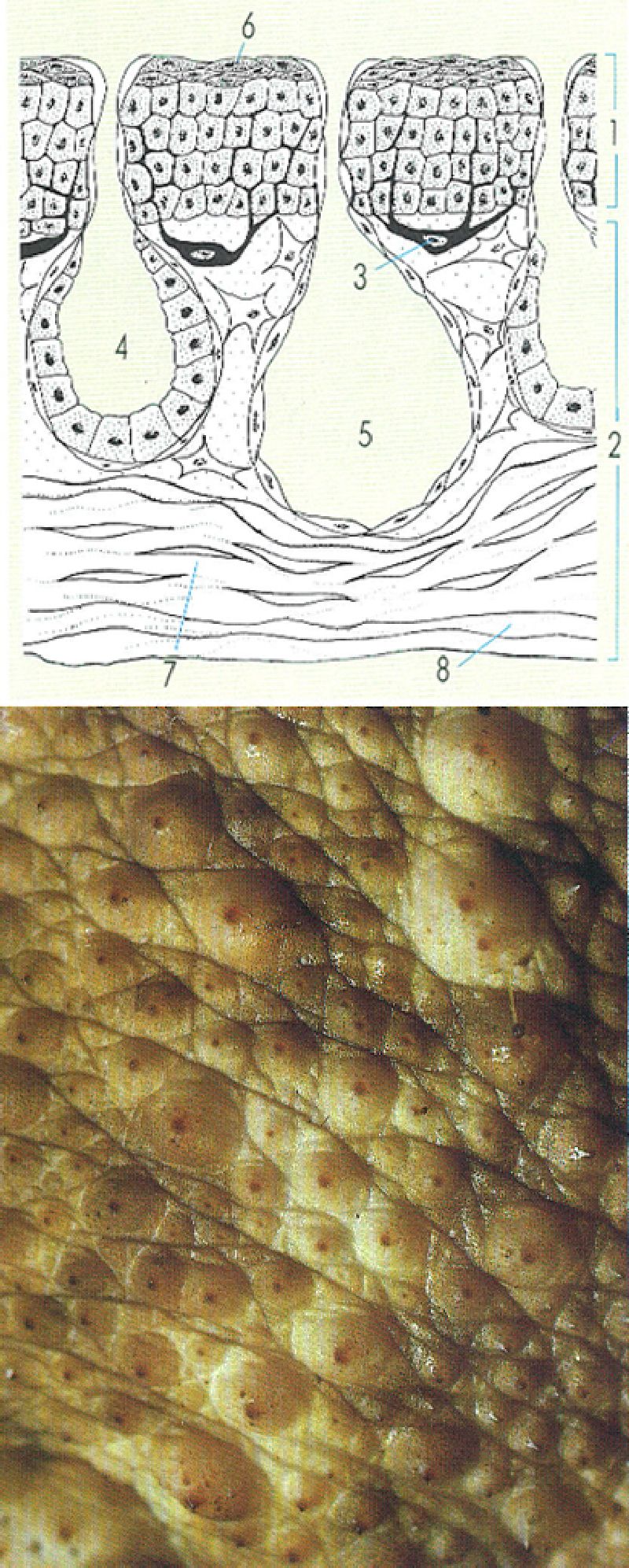
El tegument dels amfibis actuals es compon de l’epidermis i la dermis típiques dels vertebrats. La primera capa és ceratinitzada, per la qual cosa la pell resulta relativament permeable als gasos i també a l’aigua. Les glàndules, tant les mucoses (la seva secreció és responsable del tacte llefiscós dels amfibis) com les verinoses (amb diverses toxines), són molt abundants. També són nombrosos els cromatòfors, responsables de les sovint vives coloracions d’aquests animals. En el dibuix: 1 epidermis, 2 dermis, 3 cromatòfor, 4 glàndula mucosa, 5 glàndula de verí, 6 estrat corni, 7 estrat esponjós, 8 estrat compacte. A baix, veiem un detall de la pell d’un gripau comú (Bufo bufo), especialment rugosa i amb les puntes ceratinitzades.
Ferran Rodríguez, original de Goin
La pell dels amfibis és nua. Només els gimnofions hi tenen algunes escates embegudes. És formada per l’epidermis i la dermis típiques dels vertebrats, amb un estrat corni molt prim, cosa característica del tegument als animals terrestres, que evita la dessecació. La pell se sotmet a mudes periòdiques.
La muda es desprèn sencera o a trossos, i sembla que els animals se la mengen després; es tracta de cèl·lules mortes i ceratinitzades que protegeixen les cèl·lules vives de l’epidermis, disposades pluristratificadament.
La dermis subjacent és prima i consisteix en dues capes, una d’esponjosa, externa, i una de compacta, interna, recorregudes per un gran nombre de vasos sanguinis capil·lars que li permeten acomplir la funció respiratòria que té encomanada; igualment la dermis conté els cromatòfors, cèl·lules responsables de la coloració de la pell de l’amfibi, que comprenen diversos pigments.
La pell és constantment protegida per la secreció mucosa de les glàndules corresponents, pluricel·lulars, que mantenen la humitat necessària per a l’intercanvi de gasos. El mucus cutani també conté la secreció de les glàndules verinoses, que consisteix en toxines químicament molt diverses (amines, pèptids, esteroides, alcaloides) amb efectes farmacològics també diferents (cardiotòxics, miotòxics, neurotòxics i hemotòxics), la funció de les quals no és solament la defensa davant els predadors naturals, que aconsegueixen amb un cert èxit, sinó, sobretot, la defensa davant l’acció microbiana que es produeix a la pell. Normalment, les toxines es troben en una quantitat molt petita i llurs efectes en l’home, a tan baixa concentració, són menyspreables.
En alguns casos excepcionals, poden aparèixer engruiximents de l’estrat corni, ceratinitzacions a manera d’ungles falses, esperons o estructures semblants, com ocorre precisament en el nostre gripau d’esperons.
L’esquelet i la musculatura
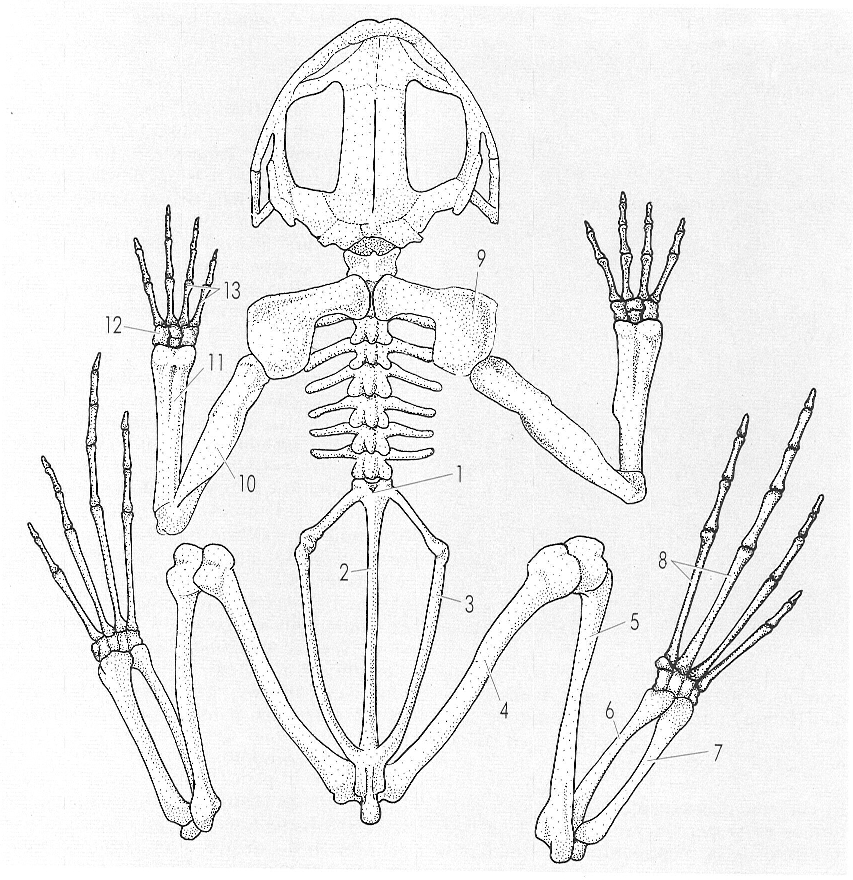
Esquelet d’amfibi, concretament d’una granota (anur). Hom ha indicat en el dibuix el nom d’aquelles parts que tenen un interès especial en aquest grup, especialment la columna vertebral i la cintura pelviana. És notable l’escurçament general del cos. 1 vèrtebra sacra, 2 urostil, 3 ili, 4 fèmur, 5 tíbio-fíbula, 6 astràgal, 7 calcani, 8 metatarsià, 9 suprascapular, 10 húmer, 11 ràdio-ulnac, 12 carp, 13 metacarpians.
Ferran Rodríguez, a partir de fonts diverses.
En el pas d’aquests animals a la terra, l’esquelet té un paper fonamental, perquè, pel fet que l’aire és molt menys dens que l’aigua, apareix la necessitat de membres ben desenvolupats per a poder suportar el pes del cos i esdevenen imprescindibles una ossificació i una solidesa general de tot l’esquelet molt superiors a les dels peixos alhora que una millor articulació. Respecte dels peixos, totes les parts de l’esquelet han sofert profundes modificacions.
El crani és aplatat i presenta un nombre d’ossos inferior al dels peixos; el condrocrani ha estat format per teixit ossi. La suspensió mandibular és autostílica, de manera que l’os articular de la mandíbula contacta amb el quadrat del crani (és la típica dels tetràpodes, excepció feta dels mamífers).
La columna vertebral presenta regions diferenciades, la qual cosa és general als vertebrats terrestres i conseqüència de l’evolució dels membres locomotors. Els amfibis tenen una vèrtebra cervical, una sèrie de vèrtebres del tronc, una vèrtebra sacra i, en el cas dels urodels, una regió caudal. La regió cervical presenta la vèrtebra atlas articulada al crani mitjançant dos còndils; les vèrtebres dorsals presenten facetes anteriors i posteriors per a un millor suport del tronc; la vèrtebra sacra presenta apòfisis transverses per a articularse amb la cintura pelviana. En els anurs —que presenten una estructura peculiar, l’urostil, amb pèrdua de la cua— existeix una forta reducció numèrica de vèrtebres. També presenten costelles, però, en general, poc desenvolupades; són més llargues en els gimnofions, més reduïdes en els urodels i pràcticament absents en els anurs, en els quals normalment només hi ha petites estructures cartilaginoses. L’estèrnum és poc desenvolupat i només es forma una mena de caixa toràcica en alguns urodels i anurs.

L’estructura esquelètica i muscular dels amfibis els permet la locomoció i el sosteniment del pes del cos en la vida a terra. Les potes són pentadàctiles i, en el cas dels anurs, com el de la fotografia, una reineta comuna (Hyla meridionalis), l’adaptació al salt implica una llargària notablement superior dels membres posteriors respecte dels anteriors. La reineta té, a més, els extrems dels dits en forma de coixinet adhesiu, cosa que li permet grimpar per superfícies completament llises.
Hàbitat/Xavier Moreno
Un aspecte molt important de l’esquelet postcranial dels vertebrats terrestres el constitueixen les cintures i les potes. La cintura pectoral és discreta, desconnectada del crani i relacionada amb la columna vertebral per un lligament muscular. La cintura pelviana és més forta i s’articula amb la columna vertebral a la vèrtebra sacra. Les potes corresponen al model pentadàctil existent a tots els vertebrats tetràpodes, amb diverses modificacions segons el medi de vida dels animals. Aquest model consisteix en un os únic (húmer o fèmur) que s’articula distalment amb dos ossos, de llargada lleugerament inferior i disposats paral·lelament (cúbit i radi o tíbia i peroné), que són completats per una sèrie d’ossos petits destinats a l’articulació (carpians i tarsians), i per una altra sèrie d’ossos alineats i allargats en cinc estructures (metacarpians i metatarsians), que acaben en els dits. En els amfibis, aquest model, llevat de les formes àpodes o d’algunes reduccions fortes, com la que es produeix en els urodels sirènids, només presenta la pèrdua d’un dit a cada mà o, en el cas dels saltadors, els anurs, l’allargament excessiu dels ossos llargs, tant del primer (fèmur), articulat a la cintura, com dels dos que el segueixen, articulats a ell (tíbia i peroné), a la pota posterior.
El caràcter més notable respecte dels pisciformes és la pèrdua progressiva de la segmentació de la musculatura. És simultàniament necessària, per a dur a terme els moviments útils obligats en el desplaçament a terra, l’aparició de músculs especialitzats a les potes i al tronc, i una coordinació molt complexa de l’activitat muscular.
Els sistemes de relació i control
El sistema nerviós
L’evolució del sistema nerviós porta a un augment, en volum i en qualitat, dels hemisferis cerebrals. Davant d’aquests, hi ha un parell de lòbuls olfactius. Les modificacions respecte de l’encèfal dels peixos són de significació discreta, però, en els hemisferis cerebrals, el pal·li, que en constitueix el sostre i els costats, comença a presentar algunes neurones disperses que, evolutivament, s’han desplaçat de l’interior a l’exterior. Els lòbuls òptics representen un volum important de l’encèfal. També el cerebel és prou desenvolupat, igual que la medul·la, amb vista a poder controlar la complexitat motora dels animals terrestres i els membres locomotors pròpiament dits. La medul·la és llarga en els urodels; en els anurs, és curta i acabada en un feix de fibres nervioses en cua de cavall.
Els amfibis presenten només deu parells de nervis cranials, a diferència de la resta de vertebrats terrestres. En incrementar-se la complexitat de la locomoció terrestre, la nervació és també molt complexa i a les regions cervical i lumbar es formen uns eixamplaments medul·lars per on entren i surten les fibres sensitives i motores que innerven les extremitats. Hi ha deu parells de nervis espinals.
Els òrgans sensorials

Les diferents formes de pupil·la constitueixen un caràcter útil en la determinació de les diverses famílies d’anurs. El gripau d’esperons (Pelobates cultripes, a l’esquerra), per exemple, la té vertical; en canvi, el gripau comúx (Bufo bufo, a la dreta) la té horitzontal. També poden ésser quasi triangulars o bé cordiformes.
Ramon Torres i Javier Andrada
L’ull presenta glàndules al seu voltant, que serveixen per a humidificar la còrnia, i parpelles mòbils (excepte a les formes completament aquàtiques i a les larves), i que són una adaptació a la vida terrestre i protegeixen el globus ocular, que és protràctil i retràctil. En els animals excavadors o manifestament hipogeus, els ulls es redueixen, com ocorre en els gimnofions. Altrament, l’estructura de l’ull dels amfibis és la típica dels vertebrats terrestres.
Les larves i urodels aquàtics conserven una línia lateral funcional, que no es troba en els amfibis terrestres.
>

El timpà, membrana que transmet les vibracions sonores, no és encara patent en tots els amfibis, però destaca clarament en algunes espècies. Hom pot apreciar, a la fotografia, el d’una granota comuna (Rana perezi), situat darrere l’ull.
Enric Curto
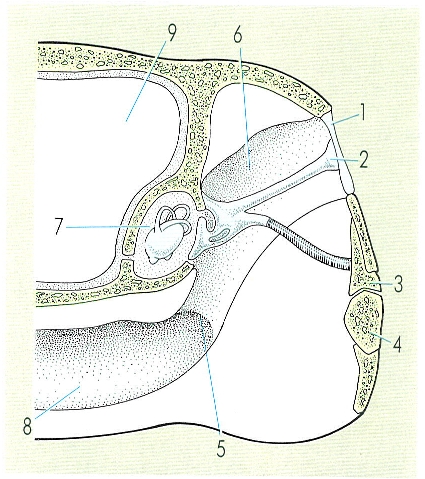
Esquema de l’oïda d’un amfibi. Hom hi indica les parts principals: 1 timpà, 2 estrep, 3 quadrat, 4 articular, 5 trompa d’Eustaqui, 6 oïda mitjana, 7 oïda interna, 8 gola, 9 cavitat cranial.
Amadeu Blasco segons Romer
Hi ha importants modificacions en l’oïda dels amfibis respecte de la dels peixos. L’oïda mitjana deriva de la primera bossa faríngia i és una cavitat plena d’aire i connectada amb la boca per la trompa d’Eustaqui, que equilibra les pressions a una banda i l’altra del timpà. Conté la columel·la, que és l’os que transmet el so. L’oïda interna, inclosa dins del crani, presenta dues finestres cobertes de membrana flexible, les quals limiten la cavitat perilimfàtica: són la finestra oval, damunt la qual es recolza la columel·la, que toca per l’altre extrem el timpà, i la finestra rodona, a la part posterior. La cavitat endolimfàtica presenta la papil·la basilar i la papil·la amfibiana sensibles al so. La vibració es transmet del timpà a la columel·la, i d’aquesta a la finestra rodona, i pot fer vibrar les papil·les i l’endolimfa; l’eficàcia d’aquest sistema és superior en els anurs, que capten vibracions d’alta i baixa freqüències procedents de l’entorn i de les emissions sonores de la mateixa espècie.
La informació química es rep mitjançant papil·les gustatives. Els amfibis semblen distingir els sabors bàsics (dolç, salat, amarg i àcid). Es troba, per primer cop a l’escala evolutiva, l’òrgan de Jacobson, consistent en dos sacs cecs connectats mitjançant els corresponents conductes amb la cavitat nasal i innervats parcialment per una branca del nervi olfactiu; la seva funció és probablement relacionada amb el reconeixement de l’aliment.
Les glàndules endocrines
No presenten peculiaritats respecte de les d’altres vertebrats tetràpodes. Les paratiroides apareixen, però, per primer cop en els amfibis, per al control del calci a la sang. La tiroide és especialment important en la metamorfosi i en l’ecdisi.
L’aparell digestiu
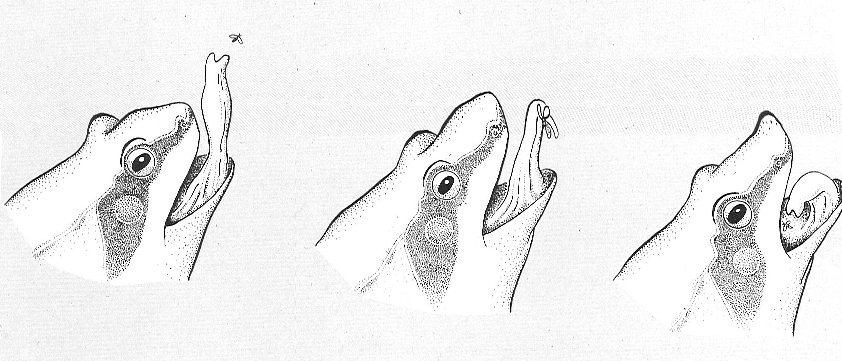
Seqüència de captura de l’aliment per una granota. La llengua. d’implantació anterior, és desplegada cap endavant fins adherir-se a l’insecte, que aleshores és transportat a la boca i engolit. Els amfibis cacen a l’aguait, encara que els anurs poden córrer o saltar per capturar llurs preses.
Amadeu Blasco.
La possessió de llengua és producte de l’adaptació a la vida terrestre, on cal humidificar i, si s’escau, mastegar l’aliment abans d’engolirlo per tal d’evitar lesions a les vies digestives. Aquesta llengua desenvolupa una musculatura pròpia que, en els amfibis, s’implanta a la part anterior de la boca, a la mandíbula, i es plega al terra d’aquella; sovint és ràpidament projectable a l’exterior per a la captura de l’aliment. Presenta, però, formes variables segons els grups, des de la llengua discoïdal dels discoglòssids fins a la quasi vermiforme dels bufònids, i les seves capacitats són relacionades amb la forma.
Les dents apareixen ja a les larves, però no són dents pròpiament dites sinó denticles cornis epidèrmics. Les dents vertaderes només apareixen en els adults —són formades aleshores per una cavitat de la polpa, amb irrigació sanguínia i terminacions nervioses, dentina i esmalt, implantades als maxil·lars o al sostre de la boca (dents vomerianes), en diferents posicions segons les famílies— o bé poden mancar completament. Els amfibis són polifiodonts, perquè renoven les dents diverses vegades, i són homodonts, perquè tenen totes les dents idèntiques. Tenen una zona feble entre la corona i l’arrel; la funció de la dent no és mastegadora, sinó de subjecció de la presa.
El tub digestiu presenta una cavitat bucal i un esòfag revestits de cilis; l’esòfag és molt curt. L’estómac és encara poc diferenciat, però presenta glàndules gàstriques amb una secreció apta per a la digestió; en els anurs, es diferencien els extrems cardíac i pilòric. L’intestí és de longitud variable, segons el tipus d’alimentació, i, per tant, més o menys sinuós. La diferenciació entre l’intestí prim i l’intestí gros, tant morfològica com fisiològica, comença a aparèixer en els urodels i només es marca bé en els anurs. L’intestí desemboca en una cavitat, l’anomenada cloaca, on van a parar igualment els aparells excretor i reproductor.
L’aparell respiratori
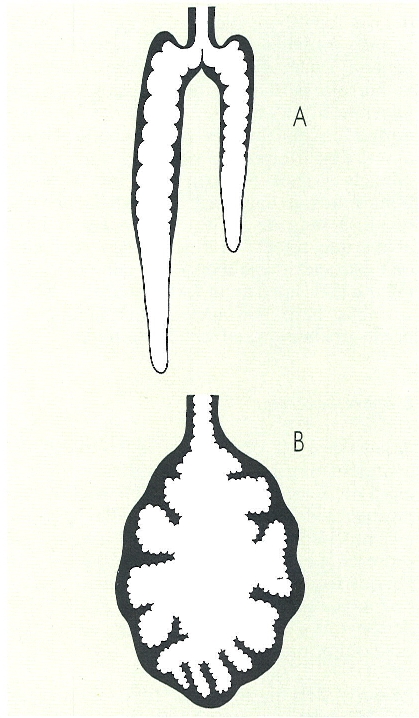
L’eficiència del parell de pulmons profundament introduïts en el cos, que es constitueixen per respirar l’aire atmosfèric evitant la deshidratació excessiva, és encara limitada en els amfibis; així, una fracció, sovint important, de l’intercanvi de gasos es realitza a la pell. El dibuix mostra les causes d’aquesta situació: el pulmó d’un urodel terrestre (A) té una saculació reduïda, mentre que en els anurs (B) els alvèols són una mica més desenvolupats.
Gustavo Hormiga, originals de Porter.
Per a l’intercanvi de gasos, són necessàries superfícies contínuament humides que posin en contacte el medi intern i el medi extern; cal, per aquest motiu, que siguin molt vascularitzades. Les larves aquàtiques dels amfibis respiren mitjançant brànquies, mentre que els adults posseeixen pulmons interns. De tota manera, la funcionalitat d’aquests pulmons no presenta una gran eficiència. Això fa que la pell i, fins i tot, el tegument de la cavitat bucal actuïn a manera d’aparell respiratori.
Les brànquies externes de les larves d’amfibi es formen damunt dels arcs branquials i són expansions tegumentàries (per tant, recobertes d’ectoderma). Són més simples en els anurs que no en els urodels. En els anurs, apareix un opercle branquial als pocs dies del desenvolupament larval i es constitueix una cambra branquial interna (que persisteix fins a la metamorfosi), oberta a l’exterior a través d’un espiracle, on es formen brànquies noves. Aquestes brànquies es reabsorbeixen durant la metamorfosi dels anurs i llur funció és substituïda pels pulmons de nova formació i pel tegument. En els urodels, les brànquies romanen externes durant tota la vida larval i, en la metamorfosi, també desapareixen en general, bé que en els urodels es donen alguns casos de neotènia, en els quals l’individu ja reproductor continua vivint a l’aigua i conserva caràcters larvals, com les brànquies. Algunes salamandres cavernícoles europees no tenen pulmons i la seva respiració és exclusivament cutània.
Els pulmons es formen durant la metamorfosi i són estructures senzilles, amb pocs alvèols i amb una superfície respiratòria reduïda, que, proporcionalment, és superior en els anurs que en els urodels. Les vies respiratòries dels amfibis són les típiques de tots els vertebrats tetràpodes. Els orificis nasals externs poden ser controlats a voluntat de l’animal, que els obre o els tanca o en varia el diàmetre, la qual cosa li permet, per exemple, de submergir-se sense problemes. Les coanes, orificis nasals interns, són característiques dels vertebrats terrestres i, en aquest cas, connecten directament amb la cavitat bucal. Segueixen la faringe, la laringe, amb la glotis per a protegir les vies respiratòries durant la deglució i els cartílags cricoide i aritenoide. Aquests presenten cordes vocals, que són dues bandes musculars, paral·leles a la glotis, que vibren amb el pas de l’aire i permeten una emissió sonora variada, especialment patent en els anurs, en algunes famílies dels quals hom troba un o dos sacs vocals, amb funció de ressonadors, en posició gular o comissural, normalment en els mascles. La tràquea, més curta en els anurs, el cos dels quals presenta un escurçament general notable, s’obre en dos bronquis, que es ramifiquen en les cavitats pulmonars simples.
El tegument és altament vascularitzat i això permet un intercanvi directe de gasos amb l’aire, sempre que es mantingui humit; en certs casos, si bé excepcionalment, la mucosa bucal també pot actuar com una superfície respiratòria. A terra ferma, per respirar, els animals tanquen la boca, inspiren l’aire pels orificis nasals i, amb una depressió del terra de la boca, l’engoleixen: els cal, aleshores, impulsar-lo encara cap als pulmons, aixecant novament el terra bucal. S’estableix, així, un moviment de deglució d’aire.
L’aparell circulatori
La circulació dels amfibis és doble i incompleta. La sang circula per dos circuits. El primer i més breu va del cor als pulmons i novament al cor, per a l’intercanvi de gasos, i el segon i més llarg va des del cor a tots els òrgans i retorna, per al transport d’oxigen a totes i cadascuna de les cèl·lules.
El cor té tres cambres o cavitats, dues aurícules i un ventricle, de manera que persisteix una certa barreja de la sang oxigenada amb la no oxigenada. L’aurícula esquerra rep la sang que ha recorregut tot el cos de l’animal i la impulsa cap al ventricle, el qual l’envia als pulmons per a l’oxigenació, d’on després retorna a l’aurícula dreta, i aquesta la impulsa novament al ventricle, que ara ha d’enviar-la als diversos òrgans. La barreja de sang és, però, relativament petita, perquè l’obertura no sincrònica de les vàlvules de comunicació entre les cavitats i les contraccions lleugerament decalades d’una banda i l’altra del cor l’eviten en bona part. D’altra banda, cal dir que a la sortida del ventricle la sang passa pel bulb aòrtic, que presenta, longitudinalment, una vàlvula espiral que contribueix a separar la sang destinada a l’arc sistèmic i al carotidi de la destinada a l’arc pulmonar. Així mateix, cal dir que la sang que arriba al cor procedent del cos rep, abans d’entrar a l’aurícula dreta, la sang provinent de la pell i, per tant, oxigenada, i que, d’aquesta manera, l’efecte de la barreja perd importància fisiològica. De l’aorta ventral, en surten quatre parells d’arcs aòrtics en els urodels i tres parells en els anurs. Les artèries que porten la sang als pulmons la dirigeixen igualment a la pell. Al sistema venós, hi ha un sistema portahepàtic que recull la sang de l’intestí i la porta al fetge abans que retorni al cor. La sang presenta una estructura semblant i una composició similar a la de la resta de vertebrats. Els glòbuls vermells, però, són nucleats i en algunes espècies arriben a tenir una bona mida.
L’aparell excretor
Aquest aparell s’ocupa de la regulació del contingut hídric i de la concentració de sals en el medi intern, a més de l’eliminació dels productes del metabolisme cel·lular, problemes tots tres molt importants per al canvi que representa el pas a terra ferma i especialment pel que fa al primer punt de regulació de continguts d’aigua. En els amfibis hi ha un parell de ronyons amb les corresponents vies d’evacuació d’orina.
El ronyó dels amfibis és de tipus opistonèfric en els adults, i correspon al segon estadi de l’evolució renal dels vertebrats. La forma i la mida són variables als diferents grups, però, en general, són relativament allargats, especialment en els gimnofions i els urodels, i més massissos en els anurs. En el mascle, els conductes del testicle penetren a la part posterior del ronyó o bé desemboquen en els conductes nèfrics, que han recollit el filtrat dels glomèruls renals i que desemboquen als túbuls col·lectors i, d’aquests, al conducte de Wolff (urèter) d’on passen directament a la cloaca; hi ha, per tant, una íntima relació entre l’aparell genital i l’excretor. Els conductes renals posteriors poden fondre’s o no al conducte de Wolff. En la femella, els dos aparells, reproductor i excretor, són independents.
La nefrona no és d’una gran eficàcia i és semblant a la dels peixos, encara que millorada mitjançant el control hormonal hipofisari de la filtració de la sang i un control simultani de la permeabilitat de la pell. Pel que fa a la bufeta urinària, és un simple diverticle de la cloaca que acumula l’orina, però és completament independent dels conductes renals.
L’aparell reproductor
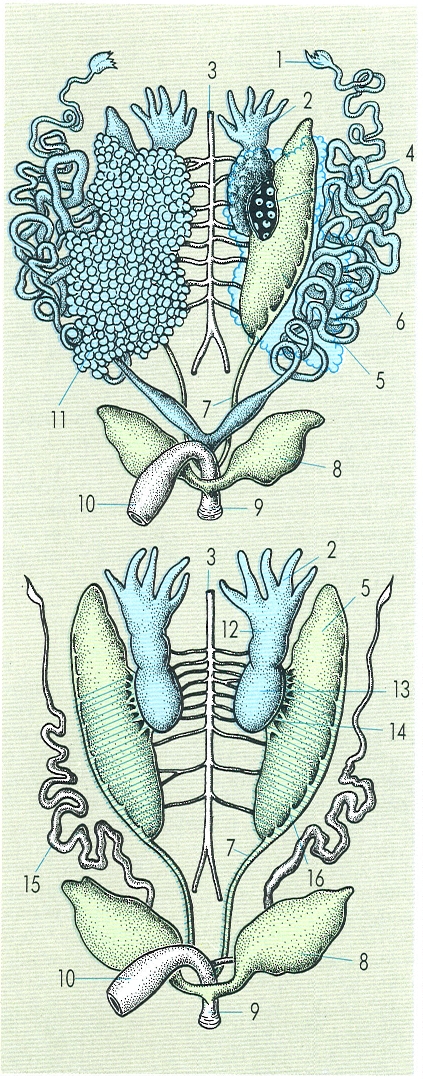
Esquema de l’aparell urogenital d’una femella (A, amb els òvuls extrets del costat esquerre, per a fer visible l’ovari), i d’un mascle (B) d’anur, concretament de gripau comú (Bufo bufo). Hom hi ha indicat: 1 pavelló de l’oviducte, 2 cos groc, 3 aorta, 4 ovari seccionat, 5 opistonefre, 6 oviducte o conducte de Müller, 7 canal de Wolf o urèter, 8 bufeta urinària, 9 cloaca, 10 recte, 11 ovari, 12 òrgan de Bidder, 13 testicle, 14 conductes eferents, 15 conducte de Müller rudimentari, 16 conductes col·lectors.
Gustavo Hormiga, original de Grassé i altres.
La femella presenta un ovari saculat, amb una cavitat limfàtica. La forma de l’ovari és variable i relacionada amb la forma del cos, i té una cavitat única, llevat dels anurs, que presenten bosses ovàriques múltiples. Els òvuls, d’un 2 m de diàmetre, presenten poc vitel·lus i són alliberats al celoma en trencarse la pròpia paret ovàrica. Existeixen cossos grassos, com a reserva energètica, que minven quan els òvuls creixen, i a la inversa. Hi ha un parell de conductes de Müller o oviductes, amb una obertura en forma d’embut, cap a la qual s’orienten els òvuls mitjançant l’acció de la paret de la cavitat interna. Durant la fase reproductora, els conductes creixen i s’entortolliguen; llur revestiment glandular segrega una substància gelatinosa i transparent que es diposita damunt de cada òvul quan aquest avança per raó dels moviments peristàltics dels conductes. Finalment, els òvuls queden emmagatzemats en una dilatació de l’oviducte fins al moment de la posta.
Els mascles presenten testicles compactes, formats pels túbuls seminífers. La forma està en relació amb la forma general del cos de l’animal i la mida és variable segons l’estació. Presenten cossos grassos associats. Un cas notable el constitueixen els bufònids, gripaus típics, en els quals l’abladó dels testicles comporta el desenvolupament d’ovaris funcionals a partir de l’anomenat òrgan de Bidder. Els túbuls deferents del testicle connecten amb el ronyó i desemboquen a conducte de Wolff. Existeixen vesícules seminals per a emmagatzemar els espermatozoides. En els mascles, hi ha també conductes de Müller rudimentaris.
Les característiques i les adaptacions fisiològiques dels amfibis
La regulació tèrmica
La producció de calor com a fruit secundari de les reaccions metabòliques és, en el cas dels amfibis, força baixa, a causa de la discreta eficàcia dels aparelli circulatori i respiratori. Aquest fet es correspon amb un metabolisme lent que requereix cercar fonts de calor externes, de tipus ambiental. Aquesta necessitat de regular ecològicament i etològicament la temperatura corporal s’anomena ectotèrmia.
Les temperatures crítiques de supervivència dels amfibis semblen variar entre –2 i 27 °C en els urodels, i entre 3 i 41 °C en els anurs. Fora d’aquests intervals tèrmics, la supervivència de l’animal és impossible, perquè esdevé incapaç de realitzar els actes vitals per manca de coordinació motora.
Pel que sembla, la majoria d’amfibis no tenen temperatures preferencials, és a dir, òptimes, de manera que dins el marge de temperatures crítiques tant els urodels com pràcticament tots els anurs suporten bé qualsevol temperatura. En els animals d’hàbits més aquàtics, el manteniment de temperatures és més fàcil, perquè l’aigua té unes variacions tèrmiques moderades, però a terra ferma aquests animals cerquen, en general, zones d’ombra i s’exposen poc o gens al sol; malgrat això, existeix un cert grau d’evaporació d’aigua per via cutània i, per aquesta raó llur temperatura corporal sempre és una mica inferior a l’ambiental; en cert sentit, això constitueix una termoregulació. Algunes granotes poden assolellar-se i, aleshores, certament, llur temperatura corporal supera en uns quants graus centígrads l’ambiental.
De tota manera, tot i l’aparent independència dels valors concrets assolits per la temperatura ambiental, sembla evident, en el cas de molts amfibis, que hi ha una relació entre l’activitat de l’animal i la situació tèrmica, ja que els amfibis són més sensibles quan es troben prop dels límits de llur interval favorable i exigeixen uns mínims per emprendre llurs activitats o per dur a terme correctament l’etapa reproductora.
L’intercanvi de gasos
En els amfibis, existeix una gran diversitat de superfícies respiratòries: brànquies, pulmons, tegument i mucosa bucofaríngia. Cadascuna funciona de manera lleugerament diferent i la seva importància relativa és molt variable. Així, les larves i els amfibis neotènics respiren fonamentalment mitjançant les brànquies i els adults ho fan pels pulmons i la pell i, en petita part, per la mucosa bucal. L’eficàcia dels pulmons varia amb la temperatura, de manera que el paper respiratori de la pell esdevé molt important a temperatures baixes; els pulmons predominen a temperatures altes i en animals actius, la pell, en canvi, és un element clau durant el repòs hivernal. En certes salamandres, com en el gènere Hydromantes, la pell és l’única superfície respiratòria, perquè no hi ha pulmons i és enormement vascularitzada i eficaç.
La captació d’aire per un amfibi proveït de pulmons es fa amb els orificis nasals oberts, la glotis tancada i per mitjà d’ascensos i descensos alternats del terra bucal; en obrir la glotis, l’aire, a pressió atmosfèrica, no penetra als pulmons (que estan a pressió superior) sinó que, aleshores, es produeix l’aspiració, amb el terra bucal deprimit, conservant l’aire engolit; un cop acabada l’aspiració, es tanquen els orificis nasals, s’aixeca el terra bucal i s’impulsa l’aire a l’interior del pulmó. Sembla que alguns amfibis poden continuar en part llur respiració dins de l’aigua; això explicaria la llarga permanència d’algunes espècies en immersió, fins unes quantes hores, i potser també la hivernació subaquàtica d’algunes granotes. Com en molts vertebrats, en les granotes ha estat verificada la quantitat superior d’hemoglobina en animals d’altitud, com a producte d’una adaptació per tal d’aprofitar millor la baixa concentració d’oxigen. Part de la regulació en l’expulsió de CO2 correspon a la pell, per incapacitat del pulmó de mantenir baixa la tensió de l’anhídric carbònic; en canvi, la tensió de l’oxigen en la sang pot ser controlada exclusivament pel pulmó.
La regulació hídrica
En animals que han esdevingut terrestres, com els amfibis, la regulació hídrica és un problema fonamental, perquè cal que compensin la pèrdua d’aigua per evaporació, que és inevitable en el medi aeri. La pell és una barrera molt permeable als gasos, com ja hem vist, però també a l’aigua, i això obliga els animals, d’una banda, a recloure’s en indrets humits i, de l’altra, a controlar acuradament les entrades i sortides d’aigua i de sals minerals de llur organisme, és a dir, a controlar tant l’activitat de la pell com la del ronyó.
En el cas de les larves, la regulació hídrica en l’aigua dolça ha de resoldre els mateixos problemes que els peixos en aquest medi. L’eliminació de residus nitrogenats en medi aquàtic es fa fonamentalment en forma de NH3, que es dilueix ràpidament en l’aigua circumdant, i l’aigua, que tendeix a entrar en excés, és utilitzada, precisament, per diluir l’orina amb amoníac. L’entrada d’aigua per òsmosi, a través de la pell, és constant, per la hipertonia del medi intern respecte de l’extern.
L’adult, en la seva vida a terra, es troba en una situació totalment diferent, perquè no pot perdre grans quantitats d’aigua amb l’orina; l’aigua ja no li entra a través de la pell, sinó que, al contrari, aquesta contribueix a la seva pèrdua. En conseqüència, els amfibis adults es veuen abocats a solucionar el problema evitant d’eliminar amoníac, reduint-lo a la urea, de manera que poden reduir la pèrdua d’aigua en forma d’orina en el procés d’eliminació dels residus nitrogenats. La nefrona renal és adaptada a la retenció superior d’aigua en els adults, per reabsorció en el túbul renal.
L’altre problema de la regulació osmòtica és la pell, que ha de ser permeable i mantenirse humida per a realitzar la funció respiratòria que té encomanada, la qual cosa comporta que el control de la sortida d’aigua de l’organisme esdevingui molt complex. En realitat, la pell és poc permeable a l’aigua, ja que existeix una capa de mucopolisacàrids amb calci subjacent a l’epidermis, situada entre la capa compacta i la capa esponjosa de la dermis, que possiblement controla, en els amfibis més terrestres almenys, la pèrdua d’aigua. En períodes molt secs, amb tot, els amfibis no poden controlar la pèrdua excessiva d’aigua i han d’aplicar altres recursos. En espècies sotmeses a secades més o menys periòdiques i serioses, en climes càlids, es presenta una estivació, que inclou la formació d’una mena de capa protectora amb la pell en muda. En biòtops més humits, un període accidentalment molt sec pot compensar-se passant a vida latent, també amb una estivació. Si l’espècie viu permanentment en clima àrid, apareixen nous mecanismes concrets, i permanents, com l’emmagatzemament d’aigua a la bufeta urinària.
D’altra banda, cal dir que els amfibis poden absorbir aigua a través de la pell, simplement mantenint-se submergits, almenys en la majoria de casos; algunes espècies fins i tot absorbeixen aigua a partir del sòl humit. Això és més evident en amfibis d’indrets àrids, que han d’aprofitar l’aigua disponible quan n’hi ha i tenen mecanismes de rehidratació ràpida molt eficaços. Cal dir també que moltes espècies sotmeses a secades poden regular l’equilibri osmòtic segons les disponibilitats d’humitat i funcionen com en aigua salada o com en aigua dolça, segons les condicions ambientals.
Pocs amfibis suporten aigües salabroses, encara que algunes larves d’anur, fins i tot en terres catalanes, poden adaptar-s’hi i sobreviuen en condicions de salinitat lleugera (com alguna espècie de Bufo o el gripau pintat, Discoglossus pictus); el mecanisme que actua en aquest cas és relacionat probablement amb la retenció d’urea i de ions minerals.
L’estratègia biològica dels amfibis
La reproducció i el desenvolupament
La reproducció dels amfibis presenta aspectes molt interessants, que són producte de la seva dependència de l’aigua, perquè l’ou no és protegit de la dessecació i a més té poques reserves nutritives, per la qual cosa el desenvolupament és indirecte. Però a part aquests condicionants fisiològics hi ha un comportament prereproductor peculiar, en què els estímuls i les respostes en la identificació sexual són molt senzills. El comportament reproductor dels amfibis és, en general, força estereotipat, amb patrons concrets i definits, normalment ben específics, que actuen, fins i tot, com una barrera reproductora. Els models bàsics funcionen amb mecanismes i estímuls molt simples. L’activació del cicle reproductor sembla seguir un rellotge intern ajustat al calendari més que a les condicions ambientals regnants, encara que aquestes poden tenir-hi una certa influència. En els gimnofions es coneix poca cosa del procés reproductor, però en els urodels i els anurs és prou conegut.
Festeig

El ritual de festeig d’aquesta parella de tritons jaspiats (Triturus marmoratus) il·lustra els preliminars de l’aparellament, que, en els urodels, són prou espectaculars.
Javier Andrada
En la major part d’amfibis, hi ha un comportament de festeig previ a la còpula. A part les cecílies, del comportament reproductor de les quals hom en coneix ben poca cosa, en els anurs i en els urodels aquesta fase ha estat ben estudiada i és prou coneguda.
En els urodels, el mascle exhibeix coloració nupcial, però no la femella, que és atreta per la vistositat de l’altre sexe. Els mascles i les femelles es dirigeixen a l’aigua per a la reproducció, llevat d’algunes salamandres que s’aparellen a terra ferma, com la salamandra del nostre país. Acostumen a arribar primer els mascles al lloc de reproducció, revestits d’una coloració nupcial més viva que l’habitual a la fase terrestre i proveïts sovint de crestes més desenvolupades al dors i a la cua. Després arriba la femella. El desencadenant de la reproducció sembla radicar en la maduresa de les glàndules sexuals, i és fonamentalment lligat al calendari i només parcialment influït per les condicions ambientals regnants en un moment determinat, de manera que l’arribada de cada espècie als seus llocs de reproducció varia poc d’un any a l’altre. El lloc ha de reunir una sèrie de condicions pel que fa al tipus d’aigua (corrent o estancada), a la temperatura (elevada o baixa), a la vegetació aquàtica (imprescindible o no), etc. En això els urodels són més selectius que no els anurs, que acostumen a aprofitar tota mena d’aigües. Es dóna el cas que moltes espècies utilitzen la mateixa localitat reproductora, però se la distribueixen, en l’espai i en el temps, entre totes elles. En climes temperats, l’estació reproductora és la primavera, però en climes freds pot ser l’estiu; en climes amb estació seca la reproducció es presenta en l’estació humida. Alguns amfibis són molt fidels a la localitat reproductora, i altres accepten i usen aigües diverses dins llur àrea. Un cop ambdós sexes es troben a l’aigua, el mascle efectua una sèrie de moviments encaminats a atreure i excitar la femella, per tal de posar-la en disposició d’acceptar i captar l’espermatòfor. És clar que aquesta norma general té excepcions, motivades per l’adaptació a cada ambient concret.


El cant dels mascles inicia espectacularment la reproducció en els anurs. L’emeten des de la vora de l’aigua o surant en aquesta, on són els primers en arribar. Les femelles reconeixen específicament aquesta veu, i s’orienten i es desplacen cap a la font sonora. Els sacs vocals o cambres de ressonància són gulars, com en la reineta comuna (Hyla meridionalis, a l’esquerra) o comissurals, com en la granota comuna (Rana perezi, a la dreta.)
Mirko Geest i Javier Andrada
Unes consideracions generals semblants són vàlides també referides als anurs, però la fase prèvia a la còpula funciona de manera diferent. En primer lloc, són els mascles els qui arriben a la localitat de posta, però les femelles no hi van immediatament, sinó que hi arriben atretes per l’emissió sonora específica dels mascles corresponents, cant que fan en cor o en solitari, i que només és reconegut per les femelles de la pròpia espècie, de manera que és un factor d’aïllament reproductor. El cant s’emet en condicions apropiades, més ben desenvolupat al pic de l’estació reproductora, i s’atenua al final tal com havia començat, més tímidament. Cal no confondre aquest cant amb altres emissions sonores, com el crit del mascle subjectat per un altre mascle, de la mateixa o d’una altra espècie (perquè el que estimula el mascle per a l’amplexus és la mida de la suposada femella); aquest crit s’anomena d’alliberament, mentre que pot emetre’s un altre crit semblant per tots els individus, que és el de l’animal capturat per un predador. Algunes espècies d’anurs, com el tòtil (Alytes obstetricans), es reprodueixen en el medi terrestre.
Acoblament

La fecundació es realitza de diversa manera en els anurs i en els urodels. En els primers, l’abraçada axil·lar, com la de la parella de granotes roges (Rana temporaria) de la fotografia superior, facilita només la fecundació externa: el mascle rega d’esperma els òvuls alliberats per la femella. En canvi, en els urodels, com en el tritó pirinenc (Euproctus asper, a baix), l’amplexus, amb entrellaçament de cues, permet acomplir la fecundació interna, amb transmissió d’un espermatòfor, per aposició de cloaques.
Josep M. Barres i Albert Montori
En els urodels, després de l’exhibició que el mascle efectua entorn de la femella, continua un ritual de comportament amb deposició final de l’espermatòfor (que conté els espermatozoides) per part del mascle, que, aleshores, obliga la femella a recollir-lo dins la cavitat genital, per a una fecundació interna. La femella efectua posteriorment, en un termini variable, la posta d’ous o de larves.
En els anurs, i també en alguns urodels, es produeix un amplexus, és a dir, una abraçada del mascle a la femella, a nivell axil·lar o inguinal, que l’excita per mitjà del moviment de les potes posteriors i li fa emetre els òvuls, els quals són regats amb l’esperma masculí; en aquest cas, la fecundació és externa. En les cecílies, la fecundació és interna, però no se’n coneix el procés.
La posta
La posta és molt diversa. En els gimnofions, la femella diposita ous fecundats, amb molt de vitel·lus, cosa que permet un llarg desenvolupament larval dins el propi ou, seguit després d’una, igualment llarga, fase de larva aquàtica. En els urodels, les salamandres van a l’aigua només per alliberar larves ja lliures, mentre que altres espècies, com els tritons, dipositen els ous, generalment un per un o en petits paquets subjectats per les plantes aquàtiques o adherits a les pedres, dintre de l’aigua, on romandran fins a l’eclosió i després per a la fase larval. En els anurs, la posta pot ésser en forma de paquet petit, de cordó llarg, de massa irregular o en alguna altra, i comprèn un nombre d’ous variable, petit o molt elevat, fins d’uns quants milers. La posta resta dipositada al fons o entorn de plantes, o bé sura a la superfície de l’aigua fins que els ous es desclouen. Hi ha, però, adaptacions a l’escassetat d’aigua, que permeten deixar la posta en nius d’escuma o entorn de les potes posteriors del mascle, i aquest la remulla periòdicament (cas dels gripaus del gènere Alytes, per exemple); o, fins i tot, hi ha casos en què la posta es conserva al dors de la femella, on cada ou disposa d’una cambra on acompleix tot el procés embrionari, fins la metamorfosi (com la granota americana Pipa pipa).
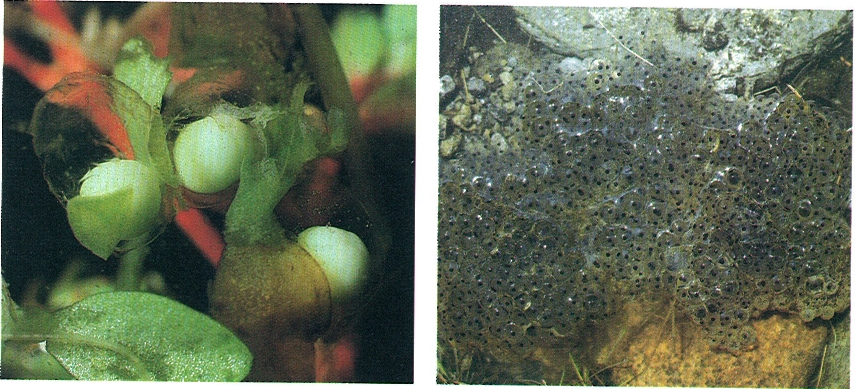

Les postes del gènere Triturus (a l’esquerra) són formades per ous independents adherits a les fulles de les plantes aquàtiques que la femella embolica entorn de cada ou. En les del gènere Rana, com la granota roja (R. temporaria, a la dreta), els ous són dipositats en paquets grossos; amb llur càpsula gelatinosa, aquests són els més grossos entre els dels anurs.
Hàbitat/Xavier Moreno i Javier Andrada

La forma i la mida de les postes són característiques de cada espècie. En el cas dels gripaus, com el gripau corredor (Bufo calamita) de la fotografia, les postes són constituïdes per cordons llargs amb dues o tres fileres d’ous; en 2,3 o 4 m de llargada poden contenir fins 4000 ous o més.
Jaume Orta
En tots els amfibis els ous són telolecítics, amb proporció moderada de vitel·lus, però variable d’un grup a l’altre segons si el desenvolupament és completament aquàtic, i aleshores són ous més petits (d’1 mm de diàmetre), o és més terrestre (fins de 4 mm de diàmetre). Si la posta té lloc a l’aigua, el revestiment gelatinós de l’ou s’infla i actua a manera de protecció, en part, probablement també, perquè el valor nutritiu que ofereix és molt baix per a resultar rendible a un predador.
Si bé la majoria d’espècies amfíbies són ovípares i els ous dipositats a l’aigua no es desclouen fins després d’un període breu, algunes formes són ovovivípares, com les salamandres, i no ponen ous, sinó que alliberen larves ben formades que completen llur desenvolupament dins l’aigua. La femella reté habitualment aquests ous dins els oviductes per un temps llarg i els embrions avancen, en el seu desenvolupament, de manera notable. Les femelles dels pípids arriben a carregar-se llurs larves al dors i les alliberen quan ja han acomplert la metamorfosi, però no hi ha intercanvi alimentari entre la mare i les cries; no es tracta de viviparisme, sinó més aviat d’incubació.
Les larves i la metaformosi
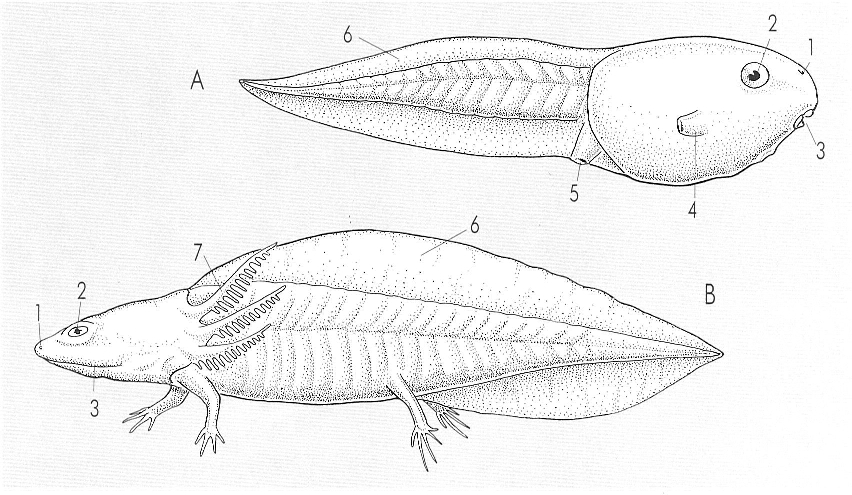
Les larves dels amfibis difereixen morfològicament en els diversos grups, bé que anatòmicament tenen caracteristiques comunes. Les d’anur (A) són àpodes, amb el cos ovoide i la cua comprimida, mentre que les d’urodel (B) són salamandriformes, tenen potes i ja són semblants a l’adult Hom ha indicat en el dibuix: 1 orifici nasal, 2 ull, 3 boca, 4 espiracle, 5 orifici anal, 6 cresta, 7 brànquies externes.
Amadeu Blasco, a partir de fonts diverses.
Les larves varien d’un grup a l’altre d’amfibis. El medi influeix en la forma de la larva: aigües corrents o estancades determinen brànquies diferents, més grosses o més petites (en relació amb el contingut d’oxigen de l’aigua); també la morfologia corporal es modifica: les larves són més comprimides en les aigües més vives. Igualment, la temperatura influeix en la durada del període larval, que és més curt com més alta sigui aquella.
En els gimnofions, el creixement intraovular és molt llarg i la larva resultant és anguil·liforme i aquàtica i ja sense brànquies, però encara necessita romandre un cert temps a l’aigua per a completar la metamorfosi.
En els urodels, les larves presenten la morfologia bàsica de l’adult en el moment mateix de la desclosa de l’ou, però són aquàtiques, amb brànquies externes i quatre potes fines. La mateixa morfologia presenta la larva de salamandra alliberada per la femella, ja lliure de les capes protectores de l’ou. La metamorfosi es produeix al cap d’uns mesos, amb reabsorció dels òrgans aquàtics, com les brànquies, i formació dels aparells terrestres, com els pulmons.
En els anurs, la larva és peculiar; el cos és gairebé esfèric o ovalat, àpode i proveït d’una cua comprimida, amb la membrana caudal ben desenvolupada. En descloure’s l’ou, la larva encara no té formada la boca i roman unes hores, o uns dies, immòbil. Després es forma la boca, amb un bec i uns denticles cornis, disposats de manera peculiar a cada espècie. Les brànquies externes inicials molt aviat són reabsorbides i es formen noves brànquies internes no homòlogues a les dels peixos, dins una cambra branquial que connecta amb l’exterior mitjançant un orifici, l’espirade. Progressivament la larva forma les potes posteriors i després les anteriors, comença a reabsorbir la cua i les brànquies i a formar els pulmons, a més de modificar l’aparell circulatori, la boca, la morfologia corporal i el tegument. En uns mesos completa la metamorfosi, i surt de l’aigua. Hom observa que els individus en metamorfosi, a més dels canvis morfològics esmentats, comencen a pujar a respirar a la superfície i romanen llargues estones subjectes a objectes flotants o situats al seu damunt fins acabar independitzant-se de l’aigua. La metamorfosi és controlada per factors genètics mitjançant les hormones però també per factors ambientals.
La neotènia

Les larves dels urodels presenten una morfologia semblant a la de l’adult, encara que en difereixen per la coloració i per les brànquies externes, que persisteixen fins a la metamorfosi, com les que mostra plenament esteses el tritó jaspiat (Triturus marmoratus) de la fotografia superior Les larves d’anur es diferencien patentment dels adults del grup pel seu cos globós i la seva cua comprimida; la larva de granota verda (Rana perezi, a baix) mostra l’absència de brànquies externes; l’individu ha iniciat la metamorfosi: persisteix la cua, les potes posteriors ja són desenvolupades i s’aprecien els monyons de les anteriors.
Salvador Bosch i Ramon Torres
En alguns urodels, els individus adults retenen caràcters larvals, el més evident dels quals és la persistència de les brànquies. Pot haver-hi una pedogènesi de base genètica i una neotènia induïda ambientalment. En el cas dels sirènids i els protèids, es tracta de pedogènesi, perquè la glàndula tiroide no obté resposta en completar la metamorfosi, mentre que en altres casos, com en el gènere americà Ambystoma, es presenta una neotènia, i els individus poden completar la metamorfosi (absorció completa de les brànquies) si les condicions són especials; normalment mantenen els caràcters larvals però, si hi ha evaporació de la massa d’aigua on viuen, passen a completar la metamorfosi per a poder migrar a una altra localitat aquàtica.
La hibridació
En condicions naturals, es produeixen en els amfibis casos d’hibridació entre espècies pròximes. Són coneguts, per exemple, entre el tritó crestat (Triturus cristatus) i el tritó jaspiat (T. marmoratus), en localitats on ambdues espècies de tritons conviuen, i donen híbrids estèrils lleugerament diferenciats segons el sexe que correspon a cada espècie progenitora. Però el cas més espectacular és el que afecta les granotes verdes europees, del grup Rana esculenta, que hom considerava tradicionalment una única espècie, a partir de la qual derivaren les altres formes per isolament geogràfic a les terres mediterrànies durant les glaciacions. Tanmateix, les anàlisis genètiques i immunològiques mostren clarament que R. ridibunda i R. lessonae, a l’occident i a l’orient d’Europa respectivament, són dues espècies diferents, i que R. esculenta és l’híbrid, fèrtil, de les dues, que és reproduïble amb ella mateixa i amb les espècies progenitores. El mecanisme genètic responsable d’aquesta situació és força complex i pressuposa l’existència de diploides i triploides de R. esculenta, que permetin l’autoreproducció i la reproducció amb una o altra de les espècies progenitores. Entre les dotacions cromosòmiques de R. lessonae i R. ridibunda, a més, sembla no produir-se mai entrecreuament genètic. D’altra banda, R. ridibunda tampoc no és una espècie única, sinó que, a més de la forma nominal, una de les seves subspècies ha passat a ésser considerada com una espècie: R. perezi, que també s’hibrida amb R. ridibunda i probablement, cal afegir R. saharica a aquest grup. La situació d’aquestes formes, molt similars en morfologia i hàbits, però biològicament ben diferenciades i, en canvi, hibridables, no és encara ben resolta, i cal atribuir la formació d’aquest complex d’espècies de granotes verdes als isolaments produïts per les glaciacions pleistocenes.
Els cicles d’activitat
Com que els amfibis són ectotèrmics, a bona part de les àrees on viuen els és necessari un període de diapausa en llur cicle actiu, perquè no poden compensar les condicions desfavorables per mitjà de mecanismes fisiològics. Només els que viuen a zones tropicals i humides poden mantenir-se actius tot l’any.
Normalment, i especialment en climes temperats, els amfibis hivernen, per bé que si el clima té una estació molt seca la diapausa correspon a aquesta època; en llocs càlids d’estiu sec, pot haver-hi una estivació. Encara que no es coneixen ben bé els mecanismes que obliguen l’animal a passar a l’estat de repòs i, més tard, a sortir-ne, pot ser que es tracti d’un mecanisme tèrmic o d’un rellotge intern. És evident que, amb les reserves de què disposa, l’animal no pot allargar excessivament el seu repòs i ha de regir-se per un calendari; d’altra banda, el fet de sortir en condicions desfavorables disminuiria les seves possibilitats de supervivència. Caldria imaginar, per tant, un rellotge tèrmic associat al rellotge intern per a determinar la sortida de la diapausa.
En clima temperat, l’animal que surt de la hivernació no triga gaire temps a entrar en període reproductor; aquest inici es regula per un mecanisme fisiològic, que és controlat per la hipòfisi i per les gònades, encara que probablement hi ha influència ambiental (potser tèrmica), i pot ésser simultani o no a tots els individus d’una població determinada. En aquesta etapa, els animals són molt gregaris, perquè les localitats de reproducció no són abundants i, a més, el gregarisme augmenta les possibilitats de reproducció pròpiament dita. Un cop acabada aquesta etapa, que inclou l’arribada a l’aigua, el festeig, la fecundació i la posta (o els processos equivalents en animals terrestres o ovovivípars), els adults es dispersen per terra ferma, on viuen individualment, i en medis sovint ben diferents del de reproducció.
Fora de l’etapa reproductora, l’amfibi adult s’ocupa d’alimentar-se. El ritme actiu és, segons les espècies, més o menys diürn, però, en general, els amfibis són més aviat crepusculars o nocturns. Aquesta etapa de llur existència és molt menys coneguda, perquè són força discrets. Pot haver-hi, amb tot, més d’un període reproductor a l’any, amb una segona reproducció a la tardor, si el clima és favorable. En regions més càlides, els amfibis poden reproduir-se quasi tot l’any, mentre que en zones fredes el cicle reproductor s’estén al llarg de dos anys, el primer dels quals correspon a l’aparellament, sovint amb la posta retardada fins l’any següent; en altres casos, les larves, que romanen a l’aigua fins assolir la metamorfosi, poden necessitar hivernar un any en fase larval si no han pogut completar llur desenvolupament.
Els joves que surten de l’aigua se sotmeten, com les larves, a una forta pressió depredadora, no selectiva sinó massiva, i pocs arriben a la maduresa sexual, que pot trigar dos anys o més, i menys encara a una edat molt elevada. Hom desconeix la longevitat màxima de moltes espècies en vida lliure.
L’adequació a l’ambient
Els factors físics
El medi físic condiciona l’existència dels amfibis en unes regions determinades, a causa, sobretot, del fet que són ectotèrmics i depenen de l’entorn per a la regulació de llur temperatura corporal. Els factors físics més importants com a condicionants de la presència dels amfibis són la temperatura, la humitat, la presència d’aigua líquida, l’existència de refugis i, en menor importància, la qualitat del substrat.
L’orientació de les superfícies i la naturalesa de llurs materials determinen, en certa manera, els factors anteriorment esmentats. Els amfibis tenen uns requeriments tèrmics relativament moderats, suporten temperatures prou baixes i exigeixen humitat i presència d’aigua, de manera que les orientacions obagues els són favorables. D’altra banda, certs tipus de materials rocosos permeten amb més facilitat que altres la formació de tolls d’aigua, així com la constitució de refugis.
Les temperatures crítiques són diferents per a cada espècie d’amfibi, però, en conjunt, són més baixes que per als rèptils. Els amfibis poden esdevenir actius entorn dels –2 °C, mentre que difícilment toleren més de 40 °C; molt sovint les màximes crítiques són molt inferiors. Existeixen espècies endèmiques d’alta muntanya i algunes formes arriben al cercle polar àrtic. No mostren preferències tèrmiques molt precises, és. a dir, moltes espècies no tenen temperatures òptimes.
Pel fet que tenen la pell permeable, la humiíat els és imprescindible. Tanmateix, alguns amfibis arriben a suportar medis força àrids, mitjançant una sèrie de recursos concrets: el primer, practicat per un bon nombre d’espècies, és la crepuscularitat o nocturnitat dels animals; en altres casos, poden acumular enormes quantitats d’aigua durant els períodes de pluja i s’inflen suficientment per a poder viure de les reserves en les etapes seques. Fins i tot, la reproducció pot arribar a independitzar-se de l’aigua, mitjançant la secreció per part dels adults d’una escuma en què es desenvolupa la posta; hi ha diversos mecanismes en diferents grups. Si hi ha aigua abundant, molts amfibis no són selectius pel que fa a la mena de massa d’aigua en ella mateixa, però altres tenen exigències precises respecte de la qualitat i la quantitat d’aigua, la seva renovació, el corrent, la vegetació aquàtica, els invertebrats existents o la profunditat.
El tipus de substrat només repercuteix directament en les formes d’hàbits excavadors, però indirectament influeix en l’existència de refugis subterranis o en altres factors biòtics o abiòtics. El paper dels refugis és fonamental, perquè és el que permet la supervivència dels animals en medis que presenten climes extremats, sigui d’alta muntanya, continentals, o altres, en què s’arriba a extrems tèrmics o d’aridesa molt notables, que l’amfibi pot evitar dins del refugi aprofitant les hores favorables per desplegar la seva activitat.
La vegetació
La vegetació d’una zona no té efecte en les comunitats amfíbiques que la formen, però certes comunitats vegetals afavoreixen la seva presència, i molt sovint la de determinades espècies, com la salamandra, freqüent als boscos, o algunes espècies de gripaus, a les clarianes i vorades; poques formes es troben en àrees massa obertes. De fet, la presència d’abundant vegetació comporta l’augment de la humitat de l’indret i, per això, les zones amb vegetació densa són més riques en amfibis, tant en diversitat d’espècies com en nombre d’individus.
Les relacions tròfiques
L’alimentació

L’alimentació dels amfibis és variada: sovint les larves, sobretot les dels anurs (a dalt), són herbívores, encara que les d’algunes espècies són carnívores; d’aquesta manera, poden repartir-se els recursos tròfics quan es reprodueixen diverses espècies a la mateixa massa d’aigua. Els adults són quasi exclusivament carnívors, com el tritó meridional (Pleurodeles waltl), que a la fotografia consumeix un oligoquet (a baix); la composició de la dieta és, tanmateix, variable segons les espècies.
Javier Andrada
Els amfibis adults són quasi exclusivament carnívors. Les preses consumides, sempre vives, són molt diverses i sovint consisteixen en diferents invertebrats, com alguns insectes, crustacis terrestres (isòpodes), oligoquets, mol·luscs gasteròpodes, etc., damunt dels quals els amfibis semblen exercir una predació no selectiva, ja que els únics factors que determinen la captura són la mida de la presa i la seva abundància dins la comunitat i, en particular, dins el biòtop de l’amfibi depredador.
La captura es realitza de diverses maneres. Els urodels aguaiten i capturen llurs preses sense projectar la llengua. Els anurs poden saltar damunt les preses i, segons la mena de llengua que presenten, poden projectar-la (com els bufònids) directament sobre la presa, embolicant-la per engolir-la, en un procés rapidíssim. La màxima precisió en la captura i en el salt l’exerceixen els rànids, que atrapen a l’aire molts insectes voladors i fins i tot algunes cries de vertebrats. La majoria de captures es realitzen en medi aeri, encara que l’espècie tingui hàbits fortament aquàtics, però també es poden capturar alguns invertebrats dins l’aigua, fet obligat en algunes formes que no presenten fase terrestre.
Les larves poden presentar un règim diferent: les d’urodel són carnívores i consumeixen petits invertebrats aquàtics sense seleccionar, ja que capturen les preses segons llur disponibilitat i llur mida. Els cap-grossos d’anur, en canvi, són predominantment herbívors i raspadors o filtradors; els primers raspen les plantes aquàtiques amb el bec i els denticles del disc oral, mentre que els segons filtren bacteris i algues en suspensió. Hom pot observar, però, que larves de règim vegetarià típic aprofiten els cadàvers d’alguns cap-grossos de llur mateixa o d’una altra espècie, i també poden observar-se aglomeracions de larves en un punt determinat entorn de la larva morta. No hi ha constància de mort produïda pels propis congèneres; es tracta, en tot cas, d’un intent d’utilitzar un recurs alimentari suplementari, perquè les poblacions de larves són molt nombroses i les fonts nutritives més aviat limitades.
La competència
La competència s’estableix en diversos camps: lluita per les mateixes fonts alimentàries, pels refugis, per les localitats de reproducció. Cal, doncs, definir el nínxol ecològic concret per a cada espècie i tenir en compte que en alguns casos l’espècie més forta o més eficaç desplaça altres espècies, que aprofitarien el mateix recurs si aquella no els ho impedís.
La competència alimentària es relaciona amb la quantitat global de preses disponibles.
Hem vist que els amfibis no seleccionen específicament llur aliment, de manera que qualsevol organisme d’una mida apropiada i no tòxic (molts eviten les abelles i animals semblants) pot ser capturat, i totes les espècies presents en una àrea es reparteixen els recursos nutritius segons l’accés que hi tenen en cada moment, és a dir, segons que les presumptes preses siguin actives a les mateixes hores que els potencials predadors. Si els invertebrats d’una zona són diversificats i abundants, la població amfíbica es mantindrà i la competència alimentària tindrà efectes més aviat reduïts. Cal, però, que els animals estableixin llur propi territori de caça i que hi cerquin el refugi per a les hores actives, tant del cicle diürn/nocturn com del cicle hivernal.
El propi refugi és un problema important; molts amfibis hivernen en solitari o reposen en petits grups (és fàcil trobar dos o tres exemplars de gripau corredor, Bufo calamita, de dia, sota una pedra) i els refugis accessibles, però fora de l’abast dels predadors i que mantinguin condicions bones de temperatura i d’humitat per a l’hivern, per a l’estiu o per a passar la nit o el dia, són poc abundants.
Les localitats de reproducció poden constituir un factor limitant molt important per als amfibis i poden determinar l’absència de les diverses espècies si no reuneixen les condicions adients. Encara que alguns amfibis són poc selectius en aquest aspecte, el cert és que tots exigeixen un mínim de condicions referents a l’estat físic (temperatura, corrent, profunditat) o, fins i tot, químic (salinitat, acidesa) de l’aigua. Diverses espècies poden dur a terme l’aparellament i la posta en el mateix estany, bassa o riu, però es distribueixen estratègicament dins la massa d’aigua; certes formes prefereixen la vora (com la granota roja, Rana temporaria), altres una mica més de profunditat (alguns Triturus), algunes precisen plantes submergides, altres volen aigua molt corrent o molt renovada o simplement només toleren aigua molt neta (com la salamandra, Salamandra salamandra). Espècies d’exigències semblants poden trobar-se al mateix riu però a diferents basses, o a la mateixa bassa però a diferents racons, amb diferent insolació, vegetació o profunditat.
Els recursos defensius

Els ambifis acostumen a ser críptics: la fotografia, de gripau comú (Bufo bufo), palesa perfectament aquesta dissimulació completa de l’animal en el seu entorn. Això no obstant, hi ha casos de coloració aposemàtica, d’advertència, com és el de la salamandra.
Jordi Berthold
Els amfibis sofreixen a la natura una important pressió de depredació. Molts vertebrats de grups diversos, rèptils, ocells i mamífers, i, fins i tot, els peixos, els ataquen en diferents fases del seu cicle biològic i en consumeixen una bona quantitat. Els peixos de riu i de llac, com les truites de riu, poden menjar un gran nombre de larves d’anur; per aquesta raó, moltes postes d’amfibi són dipositades a les vores, en aigües somes, a les quals els peixos no tenen fàcil accés. Les colobres d’aigua del gènere Natrix consumeixen tant larves com adults d’anurs, especialment granotes i gripaus, amb tots els quals comparteixen el biòtop molt sovint. Els ocells limícoles, com les cigonyes, també consumeixen amfibis, i el mateix ocorre amb alguns mamífers.

Les variacions de color són notables en alguns amfibis. En el cas de la reineta comuna (Hyla meridionalis), es presenten variacions a partir del verd brillant bàsic, normalment cap a tonalitats brunenques, que dissimulen els animals damunt del substrat (a dalt). Però també poden aparèixer mutacions anòmales, com la pèrdua de la capacitat de síntesi del pigment groc, de manera que l’animal és blau intens (a baix; l’exemplar de la fotografia presenta, tanmateix, algunes àrees verdes). Destaca, també, per la diversitat de coloració, la granota verda (Rana perezi).
Jordi Berthold
Davant d’aquest risc de predació, els amfibis han desenvolupat els corresponents mecanismes de defensa. El primer, i el més econòmic, és la coloració críptica, que correspon a la majoria d’espècies, almenys a les zones temperades. Els representants dels gèneres Rana o Hyla adopten, dins uns certs límits, variacions de coloració segons l’indret on es troben, llurs condicions fisiològiques o l’estació: les formes verd brillant, damunt d’una fulla, permeten a les reinetes de passar completament desapercebudes, sobretot si es mantenen completament immòbils; una granota damunt les llentilles d’aigua que cobreixen un estany esdevé practicament invisible. Altres espècies presenten coloracions aposemàtiques d’advertència, com la salamandra o alguns anurs tropicals: són els que presenten secrecions cutànies tòxiques. Aquestes secrecions són, en general, d’efectes moderats, encara que les d’algunes granotetes sudamericanes poden tenir efectes fulminants en animals petits i eren, fins i tot, utilitzades per les poblacions indígenes per obtenir verí. El més normal és que la concentració de substàncies tòxiques, químicament molt diverses i d’efectes molt variats, sigui molt baixa i, per tant, llur funció sigui sobretot de defensa antimicrobiana i de dissuasió del predador, perquè actuen com a irritants.
Alguns animals d’aquest grup, com els bufònids, adopten posicions peculiars de defensa, com inclinar-se cap endavant aixecant les potes posteriors, inflar-se tot el possible o fer el mort posant-se panxa enlaire, completament rígids. Aquestes posicions augmenten, en alguns casos, el volum aparent de l’animal, mentre que en altres posen en evidència parts de l’animal amb coloració d’advertència (com el ventre llampant dels representants del gènere europeu Bombina).