Els sistemes aïllats
El concepte bàsic d’illa és simple i intuïtiu: un fragment de terra envoltat d’aigua per tots costats. Però no menys intuïtivament el concepte ha estat aplicat tradicionalment a moltes altres realitats que tenen en comú el fet d’estar separades de les altres realitats equivalents per una altra realitat, en alguna mesura antitètica, que els envolta. És així com es pot parlar d’illes de cases envoltades per carrers (espai privat enfront d’espai públic), illes de vianants (espais sostrets del sistema de circulació viària motoritzada), illes de verdor (espais boscats envoltats de conreus o d’espais urbans) o bé usar l’expressió “ningú no és una illa” per expressar que ningú no pot defugir la influència dels altres quan viu en societat.
El concepte d’insularitat
Ecòlegs i biogeògrafs empren el terme illa donant per entès el concepte d’insularitat i, així, qualsevol porció de biosfera envoltada per una frontera o interfase a través de la qual els intercanvis siguin mínims i fonamentalment passius, podria ser considerada una illa. Una illa seria, doncs, un espai separat de l’immediat que l’envolta per una frontera tal que els fenòmens o els esdeveniments que es produeixen a un costat i l’altre pràcticament no tenen connexió ni s’influeixen mútuament. Així considerat, el fenomen de la insularitat és, doncs, molt estès a la natura, i es presenta en molts ambients diferents: òbviament, a les illes, però també als llacs i els estanys, i en altres ambients tan diferents com els cims de les muntanyes, els “tepuys” veneçolans, les cavitats subterrànies, els bocins de boscos que queden inclosos dins d’altres ecosistemes, zones calcàries enmig d’altres zones de substrat diferent, les fosses submarines, els voltants de les fumaroles abissals, les illes d’hàbitat, etc. En molts d’aquests casos no sol existir una interfase tan clara amb el medi que les envolta com en el cas de les illes vertaderes o dels llacs, casos paradigmàtics d’insularitat, sinó que les fronteres que limiten aquestes illes ecològiques són asimètriques i permeables, i permeten un flux de matèria i energia amb els medis circumdants que no passa exclusivament per la via detrítica.
L’espai geogràfic dels sistemes aïllats
Les illes pròpiament dites, en el sentit clàssic del mot —això és, les illes de substrat sòlid separades de tot altre medi terrestre per una extensió ocupada per un medi aquàtic—, són molt nombroses: unes 500 000, encara que el nombre exacte fa de molt mal precisar. La seva àrea total representa aproximadament el 5% de la superfície terrestre emergida.
Les illes de terra són unes àrees intrínsecament atractives, veritables mons a part, on la vida ha evolucionat seguint unes pautes particulars, diferents de les que s’han donat als continents, i sovint també a les de les altres illes. Les illes són mons aïllats, on éssers vius peculiars s’interrelacionen en xarxes ecològiques peculiars, i on l’evolució moltes vegades va més enllà de la imaginació, a tocar de la fantasia.
Els llacs —això és, les illes líquides— representen aproximadament l’1,8% de la superfície terrestre, i tant poden situar-se al si de grans masses continentals com d’illes genuïnes. Els llacs, malgrat no tenir la mateixa importància geogràfica, responen a un concepte d’organització espacial molt similar al de les illes genuïnes. Les fronteres que separen els dos tipus de medis, terrestres i aquàtics, són fronteres asimètriques, clares i de gran contrast, i l’intercanvi de matèria que té lloc a través d’aquestes fronteres sol fer-se per la via detrítica.
Altrament, les coves (illes hipogees) queden limitades a determinats substrats geològics tanmateix desigualment escampats arreu del món. Les “illes” culminals de moltes muntanyes, i altres a les quals s’ha fet referència, tenen també distribucions molt desiguals.
L’aïllament i l’evolució biològica
La biosfera es pot imaginar com solcada per innombrables vies de difusió, i plena de fronteres relatives. Aquelles faciliten el flux genètic, mentre que les fronteres el dificulten, i tot plegat depèn també de la mena d’organismes implicats. Camins i fronteres són la trama i l’ordit en el teler de la vida, que contínuament va creant noves situacions.
Conjunts de Julia
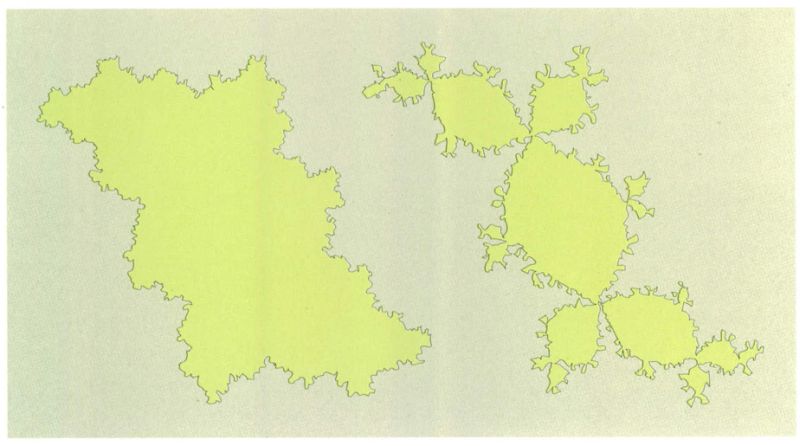
Jordi Corbera, a partir de fonts diverses
Els anomenats conjunts de Julia podrien servir d’exemple per a explicar com s’inicia el procés d’especiació. Un conjunt de Júlia es genera quan dibuixem sobre una superfície la línia que separa els punts del pla que compleixen una determinada propietat d’aquells que no la compleixen (es dona la circumstància que una línia generada d’aquesta manera té estructura de fractal). L’equació matemàtica que defineix el conjunt de Júlia de l’esquerra podria ser tan simple com Xn+1=Xn2+ C, essent Xn i Xn+1 dos punts del pla i C una constant. Variant tan sols la C d’aquesta fórmula, s’obté un conjunt de Júlia força diferent, que podria ser, per exemple, el de la dreta. Aquest model es podria equiparar a la primera fase del procés d’especiació. El conjunt de Júlia de l’esquerra representaria l’àrea de distribució d’una espècie. Si per qualsevol causa varia ni que sigui una sola constant ambiental (que equivaldria a la C), l’àrea de distribució podria convertir-se en el conjunt de Júlia de la dreta. En aquest cas, l’àrea de distribució ja no és uniforme, sinó que han aparegut diferents poblacions que no tenen contacte entre elles. Aquest és el primer pas de la formació d’una nova espècie. Si una població no és partida en dues o més poblacions separades per alguna mena de barrera (geogràfica, ecològica, genètica, etc.) que impedeix que s’encreuin els genomes respectius, no serà mai possible que les dues poblacions esdevinguin diferents, i que, per tant, a la llarga apareguin espècies diferents. Per això, els sistemes aïllats tenen un paper tan important en el procés d’especiació i en l’evolució biològica.
El fascinant comportament dels sistemes aïllats
L’estudi de les poblacions insulars ha estat sempre estimulant. Charles Darwin rebé una inspiració decisiva en fixar-se en les diferències entre espècies suposadament d’origen comú i que vivien en ambients variats de diverses illes de l’arxipèlag de les Galápagos. I l’estudi de l’aïllament de les poblacions com a agent d’especiació ha mantingut desvetllat el pensament dels biòlegs evolucionistes. Poblacions de tortugues gegants en illes del Pacífic, de peixos en diferents llacs i fins i tot en diferents espais dintre d’un mateix llac, de cargols terrestres en illes, o cargols aquàtics en llacs, i d’una gran varietat d’habitants de coves, són notables pel grau elevat d’especiació a escala relativament petita. La mateixa petitesa de les poblacions, en molts casos, n’ha facilitat la ràpida divergència, com en el cas de les sargantanes de les illes mediterrànies. És clar que aquesta manifestació tan viva de l’evolució no necessita ni illes reals i normals, ni illes invertides (és a dir, llacs), encara que totes elles ofereixen exemples molt eloqüents, especialment quan hi concorren altres causes suplementàries, com per exemple, fronteres gairebé impermeables i una extensió tan petita que exclou definitivament els grans depredadors.
Dins de conjunts d’illes, llacs i coves, també entre “tepuys” i altres ambients comparables, es reconeixen mons paral·lels, segregats i isolats, relativament tancats, en cada un dels quals l’evolució d’uns quants immigrants comuns ha seguit per vies separades i divergents en grau diferent. Les divergències evolutives reflecteixen condicions externes que s’han mantingut diferents i poc canviants, o bé semblen fruit de l’atzar. Avui dia, moltes espècies isolades es troben amenaçades d’extinció, principalment si les condicions d’entorn es ruderalitzen o suburbialitzen, i es facilita la invasió d’espècies alienes. Molts habitants de mons insulars ens semblen particularment fràgils i sempre s’esmenta amb aquest propòsit el cas del dodo de l’illa Maurici, al qual s’aplicà, en les primeres descripcions, l’epítet específic d’ineptus, que reflecteix el menyspreu amb què es veia l’absència d’unes qualitats que no li eren precises per a subsistir en el seu món abans de la irrupció del predador humà.
L’increment de la diversitat i l’alentiment de la dinàmica
L’establiment del concepte d’aïllament o d’insularitat mostra que hi ha una certa sensibilitat del naturalista envers l’operació de processos evolutius paral·lels o convergents, que es manifesten sota condicions diverses, i que produeixen poblacions relativament petites, constituïdes per individus de vida llarga, metabolisme baix sota condicions poc canviants i, molt sovint, dependència d’una font poc generosa però segura. Condicions totes de desacceleració de la dinàmica pròpia del sistema ecològic respectiu. Una conseqüència principal de la desacceleració és afavorir la diferenciació genètica i contribuir a l’augment de la diversitat.
Els artròpodes pobladors de coves acostumen a ser de mida superior a la normal en el grup al qual pertanyen. Sovint són incolors, de vida llarga i metabolisme baix, productors d’ous grossos i en petit nombre. És notable que aquestes característiques siguin compartides per animals de les grans fondàries marines, l’entorn dels quals, encara que uniforme en grans extensions, resulta comparable al de les coves per la poca densitat d’aliment. Aquesta poca densitat d’aliment és, en tots dos casos, compensada per la previsibilitat del seu subministrament, ja que, en el cas dels fons oceànics, l’aliment ve de la part il·luminada superficial, a través d’un llarg transport que n’assegura un flux migrat però segur. Per raons equivalents, el nodriment a les coves pot ser pobre, però relativament garantit.
L’aïllament en la vida dels grans fons oceànics, altrament, porta a una situació típica de dinàmica lenta i gran diversitat. Damunt d’aquest rerefons apareixen illes d’un altre gènere, illes invertides, en aquest cas els oasis hidrotermals que es formen al costat de les surgències d’aigües que, per les seves característiques, permeten l’activitat quimiosintètica d’alguns organismes. Aquests ecosistemes són típicament accelerats, de baixa diversitat i integrats per organismes de creixement ràpid, que fan un contrast ben instructiu amb les altres situacions d’aïllament. La vida que s’aplega entorn de les fonts del nostre paisatge emergit pot tenir característiques comparables.
Els culs-de-sac evolutius i els ambients fluctuants
L’aïllament entès de la manera tradicional, amb fronteres molt efectives, es pot relacionar amb escenaris d’evolució, on els límits són virtuals. En els espais caracteritzats per unes condicions més fluctuants, actua com a generador de formes taxonòmiques noves, alhora que les formes més antigues es van diversificant i se situen en posicions perifèriques, on s’arriba a acumular una gran diversitat de vida. Aquesta és una manera possible d’interpretar les fluctuacions a les zones temperades, que haurien ajudat a enriquir encara més la biodiversitat de les zones adjacents de clima més constant.
A les conques lacustres es manifesta la superior subdivisió i el potencial enriquiment dels ambients perifèrics. La subdivisió d’un gran llac en llacs menors en el curs d’episodis repetits del descens del nivell de l’aigua ha pogut portar a una considerable especiació dins de la conca lacustre general. El resultat es pot veure en les diverses nissagues de peixos i mol·luscs dels llacs africans i filipins. L’endemisme, és a dir, l’existència d’espècies pròpies de cada lloc, és relativament baix entre els pobladors de les llacunes de temporada, però mai no és absent del tot; les espècies més característiques produeixen grans quantitats de formes de resistència, que, transportades pel vent o pels ocells, mantenen un flux genètic intens que s’oposa amb èxit (encara que no sempre en el mateix grau) a la diferenciació de formes locals.
Encara que les espècies del plàncton puguin ser més antigues que les del bentos, les distribucions geogràfiques d’aquestes es veuen com si fossin més velles, perquè les del plàncton es reconstrueixen contínuament al ritme dels moviments de les aigües oceàniques. Les àrees glaciades foren netejades d’organismes cavernícoles propis (troglobionts), però els que persistiren en terres perifèriques semblen més arcaics que no les faunes recents epigees (no cavernícoles) de les mateixes regions. Per això hi ha troglobionts que es veuen més semblants als suposats descendents d’antics pobladors de la mateixa àrea, que no pas als recents, els avantpassats dels quals foren desplaçats amunt i avall. Les aigües subterrànies estan en relació les unes amb les altres i en grau molt divers per aqüífers i per sistemes càrstics locals, que poden barrejar i uniformitzar les poblacions, en relació amb les fluctuacions dels cabals que porten. La via subàlvia, subterrània i intersticial, ha estat un lloc de bescanvis entre espècies halòfiles d’origen marí costaner i espècies procedents d’aigua dolça.
Als ambients nous és fàcil la colonització per espècies alienes; aquest ha estat el cas dels embassaments espanyols poblats per formes que, en bona part, vingueren d’aigües de països més nòrdics, pròdigues en llacs de tota mena. Tot canvia, i espècies que antigament foren endemismes poden arribar a convertir-se en invasores amb tendència a esdevenir més o menys cosmopolites; aquest seria el cas, per exemple, dels mol·luscs Potamopyrgus antipodarum, un gasteròpode neozelandès que aparegué per primera vegada a Anglaterra el 1859 i que d’ençà d’aleshores s’ha estès per tota l’Europa occidental (on era conegut com a P. jenkinsi), i Dreissena polymorpha, un bivalve originari de la regió ponto-càspica que s’ha estès per les aigües interiors de gairebé tota l’Amèrica del Nord.
En ambients de característiques massa uniformes, es fa difícil trobar a prop les condicions complementàries necessàries per al desenvolupament de cicles de vida complexos, com els propis d’amfibis, insectes i molts paràsits. Tenen avantatge les espècies amb necessitats que s’han arribat a fer semblants durant tota la vida. Amfibis perennibranquis cavernícoles, com l’olm (Proteus), ho han aconseguit. Molts insectes de coves produeixen ous molt grossos i neixen en un estat larval avançat, sovint proper a la pupació. Resulta obligada una estratègia paral·lela en animals bentònics propis de les grans fondàries que han hagut de renunciar a tenir larves lliures planctonívores, perquè allà no hi ha plàncton, hi neixen, ells també, en una forma propera a la definitiva. Es produeixen menys ous, però són més voluminosos. Hi ha també l’alternativa de passar tota la vida o la major part d’aquesta en una fase semblant a la larval (anomenada neotènia) com fa l’olm, assolint, això sí, la maduresa sexual.
Aquestes regularitats i encara d’altres s’han d’avaluar de manera molt crítica. Potser sí que un augment de mida dels ous ajuda a suprimir estats larvals llargs i de naturalesa massa diferent de l’adult, en un entorn que no ofereix gaire diferenciació. Però és difícil de trobar una explicació convincent per als canvis d’al·lometria, és a dir, en les proporcions morfològiques, com quan l’allargament exagerat dels apèndixs del cos es relaciona amb un augment moderat de la massa corporal. Tant en coves com als estrats profunds del sòl i a les grans fondàries de mars i d’alguns llacs, la visió no és eficaç i sovint es perd.
Cal preguntar-se si els organismes paràsits, confinats als seus hostes respectius, no serien igualment hostatges de l’aïllament pel que fa als condicionaments imposats a la seva evolució. Certament és així, però se’n surt quan el parasitisme combina l’existència de formes larvals lliures, o de desenvolupament complicat, amb canvis d’hoste. Malgrat tot, la fidelitat als respectius hostes continua posant de manifest un aïllament relatiu.
El model de Mac Arthur i Wilson
El desig de donar expressió quantitativa o estadística a la riquesa de les biotes insulars suscità l’aproximació de Mac Arthur i Wilson (1967), nascuda de la consideració de les faunes insulars d’ocells, però aplicable d’una manera més general. Els autors es fixaren en la fauna d’ocells de les illes potser perquè la probabilitat d’immigració i extinció d’espècies està esbiaixada en relació amb el que passa als continents. Proposen una expressió general, segons la qual el valor de S, el nombre d’espècies presents en una illa, igual al nombre d’immigracions menys el nombre d’extincions (S = i - e), haurà d’estabilitzar-se entorn d’un nombre tal que dS/dt = aSi - bSe. A la pràctica sembla que S es regula de manera que les immigracions depenen poc de tal xifra, (i tendeix a 0), mentre que les extincions depenen més fortament del nombre d’espècies ja acumulades (e tendeix a 1).
Aquest model no permet prediccions gaire correctes en la seva aplicació a les illes, però ajuda a intuir els mecanismes de regulació naturals, i no solament amb relació a les illes. Serveix per a donar més valor a l’estudi dels poblaments insulars, perquè ens presenten de manera especialment efectiva i colpidora alguns aspectes de lleis que regeixen tota la diferenciació de la biosfera. Perquè hi ha estils d’evolució: la biosfera es troba sotmesa contínuament a canvis o pulsacions, que de tant en tant acceleren el flux d’energia i de tant en tant el desacceleren, o bé el mantenen generalment desaccelerat en situacions concretes o en llocs definits (com són els fons dels oceans o en els sòls, a les coves o a les illes).
Els fenòmens evolutius
Per a la comprensió dels fenòmens lligats a la insularitat cal prendre en consideració un cert nombre de processos evolutius específics dels sistemes aïllats. Tenen un rol especial les anomenades radiacions evolutiva i adaptativa.
La radiació evolutiva i la radiació adaptativa
Quan els processos d’especiació són d’una tal magnitud que, a partir d’una espècie ancestral, s’originen moltes espècies, es diu que s’ha produït una radiació evolutiva. Les radiacions es poden produir dintre d’un mega-arxipèlag, d’un arxipèlag o fins i tot dintre d’una illa (o un llac). En aquests casos, les espècies originades tant han pogut evolucionar per simpatria (dins una mateixa àrea) com per al·lopatria (a diferents àrees), si bé és més comú el segon cas. Seria, per exemple, el de les espècies, d’hàbits i ecologia similars, sovint confinades a un únic barranc o una única illa de les Canàries, dels cargols del gènere Hemicycla; de les tortugues de les Galápagos (Geochelone elephantopus); de les sargantanes Podarcis de les illes mediterrànies, o de les granotetes del grup pictissimus del gènere Eleutherodactylus de la Hispaniola.
Quan els processos d’especiació produeixen una sèrie d’espècies estretament relacionades que tenen uns requeriments ecològics diferents, moltes de les quals poden viure en simpatria, es parla de radiació adaptativa, terme que s’aplica a l’escampament d’espècies amb un ancessor comú a diferents nínxols ecològics. Les radiacions adaptatives originen grups d’espècies que sovint conviuen, i amb trets ecomorfològics que les identifiquen.
Són exemple de radiació adaptativa els pinsans de les Galápagos i de les Hawaii, els múrids de Filipines, els lemúrids, tenrècids i vivèrrids de Madagascar, les coníferes antigues de Nova Caledònia o els peixos cíclids dels llacs de l’Àfrica oriental. Un altre grup zoològic que ha donat lloc a grans radiacions adaptatives als llacs antics és el dels cargols aquàtics, amb dotzenes o centenars d’espècies endèmiques als llacs Tanganyika i Malawi (curiosament, no al llac Victòria), i també en altres llacs fora d’Àfrica: Okhrida a la frontera entre Macedònia i Albània, Baikal a Sibèria, Biwa a l’illa de Honshu (Japó), o Titicaca als Andes. Les conquilles de moltes d’aquestes espècies s’assemblen extraordinàriament a les de cargols marins, per la qual cosa hom havia pensat que aquests llacs podrien ser braços de mar aïllats; l’estudi anatòmic ha demostrat que no hi ha cap relació amb les espècies marines, sinó que es tracta d’un fenomen de convergència evolutiva sota pressions selectives semblants. El llac Baikal, d’altra banda, hostatja una tercera part dels amfípodes que es coneixen arreu del món (tant marins com d’aigües dolces), els quals, ben palesament, han radiat d’una manera extraordinària.
El cas dels pinsans de les illes Hawaii
Potser un dels casos més espectaculars de radiació adaptativa és el dels drepanidinins de les Hawaii, un grup monofilètic de passeriformes, que sembla que està relacionat filogenèticament amb les caderneres. Fins fa molt poc es considerava que la diversitat assolida per aquesta subfamília a les Hawaii incloïa vint-i-tres espècies d’onze gèneres diferents. Les espècies diferien per la forma del bec i de la llengua i pels hàbits alimentaris. Actualment, des del 1991, se sap que almenys catorze espècies i quatre gèneres més havien estat exterminats pels primers habitants de les illes: si la present varietat de becs ja és extraordinària, encara ho havia estat més abans de l’arribada dels humans.
Entre les formes conegudes d’antic hi ha una espècie, el “o’u” (Psittirostra psittacea), amb un bec convergent amb el de les cotorres, amb el qual capola fruits i flors. Una altra espècie, el pinsà de bec de cotorra de Maui (Pseudonestor xanthrophrys), que havia estat donada per extingida fins que fou retrobada el 1967 en una vall de difícil accés de l’illa de Maui, té un bec encara més robust, que fa servir per a tallar les branques on cerca els escarabats de la fusta dels quals s’alimenta. Dysmorodrepanis munroi, una espècie extingida abans del 1913, tenia un bec estrany, amb el premaxil·lar (la part superior del bec) corbat cap avall i la mandíbula (la part inferior), més curta, corbada cap amunt, de manera que mai no devia poder tancar del tot la boca; se suposa que el feia servir com si es tractés d’unes pinces. Algunes altres espècies, com ara el raspinell d’O’ahu (Paroreomyza maculata), de costums insectívors, tenen el bec llarg i estret, i la llengua allargada. Els “’amakihi” (Viridonia [= Loxops]) són també insectívors, però alhora nectarívors; l’“amakihi” gros (V. sagittirostris), espècie extingida al començament del segle XX, presentava una mandíbula convergent amb la dels trencapinyes, amb la punta del premaxil·lar i la de la mandíbula creuades. Al gènere Hemignathus, de talla superior i de bec més llarg i més corbat, s’inclouen espècies nectarívores i insectívores. Entre aquestes darreres n’hi ha una de notable, l’“’akipola’au” (Hemignathus wilsoni), que fa servir la mandíbula, curta i completament recta, per a picar la fusta dels troncs, talment com fan els picots amb el bec; el premaxil·lar, més llarg i corbat, l’utilitza per a treure els insectes dels forats que ha fet amb la mandíbula. Aquesta combinació d’una mandíbula recta i curta amb funció de martell, i un premaxil·lar llarg i corbat és única. Els gèneres més nectarívors tenen el bec molt llarg i molt corbat.
Entre les formes extingides de drepanidinins es pot destacar Chloridops regiskongi, l’espècie de bec més robust (semblant al del durbec). Hi havia espècies amb el bec de tipus pinçà, i n’hi havia amb un bec de tipus insectívor. Una espècie, Hemignathus upupirostris, tenia un bec semblant al de les puputs. Les dues espècies conegudes del gènere Vangulifer tenien un bec peculiar; de fet, no en trobem cap d’anàleg entre els ocells vivents.
El cas dels cíclids dels llacs africans
Un altre cas espectacular de radiació adaptativa és el que es dona entre els peixos de la família dels cíclids als grans llacs de l’Àfrica oriental. El llac Victòria hostatja 300 espècies endèmiques de cíclids, el Tanganyika, més de 230, i el Malawi, entre 500 i 1 000. Xifres encara més espectaculars si es té en compte que en tot el continent africà es coneixen en total unes 3 000 espècies de peixos d’aigües dolces. El tipus d’aliment disponible ha estat una pressió de selecció fonamental, atès que les diferències morfològiques, a banda una varietat espectacular de coloracions, rauen generalment en l’aparell bucal. Així, les espècies que pasturen el recobriment d’algues que creix damunt de les pedres del fons (Pseudotropheus) tenen el cap arrodonit i una dentadura en raspall, mentre que les que fan el mateix amb els revestiments algals de les plantes submergides tenen el cap afuat i les dents més fines. Les que s’alimenten de macròfits presenten una boca gran que conté una dentició poderosa, mentre els detritívors tenen el musell allargat i els fitoplanctívors pelàgics es distingeixen pel cos prim. D’altres poden allargar molt la boca per capturar animalets del zooplàncton, després de seguir-ne les migracions verticals diàries. N’hi ha que depreden mol·luscs, per la qual cosa tenen les dents bucals o bé les mandíbules faríngies, hipertrofiades i convertides en eines trituradores. Els insectívors tenen els llavis gruixuts, sovint prominents, mentre que els caçadors de gambetes tenen el premaxil·lar allargat com un bec i els caçadors de crancs tenen dents fortes i corbades i la mandíbula inferior llarga. També hi ha espècies ictiòfagues (Haplochromis, Boulengerochromis), cadascuna amb una tècnica de caça pròpia, però totes amb mandíbules grans i ben armades. Entre tanta diversitat hi ha també les que s’alimenten dels paràsits que troben damunt dels altres peixos, i no podien faltar les que, fent-se passar per netejadores, s’atansen a les seves víctimes i els arrenquen escates o els ulls. L’especialització arriba a les que mengen els ous acabats de pondre, just abans que les femelles els recullin per incubar-los a la boca; aquest extrem és superat només per especialistes a “besar”-les amb els seus llavis suaus per xuclar els ous o les larves que guarden, com fan els guramis (Helostoma). Altres espècies són d’hàbits igualment especialitzats, però encara desconeguts. Les comunitats més diverses es donen als hàbitats rocallosos, mentre que les comunitats d’aigües obertes són més pobres d’espècies.
Els cíclids dels grans llacs africans
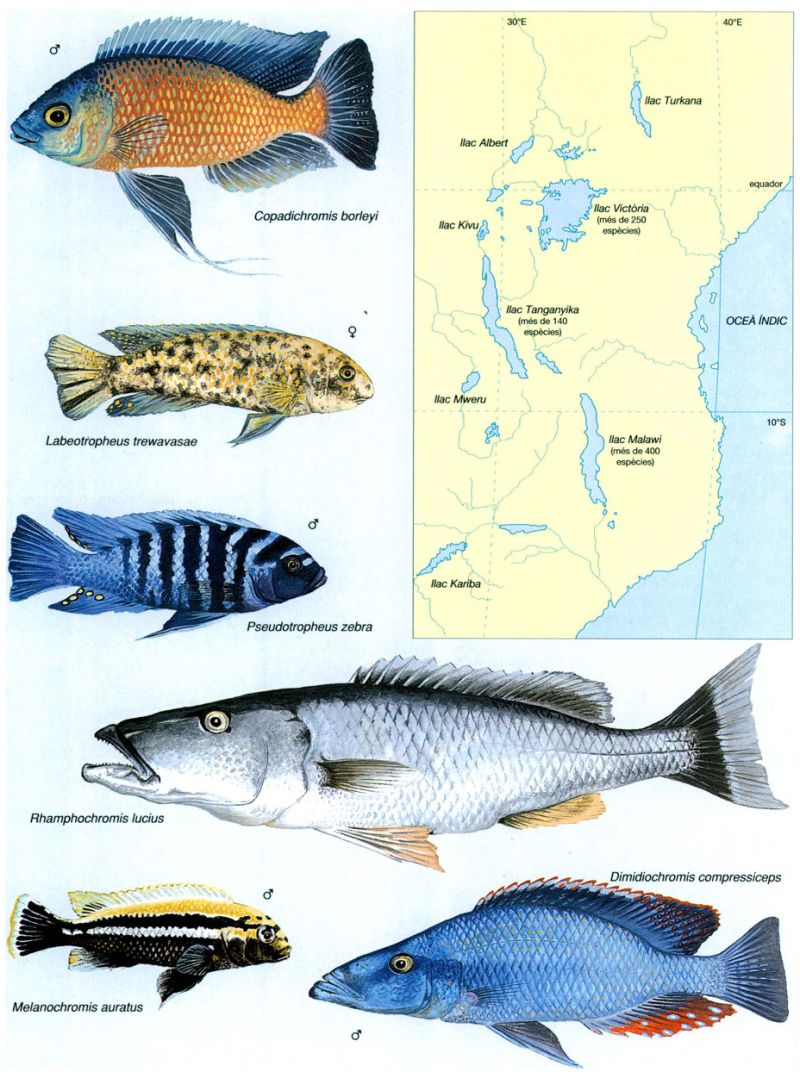
Jordi Corbera
La gran diversitat d’espècies de cíclids dels grans llacs de l’Àfrica oriental és l’exemple més espectacular de radiació adaptativa de tot el món dels vertebrats. És fàcil comprendre que els diferents llacs hostatgin espècies diferents, però el que és extraordinari és que en un mateix llac hagin pogut evolucionar i coexistir un nombre tan elevat d’espècies. Per exemple, al llac Malawi n’hi ha més de 400. Aquesta gran diversitat no hauria estat possible si els cíclids no haguessin estat capaços d’adaptar-se a aliments molt diversos, com ho demostra el fet que les principals diferències morfològiques entre les espècies resideixin, precisament, en l’aparell bucal.
Les adaptacions alimentàries, juntament amb un ampli espectre de mides i la colonització tant de les costes sorrenques i rocoses com de les aigües obertes i les profunditats, han possibilitat la coexistència en un mateix llac d’un gran nombre d’espècies. Les sis representades en són un bon exemple: Copadichromis borleyi, d’una mida màxima de 16 cm el mascle i 13 cm la femella, prefereix els fons de roca, i és predominantment planctòfag; Dimidiochromis compressiceps pot arribar fins als 21 cm i habita a les zones superficials cobertes de vegetació, on s’alimenta de petits peixos que caça a l’aguait; Labeotropheus trewavasae fa aproximadament 11 cm, es troba des de la superfície fins als 40 cm de fondària, i s’alimenta pricipalment de les algues que cobreixen les roques; Melanochromis auratus té una mida màxima de 10 cm, en el cas dels mascles, i habita en fons rocosos o mixtos, on s’alimenta, com l’espècie precedent, brostejant el recobriment algal de les roques; Pseudotropheus zebra pot arribar fins als 13,5 cm i habita exclusivament en fons rocosos, preferiblement nets de sediments, on brosteja la coberta algal de les roques, però en aigües obertes s’alimenta també de plàncton; i Rhamphochromis lucius, que fa fins a 40 cm de longitud, es troba a les aigües obertes, sovint a gran fondària, i depreda altres peixos.
L’evolució convergent
Sovint, en illes molt distants, pertanyents fins i tot a regions biogeogràfiques diferents, evolucionen espècies convergents. Això és conseqüència de diversos factors. En primer lloc, les espècies colonitzadores no són una mostra aleatòria del conjunt de les espècies continentals, de manera que fins i tot en indrets molt distants les espècies colonitzadores se solen reclutar entre els mateixos grans grups. D’altra banda, l’arribada dels mateixos grups d’espècies contribueix que les relacions ecològiques creades entre les diferents espècies no siguin gaire diferents en illes molt allunyades. Sovint hi ha espais ecològics buits en illes ben allunyades que són ocupats per espècies provinents d’estirps continentals emparentades.
Les respostes ecomorfològiques adaptatives que imposa l’evolució als ambients insulars (nesoevolució) són similars, i un resultat d’això és que de vegades diferents espècies endèmiques convergents que havien evolucionat sobre illes molt allunyades, i que mai no havien estat en contacte, han estat considerades com a una única espècie. Als llacs també s’han produït fenòmens de convergència semblants. Molts parells d’espècies de cíclids dels llacs Malawi i Tanganyika, tot i no estar directament emparentats, s’assemblen moltíssim, i al Titicaca, el Tanganyika i el Baikal es troben tricòpters no voladors pelàgics que han adoptat els hàbits dels coleòpters girínids, els quals són ben estranys als tricòpters.
Un exemple ben sorprenent d’evolució convergent a les illes és el dels elefants nans. Tots s’han extingit, però hi havia hagut formes nanes de proboscidis insulars a diferents illes mediterrànies, a les illes Channel de Califòrnia, a l’illa Wrangel del nord-est de Sibèria i també a diverses illes d’insulíndia (Timor, Flores, Cèlebes, etc.). N’hi havia que eren mamuts, altres eren elefants pròpiament dits i altres estegodonts, però tots s’assemblaven en el fet que, com a molt, feien 2 m d’alçada a la creu. Els més petits (mascles adults de menys de 80 cm a la creu) eren els de les illes mediterrànies de Sicília i Telos (Dodecanès). Algunes d’aquestes espècies s’extingiren abans de l’arribada dels humans, però moltes altres foren víctimes dels primers pobladors de les illes on vivien.
El cas de les tortugues terrestres
Un cas de convergència notable és el de les tortugues terrestres gegants. N’hi ha que són casos de gegantisme nesoevolutiu: espècies que han assolit la seva gran talla a partir d’ancessors continentals més petits. Aquest és el cas de la tortuga de les Galápagos (Geochelone elephantopus). Altres, en canvi, poden provenir d’espècies continentals encara més grosses, i per això, poden representar casos de nanisme nesoevolutiu. Aquest pot haver estat el cas de les extintes tortugues de Timor i Cèlebes, les quals potser estan relacionades amb la gegantina tortuga fòssil Indotestudo atlas.
El resultat és aproximadament el mateix: les espècies insulars de tortugues terrestres d’illes no habitades per mamífers són totes de gran talla. Sobre illes molt distants i sense relació passada ni present (Galápagos, Aldabra, Seychelles, Mascarenyes, algunes de les Antilles, Timor, Cèlebes, Canàries) han evolucionat tortugues de talla gran molt similars. Algunes d’aquestes espècies es van extingir abans de l’arribada dels humans a les illes, i moltes altres han desaparegut d’ençà de l’arribada dels humans, de manera que només en sobreviuen algunes.
La variabilitat de la tortuga gegant de les Galàpagos
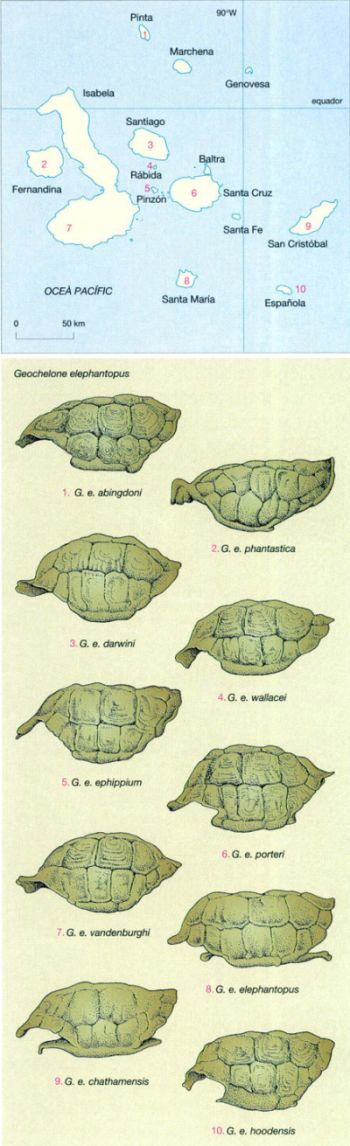
Jordi Corbera, a partir de fonts diverses
La variabilitat en el si de l’espècie Geochelone elephantopus, la tortuga gegant de les Galápagos, és tan gran que aquest queloni ha estat essencial a l’hora de comprendre l’abast de la variabilitat intraspecífica.
En total es reconeixen 15 subspècies de tortuga gegant, quatre ja extingides i una (G. e. abingdoni, de l’illa de Pinta) representada per un sol mascle (1994). Els cinc volcans de l’illa d’Isabela hostatgen cadascun una subspècie de tortuga diferent, mentre les altres es reparteixen per les illes més petites.
Una primera classificació de les tortugues es fa segons la morfologia de la closca, que pot ser en forma de sella o bé en forma de cúpula. Les tortugues que habiten les illes petites i baixes (Española, Pinzón, Pinta, etc.), on manca un estatge humit, i les que estan confinades a les regions més àrides de les illes grans i elevades (per exemple les del volcà Wolf d’Isabela), tenen la closca en forma de sella; la part anterior està aixecada de manera que els permet estirar el seu coll llarg i, com que tenen unes potes també llargues, poden arribar a les fulles més altes dels arbustos i els cactus que componen la vegetació de les àrees que habiten.
L’espècie amb una closca en forma de sella més típica és G. e. phantastica, de l’illa de Fernandina, actualment extingida. Les tortugues que habiten les terres altes humides de les illes més grans (Santa Cruz, Isabela) tenen la closca en forma de cúpula, com la de qualsevol tortuga terrestre, i el coll i les potes curtes.
A les terres altes la vegetació abunda durant la major part de l’any i les tortugues tenen menjar disponible més a prop del sòl, de manera que no els calen unes potes ni un coll llargs i la closca no s’ha modificat.
Els exemples més clars de closca en forma de cúpula són els de les subspècies G. e. darwinii, de l’illa de Santiago, i G. e. vandenburghi, del volcà Alcedo de l’illa d’Isabela.
De vegades, però, costa distingir si una tortuga té la closca d’un o altre tipus perquè també existeixen formes intermèdies.
El cas de les rates i els ratolins
Alguns casos interessants de convergència nesoevolutiva es donen en múrids, és a dir, rates i ratolins. Un de particularment significatiu seria el de la rata gegant de les Galápagos (Megaoryzomys curioi) una espècie que es va identificar l’any 1964 a partir de restes d’antigues egagròpiles d’òliba trobades a l’illa de Santa Cruz. Es va descriure com una espècie pertanyent al gènere Megalomys, gènere de rates gegants (30 a 35 cm de llargada del cos i la mateixa de cua) de les Petites Antilles, del qual només es coneixen restes de tres espècies, dues extingides respectivament a Barbuda i Saint Lucia al segle XIX (M. audreyae i M. luciae) i la tercera (M. desmarestii) a la Martinica al començament del segle XX, sembla que com a conseqüència de l’erupció i l’explosió de la Montagne Pelée el 1902. Tanmateix, s’ha hagut de concloure que l’espècie de Santa Cruz havia de ser inclosa en un gènere nou, endèmic de les Galápagos, que no pertany ni tan sols a la mateixa tribu que les Megalomys de les Antilles. Altres restes trobades a l’illa Isabela pertanyen a una altra espècie extinta, encara no descrita, del gènere Megaoryzomys.
Les rates de “malpaís” (Malpaisomys insularis), recentment descrites (1988) a partir de material subfòssil procedent de dipòsits volcànics del “malpaís” –els camps de lava recent de les Canàries orientals (Fuerteventura, Lanzarote i La Graciosa)– d’entre 5 000 i 1 000 anys enrere, són clarament convergents amb les rates dels camps de lava de les Galápagos del gènere Nesoryzomys, del qual hi ha cinc espècies extingides recentment i tres que encara sobreviuen. Es tracta de rates de cames llargues, adaptades a enfilar-se per les fissures de les roques, però ni tan sols aquesta habilitat ha permès la majoria d’espècies de resistir la influència de les rates comunes (Rattus) arribades amb els humans ni la predació per gossos i gats domèstics.
Les rates espinoses de les illes Ryukyu (Tokudaia osimensis i T. muenninki) tenen una dentició molt semblant a la de Rhagamys orthodon, un ratolí que va desaparèixer de Còrsega i Sardenya després de l’arribada dels humans. Ambdós gèneres endèmics deriven d’espècies continentals emparentades. Passa el mateix amb una sèrie de rates d’illes properes a la plataforma de la Sonda: Rattus palmarum i R. burrus, de les illes Nicobar; R. simalurensis, de l’illa Simeuluë, al nord-oest de Sumatra; R. lugens, de les Mentawai, al sud-est de la precedent; R. adustus, d’Enggano, al sud-oest de Sumatra; R. hoffmanni, de Cèlebes; R. tawitawiensis de Tawitawi, a l’arxipèlag de Sulu, al sud-oest de les Filipines. Tot i viure en illes allunyades, que mai no estigueren connectades entre elles ni amb el continent, totes aquestes espècies s’assemblen més entre elles que qualsevol d’aquestes amb les espècies de la plataforma continental.