El triomf de les poàcies
La primera sensació que tenen els qui arriben per primera vegada a l’estepa, la praderia o la pampa, és l’angúnia per la manca de recers arboris. L’absència de boscos, i fins d’arbres aïllats, sembla un error de la natura que els humans han intentat esmenar amb reiterades plantacions. Aquest esforç de ‘millora ecològica’ fou especialment important durant la segona meitat del segle XIX i la primera del XX, i tingué lloc gairebé simultàniament a Euràsia i a l’Amèrica del Sud i del Nord. Des d’aleshores, tant a les estepes com a les praderies i les pampes s’han plantat milions d’arbres que han arribat a formar franges boscoses. Les estepes i les praderies, però, continuen essent un món predominantment herbaci on el paisatge quasi sempre es veu per damunt de l’horitzó.
Els arbres esteparis: nàufrags en una mar d’herba
Existeixen pocs paisatges al planeta totalment desproveïts d’arbres. Abans que els humans comencessin a practicar l’agricultura, la vegetació arbòria cobria el 90% de la terra ferma. En molts paisatges dels que s’anomenen oberts, encara avui dia es troben alguns arbres aïllats, per exemple els baobabs (Adansonia digitata) i les acàcies (Acacia) a les sabanes africanes, els sacsaüls (Haloxylon) als deserts freds de l’Àsia central, o el bedoll nan (Betula nana), els salzes (Salix) i els verns (Alnus) a les tundres meridionals.
Estepa i praderia endins, però, només creixen plantes de veritable port arbori a les valls dels grans rius. En les condicions típiques de l’estepa, si hi ha vegetació llenyosa, només es dóna en forma d’arbustos de poca alçada que sobresurten enmig d’una mar d’herba; en són bons exemples l’espècie estepària de cirerer Prunusfruticosa, a les estepes de l’Europa oriental, les caraganes o acàcies russes (Caragana pygmaea i C. microphylla), les potentil·les (Potentilla) i les espirees arbustives (Spiraea) a les estepes àrides d’Àsia, o el sumac de Carolina o arbre vinagre (Rhus glabra) i l’avellaner americà (Corylus americana) a les praderies nord-americanes. L’absència d’arbres és un tret distintiu important dels paisatges esteparis. No és casual que a les estepes de la Sibèria meridional, Kazakhstan, Kirguizistan o Mongòlia es trobin sovint indrets habitats amb el nom kazakh (o l’equivalent en altres llengües turqueses) de “qoixaghaix”, que significa ‘adéu, arbre’.
La quantitat d’arbres o arbustos augmenta gradualment cap als marges de la zona estepària. A l’extrem occidental de les estepes eurasiàtiques, a Hongria, a l’anomenada “puszta”, pocs segles enrere encara hi havia paisatges oberts amb grans roures (Quercus), esparsos enmig de les planes cobertes d’herba. També a l’extrem septentrional de la zona estepària de l’Europa oriental, fins al segle XVIII eren habituals les anomenades boscanes parc, boscanes esclarissades amb una espessa capa d’herba sota l’estrat arbori. Pel que fa a l’Amèrica del Nord, a l’E de les Grans Planes, abans que s’iniciés l’època de l’agricultura intensiva existia un tipus de vegetació semblant de vegades anomenat ‘sabana de roures’. Als extrems meridionals de la zona d’estepes i praderies, prop del límit de contacte amb els deserts, augmenta el nombre d’arbustos i, en alguns llocs, en sòls sorrencs o gravencs, es troben densos matolls de diferents espècies de caraganes de mides diverses (Caragana bungei, C. spinosa), de potentil·les arbustives (Potentilla) i de “tar” (Nanophyton erinaceum).
En els marges d’estepes i praderies amb els biomes forestals s’havia observat, de manera reiterada, un avenç dels arbres en el si dels paisatges predominantment herbosos sense que, almenys en aparença, això fos resultat de cap intervenció humana. Habitualment, el desplaçament dels límits dels dos biomes es posava de manifest quan augmentava la pluviositat durant alguns anys consecutius. A Euràsia es poden trobar exemples d’aquesta irrupció d’arbres al marge septentrional de l’estepa, però aquest tipus d’invasió és especialment freqüent a les parts orientals de les praderies nord-americanes, on la probabilitat que hi hagi anys amb fortes precipitacions és dues o tres vegades més gran que a les parts més humides de l’estepa eurasiàtica.
Observacions particulars a llarg termini dutes a terme en el límit entre les boscanes i les praderies de l’Amèrica del Nord, en indrets protegits de la influència humana o dels animals domèstics mitjançant tanques, han demostrat que els arbustos, que són l’avançada dels boscos, prenen avantatge en poc temps sobre la vegetació herbàcia de la praderia. Els brots subterranis del sumac de Carolina o arbre vinagre (Rhus glabra), per exemple, que es troben a una profunditat de 10 a 30 cm, poden penetrar entre 7 i 9 m en un terreny ocupat per comunitats de poàcies (gramínies) de les praderies. Al cap de poc temps, els brots creixen cap amunt, emergeixen de la superfície del sòl, l’ombregen i retarden el creixement de la vegetació herbàcia. Gradualment, altres arbustos com el fruit de corall (Symphoricarpos orbiculata) i l’avellaner americà (Corylus americana), s’uneixen al sumac de Carolina per formar un tapís compacte sota el qual ja no poden créixer les poàcies. Aquestes discontinuïtats de la capa herbàcia cespitosa, abans compacta, possibiliten el creixement d’arbres més grans i, com a resultat, al cap de cinc anys una part de la superfície de la praderia és envaïda per un bosc que avança formant un front tancat. Tanmateix, en períodes de fortes secades, aquest bosc recula fins als seus límits anteriors.
Després de plantats, el creixement dels arbres durant els primers anys sol ser ràpid; només cal entrecavar la terra perquè es mantingui la humitat i desaparegui la capa d’herba. Les reserves d’humitat al sòl de l’estepa són suficients per a la transpiració dels arbres joves, però una plantació d’arbres adults necessita deu vegades més aigua per unitat de superfície que l’herba pròpia de l’estepa. Al cap de deu anys, a causa de la insuficiència hídrica, la transpiració dels arbres plantats a les estepes és entre la meitat i un terç menor que la dels arbres de la mateixa espècie en un bosc natural. Les fortes secades, que sobrevenen fatalment de manera periòdica, empitjoren la situació. Quan l’arbre arriba als 25 anys de vida, la humitat del sòl estepari sota seu ha quedat reduïda a un 14%, un nivell veritablement crític ja que en aquestes condicions els exemplars adults transpiren pràcticament tota l’aigua de què disposen. Quan els arbres assoleixen una alçada de 3-4 m, el seu sistema radicular, que creix proporcionalment a les parts aèries, no troba lloc a l’horitzó superior del sòl estepari i s’enfonsa alguns metres, aconseguint amb això que una gran part de la massa de l’arbre es trobi en els horitzons on gairebé no hi ha aigua. Els arbres de l’estepa, que necessiten grans quantitats d’aigua, es priven així ells mateixos de les últimes gotes. La insuficiència hídrica provoca primerament l’assecament de les parts arbòries superiors que accedeixen amb més dificultat a l’aigua; a continuació moren les branques inferiors i, finalment, tot l’arbre. Un arbre plantat a l’estepa viu 30 o a tot estirar 40 anys. Es pot concloure, per tant, que l’experiència de la plantació d’arbres a les estepes, les praderies i les pampes, ha demostrat que la raó principal de l’absència de bosc natural en aquest bioma és la incompatibilitat entre el funcionament biològic de les espècies arbòries i un règim d’humitat insuficient i inestable.
Les poàcies victorioses
La sequera, però, no ho explica tot. A les estepes i les praderies arriben i germinen regularment llavors i altres propàguls d’arbres transportats pel vent; tanmateix, fins i tot en condicions d’humitat suficient com les de les praderies nord-americanes d’herba alta, les plantes germinades no solen sobreviure més d’un parell d’anys. Són les poàcies, sense cap mena de dubte, les que impedeixen l’arrelament dels arbres. Les dades experimentals demostren que, si es planten llavors o plançons d’arbres en sòls llaurats i lliures de poàcies, aquests sobreviuen i creixen adequadament. En canvi, si no es destrueix la coberta graminoide, encara que es practiqui un reg artificial complementari no s’aconsegueix que les llavors germinin ni que els brots arrelin. La impotència dels arbres en la seva competència per l’aigua amb les poàcies s’explica, bàsicament, pel peculiar sistema radicular d’aquestes darreres. Les seves arrels fasciculades, amb una gran superfície total de pèls absorbents (100-200 km2/km2 d’estepa), tenen una extraordinària capacitat per absorbir la humitat dels sòls esteparis de manera ràpida i efectiva. Els arbres no gaudeixen d’aquest mecanisme de captació ràpida de l’aigua i, per això, no poden competir amb les poàcies. A més, un arbre, que té una gran massa de tiges i fulles situades a una alçada considerable per sobre del nivell del terra, necessita transpirar contínuament grans quantitats d’aigua. Una albereda, per exemple, gasta en període de creixement de 7 a 12 vegades més aigua que un gramenet xerofític de festuca (Festuca) que ocupi una superfície similar.
Els incendis, força freqüents quan el clima és sec, poden destruir temporalment la coberta graminoide d’estepes i praderies, dificultant-ne el nou desenvolupament i deixant clarianes que, aparentment, els arbres podrien aprofitar per a penetrar en l’estepa. Durant l’època de les tempestes de maig, un llamp pot inflamar en un segon l’herba seca que cobreix el sòl estepari des de la tardor, i el foc, amb la velocitat d’un antílop, corre pels espais oberts i ho converteix tot en cendres. Tanmateix, una gran part de la massa herbàcia resta amagada sota terra en forma d’arrels i altres òrgans subterranis. A part de les reserves de substàncies nutritives que ja tenen, les arrels aprofiten també les que contenen les cendres de les parts aèries que penetren en el sòl. El resultat d’això és que les poàcies cespitoses, que s’estenen centrífugament, s’emparen novament de tots els espais lliures, i de vegades, com per exemple en el cas d’alguns agròpirs (Agropyron), formen una capa d’arrels tan compacta que arriba a impedir el seu propi creixement.
Una sèrie d’experiments realitzats a les estepes i les praderies demostraren que l’incendi de periodicitat anual comporta un creixement hipertròfic de la coberta de poàcies. La protecció artificial contra el foc, en canvi, influeix favorablement en el desenvolupament de les altres plantes herbàcies, la biomassa de les quals s’incrementa unes quantes vegades en només dos anys. La massa d’herba morta s’acumula en la superfície del sòl de l’estepa i forma una capa de virosta que, al seu torn, dificulta el creixement de les poàcies. Si l’incendi és de periodicitat bianual, es formen comunitats herbàcies barrejades de poàcies i fòrbies.
L’eficiència de la producción primària
A les estepes i les praderies, el triomf de les poàcies i altres plantes herbàcies sobre les llenyoses té dues conseqüències ecològiques fonamentals. En primer lloc, la massa vegetal total és molt petita en comparació amb la de qualsevol bosc. I, en segon lloc, el quocient entre les parts subterrànies i aèries de les plantes es decanta clarament a favor de les subterrànies.
L’abundància de virosta i herba seca
La massa vegetal total a la zona de l’estepa eurasiàtica varia des d’1 kg de matèria seca/m2 en algunes regions properes als deserts freds fins a 3-4 kg/m2 a les estepes més típiques. Els extrems orientals de les praderies nord-americanes i les parts septentrionals de les estepes de l’Europa oriental, tot i ser suficientment humides, es caracteritzen per tenir uns valors moderats de fitomassa —del voltant de 2,5 kg/m2— que són, en termes mitjans, unes 100 vegades inferiors als dels boscos boreals i d’1,5 a 3 vegades superiors als dels deserts freds.
La composició de la massa vegetal a les estepes també es diferencia considerablement de la dels boscos o els deserts. La raó és que als paisatges d’herba rasa hi ha una gran quantitat de fitomassa morta i en la fitomassa viva hi predominen els brots de l’any; als boscos i als deserts, en canvi, predominen els teixits vegetals vius representats bàsicament per brots perennes. A la part aèria de la vegetació es distingeixen tres components: fitomassa verda, herba seca i virosta. L’herba seca, la constitueixen els brots que ja són morts i semisecs però encara lligats a la planta a través de l’arrel, i la virosta està formada per una catifa esponjosa de residus vegetals que han caigut a terra però que encara no s’han descompost.
A l’estepa típica, la fitomassa verda constitueix un 50% de la biomassa aèria total, l’herba seca un 35% i la virosta un 15%. A les estepes humides d’Europa, la quantitat de matèria verda de la fitomassa aèria disminueix fins a representar un 40 o un 45%, mentre que la proporció de teixits vegetals morts (herba seca i virosta) augmenta fins a ser el 55 o el 60% del total. A les praderies d’Amèrica, que reben fins a 1 000 mm de precipitacions l’any, la quantitat de fitomassa verda es redueix a un 30 o un 20%, mentre que la virosta i l’herba seca creixen fins a ser el 70 o el 80%. En contra del que pugui semblar, l’herba seca és força adequada per als consumidors heteròtrofs, ja que conté una concentració molt alta de substàncies nutritives. A més a més, d’herba verda només n’hi ha a l’estiu, mentre que de virosta i herba seca n’hi ha tot l’any. Aquesta és una de les raons que expliquen l’abundància de vertebrats i invertebrats herbívors a les estepes.
L’exigent presència dels herbívors
La massa total de vegetació aèria dels 270 milions d’ha de praderies nord-americanes pesa, en sec, al voltant de 2 100 milions de t. Amb aquesta quantitat es podien alimentar els 75 milions de bisons americans (Bison bison) —amb una mitjana de 450 kg de pes cadascun— i els 40 milions d’antílops americans (Antilocapra americana) —amb una mitjana de 70 kg de pes cadascun—, que habitaven les praderies abans de l’arribada dels europeus. El quadre, en principi, no ha canviat; al segle XX, la quantitat de caps de bestiar dels ramats domèstics és molt semblant a la d’ungulats salvatges que hi havia als segles XV i XVI. En tota la zona de la praderia, la proporció entre la massa d’animals herbívors (60 milions de t aproximadament) i la d’herba és d’1:50. Segons testimonis, la massa d’ungulats a les estepes d’Euràsia al segle passat també era molt gran i la proporció respecte a la massa aèria d’herba baixa encara era més ajustada.
Les praderies nord-americanes, que ocupen un territori més compacte que les estepes eurasiàtiques i en el passat tingueren una composició d’ungulats menys diversificada, es poden utilitzar com a exemple per a fer una valoració aproximada de la pressió de la fauna sobre la vegetació. Cada bisó americà (Bison bison) consumeix, en pes sec, 6 t d’herba en un any, i cada antílop americà (Antilocapra americana) 0,8 t. En conseqüència, la totalitat dels ungulats herbívors de les praderies nord-americanes necessita al voltant de 500 milions de t d’herba per a alimentar-se. A més d’aliment per als ungulats, la massa vegetal aèria constitueix també un plat saborós per a incomptables rosegadors, com per exemple els nombrosos gossets de les praderies (Cynomys), que poden arribar a assolir densitats de 5 000 a 15 000 individus/km2. Un experiment de camp dut a terme a les praderies demostrà que, a les zones protegides de la influència dels humans i d’aquests petits mamífers, la fitomassa era vuit vegades superior a la de les zones on dominaven els rosegadors. Un altre experiment evidencià que 256 exemplars de gosset de les praderies consumeixen en un any tanta herba com un bisó. Segons aquestes xifres, les necessitats alimentàries d’aquests rosegadors, en tota la zona ocupada per les praderies, equivalen a 700 milions de t d’herba a l’any en pes sec.
Els insectes constitueixen una competència seriosa per als mamífers. Són especialment notables les llagostes (ortòpters acrídids) que, per tal de cobrir les seves necessitats hídriques, mengen les parts verdes i sucoses de les plantes en quantitats que superen de molt el seu pes corporal. Es calcula que els eixams de llagostes, molt abundants als paisatges de tipus estepari, consumeixen anualment uns 200 milions de t de fitomassa verda i herba seca en el conjunt de l’àrea de la praderia. La seva dieta, molt variada, inclou dotzenes d’espècies herbàcies; els brots de festuques (Festuca) i d’artemísies (Artemisia) les atrauen especialment. Finalment, cal esmentar el nivell de consum dels diferents invertebrats herbívors que corren per la superfície del sòl, que pot arribar a ser anualment d’uns 1 000 milions de t de brossa estepària.
Així doncs, els vertebrats i invertebrats de la praderia necessiten en total 1 500 milions de t d’herba (en pes sec), quantitat que constitueix el 70% de tota la massa vegetal aèria. Cap altre bioma del món no experimenta una pressió constant tan forta dels heteròtrofs sobre la vegetació i, de fet, aquesta situació no seria suportable si no es donessin dues circumstàncies que la fan possible. En primer lloc, la rapidesa de la rotació biològica, que permet un creixement intensiu de la massa vegetal i reemplaça la que mor i és consumida. En segon lloc, l’existència d’una enorme massa vegetal subterrània, formada bàsicament per les arrels.
La renovació regular de la fitomassa aèria
El patró de rotació biològica en els ecosistemes esteparis es diferencia radicalment del que caracteritza els boscos boreals. Als boscos, la massa total de material vegetal és enorme (de 2 500 a 4 000 t/ha en pes sec) i està representada bàsicament per òrgans perennes; el creixement anual de teixits vegetals tendres, és a dir, la producció, se situa al voltant de 5,4 a 12 t/ha. Als ecosistemes esteparis, en canvi, la producció vegetal en un any iguala la fitomassa de l’ecosistema, és a dir, és de 25 t/ha de mitjana, i la relació entre ambdós paràmetres, producció vegetal i fitomassa, és entre 300 i 600 vegades més gran que als boscos boreals.
A les estepes i les praderies, el procés productiu pot durar de 2 a 7 mesos, segons la quantitat d’humitat disponible en el sòl. Els períodes de màxim creixement vegetal són, d’una banda, la primavera i el començament de l’estiu, moments en què s’aprofita l’aigua provinent de la fosa de la neu, i d’una altra la tardor, que és l’època en què l’arribada del fred fa disminuir l’índex d’evaporació del sòl. Així, hi ha una alternança de períodes de creixement intensiu dels teixits vegetals i de períodes en què els teixits moren. La descomposició dels residus de les arrels té lloc al llarg de tot l’any, però es realitza a diferents velocitats segons les condicions: és més ràpida quan hi ha humitat i calor, i més lenta quan el clima és fred (a l’hivern) o molt sec (la secada d’estiu). El procés de renovació del sistema radicular també és variable, de manera que les arrels que es troben a menys de 50 cm de la superfície del sòl es renoven més sovint que les situades a 1 m de profunditat, per exemple. Les plantes perennes renoven anualment un 65% de la seva massa radicular, mentre que les anuals la renoven completament. Amb tot, el fet que les plantes perennes, que tenen una massa més gran que les anuals, es descomponguin més lentament, fa que la durada de la rotació completa de totes les substàncies que formen la vegetació a les estepes s’allargui fins a 2 o 3 anys. Als boscos boreals, en canvi, la rotació completa triga dotzenes, fins i tot centenars d’anys, ja que anualment només una petita part de la fitomassa, i durant un període de temps molt curt, es diposita en forma de virosta.
El consum periòdic de massa aèria per part dels animals herbívors és un altre factor de selecció natural que estimula un creixement ràpid de les plantes d’estepes i praderies i compensa, així, la pèrdua dels òrgans fotosintetitzadors. Gràcies a la gran velocitat de producció, la vegetació de l’estepa, que té una massa insignificant, és capaç de sostenir l’existència d’un gran nombre d’animals herbívors, tant vertebrats com invertebrats.
El sòlid capital de la fitomassa subterrània
La major part de massa radicular de les plantes de l’estepa i, sobretot, de les poàcies cespitoses, es troba fins a una profunditat de 50 cm. A les estepes i les praderies, aquesta capa del sòl pot absorbir i retenir com una esponja tota la humitat procedent de les precipitacions, bé que durant un període de sequera prolongat pot arribar a assecar-se gairebé completament. En aquest horitzó superior, les fluctuacions diàries i estacionals de temperatura són més pronunciades que les de la temperatura de l’aire o dels horitzons més profunds. Això no obstant, aquesta capa sol ser la més càlida, i això fa que diverses reaccions bioquímiques s’hi produeixin de manera preferent. Precisament aquí, els processos d’acumulació i descomposició d’humus són especialment intensos i, en conseqüència, el sòl s’enriqueix en nutrients per a les plantes. No és estrany, per tant, que la major part de la fitomassa de l’estepa es concentri en aquest horitzó del sòl.
Les arrels de les plantes de la praderia
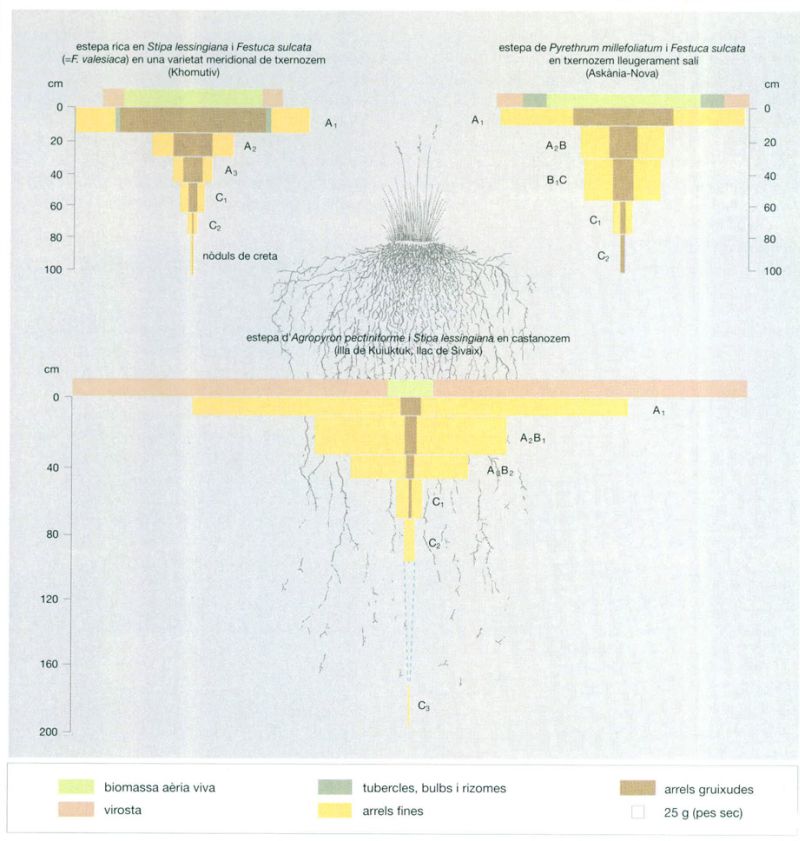
Jordi Corbera, a partir de Walter & Breckle, 1986
La massa radicular de les comunitats estepàries es concentra, sobretot, als primers horitzons del sòl, i a mesura que es va guanyant profunditat es va fent més escassa. A més, a les praderies i, especialment, a les estepes, aquesta massa radicular subterrània representa un volum molt més gran que el de la fitomassa aèria, cosa que es veu clarament en representar esquemàticament una poàcia típica d’estepa (a dalt, al centre). A les estepes d’Askània-Nova, es troben bulbs per sobre de la superfície de sòl a causa de la presència de Poa bulbosa. La major part dels hidrats de carboni i altres productes que es formen per fotosíntesi a les fulles i a les tiges verdes van a parar a les arrels. Durant el període de creixement màxim de la fitomassa verda, només un 30% de la producció és destinada a l’arrel, però a mesura que s’alenteix el creixement de les parts aèries, aquest percentatge va augmentant i pot arribar fins al 90%. No és sorprenent, per tant, que el sistema radical creixi notablement. A les estepes de la Sibèria oriental i a les del Kazakhstan, per exemple, el valor de la producció de les arrels pot assolir en un mes les 10 t/ha. Més tard, el mateix sistema radical o les parts aèries aprofiten les substàncies emmagatzemades per créixer. Des de l’arrel, l’aigua i els productes nutritius són transportats ràpidament cap a la part aèria per estimular la fotosíntesi. Tanmateix, el flux d’hidrats de carboni que passa de les arrels a les parts verdes no ultrapassa el 6% del flux que va de la part aèria a la subterrània. La reserva de substàncies nutritives roman intacta excepte en casos de necessitat extrems.
El sistema radical de les poàcies cespitoses recorda una gran barba amb nombroses arrels petites i pèls absorbents, la superfície total dels quals pot arribar als 200 km2/km2 d’estepa. Es tracta, d’altra banda, d’un òrgan extraordinàriament variable; els anys de secada, a les poàcies els creixen nombroses petites arrels xucladores destinades a la retenció d’aigua; els anys humits, per contra, produeixen arrels més gruixudes. Molt diferent és el cas de l’arrel de les dicotiledònies, que sol tenir un eix principal ben definit, que en ocasions penetra fins a 1 o 2 m de fondària. La tanarida siberiana (Tanacetum sibiricum), per exemple, pròpia de les estepes de la Sibèria oriental, té una arrel principal molt llarga que només es ramifica pels horitzons del sòl on s’acumula la humitat. El fet que les plantes de l’estepa tinguin les arrels tan llargues s’ha d’interpretar com un recurs per a emmagatzemar aigua i nutrients. Els reservoris prèviament acumulats, que poden conservar-se més d’un any, permeten a les plantes desenvolupar òrgans tant subterranis com aeris, fins i tot en condicions desfavorables. Així, els incendis de primavera o la pastura excessiva dels ramats poden destruir completament la fitomassa aèria en un indret concret, però aquesta es recupera tard o d’hora gràcies a les substàncies nutritives que les plantes emmagatzemen sota terra.
Un sistema radicular tan poderós garanteix un elevat creixement de la biomassa aèria, intensament consumida pels animals herbívors i sotmesa a incendis regulars. Així, es pot dir que la pressió dels ungulats no afecta excessivament la coberta herbàcia de l’estepa, perquè la superfície, trepitjada pels ramats fins que el seu estat recorda un paviment asfaltat, es refà fins a aconseguir la forma original només en 3 anys de supressió de les pastures. Les fortes sequeres tampoc no resulten fatals per a les plantes, ja que el 70% de la seva massa roman amagada sota el sòl. En tots aquests casos, la concentració de substàncies nutritives en les arrels constitueix un factor amortidor enfront de les condicions adverses a què estan sotmeses les parts aèries de la planta, i garanteix el seu desenvolupament.
Els privilegis dels consumidors
Paràmetres com la biomassa i el nombre d’organismes vius permeten fer una estadística de l’estat d’una comunitat en un moment determinat. Es pot caracteritzar la seva dinàmica segons la velocitat i el caràcter de la transformació de l’energia i els aliments a través de la cadena alimentària.
La quasi il·limitada oferta d’herba fresca
L’energia assimilada per les plantes a partir de la fotosíntesi es pot transformar seguint dues vies diferents. Al contrari del que passa als ecosistemes forestals, on el sistema de transformació de l’energia privilegia els consumidors sapròfags, és a dir, els animals i els fongs que s’alimenten de la virosta, de la fusta i de les restes mortes, els ecosistemes de l’estepa i la praderia es caracteritzen bàsicament per privilegiar el paper dels consumidors de fitomassa, és a dir, els animals de pastura i els herbívors en general. Aquí, la força principal que permet el desplaçament de l’energia i els nutrients d’un nivell tròfic a un altre són els ‘depredadors’ de pastures (ungulats herbívors, rosegadors, insectes), els quals, a diferència dels autèntics depredadors, es mengen la seva víctima (la planta herbàcia) lentament i per parts, i no tota de cop.
Això no obstant, si es jutgen basant-se en les interrelacions tròfiques, els animals herbívors es diferencien ben poc dels carnívors. Uns i altres mantenen relacions asimètriques amb les seves ‘preses’, fet que contrasta amb la relació més o menys igualitària que s’estableix entre espècies que competeixen per un mateix recurs tròfic; els diferents nivells d’interaccions que hi ha entre elles, però, es manifesten de manera menys clara en les estepes i les praderies que en altres biomes. Això es deu a la intensitat del procés productiu, a la presència d’una important reserva de substàncies nutritives en l’interior del sòl —inaccessible a la majoria d’herbívors— i al fet que la quantitat de veritables depredadors que limitin el nombre i l’activitat dels fitòfags és relativament petita.
En ecosistemes forestals, en particular al dels boscos boreals, els organismes fitòfags consumeixen tan sols un 10% de la producció vegetal anual. El 90% restant cau a terra en forma de fulles seques i branques, o s’acumula als troncs, a les arrels i a les branques, esdevenint així inaccessible a la major part de consumidors fins a la mort de l’arbre. Gràcies a la riquesa en elements estructurals i substàncies tànniques que presenten les plantes arbòries, tota aquesta producció, morta o dipositada en òrgans perennes, pot anar essent transformada lentament pels organismes sapròfags i reductors. A les estepes i les praderies la situació és molt diferent. Els fitòfags que viuen a la capa d’herba consumeixen del 30 al 60% de la massa vegetal abans que mori. Hi ha dues raons fonamentals que justifiquen aquest alt percentatge de consum: primer l’escassetat d’aigua, i segon la baixa concentració de substàncies nutritives en els teixits de les plantes herbàcies, d’altra banda molt suaus i suculents. A les estepes asiàtiques, per exemple, els imagos dels escarabats negres (tenebriònids) devoren diàriament uns 100 mg d’aliment per cada 50 mg de pes propi. Però els invertebrats que habiten el sòl d’estepes i praderies, malgrat la poca importància de la biomassa total que representen en l’ecosistema (de 2 a 25 g/m2), també tenen un paper fonamental en la transformació de les substàncies orgàniques. Així, en esmicolar les restes vegetals, els organismes fitòfags n’augmenten la superfície activa i preparen el terreny per a l’acció dels microorganismes.
Dels copròfags als microorganismes mineralitzadors
Hi ha tot un complex de petits animals copròfags (bàsicament larves però també imagos de coleòpters i dípters) que tenen una gran importància en el consum i el reciclatge dels excrements dels herbívors. Les larves dels escarabats piloters (escarabèids), per exemple, consumeixen 500 mg d’aliment per cada mil·ligram del seu propi pes corporal; juntament amb altres copròfags, en dos o tres dies devoren una buina de boví sense deixar-ne rastre. D’altra banda, el baix coeficient d’assimilació dels aliments que caracteritza els fitòfags de l’estepa explica que els seus excrements continguin una gran quantitat de residus vegetals semidescompostos, que, al seu torn, formen part de la dieta dels invertebrats sapròfags més petits (col·lèmbols, àcars) i de molts microorganismes. Aquesta successió de consumidors es veu afavorida pel fet que en l’intestí de fitòfags i sapròfags, com a resultat de processos bioquímics peculiars, es formen una sèrie de combinacions (vitamines del grup B i altres compostos biològicament actius) que estimulen la reproducció i l’activitat de determinats grups de microorganismes. Els excrements dels invertebrats es converteixen, per tant, en els centres de descomposició de la matèria orgànica i, per aquesta raó, els sòls de l’estepa es caracteritzen per albergar un complicat i poderós complex de reductors: fongs, protoctists i bacteris, en particular actinobacteris o actinomicets. La massa total d’aquests organismes als sòls de les estepes i les praderies arriba a assolir els 126 g/m2 en pes sec.
En els excrements més vells, amb el pas del temps, es dóna una successió dels diferents grups de microorganismes fins a arribar als que són capaços de destruir la cel·lulosa de les cèl·lules vegetals. La presència d’un gran nombre d’aquests microorganismes possibilita que la intensitat de descomposició de residus vegetals sobre els excrements dels invertebrats de l’estepa sigui de 2 a 3 vegades superior a la del sòl adjacent. En aprofitar la cel·lulosa de les parets cel·lulars com a substrat energètic, els microorganismes cel·lulosolítics redueixen dràsticament la quantitat d’hidrats de carboni emprats en la formació de les molècules dels àcids húmics.
La següent etapa en la destrucció de la matèria orgànica és la mineralització de l’humus. Als sòls de l’estepa, són principalment els actinobacteris del gènere Nocardia els que duen a terme aquesta tasca; als txernozems de les estepes típiques d’Euràsia, n’hi ha 7,2 milions d’exemplars/g d’humus, i als castanozems més clars 7,3 milions d’exemplars/g d’humus. Algunes fraccions d’húmiques dels sòls de l’estepa són mineralitzades pels microorganismes en un període d’uns 10 o 15 anys, mentre que a altres els calen de 1 000 a 1 500 anys. Per això, malgrat que la majoria dels processos de transformació són molt ràpids, els ecosistemes esteparis disposen d’una gran reserva de substàncies nutritives en forma de fraccions passives d’humus que garanteixen el funcionament estable de la comunitat.
La xarxa tròfica edàfica
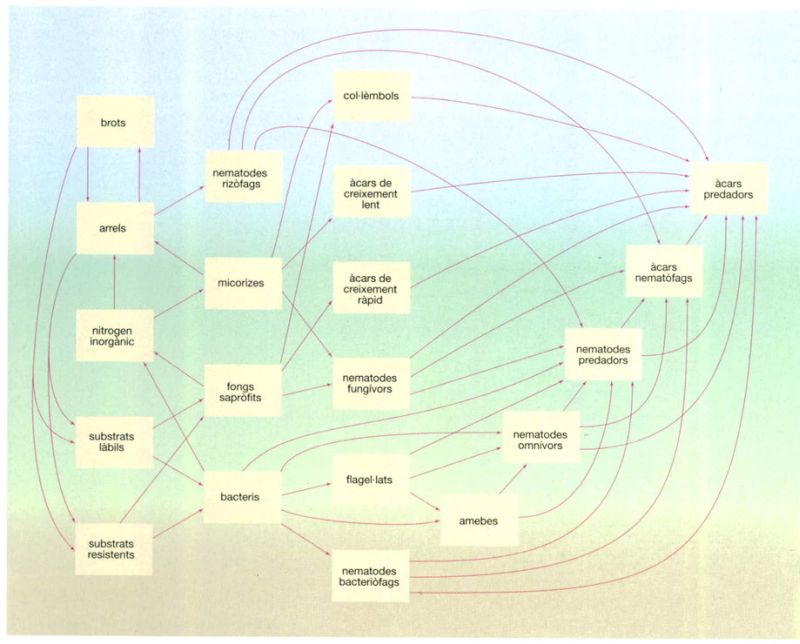
Jordi Corbera, a partir de Curry, 1994
La xarxa tròfica del sòl de les praderies és més llarga i complexa que les xarxes superficials, a causa de la major estabilitat dels ecosistemes edàfics. La major part d’organismes que habiten al sòl són detritívors, tot i que també hi són força nombrosos els animals de règim herbívor, i no hi manquen tampoc els carnívors. Tots aquests organismes estan inclosos en un dels tres grans sistemes tròfics del sòl: el microtròfic, el mesotròfic i el macrotròfic. Els organismes del sistema microtròfic es troben a la pel·lícula d’aigua que envolta les partícules edàfiques i les arrels, i són, principalment, bacteris i fongs que s’alimenten d’una gran diversitat de substrats. El sistema mesotròfic està constituït per col·lèmbols, àcars, nematodes, protozous i altres animalons que habiten als porus del sòl, on poden respirar oxigen. La seva abundància és determinada, doncs, pel grau de porositat del sòl. Els invertebrats de mida més gran, com cucs de terra (oligoquets) i milpeus (diplòpodes), que amb la seva activitat alteren l’estructura física del sòl, formen part del sistema macrotròfic. La importància relativa d’aquests tres sistemes varia molt d’una localitat a una altra, depenent de les condicions metereològiques i les propietats edàfiques, com el grau d’humitat, el pH, etc.
L’eficiència del reciclatge i la rapidesa de les successions
El torrent d’energia que flueix a través de l’ecosistema posa en moviment els cicles dels elements químics fonamentals i els seus compostos. Els nutrients que participen en els processos d’assimilació i descomposició passen alternativament de formes inorgàniques a formes orgàniques i viceversa. Així, gràcies a diversos processos d’intercanvi, el reciclatge biològic dels elements químics combina i implica tots els components de l’ecosistema.
L’elevada mobilitat dels nutrients
Les praderies, i especialment les estepes, són ecosistemes amb un alt índex d’intercanvi de carboni. La contribució dels processos abiòtics al conjunt de processos en què intervé aquest element només representa d’un 3 a un 4% de la quantitat total proveïda per la fotosíntesi. Els altres elements que limiten el creixement de les plantes, nitrogen, fòsfor i potassi, encara són més mòbils. La quantitat total de nitrogen en un ecosistema estepari típic, per exemple, és aproximadament de 6 g/m2 a les parts aèries de les plantes i de 16 g/m2 a les subterrànies. Després que s’hagi cremat la matèria orgànica, la massa dels elements minerals a les cendres és de 30 g/m2 a les parts aèries i de 120 g/m2 a les subterrànies. Si es comparen amb els dels ecosistemes dels boscos boreals, s’observa que aquests valors són lleugerament inferiors.
Per tal que els elements químics acumulats en la vegetació es puguin aprofitar novament en el cicle biològic han de tornar al seu punt d’origen, és a dir, al sòl. Poden fer-ho de quatre maneres diferents: per rentat de les parts aèries de les plantes, per deposició de les arrels (exsudació i despreniment de cèl·lules), per descomposició dels teixits morts de les parts aèries i per descomposició de les parts subterrànies mortes. El primer d’aquests processos, el rentat de carboni, nitrogen, potassi, fòsfor, sofre, sodi, magnesi i clor de les parts aèries sol tenir lloc, a les estepes, durant la segona meitat de l’estiu, període durant el qual disminueix la força de les unions entre elements en els teixits vegetals vells. Amb l’exsudació de les arrels, tornen al sòl el silici, el calci, el potassi, el sodi i el fòsfor i, en descompondre’s la capa de virosta, al voltant d’un 20% del silici i un 5% del manganès del total retornat al sòl de l’estepa tornen a ser accessibles a les arrels. La font principal de nutrients minerals per a les plantes, però, són les pròpies arrels mortes. Quan aquestes es mineralitzen, entre el 60 i el 90% de tots els elements químics retornen al sòl.
En un ecosistema d’estepa i praderia típic, el temps necessari perquè es produeixi una renovació completa de la reserva de nitrogen en la fitomassa, tant viva com morta, és d’aproximadament 0,4 anys. El temps de renovació de la reserva de carboni, en canvi, és dues vegades més llarg en la fitomassa viva i 1,5 vegades més llarg en la morta. Conseqüentment, per cada cicle complet de renovació del carboni en el sistema sòl-planta tenen lloc gairebé dos cicles complets de renovació del nitrogen. En els diferents ecosistemes d’estepa i praderia, el consum de nitrogen de les parts aèries de les plantes varia dels 2 als 10 g/m2 a l’any, mentre que el de les parts subterrànies oscil·la entre l’1,5 i els 55 g/m2 anuals. El consum d’elements minerals varia de 10 a 80 g/m2 a l’any en les parts aèries i de 20 a 325 g/m2 en les parts subterrànies.
En resum, pel que fa als cicles dels diferents elements, en ecosistemes de tipus estepari els processos biòtics són força més importants que els abiòtics. Aproximadament el 90% dels elements químics que migren dins l’ecosistema estan implicats en cicles biològics. Només el clor, el sofre i el sodi experimenten amb més freqüència transformacions abiòtiques que biòtiques. En definitiva, doncs, els ecosistemes d’estepes i praderies es caracteritzen per l’alt índex de mobilitat dels elements en els cicles biològics.
L’estalonament accelerat dels procesos de successió
A les estepes i les praderies, les successions de comunitats són un fenomen habitual. S’esdevenen com a resultat de les sobtades oscil·lacions climàtiques, de les activitats minadores dels rosegadors, del trepig i altres estralls causats pels ungulats, per la contaminació, etc. En tots els casos, l’estructura dels ecosistemes primaris es destrueix, parcialment o completament, i es va restablint després molt lentament fins a assolir un nivell màxim de maduresa anomenat clímax. Durant aquest procés de recuperació creix el nombre total d’espècies vegetals i animals, com també els lligams entre elles. Des dels estadis pioners de la successió fins al clímax, les espècies perennes desplacen les anuals en la composició de la comunitat vegetal; es van imposant les poàcies, altres plantes herbàcies i diferents formes cespitoses. Entre els insectes, els més petits, sovint caracteritzats per presentar fluctuacions sobtades de les poblacions, són substituïts per formes de grandària superior, les poblacions de les quals solen mantenir-se estables durant períodes de temps més llargs.
En les diferents successions (que es poden anomenar d’‘estepització’), el nombre relatiu d’elements químics implicats en transformacions abiòtiques disminueix en comparació amb els que participen en el reciclatge biològic. Al llarg de la successió, els cicles biològics esdevenen més tancats, la conservació de nutrients en l’ecosistema és més completa i el conjunt de processos d’intercanvi es fa més estable. A l’estepa, el temps que necessita l’ecosistema per a passar per tots els estadis de desenvolupament de la successió, des del primitiu en un sòl despullat —que, per exemple, sorgí per l’acció dels rosegadors, que el feren emergir des del fons de les seves galeries— fins a l’estadi de clímax d’estepa o praderia —amb poderoses poàcies cespitoses en sòls de tipus brunizems, txernozems o castanozems—, és d’entre 100 i 150 anys. La successió és molt més ràpida que la dels boscos boreals, que pot allargar-se fins als 300 o 500 anys, o la dels deserts freds, que pot ser de 400 a 1 000 anys.
L’aigua, cavall de batalla
En el complicat entramat de factors ecològics que determinen la diversitat dels ecosistemes esteparis, n’hi ha dos que influeixen especialment sobre la vegetació. En primer lloc, el dèficit d’humitat durant una part del període vegetatiu i, en segon lloc, les elevades temperatures diürnes, que també es donen durant un període força llarg. Sovint, aquests dos factors actuen conjuntament, complicant encara més l’existència de les delicades plantes herbàcies. El règim hídric deficitari es tradueix, d’una banda, en una limitació de la disponibilitat d’aigua, i s’enfronta, d’una altra banda, amb elevades pèrdues per transpiració que són conseqüència de la sequedat de l’aire i de les altes temperatures. Per tal de sobreviure en aquestes condicions, les plantes poden optar per diferents estratègies: en primer lloc, incrementar l’absorció d’humitat; en segon lloc, disminuir el consum d’aigua i en tercer lloc, desenvolupar un sistema que permeti suportar despeses hídriques importants.
Les barreres a l’evaporació
Per poder sobreviure, la majoria de plantes de l’estepa han desenvolupat una pubescència gruixuda a les tiges, les fulles i, de vegades, fins i tot les flors. Aquesta pubescència, especialment la de color clar, protegeix la planta de les grans pèrdues d’humitat per transpiració, ja que, en condicions de llum solar intensa, reflecteix molt bé la radiació tèrmica. D’aquesta manera, la planta suavitza el rigorós règim de temperatures i pot reduir l’evaporació —que manté la frescor de les parts aèries—. Moltes espècies, com l’herba blenera (Verbascum thapsus), la verònica blanca (Veronica incana), la nepta (Nepeta cataria), la “trudinitsa” peluda (Linosyris villosa), l’orpesa (Salvia aethiopis) i moltes altres, són tomentoses, és a dir, presenten tots els òrgans vegetatius i especialment les fulles coberts per un conjunt de pèls fins, disposats molt junts i formant com una borra (toment).
Una altra estratègia per a evitar la pèrdua d’aigua per transpiració consisteix a desenvolupar teixits tegumentaris resistents, sovint coberts d’una capa impermeable amb una cutícula gruixuda o una capa de cera. El desenvolupament de teguments protectors a les fulles explica que la coberta herbàcia de l’estepa tingui tons apagats i grisencs, que contrasten amb el verd maragda brillant de les comunitats vegetals dels prats. Entre les plantes estepàries que posseeixen una coberta de cera gris-blavosa, molt esteses a les estepes d’Euràsia, es poden esmentar, entre altres, una colla de lletereses (Euphorbia) i també el safranó rutè (Centaurea ruthenica), una herba d’1,5 m d’alçada amb enormes capítols de color groc-llimona, i el panical o card panical (Eryngium campestre), una apiàcia de fulles aspres i espinoses i inflorescència en capítol (i no en umbel·la, com és característic de la família a la qual pertany) composta per nombroses flors blavoses.
L’estalvi foliar i la suculència
La disminució general de la superfície d’evaporació també afavoreix la reducció de la pèrdua d’aigua; això s’aconsegueix amb fulles de làmines molt estretes que, si el clima és sec, fins i tot poden plegar-se en sentit longitudinal. Moltes poàcies i ciperàcies d’estepes i praderies desenvolupen aquest tipus de fulles, estretes, piloses i revolutes, en particular algunes espècies de pelaguers (Stipa lessingiana i S. stenophylla) o la festuca Festuca sulcata [=F. valesiaca].
Moltes plantes aconsegueixen un resultat similar de reducció de la superfície d’evapo-transpiració amb l’ajuda de fulles molt retallades; l’efecte és especialment evident si es comparen parelles d’espècies properes de regions humides i àrides. Només cal recordar, per exemple, els lòbuls de les fulles de la reina dels prats (Filipendula ulmaria), mesòfila i pròpia sobretot de les boscanes decídues, i comparar-los amb els de les fulles fortament retallades de la filipèndula comuna (F. vulgaris [=F. hexapetala]); s’observa que els de la primera són força més amples que els de la segona. Un fenomen semblant es dóna entre moltes espècies emparentades d’apiàcies, o amb les asteràcies del gènere Artemisia: les fulles de les espècies estepàries són més retallades i tenen els lòbuls més estrets que les d’espècies properes però pròpies de biomes més humits.
La reducció de les dimensions foliars és una altra estratègia per a disminuir la transpiració. Més concretament, és una característica de les esparregueres (Asparagus), que pertanyen a la família de les liliàcies, i de les efedres (Ephedra), un grup de gimnospermes arbustives molt peculiar. Als extensos solontxacs dels paisatges de les estepes meridionals àrides es troben també moltes plantes afil·les, en les quals la fotosíntesi no té lloc a les fulles, que no existeixen o estan extremament reduïdes, sinó a les tiges verdes. Les representants més característiques d’aquest interessant grup de plantes són les espècies halòfiles suculentes de la família de les quenopodiàcies, per exemple, la cirialera herbàcia (Salicornia europaea), algunes espècies de la barrella o “solianca” (Salsola) o el sarsasan (Halocnemum strobilaceum), un arbust d’aspecte poc habitual a causa de les seves branques carnoses i afil·les. La singular disposició de les fulles linears, en posició vertical, i no horitzontal, serveix també a algunes plantes per a evitar la pèrdua d’humitat per transpiració ja que, d’aquesta manera, disminueix substancialment la incidència dels raigs de sol sobre la superfície foliar. Aquesta disposició és pròpia de moltes asteràcies, entre altres les dels gèneres Serratula, Jurinea, Chondrilla, Centaurea i Lactuca.
En el cas de les plantes de fulla suculenta, escassament representades a l’estepa, l’estratègia per a sobreviure en condicions de dèficit d’humitat és diferent. Durant els períodes en què n’hi ha prou aquestes plantes acumulen aigua als òrgans vegetatius, especialment a les fulles; durant el període de secada, la gasten però economitzant-la. A banda dels coneguts crespinells (Sedum) i matafocs (Sempervivum), també pertanyen a aquest grup una sèrie d’espècies halòfiles de la família de les quenopodiàcies, totes elles representants dels gèneres Suaeda, Petrosimonia i Salsola, com també la cabridella (Aster tripolium), de la família de les asteràcies, i la gipsòfila salina (Gypsophila salina), de la de les cariofil·làcies, etc.