El metabolisme global de l’ecosistema
El metabolisme de les plantes pot resumir-se en els processos productius (producció primària bruta) i la respiració. La diferència entre producció primària bruta i respiració dóna la producció primària neta. Els dos processos bàsics de construcció i degradació metabòliques s’acceleren quan augmenta la temperatura i creixen també amb l’envelliment del sistema. El quocient entre les dues taxes és, així, relativament poc variable, però a mesura que augmenta la proporció de teixits que respiren sobre els que produeixen tendeix a acostarse a 1.
La producció neta global de l’ecosistema s’obté si de la producció primària neta que realitzen les plantes en restem encara la respiració de tots els organismes heteròtrofs. La productivitat o eficiència de l’ecosistema es pot expressar pel quocient entre la producció neta global i la producció primària bruta, és a dir, per la proporció de l’energia inicialment capturada per l’ecosistema que és finalment aprofitada i convertida en nova producció. En ecosistemes joves, que estan creixent ràpidament i que tenen poca biomassa vegetal no productiva, aquesta eficiència és màxima.
El metabolisme energètic de l’ecosistema va estretament associat als intercanvis materials que manté amb el seu entorn. La producció i la respiració són mesurables en termes d’entrades i sortides d’oxigen i diòxid de carboni. També es produeix fixació físico-química i biològica de nitrogen i devolució d’aquest element a l’atmosfera per desnitrificació, com també altres processos d’intercanvi gasós, als quals podem afegir les entrades d’aerosols i gasos procedents de l’activitat industrial. Amb la pluja, a més de l’aigua, arriben a l’ecosistema altres materials, sobretot ions que podran ser usats pels organismes, i les aigües de drenatge i infiltració profunda exporten materials fora de l’ecosistema. Una entrada important és la que s’esdevé per addició al sòl dels materials que procedeixen de la meteorització de la roca mare. L’entrada i la sortida d’organismes sencers o restes i d’òrgans de disseminació formen part també del balanç global, com ara l’aigua evapo-transpirada o la calor consumida en evaporació. Moltes d’aquestes entrades i sortides són quantificables amb més o menys dificultat i el seu coneixement ens diu molt sobre el funcionament de l’ecosistema en conjunt. Però també cal dir que els balanços energètics i bio-geoquímics no podran ser explicats satisfactòriament sense un coneixement detallat de moltes peculiaritats biològiques dels organismes que en formen part.
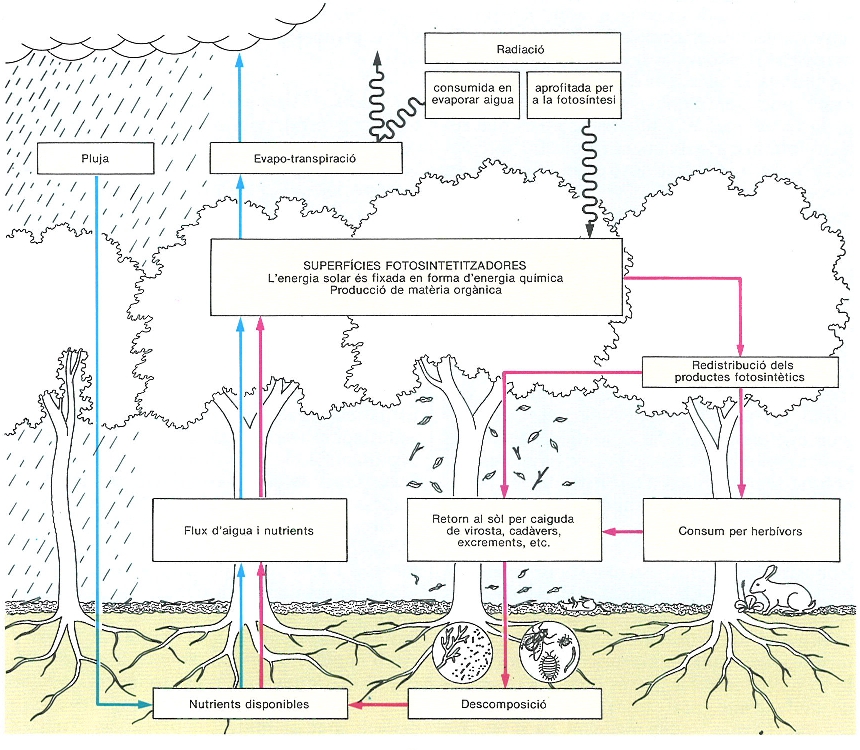
Metabolisme endosomàtic i exosomàtic en un ecosistema terrestre. A l’esquema s’aprecien dos cicles, el de l’aigua a l’esquerra i el dels materials a la dreta. El sistema funciona amb energia solar, que és fixada per les plantes en forma química (energia endosomàtica), i que mou la bomba d’aigua que porta aigua i nutrients a les parts fotosintetitzadores mitjançant l’evapotranspiració (energia exosomàtica). Aquesta darrera energia és també indispensable per a la producció, encara que no sigui fixada químicament.
Mercè Cartañá.
Qualsevol que sigui el camí metabòlic seguit, C3, C4 o CAM, l’energia que les plantes terrestres arriben a fixar és una part ben petita de la que en principi hi ha disponible: entorn d’un 1% per a la majoria de plantes i molt menys, un 1‰, si es considera el conjunt de la biosfera. Aquesta energia endosomàtica, més aviat escassa, de la qual depenen, en definitiva, tots els components vius de l’ecosistema, requereix una energia externa o exosomàtica molt més gran. En els sistemes terrestres naturals l’energia externa és sobretot la part de l’energia solar rebuda que es consumeix en l’evapo-transpiració de l’aigua. En efecte, l’aigua que s’evapora a través de les estructures aèries vegetals és la que realitza el transport de nutrients contra la gravetat, des del sòl fins a les fulles. Així, l’energia consumida en l’evapo-transpiració mou la bomba que fa funcionar la circulació dels materials dins l’ecosistema. La proporció entre l’energia consumida en aquest procés i l’energia fixada per la fotosíntesi varia normalment entre 25 i 150 (és a dir que calen de 200 a 1000 litres d’aigua per cada quilo de matèria orgànica construïda per les plantes).
Sovint s’ha presentat el funcionament dels ecosistemes lliures de pertorbacions i en fase avançada del seu desenvolupament natural espontani, d’una forma una mica utòpica, segons la qual els cicles materials tendirien a ser molt tancats (amb un màxim aprofitament dels recursos nutritius, per exemple) i el sistema, molt divers i estable en la seva composició i estructura. Recordem que la retenció de materials mai no és completa, i fins i tot que poden incrementar-se les pèrdues quan el sistema perd velocitat de creixement. A més, de forma natural sempre es produeixen pertorbacions, com a resultat de fenòmens esporàdics o cíclics del medi físic o biològic (ultra les que pugui provocar l’home, és clar). El metabolisme global pot ser, per tant, sotmès a pulsacions més o menys intenses. En els ecosistemes terrestres mediterranis, per exemple, el règim irregular de pluges, amb precipitacions totals escasses, però amb tempestes molt fortes i relativament freqüents, no hi ha dubte que aquestes tempestes tenen un efecte marcat en la funcionalitat. Molts dels processos de canvi, des del transport horitzontal de materials sòlids o dissolts fins a l’activació dels microorganismes descomponedors (i, per tant, de les taxes de descomposició) i l’increment de les taxes de producció, s’associen de forma significativa a les tempestes, de manera que el metabolisme global s’intensifica molt durant les pluges i immediatament després. Els valors globals d’entrades i sortides no ens diuen res sobre la manera com aquestes s’han repartit al llarg del temps.
Algunes pertorbacions són més periòdiques i previsibles, de forma que els organismes s’hi adapten més fàcilment o amb més precisió, com és el cas dels ritmes estacionals de temperatura, la qual cosa no exclou de tant en tant una situació anòmala prou important per a incidir fortament en el sistema i produir-hi repercussions que es prolonguen molt (les gelades excepcionals del 1956, el 1963 i el 1985 han deixat, o deixaran sentir, llurs efectes en l’estructura espacial i el funcionament de molts dels nostres ecosistemes durant molts anys). Altres tipus de pulsacions poden ser des de la caiguda d’un arbre fins a una plaga d’insectes o de fongs paràsits, un incendi, una malaltia d’una espècie important (pensem en la mixomatosi dels conills, per exemple) o la introducció d’una espècie exòtica.
Com a resultat de les pulsacions, certs processos s’acceleren i algunes reserves (biomassa viva, nutrients al sòl, etc.) es modifiquen sensiblement. L’equilibri relatiu entre construcció i degradació (fotosíntesi-producció i respiració-mineralització) canvia temporalment o definitivament. Podem observar des de petits increments o reduccions de la fullaraca acumulada a terra, fins a serioses alteracions en l’estructura vertical o horitzontal de la vegetació, seguits sempre de fenòmens dinàmics de reajustament. D’aquests fenòmens, que constitueixen la resposta dels ecosistemes a les pertorbacions, ens n’ocuparem tot seguit.
Els ecosistemes terrestres en el temps
Quan les pertorbacions són de magnitud relativament petita, s’ha dit que es produeix després una «cicatrització» de l’estructura alterada. Per exemple, després de caure un arbre es forma una clariana; la llum que hi entra estimula el creixement lateral de les branques dels arbres veïns i el de les plàntules que hi puguin germinar. En relativament poc temps, la clariana es tanca i acabarà essent impossible de reconèixer-la. Durant aquest temps, però, la clariana ha estat un punt «calent» en l’activitat metabòlica del bosc. Si la pertorbació es presenta a una escala més gran també s’origina un procés de reorganització, que els ecòlegs solen anomenar successió. Sovint, aquest procés condueix a refer un sistema molt semblant al primitiu, després d’una sèrie de canvis en la composició d’espècies i en el funcionament general de la comunitat. No sempre, però, les coses són així. Diversos factors poden incidir-hi, estroncant l’avanç en alguna fase o conduint-lo a resultats diferents, segons la intensitat i les característiques de la pertorbació inicial i les possibilitats concretes dels organismes presents.
La importància de l’estudi de les successions i cicatritzacions és prou evident. Se’n desprèn la base teòrica per a entendre la resposta dels ecosistemes a les pertorbacions de tota mena i la base factual per a ajudar a fer models realistes d’aquestes respostes en ecosistemes concrets i per a elaborar estratègies adients de gestió. Des del punt de vista teòric, l’èmfasi principal el posaríem en el fet que aquests estudis expliquen els processos autoorganitzatius dels sistemes naturals segons regles força generalitzables. Una d’elles és que la complexitat estructural tendeix a augmentar progressivament, tant pel que fa al volum d’espai aeri o subterrani ocupat i controlat pels organismes, com a la varietat de formes biològiques que s’hi van incorporant. Associada a aquesta creixent complexitat trobem també una creixent interiorització dels recursos per part dels organismes (per exemple, els nutrients minerals són cada cop en més proporció i més temps dins la biomassa) i un control més gran del medi físic inclòs en el sistema (les condicions de temperatura i humitat són menys fluctuants dins un bosc que a fora).
No cal dir, però, que, igual que passa a qualsevol sistema, inclosos els organitzats per l’home (indústries, institucions culturals i científiques, administració, etc.), una complexitat més gran comporta també que els elements productius representin una part progressivament més petita, enfront d’altres elements dedicats a garantir el transport, el sosteniment, la protecció, etc. Les fulles són una gran part de la biomassa en una comunitat d’herbes, però poden ser només un 1% del total en un bosc. Per tant, la producció primària creix menys que la biomassa, de manera que la relació producció/biomassa (l’invers de la taxa de renovació) decreix. Les necessitats de manteniment de l’estructura, en canvi, augmenten, ja que són proporcionals a la biomassa, i, per tant, la respiració total del sistema es fa cada cop més gran. El límit és determinat per una situació en què processos de construcció i processos de degradació s’equilibren i el quocient producció/respiració es fa proper a la unitat.
El màxim d’autoorganització o maduresa assolit al llarg de la successió correspon, doncs, a ecosistemes amb estructura vertical aèria i subterrània complexa, la qual modifica i esmorteeix considerablement les condicions climàtiques, amb una elevada diversitat d’organismes en els diferents nivells d’aquesta estructura, amb una taxa de renovació de la biomassa baixa i amb una producció que tot just compensa la respiració. Aquests ecosistemes (els grans boscos tropicals verges en serien un exemple paradigmàtic) tendeixen a ser relativament estables en el temps, si no s’hi produeixen canvis ambientals externs importants, i freqüentment hi trobem relacions de dependència molt intenses entre espècies. No són, però, de cap manera, sistemes immòbils; hi ha una forta dinàmica de petites pertorbacions que afecten àrees reduïdes (caiguda d’arbres, branques o grans fruits, per exemple), seguides de les corresponents reestructuracions i cicatritzacions.
A continuació, la taula mostra les regularitats en les successions ecològiques, establertes per diversos autors, en particular E.P. Odum i R. Margalef. Altres autors consideren que la base factual d’algunes d’aquestes regularitats és escassa i que part de les afirmacions que es fan són tautológiques; però el recull dóna una idea general sobre les tendències d’autoorganització dels ecosistemes que continua tenint una gran influència en el pensament ecològic actual.
Tendències de variació de diferents paràmetres en la successió ecològica
| PARÀMETRES | TENDÈNCIA DE LA VARIACIÓ EN ELS ESTADIS MADURS DE LA SUCCESSIÓ |
|---|---|
| Biomassa i producció | Creixen per separat i amb ritmes diferents |
| Producció primària/biomassa | Disminueix; l’excés de producció s’acumula com a biomassa fins que la producció s’iguala a la respiració |
| Biomassa heteròtrofs/biomassa total | Creix |
| Cadenes tròfiques | Més llargues |
| Biomassa poc activa (estructures de suport o de defensa, com ara la fusta, les closques, etc.) | Creix |
| Clorofil·la/biomassa total | Disminueix |
| Nombre de paràsits, simbionts, altres formes de coevolució intensa | Creix |
| Nombre total d’espècies a l’ecosistema | Creix |
| Temps de permanència dels nutrients fora dels organismes | Disminueix |
| Control del transport pels organismes (plantes superiors, fongs, etc.) | Creix |
| Fluctuació de grandària de les poblacions | Disminueix |
| Ritmes interns i mecanismes homeostàtics | Creixen |
| Energia disponible | Disminueix |
| Eficiència dels organismes | Creix |
| Dades de diversos autors | |
Clímax
Per regla general, es considera que l’evolució de la vegetació sota determinades condicions climàtiques tendeix a formar un tipus de comunitat i de sòl de màxima maduresa, amb les característiques que acabem d’exposar.
Els geobotànics designen aquest tipus de comunitat amb el nom de clímax i consideren que per a cada tipus de clima hi ha una vegetació i un tipus de sòl climàcics. Per tant, és possible de cartografiar un país atenent els dominis climàcics que s’hi poden reconèixer. Cada domini seria, en realitat, ocupat per la pròpia comunitat clímax corresponent o per algunes de les comunitats de la sèrie successional que hi condueix. La noció de clímax té, no obstant això, un punt d’excessiva simplificació i d’ideal platònic. En la pràctica, sovint no és possible de seguir els processos successionals reals a causa de llur durada. Les tendències es dedueixen de la coexistència sobre el terreny de comunitats més o menys madures que l’investigador ordena sobre el paper en una possible seqüència temporal. El procediment és justificat, però les conclusions en molts casos no poden sotmetre’s a prova.
Tot i admetre la realitat de les tendències successionals i el sentit o la utilitat d’una cartografia dels dominis climàcics, hi ha moltes raons per a aprofundir més en els processos successionals i per a detectar dificultats en l’aplicació universal de la teoria climàcica. A mesura que les condicions ambientals esdevenen més adverses i limitants, l’autoorganització té més dificultats per a progressar. Sovint esdevé incapaç de constituir un microclima i un sòl de característiques prou controlades i, en definitiva, quasi independents del medi extern. Aleshores, sobre diferents tipus de substrat s’arribarà a diferents màxims de maduresa i, sota un mateix clima, es tindrà un mosaic de comunitats. D’altra banda, la constitució d’un sòl madur és un procés lent. Si un sistema és sotmès a pertorbacions molt profundes, la recuperació cap a la clímax pot veure’s blocada per la necessitat de reconstruir el sòl i de disposar de llavors (cosa difícil si la pertorbació ha estat, a més, extensa).
Un altre cas que cal considerar és aquell en què es dóna un règim de pertorbació d’elevada freqüència. En aquests casos (per exemple, incendis recurrents cada 15 o 20 anys per causes naturals o antròpiques) l’evolució de la vegetació és constantment interrompuda. No s’arriba mai a depassar un cert llindar de maduresa en l’organització. Plantes i animals són seleccionats per llur capacitat de suportar aquestes altes freqüències de pertorbació, i no té gaire sentit parlar de clímax. D’altra banda, com que el clima no és estàtic, sovint es farà gairebé impossible de predir l’estat final a llarg termini de l’evolució successional. Sigui com vulgui, el que interessa a la pràctica és sempre una predicció a terminis molt més curts, d’uns pocs anys com a màxim, per poder elaborar una metodologia de gestió adient a cada cas.
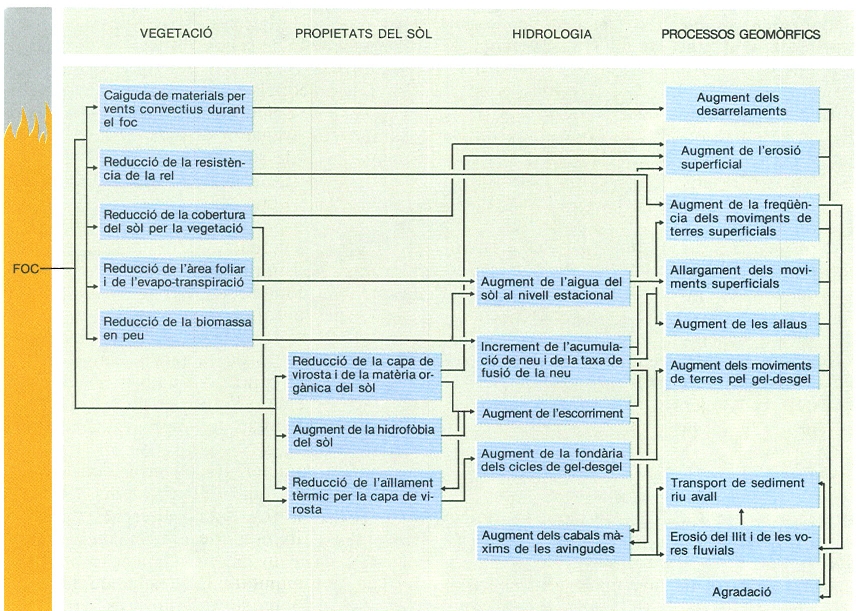
Efectes ecològics del foc, en diferents aspectes: vegetació, sòl, hidrologia, geomorfologia.
Carto-Tec, a partir de fonts diverses.
Des d’un punt de vista ecològic, és important, sobretot, d’augmentar la nostra capacitat per a trobar regles i per a establir models que ens permetin d’entendre aquests processos de canvi. Normalment, es compleixen les regles generals, abans enunciades com a tendències successionals, però la marxa concreta del procés i els tipus reals de comunitats que s’aniran desenvolupant no configuren una seqüència tan determinista com ens portaria a pensar la teoria climàcica. Després d’un canvi catastròfic, el procés d’autoorganització pot seguir fases més o menys lineals, però també poden donar-se ramificacions, punts on són possibles diverses alternatives. Sovint, el fet que se’n segueixi una o altra dependrà de factors com el moment de l’any, l’estació en què s’ha produït la pertorbació, la seva intensitat, la presència o no de llavors a terra per a iniciar la reconstrucció, que hi hagi o no pluges els dies immediats, etc. És a dir, el sistema pot seguir un camí o un altre d’acord amb fenòmens que, sovint, estan determinats des de fora del sistema, i que, per tant, podem considerar més o menys atzarosos. No cal dir la importància que té aquest enfocament si es volen abordar qüestions d’aplicació i de gestió dels sistemes naturals.
Si l’estudi de la dinàmica dels ecosistemes planteja problemes evidents relacionats amb el temps necessari per al desenvolupament de la successió, més senzill és l’estudi de les anomenades microsuccessions. Un tronc mort en un bosc, una buina de vaca en un prat són materials nous que apareixen com una illa al mig de la mar. Tot seguit, són envaïts per diversos organismes que comencen a emprar el nou recurs o a ocupar el nou hàbitat. A mesura que es modifiquen les condicions físiques i químiques del material altres organismes hi troben condicions favorables. El resultat és que s’hi succeeixen, en poc temps, veritables sèries de comunitats d’organismes, que poden ser seguides amb relativa facilitat, al llarg de les quals hom pot identificar les principals tendències que caracteritzen les successions. A qualsevol ecosistema, s’hi produeix contínuament un gran nombre d’aquestes microsuccessions, moltes de les quals són essencialment heterotròfiques. A l’inici hi ha una aportació d’energia i la microsuccessió es dóna aprofitant-la i degradant-la fins a esgotar-la. Per aquesta raó, no pot haver-hi un estat final madur, sinó que el procés s’extingeix amb la desaparició del recurs que l’ha generat, a diferència del que passa amb les successions que depenen del flux autotròfic d’energia, és a dir, de l’aprofitament directe de la radiació solar, que a la nostra escala de temps pot considerar-se com un flux de duració il·limitada.
Successió i explotació
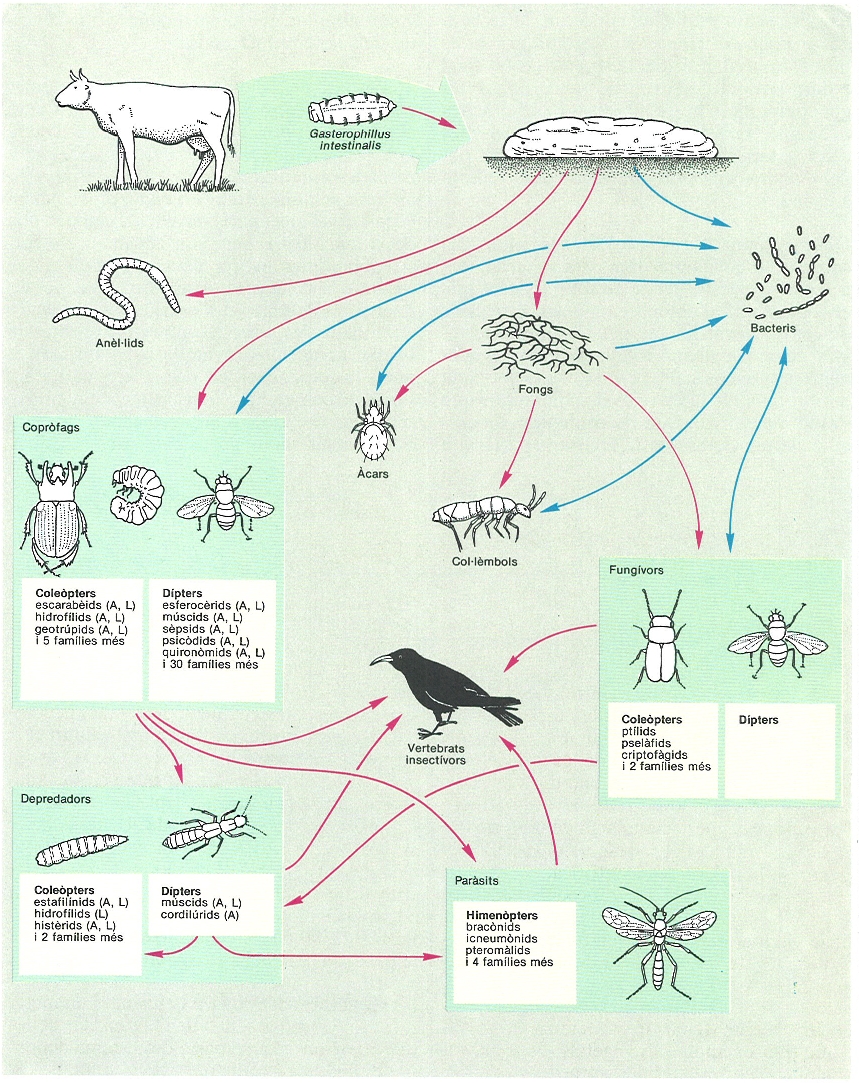
La microsuccessió que s’origina i la complexitat de relacions tròfiques que s’estableixen damunt d’una buina de vaca queden molt sintetitzades en aquest esquema. S’hi representa una xarxa tròfica ideal basada en el conjunt d’organismes trobats durant els primers 60 dies de successió en buines de la vall d’Aisa (Osca). En realitat, a cada moment de la successió, s’hi troben únicament una part dels organismes indicats. Inicialment predominen les larves de dípters i els adults i les larves d’hidrofílids. Més endavant, els coleòpters depredadors i copròfags augmenten en nombre i diversitat; els frugívors apareixen també en aquesta segona fase. Cap al final del procés és freqüent trobar invertebrats edàfics que usen la buina ja seca com a lloc de refugi i cria. Els anèl·lids fan un paper important en la desaparició final de la buina, però estan molt supeditats a les condicions d’humitat del sòl. El moment de la deposició (època de l’any, dia, nit), el lloc (recobriment arbori, proximitat de l’abeurador, orientació) i les condicions climàtiques (vent, insolació, temperatures) decideixen les espècies concretes que intervindran en la successió de descomposició d’una determinada buina. En el dibuix: A adult, L estat larval.
Amadeu Blasco, original de J. Franch.
Segons una teoria recent, es pot admetre que els ecosistemes es troben en un estat estacionari si la taxa de fotosíntesi i formació de matèria orgànica és equilibrada per la taxa de mineralització de la matèria orgànica. Una pertorbació pot considerar-se com un desajustament del cicle de matèria, que produeix canvis en la biocenosi (composició específica) i el medi físic (microclima, quimisme del sòl, etc.). La variabilitat del clima, la substitució d’individus o les extraccions de biomassa per consumidors són causes de pertorbació i es tradueixen en desajustaments temporals de les taxes i situacions d’estrès. La capacitat per a retornar a l’estat estacionan del cicle material s’anomena elasticitat i depèn de les propietats del sòl, mentre que la de retornar a la mateixa composició específica sol anomenar-se resiliència i té molt a veure amb la capacitat de regeneració de les poblacions. Si la desviació és molt gran i perdura molt de temps s’esdevé una inestabilitat catastròfica. Pot produir-se per acumulació de matèria orgànica, disminució de la reserva edàfica de nitrogen o inhibició de la nitrificació de l’humus, empobriment de l’acidificació, manca de nutrients i toxicitat àcida de les rels, reducció de la vitalitat dels arbres (en funció de la susceptibilitat de les espècies i de llur tipus biològic), etc.
Quan un ecosistema, o un determinat nivell tròfic (per exemple, els vegetals) és sotmès a explotació per organismes introduïts, per factors del medi físic (per un nou nivell tròfic, per exemple d’herbívors), o per l’home, la seva taxa de renovació s’incrementa, com ho fa també la relació entre òrgans productors i estructurals, mentre que la diversitat i la complexitat estructural tendeixen a disminuir. En termes de successió, és com si el sistema es rejovenís, es fes menys madur que no era. Tota explotació té, en aquesta òptica, efectes del mateix sentit, contraris al de les tendències autoorganitzatives espontànies de l’ecosistema, i condueix a l’expansió i al predomini d’un petit nombre d’espècies poc exigents i, corresponentment, poc especialitzades. La flora i la fauna es «banalitzen» alhora que la productivitat del sistema creix.
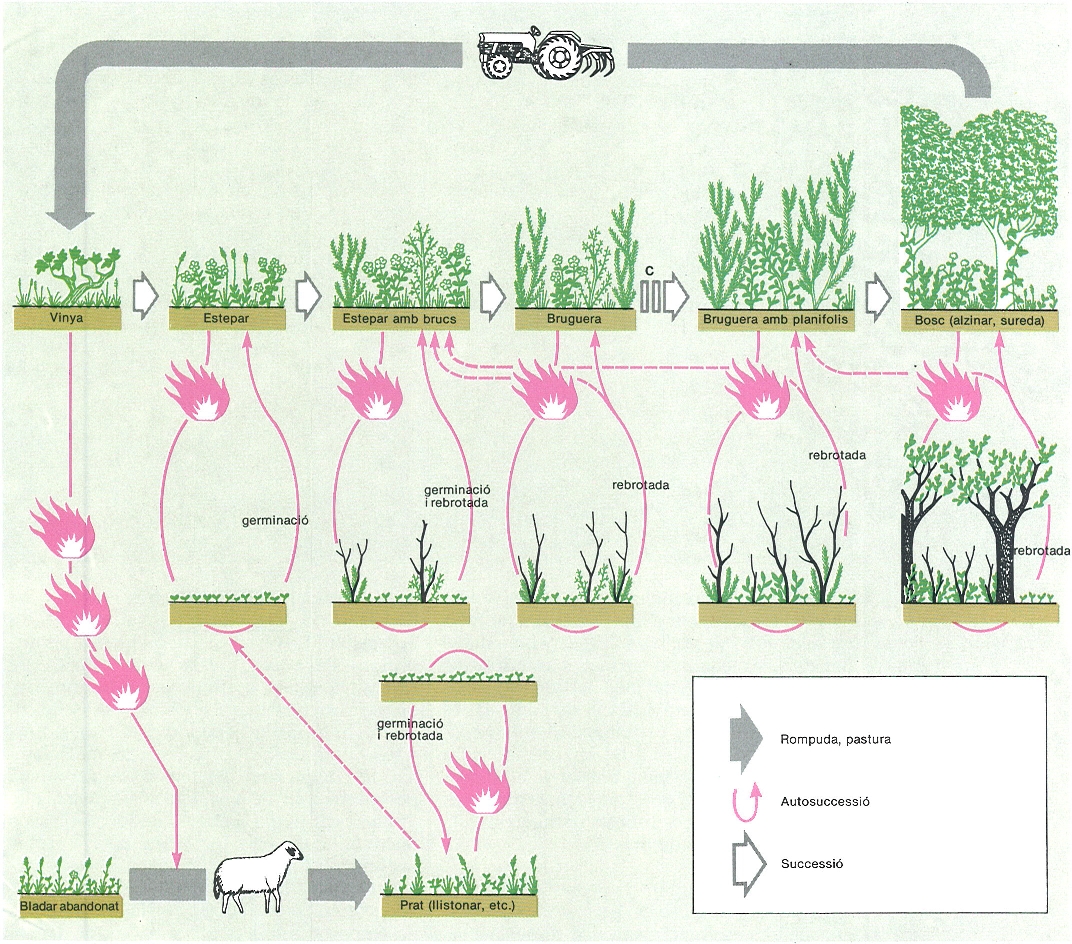
Model de successió i d’autosuccessió de la vegetació del cap de Creus (Alt Empordà). Si hom abandona la vinya (prèviament instal·lada, per exemple, en espai forestal), la successió instaura una bruguera i fins i tot un bosc, posat que hi hagi arbres llavorers a la rodalia: l’escassesa o la manca de llavors fa crític el pas C. Després del foc, per autosuccessió, sol restaurarse la mateixa comunitat cremada. La pastura reiterada manté la presència de prats (el foc de bosquines dóna brots tendres, però no pas herba pratense).
Mercè Cartañá, original de T. Franquesa.
L’explotació pot prendre la forma d’un increment de la freqüència o de la intensitat de les pertorbacions. La resposta del sistema dependrà de les seves característiques generals. Els ecosistemes més complicats, com la selva tropical o els esculls coral·lins (aparentment molt estables atès que no han sofert canvis en molts mil·lenis, ja que han arribat a interioritzar molts ritmes propis del medi i a adquirir, així, un gran poder d’anticipació i de control), es mostren, en canvi, extremament fràgils davant de pertorbacions fortes imprevisibles. La reconstrucció del sistema pot fer-se aleshores molt difícil, mentre que ecosistemes molt més senzills semblen més robusts perquè es reorganitzen molt més de pressa.
Els ecosistemes mediterranis presenten característiques no exclusives, però sí un xic peculiars en els seus mecanismes de resposta a les pertorbacions. Per exemple, després d’un incendi molts dels vegetals dominants rebroten amb gran força, de manera que, en pocs anys, s’aconsegueixen percentatges de cobertura del sòl similars als que precedien el foc. La composició de la vegetació pràcticament no ha variat en aquest procés. Per això, s’acostuma a parlar d’autosuccessió. Els ecosistemes d’aquesta mena són força estables, en un sentit més rigorós, gràcies al sistema de regeneració i a les reserves que les plantes són capaces d’emmagatzemar a llurs estructures de sosteniment.
Com a resultat de l’explotació i de les tendències autoorganitzatives dels ecosistemes, en un moment donat trobem simultàniament juxtaposades a l’espai una colla de situacions o fases d’evolució successional diferents. Això constitueix un mosaic complicat que pràcticament és sempre present als paisatges, fins i tot als més naturals, ja que sempre hi ha formes d’explotació d’una o altra mena, si bé l’escala de la pertorbació pot variar molt (als boscos plujosos tropicals hi ha moltes pertorbacions de petita escala, com caiguda de troncs, branques o fruits, però de gran freqüència, mentre que als boscos boreals de coníferes hi ha incendis periòdics cada dos-cents o tres-cents anys). Als ecosistemes mediterranis, la complexitat del mosaic és agreujada per l’antiguitat de l’explotació humana, que fa que cada petita parcel·la de territori, cada microunitat del paisatge, tingui la seva pròpia història d’ús. Aquesta «història» pesa especialment en l’estat del sòl i del banc de llavors, i condiciona fortament les possibilitats de resposta autoorganitzativa davant de noves pertorbacions.
La demografia en les plantes i els animals
Amb el pas del temps, els ecosistemes experimenten canvis en l’abundància relativa de les espècies i en el seu nombre (diversitat) i en la disposició dels organismes en l’espai. Aquests canvis es donen més de pressa en fases poc avançades de la successió i després s’alenteixen. A més, els ritmes també difereixen entre els diversos medis: els ecosistemes planctònics són molt més dinàmics que els terrestres. Però en tots els casos l’estructuració del sistema en els aspectes suara indicats és el resultat del flux d’energia que el travessa. I els elements bàsics en la relació entre organització de l’ecosistema i flux d’energia són l’estructura i funció dels individus, dit d’una altra manera, llur forma i disponibilitat d’energia.
Els organismes són unitats de matèria i estan separats entre ells. Aquesta afirmació no és trivial, ja que això ha fet possible l’evolució biològica. Dos organismes no són mai idèntics i hi ha una possibilitat no nul·la que les variacions d’un d’ells siguin transmeses a un nombre més gran de descendents que les d’un altre, segons les condicions del marc ambiental en què cadascun ha de viure. És a dir, les condicions ambientals són decisives en determinar el nombre de descendents. En aquest sentit, el sedentarisme de les plantes i la capacitat de moviment de la majoria d’animals són dues maneres molt diferents d’experimentar les condicions del medi. Mentre que els animals poden fugir de les condicions locals, les plantes no poden fer altra cosa que aguantar-les o morir-se. Per això, les plantes poden ser sotmeses a una varietat o un nombre de pressions selectives més gran que no pas els animals (més factors que en redueixen o modifiquen el creixement, la supervivència i la reproducció).
No obstant això, les plantes tenen algunes possibilitats de fugir. Una d’elles és la disseminació de propàguls lluny de la zona ocupada per la planta mare (en les plantes hi ha una gran varietat de mecanismes de disseminació, i, com a procés, és determinant per a l’estructura horitzontal —en el sòl— del sistema). Una segona possibilitat és en el mateix sistema de construcció del cos vegetatiu, i consisteix en l’acoblament seqüencial d’unitats bàsiques la mida de les quals és relativament poc variable dins de cada espècie. A més de les plantes, també mostren aquest sistema de construcció certs tipus d’animals (briozous, hidraris, coralls, etc.). Tots poden considerarse organismes modulars, en contraposició als organismes unitaris. Aquests, que són la majoria dels animals, són fets d’una sola peça diferenciada en òrgans des de molt aviat, que es desenvolupa un sol cop al llarg de tot el cicle biològic. Els modulars, per contra, creixen per formació repetitiva d’un cert nombre d’unitats bàsiques de construcció que hom pot anomenar metàmers. A les plantes, els metàmers són formats pel nus, la fulla (o fulles) i el meristema axil·lar, i l’entrenús (espai entre dos nusos consecutius). A més del metàmer, també es repeteixen durant el creixement altres unitats bàsiques de nivell superior: els mòduls, que són les agrupacions de metàmers per a formar, per exemple, una branca, una branqueta, un borró o, en general, tot allò que es produeix fins que el meristema apical atura el seu creixement (per accident o bé perquè forma una flor).
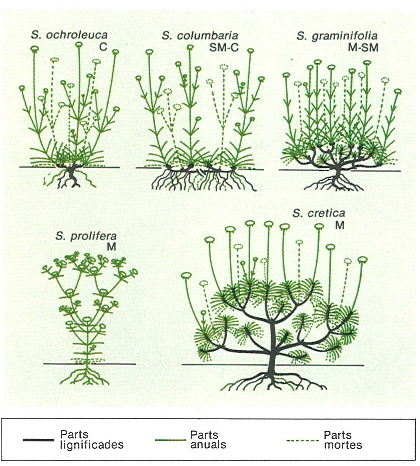
Formes de creixement en espècies d’Scabiosa de les àrees mediterrània (M), submediterrània (SM) i de l’Europa central (C). La forma de les plantes la configuren les branquetes renovables anualment i altres ja persistents. La mida que ateny la planta en un any la. determina el nombre de branquetes (mòduls) format, i d’aquest depèn el nombre de flors. És a dir, la descendència de la planta (llavors) és condicionada pel dinamisme de la seva morfologia.
Mercè Cartañá, original de W. Meusel.
El fenomen del metamerisme té propietats ecològiques interessants i ens pot ser útil per a apreciar les relacions entre estructura i funció de la vegetació i per a veure com l’organització de l’ecosistema és un efecte del flux d’energia. Considerant el cos vegetatiu de la planta com un conjunt de mòduls, es pot intuir que hi ha diverses maneres de disposar aquest cos en l’espai: en sentit preferentment vertical, en sentit preferentment horitzontal i segons la configuració unida o fragmentada del cos. Les espècies del primer grup tenen sempre una configuració unitària a l’espai; és el cas d’arbres i plantes anuals. Les del segon poden presentar una configuració unitària (amb una integració de funcions fisiològiques més o menys gran) o fragmentària, és a dir, dividida en parts genèticament idèntiques però funcionalment independents; és el cas de moltes plantes herbàcies perennes. Aquesta classificació correspon a les diferents formes ecològiques d’explorar l’espai, és a dir, de descobrir i explotar els recursos que conté. Així, un arbre que expandeix la seva capçada i creix en alçada s’apropia (i nega a altres) els recursos de llum que vénen de dalt i els de nutrients i aigua confinats pel seu sistema radical. Una gramínia cespitosa com el fenàs (Brachypodium phoenicoides) s’assegura els recursos de l’espai per a la seva expansió horitzontal produint mòduls (rizoma) amb metàmers curtíssims i fent tal densitat de tigetes aèries que pràcticament exclou tota altra planta del lloc o fa impossible el creixement de nous individus. Finalment, també és possible d’obtenir recursos horitzontalment amb la producció de mòduls (rizomes o estolons) construïts en metàmers llargs que, amb poca freqüència, es van bifurcant. Es tracta d’espècies en les quals predomina el creixement lineal en lloc del lateral, com el gram (Cynodon dactylon) o el trèvol (Trifolium).
Un aspecte interessant per a l’ecologia és la manera com es disposen els mòduls en l’espai o, si es vol, com s’exploren i exploten els recursos. La tàctica d’exploració de cada grup obeeix, d’una banda, a la programació genètica i, de l’altra, a l’oportunisme (plasticitat fenològica) de cada espècie. L’interès recent per aquests temes ha portat en particular a les regles d’ocupació de l’espai definides per Hallé i Oldemann per als arbres, però que també són vàlides per a les herbes. Aquestes regles són el resultat de considerar si el creixement és vertical o horitzontal, si és rítmic o continuat, si la floració és terminal o lateral, etc. En total s’han descrit 23 models de desenvolupament arquitectònic, és a dir, plans de desenvolupament sense que hi intervinguin accidents que lesionin els borrons. La forma que veiem en un moment donat constitueix l’arquitectura de la planta, fruit d’un procés d’acumulació de mòduls durant la seva història. El petit nombre de models, en comparació amb el gran nombre d’espècies existents, suggereix que alguns dels possibles representen síndromes adaptatives. Com a corol·lari, hom pot esperar una relació dèbil entre models i tipus de vegetació. A més, en la descripció estructural d’una comunitat cal tenir en compte la reiteració, o sia la possibilitat que es torni a reproduir el model a partir d’un lloc on s’hagin lesionat meristemes per una tempesta, un herbívor o qualsevol altra causa.
En conjunt, doncs, una comunitat vegetal té una estructura que resulta d’un creixement fins a cert punt planificat (segons un «estil» arquitectònic) i, en part, també improvisat per la contingència de cada moment. Però això és alimentat per un subministrament d’energia i canalitzat per l’arquitectura de cada individu. Quan en un bosc s’obre una clariana es donen noves oportunitats de capturar recursos. Les aprofitaran arbres joves que creixen verticalment segons el propi model i després es van ramificant amb un ritme que al final esdevindrà lent, això és, quan ja hauran estès una ampla capçada que condiciona gradients verticals d’humitat i de llum i sota la qual quedaran nous arbres joves dominats. Aquest procés de cicatrització no és sinó una alteració del nombre d’individus seguida, o no, per una substitució d’espècies amb concurrència de processos de disseminació, acció de depredadors, variació de la llum, etc. Veiem, doncs, que la persistència de l’ecosistema té lloc per mitjà d’un procés demogràfic (de naixements i morts, d’immigració i emigració), acompanyat d’una certa varietat d’estils d’estructuració per a cada component i mantingut per l’entrada d’un flux constant d’energia. Els punts de vista demogràfic i funcional són inseparables, i la relació entre ambdós s’ha de conèixer amb precisió per explicar que, per exemple, dues comunitats puguin produir la mateixa quantitat de matèria vegetal amb nombres d’espècies molt diferents, en uns casos amb unes poques espècies molt dominants i en altres amb una distribució més equitativa entre moltes espècies.
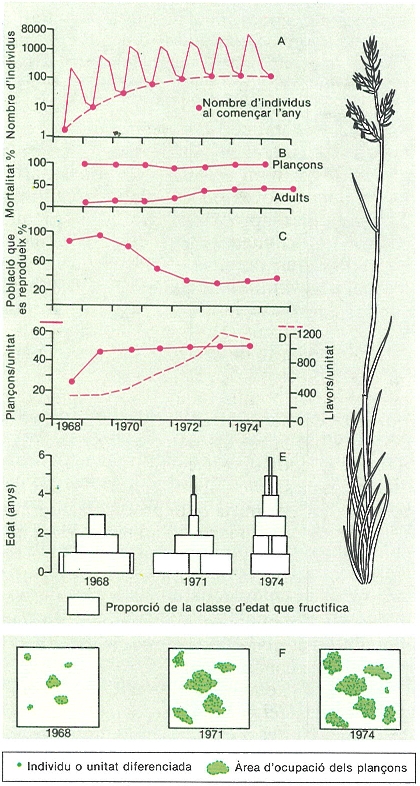
La demografia d’una població explica el procés del seu desenvolupament. Per això, cal conèixer quant canvia any darrere any (A), com afecta la mortalitat en joves i adults (B), quina és la fracció de població que es reprodueix cada any (C) i com varia la descendència per cada individu o unitat diferenciada (D). Tot això determina la composició d’edats de cada any (E) i, al seu torn, els canvis de l’ocupació i l’exploració de l’espai (F). L’exemple correspon a Corynephorus canescens.
Mercè Cartañá.
Per endinsar-nos en les relacions entre demografia i funcionalisme, convé esmentar alguns conceptes bàsics de la demografia. Els procediments que s’empren varien una mica segons que s’estudiïn organismes modulars o unitaris, però sempre es tracta d’explicar l’abundància d’individus genètics d’una espècie dins un ecosistema, com a resultat dels naixements, les morts, les immigracions i les emigracions que s’han produït en un lloc i en un temps determinats. En ecologia, interessa especialment un tipus d’estudi en què predomini un bon coneixement del cicle biològic de l’espècie (longevitat, moment de maduració sexual, periodicitat reproductiva, forma de creixement, etc.). Si els canvis d’abundància de les espècies (poblacions) els descrivim com meres diferències entre naixements i morts (cinemàtica), no sabrem gran cosa de les seves causes d’augment o disminució (dinàmica), ni dels canvis de distribució a l’espai (migracions). Per a conèixer també la dinàmica caldrà saber les causes individuals de mortalitat (per depredadors, paràsits, manca de recursos o desastres naturals) en un nombre de casos adient i tractable. D’aquesta manera sabrem, a més, el panorama aproximat de les forces selectives que incideixen en una població i podrem avançar en la previsió dels canvis futurs amb vista a intervenir en el seu control efectiu (cas de plagues) o conservació, o pensant en la biologia evolutiva de l’espècie.
La informació sobre les causes de mort s’obté seguint un col·lectiu d’individus des del naixement (en aquest sentit, és important d’assegurar el reconeixement de cada individu, i hi ha un repertori variadíssim de tècniques per a marcar els diferents tipus d’organismes). Els individus nascuts en un mateix període de temps constitueixen una cohort. El seguiment d’una cohort ens assegura una uniformitat de condicions. Es poden realitzar, després, censos periòdics (els censos són comptatges exhaustius) o estimacions mitjançant mostreigs (un altre tema sobre el qual hi ha una gran diversitat de tècniques). Així, podem calcular el nombre d’individus morts o supervivents d’un període a un altre. De tota manera, seguint una cohort no podem conèixer els individus que neixen en cada període i per això ens cal estudiar la fertilitat dels seus membres. Si combinem taxes de supervivència i fertilitat, es podrà saber la taxa mitjana de producció de descendents que esperem que tindrà un individu al llarg de la vida.
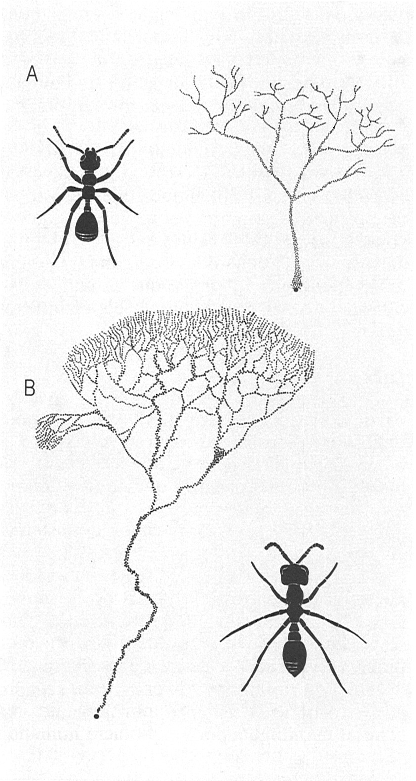
Explotació de l’espai per a cercar aliments en dos tipus de formigues: A Pheidole militicide, una formiga colonial; i B formigues guerreres que poden presentar sistemes lineals o ramificats, en aquest cas Eciton burchelli. Ambdues figures recorden la silueta d’una planta, és a dir, la pauta de ramificació que també empren les plantes per a obtenir llum i minerals.
Amadeu Blasco, a partir de fonts diverses.
Cada població real és constituïda per diverses cohorts més o menys sobreposades que en defineixen l’estructura d’edats. Aquesta estructura és modelada per les condicions ecològiques que han hagut de suportar les diferents cohorts, condicions que normalment van variant. El coneixement profund de l’estructura d’edats i de les corbes de supervivència i altres dades dinàmiques és indispensable per a la gestió de control o conservació de poblacions (des d’insectes plaga fins a grans vertebrats que hom vol protegir dins un parc natural.
Qualsevol població té un potencial de creixement pràcticament infinit. Si no el realitza és, òbviament perquè el medi sempre és limitat i no ofereix espai ni recursos més enllà d’un cert límit. Per a cada població hi ha, doncs, un límit màxim que el medi és capaç de mantenir, que s’anomena capacitat de càrrega del medi per a aquella espècie. A mesura que la població s’apropa a aquest màxim, augmenta la competència entre els individus de la mateixa població per a obtenir recursos, i el creixement es frena. En el cas de les plantes s’ha pogut establir una llei molt important: la densitat determina el nombre de mòduls que es construeixen, de manera que si la densitat és més gran s’arriba a una biomassa total similar, però a base d’individus més petits. Si la densitat de partida és molt alta (moltes llavors i molt juntes, per exemple) es produeix mortalitat o autoaclarida per causa de la competència. Si hom representa, en escales logarítmiques, la biomassa per individu enfront de la densitat, s’observa que els punts tendeixen a disposar-se sobre una línia de pendent aproximat –3/2 en tots els casos de plantes estudiats, sia herbes o arbres. En el procés de desenvolupament d’una massa vegetal, la relació entre biomassa individual i densitat segueix una trajectòria primer d’aproximació a l’esmentada línia i després al llarg de la mateixa. Aquesta relació ha rebut el nom de llei de Yoda i té moltes implicacions importants en ecologia terrestre, amb aplicacions en els àmbits agrícola i forestal.
Les adaptacions dels vegetals al medi
Hem parlat de les bases econòmiques que regulen l’assignació de recursos a les diferents parts de les plantes, sobreimposades a les pautes de creixement que són genèticament determinades. També ens hem referit a alguns aspectes molt generals de l’adaptació dels vegetals al medi terrestre. És hora d’endinsar-nos una mica més en aquestes adaptacions i de referir-nos a les condicions específiques del nostre país. Als Països Catalans trobem, en efecte, l’àmplia varietat de sòls i climes que ha estat descrita en altres parts d’aquesta obra. Cada combinació de condicions planteja una problemàtica particular per als productors primaris.
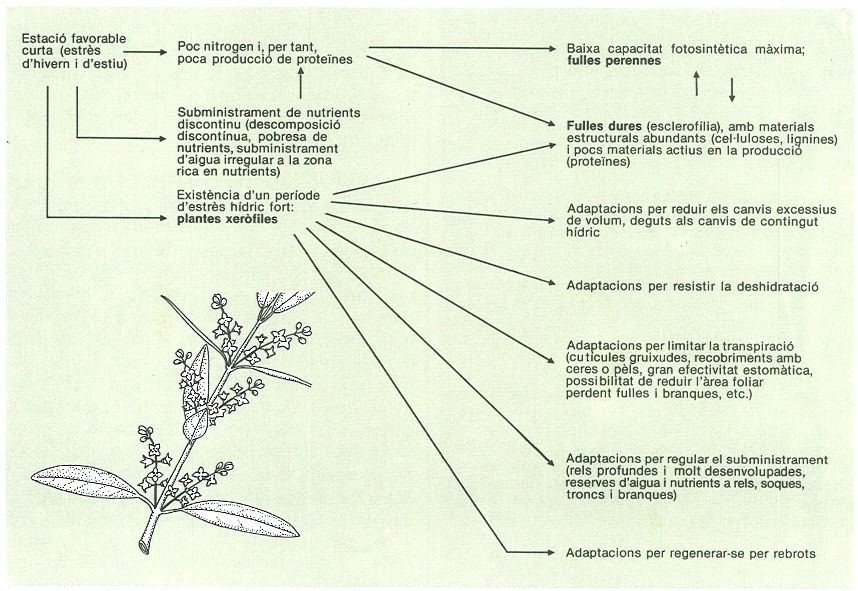
Respostes adaptatives dels vegetals dominants a la regió mediterrània septentrional a les condicions ambientals: fulles perennes, esclerofil·lisme, xerofília, rels molt desenvolupades. Algunes d’aquestes característiques podrien haver estat afavorides no sols per la importància de l’eixut estival, sinó també per l’acció dels herbívors i per una freqüència elevada d’incendis, com també per la pobresa de nutrients.
Carto-Tec / Mercè Cartañá.
La funcionalitat i l’estructura dels diferents ecosistemes reposa, sobretot, en les solucions morfològiques i fisiològiques que han trobat els productors primaris per a cada conjunt de condicions del medi. Sense entrar en el detall de totes i cadascuna de les comunitats descrites abans, sí que és convenient d’avançar en el coneixement dels complexos adaptatius més rellevants.
Atès que començarem per fixar-nos en aspectes estructurals i morfològics, ens convé de partir d’una classificació també fisiognòmica dels nostres ecosistemes terrestres més importants. Entre les diverses classificacions possibles, proposem de separar quatre grans grups: sistemes de caràcter zonal, és a dir, condicionats, sobretot, per factors climàtics; sistemes permanents, determinats, sobretot, per factors relacionats amb el substrat i amb fenòmens locals de transport horitzontal intens; sistemes resultants de la degradació de qualsevol dels anteriors, en estats transitoris o de difícil recuperació a causa de la degradació del sòl, determinats per diversos tipus i règims de pertorbació; i sistemes antropogènics, en general mantinguts per l’explotació humana i per considerables entrades d’energia externa suplementària.
En les pàgines següents tractarem aspectes de l’estructura i el funcionament d’aquests diferents tipus d’ecosistemes. Però ara, com ja hem anunciat, ens interessa posar de manifest la relació entre les plantes i llurs medis, per interpretar la morfologia i la fisiologia en termes d’adaptacions i transposar el resultat d’aquestes interpretacions a nivell de l’ecosistema.
Tipus d’ecosistemes terrestres dels Països Catalans, en una classificació molt general. Els sistemes zonals són els que corresponen a l’estadi màxim de maduresa, en equilibri amb unes condicions macroclimàtiques; els azonals permanents són els de medis molt especials, que imposen llurs característiques per damunt de les del clima general; els transitoris són els que resulten de la degradació de tots els anteriors; i els antropogènics són els edificats per l’home. No usem ací estrictament aquesta classificació perquè no ens cal introduir hipòtesis successionals, però emprant criteris fisionòmico-funcionals s’arriba a unitats semblants.
Tipus d’ecosistemes terrestres dels Països Catalans| SISTEMES ZONALS | Ecosistemes de la zona alpina |
| Boscos subalpins i montans aciculifolis de coníferes | |
| Boscos caducifolis d’hivern | |
| Boscos perennifolis mediterranis, de fulla plana o aciculifolis | |
| Màquies i espinars mediterranis litorals i continentals | |
| SISTEMES AZONALS PERMANENTS | Boscos de ribera |
| Ecosistemes dels sòls arenosos litorals | |
| Ecosistemes de sòls salins i guixencs | |
| Ecosistemes d’aiguamolls | |
| Ecosistemes de roques i tarteres | |
| Ecosistemes de zones culminals | |
| SISTEMES TRANSITORIS resultants de la degradació de qualsevol dels anteriors | Landes, matolls, garrigues, brolles, timonedes, bardisses i altres ecosistemes no zonals, dominats per plantes arbustives |
| Prats i pradells | |
| Ecosistemes d’ambients ruderals i arvenses | |
| SISTEMES ANTROPOGÈNICS | Repoblacions i plantacions |
| Agroecosistemes de secà | |
| Agroecosistemes de regadiu | |
| Ecosistemes urbans | |
| Dades elaborades per l’autor | |
Tipus biològics
La morfologia de cada planta constitueix una resposta evolutiva específica al marc ambiental que ha trobat al llarg de la seva vida. Però aquest marc ambiental actua amb pressions selectives prou fortes per conduir l’evolució d’espècies diferents cap a resultats força similars. Es produeixen, així, els anomenats fenòmens de convergència adaptativa: espècies, de vegades sense cap parentiu filogenètic proper, presenten morfologies (i fisiologies) força semblants davant d’ambients també semblants. Això ha despertat des de fa temps l’interès dels estudiosos i s’han proposat diversos sistemes de classificació dels vegetals no ja segons criteris taxonòmics sinó segons similituds d’estructura i de formes de vida. La classificació més popularitzada en els textos de botànica és, sens dubte, la ideada per Raunkjaer. Nosaltres presentarem ací, lleugerament adaptat, el sistema proposat recentment per Box, que creiem que es basa en una gamma més matisada de caràcters.
Començarem per considerar els caràcters morfològics i fisiològics més importants dels vegetals que determinen llur economia hídrica i energètica, per passar, després, a una tipologia de formes biològiques sobre la base d’aquests caràcters. Per simplificar, tractarem els caràcters en els següents grups: els que fan referència a la forma general de la planta (tipus estructural i mida); els relatius a les fulles (tipus de fulla, mida i estructura); i els relatius als ritmes d’activitat, definits per la distribució del període de foliació.
Tipus estructurals
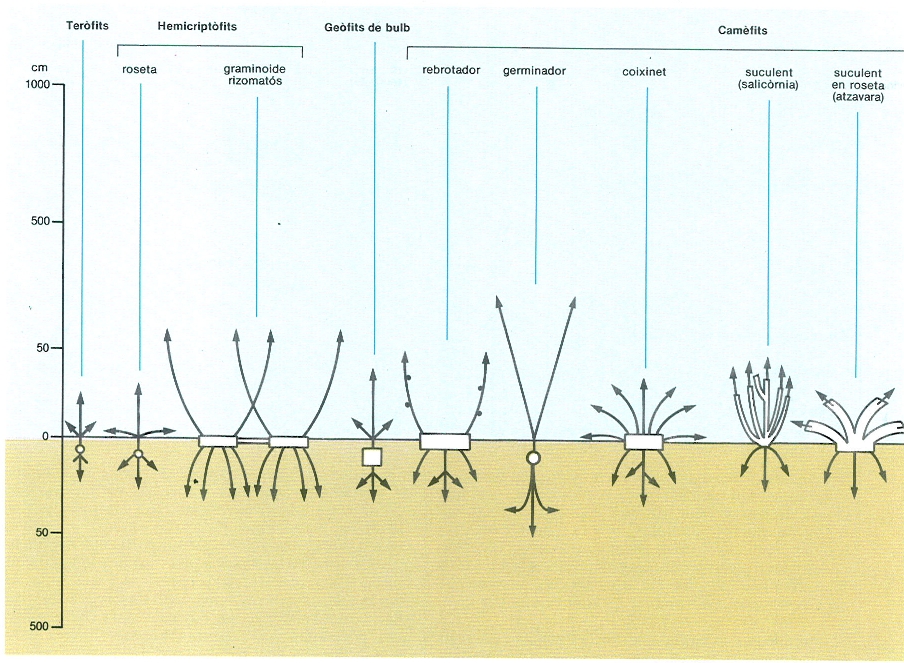
Formes de vida de vegetals superiors, considerant només l’ocupació de l’espai (fletxes) i les principals zones de magatzem de reserves (en blanc).
Mercè Cartañá, original de l’autor.
El tipus estructural d’una planta és definit, en part, per l’alçada, per la pauta de ramificació i pel fet de ser llenyosa, herbàcia o suculenta. Cada caràcter dels esmentats té fortes implicacions en l’estratègia de regulació de l’economia de recursos per part de la planta. Als Països Catalans podem trobar exemples de força tipus estructurals diferents: arbres; arbrets nans; arbustos en coixinet; arbustos normals (semblants als arbres, però amb ramificació des de terra); arbustos de fulles en roseta (com el margalló); plantes de tija suculenta; graminoides (herbes de fulles llargues i estretes que creixen per tiges laterals, com el fenàs, o formant feixos atapeïts de grans dimensions, com el càrritx); fòrbies (totes les herbes que no són graminoides, afil·les o falgueres); lianes (plantes enfiladisses, amb tiges de ràpida elongació, que pugen cap a la llum arrapant-se a altres plantes, com l’heura, les clemàtides o l’arítjol); falgueres herbàcies (de vegades poiquilohídriques); epífits (creixen damunt d’altres plantes, però continuen essent capaços de fotosintetitzar); i tallòfits (molses, hepàtiques, líquens, algues).
Hi ha, sens dubte, una relació entre el tipus estructural i la disponibilitat d’aigua, que es posa de manifest en la variació estructural que és observable al llarg dels gradients d’ariditat que es troben en el conjunt del planeta i, de N a S, en el territori que ací ens interessa. No obstant això, no solament és important la quantitat de precipitació, sinó també la duració i regularitat de les estacions plujoses, la permeabilitat més o menys gran del sòl, l’existència d’un nivell freàtic més o menys fàcilment accessible, les temperatures estivals a les estacions de clima fred, etc., de manera que no es pot establir una relació senzilla entre precipitacions i tipus estructural.
Hem d’introduir també un criteri de dimensions, no per diferenciar tipus estructurals sinó perquè per a cadascun hom pot reconèixer diverses categories de mida; i és convenient de fer-ho, ja que l’alçada de la planta reflecteix les condicions ambientals d’humitat, temperatura i nutrients.
A continuació, la taula presenta la llista de tipus biològics de les plantes dels Països Catalans i exemples principals, segons una classificació basada en la d’E.O. Box.
Tipus biològics de les plantes dels Països Catalans
| TIPUS BIOLÒGICS | EXEMPLES MÉS SIGNIFICATIUS | ||
|---|---|---|---|
| ARBRES SEMPRE VERDS DE FULLA AMPLA | |||
| Arbres planifolis de llocs càlids | Cítrics | ||
| Arbres planifolis mediterranis | Alzina, surera, ullastre, garrofer | ||
| ARBRES DE FULLA CADUCA (planifolis de fulla verda a l’estiu) | |||
| Mesofil·les de la zona temperada | Roures, aurons, faig, freixes, oms, til·lers, verns, castanyers, pollancres, salzes, moixeres, noguera, lledoner | ||
| Microfil·les d’estiu fresc | Bedoll, salzes | ||
| ARBRES DE FULLA ESTRETA O EN AGULLA | |||
| Sempre verds | De zones mediterrànies | Pi pinyer, pi blanc, xiprer | |
| De zones temperades | Pinassa, pi roig, teix | ||
| De zones de clima boreal | Avet, pi negre | ||
| Caducifolis de climes boreals | Làrix | ||
| ARBRES BAIXOS O NANS | |||
| Planifolis sempre verds de zones mediterrànies o temperades | Grèvol, llorer, fals aladern, aladern | ||
| Planifolis caducifolis d’hivern | Pruneres i afins, nesprer, arç blanc, sanguinyol, avellaner, pereres i afins | ||
| De fulla estreta | Savines i ginebres, tamarius | ||
| ARBUSTS | |||
| Planifolis sempre verds de zona temperada | Mediterranis | Garric, murta, baladre, estepa-joana, matapoll, Olivella, matabou, aladern fals, marfull, argelaga, aladern, cirerer d’arboç | |
| De zones temperades | Grèvol, boix, olivereta, lloreret | ||
| De zones d’hivern fred | Neret | ||
| Planifolis caducifolis d’hivern | Rosers, corner, riber, cotoneàster, aranyoner, noguerola, roldor, vinya, espinavessa, tortellatge, lligabosc | ||
| Perennifolis que treuen fulles després de períodes plujosos | Coralet | ||
| Perennifolis de desert càlid | Zigofil·le | ||
| Xeromòrfics | D’hivern fred | Salats, barrella | |
| D’hivern temperat | Bufalaga, estepes, romaní, estepa d’arenal | ||
| Aciculifolis sempre verds | Savines, brucs | ||
| Caducifolis d’estiu | Bufera arbustiva, arç de tanques, alfals arbori | ||
| De fulla suculenta | Salats, barrella, gavó | ||
| Afil·les o amb molt poques fulles | Amb tiges verdes fotosintetitzadores | Ginesta, gòdua, èfedra | |
| Amb tiges transformades, de morfologia foliar | Fil·locladis grans | Galzeran | |
| Fil·locladis petits perennes o caducs | Esparregueres | ||
| Arbusts nans o rastrers | Perennifolis mediterranis | Farigoles, lavandes, sàlvies, heliantems, estepa blanera, timons | |
| Perennifolis de zones temperades o d’hivern fred | Ericàcies de landes, boscos o zones alpines | ||
| Perennifolis de zones xèriques | Botja pudent, camforada | ||
| Caducifolis d’hivern de la tundra alpina | Salzes nans, nabiu | ||
| De fulla estreta, de muntanya | Savina de muntanya, ginebró | ||
| Coixinets | De muntanya, sense punxes | Arenària, dríade | |
| Xeroacàntics de muntanya | Botgeta, eriçó, eriçó groc | ||
| Socarrells de zones litorals i de muntanya balear | Eixorba-rates blanc i negre, eriçó | ||
| Arbusts de fulles en roseta | Margalló, atzavara | ||
| Arbusts de tija suculenta ramificada | Sense fulles o amb molt poques, suculència per aigua | Figuera de moro, cirialera, salat | |
| Amb fulles, suculència per làtex | Lleteresa arbòria | ||
| GRAMINOIDES | |||
| Canyes altes | Canya, canyís | ||
| Graminoides en motes | Albellatge, gesp, càrritx, càrex pèndul | ||
| Helofítiques que no formen canya | Mansega, jonc, castanyola | ||
| Graminoides curtes cespitoses | Gram, poa, llistó, pelaguer, agropir, gesp | ||
| Esclerofil·les | Cutàndia, espart, espart bord | ||
| Afil·les, amb tiges verdes | Eleocaris, jonc | ||
| FÒRBIES | |||
| Sense fulles a l’hivern | Herbes forestals, sobretot hemicriptòfits i geòfits, com moltes liliàcies, compostes, lleguminoses, ranunculàcies, umbel·líferes | ||
| Perennifoliades o semiperennifoliades de zones temperades | Rèvola, plantatge | ||
| De fulles suculentes | Crespinells, verdolaga, matafoc, barretets | ||
| Macrofòrbies o megafòrbies | Plantes sobretot d’ambients ruderals o de sòls enriquits i de força humitat, com moltes umbel·líferes, quenopodiàcies, amarantàcies, urticàcies, compostes, crucíferes, papaveràcies | ||
| Petites herbes en coixins xèrics | Saxífraga, draba, cerastis | ||
| Efímeres de desert sec | Diverses anuals, geòfits nans i graminoides | ||
| Perennifoliades de desert fred | Diversos geòfits nans i graminoides | ||
| Caducifoliades de desert fred | Cotonera, càrexs, gèum, genciana, ranuncles, saxífragues | ||
| LIANES | |||
| Planifòlies sempre verdes | Lligabosc valencià i mediterrani, arítjol | ||
| Planifòlies verdes a l’estiu | Vinya, vidalba, lligabosc etrusc i atlàntic | ||
| FALGUERES | |||
| Sempre verdes | Polypodium, Asplenium, Cheilanthes, Pellaea, Polystichum | ||
| Verdes a l’estiu | Cystopteris, Athyrium | ||
| EPÍFITS | |||
| Tal·lòfits | Líquens, molses | ||
| Vasculars | Vesc | ||
| MOLSES I LÍQUENS NO EPÍFITS | |||
| En catifa | Molses i hepàtiques dels boscos i tundra, líquens foliacis | ||
| Xèrics | Líquens crustacis, algunes molses | ||
| Dades elaborades per l’autor | |||
Tipus de fulles
Són molts els caràcters foliars que tenen un fort significat adaptatiu, des del color i la presència, o no, de pèls (que es relacionen, per exemple, amb la quantitat de radiació que les fulles poden reflectir, i, per tant, amb el balanç tèrmic de la fulla) fins a la mida i la forma més o menys retallada (que tenen a veure amb la circulació laminar o turbulenta de l’aire al voltant de la fulla, i, per tant, amb l’intercanvi gasós, la regulació de la transpiració i la pèrdua de calor). Entremig d’aquests encara trobaríem les innombrables complicacions que es deriven de la forma, disposició i abundància dels estomes, les característiques de l’epidermis, etc. En línies generals, és ben conegut que les plantes de països àrids tenen habitualment les fulles petites, gruixudes i coriàcies, mentre que les de països o llocs humits solen presentar-les més grans, primes i blanes. Establim-ne, però, algunes categories.
Pel que fa a la morfologia foliar, considerarem, seguint encara Box, quatre possibilitats: fulles amples, estretes, graminoides o absents. Per fulles estretes, en aquest sentit, entendrem només les acícules dels pins o dels brucs, les fulles esquamiformes dels xiprers i les savines o les fulles linears planes dels avets i els teixos; es tracta de fulles especialment resistents a la dessecació i a condicions extremes de calor i de fred. Podem diferenciar, entre les fulles amples, les que són més o menys enteres i les que presenten el limbe fortament dissecte. En el cas de les fulles compostes és més pràctic de considerar cada folíol com una fulla diferent, almenys en el context que ací ens interessa, per exemple a l’hora de mesurar-lo.
Si introduïm un criteri de mida de la fulla, de cada tipus foliar, en podem fer també diverses classes. Alguns autors parlen de fulles subleptofil·les (menys de 0,10 cm2 de superfície), leptofil·les (0,10-0,25 cm2), nanofil·les (0,25-2 cm2), (nano-microfil·les (2-12 cm2), microfil·les (12-20 cm2), micromesofil·les (20-56 cm), mesofil·les (56-180 cm2), macrofil·les (180-1640 cm2) i megafil·les (més de 1640 cm2). Box usa una classificació menys detallada (nano, micro, meso i macrofil·les). Les mides foliars es relacionen amb la regulació de l’intercanvi energètic i amb l’eficiència en l’ús de l’aigua (fulles massa grans poden tenir rescalfaments excessius si reben radiació directa). Als llocs freds, també exposats de vegades a altes radiacions, les fulles grans poden tenir avantatge perquè s’escalfen més que l’aire i això permet taxes superiors de fotosíntesi, però també implica més consum d’aigua, cosa que no sempre és possible. S’han proposat models força avançats que permeten de fixar les dimensions òptimes de les fulles en un ambient físic determinat. En general, l’única conclusió ben establerta és la que assenyala l’avantatge de les fulles petites en ambients de forta radiació, altes tempertures i manca d’aigua.
Finalment, l’estructura de la fulla és definida per la presència, més o menys important, de teixits esclerenquimàtics, pel grau de suculència i per la morfologia dels estomes. Podem considerar sis categories bàsiques: plantes malacofil·les (de fulles fines i toves); plantes de fulles coriàcies (de cutícules engruixides, però encara flexibles); plantes esclerofil·les (dures, però també fràgils si són doblegades); plantes suculentes (amb teixits emmagatzemadors d’aigua); plantes de fulles pubescents (toves i un xic carnoses, cobertes de pèls); i plantes afil·les o amb poques fulles, però amb tiges fotosintetitzadores. Totes aquestes diferents estructures es corresponen amb diferents funcionaments. En general, les fulles més toves són més actives, tenen taxes fotosintètiques més altes i transpiren més, però regulen poc aquesta transpiració. Les fulles perennes solen ser coriàcies o esclerificades amb taxes més baixes d’intercanvi. Les plantes suculentes i les de tiges verdes poden tenir transpiració pràcticament nul·la durant les èpoques d’eixut, però, en canvi, funcionen amb una taxa anual de fotosíntesi baixa. Els estudis ecofisiològics, que mesuren l’intercanvi gasós i els factors microclimàtics per mitjà de cambres d’aire controlat a nivell de les fulles o de petits branquillons, han donat molta informació sobre el caràcter adaptatiu de les diferents estructures foliars.
Ritmes fotosintètics estacionals i durada de la fulla
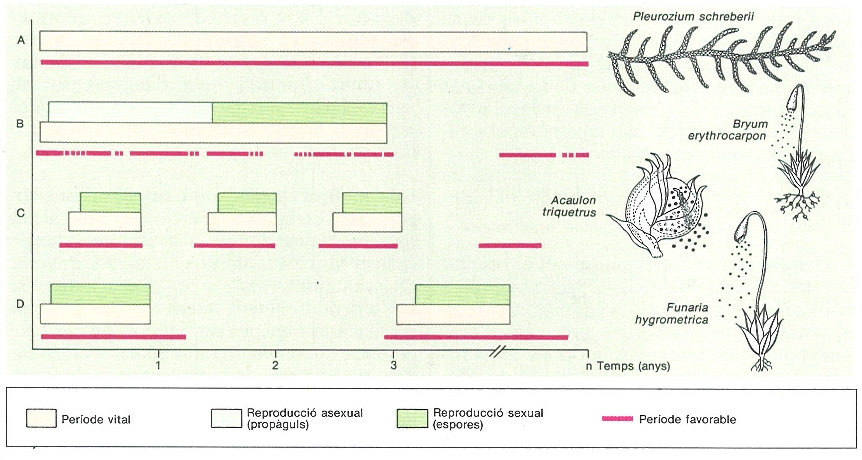
Estratègies de vida en briòfits. En les espècies perennes (A) són característics els ambients favorables continus, les plantes grans i una escassa presència d’estructures reproductores especialitzades (sexuals o asexuals). En espècies colonitzadores (B): període favorable discontinu però llarg, mida petita, resistència dels gametòfits als períodes desfavorables i gran inversió en estructures reproductores (espores o qualsevol mena de propàgul). En espècies itinerants (C): període favorable curt, però que es repeteix periòdicament al mateix lloc; la durada de la vida s’ajusta a la del període favorable; inversió important de recursos en estructures reproductores grans, força resistents i amb poca capacitat de dispersió. En espècies fugisseres (D): període favorable relativament imprevisible; durada de vida curta i gran producció de diàspores petites amb molta capacitat de dispersió.
Mercè Cartanà, original de F. Lloret.
El temps que la fulla roman activa i el període de l’any en què té lloc aquesta activitat poden variar i, d’acord amb això, és possible de diferenciar entre: plantes sempre verdes, com l’alzina, l’arboç o els pins; plantes que produeixen fulles a l’estació plujosa i les perden a l’eixuta (com és propi dels boscos caducifolis tropicals) i caducifolis d’estiu (entre els quals, s’hi compten alguns arbustos de les nostres terres meridionals); plantes verdes a l’estiu que perden la fulla a la tardor, anomenades caducifolis d’hivern, com el faig o els pollancres; plantes sufruticoses (llenyoses només a la base), que perden les tiges herbàcies o una mica lignificades en acabar l’estació de creixement, tot i conservar la base llenyosa; plantes de fulles marcescents, que mantenen les tiges o les fulles mortes durant l’estació desfavorable (com fan alguns roures, especialment el de fulla petita); plantes semiperennifoliades, que poden perdre facultativament totes o part de les fulles en avançar l’estació desfavorable (com fa l’arçot); i plantes efímeres, de cicle de vida anual (teròfits) o bé perenne (geòfits), amb tiges herbàcies que desapareixen en els períodes desfavorables.
La durada de la fulla és el resultat d’una selecció i es relaciona amb altres conjunts d’adaptacions. Una fulla amb baixa capacitat fisiològica tindrà més ingressos de carboni si dura més temps i pot aprofitar tots els moments favorables al llarg de l’any, i una capçada amb fulles de diferents edats reforçarà aquest avantatge. L’ús dels nutrients serà més eficient ja que, com que la fulla dura més, es gasta menys nutrient en construir-la per unitat de pes sec que la fulla produirà. L’esclerofil·lisme pot ser necessari per reduir l’herbivorisme i el rentatge de nutrients, com també el cost de construcció. En contrapartida, les fulles esclerofil·lès tenen baixa taxa fotosintètica màxima. Les coníferes tenen avantatge en climes subàrids i boreals, com a resultat de l’arquitectura vascular (les traqueides de petit diàmetre no s’omplen d’aire ni sota l’estrès hídric ni a baixes temperatures, però això limita la taxa fotosintètica a valors baixos, i fa necessari que les fulles siguin de vida llarga). Els ritmes d’activitat es relacionen amb els caràcters d’estructura foliar. Per exemple, l’esclerofil·lisme i, sovint, el microfil·lisme solen acompanyar el funcionament perennifoliat. El més important, però, és que els diferents ritmes d’activitat van associats a diferències fisiològiques importants.
La temperatura i les plantes
Les plantes són organismes incapaços de regular directament llur pròpia temperatura i de mantenir-la dins un interval molt constant. Això, com sabem, només ho fan els mamífers i els ocells. Malgrat tot, no s’ha de caure en l’error de creure que la temperatura d’una planta serà sempre igual a la de l’aire que l’envolta; ben al contrari, poden haver-hi diferències importants que depenen de la morfologia, l’anatomia i la fisiologia particular de cada espècie. Per tant, la temperatura que cada planta experimenta realment i la seva capacitat per a suportar-la es relacionen amb un conjunt complicat d’adaptacions de tota mena.
La temperatura d’un organisme és el resultat del seu equilibri energètic. L’organisme rep o perd calor per convecció o per conducció cap al seu medi; en perd en els processos d’evaporació d’aigua que tenen lloc a les seves superfícies (calor latent d’evaporació), per l’emissió d’energia d’ona llarga (com fan tots els cossos, en quantitat proporcional a la quarta potència de la seva temperatura absoluta); i, finalment, en guanya per l’alliberament de calor que té lloc en els processos metabòlics propis i sobretot per la radiació solar que incideix al seu damunt, directament o reflectida des d’objectes veïns. Si la suma d’entrades és més gran que la de sortides, la temperatura del cos puja i, en canvi, si és més petita, disminueix.
Les entrades de radiació depenen de l’absorció que en fa l’organisme o la part que considerem, ja que una proporció variable és reflectida o transmesa. Determinats caràcters de les plantes modifiquen la proporció que és absorbida. Per exemple, una forta tomentositat, amb pèls blancs, crea unes superfícies platejades que reflecteixen una part important de la radiació rebuda, la qual cosa serà útil en certs ambients per a evitar un escalfament massa fort i mantenir la temperatura prop de l’òptim per a la fotosíntesi. Les fulles que es mouen durant el dia, modificant llurs angles d’exposició a la radiació incident i oferint en els moments de radiació més forta una superfície més petita, poden trobar també avantatges. Durant la nit, per a reduir les pèrdues per radiació emesa, algunes plantes poden plegar les fulles i així disminueixen la superfície d’intercanvi.
Una influència molt forta en el balanç energètic és la de la transpiració, ja que es calcula que, normalment, fins la meitat de l’energia radiant absorbida no fa sinó evaporar aigua. Quan fa molta calor, la fulla pot mantenir-se a temperatures molt inferiors a les de l’aire gràcies a la transpiració, mentre que, a temperatures normals, la radiació solar escalfa la fulla per damunt de la temperatura de l’aire. No obstant això, s’ha pogut veure que les fulles presenten, pel que fa a la transpiració, comportaments molt diferents segons llurs mides.
A les fulles petites les temperatures mai no pugen tant, sota una radiació intensa, com a les fulles grans. En aquestes, i precisament per llur mida, es forma a la superfície una capa d’aire relativament quiet (capa laminar) a través del qual el vapor d’aigua circula només per difusió, de forma lenta; a les fulles petites, en canvi, tot l’aire que les envolta és sotmès a un règim turbulent, no hi ha capa laminar i el vapor d’aigua circula amb més facilitat permetent la refrigeració. Per mantenir una mateixa temperatura, les fulles petites gasten menys aigua que les grans i, si la radiació és molt forta, a les petites els és més difícil de tenir temperatures perjudicials. De tota manera, la regulació tèrmica ha d’arribar a un compromís amb l’economia de l’aigua: no sempre es pot evaporar tota l’aigua necessària per a regular la temperatura, perquè en manca. Hi ha plantes que, en lloc de transpirar més, han desenvolupat mecanismes de resistència a les altes temperatures.
La dissipació de calor per conducció o convecció des de la fulla és un aspecte que pot ser modificat per l’estructura i la morfologia foliar, i no solament per la mida. La formació d’una capa laminar, a igual superfície, és molt més fàcil en fulles arrodonides que en fulles molt allargades o amb lòbuls profunds.
La resistència que s’obté a temperatures massa altes o massa baixes en les diferents fases i parts d’una planta també és el resultat de moltes adaptacions de caràcter fisiològic i de comportament (la fenologia de les plantes constitueix una resposta a l’estacionalitat sobretot tèrmica del clima), que no podem analitzar ací. Però sí que voldríem deturar-nos en algun aspecte concret que té especial importància.
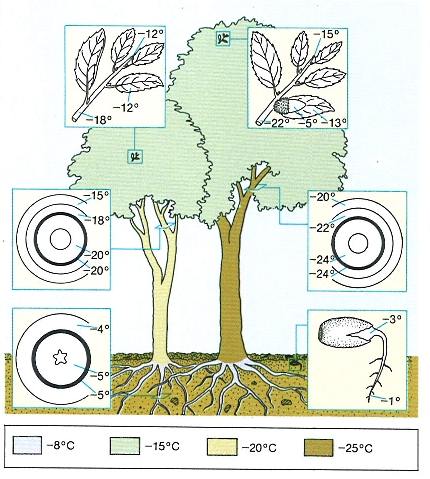
Resistència al fred de l’alzina (Quercus ilex) i temperatures a partir de les quals les gelades afecten les diferents parts del bosc.
Mercè Cartañá, amb dades de W. Larcher.
La resistència al fred i a la congelació és un caràcter que no solament varia d’una espècie a l’altra, sinó que també ho fa en les diferents fases de la vida de cada espècie: les llavors encara seques aguanten temperatures molt més baixes que les llavors ja impregnades d’aigua i que les de les plantes en creixement, i això les converteix en formes de resistència; al llarg de l’any normalment les plantes s’endureixen progressivament amb el fred i tenen una resistència màxima quan arriba l’hivern; els continguts hídrics elevats afecten negativament la resistència; els borrons en repòs són molt més resistents que les fulles en expansió; les esquames que els protegeixen sembla que poden reduir la radiació emesa durant la nit, i ho fan millor si són negres (com les del freixe); etc. L’enduriment dels teixits amb el fred al llarg de l’any es relaciona amb canvis químics en la composició dels sucs vacuolars. A mesura que la temperatura baixa d’un dia per l’altre, les plantes acumulen soluts als vacúols, la qual cosa en determina un descens fins al punt de congelació. Aquest descens no és, de tota manera, gaire important i calen altres mecanismes; per exemple, algunes plantes no deixen que es formi gel al seu interior perquè els cristalls de gel queden aïllats, en certa manera, del contingut vacuolar. S’ha dit que el marciment de les fulles de neret és un d’aquests mecanismes i sembla que el boix també limita amb èxit la formació de cristalls de gel als espais intertissulars. La conversió dels continguts cel·lulars en formes gelificades fa el protoplasma més resistent als canvis de volum i a la deshidratació, alhora que en dificulta la congelació. Sembla que els danys per fred i congelació afecten sobretot la membrana cel·lular, en part per una acció sobre la configuració de les proteïnes. Aleshores, certes plantes, en virtut d’una peculiar composició química (menys grups SH— a les molècules de proteïna), resultarien més resistents a aquests canvis.
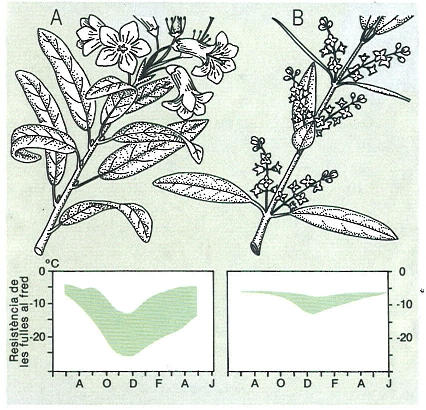
Evolució estacional de la resistència al fred en dues espècies d’ambients molt diferents: A el neret (Rhododendron ferrugineum), muntanyenc, i B l’ullastre (Olea europaea), mediterrani. Les àrees acolorides estan limitades, per dalt, per la resistència mínima, quan l’individu ha perdut l’aclimatació després de diversos dies en un lloc càlid, i, per baix, per la resistència màxima, quan l’individu ha estat aclimatat progressivament. La resistència real de cada instant queda en algun lloc a l’interior de la zona. Hi ha una variació estacional considerable en els límits superior i inferior, i molta diferència en el comportament d’ambdues espècies.
Mercè Cartañá, original de W. Larcher.
Pel que fa a les temperatures massa altes, també s’observen diferències en la capacitat de resistència d’unes plantes o altres i en una mateixa espècie segons les estacions. Molts xeròfits són heliòfils i termòfils, però els mecanismes precisos que determinen aquests comportaments són mal coneguts. Les plantes resistents tenen el punt de compensació (temperatura en què respiració i fotosíntesi tenen lloc a igual velocitat) molt més elevat. Al llarg de l’any, també hi ha una aclimatació que afavoreix que la màxima capacitat de resistència es doni a l’estiu. La fotosíntesi es fa en condicions òptimes a temperatures diferents, segons les espècies, però la respiració sempre augmenta més que la fotosíntesi en augmentar la temperatura. Per això, la producció primària neta té sempre l’òptim per sota de la temperatura que dóna la màxima taxa de fotosíntesi instantània. Les plantes C3 tenen els valors màxims de producció neta entre els 10°C i els 30°C de temperatura diürna, mentre que les C4 els tenen per sobre dels 30°C i algunes s’acosten als 50°C; les plantes CAM depenen més de les temperatures nocturnes. Alguns vegetals, com els líquens de les zones alpines i de la tundra, tenen òptims inferiors als 10°C. Per sota dels —5°C la majoria dels arbres són incapaços de fixar el diòxid de carboni, mentre que els líquens esmentats poden continuar fent-ho fins per sota dels — 15°C, i les C4 no baixen de 5°C.
La sequedat i les plantes: xerofitisme
Aquesta denominació s’aplica bé als trets adaptatius d’un contingent molt ample de la nostra flora, marcada per la necessitat de suportar períodes d’eixut relativament prolongats. Caracteritza els xeròfits el fet de mantenir vives les parts aèries (almenys tiges i branques), ni que sigui en estat d’activitat restringida, i no presentin magatzems d’aigua. Podem distingir-ne diverses menes, amb conjunts d’adaptacions tan diferents que fan de mal reunir en el mateix tipus general. En rigor, són pocs els caràcters comuns entre uns xeròfits i els altres, fora de respondre abans que res a un estrès hídric, però mirarem de considerar-ne diverses categories.

Els xeròfits malacofil·les són abundants entre els matolls mediterranis heliòfils. Les fulles toves, sovint amb una densa pubescència, poden variar força llur contingut hídric i, quan l’eixut es prolonga, poden caure en una proporció més o menys gran, amb la qual cosa es redueixen l’àrea foliar i la transpiració. Aquestes plantes són hidrolàbils, pel fet que tenen un sistema radical poc desenvolupat. A la fotografia veiem fulles d’estepa blanca (Cistus albidus).
Fototeca / MC
Els xeròfits malacofil·les presenten fulles toves i quasi sempre cobertes de pèls. La quantitat i les dimensions d’aquestes fulles solen variar al llarg de l’any, de forma que a l’estiu acostumen a ser més petites i menys nombroses, la qual cosa comporta, és clar, una reducció de l’àrea foliar total; la botja pudent (Artemisia herba-alba) perd la major part de les fulles inferiors, més grosses, i l’arçot (Rhamnus lycioides), als Monegros, pot perdre, algun any, entre el 50% i el 75% de l’àrea foliar total. Ens alguns casos hi ha substitucions de fulles més grans per altres de noves, més petites i xeromòrfiques. Les fulles estivals, sovint, són també més piloses, de manera que es redueix la transpiració ja que l’aire relativament quiet que queda entre els pèls té més resistència a la difusió del vapor d’aigua des de la fulla. El contingut d’aigua de les fulles varia també fortament al llarg de l’any, i això pot afectar la inclinació de la fulla respecte de la vertical i pot fer també que el limbe estigui més o menys estès. Molts cops, aquestes plantes produeixen essències volàtils que, ultra altres efectes que ja veurem, redueixen la transpiració. Entre els malacofil·les trobem moltes de les plantes que hem definit abans com a arbustos nans sempre verds de les zones mediterrànies, com ara les estepes i altres cistàcies, moltes lamiàcies (romaní, sàlvia, etc.) i també plantes d’altres famílies. Ho són, així mateix, alguns arbustos xeromòrfics d’hiverns freds (salat), i perennifoliats de zones xèriques (com la botja pudent), alguns coixinets, etc.

L’esclerofil·lisme ha estat considerat com una de les característiques més típiques de la vegetació mediterrània. L’aladern (Rhamnus alaternus) és un bon exemple d’un xeròfit de fulla esclerofil·la. Moltes espècies de l’alzinar i de les màquies i garrigues mediterranis tenen fulles semblants, amb una bona regulació estomàtica i una cutícula gruixuda que, ensems amb un aparell subterrani molt desenvolupat, fan que el contingut hídric sigui força constant, malgrat que sovint hi ha situacions de forta demanda evaporativa de l’aire.
Fototeca / MC
Els xeròfits esclerofil·les inclouen les plantes llenyoses de fulles fortament esclerificades, és a dir, riques en materials estructurals com la lignina i la cel·lulosa, i sovint recobertes de cutina. És freqüent que aquestes fulles siguin de mida força reduïda, associant esclerofil·lisme i microfil·lisme. La reducció de la transpiració durant el període eixut no es produeix tant per disminució de l’àrea foliar, com en el cas precedent, sinó sobretot per un control estomàtic més efectiu i una resistència cuticular a la transpiració més gran. La capacitat de les fulles esclerofil·les per a sobreviure sense aigua és molt superior que la de fulles blanes. Els estomes es tanquen quan encara hi ha força reserves d’aigua i poden suportar dèficits hídrics molt més alts sense mostrar símptomes de danys. Presenten rels capaces d’arribar a capes profundes, on el sòl conté sempre força humitat. El contingut hídric de les fulles és més baix que en el cas dels malacofil·les i també força més estable. Si l’eixut és molt intens, poden morir parts senceres de la planta, que funciona com si es tractés d’un feix de sistemes rel-tija-branques independents. Assenyalem que, tot i que l’esclerofil·lisme s’interpreta usualment com una adaptació relacionada amb l’economia hídrica, hi ha indicis que la deficiència en nutrients, concretament en fòsfor, pot determinar també uns nivells elevats d’esclerofil·lisme. Alguns autors consideren que quan aparegué el clima mediterrani les plantes adaptades a viure en ambients pobres en nutrients, que tenien fulles esclerofil·les, se’n beneficiaren perquè aquesta estructura de la fulla els facilitava la resistència als períodes de dessecació estival. Entre els xeròfits esclerofil·les es poden comptar moltes espècies dominants als nostres ecosistemes mediterranis, començant per arbres planifolis com l’alzina (Quercus ilex) i la surera (Q. suber), i arbustos planifolis com el garric (Q. coccifera), els aladerns (Rhamnus alaternus, Phillyrea), el boix (Buxus sempervirens), el llentiscle (Pistacia lentiscus) i molts d’altres. Algunes gramínies, com l’espart bord (Lygaeum spartum) o els pelaguers (Stipa), i també arbustos de fulles ericoides tenen, a més d’una estructura esclerofil·la, les fulles amb un canal central i poden plegar-se fins a adquirir secció circular, reduint molt, així, la transpiració.

Les tiges verdes fotosintetitzadores i la reducció, o total desaparició, de les fulles poden resultar avantatjoses per la disminució de les superfícies transpirants i dels costos de construcció. Aquests caràcters els presenten entre altres les plantes d’aspecte genistoide, com la mateixa ginesta (Spartium junceum), que veiem a la fotografia, o el ginestell (Sarothamnus scoparius). Fototeca / MC
Fototeca / MC
Els xeròfits afil·les comprenen espècies que han perdut les fulles o en conserven molt poques, i que, en canvi, tenen tiges verdes fotosintetitzadores. Són, com les anteriors, capaces de mantenir força constant l’estat hídric, controlant eficaçment la transpiració i explorant el sòl a fondàries considerables. En aquest grup s’hi troben moltes fabàcies, com ara la ginesta (Spartium junceum), el ginestell (Sarothamnus scoparius) i altres, però també s’hi poden incloure espècies de grups ben diferents, com les efedràcies. Moltes espècise de fabàcies són capaces de fixar nitrogen atmosfèric, gràcies a la seva associació amb bacteris simbionts. Malgrat aquest avantatge, sembla que normalment són plantes de taxes fotosintètiques relativament reduïdes.
El més característic dels xeròfits suculents és llur capacitat per a guardar aigua durant els períodes humits i gastar-la lentament durant els eixuts. Això els permet de sobreviure en condicions molt difícils. Tenen un sistema radicular molt superficial, que perd les parts absorbents durant els períodes secs i les torna a reconstruir molt de pressa per aprofitar qualsevol pluja, encara que sigui de poca intensitat. Això té sentit quan l’aigua subterrània es troba massa lluny, de manera que per a arribar-hi caldria un sistema d’arrels impossible de produir o massa costós. L’activitat fotosintètica segueix la via CAM, amb la captació del diòxid de carboni durant la nit, estratègia que permet de reduir molt les pèrdues d’aigua. A la nostra flora espontània manquen els xeròfits de tija suculenta, entre els quals destaquen, a Amèrica, les cactàcies i, a l’Àfrica, moltes euforbiàcies. Només un gènere, Caralluma, i encara de distribució molt restringida, s’ha trobat a la península Ibèrica, concretament a Andalusia. La figuera de moro (Opuntia) pot incloure’s en aquesta categoria, però és una planta introduïda. Trobem, en canvi, xeròfits de fulles suculentes. Les plantes d’aquesta mena no són pas rares als ambients rocosos. És el cas de força crassulàcies (Sedum, Sempervivum). També cal comptar-hi alguna altra espècie introduïda esdevinguda subspontània, com l’atzavara (Agave americana). No obstant això, és evident que es tracta d’una síndrome adaptativa que correspon, en conjunt, a països més àrids que el nostre i, tot i que no ens en manquen exemples, no té un paper de gaire relleu en el nostre paisatge. Es més important la suculència per causa de la presència de sals al sòl, de la qual ens ocuparem en parlar dels halòfits.
Tot i que alguns dels grups suara descrits podrien ser considerats com a xeròfits estenohídrics (per exemple, els esclerofil·les i afil·les), reservarem aquesta denominació per a aquells vegetals que regulen llur contingut hídric mitjançant la producció de substàncies especials, com el làtex de les euforbiàcies, d’algunes umbel·líferes i asteràcies (com Launaea i Lactuca). En aquestes plantes les fulles no s’assequen durant el període eixut, tot i que de vegades poden esgrogueir-se i caure. És clar que el làtex no és, com tampoc no ho són moltes de les que hem vist fins ara, una adaptació monovalent per a resoldre problemes d’economia hídrica, sinó que constitueix ensems un sistema de defensa contra els herbívors.
Les sals i les plantes: halofitisme
Quan la solució del sòl presenta una conductivitat elevada i hi ha una concentració forta de sodi, les plantes s’enfronten amb dificultats especials. Aquesta mena de condicions les trobem en zones litorals amb influència d’aigua salada i, també, en depressions tancades de territoris endorreics de les comarques interiors més àrides. Les plantes especialment adaptades per a viure-hi són considerades halòfits. Hi ha halòfits de morfologies molt variades, que comprenen arbres baixos com els tamarius (Tamarix), arbustos xeromòrfics d’hivern fred, com els salats (Atriplex), arbustos de tija suculenta ramificada, com les cirialeres (Arthrocnemum, Salicornia), arbustos de fulles suculentes, com les barrelles i altres (Suaeda, Salsola, Halopeplis), graminoides de diversos tipus, etc.
El problema essencial dels halòfits és extreure aigua de la solució del sòl, ja que per causa de l’elevada salinitat l’aigua és retinguda amb molta força al sòl (potencial osmòtic de valors absoluts molt elevats). Les plantes poden acumular sals a les fulles per mantenir un gradient de potencial que els permeti de xuclar aigua, però només fins a un cert punt, ja que es produeixen fenònems de toxicitat si la concentració salina creix massa als teixits vegetals. El clorur sòdic determina una hipertròfia de les cèl·lules. Els halòfits que toleren elevades concentracions de sals als teixits tenen, per aquesta raó, aspecte suculent a les fulles, a les tiges o a totes les parts aèries.
Alguns halòfits són obligats, altres facultatius, i apareixen diverses estructures per a reduir l’absorció de sodi o la seva acumulació. Són halòfits veritables espècies dels gèneres Suaeda, Salsola i Zygophyllum, de fulles suculentes, i les cirialeres (Salicornia i Arthrocnemum), de tiges suculentes. Hi ha halòfits que excreten sal de forma activa a través de glàndules o pèls transformats. En plantes com les estàtices (Limonium), el tamarius (Tamarix), Limoniastrum, Atriplex i altres quenopodiàcies, i fins i tot en algunes monocotiledònies (Aeluropus, Juncus), hi ha comportaments d’eliminació de sals per diversos mecanismes. Les formes de regulació per eliminació de sals solen correspondre a una suculència menys desenvolupada, mentre que els halòfits que no excreten poden reduir la salinitat per dilució, a través de la suculència. Alguns halòfits resisteixen un increment progressiu de la concentració salina als seus teixits al llarg del període vegetatiu (és el cas de certs Juncus), i renoven de tant en tant els òrgans on s’han obtingut concentracions massa elevades.
Hi ha halòfits que han de combinar la resistència a la salinitat amb la resistència a l’eixut. Hom els pot considerar simultàniament xeròfits. És el cas de Zygophyllum i d’alguns Atriplex. D’altres, en canvi, són sotmesos a períodes d’inundació importants, com les barrelles i les cirialeres, la qual cosa genera tota una altra sèrie de problemes. Alguns s’especialitzen a acumular clorurs, d’altres, clorurs i sulfats (aquests solen presentar menys suculència). Entre els halòfits no són rares les plantes que segueixen la via C4.
L’elevada salinitat crea, també, problemes per a la germinació. De fet, la majoria d’halòfits germinen més bé amb concentracions salines normals que amb les que es troben al seu medi. Potser els halòfits més estrictes (Suaeda, Salicornia) són els únics que poden requerir certs nivells de salinitat per a una germinació òptima. Potser per aquesta raó, com que la reproducció vegetativa sol veure’s molt menys afectada per la salinitat, la majoria de les espècies (Limonium, Aeluropus) la presenten preferentment.
La inundació i les plantes: helofitisme
Certes plantes estan adaptades a resistir llargs períodes d’inundació, en què es produeixen situacions d’anòxia prop de les rels. La anòxia va acompanyada de processos anaeròbics de descomposició, alliberament de nitrogen o d’òxids de nitrogen (N2O) en forma gasosa, acumulació de sulfhídric, altes concentracions de ferro i manganès en forma soluble i alliberament d’àcids carboxílics i hidrats de carboni (com l’etilè, que és un regulador del creixement vegetal). Per tant, es tracta d’un medi químic molt peculiar, que planteja greus problemes per a l’activitat dels vegetals superiors i llur adaptació.
Hi ha plantes que poden transportar oxigen de les tiges (des dels estomes o les lenticel·les dels troncs) a les rels, gràcies a l’existència al còrtex d’un aerènquima o sistema continu d’espais aeris grans. És el cas de l’arròs, del blat de moro (que només en té quan es cultiva sotmetent-lo a inundacions) i de moltes espècies de les zones humides; és habitual en aquestes espècies que les tiges siguin fistuloses, és a dir, que presentin una cavitat interior. Una altra adaptació consisteix a formar rels molt superficials, com fa l’arròs ja que, si són profundes, a més del transport d’oxigen des de les tiges s’ha de superar el problema de la fermentació. Hi ha diverses possibilitats: que les plantes puguin resistir períodes de fermentació relativament prolongats, que redueixin la taxa de fermentació per processos bioquímics, que eliminin els productes de la fermentació (acetaldehid i etanol, essencialment) a través de les rels i de les lenticel·les, o que n’evitin la formació per modificació del procés fermentatiu en el sentit d’acumular substàncies no tòxiques (malat, glicerol, aminoàcids). El ferro pot ser oxidat a formes fèrriques insolubles a la rizosfera o dins els espais intercel·lulars de l’escorça, i això fa que es formin dipòsits ben visibles d’hidròxid fèrric. El sulfhídric també pot ser oxidat, i la capacitat d’unes plantes o unes altres per a fer-ho més o menys poden determinar-ne la distribució (Molinia coerulea, per exemple, no suporta gens de sulfhídric al sòl). Alguns microorganismes poden contribuir a l’oxidació d’aquestes toxines.
En conjunt, hi ha, doncs, adaptacions anatòmiques, morfològiques, bioquímiques i de fisiologia molt diverses entre els helòfits. Els canyissars i els canyars són formacions helofítiques típiques, i també ho són diverses comunitats d’altres graminoides dels aiguamolls. No obstant això, el problema de la inundació afecta també formacions arbòries i arbustives. Als tròpics, els manglars formen ecosistemes forestals caracteritzats pel fet de desenvolupar-se en medis inundats, que presenten conjunts d’adaptacions, certament espectaculars, a la manca d’oxigen i a la salinitat. A casa nostra, les salzedes, les pollancredes, les vernedes i els tamarigars pateixen també inundacions prolongades, que estan preparats a resistir. Finalment, ja hem vist, en parlar dels halòfits, que també en aquest grup es dóna, de vegades, la necessitat de sobreviure en uns ambients periòdicament inundats. Molts vegetals són simultàniament halòfits i helòfits.
El foc i les plantes: piroftiisme

L’incendi destrueix l’estructura aèria del bosc, però aviat comença a manifestar-se la resposta biològica: rebrots d’alguns arbusts (a), plàntules de llavors enterrades (b), pinyons que cauen de les pinyes i germinen (c), etc. Queden els esquelets calcinats de les plantes que no rebroten i les branques cremades dels arbusts: un paisatge trist en el qual, però, no hi manca la vida, i on al més sovint és possible l’autoregeneració sense intervenció humana.
Mercè Cartañá, original de l’autor.
El foc, en certs ecosistemes, ha tingut de sempre una presència recurrent: en sediments lacustres de zones de bosc boreal de coníferes, hom ha pogut constatar incendis cíclics, amb una periodicitat propera als 200 anys repetida durant mil·lennis; i en zones de forta activitat volcànica, els incendis espontanis de la vegetació han estat també un fet freqüent. És difícil avui de determinar la freqüència dels incendis espontanis en zones de vegetació mediterrània o en praderies com a resultat, sobretot, de tempestes perquè les causes humanes són molt més habituals i modifiquen la dinàmica natural. S’ha especulat, però, sobre la possibilitat que els focs, tot i ser molt menys freqüents que avui, poguessin ser molt més extensos, com a resultat d’una acumulació més gran i més contínua de biomassa. L’acció pirogènica de l’home a la Mediterrània té també una gran antiguitat, ja que hi ha vestigis de l’ús del foc que es remunten a 100 000 anys o més. Això fa pensar que els focs antropogènics poden haver-se afegit als naturals i, tot plegat, fer que el foc hagi pogut ser un factor selectiu important en alguns ecosistemes. No obstant això, algunes adaptacions que s’expliquen de vegades com a resposta al foc podrien interpretar-se també d’altres maneres. De fet, tota resposta adaptativa ho és en un entorn complex, multifactorial, i potser no té sentit de discutir si hi ha caràcters que han evolucionat com a adaptacions al foc o no; és segur, però, que hi ha caràcters que han esdevingut avantatjosos competitivament en règims d’elevada freqüència d’incendis.
Entre els mecanismes per a protegir-se del foc que presenten les plantes tenim, en primer lloc, el desenvolupament d’una escorça gruixuda, que redueix la difusivitat tèrmica i pot tenir característiques que la facin menys inflamable. Sota l’escorça poden subsistir meristemes amb capacitat per a rebrotar i donar noves fulles. També les bases foliars rígides, que persisteixen sobre la tija després de la mort de la fulla, com passa a les palmes, les gramínies perennes i moltes falgueres, poden actuar com a protectors. A partir de les bases foliars protegides, les gramínies perennes poden rebrotar i regenerar-se ràpidament. Això fa de les praderies sistemes força estables sota un règim freqüent d’incendis i ha permès a l’home d’usar el foc com a sistema de gestió per a afavorir la producció d’herba verda per al bestiar. Altres formes de protecció es basen en el comportament del sòl com a aïllament tèrmic. Els rizomes i els estolons, les bases dels troncs i les rels poden tenir meristemes capaços de donar rebrots; les llavors enterrades solen quedar protegides del foc. En algunes espècies, les llavors dures tenen dificultat per a germinar, i es pot donar aleshores fins i tot una estimulació de la germinació amb l’acció del foc. Així, doncs, hi ha espècies que presenten un extraordinari increment de la germinació després d’un incendi (algunes estepes, per exemple). Germinar immediatament després del foc té l’avantatge de disposar d’una gran abundància de cendres (nodriment mineral) i d’una manca de competència per la llum. Les llavors també poden perdurar damunt la part aèria de la planta, quan estan incloses en estructures molt resistents, com és el cas de les pinyes seròtines que s’obren després dels focs, les quals, de passada, ajuden a la disseminació, ja que les pinyes enceses poden fer trajectes relativament llargs en pendents.
La destrucció de la cobertura vegetal pel foc és beneficiosa en general per a les espècies heliòfites. En alguns casos, l’elevació de temperatures a causa del foc determina la inducció de la floració, i aquesta es reforça per l’augment de llum. Així passa tant amb els geòfits, com amb les orquídies.
S’ha indicat també la possibilitat que algunes plantes puguin afavorir el foc com a estratègia per a deslliurar-se de la competència d’altres que poden interceptar més la llum, però que no tenen tanta capacitat per a regenerarse després del foc. Aquesta hipòtesi explicaria l’acumulació de resines i essències volàtils en espècies com els pins i molts arbustos mediterranis. L’avantatge de ser més inflamables i alhora més ràpides en la recolonització de la zona cremada conduiria a aquestes adaptacions, destinades, en definitiva, a establir un règim de pertorbació a mida del piròfit. Ja hem assenyalat, no obstant això, una altra explicació possible per al caràcter adaptatiu de la presència d’aquests. tipus de substàncies, amb relació a la reducció del flux transpiratori. Un punt important és que una alta inflamabilitat només pot ser un avantatge real si el règim de pertorbació no és tan intens que, entre foc i foc, l’espècie no tingui temps de desenvolupar el seu cicle complet fins a la producció de noves llavors fèrtils. Es tracta d’un problema real, especialment, com és obvi, per als arbres. Els pins mediterranis tenen tendència a presentar un temps de maduració (des de la germinació fins a la producció de llavors fèrtils) reduït respecte d’altres congèneres. El cas del pi de Canàries (Pinus canariensis) és interessant perquè té la capacitat de rebrotar, fet insòlit entre els pins i que podria relacionar-se amb l’elevada freqüència de focs espontanis amb què pot haver-se enfrontat al llarg de la seva evolució, en una regió eminentment volcànica.
Entre els piròfits, com es dedueix dels exemples que hem anat donant, hom pot reconèixer estratègies molt diverses. Repassarem ara, succintament, les definides més clarament.
Piròfits rebrotadors

Les dues estratègies principals de supervivència dels piròfits, plantes que no solament resisteixen el foc sinó que fins i tot són afavorides pels incendis, poden ser exemplificades per aquestes dues fotografies. A l’esquerra, la soca inflada del bruc boal (Erica arborea), una soca veritable o lignotúber, que és un òrgan diferenciat i especialitzat en la producció de rebrots, ja que presenta una estructura histològica característica, amb vasos espiralats. A la dreta, el cas de les estepes (Cistus), germinadors preferents o obligats: la planta es consumeix en l’incendi, però deixa càpsules amb nombroses llavors que solen germinar abundosament amb les primeres pluges i així asseguren la ràpida regeneració de les poblacions.
Mireia Abril, Jordi Vidal
Després del foc (i també després d’una estassada) algunes plantes conserven als troncs, a les soques o a les rels, borrons que poden estar en dormició fins a sortir-ne per l’estímul que determina la pertorbació. També pot ser que la pertorbació desencadeni la formació de nous meristemes i borrons. En tot cas, dels borrons estimulats o formats s’originaran rebrots que tendeixen a refer el model estructural destruït. La formació dels rebrots es pot fer en uns casos absorbint reserves d’aigua i glúcids que es trobaven al mateix cos del vegetal, i aleshores pot progressar molt de pressa, o bé cal esperar que hi hagi un bon subministrament exterior d’aigua, i aleshores el creixement s’iniciarà amb les primeres pluges significatives després del foc. El garric (Quercus coccifera) i el bruc (Erica arborea, E. multiflora) són exemples, respectivament, de la primera i la segona forma de fer. El punt des d’on es produeixen els rebrots també és important. Els arbustos que, a més de rebrotar del coll de la rel o de la soca, poden fer-ho des de les rels, recuperen molt de pressa l’espai que inicialment cobria cada individu, i, si són plantes dominants, l’autosuccessió és molt ràpida. En són exemples el garric (Quercus coccifera) i el roldor (Coriaria myrtifolia); aquest darrer, com el fenàs (Brachypodium) i la falguera aquilina (Pteridium aquilinum), rebrota a partir de rizomes i alguns Vaccinium ho fan a partir d’estolons subterranis, estructures que queden protegides del foc. Altres plantes rebroten només de soca, amb un creixement vertical molt ràpid però amb poca cober tura superficial durant els primers anys, com fan els brucs (Erica) i l’arboç (Arbutus unedo). N’hi ha, però, que tot i rebrotar de soca aconsegueixen una cobertura aviat important perquè els brots creixen amb un angle molt inclinat cap al sòl; és el cas del llentiscle (Pistacia lentiscus). Els geòfits, com els asfòdels (Asphodelus), l’abellera (Ophrys) i el gladiol (Gladiolus), creixen ràpidament a partir dels bulbs i floreixen molt aviat després del foc.
Els vegetals que rebroten i s’autoregeneren fàcilment són sempre organismes de vida llarga. Al més sovint, la producció de llavors és escassa i només serveix per a substituir els individus morts de vells o malalts, i es dóna aleshores una germinació en massa de plàntules molt localitzada al lloc buit. Aquestes plantes creixen després lentament, ja que tendeixen a desenvolupar molt les parts subterrànies. Arriba, així, tard la primera floració, però en plantes sotmeses a règim d’alta freqüència de pertorbacions la bona estructura subterrània és millor garantia de pervivència individual i de domini de l’espai que l’arribada ràpida a la maduració. Alguns rebrotadors poden ser considerats com a obligatoris, ja que pràcticament no produeixen llavors. Constitueixen clons, en els quals els individus se separen per fraccionament de rels o d’òrgans subterranis.
Piròfits germinadors
Tot un altre conjunt important d’espècies es regenera després del foc per llavors. Naturalment, la germinació sol necessitar unes condicions apropiades d’humitat, a més dels efectes desinhibidors o de xoc tèrmic i de l’enriquiment de nutrients que pugui haver produït l’incendi, i, per tant, caldrà esperar un període plujós perquè es produeixi, a part els requisits de temperatura propis de cada espècie. L’efecte perjudicial o estimulant del foc sobre la germinació depèn de la fondària en què es trobin les llavors en el sòl. Les estepes (Cistus), el romaní (Rosmarinus officinalis) i la gatosa (Ulex parviflorus) són exemples d’arbustos de comportament pirofític amb gran capacitat de producció de llavors i de germinació després del foc, i són també vegetals heliòfits per als quals l’increment de llum per desaparició de la coberta vegetal resulta un estímul en el procés de regeneració. En el mateix grup es pot classificar el pi blanc (Pinus halepensis), tan important als nostres paisatges, per bé que les llavors quedin sovint protegides no pas pel sòl sinó per les pinyes. La majoria d’aquests vegetals es regeneren obligatòriament per llavors, ja que són incapaços de produir rebrots o en fan molt pocs. Quasi totes les espècies floreixen i fan llavors fèrtils força aviat dins el seu cicle vital, i inverteixen un esforç proporcionalment molt gran en aquests processos, amb floracions molt abundants.
Tot i compartir aquests caràcters generals, podem distingir uns piròfits germinadors efímers i uns altres de vida llarga. És en aquest segon grup, és clar, que s’inclouen les plantes llenyoses abans esmentades. També són, de tota manera, essencialment espècies oportunistes que mantenen bancs de llavors molt importants al sòl, la qual cosa els permet d’ocupar aviat l’espai després de qualsevol pertorbació. En tots aquests casos, com veiem, hi ha pocs caràcters pirofítics que no puguin considerar-se merament oportunistes, si no és l’afavoriment de la germinació pel foc.
Piròfits rebrotadors facultatius
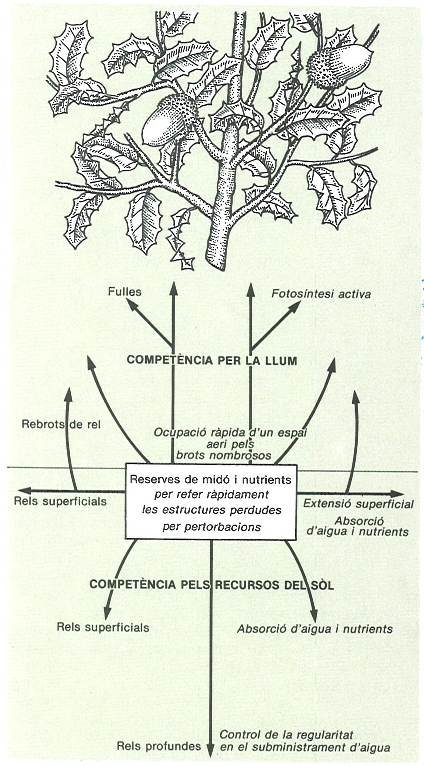
El domini de l’espai comporta el dels recursos més importants: llum en l’espai aeri, i aigua i nutrients en el subterrani. El garric (Quercus coccifera), adopta una estratègia de control i conservació de l’espai en un medi força fluctuant (recursos estacionals, incendis freqüents, etc.). S’estén multiplicantse vegetativament i formant denses masses que es regeneren per rebrots de tija, coll de rel i rel després de les pertorbacions, i assegura així el manteniment constant de l’espai.
Mercè Cartañá, a partir de fonts diverses.
Podem incloure en aquesta categoria intermèdia un conjunt de vegetals de vida relativament curta, que tenen una capacitat més reduïda que els rebrotadors típics per a sobreviure al foc, però que mostren més efectivitat en la reproducció per llavors, el creixement posterior de les plàntules i la ràpida maduració. No obstant això, són capaços de rebrotar quan el foc no ha afectat excessivament l’individu. Poden servir-nos d’exemples la farigola (Thymus vulgaris), la lledània (Teucrium polium), la botja (Dorycnium pentaphyllum) o l’espígol (Lavandula angustifolia).
L’antinòmia rebrotadors-germinadors és, sens dubte, un fenomen molt més general que la possible adaptació a medis amb alta freqüència de focs. El pirofitisme es limita essencialment, en tot cas, a afavorir certs aspectes (per exemple, que el foc estimuli realment la formació de rebrots o la germinació de les llavors), dins unes vies evolutives probablement en tots els casos ja predeterminades en la història de cada espècie.
L’especialització nutritiva
El comportament de les plantes enfront del quimisme del substrat varia d’unes espècies a les altres i això es tradueix en diferències de composició química dels teixits vegetals. Sabem que alguns halòfits poden acumular sals en quantitats superiors a les normals, i que això és precisament el que els permet de mantenir l’absorció per les rels, malgrat la força amb què l’aigua és retinguda al sòl. Hi ha molts altres casos de plantes que concentren diversos elements o que, simplement, són capaces de tolerar concentracions molt altes al medi, les quals solen ser tòxiques per a altres plantes. El comportament concentrador actiu o acumulador passiu crea ja una divisió interessant, però, ara com ara, no és possible de tenir informació suficient per a caracteritzar moltes espècies des d’aquest punt de vista. En tot cas, hi ha plantes especialistes de certs substrats que poden ser-ne indicadores per raó de llur fidelitat, com és el cas de moltes metalòfites de les regions mineres (flores cupríferes, plantes indicadores de zinc, plom, manganès, urani; flores dels sòls serpentínics indicadores de crom i níquel, etc.). A més d’aquests casos relativament rars d’especialització, hi ha fenòmens molt més corrents que ara ens cal considerar.
Característiques nutricionals de diverses plantes de l’alzinar de les muntanyes de Prades. Els histogrames mostren les desviacions percentuals de les concentracions de diferents nutrients a les fulles (a l’esquerra) i branques (a la dreta) de cada espècie, respecte dels valors mitjans per al conjunt de la fitomassa del bosc.
Carto-Tec, original d’A. Escarré i col·laboradors
Potser la gran divisió tradicional de les plantes, pel que fa als requeriments nutritius, és la que es fa entre aquelles que prefereixen els sòls calcaris (calcífiles) i aquelles que són pròpies de sòls no calcaris (calcífugues). Els sòls calcaris tenen un pH que, normalment, és proper a la neutralitat i es manté tamponat per les reserves de carbonat càlcic, mentre que els sòls no calcaris són àcids. Però el que resulta més decisiu per a les plantes no és directament el grau d’acidesa, sinó el fet que el pH determini la solubilitat dels diferents ions. Cal també afegir diferències de caràcter físic, ja que els sòls calcaris són més permeables, més secs i més calents. Als sòls àcids, el potassi és progressivament substituït en el complex d’intercanvi catiònic i després eliminat per lixiviació. Per la seva banda, el manganès i el ferro (en estat fèrric) s’alliberen en concentracions que poden arribar a ser tòxiques a pH baixos. Les plantes de sòls calcaris transplantades a sòls àcids solen presentar també símptomes de toxicitat per alumini. Si el pH baixa per sota de 4, l’alumini esdevé el catió dominant, i, com sabem, és extremament tòxic per als organismes. Aquest és precisament un dels fenòmens que, amb més insistència, han estat invocats per a explicar la decadència dels boscos centreuropeus atribuïda a les pluges àcides. En els sòls àcids, d’altra banda, la disponibilitat de nitrogen es redueix perquè baixa l’activitat microbiana i, per tant, la taxa de mineralització (i la descomposició s’alenteix perquè manquen quasi del tot els cucs de terra); el fòsfor assimilable, en presència de ferro i alumini, precipita. Entre les espècies vegetals calcífugues, dites també acidòfiles, s’observen certes peculiaritats en la composició química; a més d’una pobresa general de nutrients, presenten acumulacions de metalls com ferro, manganès i alumini. La menor abundància de calci al sòl la compensen, en part, per una eficiència més gran en l’extracció d’aquest element. Als sòls calcaris els problemes més importants no són per causa de la toxicitat, sinó de deficiències: elements com el manganès o el ferro poden serhi per sota dels requeriments de les plantes. Això és més evident en plantes adaptades a sòls àcids i transplantades a sòls calcaris, que aviat mostren clorosi per manca de ferro. D’altra banda, l’excés de calci pot interferir seriosament en l’absorció de potassi, i ser, d’aquesta manera, causa d’una altra deficiència. També el fòsfor pot ser-hi poc accessible, de vegades.
La pobresa de nutrients del sòl o oligotròfia, és relativament freqüent a casa nostra, molt sovint lligada a l’acidesa. En condicions oligotròfiques, s’afavoreixen alhora l’esclerofil·lisme, les fulles de vida relativament llarga i les plantes llenyoses o perennes de creixement lent, enfront dels caràcters oposats. Les plantes han de desenvolupar diverses facultats, com són: competir amb eficàcia pels nutrients poc accessibles o només disponibles de forma transitòria al sòl o a la virosta; poder funcionar i créixer en medis on els nivells de nutrients són més baixos que els que poden emprar altres possibles competidores; poder retenir per retranslocació proporcions substancials dels nutrients continguts a les parts del propi cos que sofreixen mortalitat estacional per causa de l’estrès o de l’envelliment; i, finalment, concentrar reserves a estructures de sosteniment i emmagatzemament aèries o subterrànies i a les llavors. Totes aquestes facultats s’obtenen mitjançant mecanismes diversos que configuren una colla d’adaptacions interessants i, per desgràcia, massa sovint poc conegudes.
Quan la composició del sòl té característiques molt especials, els canvis en la composició química de les plantes poden ser espectaculars. A les plantes halòfites (com els salats i les cirialeres, Suaeda, Arthrocnemum, Halopeplis, etc.) i a les pròpies de sòls guixencs (gipsòfiles) més típiques i a les gipsohalòfites (Gypsophylla, Ononis tridentata, Frankenia reuteri, Lithospermum fruticosum, Limonium, Tamarix, etc.) podem trobar una proporció molt elevada de minerals —cendres que resten després de la combustió al forn de la matèria orgànica dels teixits vegetals—, que de vegades sobrepassa el 50% (quan el contingut normal en cendres sol oscil·lar entre el 5% i el 15%). És freqüent que aquestes plantes siguin acumuladores de certs ions, com ara sodi i clorur les primeres o sulfat i calci o magnesi les segones, i que aquests ions determinin alts potencials osmòtics dels sucs cel·lulars i crassulència, que té característiques diferents en un cas i l’altre. També poden presentar proporcions molt altes de cendres algunes plantes de llocs sorrencs (psammòfits) prop de la mar, com la barrella (Salsola kali) o el rave de mar (Cakile maritima), que es mostren acumuladores de potassi. Altres halòfits que són també acumuladors de potassi mostren preferència per medis relativament ruderalitzats. Destaquem que l’acumulació de sals, a part la funció explicada, pot tenir un interès defensiu, perquè fa que la planta resulti poc agradable o immenjable per als animals.
Els sòls serpentínics també tenen característiques peculiars. S’hi fa una vegetació especialitzada els components de la qual poden acumular concentracions enormes de magnesi, en proporció molt superior al calci, la qual cosa és totalment anòmala.
Un cas més interessant per la seva freqüència és el de les plantes nitròfiles, pròpies de sòls amb taxa de nitrificació elevada i disponibilitat de nitrats abundant. Les grans herbes d’ambients ruderals solen ser plantes nitròfiles per excel·lència, i, en general, presenten concentracions de nitrogen relativament altes a llurs teixits. És el cas de l’ortiga gran (Urtica dioica), el card marià (Silybum marianum), Peganum harmala i moltes altres. A les comunitats d’ambients ruderals són especialment rellevants al nostre país les quenopodiàcies i amarantàcies i algunes compostes (Onopordon, Carduus, Cirsium, etc.). No obstant això, la nitrofília no queda restringida a les plantes dels ambients ruderalitzats per l’acció humana. Alguns ambients salins propicien també la nitrificació i això fa que els halòfits presentin sovint nitrofília (Atriplex). A les zones subalpines de la regió temperada, i en particular als Pirineus, les comunitats de megafòrbies ocupen llocs d’aire i sòls molt humits, engorjats o propers a salts d’aigua, o alguns pedruscalls sobre sòls profunds i rics en nitrats. Les plantes dominants d’aquestes comunitats han de considerar-se nitròfiles, com la imperatòria (Peucedanum ostruthium), el veladre (Veratrum album), l’adenòstil (Adenostyles alliariae), les tores (Aconitum paniculatum, A. napellus), el rovell d’ou (Trollius europaeus), la bistorta alpina (Polygonum alpina), etc. Un cas particular és el de l’enriquiment en nitrats per l’aportació de productes d’excreció dels animals. Això és sobretot important als llocs de repòs dels grans herbívors o dels ocells. De fet, algunes plantes d’aquests ambients, com les paradelles (Rumex), els blets (Chenopodium), el jusquiam negre (Hyosciamus niger), Sisymbrium, etc. poden haver-se expandit després cap als llocs ruderalitzats per l’home, però les trobem originàriament als peus dels penya-segats, les entrades de coves i altres llocs amb fortes aportacions d’excrements i orina, on el sòl té, més que nitrats, amoníac. Als prats hi ha nitròfiles associades als llocs més freqüentats pel bestiar com els pixallits (Taraxacum), les poes (Poa), el trèvol (Trifolium repens), etc. A les plantes de roca s’observen diferents graus de nitrofília relacionats amb els excrements dels ocells, essent els més elevats els corresponents a certes comunitats liquèniques. D’altra banda, els troncs vells dels arbres, els caus d’alguns animals i els talussos poden presentar també plantes de caràcter nitròfil.
Entre les herbes forestals hi sol haver importants diferències de comportament, de manera que algunes prefereixen els indrets més rics (eutròfics) en nutrients essencials i, sobretot, amb una taxa de nitrificació més alta. Així, el buixol (Anemone nemorosa) o el melcoratge de bosc (Mercurialis perennis) són herbes indicadores d’eutròfia a les fagedes, i també la gatassa (Ranunculus ficaria), la pulmonària (Pulmonaria officinalis), la prímula vera (Primula veris) i la gran (P. elatior), l’ortiga pudent (Stachys silvatica), etc. En canvi, altres són pròpies de sòls més oligotròfics, com la descàmpsia (Deschampsia), les lúzules (Luzula), la mèlica (Melica uniflora), l’escordi (Teucrium scorodonia), la falguera aquilina (Pteridium aquilinum) etc.
Algunes diferències de composició es relacionen amb l’estructura general de les plantes. Ja hem vist que les fulles de vida llarga solen tenir concentracions de nutrients essencials més baixes que les caduques, en correspondència amb el caràcter esclerofil·le i altres formes d’adaptació. No obstant això, en altres parts del vegetal la situació pot canviar. L’alzina, per exemple, té la fusta de les branques i, sobretot, l’escorça més riques en nutrients que la mitjana dels caducifolis (al contrari del que passa amb les fulles) i acumula grans quantitats de calci a l’escorça, fins i tot damunt de substrats silicis. Les herbes, en general, solen tenir més potassi, nitrogen i fòsfor que les fulles dels arbres i arbustos. No obstant això, algunes, com ara les gramínies, són relativament pobres i tenen una proporció baixa de cendres, fins i tot en medis salins o guixencs. En aquesta família, a més a més, és escàs el calci, mentre que adquireix més importància el silici, per raó del seu ús estructural.
La contaminació ha aportat un conjunt important de nous problemes per a les plantes. El cas més conegut és el de la presència de diòxid de sofre a l’atmosfera. Juntament amb els òxids de nitrogen, també procedents de contaminació, aquest compost és responsable de l’acidificació de la pluja i, per tant, d’efectes diversos sobre els sòls i les plantes per alteració del pH. En forma gasosa, els òxids de sofre i de nitrogen (SOx i NOx), amb el fluorhídric i l’ozó, tenen també efectes directes en els vegetals. Les plantes sotmeses a atmosferes contaminades poden acumular sofre i presentar diversos símptomes d’afectació. Es pot emprar l’índex sofre/nitrogen a les fulles per a detectar la presència de contaminants atmosfèrics. Generalment, aquesta relació té un valor de 0,03 a les fulles dels arbres. Quan hi ha contaminació per SO2 s’acumulen sulfats, a més del sofre incorporat a les proteïnes, mentre que la concentració de nitrogen roman igual i, per tant, el quocient S/N augmenta.
Al·lelopatia, defensa i comunicació
L’ambient químic dels vegetals és determinat també per les substàncies orgàniques. Un cas particularment interessant és el de l’al·lelopatia: algunes plantes alliberen al sòl substàncies que poden inhibir la germinació o el creixement d’altres vegetals. Les substàncies al·lelopàtiques poden procedir de les fulles, per rentatge per aigua de pluja o per volatilització a l’aire i adsorció al sòl, o bé de les rels, de la descomposició de restes vegetals com ara l’escorça radical o altres. Les substàncies al·lelopàtiques són sobretot terpens, fenols, alcaloides o cianides orgàniques. L’al·lelopatia sembla un fenomen molt general, no prou ben conegut si no és en alguns casos en els quals resulta particularment evident.
Són exemples ben coneguts els de les mates aromàtiques de les nostres timonedes, d’altres brolles i bosquines mediterrànies i de llocs secs amb hiverns freds, com és el cas de la botja pudent (Artemisia herba-alba), la camforada (Camphorosma monspeliaca) i d’altres. Ací hi ha pocs estudis fets, però al «chaparral» californià, Salvia leucophylla produeix terpens volàtils que, segons s’ha pogut demostrar, s’adsorbeixen sobre les partícules d’argila del sòl i hi inhibeixen la germinació i el creixement d’altres plantes. L’harmalà (Peganum harmala) produeix substàncies (alcaloides probablement) en totes les seves parts que poden inhibir el creixement i la germinació d’altres plantes.
L’efecte inhibidor de les micorizes exercit per substàncies que emet la bruguerola (Calluna vulgaris), que sembla decisiu per a evitar la invasió de la landa per arbres, i la inhibició de moltes espècies del sotabosc per la noguera (Juglans regia), per la boixerola (Arctostaphylos) i pels eucaliptus (Eucalyptus), són també exemples d’aquesta mena d’interacció. Les espècies productores d’alcaloides, com són el tabac (Nicotiana) i l’estramoni (Datura), també tenen efectes al·lelopàtics notoris; però altres menys esperables, com l’agropir (Agropyron repens), la pelosella (Hieracium pilosella), la col (Brassica oleracea), Aster canadensis, la moixera de guilla (Sorbus aucuparia), la canyota (Sorghum halepense), el gira-sol (Helianthus annuus) i arbres fruiters, com ara les pomeres i els presseguers, també produeixen diversos tipus de substàncies al·lelopàtiques. S’ha pogut veure que si s’empren residus forestals d’alzina triturats amb l’esperança de fertilitzar el sòl dels conreus, apareixen efectes inhibitoris de la producció. En definitiva, l’allelopatia ens apareix com un fenomen molt més general que hom no pensava anys enrere.
Les plantes i els animals poden usar diverses substàncies químiques com a sistema de defensa contra la depredació, que van des de la mera producció d’un gust desagradable fins a toxicitats molt fortes i d’acció rapidíssima. Els casos d’interacció química entre plantes i insectes, per exemple, són incomptables i comprenen des de l’exclusió del fitòfag a situacions de simbiosi (com passa als cecidis, que acumulen tanins, efectius per a evitar l’atac dels herbívors). També és conegut l’ús de toxines per depredadors (aranyes, serps, etc.). En general, les plantes amb fulles d’estructura persistent, com són arbres i arbustos, són pobres en nutrients i aigua i tenen compostos, com ara tanins, resines i silici, que són inhibidors del creixement. Els vertebrats solen consumir-ne, sobretot, els fruits i les llavors, els quals poden tenir característiques atractives en benefici de la dispersió, reforçades per un alt contingut de vitamines i nutrients minerals, a més de polisacàrids i lípids. Crucíferes, resedàcies i caparidàcies tenen glucosinolats que actuen com a antibiòtics, inhibeixen el creixement dels fongs, i també són al·lelopàtics, la qual cosa no impedeix que aquestes plantes tinguin força consumidors. Molts insectes semblen haver-se adaptat a aquests compostos i fins i tot els usen per a reconèixer més fàcilment el seu aliment habitual. Algunes brassicàcies tenen substàncies diferents que formen com una defensa de segon ordre (per exemple, alcaloides, com a Capsella), i així eviten els depredadors habituals de la família.
Un cas diferent és el de l’ús de substàncies químiques en la comunicació. L’exemple més conegut és, sens dubte, el de les feromones dels insectes, però hi ha exemples en el món dels animals, des dels protozous fins als vertebrats, que van des de l’atracció sexual i la senyalització del territori fins a l’alarma i l’evasió davant dels depredadors. Són menys coneguts els casos de comunicació entre plantes, però això no vol dir que no n’hi hagi. Recentment, s’ha posat de manifest que les plantes de pollancre i arç, sotmeses a l’atac d’insectes fitòfags, reaccionen produint fenols, que tendeixen a reduir atacs posteriors. El punt interessant ací és que el nivell de fenols i tanins augmenta també en plantes veïnes no atacades, per causa de l’acció d’una feromona. Una hipòtesi plausible és que els teixits lesionats produeixen etilè que, per via gasosa, afecta les plantes veïnes, induint-hi la biosíntesi dels esmentats compostos. S’ha especulat sobre la possibilitat que aquests fenòmens de comunicació puguin explicar fenòmens de sincronització d’ecosistemes.
Les relacions entre ecosistemes veïns
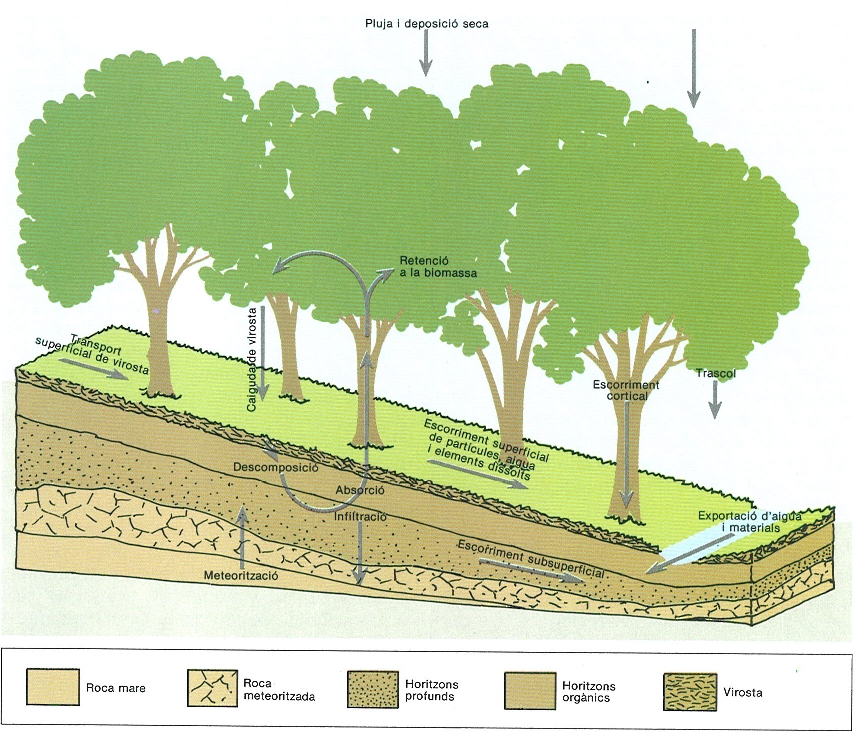
La biogeoquímica i el paisatge estan molt relacionats. A l’esquema es representen els principals processos de transport que tenen lloc al llarg d’un vessant (transport horitzontal) i a cada arbre o petita parcel·la de bosc (transport vertical). El transport horitzontal de materials per l’aigua o el vent és un fenomen bàsic, tant en geomorfologia com en ecologia.
Josep Nuet i Badia, a partir de fonts diverses.
Fins ara hem considerat l’ecosistema com la nostra unitat d’aproximació al funcionament de la natura en el medi terrestre. Hem parlat de paisatges només per explicar que constitueixen mosaics de comunitats en diferents fases d’evolució. No obstant això, és clar que entre comunitats veïnes de diferent composició i estructura hi pot haver interaccions relativament intenses. És habitual que aquestes interaccions siguin força asimètriques, de manera que un ecosistema pot beneficiar-se d’entrades d’aigua, nutrients minerals i matèria orgànica que procedeixen d’un ecosistema immediat. El resultat és un enriquiment progressiu del primer, mentre que el segon, sotmès a pèrdues (que equival a dir a explotació), es manté en un nivell inferior d’autoorganització. Els exemples més senzills d’aquests fenòmens els tenim en la influència de la topografia, el relleu. En efecte, el relleu crea una heterogeneïtat; les parts més convexes, és a dir els pendents més forts, són explotats: l’aigua de pluja i el vent arrosseguen substàncies dissoltes, partícules de pols, fulles seques i molts altres materials que tenen tendència a acumular-se al fons de les parts còncaves i a la base dels pendents.
Tota la geoquímica del paisatge depèn de processos com aquests, que se superposen a les possibles variacions de la litologia. Per regla general, trobem sòls molt més profunds, més rics i amb més capacitat d’emmagatzemar aigua a les parts planes i a les fondalades, que tenen un balanç positiu d’entrades i sortides, i sòls més prims, pobres i amb poca capacitat de retenir aigua, als vessants. Els ecosistemes que s’hi fan són també diferents, ja que damunt els primers hi haurà més capacitat productiva i més acumulació de biomassa.
Podríem trobar molts exemples en què el relleu determina, pels seus efectes sobre el transport lateral, certes pautes en la distribució de les comunitats de diferents nivells de maduresa i organització en el paisatge. L’aigua d’escorriment actua als fons de vall com un factor que pot arribar a ser d’estrès per excés. Al voltant del llit fluvial o d’un llac, hi ha un zona sotmesa a fortes fluctuacions per canvis en el cabal o el nivell freàtic i això afavoreix l’aparició d’ecosistemes amb una forta capacitat de regeneració i d’invasió. En canvi, en zones àrides de relleu torrencial (no endorreiques) la biomassa es concentra on la disponibilitat d’aigua és més elevada (contracció de la vegetació), i, per tant, hi ha un flux més gran d’energia externa.
Les relacions naturals entre ecosistemes explotadors i explotats en l’espai les retrobem, de forma encara més òbvia, en l’organització del territori sota influència humana. Les ciutats (ecosistemes urbans), treuen energia i materials dels agroecosistemes, dels llacs, els rius i les mars i dels boscos i els prats. Les carreteres i altres vies de comunicació són els camins per on circula aquesta energia. Cada parcel·la de territori queda, en la pràctica, dins l’àrea d’un mercat establert en un poble o en una petita ciutat. Uns pobles estan relacionats amb d’altres per vies de comunicació; i seguint una jerarquia de dimensions (associada al poder polític) aniríem arribant a ciutats cada cop més grans. Es tracta d’un sistema jerarquitzat d’explotació dels recursos naturals.
La societat humana, mitjançant aquesta organització en l’espai, ha incrementat els fluxos laterals de matèria i d’energia en els sistemes terrestres, i ha augmentat les dependències entre uns punts i uns altres del territori (amb un cost més gran, és clar, en termes d’energia afegida al sistema, essencialment com a combustibles fòssils). Quan volem entendre la funcionalitat d’un bosc explotat per una granja, hem d’ampliar el nostre model conceptual d’ecosistema fins a incorporar-hi ensems amb el bosc l’explotació agrícola i la casa de pagès. Sempre que les entrades i les sortides són regulades des d’un sistema veí, cal «allunyarnos-en» una mica i contemplar l’ecosistema que ens interessa com una part d’un altre de més ampli.
En aquesta perspectiva, l’ecologia del paisatge adquireix un interès creixent. Cada cop més, els ecòlegs tendeixen a pensar que només per aquest camí serà possible d’entendre el que s’ha anomenat la coevolució de l’home i el paisatge a la regió mediterrània. L’estructuració del paisatge és la resultant complexa de les relacions laterals d’explotació entre els elements del mosaic de comunitats que el constitueixen, incloses les comunitats antropogèniques; això ho hem de veure en un procés històric que pot haver condicionat canvis evolutius, fins i tot en la genètica de les espècies i en la morfologia del relleu. És habitual que tot això doni lloc a pautes repetitives. Així, encara que no hi hagi dos paisatges idèntics, és possible de trobar regularitats i d’agrupar tipus de paisatges.
Un tema crucial d’ecologia del paisatge és el del paper, progressivament minvant, que tenen els residus ja fragmentaris de sistemes poc alterats al nostre país i a grans àrees del planeta. La conservació d’aquests sistemes demana ineludiblement un disseny apropiat de l’espai que tingui en compte tant les dimensions mínimes per a permetre la subsistència de l’essencial de cada sistema i el manteniment de la diversitat biòtica, com la configuració espacial més efectiva per a la protecció. Margalef ha proposat reiteradament la idea que un reticle de comunitats més madures que emmarquen altres zones sotmeses a explotació pot resultar preferible a la preservació d’algunes grans àrees aïllades.