
Molts arbusts mediterranis rebroten activament després dels incendis, ja que la part subterrània es manté viva. El verd tendre dels rebrots de bruc contrasta amb el color encara ennegrit de les escorces dels pins, que no han resistit el pas del foc. La seva ràpida regeneració assegura una bona protecció del sòl enfront de l’erosió, una amenaça que és sempre present a les regions de clima mediterrani.
Josep Nuet i Badia
Podem trobar les formacions arbustives en medis que no permeten la instal·lació de comunitats forestals, o bé substituint aquestes quan són en fases de degradació o regeneració. En l’àmbit mediterrani són molt importants en el paisatge per dues raons: d’una banda, la llarga història d’ocupació humana, l’elevada densitat de població i els reiterats i variats processos de degradació que han sofert les masses forestals han fet que aquestes formacions arbustives ocupin actualment importants àrees de caràcter clarament forestal; d’altra banda, les condicions climàtiques d’una bona part de l’àrea mediterrània, que correspon a les zones més seques, amb una pluviositat anual inferior als 400 mm/m2, no permeten la instal·lació de formacions forestals tancades, sinó solament de comunitats arbustives amb diferents graus de desenvolupament.
Tant a l’àrea climàtica mediterrània, corresponent al domini de l’alzinar (Quercion ilicis), com a la del domini de la màquia (Oleo-Ceratonion) trobem una abundant i variada mostra de les formacions arbustives, des de les que són constituïdes per camèfits, com ara les timonedes, fins a les màquies de diferents menes, passant per les brolles, les garrigues i els espinars. A més, i per causa de la gran diversitat climàtica dels Països Catalans, on hi ha des del clima mediterrani sec de migjorn fins als climes de caràcter marcadament atlàntic o d’alta muntanya medioeuropea (sense període eixut a l’estiu), trobem també les formacions arbustives corresponents a la degradació de boscos caducifolis i de coníferes de caràcter centreuropeu, i també les formacions arbustives d’alta muntanya.
La importància quantitativa en el nostre territori de les formacions arbustives de caràcter mediterrani, enfront de les de caràcter atlàntic o d’alta muntanya, juntament amb les especials característiques de les primeres, ens ha decantat a centrar-nos-hi a l’hora de comentar i il·lustrar els diferents aspectes de la funcionalitat que tractarem. Les formacions arbustives mediterrànies presenten una fisiognomia i una funcionalitat molt particulars; hi trobem plantes xeròfites amb diferents tipus d’adaptacions per a la secada estival. A més de les particularitats relacionades amb la morfologia foliar i l’economia hídrica, ens en trobem també una colla pel que fa a l’economia dels nutrients; aquestes particularitats, lligades molt sovint a l’oligotròfia dels sòls, han estat ben estudiades en les àrees de clima mediterrani de Califòrnia, Xile i Austràlia, encara que també n’hi ha alguns estudis fets a la nostra zona.
Actualment, la isolínia dels 400 mm/m2 anuals de pluviositat marca bé el límit de la zona on el paisatge originari resta naturalment dominat per formacions arbustives; però quan hom intenta la reconstrucció de paisatges d’èpoques passades o la descripció de la vegetació potencial actual d’un lloc determinat apareixen problemes d’interpretació. Sembla que hi hagué profunds canvis climàtics associats a l’última glaciació, finalitzada fa uns 10 000 anys, però és difícil de descriure els efectes concrets a la zona mediterrània com també els canvis més subtils dels últims 5 milennis; tot i això, hom reconeix la importància de la degradació associada a l’activitat humana.
Sembla acceptat que el clima ha esdevingut més càlid i sec durant els últims milennis, però l’alt grau d’endemisme de la flora mediterrània suggereix la persistència, fins i tot en èpoques de glaciació, de condicions climàtiques no gaire diferents de les actuals, encara que potser només en àrees molt restringides. Per posar exemples podem dir que les anàlisis pol·líniques confirmen l’existència fa uns 8000 anys de formacions arbustives importants però d’altra banda, en jaciments del SE de la península Ibèrica, la zona més seca actualment i amb una flora molt característica que inclou un elevat nombre d’endemismes, s’han trobat restes de llúdrigues i tortugues d’aigua (que suggereixen l’existència de cursos d’aigua permanents), en poblats datats pels volts del 2400-1400 aC; sembla, a més, que durant aquesta època la zona estava densament poblada, cosa difícil d’imaginar en les condicions actuals. Segurament, l’home ha contribuït de manera important a la modificació d’un paisatge ja per ell mateix fràgil, amb una marcada tendència a l’erosió i l’abundància de sòls relictuals, accentuant, així, els lleugers canvis climàtics. Concretament, als Països Catalans, on les actuals condicions climàtiques permeten la coexistència de formacions forestals i arbustives, l’activitat humana dels últims segles ha d’haver afavorit clarament l’expansió d’aquestes últimes, fins a arribar a la situació actual en què la superfície que ocupen és molt extensa.
Dinàmica i funcionalitat
Tractarem els aspectes de dinàmica i funcionalitat de les formacions arbustives mediterrànies a partir de dades que fan referència a biomassa, producció, circulació de nutrients i evolució, després d’impactes com el foc a les estassades.
Biomassa
Les dades de biomassa total i la seva repartició en fraccions (fusta, fulles, fruits, rels) constitueixen paràmetres bàsics per a la descripció de la funcionalitat de les comunitats; moltes vegades, però, les dades existents es limiten a biomasses aèries i totals, sense especificar-les per fraccions. En termes generals podem dir que, òbviament, la biomassa de les comunitats arbustives és menor que la dels boscos i superior a la de formacions herbàcies; hi ha, però, altres diferències importants, com és el fet que la fracció corresponent a les fulles hi sigui proporcionalment més important que als boscos, amb totes les implicacions que això comporta, ja que es tracta de la fracció generalment més rica en nutrients i amb una taxa de renovació més elevada.
La forma d’ocupació de l’espai per una determinada biomassa també és una característica important, sobretot atenent els aspectes de recobriment i protecció del sòl contra la sequedat i l’erosió. Les formes de creixement són molt variables i ens trobem des dels arbustos amb un creixement bàsicament en alçada a partir d’una soca única (bruc, romaní, arboç) fins als de creixement més arrodonit, amb rebrots baixos i dispersos, en una superfície més gran (com per exemple els rebrots de garric). La descripció i la sistematització d’aquestes morfologies i tipus de creixements és molt senzilla i pot ser molt útil a l’hora de planificar intervencions (repoblacions, estassades) per a minimitzar els riscs d’erosió: el tipus d’espècie seleccionat per a una repoblació o la forma i la intensitat d’una estassada dependran bàsicament d’aquestes formes de creixement que fan les espècies més o menys aptes pel que fa a la protecció del sòl.
Segons quin sigui el tipus de morfologia i de creixement tindrem una biomassa més o menys compacta. Així com per un mateix tipus de comunitat es dóna una bona relació entre l’alçada mitjana i la biomassa acumulada (unes 25 t/ha de pes sec per metre d’alçada en el cas de les garrigues), quan comparem formacions diferents la simple observació de l’alçada de la comunitat pot ser molt poc indicativa de la seva biomassa acumulada; comunitats amb alçades semblants poden tenir biomasses que difereixen en un factor 4 a causa del diferent grau de compactació de la biomassa en l’espai. En formacions d’aproximadament 1 m d’alçada i recobriment total ens podem trobar amb biomasses tan diferents com 8 t/ha en un camp ocupat per olivardes, 25 t/ha en una garriga, i fins a 40 t/ha en un salobrar.
Un últim factor que determina la biomassa total és el de la densitat o recobriment de la comunitat considerada. Aquesta densitat depèn molt de la història (ús, impactes soferts) i d’algunes característiques ambientals; també són molt determinants la fondària del sòl i la disponibilitat d’aigua, que són bàsicament els paràmetres que fixen un límit a la biomassa suportable i obliguen a una distribució més o menys laxa dels individus. Aquests factors poden variar fortament en petites distàncies amb canvis associats al relleu; així, en timonedes alacantines les diferències són notables segons que la situació sigui de fondalada (recobriment d’un 80%) o de pendent o carena (recobriment d’un 15%): la biomassa en el primer cas és fins a 8 vegades superior al segon. No cal dir que el recobriment pot variar en un marge molt ampli; en comunitats de llocs molt secs o degradats (per exemple timonedes) o de llocs amb poc sòl i ambient molt dur (socarrells de muntanya mediterrània o de costes batudes pel vent) és normal trobar recobriments del 30% o inferiors, mentre que en altres situacions, i segons la història i les condicions ambientals, els recobriments poden oscil·lar entre un 100% i valors baixíssims.
D’aquestes consideracions es desprèn que només poden donar-se valors indicatius i corresponents a situacions concretes, però que tenen la virtut de donar una idea dels ordres de magnitud en què ens movem i d’acotar els valors més corrents. Fent números rodons, podem dir que ens movem entre valors d’1 i 2 t/ha en les comunitats més baixes i de menor recobriment fins a valors de l’ordre de 50 i 60 t/ha en garrigues ben constituïdes amb un recobriment total i una alçada d’uns 2 m; en màquies més desenvolupades podríem trobar valors més alts, però que difícilment superarien les 100 t/ha de pes sec. Aquests valors fan referència a la biomassa total en pes sec de les parts aèries.
La biomassa de rels i soques pot representar una part molt important de la biomassa de la comunitat, sobretot en arbustos de midapetita i adaptats a un clima on l’eixut és un problema important. Les dificultats metodològiques fan que siguin molt escasses les dades sobre biomasses subterrànies i, sovint, només es disposa de valors referents a individus aïllats, i no al conjunt de la comunitat. En arbustos de mida petita, com són les farigoles, no és estrany que hi hagi tanta biomassa subterrània com aèria, i fins i tot en arbustos grans com el bruc, la biomassa corresponent a soca i a rels representa, en un cas estudiat, comptant només les rels d’un diàmetre superior a 1 cm, un 34% de la biomassa aèria.
| COMUNITAT | LOCALITAT | EDAT (anys) |
RECOBRI- MENT(%) |
BIOMASSA (t/ha) |
|
|---|---|---|---|---|---|
| TOTAL | FOLIAR | ||||
| Timonedes | Alacantí | — | 15 | — | |
| Alacantí | — | 80 | 7 | — | |
| Socarrells | Mallorca | — | 50 | 2 | — |
| Herbassars dels guarets i deixius amb olivarda | Vallès Occidental | — | — | 8,5 | — |
| Vallès Occidental | 2 | — | 5,3 | — | |
| Vallès Occidental | 6 | — | 12,2 | — | |
| Brolles | Mallorca | — | 37 | 5,0 | — |
| Vallès Occidental | 7 | — | 5,0 | — | |
| Barcelonès | — | — | 13,4 | 4,2 | |
| Menorca | — | 90 | 21,6 | 4,1 | |
| Menorca | — | 96 | 17,1 | 3,1 | |
| Salicornar | Baix Ebre | — | — | 27,0 | 4,0 |
| Màquia | Vallès Occidental | — | — | 30,0 | 8,0 |
| Vallès Occidental | — | — | 21,0 | 3,9 | |
| Vallès Occidental | — | — | 23,0 | 3,2 | |
| Bruguera | Maresme | 15 | — | 15,0 | — |
| Maresme | 25 | — | 47,0 | — | |
| Ullastrar | Mallorca | — | 31 | 8,9 | — |
| Mallorca | — | 100 | 19,8 | — | |
| Garrigues | Garraf | 1 | — | 2,2 | 1,1 |
| Garraf | 2 | — | 5,4 | 2,8 | |
| Garraf | 3 | — | 7,1 | 2,9 | |
| Garraf | 13 | — | 10,9 | — | |
| Llenguadoc | 13 | — | 49,7 | — | |
| Garraf | 16 | — | 13,7 | 2,6 | |
| Llenguadoc | 17 | — | 23,5 | 4,0 | |
| Llenguadoc | 30 | — | 37,1 | 5,6 | |
| Dades compilades per l’autor | |||||
Parts fotosintetitzadores i parts funcionals

És general que els sistemes radiculars, en els arbusts mediterranis, siguin molt desenvolupats, de manera que poden arribar a uns quants metres de fondària. Les profundes rels asseguren el subministrament d’aigua durant els mesos eixuts i permeten una rebrostada intensa i vigorosa després dels incendis o les estassades.
Lluís López Soria
Moltes vegades, com ja hem dit, els estudis de biomassa fan referència a la biomassa total aèria sense distingir-ne fraccions. Són, però, força abundants les dades sobre biomassa de les parts fotosintetitzadores: els valors són més baixos o del mateix ordre que els que trobem en comunitats forestals i depenen fortament de la biomassa total, encara que el percentatge de parts fotosintetitzadores és més elevat quan els valors totals de biomassa són menors. Quant als valors totals, es mouen entre uns centenars de quilos per hectàrea en les comunitats menys desenvolupades, i xifres de 3-5 t/ha en brolles, garrigues o màquies ben constituïdes. Com ja hem comentat, varia fortament la proporció entre parts fotosintetitzadores (fulles i tiges verdes) i parts no fotosintetitzadores, i no solament entre espècies o formacions diferents, sinó fins i tot per a una mateixa comunitat segons quin sigui el seu grau de desenvolupament. En petits arbustos de les timonedes els valors del percentatge de biomassa corresponent a parts fotosintetitzadores oscil·len entre el 15% i el 75%; en socarrells els percentatges corresponents a fulles solen ser baixos (de l’ordre del 3% al 15%), però el conjunt de parts fotosintetitzadores (fulles i tiges verdes) arriba al 30% i 40% de la biomassa total; i en olivardes les fulles representen un 12% o un 15% de la biomassa total. Per a una mateixa formació aquests valors canvien segons el grau de desenvolupament; així, per a garrigues de diferents edats (entre 1 i 16 anys) el tant per cent de biomassa corresponent a fulles varia progressivament entre el 50% i el 20% aproximadament; per a brolles d’estepes, d’estepa blanca (Cistus albidus) i estepa borrera (C. salviifolius), entre 1 i 8 anys d’edat, aquests valors van del 35-40% a valors lleugerament inferiors al 10% respectivament.
En tot cas, els percentatges de parts fotosintetitzadores són superiors en les formacions arbustives que en les forestals, en les quals trobem quantitats sensiblement superiors de fusta; aquestes parts es renoven ràpidament (és poc freqüent que les fulles tinguin una vida mitjana superior als dos o tres anys) i són relativament riques en nutrients; això fa que el contingut mitjà (concentració) de nutrients en la biomassa sigui alt i la seva circulació més ràpida que en les comunitats forestals.
Els valors de l’índex foliar (metres quadrats de fulles per metre quadrat de terra) estan molt relacionats amb la biomassa i amb les característiques i la morfologia de les espècies presents. En individus (no comunitats) de petits arbustos com ara la farigola de Menorca (Teucrium marurum) i el socarrell (Launaea cervicornis) trobem valors d’aquest índex d’1 a 1,5, mentre que en arbustos grans com el garric (Quercus coccifera) o el llentiscle (Pistacia lentiscus) els valors poden ser de 4 o 5, o arribar fins i tot a 9. A escala de la comunitat, els marges de variació són també amplis, amb valors de 4 i 5 o superiors (en garrigues i màquies); és a dir, podem trobar índexs foliars i quantitats de pigments semblants als d’alguns boscos.
Necromassa
La quantitat de virosta present al sòl varia molt també d’acord amb el recobriment i l’edat de la formació; això últim és molt determinant ja que alguns fenòmens (com el foc) destrueixen gran part de la matèria orgànica i la virosta de les capes superiors del sòl. Com correspon a situacions on la taxa de descomposició és baixa, per causa de l’eixut, l’acumulació de virosta i matèria orgànica en el sòl pot ser molt important; així, en una garriga del S de França amb una edat de 17 anys, una biomassa total aèria de 23,5 t/ha i una caiguda anual de virosta de 2 o 3 t/ha·any, hi ha unes 80-100 t/ha de matèria orgànica acumulada en els primers 30 cm del sòl. Si parlem no de matèria orgànica sinó de virosta (dels primers centímetres del sòl), els valors seran evidentment molt més baixos tot i que poden arribar als que trobem en boscos; així, sota els coixinets d’eriçó (Erinacea anthyllis) de les parts altes de la serra d’Aitana s’acumula una quantitat de virosta que correspon a 1,5 t/ha amb un recobriment de només un 10%, és a dir que seria del mateix ordre que en un alzinar (12-13 t/ha) amb un recobriment del 100%.
Com en el cas dels boscos, es fa difícil de trobar comunitats en el seu grau màxim de desenvolupament, ja que la repetida i generalitzada acció humana directa o indirecta (explotació, incendis, erosió) manté la majoria de les comunitats en un grau de desenvolupament més o menys allunyat del màxim assolible en aquelles condicions. Això dóna una idea de l’amplitud dels marges en què ens movem a l’hora de donar dades de biomasses de comunitats arbustives, com ja hem comentat inicialment.
Economia hídrica
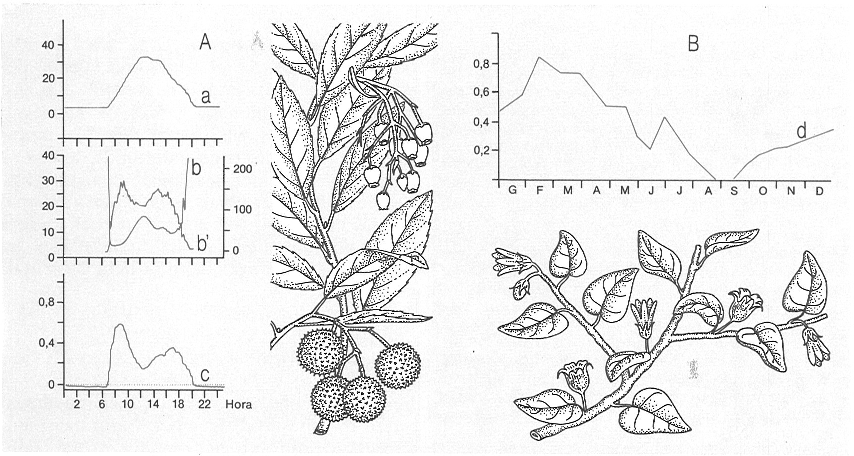
Dues estratègies oposades de les plantes enfront de l’eixut estival: A la de l’arboç (Arbustus unedo), un arbust esclerofil·le proveït d’un bon mecanisme de regulació estomàtica, i B la de la bufera arbustiva (Withania frutescens), de l’espinar alacantí, que actua com una caducifòlia estival. S’indica: a dèficit hídric (mg/l), b resistència a la difusió (S/cm), b’ transpiració (nM H2O O/cm S), c fotosíntesi neta (nM CO2/cm S), i d pes fulles/pes brots.
Mercè Cartañá, a partir de fonts diverses.
Les adaptacions dels vegetals encaminades a reduir les pèrdues d’aigua per transpiració són variades, sobretot en condicions en què l’eixut és important i l’aigua és un factor limitant; aquestes adaptacions combinen aspectes de morfologia foliar amb d’altres, com ara mecanismes fisiològics de regulació.
Els mecanismes fisiològies que asseguren una regulació eficaç d’obertura i tancament estomàtics solen presentar-se en els esclerofil·les. D’aquesta manera s’assegura una reducció de la transpiració (per tancament dels estomes) en les hores de més radiació o en qualsevol altre moment que no estigui assegurat un adequat subministrament hídric. En alguns casos, però (com per exemple en les estepes) la morfologia foliar i la regulació estomàtica no són suficients per a regular l’economia hídrica i apareixen fenòmens de reducció de la superfície foliar en l’època estival. Aquesta adaptació pot accentuar-se i donar lloc als arbustos amb dimorfisme foliar estacional o amb fenòmens de caducifòlia estival, que permeten una reducció important o total de l’àrea foliar en l’època eixuta. Les espècies amb aquestes característiques són abundants en els espinars meridionals: l’arç de tanques (Lycium intricatum), l’esparreguera de gat (Asparagus albus), per exemple.
El comportament hídric de les nostres espècies arbustives fou estudiat sobretot a partir dels anys trenta per ecofisiòlegs associats a l’escola sigmatista, al S de França. Després d’un llarg període en què l’activitat de recerca en aquest camp fou escassa, s’han tornat a iniciar estudis importants durant els darrers anys. Destaquem els realitzats per investigadors alemanys a Portugal amb espècies com el garric, l’arboç, el llentiscle i altres elements importants d’aquesta mena de comunitats, on es mesuren al mateix temps, els intercanvis d’aigua (transpiració) i de diòxid de carboni (fotosíntesi, respiració) a les fulles al llarg d’un perfil vertical que també s’analitza en variables microclimàtiques. Un dels objectius d’aquestes observacions és el d’establir models matemàtics que descriguin i permetin de preveure el comportament de la cobertura vegetal en diverses condicions. Caldwell i altres han pogut calcular, per exemple, que en un garric en què l’índex d’àrea foliar és alt (5,4) quasi totes les fulles —del 70% al 90%, segons l’estació de l’any— poden considerar-se en condicions d’ombra, i que la contribució total a l’absorció de diòxid de carboni d’aquestes fulles és tot just equivalent a la del 13% de les que ocupen els estrats superiors. De fet, aquests estrats superiors tenen un intercanvi gasós desproporcionadament alt. El consum hídric d’una comunitat de garric amb una àrea foliar més petita (2,2) ha estat avaluat per Rambal, a França, i per Caldwell i altres, prop de Lisboa, en uns 600 mm anuals en llocs amb 700 mm anuals de precipitació. L’eficiència, definida com la quantitat de diòxid de carboni fixat per molècula d’aigua consumida, disminueix en augmentar l’índex d’àrea foliar.
El garric o coscoll (Quercus coccifera) és una espècie relativament hidrostable (o sia, de contingut hídric força constant) tot i que s’han observat diferències, de manera que en llocs més secs el contingut hídric fluctua més. Les seves rels profundes li garanteixen un subministrament d’aigua més regular que en d’altres arbustos, com pot ser el cas de l’arboç.
Producció
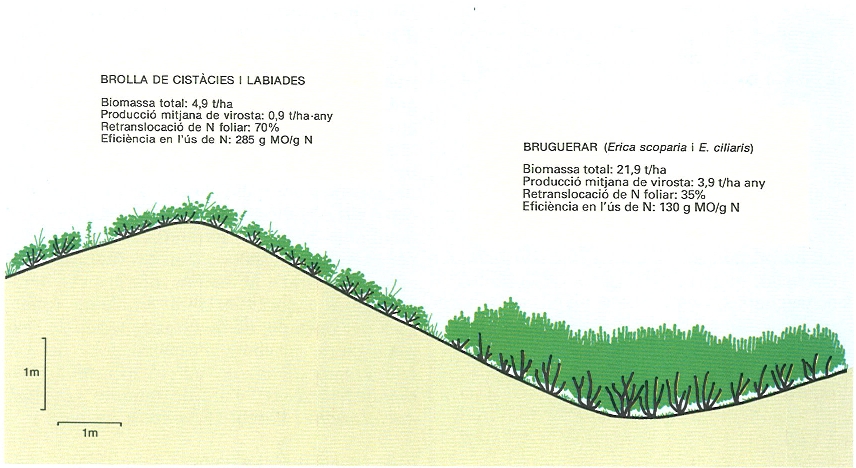
Efectes de la topografia sobre la biomassa i la producció en les formacions arbustives, com a condicionament de la disponibilitat d’aigua i de nutrients. Contrasten, en un àmbit espacialment reduït, les característiques de les formacions de carena, sotmeses a condicions d’oligotròfia i d’estrès hidric, amb les de les fondalades, sobre sòls més rics en nutrients i amb disponibilitat d’aigua. A les carenes, hi trobem formacions obertes, de baixa biomassa, alta eficiència en l’ús de nutrients i producció independent de la precipitació, mentre que a les fondalades hi ha una formació amb biomassa elevada, baixa eficiència en l’ús de nutrients i producció dependent de la precipitació. Aquest contrast ens indica que els factors limitants són la baixa disponibilitat de nutrients a les carenes i l’alta disponibilitat hídrica a les fondalades.
Josep Nuet i Badia, original de l’autor, amb dades de J. Merino i A.M. Vicente.
Les limitacions imposades per un clima com el nostre, amb eixut a l’època càlida i disponibilitat d’aigua limitada en molts moments de l’any, imposen clares restriccions en la producció d’aquestes formacions. És ben coneguda la relació directa entre evapo-transpiració i producció, i aquesta baixa producció també es posa de manifest en els boscos mediterranis, en comparació dels de zones més plujoses. D’altra banda, el sòl pot actuar com a limitant per dues vies diferents: d’una banda, en un relleu accentuat i amb propensió a l’erosió no és gens estrany de trobar sòls poc profunds, o fins i tot esquelètics, amb una baixa capacitat de retenció d’aigua, la qual cosa accentua la sequedat edàfica sota un clima de pluges escasses i irregulars, i pot imposar serioses restriccions al creixement de les comunitats; de l’altra, a la zona mediterrània, trobem sòls pobres en nutrients la disponibilitat dels quals pot ser un factor limitant no menyspreable, que es reflecteix, en els casos més extrems, en unes comunitats d’espècies adaptades a l’oligotròfia.
Cal puntualitzar, però, algunes coses sobre aquesta baixa producció, ja que en determinats casos poden aconseguirse valors elevats que fan més productives algunes comunitats arbustives que altres de forestals. Això passa en situacions molt concretes i durant períodes curts de temps, només d’alguns anys (ràpidament apareixen les limitacions morfològiques de creixement que no es donen en comunitats forestals, almenys de manera tan ràpida), en estadis inicials de la successió sobre sòls profunds (camps de conreu abandonats) o en estadis inicials de regeneració per rebrot després d’incendis o estassades, i en els quals la preexistència dels sistemes radiculars permet un creixement vigorós dels rebrots durant uns pocs anys.
Les dades de producció no són excessivament abundants en comparació dels boscos i, sobretot, en l’àmbit mediterrani. La migrada importància econòmica d’aquestes comunitats ha donat lloc a una manca de tradició en aquests estudis, a diferència del que ha passat amb els boscos, bé que actualment la situació ha canviat i hom en reconeix la importància, tant per l’àrea que ocupen com per la possibilitat d’explotar-les i pel seu paper protector del sòl després de pertorbacions. D’altra banda, l’estudi de la producció en aquestes comunitats s’enfronta amb considerables dificultats metodològiques. La majoria dels arbustos mediterranis són perennifolis i viuen sota un clima molt irregular, i aquesta combinació dóna lloc a uns ritmes de creixement també irregulars amb una brotada o més al llarg de l’any, cosa que fa molt difícil de datar tant les fulles com els brots nous. El mateix passa amb la fusta: la poca claredat amb què s’hi manifesten els anells de creixement fa dubtar de la correspondència de cadascun d’ells a un període determinat, perquè en un any se’n pot formar més d’un.
La metodologia a la qual se sol recórrer en aquests casos consisteix a datar amb la màxima precisió possible l’edat d’una determinada formació que hagués estat destruïda totalment per un incendi o una estassada, i a considerar com una bona estimació de la producció mitjana el quocient entre biomassa i edat; això és acceptable sobre períodes de temps no gaire llargs (2 o 3 decennis), ja que cal assegurar-se que no hi ha hagut una desaparició de quantitats importants d’estructures mortes per descomposició. Aquesta mateixa metodologia permet també de reconstruir de manera ràpida l’evolució del creixement i de la producció al llarg del temps si es localitzen formacions semblants que hagin estat destruïdes en moments diferents. El fet de considerar com una cronoseqüència el que en realitat no ho és (ja que no seguim una mateixa comunitat al llarg del temps, sinó que en un moment determinat fem observacions en comunitats semblants en diferents estadis de regeneració) afegeix una incertesa als resultats; el mètode presenta, però, l’avantatge de la rapidesa i es considera adequat per a descriure les tendències d’evolució en el temps. Els mètodes esmentats, però, limiten els estudis a comunitats que en un moment determinat i conegut han sofert una destrucció total per incendi o estassada i no són, doncs, aplicables a formacions que no compleixin aquesta condició. L’alternativa de marcatge i seguiment d’individus aïllats o parcel·les presenta el greu desavantatge de la lentitud.
Quant als valors, podem dir que els de producció aèria mitjana han de ser molt baixos per a les formacions arbustives baixes i poc denses, com ara timonedes, comunitats de socarrells o brolles d’estepes, i se situen per sota d’1 t/ha·any. En comunitats més denses i ben constituïdes com les garrigues o les màquies, els valors de producció depenen fortament de la disponibilitat d’aigua i de l’estadi de desenvolupament de la comunitat, però podem acotar-los aproximadament entre 1 i 4 t/ha·any de producció aèria en pes sec, de les quals la meitat o més poden correspondre a fulles; aquests valors no són menyspreables si pensem que molts boscos mediterranis (alzinars, per exemple) poden presentar produccions semblants.

L’olivarda (Inula viscosa), arbust de floració autumnal originari de les rambles mediterrànies, ha trobat un hàbit apropiat en terrenys de conreu abandonats o en llocs desproveïts de vegetació. Durant els primers anys de la colonització d’un terreny buit les olivardes assoleixen valors de producció elevats, de fins 5 o 6 t/ha l’any, semblants als de molts boscos mediterranis.
Fototeca/MC
En situacions particulars i durant períodes curts de temps poden donar-se valors més elevats. Així, per exemple, durant els 2 o 3 primers anys del procés de colonització de camps abandonats per poblacions d’olivarda (Inula viscosa) s’aconsegueixen valors de producció d’entre 5 i 6 t/ha·any; aquesta producció decau ràpidament, i als 8 o 10 anys ens trobem ja amb poblacions decrèpites i molt poc productives. Durant els primers anys de la regeneració de garrigues o bruguerars després d’un incendi o una estassada, el rebrot pot ser molt vigorós i la producció se situa entre les 3 i 4 t/ha·any, i cau ràpidament a partir dels 3-4 anys; per garrigues i en algun cas molt concret, se citen produccions sostingudes d’aquest ordre, pràcticament iguals a les de molts alzinars i pinedes.
Ja hem dit que gran part de la informació corresponent a producció s’ha obtingut a partir de cronoseqüències reals o fictícies. Al llarg d’aquests processos de recuperació del recobriment i la biomassa és molt normal que els valors més elevats de producció es donin durant els primers anys (estadis inicials) de la regeneració; quan el creixement de la població aconsegueix un determinat nivell, entren en funcionament processos de competència que frenen la taxa d’acumulació de la biomassa i determinen un augment de la proporció de biomassa morta. Això dóna, molt sovint, corbes d’acumulació de la biomassa de tipus logarítmic, amb una producció decreixent en el temps i amb tendència a estabilitzar-se. La durada d’aquest període inicial d’alta producció, el moment d’aparició dels fenòmens de competència, els valors de la producció estabilitzada i d’altres paràmetres del procés depenen de diferents variables, entre les quals podem destacar la disponibilitat d’aigua i nutrients, dependents del tipus i de la fondària del sòl; també cal destacar, com a factor important, la densitat de la població, ja que una elevada densitat d’individus provoca una ràpida acumulació inicial de biomassa, però també una ràpida aparició dels fenòmens de competència. Finalment, les espècies que constitueixen la comunitat són també determinants del funcionament global; podem considerar dos tipus bàsics i diferents d’espècies quant a la seva estratègia de regeneració: les que es regeneren per rebrot, que normalment són de longevitat i capacitat de creixement més altes, i les que ho fan exclusivament per llavors, que en general són espècies arbustives i semiarbustives de tipus colonitzador i amb baixa capacitat de desenvolupament. La combinació d’aquests diferents factors ens donarà una resposta diferent en cada cas. Per donar xifres il·lustratives podem dir que en un període de nou anys una brolla d’estepes pot acumular una biomassa d’unes 5 t/ha, mentre que en el mateix temps un bruguerar en pot acumular unes 10 t/ha i una garriga entre 15 i 20 t/ha; en el primer cas, i després d’aquest període, la biomassa ja s’estabilitza mentre que continua creixent per bruguerars i garrigues.
La circulació de nutrients
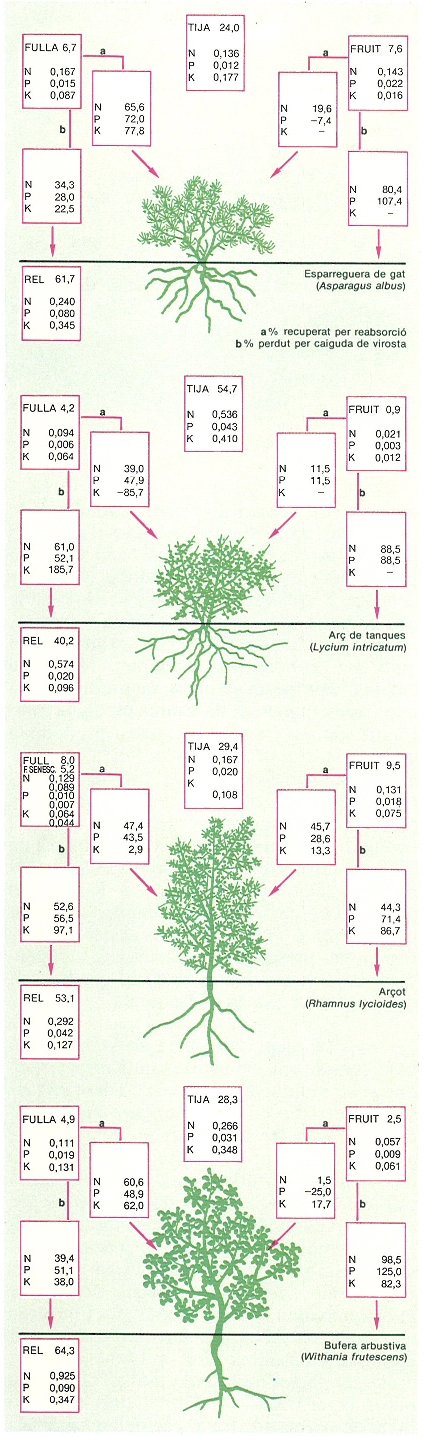
Quantitats de nitrogen (N), fòsfor (P) i potassi (K) emmagatzemades en la biomassa i percentatges recuperats per reabsorció o perduts per la caiguda de virosta en quatre espècies dels espinars alacantins. Observeu que les quantitats emmagatzemades a les rels poden ser molt altes i, també, que cada espècie té una capacitat diferent de recuperació de nutrients basics (N i P), però que, en tot cas, sempre es recupera més d’un 40% de l’element per retranslocació. Això assegura el control dels nutrients pels vegetals. En el dibuix, a representa el percentatge de reabsorció, i b el de pèrdua per caiguda de virosta.
Josep Nuet i Badia, amb dades de J. Martini.
La dinàmica dels nutrients a la comunitat és un altre aspecte important de la funcionalitat, que ha estat estudiat durant els últims anys, encara que no amb la mateixa intensitat amb què s’ha fet en boscos, com succeïa també amb la biomassa i la producció.
En les comunitats arbustives en general, el fet que hi hagi menys biomassa que en els boscos determina també una menor quantitat de minerals, però com que la proporció de les fraccions més riques en nutrients (fulles i parts fotosintèticament actives) és més gran, la concentració mitjana de nutrients en la biomassa i les taxes de circulació són més altes. Caldria esmentar la peculiaritat dels alzinars, en els quals trobem elevades concentracions d’alguns nutrients a la fusta, que donen lloc a concentracions mitjanes altes en la biomassa, més properes a les de les formacions arbustives mediterrànies que a les dels boscos extramediterranis. Quant a les taxes de circulació de nutrients, si comparem una garriga i un alzinar veiem que en la primera els nutrients circulen a doble velocitat, si prenem com una estimació d’aquesta velocitat el quocient entre les quantitats retornades cada any al sòl per la caiguda de virosta i les emmagatzemades en la biomassa.
L’estudi de la dinàmica dels nutrients en les comunitats arbustives mediterrànies ha tingut, i té, un interès no solament descriptiu sinó també relacionat amb l’adaptació a medis oligotròfics. No són estranys en l’àmbit mediterrani els sòls oligotròfics en els quals el baix contingut de nutrients representa un factor limitant per a la vegetació. De la mateixa manera que hom veu una convergència evolutiva entre espècies diferents que viuen en aquestes àrees geogràfiques separades, que s’interpreta com una adaptació al comú problema de la secada estival, s’ha buscat també alguna convergència evolutiva pel que fa al fenomen de l’oligotròfia, ja que la vida en medis oligotròfics i el manteniment d’unes taxes de producció i creixement moderadament elevades ha de comportar algun tipus d’adaptació.
Amb relació a l’oligotròfia, cal esperar una síndrome adaptativa en què es combinin característiques que permetin de reduir les necessitats de nutrients i tenir un control més gran dels seus cicles. Això comporta elevades taxes de retranslocació de nutrients de fulles i altres òrgans senescents, baixes concentracions de nutrients en la biomassa, especialment en la fracció fulles, i fulles perennes d’elevada longevitat, que alhora vol dir estructuralment resistents i poc atractives per als herbívors (esclerofil·lisme, acumulació de substàncies aromàtiques). Algunes d’aquestes adaptacions no tenen interpretació unívoca, ja que poden considerar-se també relacionades amb l’economia hídrica; és molt possible que ambdues interpretacions siguin correctes, però el pes de cada factor pot variar d’acord amb les condicions limitants; recordem l’exemple proposat en parlar de producció, en què en les situacions de carena on aigua i nutrients són, en principi, limitants, predomina el factor d’oligotròfia donant lloc a una formació que recicla el 70% del nitrogen foliar i l’usa amb una eficiència alta (285 g de virosta produïda/g de nitrogen perdut), enfront d’una comunitat de fondalada que recicla només un 35% del nitrogen foliar i exhibeix una eficiència de la meitat (130 g virosta/ g de nitrogen perdut); el caràcter marcadament limitant dels nutrients en el primer cas fa que aquesta comunitat presenti una producció independent de la disponibilitat hídrica i condicionada bàsicament per la de nutrients.
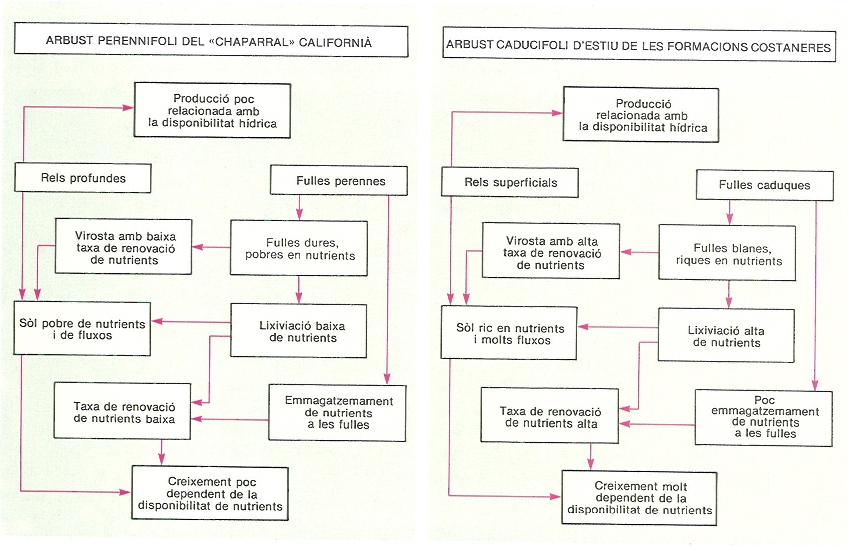
Relacions entre les característiques morfològiques i les funcionals en dos arbusts californians. L’estratègia adaptativa de l’espècie perennifòlia (Ceanothus megacarpus, del «chaparral») la fa més apta per a viure en medis oligotròfics, mentre que l’espècie caducifòlia d’estiu (Salvia leucophylla, de les formacions costaneres) queda restringida als sòls més rics en nutrients. És un exemple ben clar de la influència superior de la disponibilitat de nutrients sobre la disponibilitat hídrica en aquestes comunitats.
Mercè Cartañá, original d’A. Gray i H. Schleringer.
En totes les regions de clima mediterrani trobem formacions arbustives amb característiques que indiquen una especial adaptació als sòls oligotròfics en què es troben, i que coincideixen amb les resumides en els paràgrafs anteriors. Així, podríem citar bruguerars mediterranis, el «heath» australià, el «fynbos» sud-africà i el «chaparral» d’Adenostoma fasciculatum californià, el «mallee» australià; les característiques d’aquestes formacions contrasten amb les d’altres com la garriga o el «chaparral» de diverses espècies de Ceanothus, que viuen sobre sòls més rics i mostren una capacitat de creixement i producció més elevada. L’economia dels nutrients pot ser, doncs, un factor molt important a l’hora d’explicar la distribució a l’espai dels diferents tipus de vegetació. En el cas del «chaparral» californià s’ha demostrat la relació entre la distribució dels arbustos perennifolis (Adenostoma) i els caducifolis d’estiu (Ceanothus) i els continguts de nutrients del sòl, talment que els perennifolis se situen en sòls més oligotròfics gràcies als mecanismes abans esmentats, que els permeten un control més gran sobre els cicles de nutrients associat a unes necessitats seves més reduïdes. En altres casos, però, no s’ha detectat una eficiència especialment alta en l’ús de nutrients (en comparació, per exemple, d’espècies d’arbres caducifolis de clima temperat), ni tampoc diferències interspecífiques associades a la fenologia foliar, com per exemple en diverses espècies de l’espinar xèric alacantí: l’esparreguera de gat (Asparagus albus), la bufera arbustiva (Withania frutescens), l’arç de tanques (Lycium intrincatum) i l’arçot (Rhamus lycioides).
Cal, doncs, tenir en compte aquest doble condicionament de la producció, ja que tant l’aigua com els nutrients poden actuar com a limitants. Recordem només que l’origen filogenètic de la flora mediterrània se situa en tàxons existents sota condicions climàtiques diferents a les actuals i que podien correspondre a espècies adaptades a medis oligotròfics però amb unes característiques que les feien, també, especialment adaptables a medis secs.
La fauna
La gran abundància de grups taxonòmics diferents en la fauna de les formacions arbustives mediterrànies fa que manquin estudis complets i globals que permetin de fer-nos una idea clara de l’abundància quantitativa, i també de la producció secundària i de les influències de la fauna sobre la funcionalitat general de la formació. En la majoria dels casos, els estudis es refereixen a un o uns quants grups taxonòmics i en molts pocs casos hom passa de la descripció a la quantificació, per causa de les dificultats metodològiques.
Com a característica general de la fauna d’aquestes formacions, comuna a la de tots els ecosistemes mediterranis, podem citar la permanent activitat al llarg de l’any; tot i que tant la secada estival com el fred hivernal poden imposar restriccions a l’activitat o abundància d’algunes espècies i provocar fluctuacions estacionals en les seves poblacions, les condicions regnants al llarg de l’any poden considerar-se aptes per a una activitat ininterrompuda. Per a alguns grups, com ara la fauna del sòl, l’estudi d’aspectes com la diversitat, l’agregació i l’estratificació ha posat de manifest una gradació clara latitudinal des del tròpic fins a les altes latituds. En aquest esquema, la fauna del sòl mediterrani ocupa un lloc intermedi coherent amb la situació climàtica i geogràfica, a mig camí entre la diversificada fauna tropical i la poc diversa de les latituds elevades. Són, però, estudis restringits a uns pocs grups i que no arriben, ni de bon tros, al grau de quantificació aconseguit en l’estudi dels vegetals.
Quant a la descripció i l’enumeració de les espècies dels diferents grups presents en els diferents hàbitats o formacions cal afegir a allò que s’ha dit a la part de zoologia d’aquesta mateixa obra, que les relacions tròfiques entre plantes i animals són molt complexes i reflecteixen el grau elevat d’interacció dins l’ecosistema; una sola espècie de planta pot suportar una gamma molt variada d’espècies animals que n’aprofiten diferents parts: herbívors (menjadors de fulles, nèctar, pol·len, fruits), xilòfags, paràsits, xucladors de sava o sucs cel·lulars, necròfags, etc., a les quals trobarem també associades les corresponents espècies de depredadors i paràsits. Aquesta complexitat fa que es requereixin tècniques variades de mostratge, que, en combinació amb les dificultats d’identificació de molts grups, fan escassos els treballs globals.
Explotació i conservació
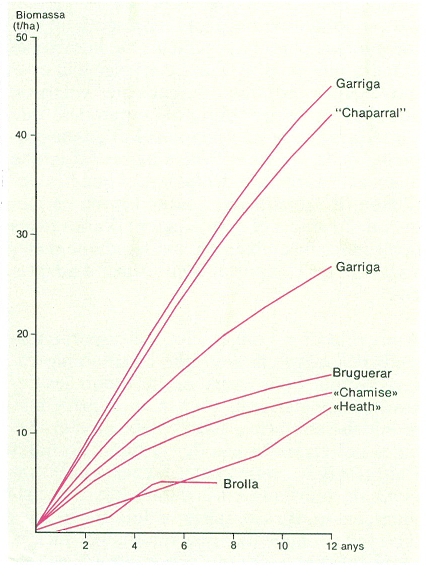
Recuperació del recobriment després d’una pertorbació, expressada en acumulació de biomassa, a diferents formacions arbustives del món. Noteu que un mateix tipus de formació (garriga) pot tenir diferents velocitats de recuperació segons les variables ambientals del lloc que ocupa.
Carto-Tec, a partir de fonts diverses.
La ràpida capacitat de regeneració de les comunitats arbustives després d’incendis, gràcies a la combinació d’estratègies de germinació i rebrot, fa que actuïn com a efectives protectores del sòl. Sota un clima com el mediterrani, en què el perill d’erosió és gran, l’existència de formacions arbustives denses i ben constituïdes assegura la protecció de sòls actualment no utilitzats (forestals marginals, conreus i pastures abandonats) però útils, tant pel seu paper de reguladors del cicle hidrològic com pel possible ús que se’n pot fer en el futur; cal pensar, a més, que el sòl és un recurs no renovable en una escala de temps humana i que la seva pèrdua és, per tant, irreversible. D’altra banda, l’erosió no és pas l’únic perill a què estan exposats els sòls; en el cas dels sòls bruns mediterranis, quan desapareix la cobertura vegetal es produeix una alteració profunda de la seva estructura per causa de la ràpida mineralització de l’humus i dels canvis químics provocats per la radiació solar; aquestes alteracions destrueixen l’estructura dels horitzons superficials que esdevenen, així, més vulnerables enfront de l’erosió. Si a això afegim que molts sòls mediterranis són relictuals (formats en altres èpoques, sota condicions més favorables que les actuals) i que, per tant, la seva regeneració actual és nul·la, podem comprendre la magnitud de la gravetat de la pèrdua d’un sòl.

El bruc boal (Erica arborea) s’ha emprat tradicionalment com a combustible en les fleques i, actualment, per a fer tanques o coberts molt més resistents que els encanyissats. La ràpida regeneració per rebrots de soca fa que es puguin fer estassades en torns molt curts (menys de 10 anys). Les soques dels mateixos brucs forneixen la fusta per a l’elaboració de pipes, encara que en aquest cas l’arrabassada de la soca comporta la destrucció de l’individu.
Miquel Riba
L’alta capacitat de regeneració d’aquestes formacions les fa especialment apropiades per a la seva explotació; en condicions en què no es comprometi la conservació del sòl, la biomassa pot ser aprofitada per a obtenir energia o transformada per processos de compostatge. Poden fer-se recomanacions molt senzilles per a determinar la intensitat i la manera d’estassar d’acord amb el pendent del terreny i amb la morfologia i l’estratègia reproductiva de les espècies dominants. Desconeixem, però, encara, molts aspectes de la biologia d’aquestes espècies que són bàsics per a planificar intervencions d’aquest tipus: quantes vegades pot rebrotar una soca?; com varia el seu potencial de producció en funció de l’edat i el nombre d’estassades sofertes?; quin és el paper de les soques i les rels com a possibles magatzems de nutrients mobilitzables?
En alguns casos l’estassada pot ser aconsellable, no per criteris de rendibilitat econòmica, sinó per evitar incendis. Això és especialment cert en brolles arbrades on la inflamabilitat de moltes espècies arbustives pot representar un seriós problema. Les consideracions fetes en el paràgraf anterior també són vàlides per a aquest cas. Cal, doncs, jugar amb diversos factors alhora (protecció efectiva del sòl, minimització del risc d’incendi, rendibilitat econòmica, etc.) i això complica la presa de decisions. Per a fer compatibles la protecció del sòl i la disminució del risc d’incendi s’han proposat estassades arreu i a curt termini en superfícies reduïdes per tal que, en cada moment, el paisatge sigui un mosaic d’àrees en diferents estadis de regeneració; cal programar les estassades de manera que, quan es facin, la quantitat de biomassa acumulada sigui important però encara no hi hagi suficient biomassa morta acumulada perquè el risc d’incendi sigui elevat, per aconseguir, així, un paisatge menys incendiable. No cal descartar tampoc l’ús de focs controlats quan aquesta pràctica permeti de reduir el risc d’incendis.
La reforestació de zones ocupades per formacions arbustives és desitjable i pot reportar beneficis econòmics, però cal que es dugui a terme donant prioritat als aspectes de protecció del sòl, com ja s’ha comentat en parlar de repoblacions i plantacions. Finalment, en algunes situacions difícils, com ara en sòls poc profunds, llocs molt eixuts, terrenys amb pendent fort, etc., cal considerar aquestes formacions com a vertaderes protectores del sòl, que vetllen per la seva conservació. Una adequada diversificació del paisatge, més gran que l’actual, pot convertir-lo en més productiu i, alhora, menys susceptible de grans incendis, tan corrents avui a la regió mediterrània.