Consideracions generals
Tractarem en aquest apartat alguns tipus d’ecosistemes sotmesos a condicions molt dures del medi físic, sobretot pel que fa al clima, com a resultat de l’altitud o del peculiar i extremat microclima de les zones culminals en muntanyes d’alçada mitjana. En aquestes situacions trobem una varietat considerable de tipus d’ecosistemes estructuralment diferenciats. En destacarem aquí únicament alguns aspectes.
El límit del bosc

El límit altitudinal del bosc a Catalunya el marca normalment la substitució dels boscos de pi negre (Pinus uncinata) per prats alpins. És el que recull aquest paisatge de la baga de Queràs, a la vall de Camprodon (Ripollès). La desaparició del bosc en aquest cas és força neta i destaca gràcies al color contrastat dels arbres. Prop del límit, els arbres es fan més petits i, sovint, perden el port arbori.
Jaume Orta
A mesura que ascendim en una muntanya, el clima esdevé progressivament més fred; això significa que l’hivern s’allarga i l’estació de creixement de les plantes s’escurça. També hi ha altres factors que es modifiquen, com són les precipitacions, la insolació, l’oscil·lació tèrmica diària i estacional i la proporció de precipitacions en forma de neu. Si ens apropem a la carena, augmenta molt la velocitat mitjana del vent. A aquests factors climàtics se n’afegeixen d’altres com el pendent, la inestabilitat del substrat, etc. A partir d’una certa alçada, la vida dels arbres es fa impossible i entrem en un estatge anomenat alpí. Massa a prop de la carena, encara que no s’hagi assolit el límit altitudinal del bosc, aquest també és substituït per altres formacions com a resultat dels factors esmentats.
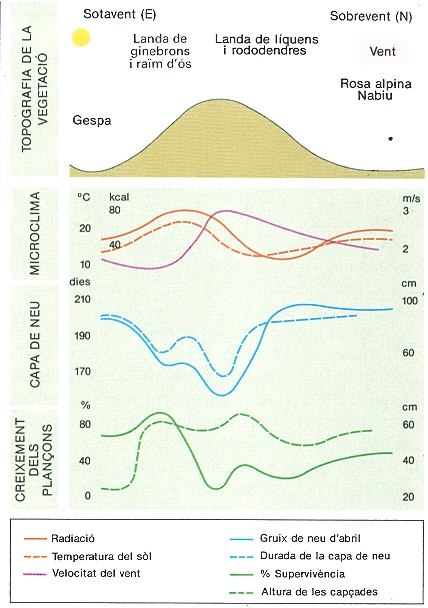
Influencia del relleu sobre la vegetació, el microclima i la innivació, en un cas estudiat als Alps suïssos (a dalt) en un vessant de 40-45° orientat al NE (les dades corresponen a valors mitjans i a l’estació vegetativa), i resultats dels cultius experimentals de Larix decidua (a baix), on es manifesten els factors més importants a l’alta muntanya: el relleu, la radiació, la temperatura del sòl, el vent, la durada i el gruix de la capa de neu, i, també, els seus efectes sobre la cobertura vegetal.
Carto-Tec, amb dades de W. Turner, citat per R.G. Barry
El límit del bosc està definit per una banda superior on les espècies d’arbres encara poden viure, però apareixen en formes de creixement no arbòries. Normalment hi trobem individus de tiges molt retorçades, sovint prostrades, de manera que els borrons de creixement romanen a menys d’un metre d’alçada. És el que en la terminologia científica internacional es coneix com a «krumholz». Aquesta forma especial de creixement és una resposta, segons sembla, a la dessecació de les fulles i a les glaçades; tots dos factors són resultants de la disminució de la temperatura mitjana anual. En efecte, el breu període de creixement fa que les fulles mostrin un desenvolupament incomplet de la cutícula, la qual resulta ser ara massa permeable al vapor d’aigua. Per tant, en moments de forta radiació solar directa o de vent intens i d’escassa disponibilitat d’aigua, les fulles poden gastar les poques reserves massa de pressa i assecar-se. D’altra banda, la prolongació del període de glaçades cap a l’estiu redueix més i més l’estació de creixement i el temps útil per a la producció activa i el creixement dels brots. L’escassa producció no pot compensar prou bé les pèrdues que es produeixen per dessecació i també per factors mecànics, que són molt importants a les zones careneres, com ara el vent i l’abrasió que produeixen les partícules que arrossega sobre els teixits vegetals, o com la neu.
Altres factors negatius es relacionen amb les condicions edàfiques: el sòl poc madur, el subministrament hídric irregular i alguns nutrients sovint poc accessibles. Les formes finalment obtingudes, en casos extrems, són similars a les que s’obtenen en jardineria amb arbres plantats amb poc sòl i podats per a limitar-ne el creixement («bonsai»). No obstant això, de vegades s’han pogut establir diferències hereditàries entre les formes nanes de muntanya i les normals. També vora la mar, arbres i arbustos de llocs exposats als vents prenen formes prostrades o en coixí. La salinitat hi contribueix, perquè actua com un aerosol sobre els extrems de les branques i sobre les fulles. Probablement sovint coexisteixen diferències purament morfogèniques i genètiques (aquestes darreres sobretot de tipus fisiològic).
La zona supraforestal
En general, hom està d’acord que el límit climàcic altitudinal del bosc es dóna, en molts territoris diferents, allà on la temperatura mitjana de l’aire del mes més càlid (juliol) és de 10°C. Més amunt del límit del bosc la radiació solar directa és forta, sovint la pluja ja comença a disminuir i les condicions varien molt d’acord amb l’exposició, que influeix sobre la durada de la neu (i, per tant, sobre les temperatures, ja que la neu protegeix el sòl durant l’hivern, i sobre el període de creixement, perquè si la neu triga molt a fondre’s el desenvolupament es retarda) i sobre l’exposició al vent. Els pendents, que poden variar molt, afecten al balanç hídric a través de la proporció d’aigua que es perd per escorriment superficial. Els cels clars determinen una forta irradiació nocturna i, per tant, una considerable oscil·lació tèrmica. A les muntanyes meridionals, el període de creixement és molt llarg tot i que les condicions d’hivern puguin ser molt severes, però apareix un risc més alt d’estrès hídric. Als llocs més secs o exposats als vents trobarem menys recobriment i un índex d’àrea foliar més baix.

Hivern i estiu en un paratge de la muntanya pirinenca: el puig de Balandrau, que separa les valls de Ribes i Camprodon. L’estacionalitat és molt marcada a l’alta muntanya, i l’activitat biològica, que hi és subjugada, es concentra molt especialment en el breu període favorable.
Josep Nuet i Badia
Les diferents combinacions de factors donen lloc a l’establiment de comunitats vegetals diverses. Així, les landes pirinenques de bàlec (Genista purgans) i boixerola (Arctostaphylos uva-ursi), amb bruguerola (Calluna vulgaris) als sòls més profunds i humífers, o de ginebró (Juniperus communis nana) sobre pedruscalls als cons de dejecció d’allaus, són pròpies de pendents forts o d’exposició a solana, on la neu no aguanta gaire, i on eviten, a més, els vents excessius. Els arbustos perennifolis de les landes viuen molts anys, creixen a poc a poc i tenen baixes taxes d’assimilació. Sembla que són normals taxes de creixement de les tiges d’1 o 2 cm l’any (nabiu, empètrum, etc.), i que aquest creixement s’esdevé, en un 80 o un 90%, en 20 o 30 dies. En general, no suporten temperatures inferiors als -30°C. Les reserves de carbohidrats són relativament escasses i la producció i la biomassa de les rels són inferiors a les aèries. Poden, en canvi, acumular lípids a les tiges. Aquestes landes formen de vegades mosaics amb retalls de bosc. A Penyagolosa, al País Valencià, les formacions del ginebró permeten la germinació, sota la protecció de la mata, dels arbres, però quan aquests creixen, el ginebró, extremament heliòfil, mor. D’aquesta manera es configura un sistema molt dinàmic, en què els arbres no arrriben a formar poblaments tancats i les taques de ginebró es desplacen per poder fugir de l’ombra d’aquells arbres que elles mateixes han contribuït a fer prosperar.

La solifluxió és una conseqüència dels processos de congelació i fusió de l’aigua del sòl en els pendents. Les gespes de festuca (Festuca) indiquen clarament el desplaçament del sòl en aquesta imatge de la serra del Port del Compte, als Prepirineus.
Jaume Orta
Les congestes presenten una zonació molt marcada de comunitats associada al temps d’innivació, des de prats on la neu no roman més de nou mesos, salzes nans (deu mesos de neu), molses com Polytrichum sexangulare (si encara s’allarga més), o bé hepàtiques com Anthelia juratzkana (els anys que la neu no s’arriba a fondre del tot), fins a arribar a neus perpètues sobre les quals pot viure l’alga Chlamydomonos nivalis, que hi dóna un color rosat degut a l’hematocrom que conté i que pot aguantar temperatures de -36°, però que, en canvi, mor a 4°C. A les congestes, la temperatura hivernal varia poc, no baixa de 0°C o 1°C, per causa de la protecció de la neu, i a l’estiu està també força regulada per l’abundància d’aigua al sòl. Alguns vegetals han mostrat una enorme resistència a la innivació, com és el cas del ranuncle glacial (Ranunculus glacialis), que pot sobreviure després de romandre 33 mesos cobert, però això no és gaire freqüent. També poden haver-hi plantes anuals de cicle ràpid (2-3 mesos), com la mucizònia de congesta (Mucizonia sedoides) i alguns geòfits. Les graminoides (joncs, càrexs, etc.) creixen així que es fon la neu, gràcies a les importants reserves de carbohidrats que tenen als rizomes (sovint de vida molt llarga) i a les bases de les tiges, i als nutrients que conserven sobretot a les poques fulles que mantenen tot l’hivern. En canvi, les plantes llenyoses troben dificultats mecàniques per resistir el pes de la neu, i queden pràcticament limitades als esmentats salzes nans, de branques totalment aplicades a terra, característica que les fa molt importants en la construcció de sòl.
És interessant d’observar que les gespes poden presentar temperatures molt elevades respecte de l’aire. Per exemple, en el cas del pèl caní (Nardus stricta) s’han mesurat a la superfície del sòl, on hi ha els meristemes de les gramínies, valors 25°C superiors als de l’aire (en dies assolellats). També les molses en coixí (Polytrichum, Drepanocladus) tenen temperatures interiors força elevades en relació amb l’aire i una humitat permanentment alta, i això és aprofitat pels petits artròpodes que hi viuen. Les formes de coixinet conserven molt la calor, de manera que les fulles d’azalea procumbent (Loiseleuria procumbens) poden allunyarse 15°C o 20°C de les temperatures ambientals els dies de sol i de 5°C a 8°C els dies núvols. Aquestes característiques tenen a veure amb la capacitat per a tenir activitat fotosintetitzadora i per a suportar temperatures molt baixes. Una de les plantes més resistents, que pot viure a les crestes més exposades i desprotegides per l’absència de neu, és precisament l’azalea procumbent. Aquest arbust aguanta fins a -36°C a l’hivern (només -8°C a l’estiu, com a conseqüència del cicle anual d’enduriment), amb les seves tiges totalment aplicades al sòl, on queda retinguda bona part de la pròpia virosta formant gruixos importants de matèria orgànica, i on les condicions de temperatura i humitat estan força ben controlades. S’ha pogut veure que la reducció de la velocitat del vent i l’alta humitat li permeten de mantenir l’intercanvi gasós (alta conductància estomàtica) de forma molt més estable que en altres arbustos, com ara la bruguerola (Calluna vulgaris). Pot, a més, formar rels adventícies que aprofiten, en el propi magatzem interior de virosta, l’aigua de fusió de la neu quan el sòl és glaçat. L’enorme capacitat de resistència té, com a contrapartida lògica, una baixa taxa fotosintètica i un creixement lent.
Per regla general, els arbustos tenen en conjunt taxes fotosintètiques més baixes que les herbes i les compensen amb una àrea foliar més gran i un període d’activitat més llarg. En les plantes alpines trobem els valors màxims de fotosíntesi a temperatures més baixes que en el cas de les d’altres procedències; i, a la mateixa temperatura, la taxa de respiració també és més alta en les primeres, com a resultat d’una taxa oxidativa superior a les mitocòndries de les cèl·lules. Un cas particular és el de les plantes de mullera que tenen la tija buida, com les cotoneres (Eriophorum), i presenten a la cavitat interior força concentració de diòxid de carboni que s’allibera en la respiració i pot ser reciclat per la planta. Aquest mecanisme contribueix probablement als ràpids creixements que solen presentar a la primavera. Un altre aspecte a considerar és la importància del substrat. D’una banda, sol haver-hi en totes aquestes comunitats una clara diferenciació entre plantes calcícoles i calcífugues. De l’altra, l’assimilació del nitrogen esdevé difícil sota les baixes temperatures dominants, i això podria explicar parcialment l’esclerofil·lisme, tan comú entre les plantes alpines. En són una notable excepció les megafòrbies que creixen, precisament, en els llocs enriquits en nitrogen.
A les clapisses, tan freqüents als ambients alpins és factible de trobar-hi exemples de successió amb diverses fases associades, en un petit espai, a les condicions d’activitat del substrat (la fixació progressa, naturalment, més de pressa a les vores d’una tartera i a les parts de menys pendent). Les pedres redueixen l’evaporació de l’aigua edàfica, cosa que afavoreix les plantes. Tot i així, hom reconeix algunes adaptacions xerofítiques i destinades a augmentar la reflexió de la radiació incident, com per exemple els toments de l’herba blanca (Senecio leucophyllus).
La vida animal al medi alpí

L’escurçó europeu (Vipera aspis) pot trobar-se a més de 2500 m d’altitud als Pirineus, on passa una hibernació més llarga que no pas a les localitats més baixes. La fotografia és feta a la serra del Catllar (Ripollès), al mes de maig.
Oriol Alamany
Les baixes temperatures i la curta durada del període favorable són els problemes més importants i determinen adaptacions interessants. En els amfibis i els rèptils, l’època de posta és retardada, augmenta la resistència a les oscil·lacions tèrmiques i es redueix el temps d’incubació. No és estrany de trobar comportaments ovo-vivípars i vivípars en espècies que són ovípares a la plana, com en la colobra llisa europea (Coronella austriaca) i en la sargantana vivípara (Lacerta vivipara).
La granota roja (Rana temporaria) pon els ous damunt la neu que cobreix els estanys, i la vida larval, que s’inicia així que la neu es fon, és molt breu. També entre els artròpodes hi ha nombroses adaptacions relacionades amb l’escurçament del cicle biològic. Una tendència general en alguns grups animals és la de presentar coloracions fosques, que augmenten l’absorció de radiació, com es pot veure sobretot en els rèptils. També són freqüents la quiescència (estat letàrgic induït pel fred) i la diapausa hivernal (estat letàrgic regit pel fotoperíode, una fórmula sovint usada en l’evolució de plantes i animals per a «predir» els canvis estacionals i avançar-s’hi). També són habituals entre els animals les migracions, de manera que la fauna resident és relativament escassa. La nidificació dels ocells sembla molt limitada per les baixes temperatures, de manera que són molt pocs els que la realitzen a l’estatge alpí.
A més del fred, la neu planteja també reptes especials en l’adaptació. Hi ha una fauna nivícola d’insectes que es manifesta en una microzonació segons el gradient d’humitat que hi ha al front de la neu en retrocés durant la fosa; aquesta fauna passa l’hivern sota la protecció de la capa blanca, com fan també alguns petits mamífers, en galeries molt superficials. Entre els insectes més característics hi ha curculiònids, estafílínids i d’altres, però destaquen els caràbids Cechenus pyrenaeus, Pyreneorites, Trechus i Timarcha. Poden considerar-se adaptacions a la neu la membrana que té l’isard entre les ungles o la coloració blanca de les plomes de la perdiu blanca a l’hivern.
Un caràcter molt freqüent entre els insectes és l’atròfia de les ales (en ortòpters i coleòpters), probablement associada a hàbits terrícoles i, fins i tot, excavadors; d’altra banda, les dimensions reduïdes i les formes aplanades, útils per a la vida a les tarteres i per a amagar-se entre les mates de gramínies i els arbustos reptants, es donen tant en insectes com en mol·luscs (per exemple, Vitrina nivalis, Helicella nubigena).
Entre els vertebrats, a més dels ja esmentats, a les zones alpines dels Pirineus es troben diversos rèptils, sobretot l’escurçó europeu (Vipera aspis), la sargantana de paret (Lacerta muralis), la pirinenca (L. monticola) i l’àgil (L. agilis) i la serp verda i groga (Coluber viridiflavus); ocells, com la perdiu blanca (Lagopus mutus), amb un plomatge blanc d’hivern que es confon amb la neu, el pardal d’ala blanca (Montifringilla nivalis), el pela-roques (Trichodroma muraria) i la merla roquera (Monticola saxatilis); amfibis com el gripau comú (Bufo bufo), la salamandra (Salamandra salamandra); i micromamífers com el talpó dels prats (Microtus arvalis), que hi és molt abundant, el talpó roig (Clethrionomys glareolus), més freqüent a les landes, el talpó de tartera (Microtus nivalis), les musaranyes cuaquadrada (Sorex araneus) i alpina (S. alpinus), l’almesquera (Galemys pyrenaicus) i la musaranya d’aigua pirinenca (Neomys fodiens). També hi ha alguns mamífers més grans, encara que són pocs els residents i la majoria només apareixen durant l’estiu. N’esmentem la llebre (Lepus europaeus), l’ermini (Mustela erminea) i la mostela (M. nivalis), la guineu (Vulpes vulpes), l’isard (Rupicapra rupicapra) i, ocasionalment, el senglar (Sus scrofa). A alguns residents, les tarteres els ofereixen refugis lliures de neu entre els blocs de pedra. En conjunt, però, són els prats els que ofereixen recursos relativament més abundants en productors primaris i gràcies a una considerable fauna estival d’artròpodes. L’escàs flux d’energia disponible limita molt els depredadors, que són pocs i poc diversos.
Els coixins espinosos de les muntanyes mediterrànies

La forma pulvinular apareix en plantes de diversos gèneres i famílies, i tant a l’alta muntanya com a la vegetació del litoral balear. La fotografía superior, presa a la serra d’Aitana (Marina Baixa), correspon a l’eriçó (Erinacea anthyllis), que és el coixinet espinós més estès a les zones culminals de les muntanyes mediterrànies i submediterrànies del nostre país. La inferior recull la presa de mesures quantitatives al llarg d’un transsecte, als socarrells del litoral menorquí, que permet de mesurar el grau de cobertura d’aquests pulvínuls, sota els quals s’amaga una activitat animal considerable.
Manuel Vicedo i Ferran Rodà
Hem vist, de passada, que algunes espècies tendeixen a adoptar formes de creixement amb les branques molt juntes i arrapades a terra. Era el cas d’arbustos com l’azalea procumbent, la boixerola, el ginebró, la savina de muntanya, el nabiu, etc., i també d’algunes molses. En tots aquests casos, la forma esmentada afavoreix la creació d’unes condicions interiors de microclima (temperatura i humitat) i de sòl millors que les de l’exterior. Un cas extraordinàriament interessant, encara que poc conegut en aspectes ecofisiològics, és el dels grans pulvínuls espinosos de l’alta muntanya mediterrània i els socarrells balears, que formen comunitats singulars i constitueixen un bell exemple de convergència adaptativa entre plantes de famílies i gèneres variats. Alguns d’aquests pulvínuls potser deuen la seva estructura a una pauta genètica de ramificació i creixement, mentre que d’altres adopten la forma de coixí com a resultat de la poda a què els sotmet sobretot l’acció eòlica. Per exemple, els socarrells balears eixorba-rates negre (Astragalus balearicus), eixorba-rates blanc (Teucrium marum subspinosum), socarrell ver (Launaea cervicornis) o socarrell bord (Centaurea balearica) presenten ramificacions amb angles molt tancats (de 10° a 15° el primer), que creixen de forma simultània i emeten eixos florífers que després s’assequen i resten sobre la planta, emergint del coixí i contribuint a la seva protecció. El socarrell retús (Dorycnium pentaphyllum varietat fulgurans) i el socarrell apiculat (Anthyllis hermanniae hystryx) mostren ramificacions en angles més grans (de 45° a 90°), similars als d’arbres o arbustos que adopten formes pulvinulars, com l’aladern fals (Phillyrea media varietat rodriguesii), l’ullastre (Olea europea) o el llentiscle (Pistacia lentiscus). Hom suposa que les plantes la forma pulvinular de les quals respon a una pauta genètica tenen menys pèrdues de teixits per l’acció del vent i resulten més efectives; també poden produir més i aconseguir majors creixements relatius, en igualtat de condicions, però el cert és que ambdues solucions poden coexistir.
Les comunitats xeroacàntiques, com se les ha denominat, solen donar valors de recobriment petits. A la serra d’Aitana, amb l’eriçó (Erinacea anthyllis) i l’eriçó groc (Vella spinosa), acompanyats de ginebre (Juniperus communis) com a dominants, el recobriment total mesurat en transsectes fou només de prop del 30%, del qual un 10% corresponia a l’eriçó i un 15% més l’ocupaven gramínies; el 70% restant era ocupat per pedres i rocs. A Penyagolosa, dos transsectes perpendiculars al pendent donaren una mitjana propera al 28% de recobriment només per l’eriçó, i fins a un 37% total; quedava, doncs, un 63% nu, però cal dir que més d’un terç del recobriment dels coixins corresponia a parts o individus morts. També en socarrells que s’estudiaren a Favàritx, amb un 35% de cobertura total, de la qual el 23% corresponia al socarrel retús (Dorycnium pentaphyllum varietat fulgurans), quasi el 86% del que ocupaven els coixins dominants eren estructures mortes que quedaven dretes. Aquestes dades fan pensar en el fort estrès ambiental a què estan sotmeses totes aquestes comunitats i, probablement, indiquen una taxa de renovació dels teixits més alta del que hom hauria pensat d’antuvi.
Com es desprèn del que hem dit sobre les condicions interiors als pulvínuls, cadascun és un veritable illot de vida en el desert pedregós que l’envolta. La virosta hi queda retinguda, juntament amb partícules de pols, gràcies a la seva estructura tancadíssima. A la serra d’Aitana, la capa de virosta a la comunitat abans esmentada (només sota els coixins) implicaria aproximadament 1,8 t/ha, quantitat que, si la cobertura fos del 100%, seria similar a la d’un alzinar ben desenvolupat (unes 12 t/ha). En aquesta capa de virosta, s’hi troba una abundant fauna de mida petita. També als socarrells balears, molts animals aprofiten les condicions creades pels coixinets, al sòl o a l’estructura aèria, com fa la formiga Tapinoma nigerrimum en construir-hi sovint nius temporals.
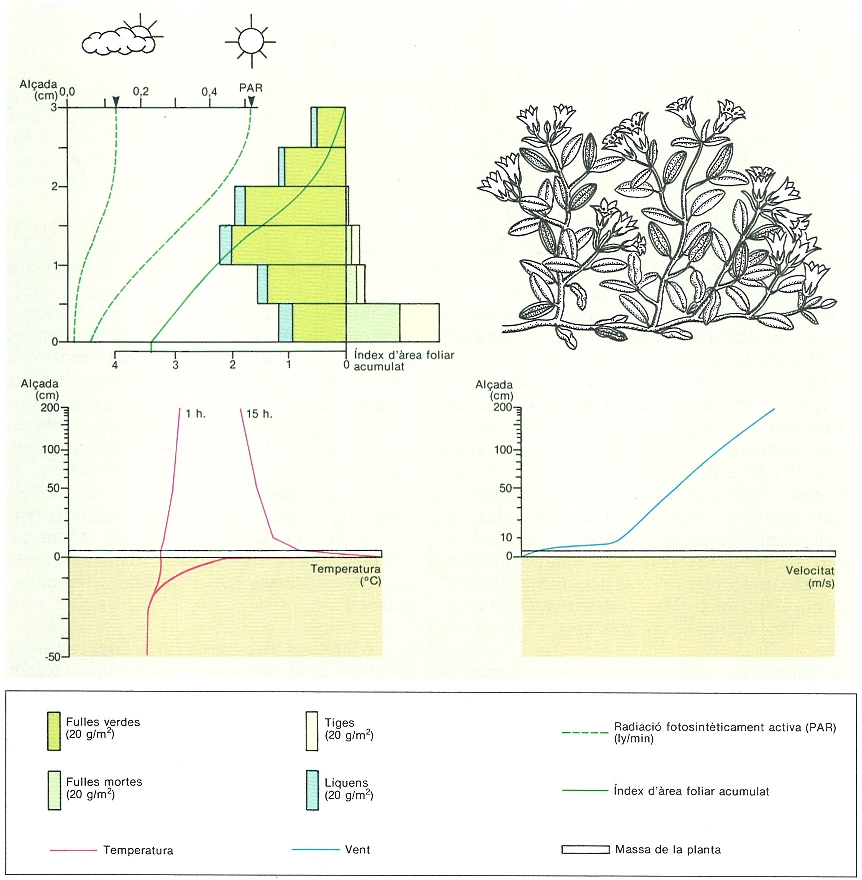
L’estructura densa dels coixinets vegetals de l’alta muntanya és una adaptació a l’adversitat del medi que aconsegueix modificar les condicions físiques a què és sotmesa la planta. L’esquema presenta els resultats d’uns estudis fets amb l’azalea procumbent (Loiseleuria procumbens) als Alps suïssos, bé que l’espècie és present també als nostres Pirineus. Mostra l’estratificació de la biomassa i l’índex acumulat de superfície foliar, juntament amb l’extinció de la llum en penetrar en el coixí. També s’hi veuen els perfils de temperatura en un dia clar d’estiu i la velocitat mitjana del vent entre mitjan juliol i mitjan octubre.
Mercè Cartañá, amb dades d’A. Cernusca
L’aparell subterrani dels coixins xeroacàntics és ben desenvolupat i, encara que no sol arribar al 50% de la biomassa total, s’hi acosta força en molts casos. En l’eixorba-rates negre (Astragalus balearicus), al pla de Cúber, a Mallorca, representava un 42%. Les rels tenen quantitats d’aigua molt més elevades que les parts aèries, on hi ha molta matèria morta (46% contra 28%), en el cas suara esmentat. Tots els coixinets tenen caràcters xerofítics, però de vegades l’estructura foliar ho és més marcadament. Així, mentre que l’eixorbarates negre té la cutícula relativament prima, estomes a les dues cares de la fulla i no té cobertura de pèls significativa, el blanc (Teucrium marum subspinosum) té la cutícula dos cops més gruixuda, estomes només a la cara inferior, que és coberta d’una densa capa de pèls, i les vores de la fulla revolutes; tot això contribueix a dificultar la difusió del vapor. Finalment, tots els pulvínuls tenen les fulles concentrades en una capa molt prima (que pot arribar a fer d’1 a 2 cm) exterior, i l’extinció de la llum cap a l’interior té lloc en un recorregut curtíssim, fent, a nivell del sòl, una ombra que és comparable a la que pot fer un bosc espès.
Encara que els ambients de la vegetació xeroacàntica presenten característiques similars als ambients alpins en alguns aspectes, també és evident que se’n diferencien en bastants d’altres. Un d’ells, important, és la reducció de la innivació. En aquest sentit, les semblances funcionals més grans podrien donar-se potser amb l’azalea procumbent, però les condicions xèriques segurament són més accentuades i el fred hivernal menys intens. No sabem per què els pulvínuls xeroacàntics són molt més alts (la forma tendeix a fer-se esfèrica) i punxosos, i és lícit de preguntar-se si aquest darrer caràcter pot haver-se desenvolupat evolutivament com una defensa contra l’herbivorisme. A l’Aties, on la vegetació xeroacàntica aconsegueix el seu màxim desenvolupament, s’ha pogut constatar que l’increment de les pastures l’afavoreix. El mateix passa amb altres espècies punxoses als Pirineus, com ara el bàlec (Genista purgans). Un altre fenomen interessant és la distribució dels socarrells, que apareixen tant al litoral balear com a zones supraforestals de la serra de Tramuntana de Mallorca. La innivació hi és sempre insignificant i el xerofitisme molt accentuat, i, sens dubte, el vent és un factor en comú. Encara hi ha moltes preguntes per respondre sobre totes aquestes comunitats. Fins i tot és possible que algunes de les seves espècies tinguin, gràcies a llur eficaç adaptació, un comportament ecològic que les faci bones colonitzadores, ja que sobre terrenys remenats per l’obertura de pistes, a les illes, hom hi pot observar que són sovint els socarrells els primers d’establir-s’hi, com fa per exemple el socarrell bord (Centaurea balearica) dins l’àmbit reduït en què es troba a Menorca.