
Les esponges o porífers tenen una gran importància en els ecosistemes bentònics marins. Són els metazous d’estructura més senzilla: sense teixits diferenciats, disposen de diferents tipus de cèl·lules especialitzades capaces de fer en conjunt totes les funcions animals. L’entrada constant d’aigua a l’interior de les seves cavitats assegura l’arribada d’aliment i l’intercanvi de gasos. L’espècie de la fotografia, Ircinia vanabilis, és corrent en els fons superficials del nostre litoral.
Enric Ballesteros
El tipus dels porífers és un grup molt ben delimitat d’animals aquàtics, amb representants a totes les mars i els oceans, i en molts rius i llacs; viuen a totes les profunditats, i des de l’equador fins als pols. La seva organització és definida per la seva manera de viure: essent bentònics i sèssils, viuen del que, per filtració, retenen de l’aigua que circula pel seu interior; tenen un sistema de canals interns i obertures superficials que permeten l’intercanvi que els és necessari per a respirar i alimentar-se. Són els metazous més primitius. Una de les seves característiques més peculiars és la plasticitat de la seva estructura. Una esponja és un organisme en continu moviment: les cèl·lules superficials (ectodèrmiques) poden enfonsar-se dins el cos (mesènquima) i transformar-se en diferents categories cel·lulars, que es desplacen mitjançant moviments ameboides. En canvi, no responen als estímuls amb contraccions sobtades, ja que no tenen veritables fibres musculars ni nervioses. No sempre han estat considerades pels naturalistes com animals metazous. Actualment, però, hi ha unanimitat en situar-los prop de la base dels metazous. La polèmica principal és en establir què és l’individu, és a dir, si les esponges són animals colonials o no. Una definició, simplificadora però artificial, considera individu tota aquella massa d’esponja delimitada per un pinacoderma continu, independentment de si té un o més òsculs.
Les esponges han tingut, des del Cambrià, un considerable paper dins els ecosistemes marins. La seva organització, encara que senzilla, els ha permès d’adaptar-se a canvis en les característiques ecològiques i de sobreviure sotmeses a condicions tròfiques difícils per a d’altres grups zoològics. La seva existència es coneix des de l’antiguitat: Aristòtil, 350 anys abans de Crist parlava de les esponges com d’animals marins dotats d’una gran capacitat de regeneració, i fins i tot n’analitzava la velocitat de creixement, aspecte que actualment encara és poc estudiat. Els primers habitants de la Mediterrània ja utilitzaven les conegudes esponges de bany: se suposa que els egipcis varen descobrir-ne aquest ús. Els soldats romans les feien servir per a beure-hi vi durant els seus viatges, en substitució de les pesades copes de metall. Es coneix, també, que la pesca d’esponges era inclosa en les proves dels antics jocs olímpics.
Tot i la seva reconeguda utilitat i el seu aprofitament comercial, de fet, de les prop de 10 000 espècies que se n’han descrit, les emprades amb diverses finalitats no arriben a 50. A l’Atlàntic septentrional les esponges acumulades a la platges després dels temporals són aprofitades com a adob per als camps de conreu. D’altra banda, fins no fa gaire, els pescadors gallecs encara brunyien els metalls dels vaixells amb esponges astrofòrides, guarnides d’espícules silícies. Però l’interès econòmic se centra en les esponges de bany, d’esquelet corni reticulat i flexible, que, això no obstant, són una minoria dins el gran grup: es tracta exclusivament dels gèneres Spongia i Hippospongia, encara que presenten diverses espècies segons les mars. Hi ha tres regions tradicionalment productores d’aquestes esponges: la Mediterrània oriental, especialment les costes de Grècia, Síria i Tunísia; el golf de Mèxic i el Carib, a l’Atlàntic septentrional; i les costes del Japó. Per als habitants de moltes de les petites illes d’aquestes regions, la pesca d’esponges va ésser l’únic mitjà de vida durant molts anys. La comercialització de les esponges al Carib se centrava en les costes de Florida, on hi havia la companyia manufacturera més important del món, que s’encarregava de distribuir fins i tot les esponges pescades a la Mediterrània. La producció de Florida va assolir fites molt altes el 1936 i el 1946, però hi va haver epidèmies el 1938 i el 1947 que la van reduir fortament, fins al punt que el 1951 es pescava només el 3% del que s’havia pescat el 1936. A la Mediterrània, la pesca abusiva va ésser la causa principal de l’enrariment dels jaciments d’esponges, ja que mai no es va tenir en compte el ritme lent de creixement d’aquests animals, ni es va establir una regulació de la pesca. Actualment, la major part de les esponges que hi ha al mercat europeu provenen del Carib (Cuba hi contribueix amb 20 t anuals) i del Japó. La pesca d’esponges, ja relativament minsa a la Mediterrània, va paralitzar-se totalment el 1988 a causa d’una nova epidèmia. Aquesta vegada, la infecció (bacteri o fong) va atacar també les fibres esquelètiques les quals feia tornar trencadisses. Al final del 1989, però, començaren a aparèixer petits exemplars d’esponges (1 cm), cosa que fa pensar en una propera recuperació dels jaciments d’esponges mediterranis. Als Països Catalans podem trobar dues esponges de bany: Spongia officianalis i Hippospongia communis, però no amb prou abundància com per a comercialitzar-les. Estudis recents estan obrint noves perspectives per a l’aprofitament de les esponges, especialment en el camp de la química orgànica, ja que s’hi han descobert esterols, terpens i isopirrols nous, que tenen aplicació en farmacologia i medicina: són substàncies tòxiques, antiinflamatòries, o amb incidència cardiovascular, gastrointestinal o antibiòtica, com l’esponjouridina, que s’extreu de Tethya cripta, de la qual ja s’han derivat tres agents antivírics i anticancerígens.
Morfologia
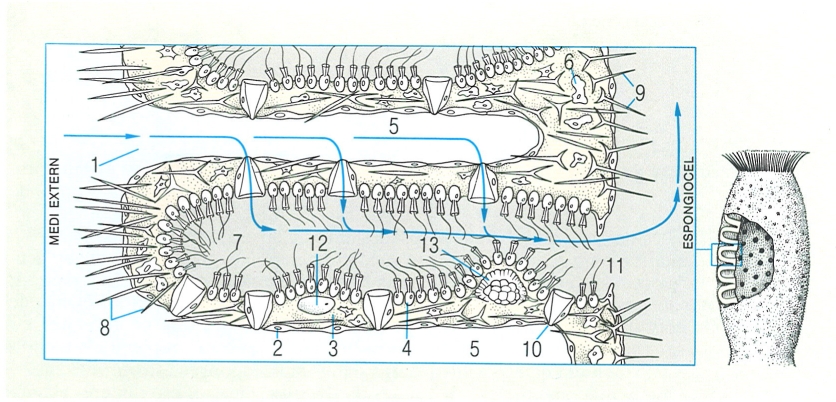
Esquema idealitzat de l’estructura, disposició cel·lular i elements reproductors en una esponja senzilla del gènere Sycon. Hom hi ha indicat: 1 ostíol, 2 ectosoma, 3 mesènquima, 4 coanòcit, 5 conducte inhalant, 6 amebòcit, 7 cambra vibràtil, 8 espícula la monaxona, 9 espícula triáxona, 10 prosòpil, 11 apòpil, 12 òvul, 13 amfiblàstula. La mateixa organització bàsica, més o menys complicada, es pot trobar en la majoria de les esponges (excepte en les hexactinèl·lides). En el dibuix, les fletxes indiquen la direcció del corrent d’aigua.
Biopunt, a partir de fonts diverses
Les esponges no mostren una simetria definida i creixen en formes molt variades, de manera que sovint el seu aspecte recorda el d’un vegetal. Les condicions ambientals, especialment la sedimentació i els corrents, influeixen particularment en llur morfologia; així, en superfícies horitzontals profundes, on la sedimentació és considerable, predominen les formes erectes, perquè això evita que el sediment n’obturi els porus, i poden ésser ramificades (Raspailia), tubulars, amb forma de copa o ventall (Phakellia) o de masses pedunculades (Rhizaxinella); en llocs batuts per les onades, creixen formant fines làmines incrustants que revesteixen el substrat. Una mateixa espècie pot presentar formes diferents segons l’hàbitat on es trobi. Quan hi ha corrents unidireccionals o continus d’aigua, es poden observar espècies, com Axinella polypoides, que es ramifiquen en un sol pla, perpendicular al moviment de l’aigua, per tal de maximitzar-ne el volum processat; aquest tipus de creixement no és exclusiu de les esponges, sinó que també s’observa en d’altres animals colonials bentònics (gorgònies). Presenten colors vistosos, vermells, ataronjats, grocs, violacis, i a vegades blaus, en alguns casos a conseqüència de l’associació amb algues simbionts (zooclorel·les verdes, zooxantel·les grogues o cianofícies blaves i vermelles).
El sistema aqüífer

El sistema aqüífer de les esponges pot tenir un grau de complexitat molt divers, peró hom n’ha definit tres tipus generals: àscon, sícon i lèucon, que sovint no s’arriben a distingir per la forma externa que adopta l’esponja. Les fotografies en mostren dos exemples. A l'esquerra, Hemimycale columella, un demospongi que viu als fons submarins del nostre país; el seu sistema aqüífer té els porus inhalants agrupats en zones ovalades o cribes. A la dreta, fotografia al microscopi electrónic d’escandallatge de l’interior de l’esponja Sycon, on es poden veure els tubs radiais i els coanòcits, equivalents a les cambres vibràtils d’altres esponges més complexes, els conductes inhalants i els orificis de sortida de l’aigua o apòpils.
Xavier Safont / M. Alba Camprubí i M. Jesús Uriz
Les esponges intercanvien gasos amb l’aigua que filtren, en retenen les partícules de què s’alimenten i hi alliberen metabòlits. Han de tenir, doncs, un mecanisme per a bombar l’aigua cap al seu interior i tornar-la al medi un cop filtrada. El cos d’una esponja conté una sèrie d’orificis i conductes de diferents diàmetres, amb cavitats entapissades de cèl·lules proveïdes d’un flagel; aquest conjunt es coneix amb el nom de sistema aqüífer. L’aigua entra dins l’esponja per uns orificis microscòpics anomenats porus inhalants o ostíols, dispersos per tota la seva superfície o agrupats en zones especialitzades; al llarg d’uns estrets conductes (inhalants o aferents), arriba fins a unes cavitats allargades o subesfèriques (cambres vibràtils), on els coanòcits (cèl·lules flagel·lades) en capturen l’aliment i impulsen l’aigua cap als conductes eferents i fins a uns altres de més amples, macroscòpics, els quals desemboquen en els òsculs.
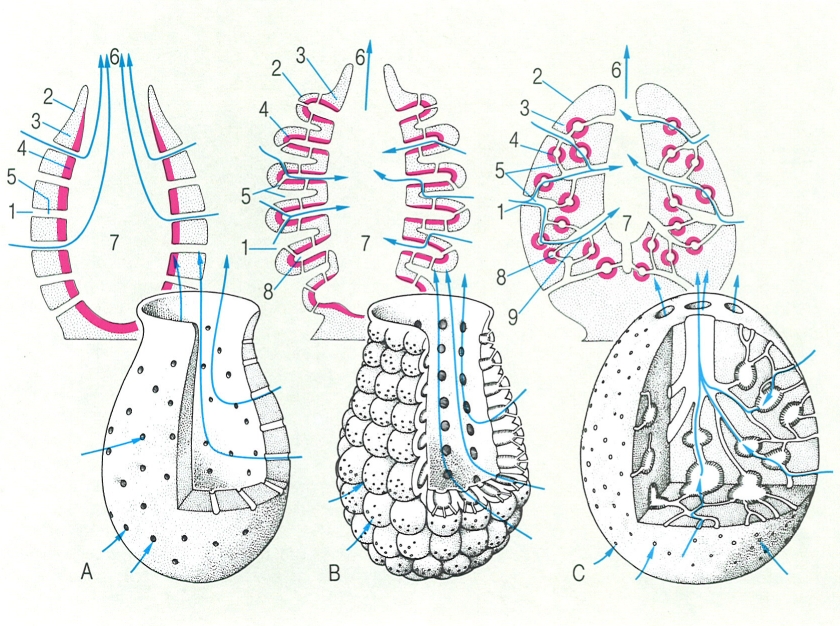
Sistemes aqüífers més senzills en les esponges: A àscon, B sícon i C lèucon. El sistema es complica extraordinàriament en la majoria de les demosponges, a partir d’algun d’aquests tres tipus bàsics. Hom ha indicat en el dibuix: 1 porus inhalant o ostíol, 2 ectosoma (pinacócits), 3 mesènquima, 4 endoderma (coanòcits, en vermell), 5 conducte eferent o inhalant (porócits), 6 óscul, 7 cavitat atrial, 8 cambra vibràtil, 9 canal exhalant.
Biopunt, a partir de fonts diverses
Hi ha tres tipus de sistemes aqüífers, coneguts tradicionalment amb la denominació d’àscon, sícon i lèucon. El tipus àscon consisteix en un cos en forma de sac o tubular, que presenta una doble paret; la de dins, formada pels coanòcits, delimita una cavitat central o atrial, que desemboca en un òscul; la de fora, o ectosoma, és formada per pinacòcits, i a la vegada és perforada pels ostíols, que comuniquen, mitjançant conductes, amb l’atri. Entre les dues capes hi ha una substància fonamental, el mesènquima, amb diferents elements cel·lulars. n’és un exemple típic Clathrina coriácea. En el tipus sícon, l’ectosoma recobreix, a més de la part externa, la cavitat atrial. Els coanòcits es troben situats dins divertides allargats que comuniquen amb l’exterior per diferents conductes aferents i amb la cavitat atrial per un de sol; podria servir d’exemple Sycon raphanus. Aquests dos primers tipus, àscon i sícon, es troben només en unes quantes espècies d’esponges calcàries. En la majoria dels demospongis adults, aquestes mateixes estructures es compliquen notablement: d’un sícon inicial que només es pot veure a la larva acabada de fixar i que es coneix amb el nom de ràgon, es desenvolupen complicats lèucons, amb diversos òsculs. El tipus ràgon correspon a un síon amb forma de con sense cmbres vibràtils a la part basal, que presenta un òscul a l’extrem d’una protuberància central. El tipus lèucon és el de màxima complexitat: els diverticles coanosòmics són globulars i es troben enfonsats dins el mesènquima, però es mantenen comunicats entre ells, amb l’exterior i amb la cavitat atrial, a través d’una munió de conductes aferents i eferents. Acanthella acuta n’és un bon exemple.
Els tipus cel·lulars

La diferenciació cel·lular no és molt gran en les esponges i pot ésser reversible. Les fotografies mostren diferents tipus de pinacòcits de la demosponja Dysidea avara: a dalt, pinacòcits allargats, al voltant d’un conducte aqüífer; a baix, endopinacòcits, flagel·lats, també entorn d’un conducte aqüífer.
Antoni Fauquet / M. Jesús Uriz
En aquests animals, la diferenciació cel·lular no és irreversible com en la resta dels metazous, i aquesta característica és molt important per a assolir la regeneració d’un individu complet a partir d’un fragment qualsevol. Presenten nombroses cèl·lules diferents, algunes amb funcions poc conegudes, de les quals citarem només les més importants. En primer lloc, els coanòcits són cèl·lules proveïdes d’un collar de microvellositats que envolta un flagel central, el moviment del qual indueix els corrents d’aigua; també intervenen en la captura de l’aliment i en la reproducció. El conjunt de coanòcits d’un individu rep el nom de coanosoma. D’altra banda, hi ha els pinacócits, cèl·lules planes que revesteixen l’exterior de l’esponja i els conductes aqüífers; formen una pel·lícula minsa anomenada ectosoma, que a vegades engloba espícules i pot reforçar-se amb col·lagen fins a esdevenir una capa gruixuda, que rep el nom de còrtex o escorça. Entre l’ectosoma i el coanosoma hi ha, dispersos dins el mesènquima, amebòcits (de funció fagocitària), espongioblasts (secretors de col·lagen), escleroblasts (secretors de l’esquelet corni, silici o calcari), miòcits (cèl·lules contràctils) i cèl·lules esferuloses (de funció excretora).

Tipus d’espícules en les demosponges: espícules dels homosclerofòrides (A), Megascleres (B), Microscleres (C) i Gemmoscleres (D). 1, 2 triactines, 3 diactina, 4 triactina monolofa, 5 triactina dilofa, 6 triactina trilofa, 7 canelobre, 8 oxa, 9 estròngil, 10 tilot, 11 estil, 12 estrongiloxa, 13 tilostil, 14 subtilostil, 15 caltropa, 16 protriena, 17 anatriena, 18 plagiotriena, 19 mesotriena, 20 ortotriena, 21 dicotriena, 22 metàster, 23 espiràster, 24 esferàster, 25 estrongilàster, 26 tilàster, 27 esteràster, 28 aspidàster, 29 sigma, 30 diancistre, 31 bipocil·lum, 32 toxa, 33 làbia, 34 amfidisc, 35 queia, 36 isoquela palmada, 37 anisoquela palmada, 38 queia ancorada espatulifera, 39 placoquela, 40 esferàncora, 41 microxa centrotilota, 42 trícodragma, 43 discorabda, 44 espinispira, 45 espira, D gemmoscleres d’aigua dolça.
Biopunt, a partir de fonts diverses

Tipus d’organització esquelètica en les esponges: 1 isodictial. 2 reticulada, 3 radial, 4 plomosa, 5 axial, 6 plumoreticulada.
Biopunt, a partir de fonts diverses
La morfologia de les espícules calcàries és poc variada; aquestes presenten dos, tres o quatre extrems més o menys esmolats, o actines, per la qual cosa hom diu que són diactinades, triactinades o tetractinades. Les espícules silícies, en canvi, presenten una gran varietat de formes i dimensions: n’hi ha des de 3 µm fins a d’alguns centímetres, i les podem separar en megascleres (generalment més grans de 100 µm) i microscleres (menys de 100 µm), encara que hi ha un petit grup, els homosclerofòrides, que presenten espícules de dimensions intermèdies); poden tenir des d’una fins a moltes actines.
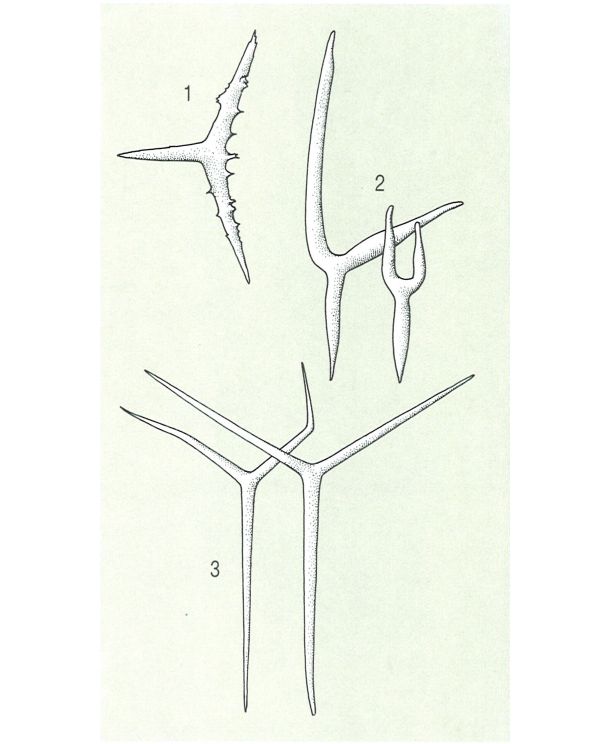
Tipus d’espícules calcàries més comunes: 1 espícula triactina espinosa, del gènere Plectronimia; 2 espícules triactines en forma de diapasó, característiques de certs feretrònides; 3 espícules triactines del gènere Clathrina.
Biopunt, a partir de fonts diverses
Les fibres d’espongina, de naturalesa proteica, formen un entramat que constitueix l’esquelet corni. Les fibres presenten una zona central, buida o plena d’una substància fonamental, que rep el nom de medul·la, i una part perifèrica, anomenada escorça, que pot ésser uniforme o en capes concèntriques (estratificada); poden englobar en el seu interior grans de sorra o restes d’espícules del sediment, i llavors hom parla de fibres empedrades o amb cossos estranys. Poden ésser dendrítiques o reticulades, i, en aquest darrer cas, se solen diferenciar les primàries (més gruixudes i amb direcció preferentment vertical) de les secundàries (transversals o obliqües). Algunes vegades aquestes fibres s’acompanyen d’espícules còrnies, com en el cas de Darwinella.

Espícula discorabda de l’espècie Latrunculia citharistae, típica de les esponges del seu grup, vista al microscopi electrònic d’escandallatge. La morfologia de les espícules és una de les bases de la classificació de les esponges.
José M. Fortuño / M. Jesús Uriz
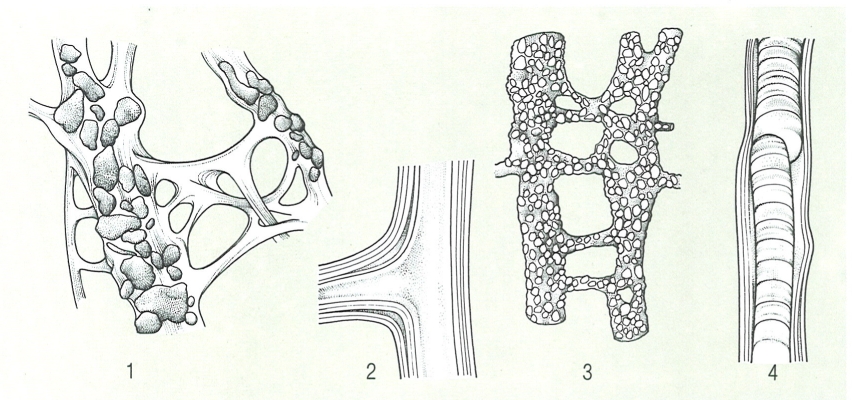
Diferents tipus de fibres d’espongina: 1 fibres primàries empedraries i secundàries, lliures de cossos estranys (lrcinia); 2 detall d’una fibra, amb l’escorça estratificada i la medul·la conspicua (Aplysina); 3 fibres primàries i secundàries totalment empedrades (Dysidea); 4 fibra amb l’escorça i la medul·la estratificades (Aplysilla).
Biopunt, a partir de fonts diverses
Ni les espícules ni les fibres no sempre estan disperses pel mesènquima, sinó que, freqüentment, formen estructures ordenades, molt variades, que es poden classificar en diferents tipus, encara que es trobin casos intermedis. En l’esquelet reticulat, les espícules o les fibres formen una xarxa de direccions i consistència variables (es parla d’esquelet renieroide o isodictial quan la malla és poligonal, més o menys uniforme en les tres direccions i formada per espícules). L’esquelet axial és format per un eix o un pla longitudinal espès d’on surten perpendicularment feixos espiculars que travessen la superfície de l’esponja. En l’esquelet plomós les espícules se situen d’una manera divergent, formant feixos ascendents. En l’esquelet dendrític els feixos o fibres d’espícules es distribueixen paral·lelament sense anastomosi. L’esquelet himedesmoide s’anomena així perquè és el típic del gènere Hymedesmia; es tracta d’una sola capa d’espícules perpendiculars al substrat. En el cas dels esquelets plumoreticulat i dendroreticulat, l’aspecte general és, respectivament, plomós i arborescent i, a més, presenta una reticulació més o menys marcada dins els feixos espiculars. L’esquelet halicondroide consisteix en un esquelet d’espícules desordenades, que no formen feixos ben definits, i és típic del gènere Halichondria.
Biologia i ecologia
La reproducció sexual i el comportament de la larva
Les esponges tenen reproducció sexual, encara que no s’hi observen gònades diferenciades. Els òvuls i els espermatozoides es troben dins el mesènquima i solen ésser el resultat de la transformació de les cambres vibràtils en oogònies o espermatogònies. La major part de les espècies són hermafrodites. Els elements de cada sexe solen madurar en èpoques diferents, per la qual cosa sembla que la fecundació és creuada. Els espermatozoides són alliberats al medi pels conductes exhalants; d’allà passen, a través dels ostíols, i juntament amb les partícules de nodriment a les cambres vibràtils d’un altre exemplar, on entren activament dins els coanòcits; poden romandrehi encistats un cert temps, fins que són injectats pel coanòcit en un òvul madur pròxim. Algunes espècies són ovípares, però la majoria incuben els ous fins en els estats larvals coneguts amb el nom d’amfiblàstula i parenquinel·la (semblant a la parenquímula d’alguns cnidaris). Les larves lliures són ciliades en tota la seva superfície, o en una part, però els desplaçaments que poden fer són limitats, per la qual cosa la dispersió és essencialment passiva. Al cap d’un temps de fer vida lliure, pelágica o, ocasionalment, bentònica, que pot variar des d’unes quantes hores fins a uns dies, les larves es fixen al substrat. En el cas de Demospongia, la larva s’aplana i dona lloc a una ràgon. En la major part dels casos, les larves presenten una certa fotofóbia: prefereixen els llocs ombrívols, i una llum intensa les indueix a seguir nedant. Necessiten substrats rugosos o pilosos per a fixar-se, encara que en el laboratori, amb l’aigua quieta, es poden establir perfectament al damunt de superfícies llises. Quan dues larves es fixen molt a prop l’una de l’altra, i en el curs del seu creixement arriben a tocar-se, pot passar que conflueixin o, al contrari, poden restar perfectament separades (fenomen de «front de no-confluència»); en general, solen confluir les larves que provenen d’un mateix progenitor.
La reproducció asexual

La preferència pels ambients il·luminats que mostra la demosponja Aplysina aerophoba és relativament exceptional dins el món de les esponges, que generalment cerquen llocs ombrius per establir-se. Aquesta espècie, de consistència flexible, és força comuna en les superfícies horitzontals de l’estatge infralitoral. Les prolongacions irregulars, que acaben per separar-se completament de l’esponja mare, són una variant de reproducció asexual típica d’aquesta espècie.
Antonio Cruz
Un cert nombre d’esponges produeixen gemmes capaces de transformar-se directament en adults. Aquestes gèmmules, hemisfèriques o esfèriques, formen una capa que resta en contacte amb el substrat i consisteixen en una massa de cèl·lules continguda dins una càpsula, amb espícules o sense. Quan la resta dels teixits moren, les gèmmules es desenvolupen i donen lloc directament a un petit adult. En el cas d’Haliclona losanoffi, les gèmmules queden adherides al voltant de les tiges de fanerògames marines, on l’adult ha crescut, de manera que quan les plantes es trenquen, se les emporten a d’altres llocs; tanmateix, la mortalitat d’aquests propàguls és molt elevada. Un altre tipus de gèmmula dona lloc a larves similars a les originades mitjançant la reproducció sexual. De fet, hi ha espècies, especialment a les zones intermareals, que poden produir larves sexuals i asexuals segons l’època l’any. Examinant aquestes esponges al llarg de diversos anys, s’ha observat que tenen cèl·lules sexuals (oòcits i espermatòcits) només durant una determinada època, mentre que tota la resta de l’any donen larves asexuals. També és freqüent la separació d’una part de l’adult com a propàgul; es coneix, per exemple, en els géneres Tethya, Aaptos i Thenea. Actualment no se sap gaire bé quins són els factors que propicien la reproducció asexual de les esponges. A les regions temperades, on els canvis estacionals de temperatura són notables, les gèmmules poden servir per a sobreviure durant l’estació freda. En la majoria d’esponges d’aigua dolça, les gèmmules fan el paper de cossos de resistència durant els períodes desfavorables.
Creixement i relacions tràfiques
La velocitat de creixement és molt variable segons les espècies. Les esponges calcàries i, en generai, les que viuen a l’espai intermareal i a les aigües dolces, presenten una alta taxa de renovació; al contrari, la resta d’espècies mostren un creixement lent i un temps de residència més llarg. En general presenten un creixement més ràpid i, per tant, una producció més alta, en els primers anys de vida; després, arriben a estabilitzar-se. Algunes, com Verongia arophoba, resisteixen bé la llum directa, encara que la majoria són esciàfil·les, és a dir, prefereixen els llocs de penombra de l’ecosistema litoral, especialment les parets rocoses protegides de la llum, l’entrada i l’interior de les cavitats submarines.
La respiració es realitza mitjançant l’absorció directa de l’oxigen dissolt en l’aigua; tant l’activitat respiratòria com la de bombeig de l’animal varien dins uns cicles diaris i anuals, rarament uniformes, i que, a més, depenen directament dels canvis de temperatura ambiental i de l’hidrodinamisme. L’excreció es fa en forma de grànuls refringents, que s’acumulen dins les cèl·lules esferuloses, que són abocades al sistema aqüífer exhalant; periòdicament també es fa l’expulsió de simbionts i cèl·lules somàtiques. La matèria orgànica alliberada per l’esponja en forma de mucus i substàncies solubles és poc coneguda.

La competència pel substrat entre diferents espècies d’esponges dona combinacions de colors bigarrades en el substrat dur litoral, que col·laboren en l’espectacularitat del paisalge submarí. En la fotografia, oblinguda en una paret del Túnel Llarg de les illes Medes (uns 20 m de profunditat), les tres tagues de color corresponen a una especie del gènere Crambe (de color vermell ataronjat). una del gènere Anchinoe (la de color blau grisós) i una tercera especie del gènere Crella (de color groc intens).
Josep M. Gili
L’alimentació natural de les esponges en el seu medi encara no és gaire ben coneguda. Tanmateix, els processos que tenen lloc per a la captura i la digestió de l’aliment han estat ben estudiats en condicions de laboratori. Subministrant partícules inerts o bacteris marcats amb un colorant fluorescent, i fent posteriorment talls histològics de les esponges, es pot veure quins tipus de cèl·lules participen en la captura, el transport i la digestió de les partícules alimentàries; les partícules més petites són retingudes pels coanòcits, mentre que els pinacòcits fagociten partícules de 5 a 50 µm, que entren pels porus, tot i que no poden travessar el petit orifici que comunica amb les cambres vibràtils. Entre les microvellositats del collar coanocitari es forma una substància mucosa o una xarxa fibril·lar capaç de retenir fins i tot els més petits col·loides. La digestió és intracel·lular, es fa en els mateixos coanòcits o els pinacòcits, o bé aquestes cèl·lules transfereixen el material alimentari als amebòcits pròxims, que són mòbils, i el porten fins a les cèl·lules internes.
La font principal d’aliment són partícules orgàniques submicroscòpiques, que són molt abundants a la mar. No obstant això, el règim alimentari de les esponges varia segons les substàncies que es troben a l’aigua, ja que es tracta d’organismes filtradors no selectius dins el marge de diàmetres dels ostíols. L’alta viabilitat del ritme de filtració entre els diferents exemplars de la mateixa espècie es relaciona no solament amb la mida sinó amb el nombre d’òsculs, el diàmetre d’aquests, i l’estat fisiològic de l’esponja: el volum d’aigua filtrat freqüentment és molt elevat, i està en relació tant amb la mateixa complexitat del sistema aqüífer com amb el cicle estacional. El percentatge de retenció en moltes esponges oscil·la entre el 68 i el 99%. Si aquesta retenció va acompanyada d’una concentració bacteriana alta o normal es pot considerar suficient per a cobrir les necessitats energètiques mínimes de l’animal i, si això no és així, l’esponja té capacitat per a capturar o absorbir altres tipus de matèria orgànica, fins i tot col·loïdal.
Es pot parlar d’una producció primària en les esponges deguda a l’associació amb organismes fotosintetitzadors: cianofícies, zooxantel·les i fins i tot fanerògames. Aquesta mena d’associació, encara que pot representar per a l’esponja uns avantatges fisiologies importants, no contribueix d’una manera significativa a la producció primària del bentos litoral mediterrani. Podria ésser que la situació hagués estat diferent en el Cambrià, on les esponges eren organismes constructors importants, però no es pot assegurar que tinguessin algues simbionts. La matèria orgànica produïda per les esponges torna al medi per diferents camins: l’alliberament directe, durant la vida de l’animal, en forma de mucus i d’elements reproductors, la biodescomposició una vegada mort, i el pas a nivells tròfics superiors (fonamentalment per depredació). Pel que fa a la degradació de l’esponja per biodescomposició cal assenyalar que les espècies amb una flora microbiana important no tenen una velocitat de descomposició superior a les altres, ja que aquesta la fan mitjançant bacteris que normalment no es troben en el seus teixits. Les fibres d’espongina són poc degradables, i romanen durant molt de temps al medi.
Són pocs els animals que s’alimenten d’esponges, especialment en les nostres mars. El seu esquelet d’espícules, i sovint la seva toxicitat, les fan poc mengívoles. Només se sap amb seguretat que serveixen d’aliment a mol·luscs, equinoderms i alguns peixos, i que la relació depredador-presa, en molts casos, és específica. Entre els mol·luscs, hi ha molts opistobranquis doridoïdeus i pleurobrancoïdeus brostejadors d’una sola espècie d’esponja, que són sorprenentment críptics quan són sobre la seva presa. Henricia sanguinolenta, entre d’altres equinoderms, és exclusivament espongiòfag. Cal assenyalar l’existència de peixos brostejadors d’esponges en zones tropicals; les poques espècies conegudes segueixen l’estratègia d’ingerir petites quantitats de diferents esponges i així eviten dosis perilloses de qualsevol toxina.

Les esponges perforants, com les de la família dels cliònids, ataquen qualsevol substrat calcari (algues, conquilles de mol·lusc), fins al punt que poden fer malbé bancs d’ostres o de coralls. En la fotografia veiem les masses verdoses perforants de Cliona viridis, damunt una alga calcària, blanca; és una espècie corrent en l’ambient infralitoral, sovint en àrees prou il·luminades.
M. Jesús Uriz
Un dels efectes més notables de les esponges en les comunitats bentòniques es manifesta en la competència pel substrat mitjançant la producció de metabòlits amb activitat citòtoxica: les esponges lluiten entre elles i amb d’altres organismes per ocupar i mantenir l’espai que ocupen, sigui impedint la install·lació de larves dels altres, sigui eliminant els adversaris. Unes quantes espècies que pertanyen a la família dels cliònids perforen i destrueixen qualsevol tipus de substrat calcari, i poden arribar a ésser perjudicials per als bancs d’ostres i coralls. D’altra banda, les esponges originen substrats nous, encara que aquesta activitat constructor és molt pobra en l’actualitat, en comparació amb el que esdevenia al Paleozoic, quan eren importants constructors d’esculls, amb els seus voluminosos esquelets calcaris (els estromatoporoïdeus), o a base de complicades xarxes silícies (litístides).
Filogènia i sistemàtica
Tradicionalment, i fins fa poc temps, horn ha considerat els porífers (Porifera) dividits en tres grans grups o classes, que se separen pel tipus d’esquelet. La classe dels calcaris (Calcarea), la dels demospongis (Demospongia) i la dels hexactinèl·lides (Hexactinellida); hom hi ha afegit, encara, la classe poc nombrosa dels esclerospongis (Sclerospongia), que alguns autors inclouen, com una subclasse, dins els demospongis. Estudis recents sobre la histologia d’algunes espècies d’hexactinèl·lides han posat de manifest que aquest grup té peculiaritats importants. En base a això, el fílum dels porífers ha estat subdividit en dos subfílums: el dels simplasmes (Symplasma) i el dels cel·lularis (Cellularia); fins i tot és dubtós que els porífers siguin un grup monofilètic, és a dir que tinguin tots un avantpassat comú i exclusiu de les esponges.
Els simplasmes comprenen únicament la classe dels hexactinèl·lides. Són porífers amb una organització senzilla, en forma de síncit, tot i que mostren la coexistència de tipus cel·lulars com arqueòcits i cèl·lules esferuloses. L’ectosoma no té pinacòcits diferenciats, però en el reduït mesènquima hi ha fibres de col·lagen i elements esquelètics i cel·lulars, bé que aquests darrers són poc nombrosos. La mesogiea és minsa, i no mostren cap mena de contractibilitat, és a dir que no tenen miòcits. Les cèl·lules flagel·lades formen un reticle anucleat que es comunica, per mitjà de ponts citoplasmàtics, amb coanoblasts nucleats. La simplicitat estructural dels simplasmes no es pot interpretar obligatòriament com a primitiva respecte de la resta dels porífers.
Els cel·lularis reuneixen les classes dels calcaris i els demospongis. Aquest gran grup d’esponges presenta una organització cel·lular definida: l’ectosoma format per pinacòcits i el coanosoma amb coanòcits individualitzats. El mesènquima, amb una matriu abundant de col·lagen, inclou elements esquelètics i diferents tipus cel·lulars ameboides. Les espícules no són mai hexaradiades.
Les esponges es troben entre els organismes més antics coneguts. Es coneixen fòssils des del Precambrià, i es pot dir que els diferents llinatges divergiren molt abans de l’Era Primària. Les espícules silícies fossilitzades, de vegades amb la sílice substituïda per altres substàncies químiques (calcita, glauconita, òxid de ferro, etc.) solen ésser tan abundants que arriben a formar roques (les espongiolites). La putrefacció, que té lloc abans de la fossilització, altera l’esquelet i fa que les espícules es dispersin. Tant sols els porífers amb espícules soldades, com els hexactinèl·lides, els faretrònides o els litístides, han conservat en estat fòssil la seva estructura esquelètica íntegra. Les esponges que són considerades més antigues són les hexactinèl·lides, que abundaren durant el Juràssic i el Cretaci. Al Carbonífer varen aparèixer les esponges calcàries i al Cambrià ho feren les demosponges monaxones. Però la filogènia de les esponges, generalment basada en la morfologia dels seus elements esquelètics, és poc coneguda; diferents teories han estat exposades sobre l’evolució dels porífers, sense que s’hagi arribat fins ara a cap acord.