Viure a la muntanya
Com en el cas del personatge de Hemingway que, en trobar-se a la neu, prop del cim del Kilimanjaro, una ròssa seca i gelada de lleopard es pregunta què havia pogut fer pujar tan amunt aquell animal, no sempre és fàcil explicar què busquen (o què han trobat) a la muntanya els animals que hi viuen. La majoria dels animals perceben les muntanyes com a barreres als seus desplaçaments; com a espais freds però amb ambients molt variats on generalment els recursos nutritius són escassos, tot i que ocasionalment poden resultar abundants. Les muntanyes limiten l’expansió d’algunes poblacions i espècies animals, mentre que n’afavoreixen l’expansió d’altres, i, en tot cas, gràcies a la seva relativa inaccessibilitat, han estat refugi on han pogut sobreviure alguns tàxons que s’han extingit a les planes.
Les adequacions al fred i a les oscil·lacions tèrmiques
Si bé a la muntanya troben principalment seguretat i aliment, els animals que hi viuen han de pagar el preu de romandre sotmesos a una duresa ambiental més gran, principalment pel que fa al fred. Ja s’han assenyalat abans els paral·lelismes (i les divergències) entre els ambients muntanyencs i els de tundra, principalment pel que fa al règim tèrmic. A les muntanyes, a diferència del que passa a la tundra, l’oscil·lació tèrmica diària és àmplia i permet cicles de glaç i desglaç. Això fa que els animals continuïn actius durant el període de l’any en què la temperatura mitjana diària es manté per sota dels 0°C a condició que la insolació diürna proporcioni, durant unes hores, temperatures més elevades. Per altra banda, gràcies a la proximitat de les zones de terra baixa de temperatures mitjanes més elevades, l’alta muntanya pot acollir una fauna molt més variada i complexa que no la tundra.
Les adaptacions dels animals muntanyencs al fred no es limiten a l’adquisició d’una capacitat de resistència enfront de les baixes temperatures hivernals. Fins i tot a l’estiu, les nits (i alguns dies) són freds a l’alta muntanya, de manera que els calen altres adaptacions, menys pronunciades, basades generalment en importants hàbits termoreguladors, per fer front a aquests freds extemporanis.
Una primera adaptació per a combatre el fred és aprofitar la radiació solar per a augmentar la temperatura corporal. La necessitat d’aprofitar la radiació solar durant el dia explica les diferències de pigmentació entre els animals homeoterms i els poiquiloterms a les muntanyes. Els primers adquireixen pelatges o plomatges hivernals, generalment blancs, que muden amb el canvi d’estació. Els segons, com que passen l’hivern en diapausa, opten per una pigmentació fosca que absorbeix més bé la radiació solar. El melanisme és, efectivament, una de les adaptacions més freqüents d’aquest ambient, ja que els pigments melànics no sols permeten l’absorció de la radiació tèrmica infraroja, sinó que actuen també de barrera per a la perniciosa radiació ultraviolada. Les papallones de muntanya, per exemple, durant el curt període reproductiu han de ser molt actives i, per tant, els seria molt nociu no poder gaudir de temperatures corporals elevades, de manera que presenten una coloració que enfosqueix proporcionalment amb l’altitud a la qual viuen. La salamandra negra dels Alps (Salamandra atra) és un altre exemple notable de melanisme muntanyenc, tot i que té altres adaptacions que tendeixen a escurçar el seu període d’activitat.
L’escurçament del període actiu és, en efecte, una de les estratègies adaptatives més esteses entre els animals de muntanya, ja que romandre actius durant l’estació freda representaria una despesa energètica massa elevada. Aprofitar la breu estació càlida pot fer-se, no únicament concentrant en un període breu totes les fases del cicle vital, sinó també fragmentant-lo i duent a terme les diferents fases en anys diferents. Aquesta darrera adaptació és freqüent tant en amfibis com en invertebrats. Així, per exemple, molts amfibis de muntanya creixen lentament i no finalitzen la seva metamorfosi durant el primer any de vida; romanen en estat larval almenys durant dos anys i arriben a assolir una mida més gran que les larves de la mateixa espècie que viuen a terra baixa. En altres casos, la fragmentació del cicle afecta els adults, que només es reprodueixen en anys alterns. Entre els insectes, molts caràbids passen diversos anys en fase de larva, apropant-se lentament al moment de la metamorfosi. L’estratègia oposada, la d’escurçar el cicle, es dóna, per exemple, en molts dípters, de vida tan curta, que, durant el breu estiu muntanyenc, poden succeir-se diverses generacions.
La presència de viviparisme en grups habitualment ovípars és una altra adaptació freqüent de les espècies muntanyenques. Un elevat percentatge d’amfibis i rèptils que viuen a les muntanyes esdevenen vivípars per tal d’escurçar el seu cicle anual i adaptar-lo a la brevetat de l’estació vegetativa. En algunes espècies l’adaptació és gradual i depèn de la severitat de les condicions ambientals. Així, la sargantana vivípara (Lacerta vivipara) —espècie estenoterma de l’Euràsia septentrional— és ovípara en el límit meridional de la seva àrea de distribució, als Pirineus i altres muntanyes de la Península Ibèrica, on l’estació càlida és més llarga. En algun cas, al viviparisme s’afegeix la supressió d’una part del cicle biològic, com en la salamandra negra dels Alps (Salamandra atra), espècie en la qual les femelles no és que pareixin cries vives, suprimint la fase d’ou, sinó que aquestes, en néixer, ja han sofert la metamorfosi i són petits adults. També són freqüents les modificacions de la conducta reproductora, i es pot arribar a la partenogènesi (reproducció sense necessitat de fecundació), tal com passa en alguns rèptils del Caucas, entre els quals Lacerta saxicola, i dels Andes. D’aquesta manera, el procés reproductor s’escurça i es fa més segur, però la flexibilitat genètica disminueix i l’espècie podria arribar fins i tot a l’extinció davant canvis ambientals molt pronunciats.
Hi ha espècies que modifiquen i adeqüen el seu interval de temperatures preferents en funció de l’interval de temperatures ambientals. Molts insectes d’alta muntanya poden suportar temperatures molt baixes (n’hi ha que poden resistir temperatures properes al zero absolut) i fins i tot les requereixen per a sobreviure. Algunes papallones que viuen en molleres no poden sobreviure si la temperatura mitjana del mes de gener roman per damunt del punt de congelació. Per a altres insectes, el factor limitant és la mitjana de temperatures del mes més càlid, que no ha de superar els 11 o 12°C. A l’estatge alpí de les muntanyes europees viuen llagostes que no suporten temperatures mitjanes que sobrepassin el llindar dels 8°C durant l’estiu; els insectes notòpters de les Muntanyes Rocalloses del Canadà viuen sobre la neu i se’ls ha de mantenir en frigorífics quan es volen conservar vius en captivitat. Les petites sargantanes de muntanya tenen una resistència a les baixes temperatures molt superior als seus congèneres de les planes i només perden la capacitat de moviment a temperatures inferiors al punt de congelació.
Els canvis adaptatius com a resposta al fred a vegades són més subtils, simples modificacions fisiològiques o etològiques poc evidents. La capacitat de controlar la posició del pèl o de les plomes per tal d’augmentar la capa d’aire aïllant que està en contacte amb la pell, la capacitat de regulació de la pèrdua de calor per evaporació, la millora de l’aïllament tèrmic de les extremitats o la modificació del metabolisme basal, són exemples típics d’aquest tipus d’adaptacions fisiològiques, però menys aparents que el melanisme o la modificació del cicle biològic. Entre els canvis etològics, que són sempre els més immediats i els més senzills en qualsevol procés d’adaptació, destaquen la recerca i la construcció de refugis subterranis, la migració a altituds inferiors o l’ús, durant l’hivern, de les reserves nutritives acumulades al cos en forma de greixos o en els dipòsits d’aliments.
Algunes adaptacions no es relacionen directament amb el fred, sinó que tenen a veure amb alguna de les seves conseqüències com, per exemple, la innivació. És el cas dels recobriments de pèls i plomes del voltant de les potes que tenen diverses espècies muntanyenques per a protegir-se de la neu i dels quals estan absents els seus congèneres de terra baixa. També s’observa aquesta adaptació en els pelatges i les plomes hivernals d’alguns dels animals, com l’ermini (Mustela erminea) o la perdiu blanca (Lagopus mutus), que, a més de ser espessos i tenir una gran capacitat aïllant, són blancs com la neu, cosa que els facilita passar inadvertits per als seus depredadors. Finalment, la neu pot donar lloc a adaptacions conductuals que serveixen a l’animal per a defensar-se del fred: la gasela tibetana o goa (Procapra picticaudata), per exemple, es protegeix dels torbs excavant una cavitat a la coberta de neu i arrecerant-se darrere de la neu amuntegada.
Molts animals de muntanya presenten també trets que semblen adaptats a unes condicions d’àmplia oscil·lació tèrmica. En alguns, les fluctuacions tèrmiques extremes els obliguen a tenir una conducta fixada per a iniciar l’activitat abans que l’ambient esdevingui massa calent, i retirar-se al refugi al capvespre, quan la temperatura disminueix. Els dies asolellats, molts animals aconsegueixen temperatures internes elevades utilitzant les pedres com a radiadors, ja que s’escalfen fàcilment amb la radiació solar. A les muntanyes tropicals s’han mesurat diferències tèrmiques a la superfície de pedres fosques que arriben fins a 85 o 90°C entre el migdia i la nit. Altres espècies, per contra, romanen en ambients que tenen la temperatura poc variable, ja sigui sota les pedres, soterrades, a l’interior de les mates herbàcies o al voltant de les congestes permanents, on la neu manté fresc l’aire circumdant. De fet, a totes les muntanyes elevades existeix una microfauna nival característica, que no es veu afectada per l’amplitud de l’oscil·lació tèrmica de l’aire.
Les adequacions al vent
Les adaptacions al vent, que, generalment per causes topogràfiques, bufa de manera assídua i intensa a les muntanyes, són menys conegudes. Els animals, durant l’hivern, es protegeixen del vent, emparant-se darrere de les roques i les parets a manera de pantalles mentre que, a ple estiu, el fan servir per a refrescar-se i per a eludir l’atac dels insectes. La capacitat dels animals de muntanya per a romandre immòbils a despit del vent és molt superior a la dels de terra baixa. L’isard (Rupicapra rupicapra) pot romandre immòbil durant hores enmig d’un torb i amb temperatures inferiors a –10°C. Aquesta resistència és possible gràcies a la coberta de pèl o ploma, que, per la posició triada per l’animal en relació amb la direcció del vent, cobreix les parts més exposades al torb: així, mentre que la majoria dels animals de muntanya avancen de cara al vent, altres, com el iac (Bosphagus grunniens), li donen l’esquena per protegir-se el musell nu amb l’espessa llana de les seves gropes.
Tanmateix, les adaptacions al vent més evidents són les que afecten els grups voladors, principalment els insectes: volar enmig d’un vent fort és arriscat, per això moltes formes muntanyenques d’insectes han esdevingut àpteres. A l’Himàlaia, la meitat dels insectes que viuen per damunt del límit del bosc han perdut la facultat de volar i, entre els que viuen prop del límit de la neu, la proporció d’espècies no voladores arriba al 60%. En molts gèneres de llagostes que inclouen espècies que viuen a diferents altituds es constata que les espècies que habiten a cotes més elevades tenen les ales més petites que les espècies de terra baixa i poden, fins i tot, haver-les perdut. Fins i tot, els insectes muntanyencs voladors acostumen a fer vols curts i de poca distància de terra per evitar que el vent els arrossegui. Contràriament al cas dels insectes, el vent és un recurs important per als desplaçaments dels grans rapinyaires que, a més de volar sense problemes, aprofiten els corrents constants per a fer vols sostinguts.
Les adequacions a l’escassesa d’oxigen
Entre les adaptacions fisiològiques a l’alta muntanya, probablement les més conegudes són les derivades de la baixa pressió atmosfèrica i, per tant, de la baixa pressió parcial d’oxigen. En els animals, les modificacions per a resoldre els problemes derivats de la hipòxia es troben fixades als gens i són, per tant, veritables adaptacions.
Els mamífers de muntanya, per exemple, presenten adaptacions, tant anatòmiques com fisiològiques: els pulmons, el cor i el volum sanguini són més grans, els eritròcits són més petits i més nombrosos, la disponibilitat d’hemoglobina és molt superior i amb una afinitat per l’oxigen més gran que la dels animals de les zones baixes. Aquestes adaptacions es manifesten molt clarament en diferents espècies d’un mateix gènere que viuen a diferents altituds. Al Caucas, per exemple, hi ha cinc espècies de talpons (Microtus) que tenen preferències diferents pel que fa a l’estatge altitudinal en el qual viuen; la grandària dels eritròcits respectius varia en relació inversa amb l’altitud de l’estatge que ocupen preferentment .
La mobilitat en relleixos i tarteres
Els espadats, les gleres i les tarteres són, possiblement, les zones menys acollidores de les muntanyes, i tan sols espècies voladores o dotades d’una extraordinària agilitat s’hi mouen amb facilitat, cosa que els aporta grans avantatges. En efecte, a les cornises i relleixos de les parets de roca hi pot créixer herba d’un gran valor nutritiu, cosa que, unida a la dificultat d’accés per als depredadors, en fa llocs d’elecció perquè molts herbívors s’hi puguin alimentar. Fins i tot quan la superfície és molt reduïda, pot servir de base per a la construcció dels nius als grans rapinyaires. No és estrany que l’àguila daurada (Aquila chrysaetos), el voltor (Gyps fulvus) o el trencalòs (Gypaetus barbatus) niïn habitualment en petits replans de les parets rocalloses.
Les gleres i les tarteres són refugis molt adients per a diferents grups d’animals, ja que els espais que queden entre les pedres formen una xarxa de túnels on troben aixopluc. Molts d’ells han perdut la capacitat d’excavar que tenen els seus congèneres de terra baixa, com el talpó de les neus (Microtus nivalis), que és excavador quan viu en zones on el sòl és ben desenvolupat, i deixa de ser-ho quan viu als tarters; la marmota europea (Marmota marmota) excava els seus caus als Alps, mentre que als Pirineus, on fou reintroduïda fa poc temps, acostuma a viure en zones d’acumulació de blocs. Les piques (Ochotona) són també animals típics dels tarters i fan el niu entre les pedres de les gleres de les muntanyes d’Àsia i de l’Amèrica del Nord.
En els espais intersticials de les gleres, a més dels animals que hi cerquen refugi, hi viuen comunitats d’animals esciòfils, que són aquells que requereixen condicions relativament constants de temperatura i d’humitat i que no resistirien valors extrems de llum i fred de l’exterior. Aquestes comunitats són constituïdes principalment per insectes àpters i aràcnids, emparentats ecològicament amb els que configuren la fauna de les coves. Les pedres tenen un paper fonamental per a aconseguir un ambient on les condicions tinguin poca variabilitat, ja que el seu paper amortidor, tant de la temperatura com de la humitat, és evident: a una hora determinada sota les pedres, la temperatura diürna no supera els 14°C, mentre que la temperatura de l’aire a l’exterior es manté al voltant de 10°C i la superfície externa de la roca s’escalfa fins a 32°C. Anàlogament, durant la nit la temperatura sota les pedres pot ser de 4°C, mentre que a la superfície externa o a l’aire exterior davalla per sota dels 0°C; la humitat relativa sota les pedres es manté elevada i amb poques variacions, entre el 75 i el 90%, tot i que a l’exterior pot oscil·lar entre el 20 i el 80%.
La dinàmica i les interaccions faunístiques
El ritme de la vida animal a la muntanya el marquen diferents “rellotges”, però, com que la temperatura és el factor ambiental dominant, els ritmes biològics se sincronitzen amb els tèrmics, de manera que les comunitats animals ajusten els seus canvis als cicles dia-nit o als cicles anuals. A aquests cicles se superposen els derivats de l’activitat reproductora, quan les generacions són massa breus per a ajustar-se a cicles anuals, o els de període més llarg, com els associats a les successions i als canvis en les comunitats animals o a la distribució dels estatges de vegetació.
Els cicles circadiaris
Els cicles diaris són els més evidents i, a les muntanyes tropicals, gairebé els únics perceptibles. Moltes espècies d’insectes i d’invertebrats de muntanya ajusten el seu horari d’activitat per a poder aprofitar les hores de temperatures més adequades i amagar-se sota les pedres o dins les esquerdes quan l’ambient és massa fred.
A muntanya, els micromamífers subterranis i els rèptils surten dels seus caus a trenc d’alba, quan el sol comença a escalfar i els permet refer-se de les baixes temperatures nocturnes; unes hores més tard, quan es desperten els depredadors, aquells ja han assolit una temperatura adequada, i romanen segurs amagats dins els caus. Durant les hores centrals del dia, quan la temperatura és molt elevada, l’activitat es restringeix al mínim per evitar pèrdues innecesàries d’aigua corporal, i no és fins més tard, quan la calor no és tan intensa, que es pot donar un període secundari d’activitat. Els refugis poden ser esquerdes a la roca viva, espais sota les pedres o entre els blocs dels tarters i zones protegides per la vegetació. La rata de muntanya de Kenya (Otomys orestes), per exemple, es refugia durant la nit sota les fulles seques de les mates de senecis arborescents (Senecio) i, a vegades, dintre d’una cavitat que excava en el tronc d’aquestes plantes.
Els ritmes diaris de les plantes en cauliròsula (els “frailejones” americans o els senecis i lobèlies arborescents africans que presenten una roseta de fulles al capdamunt d’una tija de port arbori), moltes de les quals obren les fulles verdes durant el dia i les tanquen durant la nit, ha dut diverses espècies d’insectes a aprofitar aquest refugi nocturn, adequant la seva activitat al ritme de la planta, de la qual, en un notable exemple de coevolució, també s’alimenten. Naturalment, els animals més dependents de la temperatura exterior, com els rèptils i els insectes, són els que adopten ritmes d’activitat més marcats. Els primers, gràcies a la seva elevada capacitat calorífica, sovint complementada amb pigmentacions fosques, mantenen temperatures més elevades que la de l’aire, però amb una variació paral·lela a la d’aquest. Així, les sargantanes del gènere Liolaemus, als Andes, modifiquen la temperatura interna seguint un ritme diari que coincideix amb els màxims i els mínims de la temperatura ambient, però mantenint-se sempre de 8 a 20°C per damunt d’aquesta.
No és estrany que els animals aprofitin els efectes indirectes del cicle diari com la variació de temperatura i de llum, la rosada matinal o les tempestes vespertines dels dies d’estiu per a fugir de la calor o del fred excessiu, dels insectes molestos i dels enemics, i, fins i tot, per a aconseguir aliment. Les variacions circadiàries del règim de brises de les muntanyes s’utilitzen també per a organitzar l’activitat diària i, especialment, els desplaçaments. Durant el migdia estiuenc els ungulats muntanyencs eviten la calor excessiva situant-se a llocs ombrívols, on se’ls veu sempre reposar a les mateixes hores; altres vegades, s’exposen al vent de les carenes amb la mateixa finalitat o bé amb la de deslliurar-se dels insectes. Els grans rapinyaires utilitzen els corrents tèrmics ascendents que bufen regularment cada dia per a assolir la zona més alta dels cims.
Els cicles anuals
Els cicles amb un període superior a les 24 hores, però inferior a l’any, són conseqüència d’una demografia oscil·lant. Es donen, per exemple, entre els rosegadors muntanyencs, encara que als ambients de muntanya són menys comuns que a terra baixa, tant perquè les extremes condicions ambientals imposen taxes de reproducció i creixement moderades com perquè la intensitat que assoleix la variació estiu-hivern obliga a la sincronització del cicle reproductiu amb el cicle anual. Per això, després del cicle diari, el principal regulador de la vida animal a la muntanya és el cicle anual.
Tot i la seva importància intrínseca, no és el fred la més important de les limitacions que imposa la muntanya als animals. Encara ho és més la migradesa de la vegetació (sobretot durant l’hivern), i la manca d’aliment que se’n deriva. Així, per exemple, el període de naixement de les cries cal que estigui sincronitzat amb la successió climàtica, ja que ha de coincidir amb l’època de l’any tèrmicament més favorable. La curta durada d’aquest període favorable condiciona que moltes espècies comencin l’estació reproductora molt aviat, quan el sòl és encara cobert de neu i la temperatura de l’aire és inferior fins i tot a aquella que, durant la tardor, les indueix a retirar-se al refugi hivernal.
Per a molts animals, la manera més còmoda de sobreviure a les dures condicions de l’hivern muntanyenc és fugir-ne. A les muntanyes, les migracions per cercar zones amb temperatures més suaus són relativament fàcils, ja que no cal fer grans desplaçaments per a aconseguir-ho. Baixar a nivells inferiors és a l’abast, no sols dels ocells, sinó també dels grans mamífers i de molts insectes voladors. Els desplaçaments hivernals tenen lloc amb l’arribada de les primeres neus; els isards (Rupicapra), els muflons, els urials i els argalís (Ovis), els cèrvids (Hippocamelus, Moschus, Pudu) i els porcs senglars (Sus scrofa), davallen fins als boscos situats en zones inferiors més benignes, tot i que, a vegades, es limiten a traslladar-se als vessants solans, ja que en ambdós paratges les condicions ambientals són molt similars.
Al començament de l’estiu, la muntanya és envaïda per nombrosos animals que volen aprofitar l’estació càlida. Les migracions circumscrites a la muntanya són graduals: els animals no assoleixen els cims en un sol desplaçament, sinó que es desplacen encalçant la neu que es retira, passant d’un estatge de vegetació al superior quan les condicions tèrmiques ho permeten, i reculant si el temps empitjora. A poc a poc arriben a les pastures supraforestals. Els primers a fer-ho són els ocells, tot i que durant la nit encara s’han de refugiar al bosc. Més tard, ben entrat l’estiu, arriben els insectes, a qui la davallada nocturna de la temperatura obliga a resguardar-se sota les pedres. Els grans mamífers també pugen les muntanyes en cerca d’aliment: els ungulats segueixen la neu que es retira, tot pasturant els suculents rebrots que neixen al territori descobert, mentre que els carnívors van a la recerca de ratolins i marmotes, que es desperten al punt que fon la neu. Els ocells migradors que segueixen la ruta de les muntanyes travessen els ports quan els cims ja han perdut la catifa de neu; normalment porten un cert retard respecte als que sobrevolen la mar, ja que han d’esperar fins que el clima els permeti creuar els passos més alts.
Altres animals, com els amfibis o els petits mamífers, també realitzen desplaçaments estacionals, en aquest cas més aviat relacionats amb la reproducció, bé que també amb la insuficiència d’aliment. Molts amfibis, per exemple, presenten dispersions que afecten un sol sexe o, si els afecten tots dos, de manera diferent en cada un: abans de la reproducció, mascles i femelles s’apleguen a les basses, sovint transitòries i originades per la fusió de la neu. Després de la reproducció, els mascles (i, en algunes espècies, també les femelles) abandonen el lloc de posta i es dispersen, evitant així competir per l’aliment amb la seva nombrosa descendència. Aquest mateix procés succeeix amb els petits mamífers, per als quals la dispersió postreproductora és un dels mitjans utilitzats per a evitar exhaurir els recursos en un lloc determinat. Aquestes migracions es produeixen en espècies d’elevada taxa de reproducció, ja que en cas contrari, la seva població creixeria de manera desfermada en indrets molt concrets.
El recurs de la hibernació
Per a enfrontar-se al fred de l’estació hivernal, els animals tenen també la possibilitat de la hibernació. Segons la severitat de l’hivern, aquesta serà més o menys intensa. Així, en zones d’hiverns moderats molts animals experimenten una mena d’ensopiment, fàcilment reversible quan la temperatura augmenta; llavors no es pot parlar de veritable hibernació, ja que aquesta és un procés fisiològic complex i durable, que no és susceptible d’ésser suspès per augments ocasionals de la temperatura. A més dels rats-penats, afecta unes quantes dotzenes de mamífers de latituds temperades, de les quals la marmota és, sens dubte, la més coneguda.
El factor desencadenant de la hibernació de la marmota (Marmota marmota) és l’escurçament del fotoperíode durant la tardor, cosa que provoca una resposta hormonal complexa encaminada a reduir l’activitat metabòlica general de l’organisme. L’animal es retira al seu cau, que prèviament ha condicionat amb herba i que és bastant profund, per aconseguir que la temperatura s’hi mantingui constant i sempre superior a 0°C. Mentre dorm, la temperatura corporal disminueix uns 30°C, el ritme cardíac s’alenteix almenys cinc vegades, i el respiratori ho fa com a mínim deu cops. L’animal experimenta altres canvis fisiològics, com és la modificació de la composició de la sang i el consum de les reserves de greix acumulades durant l’estiu. El son no és continuat; s’interromp de tant en tant per eliminar els productes nocius acumulats a l’organisme. Amb la primavera, el rellotge endogen de la marmota posa en marxa tot el mecanisme de recuperació de l’activitat normal. L’animal, quan es desperta i surt a l’exterior, ha perdut més de la quarta part del seu pes pre-hivernal.
Els animals poiquiloterms o de “sang freda” tenen una dependència evident de la temperatura ambiental. Els seus processos metabòlics depenen directament de la temperatura exterior: si fa fred, aquests processos són més lents, i les successives fases del cicle anual es retarden i, contràriament, quan fa calor, s’acceleren. Aquesta dependència directa de la temperatura permet considerar els animals poiquiloterms com a acumuladors de calor, que assoleixen una certa activitat quan absorbeixen un nombre prefixat de calories. Així, com que l’important és el nombre total de calories absorbides, els és indiferent obtenir-ne en uns pocs dies molt calorosos o bé durant molts dies de temperatura moderada. Simplificant una relació de dependència molt més complexa, es podria dir que si una espècie completa la seva fase vegetativa en quatre mesos quan es troba a temperatures ambientals de 7°C, només trigaria dos mesos, si la temperatura mitjana fos de 14°C, i tan sols un, si fos de 28°C.
Gràcies a la necessitat que tenen els animals poiquiloterms d’acumular un determinat nombre de calories per a poder ser actius, es pot entendre la gran diferència que hi ha en el nombre de poiquiloterms que viuen a la muntanya i a les regions de clima fred però poc variable. A la Terra del Foc, per exemple, on la temperatura mitjana anual és d’uns 7°C, amb poca variació entre estiu i hivern, pocs animals arriben a acumular prou calories per a completar el seu cicle anual, i per això el nombre d’animals poiquiloterms és escàs. En canvi, a moltes zones de muntanya, la temperatura mitjana anual és molt baixa, inferior a 0°C, però hi ha una forta estacionalitat i els dies d’estiu són calorosos, amb temperatures mitjanes superiors a 12 o 14°C, cosa que permet, durant la breu estació estival, acumular prou calories perquè el cicle anual pugui completar-se. Per això els insectes poden envair aquestes zones a l’estiu, tot i que l’hivern l’han de passar en diapausa.
La diversitat dels nínxols ecològics
La producció vegetal de l’alta muntanya contrasta amb la de la tundra. Així com durant l’estiu àrtic les plantes aprofiten la llum gairebé permanent i les temperatures no gaire fredes per a produir matèria orgànica de manera continuada, a la muntanya, la manca de llum durant la nit es compensa amb una temperatura més elevada durant el dia. El resultat és que, tant la biomassa com la producció vegetal de l’alta muntanya alpina i de la tundra àrtica són similars (de 500 a 1 000 g/m2 com a mitjana anual, tant a la tundra àrtica com a l’estatge alpí). Els herbívors de muntanya, però, cerquen la qualitat més que no pas la quantitat, s’estimen més els rebrots tendres de les plantes del començament de la primavera que les tiges endurides i resseques del final de l’estiu. Per això, tant els residents permanents com els migratoris, segueixen la retirada de la neu per tal de menjar les herbes acabades de brotar. Així, a ple estiu assoleixen les cotes més altes, just per sota de l’estatge nival.
A la muntanya, els carnívors s’especialitzen en els principals tipus de presa disponibles: ungulats, rèptils, rosegadors i insectes. Els ungulats són peces importants, però poc accessibles (llevat de les cries o quan, un cop morts, serveixen d’aliment per als carronyers). Malgrat la dificultat de caçar-los, atreuen els grans carnívors de zones més baixes. Així, el puma (Felis concolor), el lleopard (Panthera pardus) i l’especialitzada pantera de les neus (P. uncia) poden caçar, fins i tot, al nivell de les neus perpètues. Els petits carnívors, com molts rapinyaires petites o mitjanes i algun rèptil, s’alimenten preferentment de rosegadors, que compensen la petita mida amb una gran abundor; dins el grup de petits carnívors es troben caçadors especialitzats, com l’ermini (Mustela erminea), de cos prim i flexible que li permet penetrar a les galeries dels rosegadors subterranis. Els insectívors no solen ser residents permanents de la muntanya, sinó visitants estivals que aprofiten una proliferació puntual d’insectes. Tan sols alguns insectívors (caràbids, aranyes i altres petits invertebrats) que, per les seves petites dimensions, poden trobar suficient nombre de preses fora de les èpoques de proliferació, viuen permanentment a la muntanya.
Llevat dels ocells carronyaires, els animals descomponedors són menys abundants a la muntanya que en biomes on la producció de matèria orgànica és molt més alta. El fred limita la producció i també frena la velocitat de descomposició, especialment en els sòls àcids, pobres, inundats i rocallosos típics de les muntanyes. Aquesta baixa velocitat de descomposició implica la formació de torberes i zones entollades on s’acumulen les restes vegetal sense descompondre. La fauna edàfica és, però, abundant i variada: en un litre de sòl alpí, per exemple, s’hi han trobat entre 500 i 2 500 insectes edàfics, de 500 a 3 000 àcars, fins a 1 800 rotífers, uns 3 000 tardígrads, 18 000 nematodes i de 500 000 a 1 200 000 protoctists heteròtrofs.
Les xarxes tròfiques
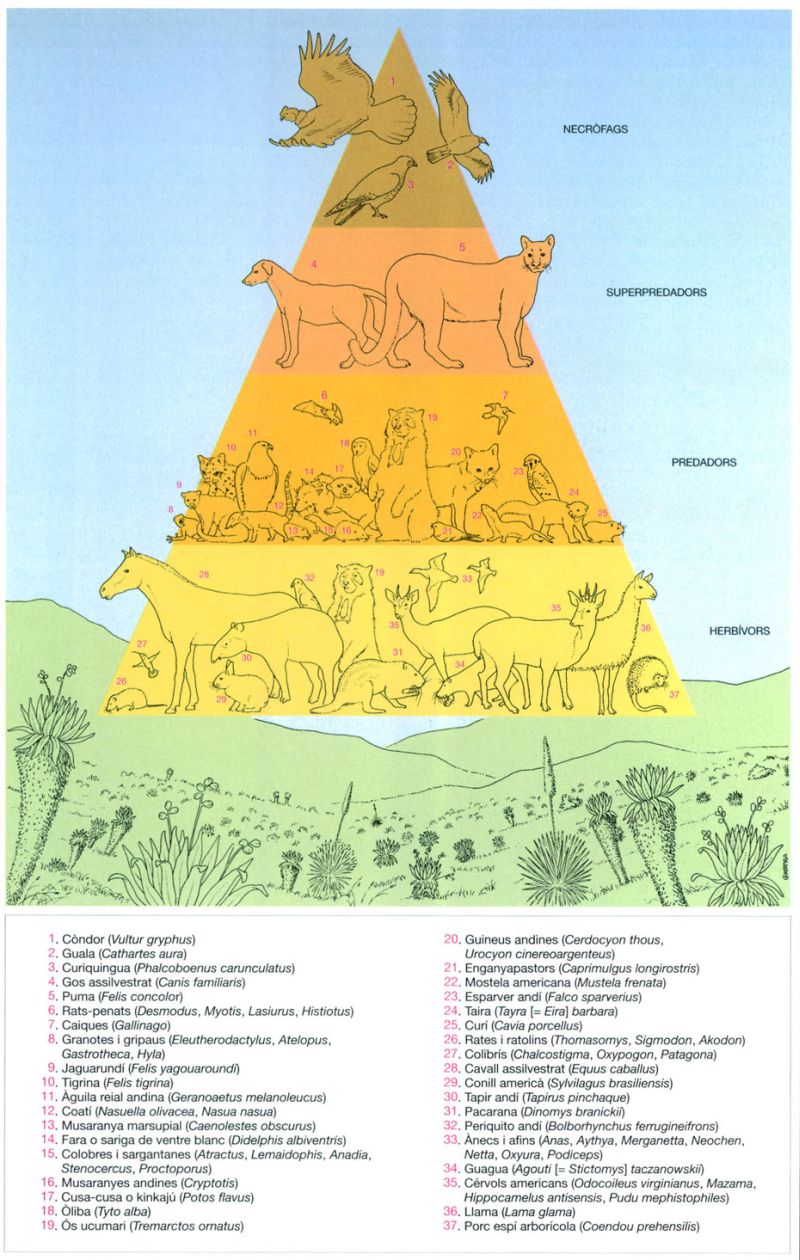
La piràmide tròfica del “páramo” andí dona exemple de la simplicitat de les xarxes tròfiques de les regions muntanyoses, en les quals intervenen un nombre relativament baix d’espècies: tres ocells necròfags, un sol gran depredador natural —el puma (Felis concolor), ja que els gossos (Canis familiaris) foren introduïts pels humans— i un nombre més elevat de petits depredadors i fitòfags.
Jordi Corbera, a partir de Del Llano, 1990
A l’alta muntanya, la riquesa específica és molt baixa i les xarxes tròfiques són formades per pocs elements. Un dels exemples més representatius és el d’un estudi fet a l’estatge alpí de les Muntanyes Rocalloses (a una altitud d’entre 3 300 i 3 800 m). Hom identificà tretze espècies de mamífers herbívors, tres musaranyes, sis espècies d’ocells herbívors i insectívors que nidifiquen habitualment a la zona, però que emigren durant l’hivern, vuit espècies de mamífers carnívors i tantes més d’ocells rapinyaires: en total, unes quaranta espècies de vertebrats terrestres amb més de set-centes interaccions possibles. L’estudi destacà que els carnívors més eurífags eren els rapinyaires, que cacen qualsevol animal que tingui una mida adient, als quals cal afegir el golut (Gulo gulo) i la guineu (Vulpes vulpes) espècies que, malgrat ser preferentment forestals, cacen també als rasos alpins i són molt poc selectives. Els petits depredadors no poden capturar les preses de dimensions mitjanes, com per exemple la marmota de ventre groc (Marmota flaviventris), mentre que les grans, com la cabra de les Rocalloses (Oreamnos americanus) o el cérvol (Cervus elaphus) només són accessibles al golut i a l’ós bru (Ursus arctos). Els herbívors tenen una alimentació molt selectiva: la majoria dels rosegadors subterranis consumeixen fulles i òrgans vegetals de reserva, com arrels i bulbs. El conill (Sylvilagus) i els esquirols de terra (Spermophilus) són folívors, és a dir, s’alimenten de fulles, i la dieta de les piques (Ochotona) es basa principalment en fruits i llavors.
Als Alps i als Pirineus les xarxes tròfiques són bastant semblants. Els talpons (Microtus) ocupen el lloc dels rosegadors subterranis de les Muntanyes Rocalloses, les marmotes (Marmota marmota) el de les seves congèneres americanes, i hi manquen els esquirols de terra (Spermophilus). Els depredadors són poc variats, amb predomini de la guineu (Vulpes vulpes) i dels petits carnívors. L’ós pràcticament s’ha extingit, i l’únic gran omnívor és el porc senglar (Sus scrofa). Falta un superdepredador com el golut.
A l’Amèrica del Sud, els “tucotucos” (Ctenomys) són els rosegadors subterranis, les xinxilles (Chinchilla laniger) són equivalents a les piques (Ochotona) i el principal ungulat és el guanac (Lama guanicoe). L’únic gran depredador és el puma (Felis concolor).
A l’Àfrica, els homòlegs dels ocotònids són els hiracoïdeus o damans (Procavia) i el caçador més important és el lleopard (Panthera pardus). Finalment, a l’Himàlaia existeixen nombroses espècies de rosegadors subterranis, que n’inclouen alguns de mida intermèdia com les marmotes (Marmota caudata), hi ha nombrosos ungulats i dos grans depredadors, l’ós tibetà (Ursus [=Selenarctos] thibetanus) i la pantera de les neus (Panthera uncia).