Aigües fredes i poc mineralitzades
Fora de les zones de neu permanent, l’alta muntanya és sovint un paratge ric en ambients aquàtics de notable singularitat. A més de l’abundant precipitació en forma de pluja o de neu per l’efecte del relleu sobre la circulació de les masses d’aire, els tipus més freqüents de roques (generalment ígnies, metamòrfiques o magmàtiques) són prou impermeables i resistents a l’acció erosiva de l’aigua. Així, l’aigua hi circula superficialment en forma de torrents i rierols i s’acumula a les concavitats i a les valls, donant lloc a la formació d’estanys.
L’origen i la conformació dels estanys i dels aiguals
Els estanys d’alta muntanya sovint deuen el seu origen al vulcanisme o als efectes modeladors de les glaciacions del Plistocè, l’agent que més llacs actuals ha produït arreu del món, en alguns casos remodelant cubetes ja existents. Alguns estanys encara avui són periglacials, i reben la influència directa de les aigües fredes, i normalment tèrboles, d’una glacera. Hi ha estanys a les muntanyes d’Amèrica del Nord (Rocalloses, Sierra Nevada, Apalatxes), Amèrica del Sud (Andes), Euràsia (muntanyes d’Escandinàvia, Pirineus, Alps, Tatra, Caucas, Pamir, Hindu Kush, Karakoram, Himàlaia), Àfrica (Kenya, Ruwenzori, Kilimanjaro) i Oceania (Alps de Nova Zelanda, serralades centrals de Nova Guinea). La importància dels estanys en el paisatge i la vida de muntanya ha fet que tinguin localment o regionalment noms particulars, diferents del terme general de llac.
L’altitud on se situen els estanys de muntanya varia molt i depèn de la latitud i la continentalitat. Van d’un centenar de metres a les altes latituds, a més de tres mil a les regions tropicals. Els llacs de més altitud del món són probablement alguns de l’Àsia central situats cap als 5 400 m. Normalment es troben situats per sobre del límit altitudinal del bosc i ben sovint en paratges de poca vegetació, quasi desèrtics en alguns casos. Aquest fet comporta que les aigües siguin més aviat pobres de materials húmics, cosa que els diferencia dels llacs de terra baixa de l’Europa nòrdica i del Canadà, amb els quals comparteixen, en canvi, moltes altres característiques. Malgrat la baixa biomassa i la producció vegetal a les conques, la descomposició és molt lenta, de manera que no és rar trobar, sobretot en els llacs de cotes inferiors dintre de cada serralada, molleres, i fins i tot torberes d’esfagnes.
Els estanys de muntanya normalment són petits, encara que es troben excepcions notables, com el llac Tahoe a la Sierra Nevada californiana, o alguns estanys de l’Himàlaia tibetà (Tso Moriri, Panggong Tso), que tenen alguns centenars de quilòmetres quadrats de superfície, o l’excepcional llac Titicaca, als Andes, amb més de 8 000; en canvi, solen ser profunds, si més no respecte a la mida. La relació entre l’àrea de la conca de drenatge i el volum del llac determina algunes característiques dels estanys: com més gran és, major és el contingut de sals de l’aigua, l’entrada de nutrients i les aportacions de materials particulats orgànics i minerals que l’estany rep des de la conca, aspectes que determinen els organismes que es trobaren a l’estany. La morfologia del llac, sobretot la relació entre el seu volum i la superfície de sediment en contacte amb l’aigua, i la periodicitat amb què es barreja l’aigua del llac, condicionen també les diferències de la biota dels estanys i la seva dinàmica.
Els llacs i els estanys
La variació de les característiques dels llacs a cada serralada en bona part va lligada a un gradient altitudinal. És més probable que els estanys situats en els circs, les calderes, o prop de la línia de carena tinguin una forma més arrodonida, més fondària amb relació a la seva àrea, una producció pròpia menor, poca entrada de material al·lòcton i una diversitat biològica menor. En canvi, els estanys situats a les valls de menor altitud és probable que rebin més aportacions de tot tipus que enriqueixen les possibilitats de vida.
En qualsevol cas, comparats amb els de terra baixa, els estanys de muntanya són sempre poc productius, d’aigües molt clares, fredes i toves. La baixa activitat biològica fa que els valors d’oxigen no s’apartin massa dels de saturació, encara que en llacs de molta altitud (més de 4 000 m) es poden trobar perfils d’oxigen similars als d’alguns llacs molt productius de terra baixa, amb una disminució molt marcada durant l’estiu a les capes fondes. Això és degut al fet que la pressió atmosfèrica en aquelles altituds és força més feble que a la terra baixa (pot arribar a la meitat de la del nivell del mar en els de més altitud), de manera que s’hi dissol menys oxigen i, tot i la menor activitat en el sediment, es pot arribar a situacions d’hipòxia de les capes fondes similars a les que es donen en llacs productius de terra baixa.
Les concentracions de fòsfor, principal limitador de la producció dels vegetals, són molt baixes, semblants a les d’àrees centrals dels oceans, a les aigües dels estanys de muntanya; a les serralades àrides, com algunes de la perifèria de l’Himàlaia, sembla que el rentat de sediments glaciars i morrenes aporta una mica més de fòsfor i, en conseqüència, els llacs són una mica més productius. Les concentracions de nitrogen varien molt geogràficament. A les muntanyes europees, a causa de la contaminació atmosfèrica, les concentracions de compostos de nitrogen són elevades i no paren d’incrementar-se, mentre que als Andes, en canvi, els valors són pràcticament indetectables, fet que atorga més protagonisme a alguns cianobacteris fixadors de nitrogen atmosfèric, fixació que ha estat ben estudiada en el llac Titicaca.
De vegades els estanys muntanyencs es troben en conques endorreiques, de manera que no hi ha cap efluent que dreni aigua cap a terres més baixes. Això, especialment en muntanyes relativament àrides, dóna lloc a aigües salabroses de proporcions iòniques poc habituals, generalment més enriquides de potassi i de sulfats que de clorurs. Hi ha exemples notables a les serralades perifèriques de l’Himàlaia, on trobem llacs a les ribes dels quals hi precipiten sals. Molts d’aquests llacs han sofert fortes oscil·lacions de nivell des del seu origen, normalment, però no sempre, en sentit regressiu. Els més salats tenen espècies pròpies de llacs de plana de països àrids, com l’artèmia (Artemia salina) o el rotífer Brachionus plicatilis.
Fora de la zona tropical és força comú que els estanys d’alta muntanya passin una bona part de l’any glaçats, fet que condiciona molt la vida, especialment allí on s’afegeix una quantitat considerable de neu al gruix de gel, la qual absorbeix la llum i deixa l’estany a les fosques. Aquesta coberta de neu i gel es converteix en un hàbitat singular per als microorganismes, especialment cap a la part final del període, quan la llum i la temperatura exterior augmenten. L’existència d’estanys permanentment glaçats no està ben documentada, però sembla que és el cas de l’Ororotse Tso, al Tibet, el qual té el fons cobert per un tapís d’una clorofícia filamentosa. En el cas dels estanys de muntanya d’origen volcànic, la composició de l’aigua és molt diferent de la dels d’origen glacial, que són la majoria; però, lluny de tenir organismes particulars, aquestes aigües generalment sustenten uns poblaments d’algues molt banals i la diversitat dels consumidors és escassa.
Els torrents i els rierols
Els rierols de muntanya, generalment abundants però de poc cabal, tenen unes característiques força uniformes: baixa temperatura (les aigües rarament sobrepassen els 10°C, sigui quina sigui la latitud), fort pendent (sovint més del 15%), que en alguns casos dóna atractius salts d’aigua, variacions sobtades de cabal a causa de l’estacionalitat de la precipitació (molt accentuades en les àrees de monsons o durant els períodes de fusió de la neu), baix contingut de sals en solució, conseqüència de la baixa meteorització química de les roques, i bona oxigenació, fruit de les temperatures baixes i la poca activitat biològica.
Els rierols connecten entre ells la resta d’ambients aquàtics que podem trobar a la muntanya, estanys i molleres, i presenten una gran superfície de contacte amb el conjunt d’ecosistemes terrestres. Això fa que les característiques dels organismes i dels materials que són arrossegats per les aigües siguin bons indicadors del conjunt de sistemes naturals de la conca que drenen.
El poblament faunístic i florístic
D’una manera general, pot dir-se que els sistemes aquàtics de muntanya són, a escala de tot el món, força uniformes, no sols pel que fa a les seves característiques físiques, sinó també a les biològiques. Això fa que el grau de cosmopolitisme de les espècies que en formen el poblament sigui més aviat alt i les diferències biogeogràfiques menors que en els sistemes limnètics de terra baixa.
La vida en els ambients fluvials
Les comunitats dels rierols, com gairebé de tots els ambients aquàtics d’alta muntanya, tenen un nombre elevat d’espècies cosmopolites, o si més no, d’àmplia distribució en àrees de condicions ecològiques similars. Les característiques del medi fan necessària una elevada taxa de renovació i una gran capacitat de dispersió. La fauna de macroinvertebrats dels Pirineus, per exemple, s’assembla a la dels Alps i totes dues a la d’àrees nòrdiques molt més baixes. Els estudis a les serralades asiàtiques són molt limitats i de poc detall taxonòmic, però sembla que l’abundància relativa de les famílies és també la mateixa d’un lloc a l’altre. Als rius de l’Himàlaia, per exemple, les larves d’efemeròpters, sobretot de la família dels bètids, constitueixen el 60% de les espècies, seguides en abundància per les larves de tricòpters, que són les dominants en els rierols de més altitud, principalment amb espècies de la família dels limnefílids.
Els torrents de les zones de capçalera, situats per sobre el límit del bosc, presenten unes condicions severes per al desenvolupament dels organismes. El fort pendent fa que l’aigua porti una gran empenta i tingui un poder arrossegador que impedeix la diversificació de microambients i, com a conseqüència, la diversitat de les comunitats. El risc de ser arrossegat riu avall (deriva) fa que les espècies, amb independència del grup taxonòmic al qual pertanyen, presentin adaptacions morfològiques i de comportament similars. D’altra banda, són molt accentuades les fluctuacions de cabal i les oscil·lacions tèrmiques, i no són rars els períodes de sequera, que comporten situacions de perill sobtat. Als salts d’aigua, l’empenta del corrent fa que la fauna, oligoquets i gasteròpodes principalment, es vegi fortament associada als briòfits. A les parts marginals, sobre les superfícies higropètriques, la diversitat de la flora i de la fauna augmenta.
La producció autòctona d’aquests rierols es basa en molses emergents i algues adherides a les pedres, que creixen aprofitant els pocs elements nutritius de l’aigua. La fauna és constituïda primordialment per omnívors, generalment animalons petits, de creixement ràpid i cicle curt, que solen tenir explosions demogràfiques durant el desglaç i representen una font d’aliment per als depredadors, primordialment plecòpters o planàries. Normalment en aquests torrents no hi ha peixos.
Aigües avall disminueix l’efecte immediat dels fenòmens meteorològics i la distribució es va fent més regular al llarg de l’any. L’ambient es fa més heterogeni i les comunitats s’organitzen en un mosaic complex. Augmenta la població dels herbívors brostejadors, principalment dels grups dels efemeròpters i dels quironòmids, que s’alimenten raspant les algues que creixen sobre les pedres. A les parts de menys pendent apareixen macròfits submergits i, als organismes anteriors, s’afegeixen gasteròpodes i tricòpters. La diversitat de depredadors augmenta paral·lelament a la dels herbívors; es tracta sobretot de plecòpters i de planàries, com en els torrents de més altitud, però hi són representats per espècies diferents, de cicle més llarg. Un ambient particular són els efluents dels estanys, on s’observa un predomini d’animals filtradors, simúlids i quironòmids, que aprofiten el plàncton i els detrits arrossegats des de l’estany.
Quan els rierols entren en una zona forestada, l’augment de l’entrada de fragments vegetals fa que els organismes trituradors, com determinats tricòpters, prenguin un paper rellevant. La caiguda de troncs, branquillons i fulles augmenta l’heterogeneïtat i crea barreres naturals on s’acumula matèria orgànica particulada fina que és utilitzada per organismes recol·lectors, com algunes famílies de plecòpters (sobretot nemúrids) o com els ostracodes. Els filtradors (simúlids i tricòpters com ara Hydropsyche) se situen a les parts amb més corrent. A la primavera, el desglaç produeix un rentat de material i organismes. En els moments de crescuda, moltes larves sobreviuen a les zones marginals o en els espais intersticials de la llera del riu. El retorn de l’activitat fotosintetitzadora de les algues adherides a les pedres fa augmentar els brostejadors. Moltes espècies omnívores es decanten cap a un règim més carnívor, i augmenten els genuïns depredadors. Fins i tot, hi ha un cert canibalisme, facilitat per l’increment d’individus malmesos durant l’hivern o en ser arrossegats pel corrent.
Molts rierols queden penjats en valls laterals on de manera natural els peixos no poden tenir accés. A molts països, però, hom ha introduït en aquests ambients espècies, ja sia autòctones o, més sovint, d’altres indrets i, fins i tot, altres continents. És comú que les espècies introduïdes, encara que sobrevisquin, no es reprodueixen. Normalment els peixos que habiten els rierols de muntanya són salmònids, principalment un o altre tipus de truita (Salmo, Salvelinus), i s’alimenten d’invertebrats. En els rius de muntanya del Nepal, però, hi ha peixos herbívors, com el ciprínid Garra, que s’alimenten d’algues sèssils.
La vida en els ambients lacustres
Les diferències biogeogràfiques de la flora i la fauna són menors entre els llacs de muntanya de les diferents parts del món que entre els de les terres baixes. El seu origen recent i les condicions extremes que s’hi donen no han permès gaire diversificació. Fins i tot a les serralades tropicals, com ara determinades parts de les muntanyes andines, hi ha molts tàxons cosmopolites. Tot i així, és possible trobar alguns endemismes, com algunes espècies del crustaci Boeckella als llacs dels Andes, o la diferenciació de mol·luscs (melaníids) i alguns peixos al llac Titicaca. Fins i tot es poden distingir algunes disjuncions, com la distribució dels diferents components bòreo-alpins a les diferents serralades europees, o identificar vies de colonització independents per a diferents grups, com en el cas dels crustacis dels Pirineus, dels quals els diaptòmids tenen més connexions amb espècies de l’Atles que amb les centreuropees, mentre el cas dels ciclòpids és l’oposat.
El fitoplàncton
Molts estanys d’alta muntanya tenen una manca notable d’organismes macroscòpics a causa de les condicions extremes físiques i químiques. No obstant això, la vida microbiana hi és molt important, encara que fins a la introducció de tècniques d’estudi acurades, cap a la meitat del segle XX, es considerava irrellevant. En el plàncton predominen les formes flagel·lades i petites: al voltant del 80% de les algues microscòpiques que hi viuen fan menys de 20 µm.
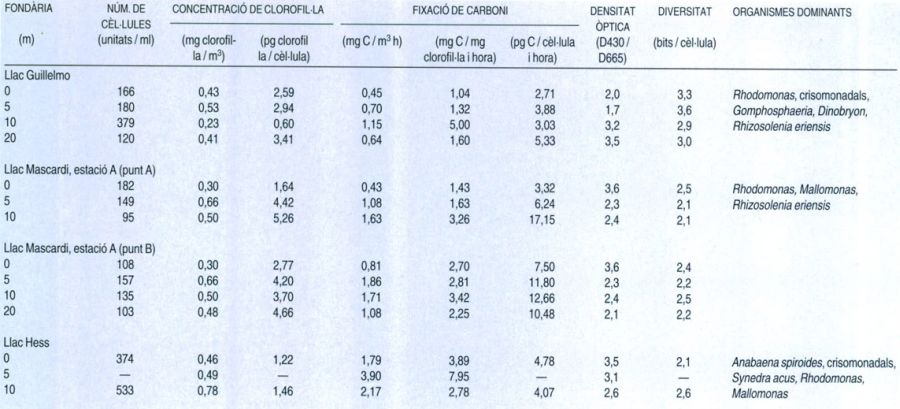
Característiques del fitoplàncton dels llacs andins argentins a una temperatura de 5,5-6,5°C (dades del 1969). Les mostres del llac Mascardi són preses a dos punts molt propers (tots dos a la mateixa estació de recerca). El fitoplàncton d’aquests tres llacs, en línies generals, presenta moltes semblances amb el d’altres llacs d’alta muntanya i amb els de les regions polars, tant pel que fa a la composició de les espècies, com a la seva concentració relativa i la productivitat. Les espècies fitoplanctòniques dels llacs d’alta muntanya són formes molt petites (no superen els 20 µm), i tant la seva biomassa com la seva activitat es mantenen a nivells força baixos a causa de l’escassetat de nutrients i les particulars condicions climàtiques. La productivitat (expressada en la fixació de carboni per unitat de volum i temps) sol ser, per tant, força baixa.
Margalef, 1983
La periodicitat anual del poblament fitoplanctònic no presenta una pauta general per a tots els estanys. A latituds temperades, normalment a la primavera, després de la fusió del gel, creixen crisòfits. El poblament d’estiu és més variat i, al llarg de la columna d’aigua apareixen, segregant-se en l’espai, criptòfits, cloròfits i dinoflagel·lats. A la tardor, és quan hi ha generalment el màxim de producció, en la qual la intervenció de diatomees és molt més variable que en els llacs de terra baixa. Sota el gel, en condicions de poca llum i baixa temperatura, les algues persisteixen força temps, i algunes espècies hi creixen activament a irradiàncies molt baixes. Si el gruix de neu deixa a les fosques la massa d’aigua, la biomassa d’algues decreix lentament però progressivament, i els flagel·lats incolors que s’alimenten de bacteris i substàncies dissoltes van prenent importància. A les àrees tropicals, l’estacionalitat és menys marcada, però normalment s’observen canvis profunds en les comunitats en el període de les pluges. Molt sovint, sobretot en els estanys amb vegetació ben desenvolupada, hi ha un percentatge significatiu d’organismes litorals que es troben a les aigües obertes d’una manera una mica accidental; és el cas de moltes desmidiàcies i diatomees de llacs andins colombians, per exemple.
El zooplàncton
A partir de la producció de les algues microscòpiques s’organitza una xarxa de consumidors i descomponedors, on els organismes petits, bacteris, ciliats i rotífers juguen un paper molt important. El poblament de crustacis planctònics és, en canvi, pobre. La composició més habitual és la d’un cladòcer filtrador i un o dos ciclòpids, als quals s’afegeix algun diaptòmid durant els períodes més favorables.
El bentos i la vegetació litoral
Les ribes de molts dels estanys d’alta muntanya no són més que un munt d’esbaldregalls. La vegetació és constituïda per molses, sovint espècies pròpies de rierols, que també colonitzen els estanys i modifiquen, en fer-ho, la seva morfologia: allarguen les tiges, la disposició de les fulles es fa més laxa i els rizoides són més prims i febles. Hi ha espècies que penetren fins a fondàries considerables, de 30 m o més, fins allà on arribi prou llum per a satisfer la seva baixa demanda respiratòria. A les aigües més toves trobem esfagnes, en alguns casos formant poblacions completament submergides. La resta de productors bentònics són algues microscòpiques, diatomees i molts cianobacteris, que es van substituint els uns als altres a mesura que augmenta la fondària. En els primers metres, la llum ultraviolada, que arriba amb més quantitat a l’alta muntanya, perjudica el recobriment de les roques per part de les algues. És molt comú observar que la part inferior de les pedres, allí on només arriba llum difosa, és molt més rica en algues que les parts exposades a les radiacions directes. Les desmidiàcies, per exemple, són abundants en llacs en contacte amb torberes o amb un litoral ric en molses, però sembla que hi ha diferències regionals que no s’acaben d’entendre bé: és més ric el poblament de desmidiàcies dels llacs andins colombians, per exemple, que el dels llacs dels Alps o, encara més, el dels estanys pirinencs.
Els estanys amb un litoral més ben desenvolupat poden tenir macròfits aquàtics submergits. En algunes serralades, com per exemple a la dels Alps, es fa difícil entendre la seva absència o escassetat. Als Pirineus, s’han estudiat amb cert detall les causes que determinen la distribució de les diverses espècies i s’ha observat que les diferències de poblament es deuen fonamentalment al contingut mineral de l’aigua. Tot i que sempre és molt baix, les diferències són suficients per a determinar canvis en la composició específica i en l’estat tròfic i així, per exemple, els estanys d’aigües molt toves tenen comunitats dominades per isoets (Isoetes). En augmentar una mica la duresa de l’aigua, aquesta comunitat és substituïda per un herbassar de potamogètons (Potamogeton), i volantins (Ceratophyllum, Myriophyllum) i ranuncles aquàtics (Ranunculus). A les basses molt productives, que generalment ho són a causa de la freqüentació del bestiar domèstic, s’estableixen poblacions de macròfits d’aigües riques de nutrients, com ara cal·lítrics (Callitriche). Es troben comunitats similars en llacs andins i de serralades asiàtiques en condicions equivalents.
Les fanerògames rarament viuen a més de 5 m de fondària. En aigües més profundes, els únics macròfits que es troben són caròfits, algues verdes macroscòpiques, principalment del gènere Nitella, o les molses abans esmentades. El límit al creixement d’angiospermes no l’imposa la llum, ja que la seva presència s’atura a fondàries amb irradiàncies per sobre del punt de compensació de la respiració. Les causes poden ser diverses, per exemple canvis en la distribució del sediment degudes al pendent. La causa última, però, quan tots els altres factors són favorables, és un augment de la concentració d’oxigen en els espais aeris interiors de les plantes a mesura que la fondària creix. La pressió hidrostàtica més alta permet una pressió parcial dels gasos (entre ells l’oxigen) més elevada a l’interior dels espais aeris de les plantes. Això afavoreix un increment de la concentració de radicals oxidants, els quals ataquen les membranes de les cèl·lules de les plantes. Plantes sense espais aeris com les molses o algunes fanerògames marines arriben a fondàries més grans.
Als estanys amb una vegetació més ben desenvolupada sol presentar-se una clara distribució del poblament vegetal en forma de cinyells concèntrics, des de la vegetació dels prats del voltant fins a la de les plantes surants aquàtiques. Als Pirineus, per exemple, la transició entre el prat alpí i les comunitats submergides és representada per una comunitat de ciperàcies (Carex, Eriophorum) i juncàcies (Juncus), junt amb algunes altres plantes característiques, com les violes d’aigua (Pinguicula) o la fetgera blanca (Parnassia palustris). Tot i que aquesta comunitat només s’inunda temporalment, sempre disposa d’aigua abundant gràcies al substrat d’esfagnes i molses que l’acull. Cap a l’interior de l’estany, ja completament inundat, pot haver-hi un altre cinyell, gairebé monospecífic de càrex inflat (Carex rostrata). Fins a uns quants metres de fondària, s’estén la comunitat típicament aquàtica, amb isoet lacustre (Isoetes lacustris), subulària (Subularia aquatica), ranuncles d’aigua (Ranunculus aquatilis, R. pseudofluitans) o potamogètons (Potamogeton). L’espargani de muntanya (Sparganium angustifolium), de llargues fulles acintades, es troba superposat a aquestes plantes de petites dimensions.
En llacs petits i soms i amb molta vegetació aquàtica, l’elevada fotosíntesi durant el dia pot fer pujar el pH de l’aigua fins a valors sorprenentment alts. Això fa que el diòxid de carboni sigui escàs, i encara més si l’aigua s’escalfa i l’altitud és gran. En aquest cas es justifica una estratègia com la que han desenvolupat els isoets (Isoetes) que consisteix a agafar diòxid de carboni a la nit i emmagatzemar-lo en forma d’àcids orgànics, per utilitzar-los de dia per a la fotosíntesi. Amb aquest metabolisme fotosintètic, que es creia característic de plantes terrestres adaptades a llocs àrids (concretament de les crassulàcies, en les quals havia estat descobert inicialment) es disminueix el risc d’una pèrdua d’aigua excessiva obrint els estomes durant la nit.
Els invertebrats
La meitat de la fauna d’invertebrats que viu a la zona litoral són insectes que no desenvolupen el seu cicle complet al si de l’estany. Moltes larves d’insectes són les mateixes que es troben a rierols i fonts. Els invertebrats que sí que compleixen tot el seu cicle a l’aigua de l’estany són nombrosos coleòpters, mol·luscs, sovint amb la closca molt feble a causa del baix contingut de calci de les aigües, esponges, briozous, celenterats, planàries i crustacis. Normalment hi ha més diversitat de crustacis bentònics que no pas de planctònics, sobretot de cladòcers del grup dels quidòrids. La fauna bentònica de les parts més fondes i sense vegetació és menys diversa que la del litoral i es limita a quironòmids, oligoquets, ostracodes i, excepcionalment, mol·luscs bivalves.
Els peixos i els amfibis
Els peixos presenten poca diversitat en els estanys de muntanya. Generalment s’hi troba una sola espècie de truita (dels gèneres Salmo, Oncorhynchus o Salvelinus, o bé, a les aigües de les terres més meridionals de l’hemisferi sud, Galaxias). Com que la pesca esportiva és un dels pocs usos d’aquests estanys, la introducció d’espècies i el repoblament d’estanys originàriament sense truites ha estat un fet comú. El principal aliment de les truites són les larves d’insectes, i també els insectes adults atrapats a la pel·lícula superficial de l’aigua. Però en moments d’escassetat poden menjar pràcticament qualsevol cosa, fins i tot ous i exemplars més petits de la mateixa espècie.
Els amfibis adults no són comuns als estanys, les fases larvals, que duren uns 20 dies, de vegades omplen les vores o, més sovint petites basses del litoral, que serveixen de refugi contra l’acció depredadora dels peixos.