L’arxipèlag planetari
En rigor, totes les terres emergides del planeta, fins els continents, són illes. Totes les masses continentals, el Vell Món, el Nou Món, l’Antàrtida, Austràlia, estan envoltades d’aigües. Però és per sota dels 10 milions de km2 que comencen a plantejar-se dubtes. Austràlia, amb els seus 7 milions i mig de km2, és un continent o una illa? S’accepta generalment que és un continent, però el seu poblament vegetal i animal té no poc d’insular i tampoc no és rar sentir-lo anomenar continent-illa. Grenlàndia, que amb més de 2 milions de km2 és generalment reconeguda com la més gran de les illes, pel seu inlandsis, fa un bon paper com a bessona petita de l’Antàrtida i sovint és tractada com un element diferenciat d’Amèrica. Nova Guinea, que no arriba als 800 000 km2, és la segona illa en grandària i la primera que ho és sense ambigüitat ni dubte de cap mena. Tot plegat, acceptant Grenlàndia en el collectiu, un 5% de la superfície terrestre estaria ocupat per illes, unes 500 000 en total.
Les illes continentals i les illes oceàniques
Hom pot considerar d’una manera general com a illes continentals aquelles que al llarg de la seva història han sofert unions amb els continents propers, i que, per tant, han homogeneïtzat la seva fauna amb la continental, independentment de l’origen geològic. Les illes continentals són la majoria, però són menys “illes” en el sentit biològic estricte, ja que la seva fauna i flora s’assemblen més a les dels continents veïns i els endemismes són escassos.
Les illes pròpiament oceàniques, en canvi, són les que no han estat mai en contacte amb el continent o que, en tot cas, ho estigueren fa molts de milions d’anys; són aquestes les que presenten les biotes insulars més extraordinàries, caracteritzades precisament per la seva pobresa taxonòmica, per la seva manca d’harmonia (no presenten els mateixos tipus adaptatius bàsics) respecte les biotes continentals de les quals deriven, i pel seu elevat índex d’endemismes.
Una tercera categoria, les illes paraoceàniques, són aquelles que, tot i haver estat en contacte amb el continent, presenten biotes comparables a les d’una illa oceànica (amb elevats graus de pobresa, manca d’harmonia i endemicitat). Illes com les mediterrànies, les illes Malvines, l’illa de Wrangel o les illes Mentawai són paraoceàniques. Tot i haver tingut un contacte amb el continent, la seva composició faunística és marcada per l’acció d’uns filtres ecològics extrems. En el cas de la Mediterrània, algunes de les illes foren colonitzades al Messinià, fa 5,5 milions d’anys, quan la Mediterrània es va assecar a causa del tancament de l’estret de Gibraltar. La fauna colonitzadora va haver de travessar autèntics deserts salins per assolir les terres insulars. Aquests deserts constituïren un filtre per a la colonització de les illes, un filtre encara més potent que la mateixa mar. En el cas d’illes com la de Wrangel o les Malvines, el filtre degué ser imposat per unes condicions climàtiques de freds extrems. Les illes Mentawai, a ponent de Sumatra, poden haver estat unides a la plataforma continental durant les glaciacions, però a través d’un istme summament estret, que també va actuar de filtre per a la seva colonització.
El conjunt de les illes oceàniques i paraoceàniques, que són el 2/3 de les illes del planeta, ha de ser considerat com un punt calent de biodiversitat a la Terra, almenys per a certs grups animals. Així, en l’1,5% de la superfície terrestre que representen les illes oceàniques i paraoceàniques es troba (o es trobava fins a l’arribada dels humans) aproximadament el 15% de les espècies modernes de mamífers, més del 20% de les d’ocells i una proporció encara més gran de les de rèptils.
El procés de colonització biològica
Les illes, com els éssers vius, tenen un naixement, una durada, i una mort. N’hi ha de joves i de molt velles. N’hi ha d’efímeres i de durables. Quan neixen, les illes ofereixen als naturalistes l’oportunitat d’estudiar els fenòmens de colonització. Tanmateix, el naixement d’una illa és un esdeveniment rar. Algunes illes que han nascut recentment són la de Surtsey, a Islàndia, que va néixer el 1963, o la d’Anak Krakatau, nascuda el 1930, a l’estret de Sunda, entre Java i Sumatra, al cràter submergit que va deixar el Krakatau després de la seva famosa explosió del 27 d’agost de 1883. La colonització de les illes s’ha pogut estudiar també després de l’aniquilació del seu poblament viu, resultat, bé de causes naturals (normalment la devastació originada per una explosió volcànica o per un tifó), bé de causes artificials (als anys seixanta es van desfaunar amb biocides alguns “cayos” del Carib amb l’objecte de poder estudiar-ne la recolonització).
Colonització de les illes del Pacífic per les plantes fanerògames
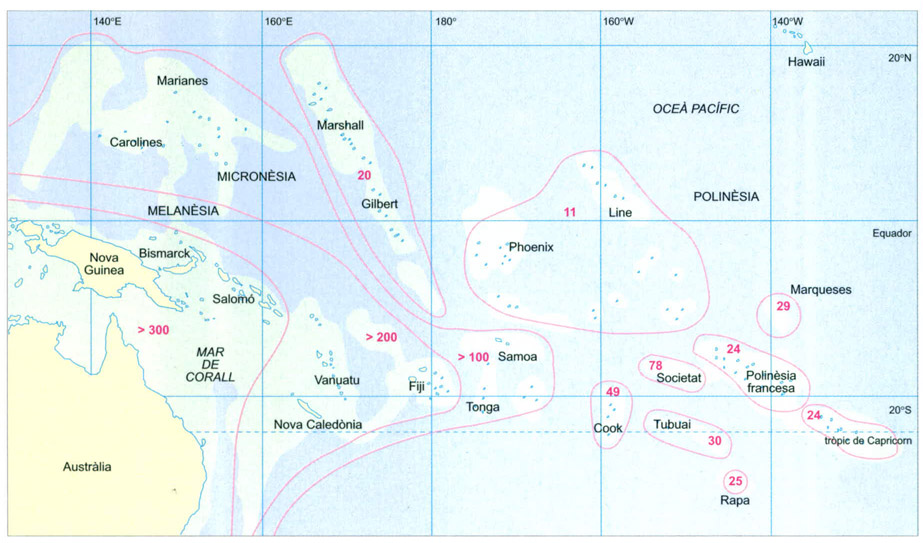
Les isolínies vermelles encerclen localitats amb el mateix nombre de gèneres.
Editrònica, a partir de B. Cox, D. Moore & P. Whitefield, 1989
Si s'analitza la colonització de les illes del Pacífic per les plantes fanorògames es posa en evidència que com més cap a l’est del continent asiàtic es troba una illa, menys plantes han aconseguit arribar-hi i colonitzar-la. La diversitat vegetal d’una illa és bàsica per a la dispersió dels altres organismes vius, ja que determina la disponibilitat de recursos ecològics i, per tant, la possibilitat que un nombre més o menys gran d’espècies animals puguin colonitzar-la. Moltes fanerògames presenten adaptacions per assegurar-se que les seves llavors es dispersin i germinin lluny de la planta mare. Algunes llavors, per exemple, presenten un plomall que facilita que se les emporti el vent; altres són incloses en fruits carnosos que són ingerits pels ocells, i així aconsegueixen superar grans distàncies; encara d’altres tenen les llavors recobertes per petits ganxos o bé per unes glàndules que segreguen substàncies viscoses que els permeten fixar-se al plomatge dels ocells i acompanyar-los en els seus viatges. Les plantes que presenten adaptacions d’aquest tipus són les que més fàcilment aconsegueixen arribar a les illes llunyanes.
La dispersió transoceànica
L’arribada d’organismes terrestres a les illes oceàniques és atzarosa i sovint passiva. D’altra banda, la capacitat de supervivència a la llarga travessia varia molt d’un tàxon a un altre, per la qual cosa la composició florística i faunística de les comunitats que es van formant és anòmala si es compara amb la dels continents. Així, dominen algunes famílies, mentre que molts altres grups manquen. Per exemple, els llimacs, els amfibis i els mamífers no voladors són pràcticament desconeguts. L’absència de mamífers depredadors de vertebrats és un dels trets més remarcables de les illes oceàniques, tant per la dificultat que tenen a arribar-hi com, sobretot, perquè els recursos disponibles en un territori petit no són suficients per a mantenir una població viable de carnívors. En canvi, les poblacions de cargols terrestres poden sobreviure en àrees summament petites durant molt de temps, la qual cosa explica l’enorme diversificació d’aquests animals en quasi totes les illes.
El vent pot transportar a les illes les llavors de les plantes en el cas d’espècies anemocores, però també dispersa les espores de les falgueres i les diminutes llavors de les orquídies. De la mateixa manera, pot dur-hi animals molt petits, insectes voladors i rates-pinyades. També es creu que és el vent el responsable de la colonització de remotes illes del Pacífic per cargols terrestres de mida petita; els cargols de la família dels endodòntids (gairebé tots molt petits) estan representats per multitud d’espècies a Micronèsia: només a la petita illa de Rapa se n’han descrit més d’un centenar com a endèmiques.
Els corrents marins són responsables de l’arribada de petites illes flotants, procedents de les desembocadures de rius cabalosos i formades per restes de vegetació de ribera acaramullats amb plantes flotants. Aquestes masses vegetals contenen multitud d’organismes, molts dels quals poden resistir un viatge transoceànic. Tots els organismes dispersats per la mar, però, han de poder resistir la salinitat exterior. No és sorprenent que la franja litoral de les illes perdudes a les regions tropicals del Pacífic i de l’Índic contingui una vegetació formada majoritàriament per espècies de distribució molt àmplia, les llavors de les quals es mantenen viables després de mesos surant. Tampoc no es pot oblidar la colonització de les illes oceàniques per organismes marins que s’adapten als ambients sense ocupar i entren a formar part de les comunitats terrestres i de les aigües dolces.
Les espècies pioneres
En una primera fase de la vida d’una illa, hi arriben el que es consideren espècies pioneres, colonitzadores d’èxit (diversos ocells, insectes voladors, etc.). Les poblacions d’aquestes espècies mantenen normalment un flux genètic amb les restants poblacions de l’espècie, de manera que no s’originen, almenys de bon començament, espècies endèmiques insulars. Entre les espècies colonitzadores eficients hi ha òbviament els “super-rodamons”, que han optat per la dispersió com a forma de vida: es troben presents en densitats espectacularment elevades en les illes disponibles per a ser colonitzades. Efectivament, es tracta d’espècies que tenen una extrema habilitat per a travessar canals marins i tenen una capacitat reproductora elevada. Solen residir en illes que són massa petites per mantenir poblacions estables d’altres espècies o en illes que han estat devastades per tifons, huracans o per explosions volcàniques. Són desplaçades per altres espècies que hi arriben més tard, però que poden utilitzar els recursos insulars d’una manera més eficient i durable.
Les illes no tenen la mateixa significació per a totes les espècies. Així, illes que per als grans vertebrats poden representar territoris molt particulars, per a un petit animal o una molsa poden no ser gaire diferents del continent. Això no vol dir que aquests organismes no s’hi especiïn, sinó que ho poden fer seguint unes pautes similars a les que segueixen al continent. Així i tot, moltes illes imposen també models propis a l’evolució. Un exemple, entre molts d’altres, seria el dels wetas, un grup d’ortòpters nocturns no voladors, que són els insectes més grans de Nova Zelanda (fins a 82 mm de llarg i 71 g de pes en el cas de Deinacrida heteracantha, el wetapunga). Un altre exemple seria la papanòvia gegant de Santa Helena (el dermàpter Labidura herculeana), o algunes aranyes gegantines, com ara algunes de les de l’illa Deserta Grande (Madeira), de São Tomé, de Vanuatu o de les Fiji.
Els relictes i les novetats
Els endemismes de les illes poden ser o bé espècies relictes, provinents d’antigues nissagues d’antuvi esteses per àrees continentals, o espècies (o tàxons més elevats) originades a illes per nesoevolució.
Perquè s’originin endemismes insulars cal que es trenqui el flux genètic amb les poblacions ancestrals, i que transcorri temps a bastament perquè es puguin donar prou canvis evolutius. Els endemismes originats per nesoevolució es produeixen a una escala relativament gran quan l’illa és suficientment lluny de les regions d’origen perquè l’arribada d’immigrants sigui un fenomen aleatori, i mantenir el contacte amb les poblacions originals sigui pràcticament impossible del tot. Les espècies que colonitzen les illes per aquesta via atzarosa cauen en una trampa tan biogeogràfica com evolutiva: són illes d’on tenen molt poques probabilitats de sortir. Sovint, algunes espècies descendents d’aquelles que hi havien arribat nedant inicien la colonització dels medis terrestres i evolucionen perdent la capacitat de nedar, com per exemple els extingits hipopòtams nans de Xipre (Phanourios minor) o el cocodril terrestre de Nova Caledònia (Mekosuchus inexpectatus). Igualment, els descendents de moltes espècies que hi han arribat volant perden la capacitat de vol, com és el cas dels extints moes (Dinornis) i dels kiwis (Apteryx) de Nova Zelanda o el dels ocells-elefant de Madagascar (Aepyornis).
La manca dels depredadors continentals habituals, la nova combinació d’espècies a les xarxes ecològiques insulars i l’oferta de recursos disponibles als medis insulars colonitzats condicionen en bona part l’evolució d’aquests estranys habitants de les illes. Els processos històrics tenen una gran importància a les illes, ja que l’ordre d’arribada condiciona sovint les diferents formes d’especiació.
El poblament biològic i les interrelacions
Tot i que l’intercanvi de matèria i d’energia que es dóna entre les illes genuïnes i les aigües que les envolten es fa bàsicament per la via detrítica, determinades espècies animals han sabut viure a cavall entre els dos medis. Aquest és el cas dels pinnípedes i els altres ocells marins, de les llúdries marines, de les tortugues i les iguanes marines i molts d’altres. Aquestes espècies relacionen els medis costaners, i les illes en particular, amb les aigües circumdants, i és mercès a aquesta mena de relacions que es pot dir que la importància ecològica de les illes ultrapassa (en alguns casos molt) la derivada de la seva simple extensió geogràfica.
Les illes de Santa Helena (122 km2) i l’Ascensió (97 km2), per exemple, malgrat la seva petita superfície, constitueixen les úniques taques de terra ferma en una àrea de 15 milions de km2 (el 3% de la superfície de la Terra). Els ocells marins que hi nien, en realitat viuen sobre una extensió molt àmplia de l’oceà Atlàntic, de manera que és a través d’aquestes espècies, que viuen a cavall entre les illes (on es reprodueixen) i les aigües circumdants (on s’alimenten), que les illes exerceixen la seva influència sobre la mar que les envolta. D’ençà de l’arribada dels humans a aquestes illes, les poblacions d’ocells marins han passat de ser de desenes de milions d’individus a només uns pocs milers. Així, la petita intervenció dels humans en només unes àrees molt reduïdes de la Terra ha modificat el funcionament ecològic d’àmplies àrees oceàniques.
Les singularitats de l’evolució insular
El resultat de l’evolució insular és sempre sorprenent, i sovint dóna lloc a fenotips “prohibits” en ambients de més extensió i diversitat. Un bon exemple és la papanòvia gegant de Santa Helena (Labidura herculeana), un dermàpter que ateny els 8 cm de llarg, i és, amb diferència, la papanòvia més gran que es coneix. Les illes oceàniques són, doncs, veritables laboratoris naturals, on les possibilitats evolutives es duen als extrems més insospitats.
Les paradoxes més freqüents
La mida reduïda de la població, la intensitat elevada dels factors de selecció i el relaxament de les limitacions que hi ha en ecosistemes més diversos sobre les possibles morfologies i comportaments fan que l’evolució adaptativa de les poblacions que s’estableixen a les illes oceàniques sigui ràpida i vagi acompanyada d’una sèrie de canvis poc predicibles, conseqüència del fenomen de deriva genètica, a causa del baix nombre de colonitzadors inicials, de la mida mitjana de la població i de les fortes oscil·lacions d’aquesta. En molts casos també actua la selecció sexual, la qual no es troba tan limitada per factors adaptatius com als continents.
L’evolució en aïllament és responsable de l’existència de plantes tan estranyes que han hagut de ser classificades en famílies pròpies com ara Lactoris fernandeziana, únic representant de les lactoridiàcies i endèmica de l’illa de Juan Fernández, o Degeneria vitiensis, exclusiva de les Fiji i únic membre de les degeneriàcies. Altres espècies vegetals, tot i que pertanyen a famílies ben conegudes als continents, han adquirit morfologies sorprenents, com les compostes arborescents de Santa Helena, de Raratonga i de Juan Fernández. Les compostes es dispersen millor que la majoria dels arbres de les zones tropicals i temperades, per això han pogut colonitzar aquelles illes i evolucionar cap a formes arborescents abans que arribessin els arbres convencionals, aprofitant aquesta oportunitat ecològica única.
Els ocells que fan migracions a grans distàncies poden fer escala a les illes més aïllades, o fins i tot establir-s’hi com a residents. L’estudi fet sobre ocells de les illes ha demostrat que una espècie isolada en una illa remota pot acabar adquirint una gran especialització alimentària i uns hàbits particulars sense cap altra pressió selectiva que l’optimització de les condicions en un ambient restringit que ofereix poca varietat de recursos. Això té una gran importància per a la teoria ecològica, car contradiu les teories que expliquen la diferenciació entre espècies properes com a conseqüència de la competència interespecífica.
En aquest sentit, és il·lustratiu el cas del víreo de Fernando de Noronha (Vireo gracilirostris), un petit ocell insectívor endèmic d’aquesta illa, situada davant de l’extrem oriental de l’Amèrica del Sud. Es tracta d’una població divergent d’una espècie d’ocell migrador que és comuna al continent americà, i que a l’illa derivà ràpidament cap a una morfologia i un comportament que li permeten capturar petits insectes d’entre el fullam, sobretot de la cara inferior de les fulles; en conseqüència, ha sofert una reducció de mida, un arrodoniment de les ales, un allargament de les potes i de la cua, i ha desenvolupat un bec més llarg i prim i una coloració diferent i més vistosa que la del seu avantpassat. Un cas semblant és el dels ulls-blancs de la Reunió (Zosterops borbonicus i Z. olivaceus); la primera d’aquestes espècies, més generalista, s’assembla més a altres congèneres, mentre que la segona, més diferenciada, és nectarívora, de manera que sembla raonable suposar que l’especialització d’aquesta darrera és considerablement anterior i per tant la seva arribada a les Mascarenyes és independent de la de l’altra i la precedí.
El cas dels ocells pelàgics
La majoria dels ocells marins pelàgics nien a les illes més llunyanes, on formen colònies nombrosíssimes. Aquests ocells, d’ales llargues i potes curtes, perfectament adaptats a volar enormes distàncies aprofitant l’oratge de la superfície de la mar, serien presa fàcil per a molts depredadors, si aterressin a les costes continentals. Per això es concentren per criar a les illes més allunyades, on estan lliures d’enemics terrestres. Allí, cada parella nidificant pon un sol ou per niada, l’incuba durant un període prolongat i té cura del poll, que creix molt a poc a poc i acumula molts greixos abans de poder-se valdre. La mida de l’àpat que rep un poll representa normalment entre un 14 i un 18% del pes de l’adult que el duu. Sota aquests condicionants, aquests ocells han desenvolupat una estratègia reproductiva que permet maximitzar l’energia neta disponible i minimitzar el nombre de viatges que fan els adults, els quals participen tots dos per igual en les tasques d’incubació i alimentació del poll. Els procel·lariformes han adquirit una adaptació addicional, afavorida per la seva dieta rica de lípids: el seu estómac té una cambra especial on el menjar és parcialment digerit per a augmentar-ne la densitat.
Els ocells pelàgics, atesa la densitat de població que atenyen i el temps que passen a les illes, són un element fonamental en l’ecologia de les illes més aïllades. Les àrees de nidificació reben la deposició d’enormes quantitats d’excrements, que s’acumulen formant l’anomentat guano. A més d’establir un lligam entre una àrea terrestre molt reduïda i una regió marina normalment força àmplia, també poden transportar d’una illa a una altra, aferrats al plomatge o dins del tracte digestiu, animals petits, propàguls i llavors. Aquest tipus de dispersió sembla, per exemple, l’única explicació plausible de la presència de Peperomia berteroana, una piperàcia que només es troba a dues illes allunyadíssimes entre elles d’oceans diferents: Juan Fernández i Tristan da Cunha. Un cas semblant podria ser el de les compostes del gènere Rhamphogyne, amb només dues espècies, una a les Mascarenyes i l’altra a Nova Guinea.
El cas dels ocells no voladors
La pèrdua de la capacitat de volar dels ocells és un fenomen que s’ha produït en repetides ocasions en moltes illes remotes, originant nombroses espècies endèmiques. Lliures d’enemics, la selecció natural hauria d’afavorir la capacitat d’emmagatzemar greixos i minimitzar les despeses metabòliques associades a activitats inútils, en absència de depredadors terrestres dels quals fugir, com ara volar. La família que inclou la majoria d’aquests casos és la dels ràl·lids, que sembla que foren molt abundants en moltes illes. Actualment en sobreviuen pocs, per tal com foren presa fàcil dels mariners i les rates que els acompanyaven. S’estima que s’han extingit més de dues mil espècies de rasclons no voladors insulars a partir de la primera arribada dels humans a les illes on vivien. És el cas dels rasclons de les Hawaii, coneguts només per testimonis arqueològics. La darrera extinció registrada d’aquest grup fou la de Gallirallus wakensis, exterminada al final de la Segona Guerra Mundial per soldats que quedaren abandonats a l’illa de Wake, al mig del Pacífic. El “takahe” de l’illa Sud de Nova Zelanda (Notornis mantellii), donat per extingit al començament del segle XX, fou retrobat el 1948.
Probablement l’habitant d’illes remotes més popular sigui un altre d’aquests ocells no voladors, el dodo (Raphus cucullatus), un ocell famós per la seva estranya forma i suposada estupidesa. Endèmic de l’illa Maurici, despertava entre els mariners europeus que arribaven a aquesta terra deshabitada una barreja d’admiració i burla: on s’havia vist mai un ocell tan gras, amb unes ales tan petites, un cap tan estrany i que no fugia dels humans? Tothom sap que els dodos van desaparèixer per sempre pocs anys després del seu descobriment, a causa de les matances de què foren objecte pels mariners i els animals que introduïren. Avui només en resten alguns dibuixos i descripcions del segle XVII, trossos d’exemplars dissecats i els pocs ossos que s’han trobat en prospeccions paleontològiques. Una altra espècie, el solitari (Pezophaps solitarius), endèmic de l’illa Rodrigues, també s’extingí al segle XVIII.
L’índex d’endemisme
Així i tot, la proporció i la quantitat d’espècies endèmiques, generalment elevades, varia molt entre la multitud d’illes disperses als oceans. Situació geogràfica, dimensions, clima, relleu i història geològica són els factors fonamentals que ho expliquen: una illa gran, muntanyosa i tropical contindrà moltes més espècies i una fracció més gran d’endemismes que no una illa petita, baixa i situada fora de les regions intertropicals. Un exemple d’aquest contrast, el forneixen la fauna de caragols terrestres d’Islàndia, on només hi ha una vintena d’espècies àmpliament distribuïdes a les regions boreals d’Europa, comparada amb les comunitats de cargols equivalents de les illes altes de Polinèsia. Per exemple, el gènere Partula a l’illa de Moorea es diversificà almenys en onze espècies, dues de les quals donaren lloc a subspècies ben definides. El clima suau i càlid d’aquesta illa abrupta i coberta de boscúries permeté un procés d’evolució al·lopàtrica comparable al d’altres cargols a Hawaii i les Canàries.
Les illes oceàniques més típiques (excloent les més grans, Madagascar i Nova Zelanda) són molt joves a escala geològica. Les poques que es podrien anomenar antigues tenen com a màxim vint milions d’anys d’edat. Poques, però, en tenen més de deu. Les més velles han hostatjat els grups més diversificats, incloses les espècies més estranyes, que són les que han tingut més temps per a evolucionar en aïllament. Així, el 80% de la flora fanerogàmica de les Mascarenyes la constitueixen gèneres endèmics, cosa que testimonia l’antiguitat d’aquestes illes.
La història geològica de cada illa afecta decisivament l’evolució dels seus habitants. Moltes illes oceàniques situades a les regions àrtiques i antàrtiques tenen biotes pobres i sense endemismes a causa de les glaciacions. La submersió repetida d’Aldabra i d’altres atols durant els períodes interglacials fa que les faunes quaternàries d’aquestes illes estiguin formades per episodis independents de colonització, diversificació i extinció. A les Bermudes, on al llarg del Plistocè sempre hi hagué alguna part emergida, la situació és més complicada; els cargols del gènere Poecilozonites hi han deixat un excel·lent registre fòssil, que testimonia un patró de diversificació iteratiu causat per la fragmentació illenca i l’adaptació als canvis ambientals.
Variabilitat dels cargols del gènere Partula
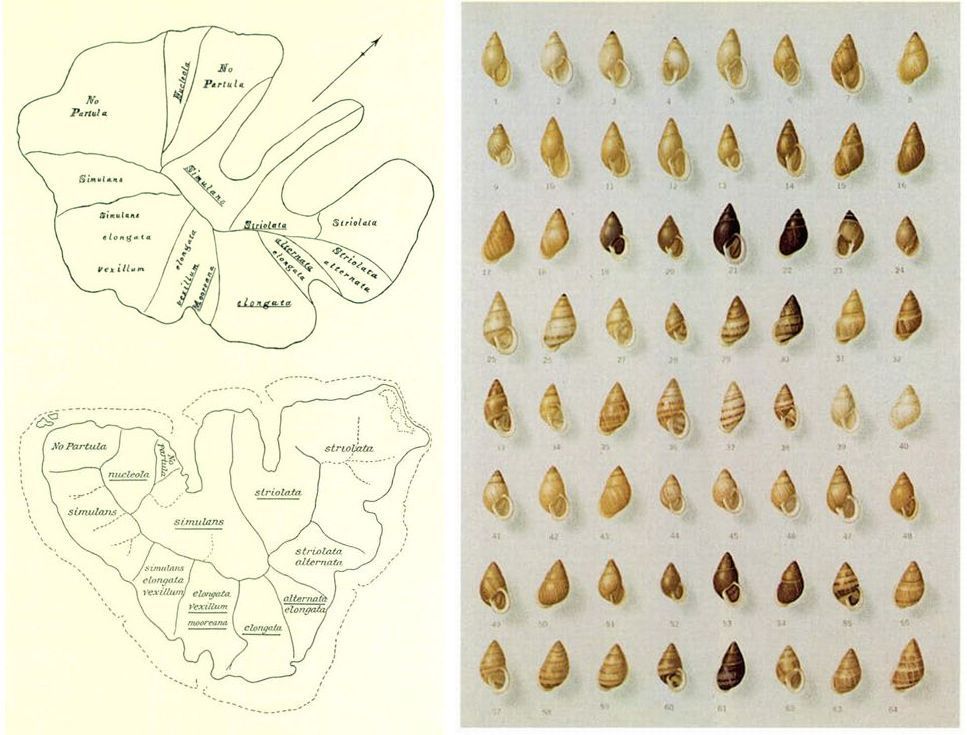
Cortesia del Museu de Zoologia de Barcelona / Jordi Vidal
Onze espècies de cargols terrestres del gènere Partula, la majoria de les quals es troben actualment en perill d’extinció, viuen o vivien a l’illa polinèsia de Moorea. Aquesta gran diversitat d’espècies es deu a la singularitat del relleu de l’illa. En efecte, una cadena muntanyosa en forma de mitja lluna, part de la paret d’un antic cràter volcànic, recorre l’illa del NW al NE, amb la part convexa cap al sud, mentre que perpendicularment a l’eix principal d’aquesta cadena s´obren nombroses valls, que queden ben delimitades per unes crestes muntanyoses.
El relatiu aïllament de cada vall determina en gran manera la distribució de la vegetació, que al seu torn proporciona hàbitats adequats a les diferents espècies de cargols. Una mateixa espècie pot trobar-se en diferents valls, amb subspècies i varietats diferents a cadascuna, o bé estar restringida a una determinada zona dins una de les valls. La distribució de les diferents espècies, subspècies i varietats fou estudiada per Garrett del 1866 al 1888. H. E. Crampton reproduí el mapa de la distribució de Garrett (a dalt) a la seva obra “Studies on the variation, distribution and evolution of thegenus Partula” (Carnegie Institution of Washington, 1932), al costat d’una versió millorada feta per ell mateix (a baix), ja que en l’època de Garrett no es disposava de mapes gaire exactes de l’illa de Moorea (el mapa que Garrett dibuixà està clarament esbiaixat; la part occidental de l’illa, que estudià amb més detall, queda desproporcionadament gran). A la mateixa obra, Crampton proporciona una mostra de la gran diversitat de formes de les subspècies P. taeniata simulans (números de l’1 al 38) i P. t. striolata (números del 39 al 64), dues de les més esteses. Segons la localitat d’on procedeixin, els cargols tenen diferent mida, amplada de l’obertura de la conquilla, patró cromàtic, etc. Les varietats corresponents als números 4 i 23 són mutants que enrotllen la conquilla cap a l’esquerra.
El fenomen del gegantisme
El dodo (Raphus cucullatus) forneix un exemple valuós de l’evolució en illes oceàniques. Indubtablement es tractava d’un colom gegantí que havia evolucionat sense depredadors durant milions d’anys. A part el dodo de Maurici, altres coloms gegantins i no voladors, anomenats solitaris (Pezophaps solitarius), que foren exterminats de la mateixa manera, visqueren en una altra de les Mascarenyes: l’illa Rodrigues. És interessant que en cadascuna d’aquestes illes perdudes a l’oceà Índic els coloms experimentessin una evolució independent però paral·lela, des de les formes voladores capaces de colonitzar-les fins als famosos ocells extingits. El temps disponible per a sofrir aquestes modificacions sembla decisiu, atès que les Mascarenyes es troben entre les illes oceàniques més antigues. Un gènere endèmic de tortugues gegants, Cylindraspis, més diferenciades entre illes que en el cas de les Galápagos, tingué a les Mascarenyes la mateixa sort que els dodos i els solitaris.
També a les illes coral·lines situades al nord de Madagascar hi hagué tortugues gegants. En aquest cas, probablement transportades fins allí pels corrents marins des de Madagascar, ja que estaven emparentades amb les que es coneixen com a fòssils d’aquesta gran illa. Només en sobreviu una sola espècie, la tortuga d’Aldabra (Aldabrachelys gigantea), que amb 105 cm de longitud de la closca i un pes de 120 kg, presenta com a tret característic una mena de llavi a la cambra nasal anterior que li permet beure pel nas; aquesta adaptació tan curiosa li permet aprofitar els tolls petits i poc profunds que són el seu únic subministrament d’aigua dolça.
Ocells i tortugues gegants tingueren amb seguretat una gran importància als ecosistemes de les illes més isolades. Eren els principals herbívors i frugívors, i tots els relats contemporanis en parlen en termes d’abundància. Són pocs, però, els estudis dirigits a esbrinar el paper d’aquests gegants sobre l’evolució de la flora local. Pel que se sap de l’únic supervivent d’aquests gegants, la tortuga d’Aldabra, té efectes devastadors sobre la vegetació natural. Com que li cal l’ombra dels arbustos per a suportar la calor del dia, sovint excava un clot per trobar-se més a gust, amb la qual cosa causa la mort de molts arbres i impedeix el creixement dels plançons. L’acumulació de materials fecals al peu i als voltants dels arbustos alimenta una elevada densitat de crancs ermitans terrestres. La voracitat d’aquestes tortugues ha estat sens dubte un factor de selecció determinant sobre les plantes que formen l’anomenada “gespa de tortuga”, les quals són varietats nanes que rarament superen els 2 cm d’alçada, adaptades a una intensa pressió d’herbivorisme. D’altra banda, les tortugues mantenen una relació ambivalent amb el ràl·lid no volador Dryolimnas cuvieri aldabranus, que és un depredador de les tortugues joves però que lleva els paràsits externs de les adultes. Actualment es calcula que hi ha més de 150 000 tortugues gegants a Aldabra, sens dubte un factor essencial en l’ecosistema terrestre d’aquest atol.
Un cas ben conegut que implica un gegant extingit és el que relaciona el dodo de Maurici (Raphus cucullatus) amb el tambalacoc o arbre de ferro de Maurici (Sideroxylon grandiflorum), una sapotàcia endèmica de grans dimensions i en perill d’extinció, representada ara quasi exclusivament per exemplars nascuts abans de l’extinció dels dodos. Les llavors del tambalacoc necessiten passar una escarificació superficial per a germinar, cosa que s’ha comprovat que s’esdevé quan passen pel tracte digestiu d’un gall dindi. No hi ha gaire constància del comportament del dodo, però se sap que s’empassava pedretes per ajudar a la trituració de les fruites que ingeria. Totes les proves apunten cap a una relació de mutualisme entre l’arbre que produïa fruits i l’ocell que en dispersava i n’activava les llavors. De tota manera, encara es discuteix sobre l’obligatorietat d’aquest mutualisme, desenvolupat en una comunitat formada per poques espècies.
Els casos extrems d’aïllament
La composició florística de les illes Mascarenyes, per altra banda, permet copsar la importància de la dispersió a través de l’oceà en illes oceàniques que no formen part de grans arxipèlags. Les espècies terrestres que hi viuen són aquelles que hi han pogut arribar (sempre aportacions escasses i esporàdiques), han tingut èxit en colonitzar el nou territori i han pogut mantenir poblacions demogràficament viables. Aquest grup d’illes, situades a l’Índic sud-occidental, és format per tres edificis volcànics emergents molt allunyats entre ells. La Reunió conté un gran volcà actiu, mentre que Maurici i Rodrigues són terres ja força erosionades. Malgrat la gran quantitat d’endemismes que hom ha descobert en cada una d’elles, la seva situació geogràfica ha determinat que les biotes de totes tres comparteixin molts elements faunístics i florístics. És natural, si es pensa que el 36% dels gèneres de fanerògames de la seva flora procedeix de Madagascar, i que encara s’hi ha de sumar un altre 44% de gèneres comuns a l’Àfrica oriental i a Madagascar: el 80%, doncs, de la flora d’aquestes illes arribà de l’oest, seguint la trajectòria predominant dels corrents marins; de la resta de gèneres, el 20% arribà amb el corrent equatorial de terres molt més llunyanes (el 12% de la regió tropical indopacífica i només el 8% de la regió oriental).
Els isòpodes terrestres de l’illa de l’Ascensió són un altre bon exemple de la història dels colonitzadors de les illes remotes. Se n’han trobat 13 espècies, de les quals n’hi ha nou que han estat introduïdes amb el comerç des de regions tan llunyanes com l’Àsia sud-oriental, la Mediterrània, Sud-àfrica i l’Amèrica del Sud. Una desena espècie és de distribució pantropical, però es pot afirmar que arribà sense l’ajut humà, ja que és halòfila i es pot trobar sobre fustes flotants. Les tres espècies restants són endèmiques d’aquesta illa, però estan emparentades amb grups africans, com es podria esperar tenint en compte que l’illa de l’Ascensió es troba al bell mig del recorregut del corrent sud-equatorial atlàntic. Aquest corrent procedeix de les costes del golf de Guinea, on hi ha grans rius pels quals haurien pogut arribar a la mar els rais de vegetació que transportarien els avantpassats dels endemismes.
Els grans mons a part
Les parts emergides d’alguns fragments a la deriva de plaques continentals o de microplaques d’una certa extensió poden constituir grans mons a part, illes de grandària considerable, en algun cas gairebé comparables a microcontinents. Es tracta de terres que ja fa molt de temps que se separaren de les masses continentals a les quals estigueren unides i iniciaren la seva singladura per la mar. En són bons exemples Madagascar, Nova Zelanda, Nova Caledònia i el massís cirno-sard, que forma el que avui són les illes de Còrsega i Sardenya.
El cas de Madagascar: lèmurs i camaleons
Amb una àrea de 590 000 km2, 1 600 km de llarg i 580 km d’amplada màxima, Madagascar és l’illa oceànica més gran del món (la quarta en termes absoluts si comptem les illes continentals, les tres més grans de les quals, Grenlàndia, Nova Guinea i la Terra de Baffin, superen Madagascar en extensió). Separada del continent africà fa 165 milions d’anys pel que avui és el canal de Moçambic (que té una amplada que varia entre els 350 i els 1 200 km), es troba quasi enterament a la zona tropical, entre els 12 i els 26° de latitud S. El clima, però, no és homogeni, sinó que té una franja oriental molt humida, que és per això ocupada per una ampla varietat de selves tropicals, mentre que a la part occidental el clima és molt més sec, i a l’extrem meridional s’assoleixen unes condicions subdesèrtiques. És precisament en aquest extrem meridional on es troba la més elevada proporció d’endemismes vegetals de l’illa: el 95% de les espècies vegetals són endèmiques (al conjunt de l’illa la proporció és del 85%). A les àrides terres del sud i del sud-est malgaixos s’hi fan comunitats singulars de plantes suculentes, entre les quals es troben les didieràcies, una família endèmica de l’illa, que recorden les cactàcies per la seva morfologia però amb les quals no tenen cap relació.
La fauna i la flora de Madagascar provenen d’un nombre limitat de nissagues que colonitzaren l’illa en el passat. Dels mamífers, només les rates-pinyades, els insectívors (amb una família endèmica, la dels tenrècids, amb 21 espècies), els rosegadors (amb una única subfamília endèmica amb no menys de deu espècies vivents i dues o tres descomparegudes), els primats, els carnívors i els hipopòtams (amb dues espècies nanes, avui extingides) han format part de la fauna autòctona. La presència de carnívors autòctons, tanmateix, és una raresa en una illa. Entre els carnívors malgaixos cal destacar la fossa (Cryptoprocta ferox), un vivèrrid de grans dimensions que convergeix força amb els felins americans, en particular el puma, i és de fet el superdepredador terrestre de la fauna malgaixa. En el passat hi vivia una espècie propera, més gran, la fossa de les cavernes (C. spelaea), que devia depredar els grans pro-simis malgaixos, avui extingits.
Els lèmurs són un grup de primats endèmics de Madagascar, que havien estat molt nombrosos en el passat: sembla que eren una cinquantena d’espècies quan hi van arribar els humans. Hi havia des d’espècies de la mida d’un ratolí, com el lèmur ratolí (Microcebus murinus, d’uns 50 g de pes, el més petit dels primats), fins a espècies de la mida d’un goril·la (com l’extingit Archaeoindris fontoynonti, amb un pes estimat de més de 170 kg). Els lèmurs ocupaven una àmplia varietat d’hàbitats, des de la selva plujosa tropical fins a les formacions llenyoses obertes, talment com ho fan els primats superiors a l’Àfrica. Estaven representats per molts de tipus ecològics. Algunes espècies (Palaeopropithecus ingens, Babakotia radofilai) eren convergents amb els peresosos arborícoles sud-americans; altres (Megaladapis) ho eren amb els coales australians, encara que més grossos. N’hi havia de convergents amb els papions africans (Archaeolemur majori i A. edwardsi) mentre que els ei-ei (Daubentonia madagascarensis i l’extingit D. robusta) ocupen a Madagascar l’espai ecològic dels picots. Després de l’arribada dels humans, almenys 15 espècies de lèmurs s’han extingit, entre els quals es troben totes les de talla gran i d’hàbits especialitzats.
D’entre les 21 espècies de tenrècids, algunes, com el sokina (Setifer setosus) i el tambotriky (Echinops telfairi), són convergents amb els eriçons; altres, per exemple moltes espècies del gènere Microgale, amb les musaranyes; altres, com els voalavonarabo (Oryzorictes talpoides i altres espècies del mateix gènere), amb els talps. Plesiorycteropus madagascariensis, un mamífer extingit probablement als segles VIII o IX, que durant molts anys havia estat considerat una espècie pròxima al porc formiguer i posteriorment un insectívor emparentat amb els tenrècids, ha estat separat recentment (juny del 1994) en un ordre particular, els bibimalagasis, del qual seria l’únic gènere representant.
A Madagascar viuen més d’un centenar d’espècies endèmiques d’ocells entre ells els “vanga”, una família (vàngids) endèmica de l’illa amb una radiació adaptativa espectacular, comparable amb la dels pinsans de les Galápagos o la dels de les Hawaii: cada espècie té un bec característic, relacionat amb la seva alimentació. Entre els ocells extingits s’ha d’esmentar la família endèmica extinta dels epiornítids, els ocells-elefant (Aepyornis), ocells de gran talla, no voladors sinó corredors, similars als emús australians. L’espècie més gran, A. maximus, feia 3 m d’alçada i prop de 500 kg de pes, i ponia uns ous d’un volum de 12 l.
Entre els rèptils, destaquen les 56 espècies de camaleons malgaixos (dues terceres parts de les que viuen actualment arreu del món): 34 són camaleons típics (Chamaeleo), anomenats localment “tanala”, “sangorita” o “tarondro” i 22 són camaleons nans o “anjava” (Brookesia). També hi ha una gran diversitat de llangardaixos cordílids (12 espècies endèmiques), escíncids (46) i gecònids (més de 60), amb moltes espècies diürnes i de colors vistosos del gènere Phelsuma. Madagascar hostatja igualment 7 espècies endèmiques d’iguanes, unes quantes de serps (3 boes, 8 tiflòpids i 50 colúbrids) i algunes de tortugues. Es coneixen restes subrecents de tortugues gegants. La presència de boes i d’iguanes resulta sorprenent, perquè són grups absents del continent africà, i en canvi no es troben a Madagascar molts grups de rèptils ben representats a Àfrica (amfisbènids, agàmids, varànids, lacèrtids, elàpids i vipèrids). Entre els amfibis destaquen les 150 espècies de granotes.
A diferència del que s’esdevé a la majoria d’illes oceàniques, a Madagascar viuen peixos d’aigua dolça (28 espècies endèmiques a les quals cal sumar-ne 10 més de peixos eurihalins igualment endèmics). La majoria d’elles es troben en franca regressió, principalment a causa de l’alteració dels seus hàbitats i la introducció de peixos exòtics.
El cas de Nova Zelanda: kiwis i cucs gegants
Aïllades des de fa uns 80 milions d’anys, al Pacífic meridional, les dues illes de Nova Zelanda, unides i separades en diverses ocasions al llarg de la seva història, i que biogeogràficament componen una unitat, constitueixen el més extens d’aquests “mons a part” després de Madagascar. La fauna i la flora contenen molts elements originaris de l’antic continent de Gondwana i el nivell d’endemisme assoleix el 100% en molts grups.
Nova Zelanda difereix de Madagascar no sols per la mida més petita i la llunyania de tot continent i fins i tot de tota altra illa, sinó també perquè presenta un clima de caràcter temperat i ocasionalment subantàrtic, enfront de la zonació climàtica malgaixa, més càlida, que inclou zones semidesèrtiques i tropicals. Difereix d’altres illes oceàniques amb faunes extraordinàries amb les quals sovint és comparada (principalment Galápagos, Hawaii i Fiji) per la seva gran extensió (270 000 km2), per la seva antiguitat, pel seu clima i per haver sofert l’efecte de les glaciacions. Tal vegada el seu clima temperat i fred hagi impedit les grans radiacions evolutives trobades a les Hawaii.
A Nova Zelanda no hi ha mamífers terrestres autòctons, encara que sí una família endèmica de rates-pinyades, la dels mistacínids, amb dues espècies, Mystacina tuberculata i l’aparentment extingida M. robusta, els ancessors de les quals van colonitzar Nova Zelanda presumiblement fa uns 35 milions d’anys, des de l’Amèrica del Sud. Poden fer servir les ales com a potes anteriors, capacitades per a “caminar” amb una certa habilitat pels arbres i les roques. S’ha dit que tenen uns hàbits marxadors convergents amb els d’alguna mena de ratolí. Aquestes dues espècies, juntament amb el vespertiliònid Chalinolobus tuberculatus, són els únics mamífers no marins autòctons de Nova Zelanda. Mystacina tuberculata pol·linitza les flors de Dactylanthus taylorii, una balanoforàcia no fotosintetitzadora, paràsita de les arrels d’arbres i arbustos, l’única planta del món pol·linitzada per una rata-pinyada caminadora.
A diferència del que s’esdevé amb els mamífers, la diversitat és relativament elevada entre els ocells i els rèptils, dues classes de vertebrats que inclouen espècies amb una elevada capacitat de colonització d’illes mitjançant migracions ultramarines. Hi ha 62 espècies de rèptils, dues d’elles, els tuatares (Sphenodon punctatus i S. guntheri), d’una família (esfenodòntids) i un ordre (rincocèfals) endèmics de Nova Zelanda, mentre que les restants es reparteixen entre dues famílies, els escíncids i els gecònids. Els tuatares sobreviuen actualment només a 24 petites illes del golf Hauraki i de la badia de Plenty, al nord de l’illa Nord, i a les illes Stephens, North Brother i Trios, a l’estret de Cook. Hom havia cregut que només n’hi havia una espècie fins que el 1990 es va descobrir que la població de l’illa North Brother està molt diferenciada genèticament i representa una espècie particular (S. guntheri), en greu perill d’extinció. Els tuatares viuen pròsperament a illes que no han estat colonitzades per rates introduïdes i on crien ocells marins. Bé que poden excavar les seves pròpies llorigueres, fan servir les d’alguns petrells (per exemple, Pachyptila turtur). Són nocturns i poden viure més de 70 anys. Molts altres rèptils de Nova Zelanda són també força extraordinaris; els gecònids hi arribaren durant el Miocè i els escíncids durant el Plio-Plistocè, i hi van tenir una radiació important. L’únic exemplar conegut del dragó Hoplodactylus delcourtii fa 37 cm de llargària del musell a la cloaca, i és un dels gecònids més grans del món.
Fins a l’arribada dels humans a Nova Zelanda hi havia no menys de 89 espècies d’ocells terrestres, de les quals s’han extingit 36, és a dir, el 40%. Potser els més característics i extraordinaris eren els moes (Dinornis), 11 espècies d’ocells de talla entre mitjana i gegantina (3 m d’alçària l’espècie més gran, D. giganteus, d’uns 270 kg de pes), que desaparegueren amb l’arribada dels maoris, com també ho féu el seu depredador, l’àguila caçadora de moes o àguila de Haast (Harpagornis moorei). Altres ocells destacables són els kiwis (Apteryx), no emparentats amb els moes, ben al contrari del que es creia fins fa poc; són ocells corredors de la mida d’una gallina, d’hàbits nocturns, els únics del món que tenen un bec que funciona realment com un nas. En declivi a les illes principals, viuen més de 30 anys i en cada posta les femelles fan un únic ou d’una lliura de pes (proporcionalment enorme, ja que és més del 25% del pes de la mare), que coven durant quasi 3 mesos. S’ha suggerit que podrien ser descendents d’ocells voladors que, en romandre a Nova Zelanda, evolucionaren cap a formes sense capacitat de vol. El kakapo (Strigops hapbroptilus) és el més gran dels lloros (els mascles adults fan 2,5 kg), i l’única espècie no voladora del seu ordre; d’hàbits nocturns, en queden menys d’un centenar d’exemplars. Altres ocells particulars de Nova Zelanda són el weka (Capellirallus australis), la kea o lloro rapinyaire (Nestor notabilis) i el “takahe” o gall faver neozelandès (Porphyrio mantelli).
A Nova Zelanda, talment com a Madagascar, també trobem amfibis endèmics insulars, un element del qual solen estar mancades les illes oceàniques: les quatre espècies vivents i les tres aparentment extingides de granotes del ja esmentat gènere Leiopelma.
Però no sols els vertebrats són extraordinaris a Nova Zelanda. Entre els invertebrats abunden també els endemismes, i molts de singulars. A més dels ja esmentats wetas (Deinacrida) també hi ha cucs de terra gegants com Spenceriella gigantea, que fa prop d’1,5 m de llarg i més d’1 cm de diàmetre i cargols gegants (15 espècies dels gèneres Powelliphanta i Paryphanta). Grups sencers han radiat, originant grans nombres d’espècies endèmiques (cargols i llimacs, cucs de terra, planàries, papallones, escarabats, etc.).
El caràcter endèmic de la fauna i la flora neozelandeses, unit a la manca d’harmonia taxonòmica que d’una manera diferencial es dóna a cada grup, fa que les relacions ecològiques entre les diferents espècies sovint siguin també peculiars. La biologia reproductora de moltes plantes neozelandeses presenta moltes característiques inusuals: flors petites, d’estructura simple i gens vistoses, i fruits carnosos o glans. Entre les plantes d’origen arcaic es troben les winteràcies, que tenen les estructures reproductores i vegetatives més primitives de totes les angiospermes, cosa que les apropa als seus primitius i extints avantpassats. A banda el cas inusual de la pol·linització de Dactylanthus taylorii, per la rata-pinyada caminadora Mystacina tuberculata, algunes flors neozelandeses són pol·linitzades per saures, un fet insòlit al món, que només es repeteix a l’illa de Cabrera, a les Balears, gairebé als antípodes exactes de Nova Zelanda, on algunes plantes són pol·linitzades per la sargantana endèmica Podarcis lilfordi.
El cas de Nova Caledònia: araucàries i cagús
L’illa de Nova Caledònia és un dels més extraordinaris d’aquests grans mons a part. A 1 800 km al nord-oest de Nova Zelanda, amb 16 750 km2, hostatja una fauna i una flora quasi totalment endèmiques i d’origen molt antic.
N’és característica la gran diversitat i abundància de gimnospermes (43 espècies endèmiques i una de caràcter litoral indopacífic): es tracta de la concentració més gran d’espècies de coníferes que hi ha al món. De les 19 espècies d’araucàries (Araucaria) conegudes, totes de l’hemisferi meridional, 13 són neocaledonianes. L’anomenada araucària o pi columnar (Araucaria columnaris), abundant en algunes badies de Nova Caledònia i a les veïnes illes Loyauté, amb més de 50 m d’alçària, és l’arbre tradicional dels melanesis. Una podocarpàcia, Parasitaxus rustus, és l’única gimnosperma paràsita que es coneix al món. Dels 17 gèneres de palmeres que viuen a Nova Caledònia, 16 són endèmics, amb una trentena d’espècies, algunes en perill d’extinció. També s’hi troben falgueres gegants, per exemple les del gènere Cyathea, amb 7 espècies endèmiques, dues de les quals fan més de 25 m d’alçària. En total a Nova Caledonia es troben 1 575 espècies de plantes, amb un 89% d’endemismes.
La fauna, tan original com la flora, no té cap mamífer terrestre, encara que sí 6 espècies endèmiques de rates-pinyades. La fauna d’ocells també és altament endèmica i ha sofert l’onada d’extincions relacionada amb l’arribada dels humans, ja que el 40% de les espècies ha desaparegut. L’espècie més original de l’illa, Sylviornis neocaledoniae, exterminada pels primers pobladors melanesis i de la qual s’han descobert recentment restes, era un ocell no volador d’uns 30 kg i 1,6 m de llargària, que va ser descrit primer com un ocell corredor; més tard es va suggerir que era un gal·liforme molt modificat, un megapòdid gegant, gràcies a la descoberta de grans túmuls de terra (els megapòdids construeixen, per criar, túmuls de terra de grans dimensions), de fins a 40 m de diàmetre, que els arqueòlegs havien estudiat, intentant, sense èxit, trobar-hi traces d’enterraments humans. Entre els ocells vivents de l’illa cal destacar el cagú (Rhynochetos jubatus), que juntament amb l’extint cagú de terra baixa (R. orarius) formen una família d’ocells gruïformes endèmica de Nova Caledònia. El cagú, l’ocell emblemàtic de Nova Caledònia, és un ocell no volador, que nia a terra, on posa un únic ou; la seva població actual s’estima en uns pocs centenars d’individus. La fauna reptiliana patí durament l’impacte de l’arribada dels humans: una tortuga gegant banyuda (Meiolania mackayi), un cocodril terrestre (Mekosuchus inexpectatus), un varà de metre i mig i alguns gecònids i escíncids, dels 33 que hi havia inicialment, s’han extingit.
El grau d’endemisme també és extraordinari entre els invertebrats. Els cargols (més de 300 espècies) pertanyen a dues famílies endèmiques, una d’elles amb espècies carnívores. Les primitives papallones del gènere Sabatinca han radiat i han originat més de trenta espècies.
El cas de les Seychelles: un petit gran món a part
L’arxipèlag de les Seychelles, situat prop de l’equador al mig de l’oceà Índic occidental, constitueix un cas particular des del punt de vista biogeogràfic. Es tracta d’unes quaranta illes i illots, que tenen una superfície total d’uns 400 km2, escampades pel banc de les Seychelles, una plataforma de més de 43 000 km2 i menys de 100 m de fondària, voltada de profunditats que superen els 4 000 m. Són integrades per granits formats al Pre-cambrià a l’escorça continental, i en menor grau per altres roques ígnies d’origen més recent i afinitats més oceàniques. L’origen d’aquest microcontinent rau en el trencament de Gondwana, quan l’Índia derivà vers el nord i Madagascar s’allunyà de l’Àfrica. A mig camí entre l’Índia i Madagascar, aïllat des del començament del Terciari, fa uns 50 milions d’anys, quedà aquest petit món, en el qual l’evolució de la biota prosseguí sense més contactes amb la resta de les terres emergides. Des de llavors fins al seu descobriment per navegants àrabs, xinesos i europeus, només es produirien arribades ocasionals d’ocells i d’organismes nàufrags, com a les veritables illes oceàniques.
Les Seychelles hostatgen avui una flora i una fauna sorprenents per la seva composició i diversitat, malgrat les pertorbacions causades pels tres darrers segles d’ocupació humana. Les terres baixes estaven cobertes per selves de “bois-de-fer” (Vateriopsis [=Vateria] seychellarum), una dipterocarpàcia de grans dimensions de la qual resten actualment molt pocs exemplars (una cinquantena a l’illa de Mahé), habitades per abundants tortugues gegants (Dipsochelys) emparentades amb les que encara sobreviuen a Aldabra, mentre que als rierols hi havia cocodrils de grans dimensions; tots aquests rèptils gegants foren caçats fins a l’extinció.
La vegetació original es conserva només als cims i els vessants superiors de les muntanyes, que atenyen fins als 914 m d’altitud. De les 766 espècies de fanerògames catalogades, només 222 són nadiues, entre les quals es mantenen encara 69 endemismes. L’antiguitat de l’arxipèlag queda palesa en constatar l’abundància de gèneres endèmics: deu gèneres de fanerògames, entre els quals sis de palmeres. Cal remarcar el cas de Medusagyne oppositifolia, l’única espècie de la família de les medusaginàcies, un arbrissó raríssim del qual es coneixen avui una desena d’exemplars espontanis (creix bé en conreu a partir de llavor) però que s’havia donat per extingit del 1903 al 1970, quan fou trobada a l’illa de Mahé, a 250 m d’altitud, la població que ara es considera la darrera representació espontània de l’espècie. El cocoter de mar (Lodoicea maldivica) era conegut d’antic per les seves llavors, de fins a 20 kg de pes, que excepcionalment atenyen les costes índies; l’especialització insular és evident en aquesta espècie, que produeix pocs descendents, triga dos anys a germinar, necessita 25 anys per a produir els primers fruits i ha d’esperar-ne 7 entre la fertilització i la maduració del fruit. La vegetació culminal de les Seychelles també és peculiar, formada en bona part per pradells de plantes carnívores del gènere Nepenthes, els únics vegetals que poden créixer damunt mateix del dom de granit.
Els cargols terrestres són molt diversificats, però quasi tots estan limitats als ecosistemes originals i per tant es troben en perill d’extinció, o ja extingits. La majoria de les espècies de l’arxipèlag pertanyen a gèneres endèmics, de manera que les seves afinitats taxonòmiques amb altres regions són febles, i repartides entre Madagascar i l’Índia. Més de la meitat de les espècies d’insectes, conegudes encara molt parcialment, són endemismes; la majoria de les espècies nadiues estan associades íntimament amb les plantes autòctones, per la qual cosa es troben només a les zones més abruptes.
Un dels trets més sorprenents de les Seychelles és la presència de nombrosos amfibis, un grup absent de totes les altres illes remotes. Hom ha descobert quatre espècies de cecílies, un grup poc conegut d’amfibis subterranis àpodes que recorden cucs de terra, d’origen molt antic i afinitats desconegudes. Una altra família endèmica és la dels sooglòsids, formada per tres espècies de petites granotes arborícoles (Nesomantis thomasseti, Sooglosus gardineri i S. seychellensis). Una altra granota arbòria endèmica (Tachycnemis seychellensis) és un representant força primitiu de la família dels hiperòlids, comuna amb Madagascar i Àfrica. El catàleg d’amfibis es completa amb un rànid introduït en temps històrics.
A part els cocodrils i les tortugues gegants, entre els rèptils hi ha escíncids endèmics (notablement els del gènere Mabuya) molt divergents respecte als seus parents continentals. Només una espècie de camaleó i les tortugues d’aigua dolça mostren una afinitat africana. Hom coneix 15 espècies d’ocells endèmics, tres de les quals (i segurament moltes més) són extingides i la majoria de la resta es compten entre els més amenaçats del món. Curiosament, un nectarínid s’ha adaptat als ambients humanitzats, on proliferen els jardins amb flors. Els mamífers autòctons són dues espècies endèmiques de rates-pinyades: una insectívora i l’altra frugívora.
Els megaarxipèlags intercontinentals
A les zones de fricció entre les grans plaques continentals s’han generat grans conjunts d’arxipèlags amb roques continentals i volcàniques. Les seves faunes i flores solen ser estranyes mescles d’elements provinents dels continents que els envolten. Els exemples més notables són les illes mediterrànies, les Antilles (Petites Antilles, Grans Antilles i Bahames) i les illes orientals oceàniques d’Insulíndia (Filipines —llevat de Palawan i Calamian, que són la prolongació continental de Borneo—, Cèlebes, Moluques, Timor, Flores, etc.).
Les illes mediterrànies: sargantanes a dojo
La Mediterrània és com un oceà en miniatura, amb corrents marins, àrees de productivitat elevada i illes tant continentals com paraoceàniques. Les illes mediterrànies paraoceàniques, amb una extensió conjunta de més de 80 000 km2, tot i haver estat en contacte amb les terres circumdants, tenien, fins que els humans les van colonitzar, unes faunes i unes flores que rivalitzaven, pel seu caràcter extraordinari, amb les de les illes oceàniques. De clima àrid i marcadament estacional, tot i ser riques en endemismes, no suporten una diversitat biològica tan elevada com les Antilles o les Filipines. Tots els mamífers terrestres autòctons de les illes mediterrànies al Plistocè, unes 45 espècies, eren endèmics. Hi havia elefants nans (Elephas falconeri) d’un metre d’alçada a la creu, hipopòtams nans (Phanourios) de la mida d’un porc, cérvols nans (Praemegaceros) amb una alçada a la creu d’uns 50 cm, un gòral nan (Myotragus) amb ulls frontals i incisives de rosegadors, rates cellardes gegantines (Leithia, Hypnomys), etc.
Les illes mediterrànies no han estat prou aïllades per a la majoria d’ocells. Pocs grups s’han especiat originant endemismes insulars, i els més espectaculars ja han desaparegut: un duc nan (Bubo insularis) que va viure a Còrsega i Sardenya, i un cigne gegant no volador (Cygnus falconeri), a Malta. Els pocs endemismes insulars mediterranis que han sobreviscut són petits passeriformes, com la tallareta sarda (Sylvia sarda), de Còrsega, Sardenya, Mallorca i Eivissa, la tallareta de Xipre (S. melanothorax) i els pica-soques Sitta whiteheadi, de Còrsega, i S. krueperi, de diverses illes gregues. Són ocells que han evolucionat conjuntament amb la vegetació mediterrània i dispersen les llavors dels fruits carnosos de diverses espècies, com ara llentiscles (Pistacia), llorers (Laurus), aladerns (Phillyrea) o oliveres (Olea).
A les illes mediterrànies s’ha produït una petita diversificació insular a les sargantanes del gènere Podarcis. Una desena d’espècies diferenciades en alguns centenars de subspècies, amb formes melàniques, cianòtiques, verdes, marrons, de disseny variable, robustes, gràcils, de potes curtes, de potes llargues, etc., escampades per totes les illes de la conca reflecteixen una bona varietat d’adaptacions als diferents medis microinsulars, que són més heterogenis del que semblen de cop d’ull. També hi ha algunes espècies relictes miocèniques, com la sargantana de Bedriaga (Archaeolacerta bedriagae) o l’algiroide nan (Algyroides fitzingeri), ambdós de Còrsega i Sardenya. Les illes mediterrànies estan mancades d’ofidis autòctons. No obstant això, Còrsega i Sardenya són de les poques illes del món que tenen urodels, concretament salamandres cavernícoles (4 espècies del gènere Speleomantes [=Hydromantes] que viuen a diferents massissos càrstics) i tritons (2 espècies del gènere Euproctus). Aquestes espècies són testimonis de les antigues connexions de la microplaca cirno-sarda, i no de colonitzacions ultramarines.
També trobem alguns amfibis anurs a les illes mediterrànies: alguns discoglòssids (com els ferrerets Alytes muletensis de Mallorca i l’extint A. talaioticus de Menorca, i els calàpets pintats (Discoglossus sardus i D. montalentii) de Còrsega i Sardenya), que assoliren les illes en el moment de la dessecació de la Mediterrània, i un hílid, la reineta sarda (Hyla sarda), un amfibi els ancestres del qual arribaren a Sardenya mitjançant migració ultramarina.
Les Antilles: llangardaixos, huties i almiquins
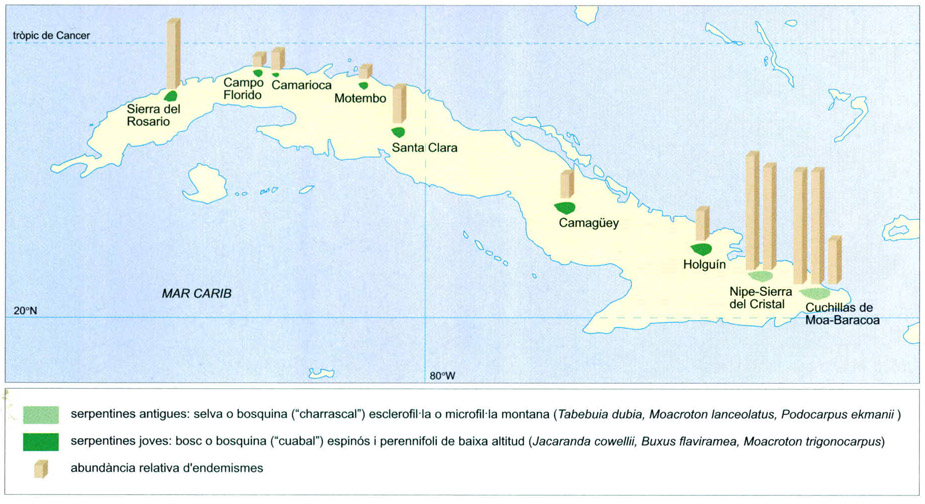
Les situacions de doble insularitat incrementen l’endemicitat, com és el cas, a Cuba, de la vegetació dels afloraments de serpentines (roques amb un alt contingut de magnesi, generadores de sòls oligotròfics). La vegetació serpentinícola, de caràcter espinós, microfil·le o esclerofil·le, forma illes enmig de la vegetació basòfila, perennifòlia estacional o semicaducifòlia, de les calcàries predominants a l’illa. Per aquest doble aïllament, els claps serpentinícoles presenten uns índexs d’endemicitat elevadíssims. El mapa mostra la distribució d’aquests claps, i l’abundància relativa d’endemismes, proporcional a la llargada de les barres.
Editrònica, a partir de Borhidi, 1993
Fent de pont entre l’Amèrica del Nord i l’Amèrica del Sud, les Antilles tenen una història geològica complexa, encara no del tot aclarida. Consten de tres arxipèlags principals, les Grans Antilles, les Petites Antilles i les Bahames, amb una superfície total d’uns 240 000 km2. Es troben a la zona tropical, amb temperatures altes i relativament uniformes al llarg de l’any. Les Grans Antilles són d’origen antic, si bé encara està en debat si mai estigueren unides als continents o no; el cert és que per a comprendre la natura de les seves biotes no es fa necessari postular antigues connexions, de tan particulars com són. Les Petites Antilles no semblen tan antigues i tenen volcans actius. Les Bahames, illes coral·lines, planes i situades en una extensa zona d’aigües poc profundes (el banc de les Bahames), han sofert uns canvis més grans en la seva extensió des de la darrera glaciació fins a l’actualitat: han vist reduïda la seva àrea de 129 000 km2 a 11 000 km2.
La fauna de les Antilles és en bona part endèmica. Abans de l’arribada dels humans hi havia unes 90 espècies de mamífers terrestres no voladors, tots endèmics (més d’un 80% de les quals s’han extingit), a més d’unes 45 espècies de rates-pinyades (amb una família endèmica). Hi havia 12 espècies d’insectívors, 18 de peresosos (tant terrestres com arborícoles), entre 4 i 7 de simis, i més de 55 de rosegadors, principalment huties (Capromys i gèneres propers). Dels insectívors, que pertanyien a dues famílies molt primitives (solenodòntids i nesofòntids), n’han sobreviscut els almiquins (Solenodon cubanus, l’almiquí cubà, i S. paradoxus, l’almiquí de la Hispaniola). De peresosos n’hi havia des de terrestres de talla relativament gran, com Megalocnus rodens, de Cuba, amb una dentició molt diferenciada, fins a arborícoles de talla petita, com els actuals peresosos sud-americans; no n’ha sobreviscut cap. Hi havia simis a Cuba, a la Hispaniola i a Jamaica, on vivia la mona de Jamaica (Xenothrix mcgregori), d’afinitats desconegudes, cap dels quals no ha sobreviscut. Els rosegadors havien sofert una radiació adaptativa de la qual derivaren espècies arbòries que ocupaven nínxols similars als de simis i esquirols, altres de més terrestres que ocupaven nínxols similars als dels porcs espins i d’altres igualment terrestres però de talla gran. Els aparells mastegadors d’aquests rosegadors es relacionaven amb la seva radiació adaptativa, i s’han descrit diferents models de masticació. Sobreviuen només un dotzena d’espècies d’huties, la majoria a l’illa de Cuba.
Entre els ocells de les Antilles destaca una família endèmica, la dels tòdids, d’afinitats desconegudes, amb 5 espècies. Hi ha unes 160 espècies d’ocells endèmics de les Índies occidentals, i un mínim de 40 més han desaparegut d’ençà de l’arribada dels humans, entre elles mussols gegantins (fins a 1,5 m d’alçada) de cames llargues, òlibes de gran talla, rasclons no voladors, caracaràs, falcons buteonins gegantins, pinsans, falziots, etc. La diversitat actual dels ocells de les Antilles és una miniaturització de la biodiversitat real que assoliren en el passat.
La fauna herpetològica de les Antilles és d’una diversitat extraordinària, molt més gran que la que es troba en qualsevol altre arxipèlag del món. Hi viuen actualment més de 480 espècies de rèptils i amfibis. Hi són representades sis famílies de saures, principalment gecònids i iguànids, i en concret dos gèneres: Anolis, un gènere d’iguànids amb una interessant radiació evolutiva, de la qual han resultat 110 espècies i Sphaerodactylus, un de gecònids amb prop de 80 espècies. Hi ha també 41 espècies de colúbrids, boes de les Antilles (Epicrates) i uns quants cocodrils. Entre els anurs hi ha estranys gripaus cuirassats (Peltophryne) i més de 110 espècies d’Eleutherodactylus.
La Wallàcia: rates, ratolins i tarsis
Entre les regions biogeogràfiques australiana i oriental es troben diversos arxipèlags d’illes oceàniques que no se situen exactament ni a la plataforma de la Sonda ni a la de Sahul. Les illes situades fora d’aquestes plataformes presenten faunes i flores altament endèmiques, que no es poden assignar totalment, des del punt de vista biogeogràfic, ni a la regió oriental o indomalaia com les de la plataforma de la Sonda, ni a l’australiana com Nova Guinea i les altres illes de la plataforma continental de Sahul.
Aquest veritable megaarxipèlag intercontinental, al qual s’aplica sovint el nom de Wallàcia en record del naturalista que per primera vegada reconegué l’especificitat de la regió, comprèn, al nord, les illes Filipines (excepte Palawan i Calamian, que representen una prolongació de Borneo cap al nord-est), al centre, les Cèlebes, les Moluques i illes properes, i, més cap al sud, la cadena formada per Lombok, Flores, Arum i Timor. En aquestes illes s’han produït radiacions d’espècies, s’han originat estranys endemismes i grups taxonòmics sencers no hi han arribat. Com en el cas d’altres megaarxipèlags intercontinentals, l’origen de la seva fauna és mixt, amb espècies provinents tant de la regió australiana com de l’oriental.
La diversitat de rates i ratolins posa de manifest la radiació que han sofert a Wallàcia els múrids (més de cent espècies endèmiques en total, de les quals no menys de 46 a les Filipines, i no menys de 36 a Cèlebes). N’hi ha espècies molt grans, altres de minúscules, algunes d’arborícoles, altres d’insectívores, i també n’hi ha de frugívores, de granívores i de Polífagues.
Els súids van colonitzar diferents illes, i hi originaren formes endèmiques: el senglar de les Cèlebes (Sus celebensis) i l’extingit Celebochoerus heekeneri, a les Cèlebes; Sus philippensis i S. cebifrons, considerades antigament subspècies respectivament del porc comú (S. scrofa) i del porc barbat (S. barbatus), a les Filipines; S. heureni i S. timorensis, també considerades subspècies del porc comú, respectivament a Flores i a Timor. Talment com en algunes illes mediterrànies, a les Filipines i les Cèlebes es troben espècies nanes de bòvids, els anoes de terra baixa (Bubalus depressicornis) i muntanya (B. quarlesi) a les Cèlebes, i el tamamaw (B. mindorensis) a les Filipines. Algunes illes (Timor, Flores, Cèlebes) foren colonitzades per estegodonts, que originaren formes nanes insulars, totes desaparegudes ja fa temps.
A les Filipines i les Cèlebes viuen quatre espècies endèmiques de tarsis (Tarsius), uns primats de relacions filogenètiques encara no prou aclarides, com també altres primats més moderns, i ara set espècies de Macaca, només a les Cèlebes. A les Filipines es troben dermòpters endèmics, que són una de les preses favorites de l’àguila menjamones (Pithecophaga jefferyi), també endèmica.
Els grans arxipèlags oceànics
Sovint, sobre punts calents del mantell terrestre s’han originat progressivament, al llarg de milions d’anys, arxipèlags d’illes volcàniques. Aquests arxipèlags són molt diferents, per l’origen i pels materials que els formen, dels mons originats per la fragmentació antiga de l’escorça i inclouen illes de diferent antiguitat, de vegades ordenades precisament segons l’edat. En són exemples les Hawaii, les Galápagos, les Canàries, les illes del Cap Verd, i molts altres arxipèlags aïllats.
El cas de les illes Canàries: rates de “malpaís” i coloms de laurisilva
Les illes Canàries formen un arxipèlag volcànic d’uns 7 500 km2 separat poc més de 100 km de les costes africanes. Hi ha 7 illes grans, habitades des de fa prop de 2 000 anys, i altres illes menors. Les illes més antigues són les més properes al continent, Fuerteventura i Lanzarote, amb una edat de més de 25 milions d’anys, i les més recents són Hierro i La Palma, amb una edat estimada de menys de 2 milions d’anys. La flora i la fauna de les illes és altament endèmica.
Les quatre illes més properes al continent arribaren a ser colonitzades per mamífers terrestres. Hi ha hagut com a mínim dues espècies de rates gegants (Canariomys), una rata de “malpaís” (Malpaisomys insularis) i dues espècies de musaranyes (Crocidura) endèmiques, però només sobreviuen aquestes darreres.
Entre els ocells endèmics destaquen el pinsà blau (Fringilla teydea), els coloms de laurisilva (Columba junoniae), la “paloma rabiche”, de Tenerife, La Palma i La Gomera, i C. bollii, la “paloma turqué” d’aquestes mateixes illes), el bitxac de Fuerteventura (Saxicola dacotiae) i la garsa de mar negra (Haematopus meadewaldoi), recentment extinta. Altres dues espècies tenen un estatut taxonòmic confús: una òliba petita (Tyto alba gracilirostris) i un reietó (Regulus regulus teneriffae). Es coneixen dues altres espècies d’ocells terrestres (una guatlla de cames llargues i una cadernera), i dues baldrigues desaparegudes. Com a les Galápagos, també hi ha un aligot endèmic (bé que en aquest cas és una subspècie).
Un dels elements més extraordinaris de la fauna vertebrada canària són els llangardaixos del gènere endèmic Gallotia (derivat probablement de les sargantanes de cua llarga del gènere Psammodromus). L’espècie més gran de les quatre vivents, G. simonyi, sobreviu actualment en uns penya-segats de l’illa de Hierro, però es creu que havia estat escampada per totes les illes occidentals de l’arxipèlag. L’extingit G. goliath (probablement conspecífic amb G. simonyi) era el lacèrtid més gran i robust del món (arribava a fer 1,40 m de llargària total), i sembla que era una menja molt apreciada pels guanxes, la primera població humana de les illes. A les Canàries també s’han produït radiacions específiques d’altres rèptils (gecònids del gènere Tarentola i escíncids del gènere Chalcides).
El cas de les illes Hawaii: pinsans, drosòfiles i cargols
Hawaii, l’arxipèlag oceànic més aïllat de tot el món, és format per 8 illes principals i 124 illetes, amb un total de 16 759 km2 i es prolonga, sota les aigües del Pacífic, en la cadena de muntanyes submarines Emperor. Les illes hawaianes s’arrengleren seguint una línia de quasi 2 700 km, pràcticament en la direcció dels paral·lels. Les més grans i modernes se situen a l’extrem oriental i com més a l’oest són més antigues i, en conjunt, més petites. Les terres continentals més properes, les costes californianes, es troben situades a uns 3 000 km de l’illa gran de Hawaii. És un arxipèlag relativament jove; les illes principals de l’arxipèlag, les orientals, tenen menys de 5 milions d’anys (l’illa gran de Hawaii en té menys d’un!). Les petites illes occidentals, quasi totalment desmantellades, tenen edats molt superiors. Amb un clima subtropical, les Hawaii contenen la fauna més disharmòniques de tot el món.
A les Hawaii no es troben mamífers terrestres, ni rèptils, ni amfibis. Només hi ha una espècie de rata-pinyada, autòctona, però no endèmica, Lasiurus cinereus, arribada des de l’Amèrica del Nord. En canvi, han estat el paradís dels ocells: l’ornitofauna terrestre originària, provinent d’una vintena de colonitzacions des de diferents continents i des d’altres illes polinèsiques, era endèmica en un 100%. Ja s’ha fet esment de la radiació evolutiva dels pinsans. No han estat, però, els únics ocells que han experimentat una radiació semblant. A les illes de Molokai i Maui, per exemple, hi van viure uns ibis no voladors, els Apteribis, convergents amb els kiwis de Nova Zelanda. Es coneixen quatre espècies extintes de moa-nalos, ànecs de gran talla, no voladors, i amb uns becs peculiars. El més estrany dels moa-nalos era Chelychelynichen quassus, que tenia un bec similar al d’algunes tortugues. També hi havia com a mínim una dotzena d’espècies de rasclons no voladors; però només dues, ara extingides, sobrevisqueren fins a temps històrics; totes les altres s’extingiren de resultes de l’arribada dels primers humans, els polinesis, i la fauna associada amb ells, abans que arribessin els europeus. També els depredadors havien de ser ocells: dues espècies d’aligots (Buteo); una arpella (Circus dossenus), convergent amb els esparvers (Accipiter), ornitòfags com ella; la temible àguila marina (Haliaaetus), i quatre espècies de mussols del gènere endèmic Gallistrix. De tots ells només sobreviu actualment l’anomenat aligot de Hawaii (Buteo solitarius).
L’originalitat de la fauna de les Hawaii també és palesa entre els invertebrats. S’estima que el 99% dels insectes (més de 10 000 espècies conegudes) són endèmics. Se suposa que deriven d’uns 400 immigrants. Molts grups han radiat considerablement. Un gènere de papallones, Hyposmocoma, inclou més de 220 espècies endèmiques. De drosòfiles (Drosophila) n’hi ha més de 700, producte de dues colonitzacions. De mol·luscs terrestres n’hi ha més de 1 200 espècies, que poden haver derivat d’una vintena d’espècies colonitzadores. També entre els invertebrats hawaians, n’hi ha que han evolucionat adaptant estils de vida insòlits per al seu grup. Així, les larves d’una espècie d’odonat zigòpter, Megalagrion oahuense, són les úniques del món que no són aquàtiques sinó terrestres, i viuen als sòls dels boscos humits de muntanya. Un grill del gènere Caconemobius ha deixat de ser terrestre per adoptar un estil de vida parcialment marí. Les erugues del gènere de lepidòpters Eupithecia han deixat de ser herbívores, per transformar-se en depredadors a l’aguait.
El cas de les illes Galápagos: tortugues, iguanes i pinsans
Situades a 1 000 km de les costes de l’Amèrica del Sud, arran de l’equador, les illes Galápagos, amb 9 illes principals i 10 de menors i una superfície total d’uns 8 000 km2, constitueixen l’arxipèlag oceànic que més ben conservat ha arribat fins als nostres dies. Aparentment mai no foren colonitzades pels indis sud-americans ni pels polinesis. La primera referència coneguda del seu descobriment es deu a fra Tomás de Berlanga, bisbe de Panamà, que les visità el 1535. Els avatars històrics les privaren d’una colonització primerenca, i fins el 1832 no s’hi establí cap assentament humà significatiu, fet que contribuí a salvaguardar la seva natura. Encara avui (1994) la seva població no arriba a les 10 000 persones. Algunes de les illes grans estan deshabitades i no tenen mamífers introduïts, un fet actualment excepcional a les illes oceàniques.
L’origen de l’arxipèlag és recent. Les illes més antigues, les orientals, s’estima que tenen entre 3 i 5 milions d’anys, bé que algunes datacions recents suggereixen que ja hi havia alguna illa fa 9 milions d’anys. Les més recents són probablement les grans illes occidentals (Isabela i Fernandina), que sembla que van emergir fa menys d’un milió d’anys. Les Galápagos foren poblades totalment per una fauna immigrant ultramarina, part de la qual, com ara els pinsans que Darwin féu famosos, va sofrir radiacions adaptatives.
A les Galápagos hi ha 625 espècies vegetals, de les quals més de 200 són endèmiques. La vegetació és dominada per plantes cactiformes com les del gènere Opuntia, que s’ha diversificat molt, i inclou algunes formes gegants (fins a 4 m d’altura i amb troncs de més d’1 m de diàmetre en algun cas). Els fruits d’aquestes plantes (figues de moro) són consumides per alguns vertebrats de les Galápagos, com ara les tortugues gegants o les iguanes terrestres. També mereixen ser destacades algunes espècies arbòries de Scalesia un gènere de compostes endèmic la radiació adaptativa del qual (14 espècies al conjunt de les illes), encara que menys coneguda i espectacular que la dels pinsans, no és pas menys significativa. Com a curiositat, la tomaquera de les Galápagos (Lycopersicon cheesmanii), es pot regar amb aigua de la mar.
Les Galápagos van sofrir tres invasions de rates i ratolins abans de la darrera, provocada per la implantació humana. Una va donar origen a un gènere de rates de mida grossa (Megaoryzomys), actualment extingides. Una segona invasióva originar un grup de rates més petites típiques dels camps de lava, amb algunes espècies vivents de les illes de Santa Cruz (Nesoryzomys darwini), Fernandina (N. fernandinae, N. indefessus), Santiago (N. swarthi) i d’altres de recentment extingides de les illes de Santa Cruz, Isabela, Baltra i Rábida. Una tercera invasió és representada per una espècie endèmica de rata arrossera (Oryzomys galapagoensis).
A diferència de la majoria d’arxipèlags oceànics, no sembla que l’arribada dels humans hagi suposat a les Galápagos la pèrdua de cap espècie d’ocell. Una de les espècies més notables que hi viuen, sense cap paral·lelisme vivent, és el corb marí de les Galápagos (Compsohalieus [=Nannopterum] harrisi), no volador, del doble de pes que els seus congèneres voladors i molt més bon cabussador que ells (pot arribar als 45 m de profunditat); proveït d’unes ales petitíssimes, és l’única espècie vivent que ens pot donar una idea de com devien ser els famosos Hesperornis del Cretaci. A més dels famosos pinsans, a les Galápagos, hi crien 10 espècies autòctones més d’ocells terrestres, entre les quals es troben els dos principals depredadors de petits vertebrats, l’aligot de les Galápagos (Buteo galapagoensis) i la petita òliba de les Galápagos (Tyto punctatissima). Hi ha una tórtora endèmica (Zenaida galapagoensis); quatre espècies endèmiques de mímids, els “cucuves” (Nesomimus); un ràl·lid, el “pachay” (Laterallus spinolotus); un muscicàpid, el papamosques de les Galapágos (Myiarchus magnirostris), i un hirundínid, l’oreneta de les Galápagos (Progne modesta).
Les Galápagos són un veritable paradís per als rèptils. Foren precisament uns rèptils els que els donaren el nom i la fama en èpoques passades. Durant els segles XVII i XVIII, les illes foren visitades per corsaris i pirates, que sabien que hi obtindrien aigua dolça i carn de tortuga. Hi havia poblacions nombroses de tortugues gegants (Geochelone elephantopus) a quasi totes les illes, bé que actualment són abundants sobretot al volcà Alcedo, a l’illa d’Isabela.
Altres rèptils notables de les Galápagos són les iguanes marines i les terrestres. Les iguanes marines (Amblyrhynchus cristatus), que arriben a fer 1,20 m de llarg, són dels pocs saures que han estat capaços de colonitzar les aigües costaneres; de règim vegetarià, com les terrestres, s’alimenten d’algues i poden beure aigua de la mar, gràcies a unes glàndules que els permeten excretar l’excés de sal. Les iguanes terrestres (Conolophus subcristatus i C. pallidus) són molt menys nombroses: només queden dues poblacions intocades, les de l’illa Fernandina (malgrat els efectes d’una devastadora explosió volcànica recent) i les de Plaza Sur, a l’illa Santa Cruz; les poblacions restants semblen estar en declivi i algunes ja han desaparegut.