Consideracions generals

Els boscos han esdevingut als nostres ulls l’exemple per antonomàsia de l’èxit de la natura: cap altre ecosistema no iguala la seva capacitat d’acumular biomassa i de modificar les condicions ambientals. Malgrat la dominancia que hi assoleixen els vegetals arboris, els boscos són ecosistemes complexos i no mers conjunts d’arbres. Els boscos cobreixen encara més d’una quarta part dels Països Catalans. Aquesta avetosa de la mata de València (Vall d’Àneu) és una bella mostra del nostre patrimoni forestal.
Fototeca/MC
Els boscos són ecosistemes terrestres dominats per arbres. Malgrat la pressió humana, ocupen encara el 40% de la superfície de les terres emergides, si descomptem els deserts extrems i els gels permanents. Des de les pluviïsilves equatorials fins a la taigà trobem boscos en un ventall molt ample de climes i sòls. De fet, podem esperar que un tipus o altre de bosc domini qualsevol paisatge on l’estació vegetativa no sigui gaire curta i on la disponibilitat d’aigua no sigui inferior a cert llindar.
Els boscos destaquen de la resta dels ecosistemes per la seva gran acumulació de biomassa: la biomassa total (aèria més subterrània) d’un bosc madur pot superar les 300 tones de pes sec per hectàrea, xifra unes 30 vegades superior a la biomassa de la majoria dels ecosistemes herbacis i unes 1000 vegades més gran que la dels ecosistemes planctònics. Al mateix temps, els boscos són els ecosistemes terrestres més productius, amb uns valors de producció neta mitjana que varien de 8 a 20 t/ha·any segons els biomes. Recordem que, en comparació, la producció neta mitjana dels conreus s’ha estimat en 6,5 t/ha·any, i la dels oceans, en 1,5 t/ha·any. El grau de producció forestal només és igualat o superat per alguns ecosistemes aquàtics o terrestre-aquàtics especialment productius, com ara estuaris, esculls i aiguamolls.
L’èxit ecològic que han tingut els ecosistemes forestals no és pas recent. Boscos de pteridòfits gegants existien ja al Devonià mitjà superior, fa uns 370 milions d’anys, i dominaren els paisatges pantanosos del Carbonífer (fa entre 345 i 280 milions d’anys). La major part dels jaciments de carbó actuals són el resultat de l’acumulació de matèria orgànica deguda a la producció d’aquests boscos del passat. Posteriorment, les gimnospermes, i entre elles les coníferes, dominaren els boscos del Triàsic i el Juràssic (fa entre 225 i 136 milions d’anys). A partir del Cretaci mitjà, fa uns 90 milions d’anys, les angiospermes es diversificaren i s’expandiren, i els boscos d’arbres amb flors relegaren les coníferes als llocs menys favorables.
La forma de vida arbòria
La forma de vida arbòria ha estat assajada amb èxit per tots els grans grups de plantes vasculars. Els boscos són ecosistemes terrestres dominats per arbres. Malgrat la pressió humana, ocupen encara el 40% de la superfície de les terres emergides, si descomptem els deserts extrems i els gels permanents. Des de les pluviïsilves equatorials fins a la taigà trobem boscos en un ventall molt ample de climes i sòls. De fet, podem esperar que un tipus o altre de bosc domini qualsevol paisatge on l’estació vegetativa no sigui gaire curta i on la disponibilitat d’aigua no sigui inferior a cert llindar.

Els arbres caducifolis, com aquests bedolls (Betula pendula) en fullatge tardoral, renoven cada any totes les fulles d’una manera més o menys simultània. D’aquesta manera l’arbre té fulles verdes només quan la llum, la temperatura i la disponibilitat d’aigua permeten una elevada producció neta, i queda desproveït de fulles durant l’època desfavorable o de repòs. Per contra, els arbres perennifolis conserven fulles verdes durant tot l’any, les quals han de poder resistir les condicions adverses de fred o d’eixut que es presentin.
Fototeca/MC
Els boscos destaquen de la resta dels ecosistemes per la seva gran acumulació de biomassa: la biomassa total (aèria més subterrània) d’un bosc madur pot superar les 300 tones de pes sec per hectàrea, xifra unes 30 vegades superior a la biomassa de la majoria dels ecosistemes herbacis i unes 1000 vegades més gran que la dels ecosistemes planctònics. Al mateix temps, els boscos són els ecosistemes terrestres més productius, amb uns valors de producció neta mitjana que varien de 8 a 20 t/ha·any segons els biomes. Recordem que, en comparació, la producció neta mitjana dels conreus s’ha estimat en 6,5 t/ha·any, i la dels oceans, en 1,5 t/ha·any. El grau de producció forestal només és igualat o superat per alguns ecosistemes aquàtics o terrestre-aquàtics especialment productius, com ara estuaris, esculls i aiguamolls.
L’èxit ecològic que han tingut els ecosistemes forestals no és pas recent. Boscos de pteridòfits gegants existien ja al Devonià mitjà superior, fa uns 370 milions d’anys, i dominaren els paisatges pantanosos del Carbonífer (fa entre 345 i 280 milions d’anys). La major part dels jaciments de carbó actuals són el resultat de l’acumulació de matèria orgànica deguda a la producció d’aquests boscos del passat. Posteriorment, les gimnospermes, i entre elles les coníferes, dominaren els boscos del Triàsic i el Juràssic (fa entre 225 i 136 milions d’anys). A partir del Cretaci mitjà, fa uns 90 milions d’anys, les angiospermes es diversificaren i s’expandiren, i els boscos d’arbres amb flors relegaren les coníferes als llocs menys favorables.
El disseny bàsic d’un arbre consta d’un sistema de rels que fixa l’arbre al terreny i que absorbeix l’aigua i els nutrients del sòl; d’un element columnar, el tronc, que serveix de suport i de transport; i d’un aparell fotosintetitzador enlairat, la capçada. El disseny anterior va generalment associat a la capacitat de créixer fins a assolir grans proporcions, tant en alçada com en diàmetre, i a una longevitat considerable, sovint gairebé de segles. El «General Sherman», un exemplar de Sequoiadendron giganteum, té un pes estimat en unes 2030 tones, equivalent a uns vint rorquals blaus. Exemplars d’arbres de diferents espècies (Sequoia sempervirens, Pseudotsuga menziesii, Eucalyptus regnans) s’acosten als 100 m d’alçada o els superen; el fet d’haver resolt els problemes d’estabilitat mecànica i de transport d’aigua fins al cim d’una tal capçada representa una de les fites evolutives més sorprenents. En les dures condicions d’una carena a 3280 m d’altitud, en les White Mountains de Califòrnia, un tortuós exemplar de Pinus longaeva té una edat de 4900 anys, segons indiquen els seus anells de creixement; era ja un arbre madur quan es construïren les grans piràmides. Moltes espècies d’arbres poden arribar als 500 anys d’edat.
Els arbres dels Països Catalans queden lluny dels rècords anteriors, però així i tot moltes espècies poden assolir edats i dimensions ben respectables, si les condicions són favorables i l’home no els talla abans. Un avet majestuós en una obaga pirinenca pot tenir 300 anys d’edat, fer 30 m d’alçada i tenir un tronc d’un metre de diàmetre. Un arbre així pot pesar, sense les rels, unes 5 tones de pes sec, biomassa que podria equivaler a la d’unes 50 000 plantes herbàcies més aviat robustes o a la de 1016 (deu mil bilions) algues unicel·lulars del plàncton. La importància que això té per al funcionament dels ecosistemes respectius es posa de manifest quan pensem que la biomassa d’aquest únic avet seria la de tota una hectàrea d’un herbei alt, o la del fitoplàncton de 500 000 000 m3 d’una aigua més aviat pobra, com la de la Mediterrània.
Els avantatges de la forma de vida arbòria són diversos: una capçada elevada permet rebre i utilitzar la llum abans que no ho facin altres vegetals de talla menor; una grandària considerable i una vida llarga afavoreixen l’ocupació permanent de l’espai, un recurs essencial per als organismes sèssils; un sistema radicular horitzontalment extens i més o menys profund permet explorar un gran volum de sòl i garanteix el subministrament d’aigua i de nutrients. Nogensmenys, ser un arbre també té els seus inconvenients: els arbres alts són vulnerables al llamp, als vents forts i a les tempestes de neu; la gran acumulació de biomassa morta que representa la fusta central dels troncs ha de ser defensada dels organismes descomponedors; els arbres grossos tenen grans quantitats de teixits vius que respiren però no fotosintetitzen (càmbium, floema, parènquima de l’albeca, rels), la qual cosa grava el balanç de carboni de l’arbre; la longevitat va associada a un temps mitjà de generació llarg i a una taxa d’augment poblacional que és petita comparada amb la de vegetals més petits i de vida més curta. En termes ecològics, els avantatges de la forma de vida arbòria han pesat més que els inconvenients, com ho demostra la dilatada dominància dels arbres en tots aquells paisatges on les condicions són més favorables per a la vida dels vegetals. En termes evolutius, el balanç no és tan nítid. Només cal recordar que, entre les angiospermes, les plantes herbàcies són més evolucionades que les llenyoses, i que els arbres constitueixen només un percentatge moderat de totes les espècies actuals de plantes vasculars. Als Països Catalans, per exemple, hi ha unes 85 espècies d’arbres autòctons o naturalitzats (incloent-hi espècies com ara l’arboç, el llentiscle, el margalló i molts altres que generalment són arbustius, però que poden assolir port arbori); això representa menys del 3% de la flora vascular del nostre territori, estimada en 3500 espècies.
L’estructura i l’ambient forestals
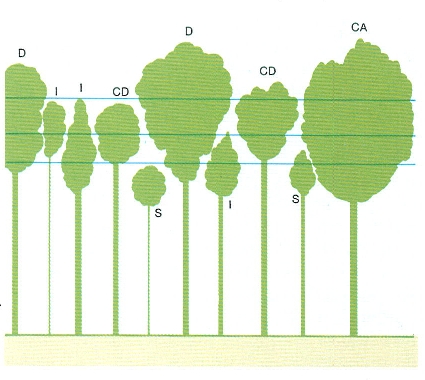
Classes d’arbres segons la seva posició en l’estrat arbori: D dominant, CD codominant, CA crescut aïllat, I intermedi, S suprimit. Els arbres de les tres primeres classes estan ben il·luminats i poden créixer. Els arbres intermedis i els suprimits estan ombrejats pels altres i el seu creixement és petit o nul. La classificació és d’H.W. Hocker.
Josep Nuet i Badia.
Com a la resta dels ecosistemes terrestres, l’element que domina l’estructura dels boscos és la vegetació. Segons la seva alçada, les plantes d’un bosc poden disposar-se en estrats més o menys diferenciats. Pot haver-hi un o més estrats arboris que ombregen amb intensitat variable els vegetals no arboris, els quals formen el sotabosc. Aquest pot estar format d’un o més estrats arbustius, d’un estrat herbaci i un estrat muscinal, a més de plantes enfiladisses i d’epífits. És clar que molt rarament els trobarem tots en un mateix bosc. Cadascun dels estrats pot ser horitzontalment continu o discontinu.
La disposició i la continuïtat dels estrats
Si el bosc és obert, és a dir si les capçades dels arbres veïns no es toquen entre elles, arribarà a terra una quantitat important de llum i el sotabosc podrà ser abundant; els boscos tancats, sobretot si són d’arbres de capçada espessa, deixen passar molt poca llum fins a terra (almenys durant l’estació vegetativa) i el sotabosc acostuma a ser ben minso. La major part dels nostres boscos són oberts. Uns perquè són boscos joves que encara estan colonitzant camps o pastures abandonats durant les darreres dècades. Altres, a les contrades més seques, perquè no hi ha prou aigua disponible per a mantenir un estrat arbori tancat. Altres, perquè el bosc s’està regenerant després d’alguna pertorbació natural o induïda per l’home. I gairebé tots, perquè el règim predominant de gestió es basa en el sistema d’aclarides, amb torns de tala de 15 a 30 anys segons les espècies i els llocs; això manté obert cada rodal durant gran part del temps. En aquestes condicions no és estrany que molts dels nostres boscos tinguin un sotabosc notable i format sovint per vegetals que no toleren l’ombra. Aquest sotabosc pot ser predominantment herbaci o arbustiu. Del primer cas tenim, com a exemples més destacats, els fenassars de les pinedes mediterrànies, les joncedes de rouredes i pinedes submediterrànies o els prats d’ussona dels boscos assolellats de pi negre. Del segon cas tenim, entre altres, les brolles caldcoles o silicícoles de pinedes mediterrànies, alzinars i suredes; les boixedes submediterrànies; o les catifes denses però baixes de neret a les obagues silícies de pi negre.

Els boscos són sistemes dinàmics. Molts dels boscos dels Països Catalans no existien fa cent anys, s’han originat per regeneració natural de camps o pastures abandonats. A la fotografia, ginebres (Juniperus communis) i alzines (Quercus ilex) colonitzen el falguerar d’una clariana en l’estatge de la fageda al Montseny. Quan finalment s’hi estableixi el faig (Fagus sylvatica), la densa ombra d’aquest farà que ginebrons i alzines minvin fins a morir, tot i que alguns exemplars poden persistir molt de temps. Els estrats i la composició floristica d’un bosc depenen en part de la seva història.
Ferran Rodà
De vegades el sotabosc pot assolir un desenvolupament extraordinari, com succeeix als terrenys silicis de la Serralada Litoral i de la Serralada Prelitoral. Aquí, sota un estrat arbori obert, típicament de pi pinyer o de surera, hi ha una massa de bruc boal i arboç que, si no ha estat estassada durant els darrers vint anys, arriba als 5 o 6 m d’altura. Un sotabosc així pot tenir una biomassa igual o superior a la del mateix estrat arbori. En aquestes condicions, i també amb un sotabosc menor, és molt probable que arbres i arbustos competeixin per l’aigua i els nutrients, i que la producció dels primers augmenti si s’elimina el sotabosc, tal com ha estat demostrat en altres països.
En els boscos tancats, la situació és ben diferent. El terra pot estar cobert principalment de fullaraca, i les poques plantes que trobem al sotabosc poden ser d’espècies que toleren l’ombra. Aquestes són típicament herbàcies i, a causa de la poca radiació disponible, estan «afamades de carboni»; acostumen a disposar les fulles horitzontalment per interceptar la màxima radiació possible, i en una sola capa perquè no s’ombregin les unes a les altres. En els boscos caducifolis, alguns geòfits fugen de l’ombra avançant el seu cicle a la sortida de les fulles dels arbres. Per tancat que sigui un bosc, hi ha una certa proporció de taques de sol, degudes a forats entre les branques o entre els arbres; aquest mosaic de taques de sol permet l’existència en el sotabosc de plantes que no toleren gaire l’ombra.
Molts dels nostres boscos tancats, ho són de fa poc temps; sovint en el sotabosc hi ha plantes heliòfiles que havien crescut quan el bosc era obert o quan encara no n’hi havia. Aquestes plantes malviuen en el bosc tancat i, si no s’obren noves clarianes, al final es moren. Per exemple, a les fagedes del Montseny és freqüent trobar-hi troncs morts de ginebró, una espècie poc resistent a l’ombra; això ens indica que aquestes fagedes eren més obertes en el passat o que el faig ha colonitzat no fa gaires dècades una artiga o d’altres espais oberts.
La composició i les dimensions de l’estrat arbori
Un bosc alt o gran és el que formen els arbres nascuts de llavor. Un bosc baix o menut és el que formen els arbres rebrotats de soca o de rel. Diem que un bosc és regular si tots els arbres tenen la mateixa edat, o irregular si els arbres són d’edats diverses. Les nostres coníferes no rebroten en ser tallades o cremades, de manera que sempre faran bosc alt. En canvi, els planifolis acostumen a rebrotar o tanyar després de la tala, i els trobarem fent bosc alt o bosc baix segons que el règim d’explotació afavoreixi els arbres de llavor o de rebrot. Els boscos regulars es generen amb tallades arreu, seguides de reforestació natural o artificial; es poden generar també, però, per pertorbacions naturals que destrueixen tot un rodal, seguides de regeneració sincrònica. Els boscos irregulars s’originen per aclarides periòdiques, si aquestes són prou intenses perquè hi hagi regeneració; ho poden fer també de manera natural si el règim de pertorbació és tal que ha permès diverses etapes de regeneració. Els tipus de bosc més estesos als Països Catalans són el bosc alt irregular i el bosc baix irregular. Com a bosc alt irregular, es gestionen les bones suredes, algunes rouredes, les fagedes i la major part de pinedes de tota mena i avetoses; i com a bosc baix irregular, els alzinars a les terres barcelonines i gironines i la major part de les rouredes. Com a bosc alt regular es porten les pinedes de pinastre, les plantacions de pi insigne, molts boscos de pinassa i alguns boscos de pi roig, de pi negre i d’avet; i com a bosc baix regular es menen les castanyedes, i els alzinars a les terres meridionals.
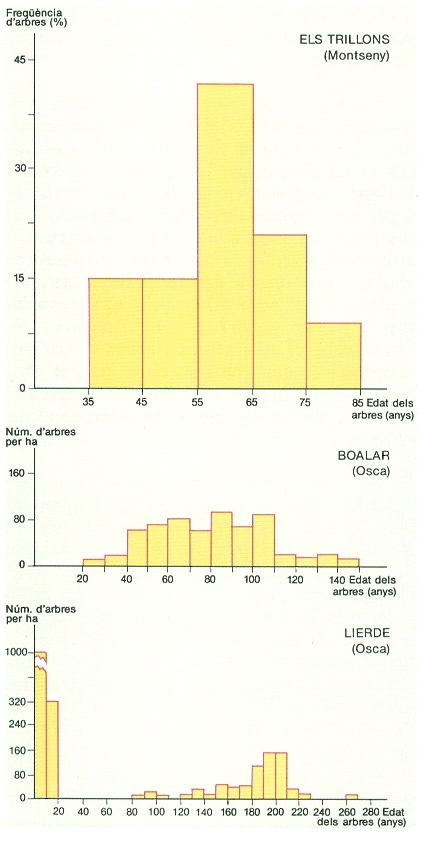
Estructures d’edats en tres boscos de coníferes: de dalt a baix, pi roig (Pinus sylvestris) al Montseny, i pi roig i pi negre (P. uncinata) als Pirineus d’Osca. Les dades procedeixen d’una parcel·la de 0,1-0,25 ha en cada bosc. Els gràfics ens diuen les proporcions d’arbres de diferents edats, determinades pels anells de la fusta. Cap de les estructures d’edats correspon a una població estable. És a dir, la composició per edats canviarà forçosament amb el temps als tres boscos, perquè en tots les classes d’edat més abundants són seguides per classes d’arbres més joves i menys abundants. Als Trilions i a Boalar no hi ha hagut una regeneració eficaç en els darrers 20 o 35 anys. A Lierde hi ha una regeneració abundant els darrers 20 anys, però no n’hi va haver durant els 60 anys anteriors. La regeneració polsant és molt freqüent en els boscos. La història del bosc i el tipus d’aprofitament forestal deixen la seva empremta en l’estructura d’edats dels arbres.
Carto-Tec, amb dades de F. Rodà i J. Puigdefábregas.
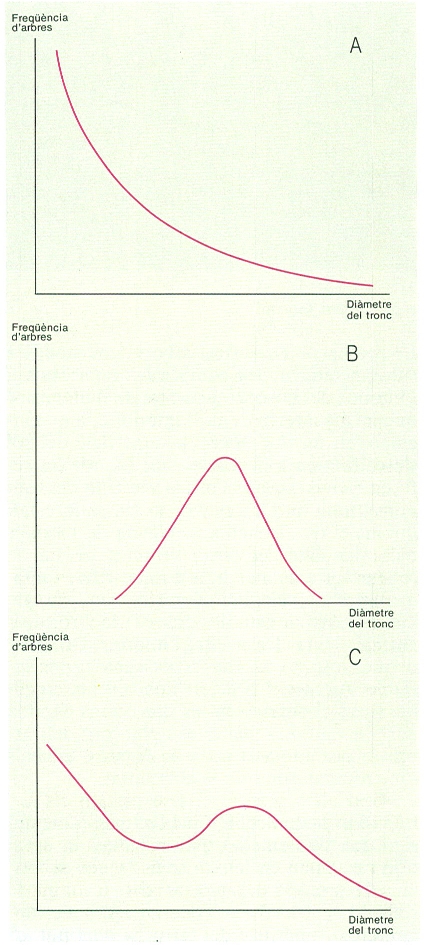
La composició diamètrica d’un bosc ens diu les proporcions d’arbres de diferents diàmetres de tronc, mesurats a 1,3 m del terra. La figura il·lustra tres estructures idealitzades de diàmetres: A disminució progressiva d’abundància des dels arbres petits fins als grans; B màxima abundància d’arbres de diàmetres propers a la mitjana; i C distribució bimodal. La situació A podria indicar un bosc amb regeneració continuada, o un bosc regular on sobreviuen els arbrets suprimits. La situació B és típica dels boscos regulars, però també es pot donar en un bosc irregular sense regeneració i amb mortalitat dels arbres suprimits. La situació C pot resultar de l’existència de pulsacions de regeneració o del creixement diferencial de dues classes d’arbres; per exemple, els que formen el sostre del bosc estan il·luminats i creixen, i els que no formen el sostre del bosc estan ombrejats i queden petits. A causa de les diferències de creixement i supervivència, l’estructura de diàmetres no sempre reflecteix l’estructura d’edats d’un bosc.
Carto-Tec, original de l’autor.
Les coníferes representen les tres quartes parts dels nostres boscos; la quarta part restant és de planifolis. Aquesta relació només s’inverteix a les comarques gironines, on els planifolis ocupen una mica més de la meitat de la superfície arbrada. El pi blanc és, amb molta diferència, l’espècie forestal més estesa ja que els boscos dominats per aquesta espècie representen el 33% de la nostra superfície boscosa (dades dels anys seixanta). Els incendis dels darrers 20 anys han afectat particularment els boscos de pi blanc, sobretot a les terres tarragonines i al País Valencià, de manera que és possible que la representació d’aquesta espècie hagi disminuït. Pel que fa als planifolis, els boscos més estesos són els d’alzina o carrasca, que ocupen conjuntament un 10% de la superfície arbrada.

La major part dels boscos dels Països Catalans són fortament monospecífics, és a dir, estan dominats per una sola espècie d’arbre, tot i que no hi falten exemples de boscos mixts. Entre les mescles arbòries més freqüents tenim les constituïdes per una o més espècies de pins amb una o més espècies de Quercus, com aquest bosc de pi roig (Pinus sylvestris) i roure martinenc (Quercus pubescens), en els contraforts del Montsent, al Berguedà.
Fototeca/MC
Les nostres masses forestals són fortament monospecífiques, pel que fa als arbres dominants. Un 73% de tota la superfície boscosa és netament dominada per una o altra espècie d’arbre. De fet, estem molt acostumats que els nostres boscos estiguin formats gairebé per una única espècie d’arbre; les expressions del tipus «bosc de faig» bé ho recullen. Però aquesta situació, malgrat ser compartida amb la major part d’Europa, no deixa de ser sorprenent. Per contrast, alguns boscos temperats de caducifolis de l’E de l’Amèrica del Nord i del Japó contenen 20 o 30 espècies d’arbres en una sola hectàrea. Tot i que les flores arbòries d’aquestes darreres regions són força més riques que la nostra (potser en part perquè les extincions d’arbres durant les glaciacions pleistocèniques van ser més nombroses a Europa), no podem deixar de preguntar-nos fins a quin punt el caràcter generalment monospecífic dels nostres boscos és un fet natural o ha estat, al contrari, el resultat de l’actuació de l’home sobre boscos més mixtos que els actuals, que ha afavorit en cada cas intencionadament o inadvertidament una de les espècies arbòries sobre les altres.
La grandària d’un arbre es reflecteix en les seves diferents dimensions, les més útils de les quals són l’alçada, el diàmetre del tronc i el diàmetre de la capçada. No tots els arbres d’un rodal tenen les mateixes dimensions. Arbres propers d’una mateixa espècie poden diferir bé perquè no tenen la mateixa edat, bé perquè han crescut a velocitats diferents. Per tant, trobarem determinades estructures o distribucions de grandàries no sols en els boscos irregulars sinó també en els regulars. Les velocitats de creixement d’arbres veïns de la mateixa edat poden no ser les mateixes a causa principalment de les diferències en els genotips, en el grau de competència experimentat per cada arbre o en la qualitat del sòl.
La distribució de freqüències dels diàmetres dels troncs (mesurats a l’alçada convencional d’1,3 m del terra o diàmetre normal) és un dels aspectes bàsics de l’estructura del rodal. Aquesta distribució o composició diamètrica es pot utilitzar per a calcular la biomassa i la producció del bosc. D’altra banda, l’estructura de diàmetres d’un rodal en un cert moment resulta d’una sèrie de processos dinàmics que inclouen tant el creixement dels arbres actuals com la mortalitat d’arbres preexistents i la posible aparició de nous individus. Però cal recordar que l’estructura d’edats pot ser ben diferent de l’estructura de diàmetres. Un arbre petit no ha de ser necessàriament un arbre jove: pot ser un arbre d’edat considerable que hagi crescut poc.
L’ambient forestal

El microclima forestal pot ser estudiat amb l’ajut de torres d’instrumentació com aquesta, situada en una parcel·la experimental a la fageda de Santa Fe de Montseny. Al llarg de la torre poden instal·lar-se sensors que mesurin les variacions de llum, temperatura, humitat relativa de l’aire i velocitat del vent que tenen lloc a diferents alçades, la qual cosa permet conèixer els efectes de les capçades sobre el medi físic. La torre pot utilitzar-se també per agafar mostres de fulles i branques a diferents alçades i estudiar-ne les variacions anatòmiques o fisiològiques segons la posició que tenen dins l’arbre. La figura 218 és un exemple de la informació que es pot obtenir gràcies a aquest tipus de torres.
Ferran Rodà
La presència d’un estrat arbori dens modifica substancialment les condicions ambientals a l’interior del bosc: la quantitat de llum i calor que arriba a terra és molt inferior a la que incideix en un lloc obert; la superfície del sòl s’escalfa menys durant el dia, i també es refreda menys durant la nit; la velocitat del vent minva molt en l’interior de la massa forestal; durant el dia, la humitat relativa de l’aire és més alta dins del bosc, sobretot perquè la temperatura és més baixa; aquest fet, juntament amb la velocitat menor del vent, fa que l’evaporació directa des del sòl i la virosta sigui més lenta. En resum, l’interior d’un bosc dens és fosc, fresc, humit, poc ventós i poc variable. Aquestes característiques del microclima forestal defineixen les condicions de vida per a les pròpies plàntules i plançons dels arbres, i per a tota la resta de plantes, animals i microorganismes de la comunitat.
La raó principal del fet que l’ambient forestal sigui tan diferent del que hi ha en un espai obert es troba en les característiques del balanç de radiació dels boscos. En un sòl nu, la reflexió i l’absorció de l’energia radiant es donen en la superfície del terra, la qual pot arribar a escalfar-se molt per damunt de la temperatura de l’aire. En un ecosistema herbaci o arbustiu la superfície activa en els bescanvis de radiació passa a ser la part superior de les plantes, encara no gaire allunyada de terra; de fet, més que d’una superfície, es tracta d’un volum d’uns 20 a 50 cm de gruix, en el qual es concentren la major part de les fulles. En un bosc tancat, la zona on hi ha més intercepció de la radiació és l’extrem superior de les capçades, que pot estar, per exemple, a 10 m del terra i tenir un gruix de 2 o 3 m. Els sòls forestals es troben, per tant, distanciats del punt de l’ecosistema on té lloc l’intercanvi de radiacions.
Els boscos es diferencien d’altres ecosistemes no sols per aquest enlairament de la superfície de radiació activa sinó per la destinació de la radiació incident. El percentatge de radiació solar reflectida per les capçades d’un bosc depèn de la longitud d’ona, de l’angle d’incidència de la radiació, de les propietats òptiques de les fulles i de la quantitat i disposició de fulles i branques. En general, els boscos tenen un albedo més petit (reflecteixen menys radiació) que altres tipus de vegetació. Els valors típics d’albedo dels ecosistemes forestals són entre el 5 i el 15% de la radiació d’ona curta incident. L’albedo és mínim en boscos densos de coníferes de fullatge fosc, com el pi negre, i més gran en boscos de fullatge clar, boscos amb clarianes, caducifolis sense fulla o boscos nevats. Amb cel serè, les capçades de l’alzinar de Le Rouquet (prop de Montpeller) reflecteixen un 12% de la radiació d’ona curta incident; però aquest percentatge resulta d’una reflexió diferencial segons la longitud d’ona: es reflecteix un 20% de l’infraroig i només un 4% de la radiació fotosintèticament activa.
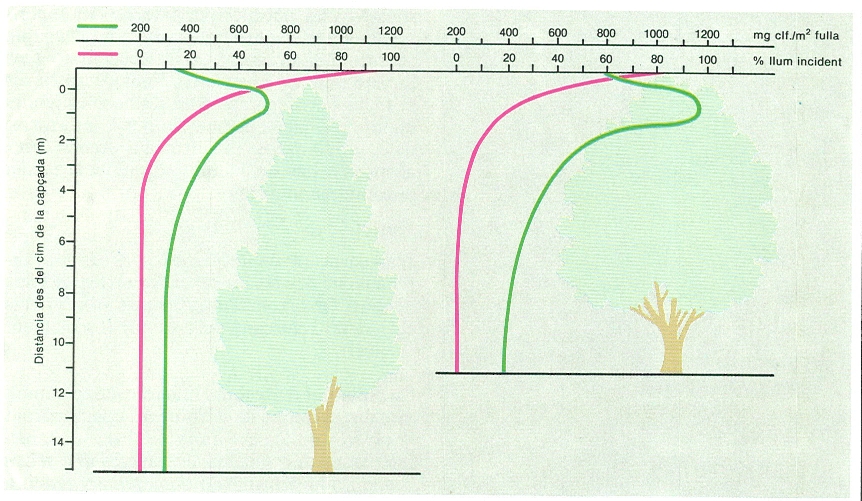
Perfils verticals de llum (en vermell) i clorofil·la (en verd) en una fageda (esquerra) i un alzinar (dreta) del Montseny. A mesura que descendim a través de les capçades, la intensitat de llum disminueix exponencialment: a 2 m per sota de l’extrem de les capçades, només hi arriba el 20% de la radiació fotosintèticament activa que hi ha a l’exterior. La concentració de clorofil·la a les fulles té un màxim destacat en aquest nivell. Les variacions són semblants en ambdós boscos, però la concentració de clorofil·la és molt més alta a l’alzinar.
Josep Nuet, amb dades de C. Gracia.
Un bosc dens transmet fins a terra un percentatge molt petit de la radiació incident. En alzinars i fagedes tancats, a les taques d’ombra, hi arriba sovint menys del 2% de la radiació fotosintèticament activa que hi ha a l’exterior del bosc. La manca de llum esdevé així un factor limitant de primer ordre per a les plantes del sotabosc. Altres boscos, per exemple els de coníferes de capçades poc espesses, com el pi blanc, deixen passar un percentatge més alt de radiació.
Si els boscos reflecteixen i transmeten poca radiació, necessàriament han d’absorbir-ne molta. En efecte, segons el tipus de bosc i la quantitat de clarianes, els arbres absorbeixen del 50 al 90% de la radiació global incident. Hi ha una absorció preferent de les longituds d’ona visibles: per exemple, l’alzinar de Le Rouquet absorbeix el 77% de la radiació fotosintèticament activa, però només el 54% de l’infraroig. Això implica que la radiació que arriba a l’interior del bosc és no sols quantitativament escassa sinó espectralment diferent, amb un gran predomini de l’infraroig.
Les capçades poden absorbir una quantitat tan gran de radiació, a causa de la seva complexa estructura. La superfície que intercepta l’a radiació és subdividida en milers de fulles i branques de diferents orientacions que propicien les reflexions múltiples, de manera que un foto incident té moltes possibilitats de ser finalment absorbit. Les capçades dels arbres són trampes eficaces de radiació.
Si les capçades absorbeixen un percentatge elevat de radiació sembla que s’haurien d’escalfar molt. Realment, en un dia de sol les temperatures màximes del bosc se solen trobar a la part subterminal de les capçades, on el fullatge és dens. No obstant això, les capçades s’escalfen molt menys del que esperaríem per tres motius diferents. Primer, la zona d’absorció de radiació no és una superfície sinó un volum d’alguns metres de gruix. Segon, pel fet que l’àrea foliar total és molt retallada en petits elements interceptors, s’afavoreix enormement la dissipació convectiva de calor, mitjançant la qual l’aire refreda les capçades escalfades pel sol. Tercer, una part important de la radiació absorbida s’utilitza per a evaporar aigua en la transpiració de les fulles. L’energia radiant invertida en la transpiració d’un bosc pot representar el 25% de la radiació solar rebuda durant tot l’any. L’eficàcia combinada d’aquests mecanismes de refrigeració dóna com a resultat que les capçades assolellades no s’escalfin gaire per damunt de la temperatura de l’aire i que fins i tot puguin estar per sota de la temperatura d’aquest, si hi ha suficient aigua en el sòl per a poder mantenir una elevada transpiració, tal com s’ha comprovat en estudis detallats duts a terme a l’alzinar del Montseny.
L’economia hídrica en els ecosistemes forestals

Boira estival a la fageda. A les muntanyes mediterrànies el dèficit estival de pluja és ben apreciable fins i tot a les altituds on es fan les fagedes. Les boires estivals produïdes per la marinada són freqüents en aquestes muntanyes, sobretot als vessants marítims, i hom diu que la persistència de les fagedes a les muntanyes mediterrànies es deu a les boires. La boira permet al faig reduir la transpiració i estalviar aigua, però a aquest avantatge cal contraposar l’inconvenient d’una fotosíntesi neta disminuïda per la feble intensitat lumínica mentre dura la boira.
Ferran Rodà
Com tots els ecosistemes terrestres, els boscos necessiten quantitats més o menys grans d’aigua per a subsistir. Quan hi ha abundant aigua disponible en el sòl, els arbres d’un bosc poden transpirar 40 o 60 tones diàries d’aigua per hectàrea. Les entrades d’aigua en un bosc permeten que l’absorció per les rels compensi les pèrdues hídriques per transpiració, mantenint la turgència cel·lular i l’activitat dels arbres. Excepte per als boscos de ribera, la precipitació constitueix la principal entrada d’aigua en els ecosistemes forestals.
Als Països Catalans, la pluviositat anual mitjana de les zones forestades varia des d’uns 400 mm en els boscos més secs de pi blanc fins a 1500 mm en alguns boscos subalpins. Gran part dels nostres boscos reben una precipitació mitjana inferior als 800 mm anuals. Si tenim en compte la gran variabilitat interanual i la forta irregularitat estacional de les precipitacions en els nostres climes, amb un eixut estival més o menys acusat, les xifres anteriors impliquen que la disponibilitat d’aigua deu ser el principal factor limitant en la major part dels nostres boscos, particularment a la terra baixa i als sectors no especialment plujosos de la muntanya mitjana.
La distribució de la pluja en el bosc
Pèrdues per intercepció
Una part de l’aigua precipitada damunt d’un bosc s’evapora directament des de les capçades molles sense arribar a terra: és el que s’anomena pèrdues per intercepció. En pluges poc abundoses (2) la major part de la precipitació pot evaporar-se des de les capçades. Amb pluges progressivament més abundants, el percentatge d’aigua que és interceptada i evaporada disminueix ràpidament, però en canvi la quantitat total de pèrdues per intercepció augmenta. Aquestes pèrdues seran més importants com més llargues i menys intenses siguin les precipitacions. Els boscos densos de coníferes, amb fulles aciculars i índexs elevats d’àrea foliar, experimenten pèrdues per intercepció particularment elevades.
En termes anuals, les pèrdues per intercepció en un bosc representen sovint entre un 10% i un 30% de la precipitació. Per tant, l’interior del bosc rep al cap de l’any força menys aigua que un espai obert adjacent. El règim de pluges dels Països Catalans, on un percentatge important de la precipitació anual cau en pocs dies, afavoriria que les pèrdues per intercepció fossin menors que en climes atlàntics o centreuropeus, on les pluges cauen menys concentrades en el temps. Malgrat això, l’única xifra d’intercepció que es coneix als nostres boscos no és gaire baixa: l’alzinar de Prades intercepta un 18% de la precipitació anual.
L’estrat arbori no és l’únic que pot interceptar aigua. Si al bosc hi ha estrats arbustiu, herbaci o muscinal ben desenvolupats, caldrà sumar-hi les pèrdues per intercepció experimentades en cada estrat. Fins i tot la capa de virosta té una capacitat d’intercepció no gens menyspreable. El resultat final de totes les pèrdues per intercepció en un bosc és que la quantitat d’aigua disponible per a infiltrar-se en el sòl, i eventualment per a drenar o ser absorbida per les plantes, disminueix en un percentatge important.
Trascol i escolament cortical
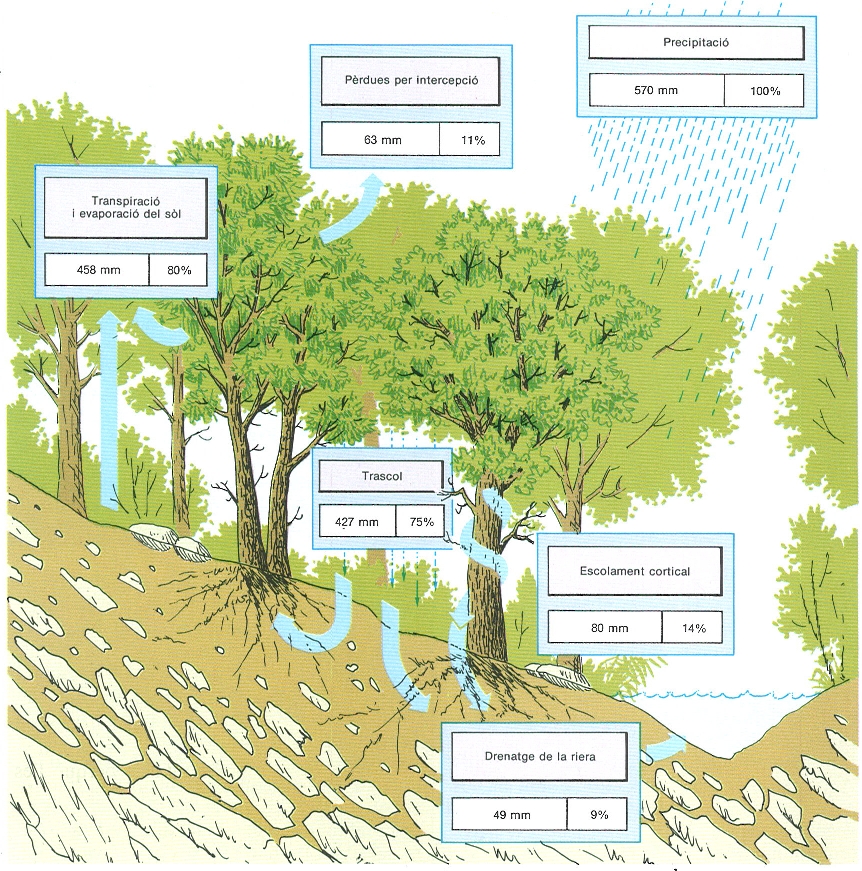
Hidrologia forestal d’un alzinar a les muntanyes de Prades. La figura quantifica les principals vies de circulació de l’aigua en un alzinar dens de rebrot. La magnitud de cada flux hidrològic és la mitjana de diversos anys d’estudi, i s’expressa en mil·límetres (o l/m2) per any i en percentatge respecte la precipitació. La pèrdua per intercepció és la quantitat d’aigua de precipitació que s’evapora directament des de les capçades o els troncs, sense arribar a terra. El trascol és l’aigua que degota a través dels arbres, mentre que l’escolament cortical és l’aigua que arriba a terra relliscant pels troncs. El drenatge s’ha mesurat en una conca de 51 ha de litologia impermeable i coberta tota ella pel mateix tipus d’alzinar. L’evapo-transpiració anual d’aquest bosc (incloent-hi les pèrdues per intercepció) és el 91% de la precipitació rebuda, xifra que posa de manifest la importància de l’aigua com a factor limitant en els boscos mediterranis i el paper crucial d’aquests en el cicle hidrològic.
Román Montull, amb dades de J. Bellot, M.J. Lledó, J. Piñol i A. Escarré.
L’aigua de precipitació que no s’evapora directament des de les capçades arriba al terra del bosc bé degotant a través dels arbres (trascol) o bé relliscant pels troncs (escolament cortical).
La quantitat d’escolament cortical depèn molt de l’arquitectura de l’arbre, de la grandària i la posició de la seva capçada, i de les característiques de l’escorça. Els arbres que tenen branques horitzontals o descendents generen poc o gens escolament cortical; és el cas de moltes coníferes. Els volums més grans d’escolament cortical són produïts per arbres de branques ascendents, troncs rectes i escorces llises, com el faig crescut en un bosc dens: un faig dominant de 30 cm de diàmetre normal pot reportar més de 100 litres d’escolament cortical en una pluja de 40 mm. Les alzines generen també bastant escolament cortical, sobretot si compleixen les condicions anteriors. En canvi, la major part dels roures ho fan menys abundosament, perquè solen tenir les branques més horitzontals i perquè l’escorça clivellada dificulta el flux de l’aigua. En base anual, l’escolament cortical pot representar des de menys de l’1% fins al 15 o 17% de la precipitació, segons el tipus de bosc: per exemple, a l’alzinar de Prades, amb una alta densitat de peus més aviat petits, l’escolament cortical és un 13% de la precipitació anual; a l’alzinar de Le Rouquet (prop de Montpeller), menys dens i amb peus més grans i més tortuosos, és només del 4%. El paper de l’escolament cortical probablement resulta subestimat en els anteriors percentatges, que consideren tota la superfície del bosc. Però l’aigua de l’escolament cortical va a parar a una àrea molt restringida al voltant de la base de cada arbre. L’efecte és el d’un rec localitzat de considerable intensitat. Aquesta aportació concentrada d’aigua (i nutrients) pot ser important per als arbres que la reben, ja que pot modificar les propietats del sòl i generar àrees saturades productores d’escorriment subsuperficial.
El trascol és la fracció quantitativament més important de la precipitació ja que representa entre el 60% i el 85% de la precipitació anual fora del bosc. Podem considerar que és l’aigua precipitada que queda després que se n’evapori una part des de les capçades i una altra rellisqui pels troncs. Per tant, tots aquells factors que tendeixin a augmentar les pèrdues per la intercepció o l’escolament cortical tendiran a disminuir el trascol, i viceversa. Als tres alzinars que hom ha estudiat en detall (Prades, Montseny i Le Rouquet), el trascol és només el 65-69% de la precipitació anual; això vol dir que a la major part del bosc no directament afectada per l’escolament cortical només arriben a terra els dos terços de l’aigua que plou anualment. El trascol es reparteix d’una manera molt heterogènia dintre del bosc i això obliga a instal·lar un nombre elevat de pluviòmetres (més de 20) per conèixer amb certa fiabilitat la quantitat mitjana que representa. Aquestes xarxes de pluviòmetres permeten estudiar la distribució espacial de la quantitat de trascol. Alguns punts recullen molt poca aigua, potser perquè les capçades són localment més espesses, mentre que altres punts en recullen tanta o més de la que plou fora del bosc, probablement perquè hi ha rutes preferents de degoteig. En la mesura que aquestes pautes tinguin certa persistència i no variïn aleatòriament d’una pluja a una altra, la variabilitat espacial de la quantitat de trascol pot ser un factor important en el manteniment de l’heterogeneïtat dels sòls forestals.
L’aigua edàfica

La importància de la disponibilitat d’aigua en el sòl es posa de manifest en la fageda de l’obaga de Matagalls (Montseny), fotografiada el 20 d’agost de 1988. Després de dos mesos d’eixut, la fageda sobre sòl col·luvial relativament profund es manté ben verda (centre de la imatge), mentre que les fagedes que la flanquegen, sobre sòls més esquelètics i amb menys capacitat de retenció d’aigua, tenen la fulla morta dos mesos abans del que els correspondria.
Ferran Rodà
L’aigua de precipitació que arriba al terra del bosc pot infiltrar-se en el sòl o escórrer-se superficialment pendent avall. Els sòls forestals acostumen a tenir elevades taxes d’infiltració que rarament la intensitat de la precipitació depassa. Això fa que en els boscos no pertorbats la infiltració sigui molt més important que l’escorriment superficial, i que la hidrologia dels ecosistemes forestals estigui dominada pels processos subsuperficials. Conseqüentment, l’erosió hídrica dels vessants forestats acostuma a ser escassa perquè el flux subsuperficial de l’aigua té una velocitat molt més petita, per la qual cosa disposa de menys potencial erosiu que no pas l’escorriment superficial.
L’elevada capacitat d’infiltració dels sòls forestals es deu a la porositat dels seus horitzons superficials, rics en matèria orgànica. Aquesta porositat resulta en gran part de l’activitat biològica: la producció i la mortalitat de rels fines juntament amb l’acció excavadora de la fauna edàfica mantenen una xarxa de canalicles de diferents dimensions, els quals transmeten l’aigua cap als horitzons inferiors. Aquesta no és una propietat limitada als boscos, sinó que la majoria dels sòls sota una vegetació densa, sigui herbàcia o arbustiva, tenen una bona capacitat d’infiltració. Per tant, la regulació hidrológica i erosiva no és exclusiva dels sòls forestals, i la comparació de boscos, matollars i prats revela sovint, en aquest sentit, menys diferències de les que a primera vista es podrien esperar. El factor crucial per a aquella regulació no és l’alçada de la vegetació sinó l’existència d’horitzons superficials biològicament actius que donin una infiltració elevada.
Un cop infiltrada, l’aigua del sòl pot drenar de pressa o a poc a poc, evaporar-se o ser absorbida per les rels. En els sòls forestals, l’evaporació és alentida per la capa de virosta i per l’ombra dels arbres, de manera que en un bosc la transpiració és molt més gran que l’evaporació directa des del sòl. Nogensmenys, aquesta pot ser apreciable en boscos oberts, en llocs sense virosta o després de pluges que només arriben a mullar els primers centímetres del sòl.
La quantitat d’aigua disponible en el sòl és un dels factors que més pesen en l’ecologia dels nostres boscos. Les dimensions singulars que assoleixen els pins i les alzines que creixen a la vora de fonts i rierols testifiquen la importància d’un bon subministrament hídric. La quantitat d’aigua utilitzable per les plantes que pot contenir un sòl depèn fonamentalment de la seva profunditat, pedregositat i textura, i també de la seva posició topogràfica, que determina la velocitat a la qual drena. Aquestes propietats del sòl varien d’una manera més o menys predictible al llarg de catenes repetides en el paisatge: la màxima disponibilitat d’aigua i la major producció forestal són a les parts baixes dels vessants i en el llocs còncaus, mentre que les part altes i els llocs convexos són molt més-secs i menys productius.
La reserva d’aigua del sòl es consumeix per drenatge, evaporació i transpiració, i es recarrega per infiltració i, en les posicions topogràfiques adequades, per drenatge lateral. Com que tots aquests processos no actuen simultàniament, ni amb la mateixa intensitat en les diferents fondàries del sòl, es genera un determinat perfil vertical de contingut hídric en el sòl. Les variacions temporals d’aquest perfil d’humitat edàfica en un lloc concret constitueixen el seu règim hídric. Malauradament, hi ha molt poca informació detallada sobre els règims hídrics dels nostres sòls forestals. En comentarem dos casos generals, de particular interès forestal.
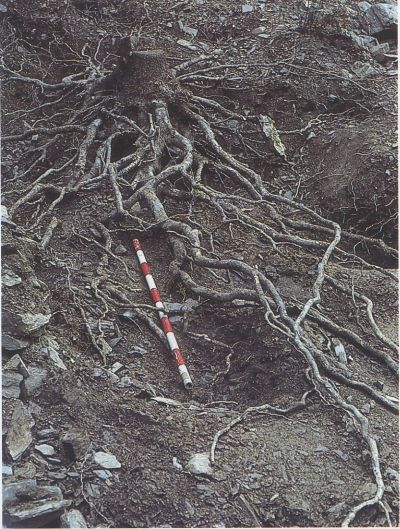
Les rels són una part vital de l’arbre perquè el fixen al terreny i absorbeixen l’aigua i els nutrients minerals. Els sistemes radiculars difereixen molt en extensió i en profunditat segons l’espècie d’arbre i, dins una mateixa espècie, segons les condicions de creixement. Molts arbres i arbusts mediterranis poden tenir rels profundes que ajuden a suportar l’eixut estival, però poden tenir també sistemes radiculars força superficials quan creixen en sòls prims sobre roques no fissurades, com aquesta alzina de la Castanya (Montseny).
Ferran Rodà
El primer fa referència als avantatges que proporciona a un arbre el fet de tenir les rels superficials o profundes. Els primers 50 cm del sòl contenen aigua disponible amb més freqüència, perquè sovint una pluja no és prou abundosa per generar una percolació més profunda. Aquest fet recomana tenir rels superficials, ja que poden absorbir l’aigua abans que no arribi a un vegetal de rels profundes. D’altra banda, els horitzons superficials són els que s’assequen primer. Per sobreviure l’eixut estival, un arbre de rels superficials ha de tenir un sistema radicular molt extens o ser capaç de resistir un fort estrès hídric o d’utilitzar reserves d’aigua en el tronc. En canvi, un arbre de rels profundes pot passar l’eixut estival sense gaire estrès hídric si hi ha suficient aigua a les capes profundes. L’alzina i altres vegetals llenyosos mediterranis poden tenir rels molt profundes, si el substrat ho permet. A les calcàries fissurades de Le Rouquet, les rels d’alzina arriben a 10 m de fondària. A la zona de clima mediterrani de Califòrnia, alguns arbustos esclerofil·lès poden tenir rels fins a 26 m de profunditat. Segurament, dades similars es podrien obtenir per als vegetals mediterranis dels Països Catalans. Perquè resulti profitós produir i mantenir aquestes rels cal que la pluviositat o la topografia del lloc permetin la recàrrega, almenys ocasional, de la reserva profunda d’aigua.
El segon cas es refereix a l’extrem oposat del gradient d’humitat: els sòls entollats. Si un sòl es manté saturat d’aigua, esdevé anaeròbic, mentre que les rels dels arbres respiren oxigen. Algunes espècies d’arbres han resolt el problema de subministrar oxigen a les rels en un sòl anaeròbic i poden viure, per tant, en llocs temporalment o permanentment inundats: són els arbres de ribera, com el vern, els salzes i els pollancres. Altres arbres, com el faig, tenen rels molt sensibles, sigui a la anòxia sigui a la mobilització de metalls (com ara el manganès, que té lloc en condicions reductores), i defugen els sòls que romanen saturats gaire temps.
El balanç hídric
L’estudi de petites conques sobre substrat rocós impermeable permet conèixer el balanç entre la quantitat d’aigua que entra a l’ecosistema amb la precipitació i la quantitat d’aigua que en surt per la riera que drena la conca. La impermeabilitat de la roca mare és necessària per a poder suposar que les sortides d’aigua per infiltració profunda són menyspreables. Si la conca és coberta per un ecosistema relativament homogeni, el drenatge anual ens diu quina és la «producció» d’aigua d’aquest ecosistema. A més, si suposem que la reserva hídrica del sòl és sensiblement igual al principi i al final del període d’estudi, la diferència entre la precipitació i el drenatge anuals ens dóna l’evapo-transpiració real de l’ecosistema, pel que fa a la conca. Aquesta possibilitat és prou interessant, ateses les dificultats metodològiques a l’hora de mesurar l’evapotranspiració d’un bosc a escales d’espai i de temps rellevants per al paisatge.
Als Països Catalans funcionen dues àrees experimentals amb petites conques aforades i instrumentades per a obtenir els balanços d’aigua i nutrients. Una d’aquestes, una conca de 55 ha d’alzinar, a Prades, produí com a mitjana 40 mm/any de drenatge, en un període de 5 anys amb una precipitació de 535 mm anuals; l’altra, una conca d’alzinar de 4,3 ha, al Montseny, produí 404 mm de drenatge en un any que plogueren 873 mm. Aquestes dues localitats poden ser representatives, almenys a grans trets, de dos tipus de condicions molt generalitzades als Països Catalans. El drenatge anual obtingut a la conca de Prades podria ser semblant al de molts dels nostres boscos secs, amb precipitacions inferiors als 600 o 700 mm/any, els quals destinen a l’evapo-transpiració la major part de l’aigua rebuda i produeixen, per tant, un drenatge escàs; en aquests boscos, propis de la terra baixa i de la muntanya mitjana seca, la quantitat d’aigua que percola a través de tot el sòl és petita, la qual cosa implica que els sòls són poc rentats, poc àcids i que, fins i tot en àrees silicatades, la saturació de bases és alta. També són els boscos més fortament limitats per la disponibilitat d’aigua. Al contrari, el drenatge més gran obtingut al Montseny podria representar aquells boscos de la muntanya mitjana plujosa, amb precipitacions entre 700 i 1000 mm/any, no tan fortament limitats per l’aigua, on almenys en alguna part de la conca i durant un temps hi ha excedents hídrics que alimenten un drenatge considerable; els sòls hi són més lixiviats, amb menys cations bàsics i més àcids.
Els boscos produeixen menys drenatge que els ecosistemes herbacis o arbustius. Això es deu al fet que els ecosistemes forestals tenen més pèrdues per intercepció i transpiren més. Tots els experiments d’eliminació o de reducció de la cobertura forestal que s’han fet arreu del món en molt diversos tipus de bosc han donat sempre com a resultat que el drenatge anual en conques on el bosc s’ha tallat totalment o en part és més gran que en conques de control amb el bosc intacte. Aquest és un fet que s’ignora sovint quan es considera el paper hidrològic dels boscos.
El balanç hídric de la conca ens dóna una evapo-transpiració de 495 mm anuals a l’alzinar de Prades, i de 469 al del Montseny. Aquestes estimacions inclouen les pèrdues per intercepció que, com hem vist, poden ser d’uns 70-80 mm anuals en aquests alzinars. Cal recordar també que aquestes xifres d’evapotranspiració són valors mitjans per a la conca, i que segurament l’evapo-transpiració és menor a les parts altes del vessant, que drenen més ràpidament, i major a les parts baixes. Resulta sorprenent que dos alzinars, on la disponibilitat d’aigua (indicada per la precipitació) és tan diferent, tinguin pràcticament la mateixa evapo-transpiració. Probablement, com ja hem comentat, a Prades l’evapo-transpiració és molt limitada per la disponibilitat d’aigua, fet que és corroborat per la forta correlació que hi ha entre la precipitació i l’evapo-transpiració anuals en aquesta conca. Al Montseny, sense que desaparegui del tot la limitació hídrica, l’evapotranspiració és probablement limitada també per la disponibilitat d’energia per evaporar aigua, sobretot durant els mesos d’hivern, quan la conca estudiada, que és molt obaga, no rep radiació solar directa.
La matèria orgànica

Els boscos capitalitzen en forma d’increment de biomassa bona part de la seva producció primària neta. Aquest increment de biomassa és constituït principalment per fusta i escorça de troncs, branques i rels. Els aprofitaments forestals ben portats utilitzen aquesta propietat i retiren del bosc, com a mitjana, una quantitat de fusta igual a la que el bosc pot produir anualment. Si es respecta aquest principi de la productivitat sostinguda i es realitzen els tractaments de manera que s’afavoreixi la regeneració del bosc, els aprofitaments són compatibles amb la persistència indefinida de l’ecosistema forestal. Als Països Catalans, la major part dels boscos es tracten pel sistema d’aclarides. A la fotografia en veiem un exemple en un alzinar del Montseny, utilitzat per a l’obtenció de llenya. Els troncs i les branques gruixudes, trossejats, han estat apilats a la corredora o estimball per extreure’ls de! bosc.
Ferran Rodà
El funcionament de l’ecosistema forestal té dos aspectes bàsics: el flux d’energia i la circulació de nutrients. En parlar de l’ambient forestal hem vist què succeïa amb la radiació solar que arriba al bosc. Una quantitat petita, però altament significativa, de l’energia radiant incident és fixada pels arbres i altres vegetals en forma d’energia química en els enllaços de la matèria orgànica produïda per la fotosíntesi. A partir d’aquí, l’energia flueix a través de l’ecosistema en forma de matèria orgànica. Un gram de matèria vegetal seca té un contingut energètic mitjà de 4 o 5 kilocalories, la qual cosa ens permet transformar les transferències de matèria orgànica en fluxos d’energia.
La producció primària bruta és el total de matèria orgànica produïda pel conjunt dels vegetals del bosc. Bona part d’aquesta matèria orgànica és consumida en la respiració dels mateixos vegetals. La resta, o la producció primària neta, és el que els vegetals poden destinar al seu creixement i reproducció, i és també l’energia disponible per a ser utilitzada pels altres nivells tròfics, com els fitòfags, els detritívors i els descomponedors. Dos punts clau diferencien els boscos dels altres ecosistemes. En primer lloc, els seus productors primaris (els arbres) capitalitzen gran part de la seva producció neta en la formació de fusta, un teixit persistent; això condueix a una notable acumulació de biomassa. En segon lloc, la major part de la matèria orgànica produïda segueix tard o d’hora la via energètica dels detritívors i descomponedors en lloc de la dels fitòfags.
La producció primària no és possible sense certes quantitats de determinats elements químics o nutrients. Els arbres prenen del sòl, en forma inorgànica, la majoria dels nutrients que necessiten. Dins l’arbre, part d’aquests nutrients són incorporats a la matèria orgànica. Malgrat que hi ha fluxos importants de nutrients que tenen lloc en forma inorgànica, moltes de les transferències de nutrients a través de l’ecosistema van lligades a la circulació de la matèria orgànica. En particular, els nutrients que els arbres han absorbit tornen al sòl generalment mitjançant la matèria orgànica morta que es desprèn d’aquests, i són alliberats un altre cop en forma inorgànica per l’activitat dels deseomponedors. Aquest reciclatge actiu de nutrients entre els arbres i el sòl és un dels trets principals del funcionament del bosc i és fonamental per a mantenir-ne la productivitat. Tractarem a continuació de les característiques bàsiques de la distribució i circulació de la matèria orgànica en els ecosistemes forestals, i després de la circulació de nutrients.
Producció
En general, els boscos són ecosistemes altament productius. Això es deu, d’una banda, al fet que potencialment els boscos solen ocupar els climes i sòls més favorables i, de l’altra, a la possessió de troncs, brancams i sistemes radiculars importants que són veritables infrastructures de suport, d’adquisició de recursos, d’emmagatzematge i de transport.
La producció primària neta mitjana de les pluviïsilves tropicals s’estima en 20 t/ha·any, en pes sec; la dels boscos temperats de coníferes o de caducifolis, en 12 t/ha·any; i la dels boscos boreals de coníferes o taigà, en 8 t/ha·any. Aquestes xifres es podrien considerar produccions totals, ja que inclouen les rels, però el reconeixement recent de la importància de la producció de rels fines fa pensar que més aviat deuen ser subestimacions. Tot i això, els valors anteriors indiquen prou bé que els principals factors limitants de la producció primària dels boscos, a l’escala dels continents, són la disponibilitat d’energia i la disponibilitat d’aigua. La producció és màxima en els boscos de climes càlids i humids, i mínima en climes freds o secs. La producció primària mitjana sota un clima determinat es pot predir amb certa aproximació a partir de l’evapo-transpiració real, ja que aquesta integra la disponibilitat d’aigua i d’energia per a evaporar-la. També dins dels Països Catalans es posen clarament de manifest els determinants climàtics de la producció. Podem esperar que la producció primària dels nostres boscos augmenti de S a N, a mesura que augmenten les precipitacions.
A l’escala del rodal les produccions poden variar molt en distàncies d’un centenar de metres, perquè depenen fortament de la qualitat del lloc, de l’espècie d’arbre i de les característiques de la massa. Entre els factors importants per la qualitat de lloc hi ha l’orientació, la inclinació, la posició topogràfica i el tipus i la profunditat del sòl. Tots poden influir en el creixement dels arbres perquè modifiquen el microclima, el règim hídric del sòl o la disponibilitat de nutrients. Entre les característiques de la massa arbòria que poden determinar la producció figuren els genotips de l’espècie o espècies dominants, i l’edat, la grandària, la densitat i la morfologia dels arbres. Aquestes característiques poden variar molt entre llocs propers, ateses les diferències en la qualitat del lloc o, més generalment, les diferències històriques, com el tipus de gestió silvícola o l’estat successional.
Quan un bosc s’estableix en un lloc, la producció primària bruta augmenta ràpidament fins que es tanca el sostre del bosc. A partir d’aquest moment, la biomassa foliar roman més o menys constant i també la producció bruta. La respiració dels arbres va augmentant de manera continuada perquè es van acumulant teixits vius als troncs i a les rels. Com a resultat, la producció primària neta presenta un màxim al voltant del moment en què es tanca el sostre del bosc, i després disminueix. Si una pertorbació destrueix els arbres, el cicle comença un altre cop. Si en un bosc tancat una pertorbació mata només una certa proporció d’arbres, pot ser que la producció primària neta augmenti. Això es deu al fet que la respiració disminueix en ser eliminada part de la biomassa, però en canvi l’àrea foliar i la producció primària bruta poden recuperar en poc temps els valors anteriors a la pertorbació perquè els arbres que sobreviuen reben més llum i disposen de més aigua i nutrients cadascun. Aquesta paradoxa indica que la competència per excés de densitat limita sovint la producció d’un rodal. La pràctica silvícola d’aclarides periòdiques es basa en aquest principi.
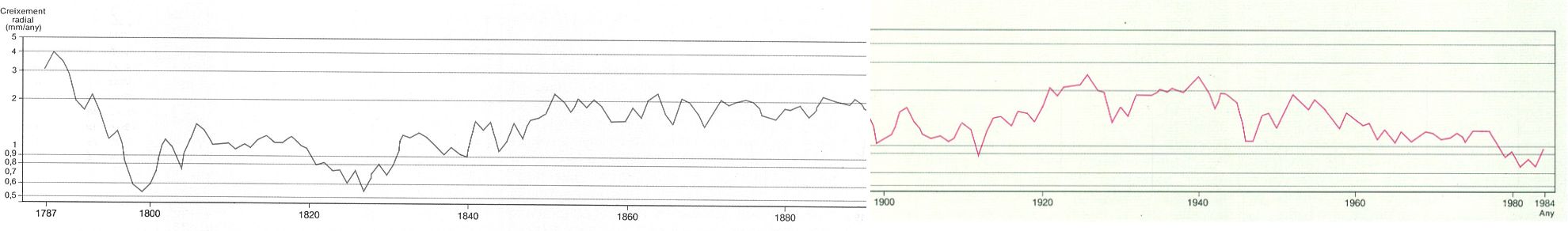
Creixement anual dels avets de Passavets (Montseny) durant els darrers 200 anys. La corba és la mitjana del creixement radial del tronc de 10 avets. Noteu que el creixement es representa en escala logarítmica.
Carto-Tec, original de C. Tomàs.
La producció primària neta es reparteix en fulles, inflorescències, fruits, brots, fusta i escorça de branques, de troncs i de rels gruixudes, formació de rels fines i exsudats. El mesurament de la producció neta d’un bosc és molt laboriós ja que cal incloure-hi tots aquests components i enfrontar-se amb una considerable variabilitat entre arbres. A més, algunes de les fraccions citades plantegen problemes metodològics seriosos, com les rels. Per aquests motius, són ben poques les dades sobre la producció primària dels nostres boscos que tinguin un fonament sòlid. L’estudi més complet realitzat fins ara als Països Catalans és el d’una parcel·la experimental d’alzinar muntanyenc a la Castanya (Montseny). En aquesta parcel·la, s’obtingué una producció primària neta aèria de 9,7 t/ha·any de pes sec, de les quals 9,3 corresponen a l’estrat arbori. La producció de fulles és de 3,1 t/ha·any; la de fruits (glans i cúpules), 0,6; la de troncs i branques gruixudes (d’un diàmetre més gran de 5 cm), 3,4; i la de branques primes (d’un diàmetre més petit de 5 cm), incloent-hi els brots de l’any, 2,3 t/ha·any. La producció de rels no es mesurà, però si les rels gruixudes (d’un diàmetre més gran d’1 cm) d’aquesta parcel·la tinguessin la mateixa taxa de renovació que la biomassa llenyosa aèria, la producció de rels gruixudes representaria 1,5 t/ha·any. Això eleva la producció d’aquest bosc a 11,2 t/ha·any, i caldria afegir-hi diverses tones més per la producció de rels fines: en els boscos europeus i nord-americans, on aquesta ha estat mesurada, ha resultat semblant o considerablement superior a la producció de fulles.
Els valors de producció que acabem de veure poden semblar alts per a un alzinar i podrien reflectir les condicions climàtiques i topogràfiques favorables en què es troba aquesta parcel·la del Montseny. No obstant això, les xifres de producció que hem donat serien aplicables a molts boscos temperats, i, probablement, gran part dels nostres boscos de caducifolis i de coníferes no s’allunyarien gaire d’aquests valors, almenys en climes plujosos no excessivament freds.
Mort i descomposició
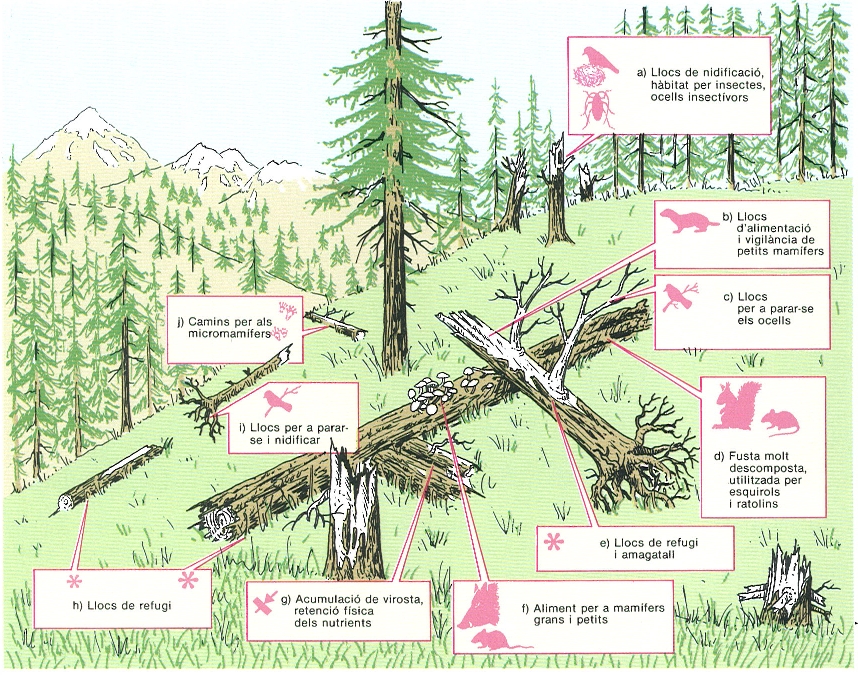
La fusta gruixuda morta té un paper molt important en els ecositemes forestals, tant si es tracta de troncs drets (a) com de troncs o branques grosses caiguts al terra del bosc (b-j). La fusta gruixuda morta és quasi inexistent en la major part dels boscos gestionats, mentre que en els boscos madurs arriba a ser un component principal de l’ecosistema. En boscos verges de coníferes de l’oest de l’Amèrica del Nord, els troncs en descomposició poden ocupar fins el 20% de la superfície del terra. La figura il·lustra algunes de les funcions de la fusta morta i posa èmfasi en el seu paper com a hàbitat per als vertebrats.
Román Montull, simplificat a partir d’originals de F.J. Triska i K. Cromack.
Com hem vist, part de la producció primària neta es destina a construir òrgans de vida relativament curta, com ara fulles, inflorescències, fruits i rels fines. D’una manera periòdica o continuada, aquests òrgans moren, es desprenen de l’arbre i passen a formar part de la matèria orgànica morta del sòl. La caiguda de virosta i la mortalitat de rels fines constitueixen l’entrada d’energia per als animals detritívors i per als microorganismes descomponedors.
En la caiguda de virosta és habitual distingir una fracció fina (formada pèr fulles, branquillons, inflorescències, fruits, fragments d’escorça o d’epífits, restes d’insectes, etc.) i una fracció gruixuda (formada per les branques que superen una grandària determinada). La caiguda de virosta fina es mesura amb cistelles de superfície coneguda, però les branques gruixudes requereixen unitats de mostreig més grans. La caiguda de virosta sol ser fortament estacional. Això és evident en el cas de les fulles dels caducifolis, però també en les fulles dels perennifolis. Les fulles de l’alzina cauen sobretot en el moment que broten les fulles noves, pel maig a la terra baixa i pel juny a muntanya. Les acícules del pi blanc i del pi pinyer cauen principalment al juliol i a l’agost. Al Montseny, el màxim de caiguda de fulles d’avet és a l’agost, igual que el teix i el boix grèvol. Al Pirineu, el pi roig té la màxima caiguda d’acícules a l’octubre. Per als perennifolis mediterranis s’ha especulat que aquelles espècies que són més limitades per l’aigua perden la fulla a l’estiu, per reduir l’àrea foliar en el moment de més demanda evaporativa; contràriament, les espècies més aviat limitades pels nutrients perden la fulla vella en brotar, per cobrir la demanda de nutrients dels brots i de les fulles noves amb els nutrients recuperats de les velles. El pi blanc i el pi pinyer serien en el primer cas, i l’alzina, de rels profundes que li permetrien de pal·liar l’eixut estival, en el segon.

Els fongs són agents essencials en la descomposició de la fusta morta. El miceli dels fongs lignícoles creix per l’interior de la fusta i degrada la cel·lulosa, la lignina i altres compostos orgànics en els seus components més senzills, utilitzant l’energia continguda en el material de partida. En la fotografia veiem els cossos fructífers del bolet de soca Trametes versicolor sobre una branca morta de roure.
Anna Àvila.
L’estacionalitat en la caiguda es dóna també en altres fraccions de la virosta, sobretot en inflorescències i fruits. Glans i fages cauen predominantment a la tardor i formen part important de l’alimentació de molts ocells i mamífers durant la tardor i l’hivern. En alguns casos aquesta relació tròfica és ben forta: la demografia de les poblacions de ratolins de bosc (Apodemus) pot trobar-se condicionada per la quantitat de glans produïdes cada any.
La quantitat anual de virosta produïda varia segons el tipus de bosc i es correlaciona amb la producció primària neta, de manera que la caiguda de virosta és més gran en els boscos més productius. A les nostres latituds la major part dels boscos produeixen de 3 a 6 t/ha·any de virosta, en pes sec. L’alzinar de la Castanya produeix 4,8 t/ha·any de virosta fina, mentre que l’alzinar més xèric de Prades i dos altres alzinars situats al S de França i a Sicília tenen tots tres unes caigudes de virosta entre 3,6 i 3,8 t/ha·any. També al S de França, un bosc dens de pi blanc produí 3,9 t/ha·any de virosta, valor molt semblant al dels alzinars, però un bosc de pi pinyer produí el doble (7,9 t/ha·any). El bosc de pi roig amb grèvol de San Juan de la Peña, a l’alt Aragó, produí 4,0 t/ha·any de virosta. Al Montseny, la caiguda de virosta fina d’una fageda fou de 4,1 t/ha·any, i la d’una avetosa 3,3 t/ha·any, xifres sensiblement inferiors a la de l’alzinar muntanyenc de la Castanya. Un recull bibliogràfic dóna una producció mitjana de virosta de 4,4 t/ha·any per als boscos temperats de coníferes, i 5,4 t/ha·any per als boscos temperats de caducifolis.
Les fulles dels arbres constitueixen la fracció dominant de la caiguda de virosta i representen com a mitjana al voltant del 70% del pes total de virosta fina; aquest percentatge tendeix a davallar a mesura que el bosc madura i augmenta la caiguda de branques. La quantitat de fulles caigudes és una mesura útil de la producció de fulles dels estrats arbori i arbustiu alt. Això és clar en el cas dels caducifolis, però també es pot aplicar als perennifolis si suposem que la biomassa foliar és en estat estacionari, cosa raonable si el bosc no és gaire jove. Tant per a caducifolis com perennifolis, si considerem que la caiguda de fulles en mesura la producció, no tenim en compte el consum pels fitòfags, però aquest representa generalment un percentatge petit de la producció foliar. En la majoria dels boscos mediterranis i temperats la caiguda de fulles és de 2 a 4 t/ha·any.

L’autoaclarida és el procés natural de mortalitat deguda a la competència derivada de l’excés de densitat. Aquesta mortalitat és més gran en els arbres menys vigorosos, que sovint són els de dimensions més petites. El faig arquejat del centre de la fotografia és mort, probablement perquè va quedar durant molt de temps ombrejat pels seus veïns dominants. Els arbres suprimits tenen una producció neta molt baixa que no els permet dotar-se de les reserves de carbohidrats o de les defenses químiques necessàries per a sobreviure en condicions adverses.
Ferran Rodà
Després de les fulles, les fraccions més importants de la caiguda de virosta solen ser els branquillons i els fruits. La producció de fruits que fan els arbres forestals és extraordinàriament variable d’un any a l’altre. Un any d’elevada producció de fruits és seguit típicament d’un o més anys de producció escassa. En dos anys d’estudi a la fageda de Santa Fe del Montseny, el primer any es recolliren 0,72 t/ha·any de fruits (fages i cúpules) i el segon només 0,007 t/ha·any. A l’alzinar de Prades, en tres anys d’estudi, es recolliren successivament 1,7, 0,05 i 0,2 t/ha·any de glans més cúpules. A l’alzinar de Le Rouquet, prop de Montpeller, la caiguda d’inflorescències i fruits durant set anys varià entre 0,3 i 1,5 t/ha·any. En el faig, es diu que hi ha un any de producció molt elevada de fruits cada 4-6 anys o cada 5-7 anys, segons els autors. Malauradament, aquestes freqüències depenen de com es defineixi que un any és de producció extraordinària (una anyada) i, a més, calen sèries molt llargues de dades per a establir estadísticament quin és el període de recurrència de tals anys. Tant en roures (Quercus robur i Q. petraea) com en el faig, a Europa hi ha una considerable variació geogràfica en la duració dels intervals entre anyades. S’ha interpretat aquesta gran variabilitat interanual en la producció de fruits dels arbres forestals com una adaptació per evitar que les poblacions dels depredadors de llavors, tant vertebrats com insectes, es puguin mantenir constantment a una densitat alta a la qual podrien consumir gairebé totes les llavors. Les anyades saturarien els depredadors de llavors i permetrien que en germinessin una quantitat important. De fet, la regeneració eficaç per llavor en molts arbres forestals és sovint associada al fet que coincideixin un any d’elevada fructificació amb la disponibilitat d’espais adequats per a la germinació i la supervivència de les plàntules.
La virosta que cau a terra tarda un cert temps a descompondre’s. Això vol dir que, si la virosta no és exportada pendent avall pel vent o l’aigua, se n’acumularà una capa damunt del sòl. La quantitat de matèria orgànica acumulada en aquest horitzó orgànic dependrà del balanç entre les taxes de caiguda de virosta i de descomposició. La velocitat de descomposició depèn del clima i de la qualitat del substrat. L’activitat dels descomponedors s’incrementa amb la temperatura i amb la humitat, de manera que la quantitat de virosta en descomposició damunt de terra disminueix des de la taigà fins a la pluviïsilva tropical. Novament, es pot utilitzar l’evapo-transpiració real per a predir la magnitud de la velocitat de descomposició d’un material de certes característiques. A més evapo-transpiració real més ràpida és la descomposició. Entre les propietats de la virosta que influeixen en la velocitat de descomposició figuren, com a més importants, els seus continguts de lignina i nitrogen. La descomposició és més ràpida en materials rics en nitrogen i pobres en lignina. En general, per aquestes raons, la virosta dels caducifolis es descompon més ràpidament que la virosta de coníferes. Tanmateix, hi ha força variabilitat entre les taxes de descomposició de les fulles de diferents espècies de caducifolis i de coníferes. Entre els primers, les fulles de faig i de roure es descomponen molt més lentament que les de freixe, blada, om o saüquer, a causa del contingut més alt de lignina i de la concentració més baixa de nitrogen d’aquelles.
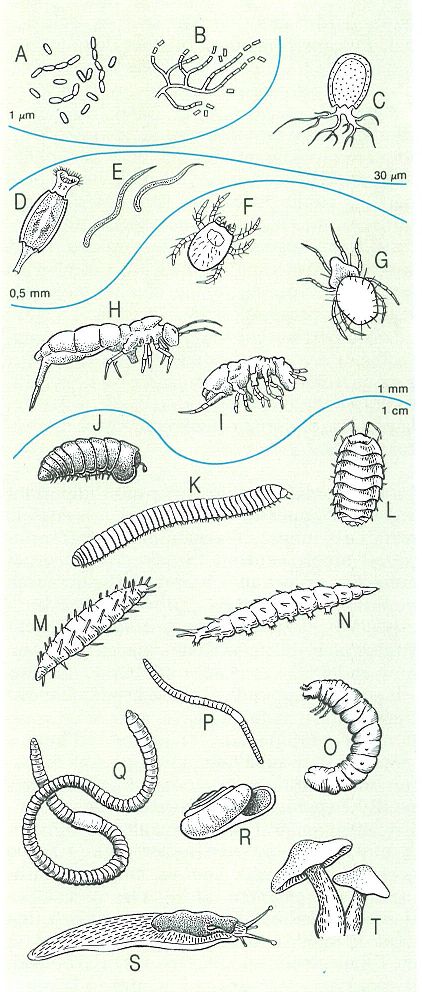
Representants dels principals grups d’organismes descomponedors o detritívors de la virosta i el sòl forestals. Els dibuixos no estan fets a la mateixa escala, però s’han agrupat els organismes segons la seva mida. A bacteris, B actinomicets, C tecameba (protozou), D rotífer, E nematodes, F-G criptostigamates (àcars oribàtids), H-I col·lèmbols, J-K diplòpodes, L isòpode, M-N larves de dípter, O larva d’escarabèid (coleòpter), P-Q anèl·lids (P enquitrèid, Q lumbrícid), R-S mol·luscs, T bolet.
Amadeu Blasco, modificat a partir d’un original de J.R. Packham i D.J.L Harding.
La descomposició és un procés complex. Les fulles i branques en descomposició poden perdre pes per lixiviació, fragmentació, respiració dels heteròtrofs i emigració d’adults emergents d’insectes xil·lòfags. La descomposició comença ja a les capçades, sobretot per les branques que poden romandre molts anys mortes en els arbres i que quan cauen poden haver perdut ja un terç del seu pes sec original. En canvi, les fulles caigudes de fa poc estan gairebé intactes i la colonització microbiana és encara incipient. La descomposició en aquesta capa superficial de fulles fresques, o capa L, és sovint lenta perquè les fulles s’assequen ràpidament després de cada pluja. Durant els primers mesos, la pluja lixivia gran part del contingut orgànic soluble i d’aquells nutrients més fàcilment rentables, com el potassi. L’any següent, les fulles són enterrades sota una nova capa de fullaraca i esdevenen la capa F. Aquesta roman humida molt més temps i la descomposició s’accelera. La colonització pels micelis fúngics i la respiració microbiana són màximes en aquesta capa. Per sota es troba una capa orgànica on l’estructura del material original ja no és identificable, la capa H, enriquida en els compostos més recalcitrants.
La fauna edàfica té un paper molt important en la descomposició de la virosta. Els cucs de terra i els milpeus fragmenten i enterren les fulles, acceleren molt la incorporació de la materia orgànica al sòl mineral i generen un humus de tipus «mull». En sòls molt àcids, i especialment amb virosta de coníferes, l’activitat d’aquesta macrofauna manca o es veu molt reduïda, i es genera un humus de tipus «mor» on les capes L, F i H són clarament distingibles. Un mateix tipus de fulla es descompon més ràpidament en un sòl d’humus «mull» que en un d’humus «mor», sota el mateix clima. També l’activitat d’animals més petits, l’anomenada mesofauna, és crucial. Un metre quadrat de sòl forestal pot contenir alguns milions de nematodes, i fins 400 000 àcars i colèmbols. Aquests animals consumeixen i fragmenten les fulles en descomposició i d’altra matèria orgànica, i augmenten molt la superfície del substrat exposada; a més, amb els seus desplaçaments, afavoreixen la inoculació microbiana i mantenen la porositat del sòl. Altres, com ara molts àcars oribàtids, s’alimenten del miceli dels fongs i sembla que afecten la taxa de descomposició en depredar i rejovenir les poblacions fúngiques. Si s’exclou experimentalment la fauna edàfica per mitjans físics o químics, la velocitat de descomposició de la matèria orgànica disminueix considerablement.
La circulació de nutrients
Els nutrients en la vegetació
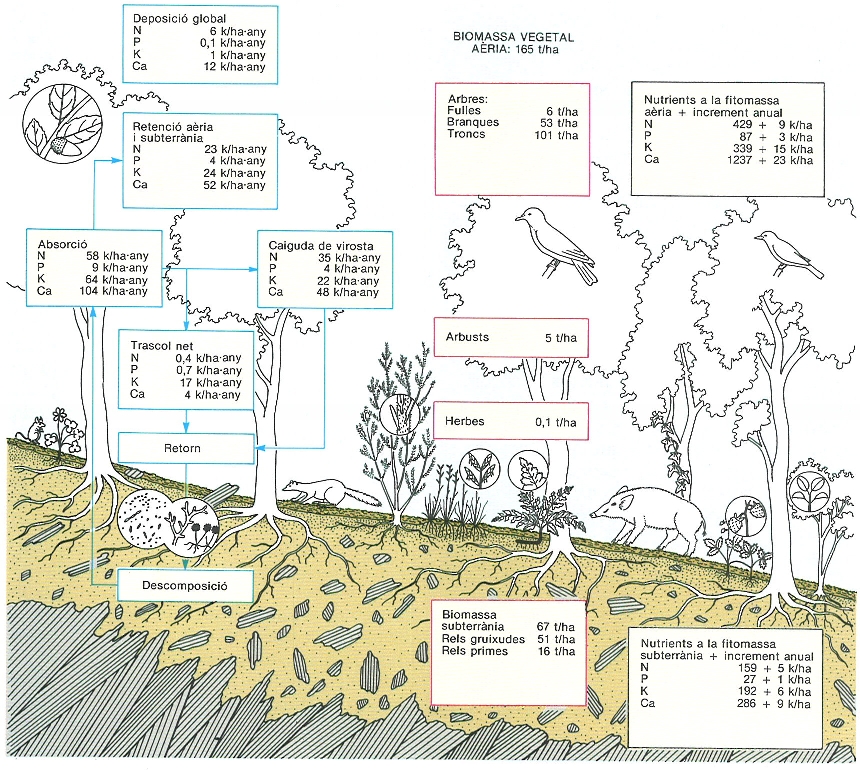
Distribució de la biomassa vegetal (blau), dels nutrients continguts en aquesta biomassa (negre) i principals fluxos de nutrients (vermell) en la parcel·la d’alzinar muntanyenc de la Castanya (Montseny), L’estrat arbori domina de molt la distribució de la biomassa i dels nutrients, ja que el sotabosc és ben migrat. Els boscos absorbeixen cada any quantitats importants de nutrients del sòl. En aquest alzinar, aproximadament la meitat dels nutrients absorbits retorna al terra del bosc i la resta s’incorpora a la producció anual de fusta i escorça (retenció). Però l’increment anual net de nutrients a la biomassa és inferior a la retenció perquè també cauen a terra branques produïdes en anys anteriors. Tot i així, les quantitats de nutrients contingudes a la biomassa augmenten (segons de quin nutrient es tracta) d’un 2 a un 4% anual a causa de l’acumulació de biomassa en aquest bosc en creixement.
Mercè Cartañá, original de l’autor, amb dades de J. Canadell, L. Ferrés, F. Rodà i A.M.C. Verdú.
Dins d’un arbre, la concentració de nutrients varia força d’una part a una altra. En general, els òrgans de més activitat metabòlica són els més rics en nutrients. Així, les fulles, brots joves, inflorescències i fruits en tenen concentracions relativament altes, especialment de nitrogen, fòsfor, potassi i magnesi. La fusta és molt més pobra de nutrients, sobretot la fusta central del tronc, que és xilema mort. L’escorça sol tenir-ne concentracions intermèdies entre les fulles i la fusta, però és molt més rica en calci que aquestes, sobretot al gènere Quercus.
Les fulles de diferents espècies d’arbres poden presentar diferències notables en la seva concentració de nutrients. En general, les dels caducifolis en són molt més riques que les dels perennifolis; per exemple, les fulles de faig tenen un 2% de nitrogen respecte al seu pes sec mentre que les de l’alzina superen per poc 1’1%. Aquestes diferències són importants perquè influeixen tant en la capacitat fotosintetitzadora màxima del vegetal com en la quantitat de nutrients necessaris per a construir la biomassa foliar produïda cada any. Les concentracions foliars de nutrients poden variar segons l’edat de la fulla i la seva posició dins de la capçada. De particular interès en els cicles biogeoquímics és la reabsorció o retranslocament de nutrients des de les fulles velles cap a altres parts de la planta. Aquest retranslocament permet a l’arbre retirar part dels nutrients que contenen les fulles abans que no caiguin, i transportar-los a òrgans en creixement o emmagatzemar-los per a un ús futur. Al Montseny, l’alzina retransloca la meitat del nitrogen i un terç del fòsfor de les seves fulles abans que no caiguin, mentre que el faig retransloca a la tardor el 46% del fòsfor foliar. Mitjançant aquest procés, l’arbre adquireix un control intern més alt sobre la disponibilitat de nutrients, i evita que els nutrients reabsorbits puguin ser captats per un altre arbre, immobilitzats pels microorganismes del sòl o exportats del bosc amb l’aigua de drenatge. El retranslocament fa que la fullaraca produïda pels arbres sigui més pobra en nutrients. Com hem comentat, això en retarda la descomposició, amb la qual cosa aquest retranslocament té efectes per a l’ecosistema perquè afecta aquest pas crític del reciclatge de la matèria orgànica.
Les branques fines d’alzina són més riques en nutrients que les d’altres arbres temperats, tant coníferes com caducifolis. En la mesura que aquests nutrients siguin mobilitzables, això independitzaria, fins a un cert grau, l’arbre enfront de les variacions ambientals en la disponibilitat de nutrients. Aquesta característica resultaria adaptativa per als vegetals mediterranis que viuen sovint en sòls pobres de nutrients i sotmesos a règims de precipitacions molt irregulars.

Les parcel·les experimentals permanents, com aquesta de l’alzinar muntanyenc de la Castanya (Montseny), permeten l’estudi intensiu del funcionament de l’ecosistema forestal. Amb l’utillatge i els mostreigs adients es poden determinar els fluxos d’energia entre els diferents components de l’ecosistema, i es pot quantificar la distribució i la circulació de materials essencials com l’aigua, la matèria orgànica o els nutrients minerals. La cistella verda que veiem a la fotografia, una de les moltes que hi ha a la parcel·la, serveix per a mesurar la caiguda de virosta per unitat de superfície i de temps, i poder avaluar així la magnitud del retorn de matèria orgànica i nutrients des dels arbres al sòl.
Ferran Rodà
La mineralomassa o contingut de nutrients és la quantitat de nutrients que conté la vegetació per unitat de superfície del bosc. Aquesta quantitat de nutrients que la vegetació emmagatzema o immobilitza depèn de la seva biomassa i de la seva concentració mitjana de nutrients. Com que els boscos acumulen molta biomassa, les mineralomasses forestals són també molt més grans que les dels altres ecosistemes. A les pluviïsilves equatorials la vegetació conté la major part dels nutrients de l’ecosistema. Als boscos temperats, el sòl acostuma a tenir més pes quantitatiu en la distribució de nutrients, tot i que la vegetació en conté quantitats molt considerables. Els boscos mediterranis tenen sovint biomasses només moderades, però les concentracions mitjanes de nutrients als arbres són més aviat altes, amb la qual cosa les seves mineralomasses poden ser iguals o superiors a les dels boscos temperats. Per exemple, la biomassa aèria de l’alzinar de la Castanya, sobre sòl silicatat, conté 1240 kg/ha de calci, i 430 kg/ha de nitrogen. A causa de la seva elevada biomassa, els troncs contenen un percentatge alt dels nutrients dels arbres. No obstant això, les fraccions més riques en nutrients com són ara les fulles i branques fines tenen més participació en la mineralomassa de la que indica la seva petita biomassa. Així, a l’alzinar de la Castanya, les fulles representen només el 4% de la biomassa aèria, però contenen un 20% del nitrogen. En números rodons, els troncs contenen sovint la meitat dels nutrients de la biomassa aèria del bosc. Això vol dir que aquells sistemes de gestió forestal que comporten una utilització de tot l’arbre (tronc, branques i fulles) ocasionen una exportació de nutrients del bosc que és aproximadament el doble d’un sistema convencional on només es retiren els troncs. L’escorça pot tenir igualment un paper destacat en la distribució dels nutrients. A l’alzinar de la Castanya, l’escorça de troncs i branques conté el 68% de tot el calci de la biomassa aèria.
Els cicles biogeoquímics
En un bosc hi ha fluxos de nutrients que entren a l’ecosistema o en surten. Aquests fluxos constitueixen el cicle extern de nutrients o cicle intersistèmic, perquè connecten l’ecosistema amb altres ecosistemes adjacents o llunyans. Al mateix temps, dintre del bosc tenen lloc fluxos de nutrients entre les diferents parts o compartiments de l’ecosistema forestal: és el cicle intern de nutrients o cicle intrasistèmic. Una de les característiques més destacades dels ecosistemes forestals relativament poc pertorbats és que les quantitats de nutrients essencials que circulen anualment dins del bosc són molt més grans que les quantitats de nutrients que al cap de l’any han entrat a l’ecosistema o n’han sortit. El funcionament continuat del bosc depèn d’aquesta capacitat de reciclar eficientment els elements nutritius tot i mantenir-ne baixa l’exportació o pèrdua.
El cicle intern
La circulació interna de nutrients en l’ecosistema forestal té cinc passos o fluxos principals: l’absorció de nutrients del sòl per les rels o les micorrizes, i el seu transport fins a les parts de l’arbre on seran utilitzats; el retranslocament de nutrients dels òrgans vells per ser reutilitzats o emmagatzemats; la retenció de nutrients en la fusta i l’escorça produïda cada any; el retorn al sòl dels nutrients absorbits que no hagin quedat retinguts en la fusta i l’escorça noves; i la mineralització que els descomponedors fan dels nutrients retornats al sòl en forma orgànica, nutrients que són alliberats en forma inorgànica i poden ser absorbits novament pels arbres.
A escala anual els nutrients absorbits poden ser, doncs, retinguts en la nova producció d’òrgans persistents que s’acumulen en l’arbre (fusta i escorça de branques, troncs i rels) o destinats a la producció d’òrgans de renovació més ràpida (fulles, inflorescències, fruits i rels fines) i retornats al sòl. Retenció i retorn tenen, per tant, un efecte molt diferent sobre la destinació dels nutrients. Mentre que els nutrients retornats circulen activament per l’ecosistema, els nutrients retinguts incrementen la mineralomassa del bosc i queden fora del cicle general durant dècades o segles fins que les parts llenyoses que els contenen es moren i es descomponen. La repartició dels nutrients absorbits entre retenció i retorn depèn del nutrient considerat, del tipus de bosc i de l’estat successional. Els boscos joves retenen en les seves fraccions llenyoses un percentatge molt elevat dels nutrients que absorbeixen anualment, mentre que en els boscos madurs, amb poca acumulació nova de biomassa, el retorn sol ser més important. A l’alzinar de la Castanya, la retenció en troncs i branques hi representa els següents percentatges de l’absorció aèria anual de nutrients: nitrogen 33%, fòsfor 42%, potassi 31%, calci 46% i magnesi 38%. Veiem que el potassi i el nitrogen queden retinguts en menys proporció i són retornats en més proporció que el calci i el fòsfor. Les diferènciés entre elements depenen de les concentracions respectives en la fusta i l’escorça i de la magnitud del retorn de cadascun.
El retorn de nutrients al sòl es realitza per via aèria i subterrània. El retorn subterrani té lloc mitjançant els exsudats de les rels, la mort i descomposició de rels gruixudes, el despreniment del còrtex radicular a mesura que la rel creix, i la mortalitat de rels fines. El retorn subterrani de nutrients és quantitativament molt important i, a més, deu tenir una gran influència en les propietats i la dinàmica de la rizosfera. No obstant això, els problemes metodològics fan que hi hagi poca informació sobre la magnitud dels fluxos de nutrients implicats.
El retorn aeri té lloc per lixiviació de capçades i troncs, i per la caiguda de virosta. La lixiviació produïda per l’aigua de pluja és especialment important en el flux de retorn de potassi i, en menys grau, en el de fòsfor i magnesi. La major part dels nutrients lixiviats dels arbres arriben al terra del bosc amb el trascol, mentre que l’escolament cortical pot aportar des de quantitats negligibles fins a un 30% dels elements lixiviats. Mentre que per al potassi el flux per lixiviació pot ser el mateix que per a la caiguda de virosta, per a tots els altres nutrients el retorn aeri és dominat per aquest darrer procés. S’ha dit que els nutrients que retornen anualment al terra del bosc, i en particular els que conté la caiguda de virosta, són el capital circulant de la fertilitat del sòl. Prèvia descomposició de la matèria orgànica que els conté, aquests nutrients poden estar novament disponibles per ser absorbits pels arbres, i la seva circulació continuada manté la productivitat del bosc. El flux de nutrients amb la caiguda de virosta es pot considerar un índex de la disponibilitat de nutrients en l’ecosistema. La major part dels alzinars i de les pinedes mediterranis retornen al sòl amb la caiguda de virosta una quantitat de nitrogen que pot oscil·lar entre els 25 i els 35 kg/ha any, xifra semblant als 34 kg N/ha. any que retornen com a mitjana els boscos temperats de coníferes, però sensiblement inferior als 51 kg N/ha ·any que cauen, també com a mitjana, amb la virosta dels boscos temperats de caducifolis. Els boscos d’una mateixa espècie poden retornar quantitats diferents de nutrients segons la qualitat del lloc, l’estat successional i altres factors. Per exemple, la fageda de Santa Fe del Montseny retorna amb la virosta només 25 kg de nitrogen/ha any, la meitat del que retornen com a terme mitjà les fagedes centreuropees. Això no es deu al fet que la fageda del Montseny produeixi menys virosta que les fagedes forànies sinó que la virosta que produeix és molt pobre en nitrogen. És possible que l’escassa circulació de nitrogen a les fagedes de les muntanyes circummediterrànies es relacioni amb una menor velocitat de descomposició de la fullaraca que resultaria de la manca recurrent d’aigua.
El cicle extern

Els fitòfags, o animals que mengen vegetals vius, poden tenir una gran importància en el funcionament dels ecosistemes forestals. Un exemple espectacular són les defoliacions produïdes per erugues que s’alimenten de fulles. Mentre que algunes espècies d’arbres moren en ser totalment defoliades, les branques d’altres rebroten i reconstitueixen les capçades en poc temps, de manera que la defoliació hi té un efecte molt menys dràstic. És el cas d’aquesta sureda de Sant Feliu de Buixalleu (Selva), defoliada per erugues de Lymantria dispar al principi del mes de juliol de 1983. La defoliació d’un bosc fa que augmenti molt la llum que hi penetra, accelera el cicle de nutrients i afecta de manera subtil·l’organització de les capçades que es regeneren.
Ferran Rodà
Ja hem descrit l’ús de petites conques aforades sobre roca impermeable per a mesurar els fluxos d’entrada i sortida de nutrients en un ecosistema forestal, i hem comentat les principals característiques dels balanços de nutrients en dues conques d’alzinar al Montseny i a Prades. Recordem aquí només que l’atmosfera és una font apreciable de nutrients, i contaminants, per als ecosistemes terrestres. La precipitació, la sedimentació de pols, la impactació d’aerosols i l’absorció de gasos aporten nutrients als boscos. Malauradament és molt difícil de mesurar els fluxos d’elements dipositats per impactació d’aerosols o absorció de gasos sobre un sostre forestal d’arquitectura complexa.
Sovint es mesura només una part de la deposició atmosfèrica: l’anomenada deposició global, que és la suma dels elements aportats per la precipitació i per la sedimentació de pols. La deposició global que reben els boscos dels Països Catalans es caracteritza pel fet de ser bàsica o només moderadament àcida. Excepte en zones concretes, les aportacions atmosfèriques de sofre i nitrogen són igualment molt menors que les que predominen a l’Europa central i septentrional. A més, la major part del sofre atmosfèric entra als nostres boscos en forma de sulfats neutres i no com a àcid sulfúric. El conjunt dels boscos dels Països Catalans no es troba, doncs, sotmès a un estrès ambiental especialment fort que es derivi de la deposició atmosfèrica de substàncies àcides o acidificants, situació que es reforça amb l’elevada capacitat de neutralització que tenen la majoria dels nostres sòls, tan carbonatats com silicatats. La deposició global és una via d’entrada de nutrients als boscos. Pels estudis fets a Prades i al Montseny, sabem que els boscos catalans reben cada any 5 o 6 kg/ha de nitrogen inorgànic en la deposició global, i caldria afegir-hi una quantitat indeterminada de nitrogen orgànic. L’aportació atmosfèrica de nitrogen és energèticament gratuïta per als arbres i té lloc en formes fàcilment assimilables. Aquest subsidi atmosfèric pot tenir un paper destacat en la nutrició nitrogenada dels ecosistemes forestals. A l’alzinar de la Castanya, el nitrogen inorgànic de la deposició global representa l’11% de l’absorció aèria anual, però el 44% de l’increment anual de la mineralomassa aèria d’aquest element. La deposició global aporta també altres nutrients. Les entrades de calci per aquesta via als nostres boscos són de 10-15 kg/ha any, i podrien tenir una certa significació en sòls pobres de bases.
La fixació biològica de nitrogen és una altra possible font de nitrogen atmosfèric per als ecosistemes. Però a diferència de la deposició atmosfèrica, la fixació biològica és energèticament costosa, i només la poden realitzar certs microorganismes. La fixació biològica de nitrogen deu ser important a les vernedes, a causa de la simbiosi del vern amb un actinomicet fixador, i en aquells boscos on abundin les lleguminoses. A la resta dels nostres ecosistemes forestals pot haver-hi fixació de nitrogen per microorganismes de vida lliure (no simbiòtics) en el sòl o en la fusta morta, però aquest flux no ha estat mai quantificat als boscos dels Països Catalans.
Els principals fluxos de sortida de nutrients dels ecosistemes forestals són les emissions gasoses de sofre i nitrogen, l’exportació d’elements dissolts o particulats en les aigües de drenatge i l’extracció de nutrients que té lloc en els productes forestals que l’home retira. No hi ha informació sobre les emissions gasoses als nostres boscos. La sortida de nutrients en les aigües de drenatge ha estat considerada en un altre article amb les dades de dues conques d’alzinar a Prades i al Montseny. Com altres boscos relativament poc pertorbats, aquests alzinars són altament retentius per als nutrients essencials. En particular, les conques forestades solen acumular el nitrogen i el fòsfor d’origen atmosfèric. L’exportació de nutrients pot augmentar després d’una pertorbació, perquè aquesta redueix l’absorció de nutrients per la vegetació mentre que la descomposició i mineralització de la matèria orgànica continua a una taxa igual o més gran que la que es donava abans de la pertorbació. Això genera una acumulació de nutrients disponibles en el sòl que, si no és contrarestada per altres processos biogeoquímics, poden perdre’s amb l’aigua de drenatge o en forma gasosa. Experiments recents a l’alzinar de la Castanya, al Montseny, han demostrat que els boscos esclerofil·les mediterranis poden tenir una pèrdua molt elevada de nitrogen quan una pertorbació ha eliminat l’absorció d’aigua i de nutrients per les rels dels arbres.
Els boscos dels Països Catalans

Els boscos dels Països Catalans han tingut una història complexa, un component important de la qual ha estat la regeneració de les masses forestals durant els darrers cent anys, com a conseqüència de l’abandó de conreus i pastures marginals i de la caiguda en desús de pràctiques de gestió més intensives que les actuals. La regeneració del bosc, en aquest cas un alzinar muntanyenc, és evident en els vessants de la vall de la Castanya (Montseny), fotografiats el 1935 (a dalt) i el 1981 (a baix). En primer terme, la masia del Boscàs. La causa probable d’aquesta regeneració ha estat la fi del carboneig, anteriorment molt estès, que comportava la tallada arreu dels alzinars en torns curts. L’estructura dels nostres alzinars actuals, majoritàriament de rebrot, duu encara l’empremta del carboneig.
Hereus del Boscàs i F. Rodà.
Segons l’Inventario Forestal Nacional, els boscos dels Països Catalans espanyols ocupaven 1 680 000 ha quan es va fer l’inventari, entre 1966 i 1971. Aquesta extensió arbrada representa el 28% de la superficie implicada. Hi ha, però, grans variacions en tal percentatge, que va disminuint en general de N a S. Així, mentre que a les comarques gironines els boscos ocupen el 54% del territori i a les barcelonines el 47%, a les Illes ocupen el 21% i a Alacant només el 12%. Els boscos dels Països Catalans són molt variats, com correspon a un país muntanyós a la vora de la Mediterrània on hi ha una gran diversitat de climes i sòls.
Si considerem globalment tots els Països Catalans, el clima és el determinant principal dels tipus de bosc que trobem en cada contrada. Així, les pinedes de pi blanc (Pinus halepensis) són gairebé l’únic bosc de les terres meridionals, càlides i seques. Al N del Llobregat, amb un clima mediterrani subhumit, els boscos d’altituds moderades poden ser principalment de pi blanc (P. halepensis), pi pinyer (P. pinea), pinastre (P. pinaster), alzina (Quercus ilex), surera (Q. suber) o roure martinenc (Q. pubescens). A la muntanya mitjana poc plujosa, de clima continental, trobem sobretot boscos de pinassa (P. nigra), pi roig (P. sylvestris), carrasca (Quercus rotundifolia), roure de fulla petita (Q. faginea) o roure martinenc (Q. pubescens). A la muntanya mitjana plujosa els boscos dominants poden ser de pi roig (P. sylvestris), roure martinenc (Q. pubescens), roure de fulla gran (Q. petraea), roure pènol (Q. robur), faig (Fagus sylvatica) o avet (Abies alba). Finalment, a l’alta muntanya els boscos subalpins són principalment de pi negre (P. uncinata) i de vegades d’avet (Abies alba) o bedoll (Betula pendula). Les propietats del sòl poden influir també sobre el tipus de bosc, però sovint aquest efecte s’expressa a escala més localitzada que no pas el clima. La surera, el pi pinastre i el castanyer (Castanea sativa) requereixen sòls silicis, sense carbonats; la pinassa, en canvi, viu millor en sòls calcaris. Verns (Alnus glutinosa), salzes (Salix), pollancres (Populus) i oms (Ulmus) necessiten sòls molt ben proveïts d’aigua i per això es troben restringits a les fondalades i a les riberes dels cursos d’aigua. Sense negar el pes dels determinants climàtics i edàfics, cal tenir present que en un país com el nostre, amb molts milennis d’ocupació i utilització humana del territori, l’acció de l’home ha estat i és un dels factors principals en l’ecologia dels nostres boscos. En gran part, podem afirmar que l’estat actual dels boscos dels Països Catalans respon a l’activitat que hi ha desenvolupat l’home.

Els boscos de pi blanc (Pinus halepensis) són la formació forestal més estesa dels Països Catalans. Abundant tant sobre sòls calcaris com silicis, aquest pi de la terra baixa mediterrània constitueix masses més o menys obertes que solen dur com a sotabosc brolles diverses. El pi blanc és molt resistent a l’eixut i pot colonitzar sòls que semblen massa pobres per a altres arbres. Durant aquest segle, les pinedes de pi blanc s’han expandit molt arran de l’abandó dels conreus i de les pastures marginals. Els incendis dels darrers vint anys han castigat fortament aquestes pinedes, sobretot al Païs Valencià i a les comarques tarragonines.
Jordi Vidal.
Els boscos han estat sempre una font de primeres matèries gairebé del tot imprescindibles: fusta gruixuda per a construir edificis i vaixells; fusta per a fer mobles, carros, tanques i utensilis; llenya i carbó per a forns, fargues i llars; fibra per a fer paper; suro; fruits per a les persones i el bestiar; herba per als ramats; caça; tanins per a adobar les pells; i molts altres productes. En cada època i en cada bosc, hi predominava un o més d’aquests usos sobre els altres, el qual requeria sovint una gestió forestal específica. Més que en qualsevol altre ecosistema, el pes de la història és palpable en els boscos. Les diferents modalitats de pràctiques silvícoles, tant les passades com les presents, poden deixar en els ecosistemes forestals empremtes que perduren dècades o segles. Els boscos fan seva la frase de Margalef: «les estructures del present són obra del passat que projecten informació cap al futur».
El cas dels boscos de ribera

La inundació produïda per les grans avingudes afecta molt sovint el bosc de ribera (a dalt, el Segre, sortint de l’embassament d’Oliana al juny de 1981), els arbres i arbustos del qual solen presentar adaptacions fisiològiques que els permeten suportar l’ambient poc oxigenat que té el sòl sota la coberta d’aigua. Els efectes d’aquestes avingudes són força variables, sobretot segons el cabal d’aigua excedent i l’energia que porta associada. Els arrossegaments orgànics i inorgànics (a baix, el Fluvià a Esponellà, a l’octubre de 1987) creixen de manera exponencial amb l’augment del cabal. Així, els boscos de ribera són sotmesos a forts processos de pèrdua i d’intercepció de materials. La presència de materials orgànics interceptats resta com un testimoni de l’alçada màxima assolida per l’aigua en els moments de més descàrrega, cosa que, atesa la curta durada de les avingudes, seria difícilment detectable per altres sistemes.
Ernest Costa / Josep M. Barres
Al llarg dels cursos fluvials, i amb una certa independència de la vegetació zonal de l’indret, es donen formacions d’arbres i arbustos que representen una resposta a una major disponibilitat d’aigua, la qual va minvant a mesura que ens allunyem de les vores. En aquest biòtop les situacions més difícils per a la vegetació són a causa de dos efectes diferents de l’aigua: la inundació, que com en altres aiguals pot donar situacions d’anaerobiosi al sòl, i l’arrossegament a causa de les avingudes que afecten l’estructura i l’estabilitat de les plantes. Les comunitats de ribera pateixen aquests efectes en una proporció diversa que depèn, sobretot, del règim hidrològic. Molt sovint aquesta vegetació es dóna en bandes que segueixen els cursos d’aigua i representen diverses capacitats per a resistir els efectes esmentats de l’aigua. Els requeriments hídrics, en relació amb la fondària i la constància del mantell freàtic, són un tercer condicionament d’aquesta estructura zonal dels boscos de ribera.
Les característiques de les xarxes hidrogràfiques depenen força del substrat geològic per on discorren, el qual condiciona la permeabilitat dels llits i, per tant, el cabal, i també la composició química de l’aigua. Els sòls on creixen els boscos de ribera són usualment al·luvials, sense un perfil ben diferenciat i amb una textura força heterogènia, normalment amb un llit de materials d’un cert gruix (sorra o grava) disposats sobre materials més fins. Aquests sòls tenen una elevada porositat i encara que sempre hi hagi un mantell freàtic amb fortes oscil·lacions, l’ambient no sol ser reductor, o ho és només en els llocs on l’aigua es renova amb un ritme molt lent. L’existència d’una forta activitat d’humificació és una altra característica dels sòls de ribera.

Esquema de les comunitats del bosc de ribera del delta de l’Ebre, actualment restringit a àrees molt localitzades. Hom hi ha indicat els organismes següents: 1 salze blanc (Salix alba), 2 plàtan (Platanus hybrida), 3 pollancre (Populus nigra), 4 àlber (P. alba), 5 pollancre del Canadà (P. canadensis), 6 vern (Alnus glutinosa), 7 i 8 freixe de fulla petita (Fraxinus angustifolia, F. oxycarpa), 9 om (Ulmus minor), 10 tamariu gàl·lic (Tamarix gallica), 11 canya (Arundo donax), 12 canyís (Phragmites australis), 13 esbarzer (Rubus), 14 algues filamentoses, 15 i 16 potamogètons (Potamogeton natans, P. crispus), 17 volantí (Ceratophyllum demersum), 18 lligabosc valencià (Lonicera biflora), 19 aloc (Vitex agnus-castus), 20 heura (Hedera helix), 21 corretjola gran (Calystegia sepium), 22 corretjola borda (Cynanchum acutum), 23 martinet de nit (Nycticorax nyeticorax), 24 gamba roja (Tringa totanus), 25 xot (Otus scops), 26 ànec collverd (Anas platyrhynchos), 27 teixidor (Remiz pendulinus), 28 eriçó comú (Erinaceus europaeus), 29 balquer (Acrocephalus arundinaceus), 30 cargolet (Troglodytes troglodytes), 31 blauet (Alcedo atthis), 32 llop de riu (Esox lucius), 33 llobarro (Dicentrarchus labrax), 34 carpa (Cyprinus carpio), 35 guineu (Vulpes vulpes), 36 rata comuna (Rattus norvegicus), 37 polla d’aigua (Gallinula chloropus), 38 tórtora (Streptopelia turtur), 39 colobra de collar (Natrix natrix), 40 granota verda (Rana perezi), 41 mostela (Mustela nivalis), 42 merla (Turdus merula), 43 llúdria (Lutra lutra).
Román Montull; dades del Parc Natural del Delta de l’Ebre.
Malgrat l’escassa extensió que tenen aquestes comunitats als Països Catalans, llur diversitat és considerable, atesos els gradients climàtics i de resposta hidrològica existents entre les riberes altes pirinenques i les rambles del País Valencià. De comunitats caducifòlies, com ara vernedes, alberedes, omedes i salzedes es passa a una vegetació d’arbustos de fulla perenne, com són els tamarigars i baladrars. Les característiques fitosociològiques d’aquestes comunitats, com també les dels medis on es donen, s’han tractat ja en aquesta mateixa obra. D’una manera molt simplificada, la posició de les comunitats caducifòlies respecte de l’eix del riu pot servir per a resumir l’exigència hídrica i la resistència a les avingudes. Aquesta posició segueix el següent ordre: salzedes, vernedes, alberedes i omedes. Als vegetals que formen aquestes comunitats, hom hi reconeix alguns trets d’adaptació a un medi tan fluctuant com el que s’ha descrit. Molts són espècies pioneres de creixement ràpid. En algunes espècies concretes, com ara el vern (Alnus glutinosa), s’ha demostrat la capacitat de fixar nitrogen atmosfèric mitjançant la simbiosi amb el fong Frankenia alni; aquesta és també una característica d’espècies amb el caràcter de colonitzadores de medis oligotròfics. Certes espècies de ribera presenten característiques metabòliques especials similars a les descrites pels helòfits, que les fan resistents a la manca d’oxigen. Les vies metabòliques anaeròbiques acaben donant acetaldehid o etanol, que en aquestes plantes més adaptades al medi fluvial poden acumular-se a nivells més alts dels normals sense resultar tòxics. L’especial flexibilitat de les tiges de salze els representa també un cert avantatge com a espècies de primera línia i que reben la part més important de l’energia cinètica de les avingudes.
Hi ha poques dades sobre aspectes estructurals i funcionals dels boscos de ribera, amb excepció de les alberedes i pollancredes que tenen un gran interès silvícola. Sobre les vernedes hi ha un estudi força complet als Estats Units d’un bosc d’Alnus rubra d’uns trenta anys que presentava una biomassa aèria de 127,5 t/ha i 35,2 t/ha de rels. En altres vernedes europees d’Alnus glutinosa, de densitats que varien entre 1600 i 5700 peus/ha, s’han mesurat biomasses aèries de 62 i 109 t/ha. Pel que fa a les alberedes i pollancredes, i segons una recopilació de dades de dotze plantacions, mentre que la biomassa aèria de fusta varia de 21,5 a 198 t/ha, la de fulles ho fa entre 1,5 i 5,5. L’única dada trobada de biomassa de rels és de 24,7 t/ha, i correspon a una plantació de 29 m2/ha d’àrea basal i una biomassa aèria de 42,1 t/ha. Els valors de producció primària estimats per a les vernedes varien entre 6,4 i vora 12 t/ha·any, i per a les alberedes i pollancredes els valors poden arribar fins a prop de 25 t/ha·any. Per a fer-se una idea dels elevats valors de producció que poden donar aquests últims tipus de boscos es pot citar una pollancreda canadenca de 15 anys, 9400 peus/ha i una biomassa aèria de 53,8 t/ha, que té una producció total de 10,8 t/ha. Estudis sobre els cicles de nutrients en aquestes comunitats són encara menys freqüents. Per les concentracions dels diversos òrgans vegetals, aquest bosc es pot considerar intermedi entre les fagedes i les rouredes, i presenta, això sí, la particularitat de la fixació de nitrogen atmosfèric que en altres treballs s’ha vist que pot arribar a valors de fins 300 kg/ha·any.
| BOSCOS DE RIBERA | PRODUCCIÓ (t/ha·any) |
||||
|---|---|---|---|---|---|
| COMUNITAT | INTERVAL D’EDAT DELS ARBRES (anys) |
ÀREA BASAL DELS ARBRES (m2/ha) |
FULLES | BRANQUES I TRONCS |
TOTAL |
| VERNEDES | |||||
| Verns (55%) amb bedolls (44%) (Durham, Anglaterra) |
5-80 | 21,0 | 1,78 | 4,60 | 6,38 |
| Verns (20%) amb càrpinus (Daverdisse, Bèlgica) |
13-18 | 31,7 | 2,79 | 5,87 | 8,66 |
| Verns amb salzes (Fairbanks, Alaska) |
20 | 28,9 | 1,89 | 5,93 | 7,82 |
| POLLANCREDES | |||||
| Trèmols amb bedolls (Voronezh, Rússia) |
45-55 | — | 3,90 | 8,50 | 12,40 |
| Populus daviniana (Hokkaido, Japó) |
25-40 | 26,7 | 2,17 | 6,50 | 8,67 |
| Dades de M.J.R. Cannell | |||||
Les repoblacions i les plantacions

Les perxades de castanyer, plantacions tractades amb règim de tallada arran cada 20 anys i regeneració per rebrot, han conformat molts paisatges de la muntanya mitjana. Actualment, coneixen una certa decadència a causa de la davallada del valor de la fusta combinada amb l’aparició de malures greus (tinta i xancre) de procedència extramediterrània.
Lluis Ferrés
La repoblació i la plantació de boscos i arbredes no és pas una activitat recent sinó que ja s’ha dut a terme en èpoques passades, encara que no amb la intensitat dels últims decennis. Els canvis de l’ús del territori han provocat, sobretot en els últims 100-150 anys, l’abandó de conreus i pastures situats en zones poc productives. En alguns casos aquestes zones han estat senzillament abandonades, però en d’altres s’han dut a terme activitats de repoblació o plantació amb espècies autòctones o exòtiques. Recordem, només per la seva magnitud, l’abandó d’extensions importants de conreus de la vinya que van ser repoblades, en alguns casos, amb coníferes o amb sureres, aquests últims a les zones silícies. Altres vegades la puixança econòmica d’un tipus d’arbre n’ha provocat la plantació, sia en terrenys abandonats o fins i tot substituint altres tipus de boscos; aquest seria el cas del castanyer, molt rendible econòmicament fins fa 20 o 30 anys i plantat sobre grans extensions d’antics conreus (fins i tot substituint alzinars, rouredes i fagedes), que encarà avui podem veure a la zona compresa entre el Montseny i les Guilleries. Els canvis en el valor econòmic de les fustes han propiciat també les substitucions de boscos o les reconversions de terrenys agrícoles en plantacions, que han canviat el paisatge; pensem només en l’increment de les plantacions de coníferes durant els últims decennis, per la seva rendibilitat. La repoblació i la plantació es fan ara amb una barreja d’espècies autòctones i introduïdes seleccionades segons el creixement i el valor econòmic de la fusta.
Encara que només unes poques espècies ocupen grans extensions, s’ha assajat ja al nostre país la incorporació d’unes 300 espècies exòtiques, incluint-hi coníferes i planifolis; espècies actualment tan comunes com el pi insigne (Pinus radiata), d’origen americà, i diversos eucaliptus (Eucalyptus), d’origen australià, van ser introduïdes industrialment al nostre país al començament de segle. Actualment, les àrees susceptibles de ser ocupades per repoblacions o plantacions són extenses, ja que als antics conreus i pastures abandonats cal afegir les importants superfícies de bosc cremat (a la ratlla de desenes de milers d’hectàrees anuals). Cal, però, actuar amb cura, ja que no podem basar-nos únicament en valoracions econòmiques actuals a l’hora de portar a terme inversions a llarg termini.
Dinàmica i funcionalitat
Algunes repoblacions impliquen la regeneració de masses forestals degradades pel foc, excés de pastura o mala gestió forestal; un cop regenerades, aquestes masses poden restar com a bosc de protecció o ser explotades, de forma més o menys intensa segons la seva situació, amb els mètodes aplicats al bosc natural o autòcton (aclarides o tallades arreu).
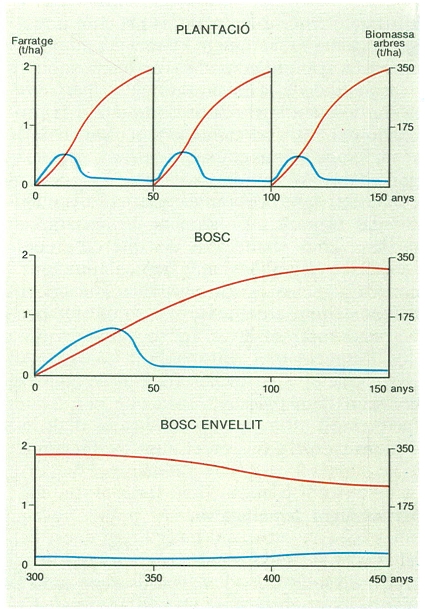
Evolució comparada en una plantació i un bosc de la biomasa d’arbres (en vermell) i la quantitat de recursos disponibles per als ungulats en forma de farratge (en blau). El caràcter periòdicament rejovenit de la plantació afavoreix durant curts períodes l’elevada producció de recursos consumibles pels ungulats. Un paisatge amb plantacions de diferents edats pot ser clarament més favorable per a algunes espècies animals que no aquells on domina el bosc natural o on les plantacions són sincronitzades.
Carto-Tec, original de J.A. Rochelle i F. Bunnell.
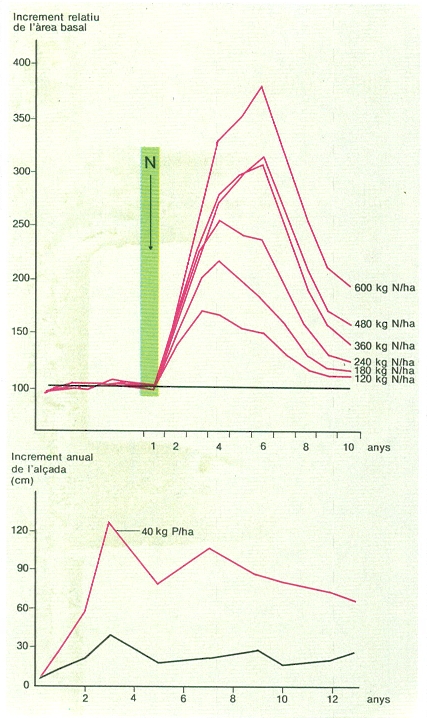
Efectes de l’adob (línia vermella) sobre el creixement d’arbres en plantacions (la línia negra correspon a la plantació sense tractar amb adob). En el gràfic superior veiem que l’adob amb compostos nitrogenats provoca un augment del creixement en el pi roig (Pinus sylvestris), però que el màxim no s’assoleix fins de 4 a 6 anys després de l’aplicació; el creixement, proporcional a la quantitat d’adob, decau després ràpidament. En el gràfic inferior veiem que per a una altra espècie de pi l’efecte de l’adob amb compostos de fòsfor es manté durant un període més llarg.
Carto-Tec, original de R. Ballard.
Serà diferent, però, el cas de les plantacions regulars en antics conreus o pastures o que han substituït masses forestals autòctones degradades o poc productives. Aquestes plantacions regulars són com conreus, en els quals la collita no és anual sinó que pot fer-se per períodes de 15 a 150 anys d’acord amb les espècies i usos de la fusta. El cas més semblant a un conreu, quant a torn i intensitat dels tractaments, el trobem en els pollancres: les pollancredes es reguen, s’adoben, es llauren (un parell de cops l’any) i, a vegades, s’hi fan tractaments de poda per millorar la qualitat de la fusta. En el cas de les coníferes, amb rotacions més llargues, que oscil·len entre els 30 i els 150 anys, els tractaments són menys intensos però més variats. Aquests tractaments inclouen les successives aclarides, que són encaminades a rebaixar la densitat de la plantació fins a deixar-la al nivell adequat per a la tala final, i la poda de les branques mortes (en les espècies que no fan autopoda), que contribueix a millorar la qualitat de la fusta.
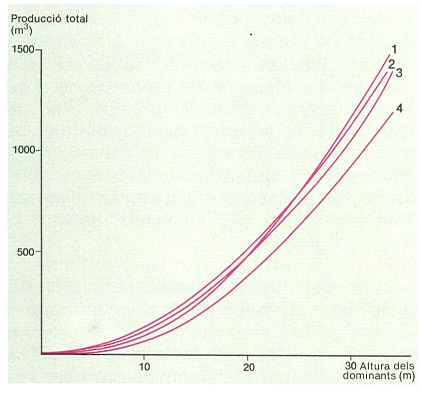
Relació entre la producció total de fusta i l’alçada dels arbres dominants en quatre plantacions de Picea excelsa d’àrees geogràfiques diferents 1 Massís Central, 2 Alps, 3 Jura i 4 Suàbia, RFA). Per a arbres d’una mateixa alçada, la producció total varia entre 900 i 1150 m3, segons l’àrea geogràfica i el temps que es triga a aconseguir una alçària determinada. La determinació d’aquesta relació (llei d’Eicchom) permet de definir l’índex de qualitat-producció de les parcel·les de plantació.
Carto-Tec, original d’A. Parde
Tractaments d’aclarida de plantacions de la pícea Picea excelsa proposats per dos autors diferents (Bartet 1976, en blau, i Braastad 1975, en vermell), en els quals varien les intensitats inicials de plantació (entre 2200 i 3000 arbres/ha), com també el nombre i la intensitat de les diferents aclarides. El factor d’espaiament o separació dels arbres (S) es defineix com la relació entre la distància mitjana entre arbres (plantats en xarxa triangular) i l’alçada dels arbres dominants, és a dir, S (%) = distància / alçada dels dominants × 100. Cada tipus de tractament té uns costos diferents i també dóna lloc a un producte final diferent. Per això, per a cada situació cal avaluar experimentalment el tractament més adequat.
Carto-Tec original de J. Pardé.
Importantíssims són els tractaments apropiats per a les diferents espècies (sobretot pel que fa a densitats inicials de plantació, com també a la intensitat i la periodicitat de les aclarides), ja que condicionen tant la producció global de la plantació com el seu rendiment econòmic. Cal tenir en compte tant el factor d’espai (relació entre l’alçada dels dominants i la distància entre els arbres) com els factors de morfologia i estabilitat (relació entre diàmetre i alçada dels dominants); és ben conegut que una excessiva densitat pot disminuir tant la producció global, per l’aparició de fenòmens de competència, com la qualitat de la fusta, a causa dels menors diàmetres individuals aconseguits.
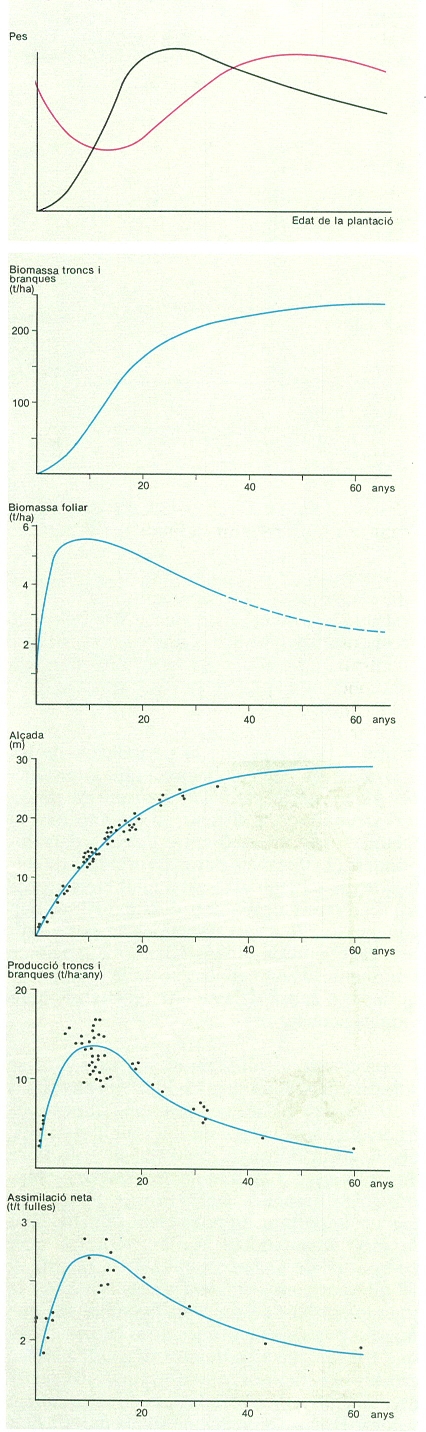
Els principals paràmetres que es relacionen amb la producció i el creixement de les plantacions tenen una evolució diferent dels seus equivalents en boscos naturals o fins i tot en boscos explotats, ja que varien en avançar l’edat de la plantació. La figura mostra aquesta variació en diferents paràmetres. En el primer cas (a dalt), es tracta de les pautes hipotètiques de caiguda de virosta (en negre) i de pes de matèria orgànica acumulada en el sòl (en vermell) durant el segon període de rotació d’una plantació, on s’observa que és un procés polsant, que es va truncant en arribar al final de cada torn. La resta dels gràfics es refereixen a l’evolució temporal de cada paràmetre en una plantació del vern Alnus rubra, des del moment de la plantació fins al final del torn.
Carto-Tec, original d’H.G. Milleri E.D. Ford
La producció assolida en molts casos, que se sol situar al voltant dels 15-20 m3/ha·any (unes 7-10 t/ha·any de fusta en pes sec) i que pot arribar, en torns curts i en llocs molt concrets, als 30-40 m3/ha·any (15-20 t/ha·any de fusta en pes sec), i a la qual caldria sumar la producció de branques, fulles i fruits, pot provocar un important empobriment del sòl; això fa molt aconsellable l’adob, sobretot amb compostos de fòsfor, després de la tala final. El diferent comportament de les espècies i la gran quantitat de situacions possibles fan que no se’n puguin donar unes normes fixes, tot i que cal dir que s’ha demostrat que l’adob té efectes beneficiosos, entre els quals cal destacar l’augment de la producció; en cada cas cal valorar els costos, la magnitud i la durada dels efectes per poder prendre una decisió racional. Cal no oblidar tampoc que les coníferes i alguns planifolis tenen una fullaraca amb una marcada tendència a acidificar el sòl, cosa que augmenta el grau d’oligotròfia. Pot ser aconsellable no solament un adob entre plantació i plantació, sinó també una rotació d’espècies o una barreja (diferents espècies arbòries o una combinació d’arbòries i arbustives) destinades a millorar, o almenys a mantenir, la qualitat del sòl.
La funcionalitat de les plantacions difereix de la del bosc en molts aspectes quantitatius i qualitatius. Les pràctiques intensives de conreu i l’ús de varietats altament productives fan que a les plantacions s’aconsegueixi una producció més elevada que en els boscos de les mateixes espècies; en el cas de l’avet de Douglass (Pseudotsuga menziesii) la producció de les plantacions és fàcilment el doble de la dels boscos de la mateixa espècie i en altres casos pot arribar a ser del 300-400%.
El caràcter intermitent de la funcionalitat de les plantacions, enfront del més continu del bosc, fins i tot del bosc explotat, fa que molts aspectes de la funcionalitat canviïn marcadament al llarg de l’existència de la plantació. La biomassa, la producció, la caiguda de virosta i la seva acumulació al sòl, la taxa de descomposició de la matèria orgànica, l’absorció d’aigua i nutrients, la penetració de la llum, etc. van canviant alternativament de forma gradual i brusca. Això afecta no solament el funcionament dels arbres de la plantació sinó també el de tots els organismes que en depenen. En casos de fauna molt especialitzada que requereix uns recursos molts concrets, la seva conservació pot veure’s compromesa; això passa amb espècies que necessiten arbres vells per a fer-hi el niu o amb algunes espècies de descomponedors, inclosa la fauna d’artròpodes associada a les soques i als troncs caiguts.
Alguns d’aquests problemes poden agreujarse perquè les plantacions solen fer-se en terrenys agrícoles marginals o en sòls no aptes per a l’agricultura per causa del pendent, la poca fondària o l’escassa capacitat de retenció d’aigua. Són, doncs, sòls relativament pobres i que poden patir de forma més marcada els efectes d’un conreu intensiu d’arbres.
El tractament de les plantacions com a masses regulars és clarament el més adequat si es tenen en compte només els costos econòmics i la qualitat de la fusta obtinguda, però presenta desavantatges, sobretot quant a la protecció del sòl. Cal evitar la sincronització de plantacions sobre grans extensions que obligui a tales arreu molt extenses. Com ja hem comentat, tant econòmicament com ecològicament és molt més adequat un paisatge en mosaic, amb plantacions de diferents edats i espècies alternades amb boscos autòctons i conreus, que assegurin una diversificació suficient del paisatge; això pot comportar també més rendibilitat, pel fet de situar en cada lloc l’espècie més apropiada, i assegura, a més, gràcies a la asincronia, un rendiment constant d’un paisatge, cosa que pot ser un factor molt estimable en condicions d’alt risc d’incendi com passa en el món mediterrani.
Metodologia de plantació

Les repoblacions o plantacions de zones desforestades situades en terrenys de fort pendent amb un tractament previ d’afeixament fet amb maquinària pesada constitueixen una pràctica discutible en regions de clima mediterrani: la pèrdua de sòl a causa de l’erosió provocada per les pluges torrencials representa una pèrdua gran, encara que difícilment avaluable en termes econòmics. La fotografia correspon als altiplans del Baix Cinca.
Jordi Vidal.
El procés de repoblació comença amb la selecció de la grana o l’esqueix i acaba amb la plantació en el terreny, un cop preparat. Cadascun dels passos és, evidentment, important per a aconseguir la finalitat proposada.
La selecció de la grana o l’esqueix és un pas bàsic en què cal tenir en compte tant la varietat o raça com la procedència geogràfica. Els responsables de vivers, fins i tot els dels organismes de l’administració, gairebé mai no en coneixen la procedència, i s’arriba a donar el cas que compren la grana a pes, sense demanar més informació, estimulant, així, que es recullin d’arbres petits i de llocs on les males condicions ambientals afavoreixen la producció de grans quantitats de llavors de baixa qualitat. En el cas de les espècies que es reprodueixen per esqueix, pot haver-hi una gran quantitat de clons adaptables a diferents condicions. Sobre aquests aspectes cal realitzar un esforç important en el camp de la recerca que permeti d’augmentar la quantitat i la qualitat de la fusta obtinguda.
Els tractaments de viver (sembra, selecció) són també importants per a assegurar la qualitat i la supervivència dels plançons emprats en la plantació. Els procés de plantació o repoblació pot ser més o menys costós segons les característiques del terreny, però no solament cal tenir en compte les variables econòmiques a l’hora de l’elecció, ja que algunes tècniques ràpides i poc costoses poden tenir efectes nocius perquè s’incrementa el risc d’erosió.
En terrenys plans o ja afeixats, la plantació en sots o en solcs són mètodes ràpids i poc costosos que no provoquen un augment del risc d’erosió. En terrenys amb fort pendent és on cal tenir més cura en la repoblació, ja que els processos erosius poden comprometre el futur de les plantacions; la plantació en sots, a pesar del seu cost, pot ser aconsellable ja que permet de mantenir la cobertura arbustiva. L’obertura de solcs seguint les corbes de nivell pot fer-se mecànicament i augmenta la taxa de supervivència dels plançons en remoure profundament el sòl; l’alternança de bandes de vegetació inalterades pot constituir una protecció efectiva enfront de l’erosió.
El mètode de l’afeixament o aterrassament amb maquinària pesant ha estat molt emprat, tot i que és el que provoca un impacte més gran sobre el medi. Implica una destrucció total de la vegetació preexistent i una remoció profunda del sòl, que queda al descobert i profundament alterat. La polèmica sobre els avantatges i els inconvenients d’aquest mètode ha estat i és forta, i s’ha arribat a assegurar que l’afeixament augmenta la infiltració de l’aigua i pot representar una protecció en front de l’erosió. Sembla evident, però, que la construcció de terrasses amb maquinària pesada no té res a veure amb l’aterrassament agrícola, on les feixes són protegides per parets, i, també, que difícilment es pot aconseguir una millor protecció del sòl que la fornida per una cobertura vegetal, encara que sigui arbustiva. La torrencialitat de la pluja de la regió mediterrània fa poc aconsellables aquests mètodes que provoquen la destrucció de la vegetació i la destrucció de l’estructura del sòl fins a fondàries importants en pendents acusats, encara que siguin mètodes poc costosos econòmicament.
Espècies de repoblació

Repoblació amb eucaliptus feta en terrenys de fort pendent i sòl sorrenc ocupats anteriorment per un bosc mixt d’alzina i surera que va ser destruït per un incendi. L’aspecte marronós dels arbres és degut als greus efectes de les fredorades intenses de l’any 1985. Aquesta plantació és un exemple complet: substitució d’un bosc autòcton de planifolis que haurien rebrotat, amb un mètode agressiu i emprant una espècie d’eucaliptus que no suporta els ocasionals freds intensos mediterranis. Un bon exemple d’una actuació forestal inadequada.
Jordi Vidal
Fins al començament d’aquest segle per a la repoblació s’empraven majoritàriament espècies autòctones de planifolis (castanyers, pollancres, plàtans) o de coníferes (pinassa, pi blanc, pi roig, pinastre). Més tard (1910-20) va començar el procés d’introducció d’espècies a gran escala, acompanyat d’un augment de l’interès i la rendibilitat econòmica de les coníferes. La creixent demanda de fusta per a pasta de paper ha comportat també l’augment de les superfícies dedicades als planifolis aptes per a aquesta finalitat, com és el cas dels eucaliptus, els quals, juntament amb el pi insigne, van ser introduïts industrialment al començament d’aquest segle.
Actualment són molt importants les plantacions de coníferes exòtiques, entre les quals destaquen el pi insigne (Pinus radiata), plantat en llocs de sòl profund i preferentment obacs fins a uns 600 m, molt corrent sobre antics conreus o substituint perxades de castanyer afectades per malures (xancre o tinta) o, senzillament, per la pèrdua del valor econòmic. L’avet de Douglas (Pseudotsuga menziesii) es planta en situacions semblants per sobre dels 600 m. Caldria citar també algunes varietats forasteres de pinassa (Pinus nigra, varietats de Còrsega i d’Àustria) i del pinastre (Pinus pinaster, varietats de Còrsega i del sud de França), plantades preferentment en solells per sobre i per sota dels 600 m, respectivament. S’han introduït moltes altres espècies de coníferes (aproximadament unes 200) però en plantacions que ocupen àrees molt restringides.

Les pollancredes, plantacions d’arbres idèntics genèticament, ja que es reprodueixen clònicament a partir d’estaques obtingudes d’exemplars seleccionats, són un magnífic exemple de silvicultura racional: ben integrades en l’ambient i amb una alta productivitat. Per a completar-ho, el seu valor paisatgístic és difícilment superable des del punt de vista estètic: el contrapunt canviant en un paisatge immutable, sempre verd.
Marcel·la Chinchilla
Quant als planifolis, cal destacar els eucaliptus, encara que als Països Catalans les superfícies ocupades no són gens importants comparades amb les que hi ha en altres llocs de la península Ibèrica. El fet que els eucaliptus rebrotin bé després del foc, com també la gran varietat d’espècies dins del gènere (unes 600) que s’adapten a condicions molt diferents, fa que siguin una alternativa a considerar, i més encara si pensem en la seva elevada producció. Cal tenir en compte, però, els fenòmens d’al·lelopatia provocats per la virosta d’algunes d’aquestes espècies, que donen lloc a la desaparició de la flora herbàcia i arbustiva i també a una acidificació i empobriment del sòl; caldrien estudis detallats sobre aquests extrems. S’han provat també altres espècies de planifolis (blades i roures americans, entre d’altres), però, fins ara, només en superfícies molt reduïdes.
Comentari a part mereixen els pollancres, les plantacions dels quals en terrenys de ribera ja són tradicionals al nostre país. Durant els últims anys s’ha avançat molt en la millora i la selecció de clons de pollancre (sobretot gràcies a les recerques fetes pels instituts d’investigació d’Itàlia i dels Països Baixos), derivats alguns de l’espècie europea (Populus nigra) però majoritàriament dels híbrids euroamericans (Populus nigra × Populus deltoides), de l’espècie americana Populus deltoides, o dels híbrids interamericans (Populus deltoides × Populus trichocarpà), molt ben adaptats aquests últims a les condicions de muntanya. S’han aconseguit clons resistents a les malalties més típiques (per exemple, el fong foliar Marsonina), clons adaptats a les condicions més fredes de muntanya i clons altament productius.
La recerca constant d’espècies d’alta productivitat, la desaparició d’antics usos de la fusta (bótes, rodells, carbó, mànecs, rodes de carro, etc.), l’aparició d’usos nous (més demanda de fusta per a pasta de paper, trituració, desenrotllament) i la gran quantitat de superfícies desforestades (antics conreus i pastures, boscos cremats) ha provocat la reforestació i la plantació de superfícies considerables, com també la introducció d’espècies forasteres. Tot plegat implica un canvi considerable del paisatge, i cal recordar que aquestes reconversions es fan sempre a llarg termini i que és, per tant, convenient de no guiar-se només per criteris econòmics actuals. Mantenir una certa diversitat d’espècies no és solament una bona estratègia ecològica i paisatgística, sinó també econòmica.