L’especial interès biogeogràfic que ofereixen els diferents grups de pisciformes consisteix en la llarga evolució que han sofert des de llur aparició a l’Ordovicià, ara fa uns 500 milions d’anys, com també en la seva àmplia distribució geogràfica, gairebé mundial. Pel que fa als peixos d’aigua dolça, s’hi afegeix la seva limitada capacitat de dispersió, que els condiciona a una distribució força supeditada a les diferències geogràfiques. Els peixos han mostrat una notable aptitud d’adaptació als diversos ambients aquàtics. N’hi ha per totes les mars i oceans de les diferents latituds, des de les regions àrtiques fins a les costes antàrtiques. Semblantment, les espècies d’aigües epicontinentals colonitzen gairebé totes les zones aquàtiques de la superfície terrestre, per insòlites que siguin. Així, els ciprinodòntids, com els fartets (Aphanius), Tellia, etc., poden viure fins i tot als oasis africans, i d’altres espècies similars que poblen les cavernes subterrànies d’Amèrica poden tenir una distribució molt restringida, com ara Cyprinodon diabolis, que només viu en una font de l’estat de Nevada, als Estats Units. Així i tot, no solen colonitzar les illes petites, ni tampoc les aigües hiperhalines (llevat dels ciprinodòntids), ni el continent antàrtic. Tant en les aigües epicontinentals com en les marines, el nombre més elevat d’espècies s’enregistra a les regions tropicals i minva progressivament vers els pols. Per contra, les aigües de latituds més boreals i australs solen tenir les poblacions més importants d’individus, tret dels peixos batipelàgics i batibentònics, que presenten els nombres més grans d’individus en àrees tropicals i subtropicals, prop dels continents; en aquest cas, les espècies tendeixen a ésser de mides més grans que els seus parents tropicals, i també s’hi observen més espècies migradores.
Conceptes generals
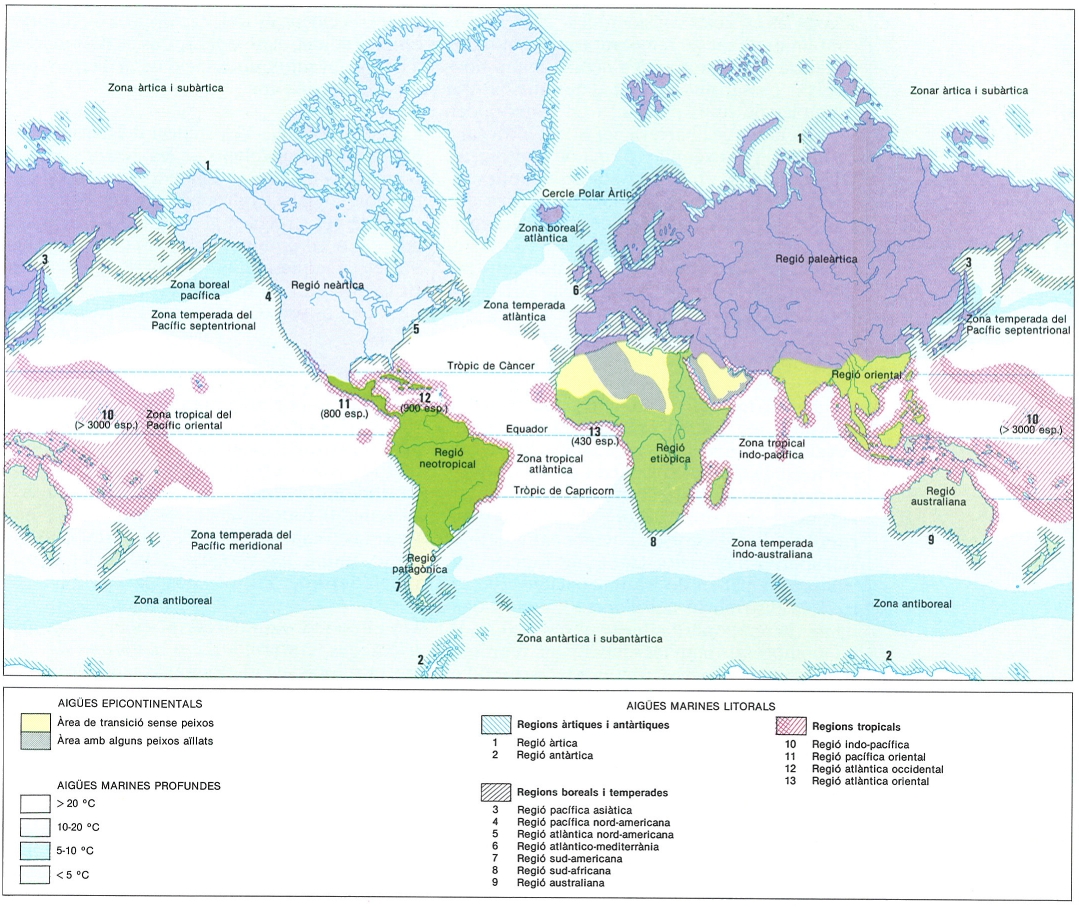
Regions ictiogeogràfiques del món. Pel que fa als peixos epicontinentals, les regions s’han indicat en el mapa en colors diferents, però mantenint les mateixes gammes de color per a les superregions (gamma dels liles per a la superregió holàrtica, dels grisos per a la panaustral, i dels verds per a la tropical). En el cas dels peixos marins, les regions litorals s’han marcat segons els colors indicats en la cartel·la; les oceàniques s’indiquen directament sobre el mapa (noteu la correspondència entre els límits de distribució dels grups d’espècies i les isotermes mitjanes anuals a la superfície dels oceans).
Carto-Tec, original de l’autor.
L’àrea de distribució d’un tàxon és l’ocupada d’una manera natural per les seves poblacions (espècies, gèneres o altres membres del tàxon), sense tenir en compte les introduccions fetes per l’home. La seva representació cartogràfica és un mapa de punts corresponents a les localitats on es troba el tàxon, o bé, d’una manera més general i fent l’aproximació d’uniformitat a la distribució, un mapa on es dibuixen superfícies, el contorn de les quals representa els límits de distribució; en realitat, aquest contorn depèn bastant de factors ecològics i geogràfics que, si més no, confereixen un caràcter dinàmic a les àrees de destribució. L’extensió de les àrees oscil·la entre el cosmopolitisme més ampli i l’endemisme més local; d’altra banda poden tenir un caràcter continu o bé ésser fragmentades; en aquest sentit, dos tàxons filogenèticament pròxims però que ocupen àrees espacialment disjuntes es consideren vicaris l’un de l’altre. Tanmateix, la repartició de les àrees constitueix una bona part dels estudis biogeogràfics. Aquests estudis també comporten de cercar explicacions satisfactòries per a la configuració de les àrees en el passat, a més de determinar-ne l’origen i llurs mecanismes evolutius.
Hom pot considerar que la biogeografia històrica segueix una evolució que passa per tres fases. La primera, la descriptiva, és necessària per a qualsevol estudi posterior i, històricament, assolí el seu apogeu amb les grans subdivisions biogeogràfiques fetes al segle passat. La segona fase, la narrativa, proposa uns arguments i unes teories per a explicar les observacions descrites; tanmateix, aquestes proves s’acostumen a invocar per reforçar o verificar la teoria inicialment suggerida per les observacions. Aquests criteri d’introducció a partir d’unes dades empíriques mena a una pobresa en el contingut de les teories formulades. Actualment, hom s’esforça a situar-se en una fase més madura, l’analítica, on només es plantegen hipòtesis que siguin refutables; aquest programa té un rigor científic més gran i substitueix el procés deductiu per un procés hipotètico-deductiu, en el qual la dinàmica amb què se succeixen les noves teories a partir de la falsificació de les ja existents, dona lloc a un veritable progrés científic.
Els coneixements actuals sobre les espècies vivents són incomplets, sobretot pel que fa a les regions tropicals. Aquest desconeixement s’estén a l’ecologia i a la capacitat de dispersió real de nombroses espècies; i encara és força més profund amb relació als mecanismes de diferenciació i evolució de totes les espècies en general.
El camp de la biogeografia ha estat dividit en tres escoles especialitzades: la dels factors ecològics, la de la dispersió i la de la vicariància dels organismes. Tot i haver estat propugnada i mantinguda per nombrosos autors, aquesta divisió és artificial i s’hi pot apreciar una complementarietat entre els diferents punts de vista. Actualment els esforços semblen encaminats a apropar les posicions de les diferents escoles i a intentar arribar a una síntesi entre totes elles.
La primera d’aquestes branques, l’ecologia geogràfica, posa l’èmfasi en l’estudi de les poblacions i en les relacions entre llurs caràcters adaptatius (és a dir, aquells que són influïts per la selecció natural) i el medi, a fi de determinar perquè una espècie colonitza una àrea. L’absència d’una espècie en una àrea ecològicament equivalent a una altra on és present depèn més de factors històrics que no pas de factors ecològics.
La biogeografia històrica s’ocupa de saber com s’han produït les distribucions actuals, basant-se en la història de la Terra i en l’evolució dels organismes. Els dispersalistes concentren la seva atenció en les possibilitats de dispersió dels organismes, i cerquen els centres d’origen de les espècies i les seves vies de dispersió. Malgrat la importància de les seves contribucions, aquesta escola ha tingut tradicionalment, com a aspectes deficients, d’una banda la freqüent construcció de teories narratives i de l’altra la resistència a considerar hipòtesis alternatives. Com a exemple del primer punt recordem l’ús de regles fixades per interpretar les distribucions en el passat, com ara la que situa el centre d’origen d’un grup taxonòmic en la seva àrea de més diversitat o també en la que conté les espècies filogenèticament més antigues (altres autors sostenen que les espècies noves són d’origen perifèric i envaeixen els nuclis de les ancestrals i les desplacen cap a la perifèria i, que, per tant, els centres d’evolució i de dispersió poden coincidir amb els d’extinció); en realitat, aquestes regles i d’altres de similars són vàlides per a problemes concrets, però no es poden generalitzar sense entrar en contradiccions. Pel que fa al segon punt, els dispersalistes van crear una sèrie de ponts intercontinentals hipotètics, actualment desapareguts, per tal d’explicar satisfactòriament les actuals disjuncions transoceàniques a partir de migracions de faunes terrestres senceres; aquesta argumentació fou paral·lela a una ferma oposició a la teoria de la deriva dels continents, fins que aquesta no va quedar definitivament demostrada pels camps de la física i de la geologia.
La biogeografia de la vicariància, que va aparèixer als anys setanta, proposa que la distribució de cada espècie és el resultat del procés següent: en primer lloc, l’espècie es dispersa per una àrea àmplia; en segon lloc, l’àrea es fragmenta en poblacions al·lopàtriques aïllades; després, les poblacions al·lopàtriques es consoliden com a espècies vicàries; finalment, les noves espècies poden desplaçar-se i sobreposar la seva àrea de distribució a les d’altres espècies. A diferència de l’escola de la dispersió, fonamentada en la sistemàtica evolutiva tradicional, la biogeografia de la vicariància es basa en la cladística o sistemàtica filogenètica formalitzada per W. Henning (1950), però els seus orígens es troben en l’obra de fitogeògraf Augustin P. de Candolle (1855). La cladística és un mètode de classificació que dona lloc a arbres d’afinitats relatives (anomenats cladogrames), en els quals s’identifiquen les parelles d’espècies germanes que tenen una semblança més gran sobre la base d’un sol criteri objectiu en general, i seguidament, per a cada grup, hom busca l’avantpassat immediat del qual és l’únic descendent. Substituint els tàxons representants en els cladogrames per les localitats geogràfiques que ocupen, s’obtenen els corresponents cladogrames d’àrees. Aquesta transformació, que és bàsicament una projecció d’una estructura filogenètica sobre l’espai geogràfic, és interessant perquè permet de plantejar hipòtesis biogeogràfiques per a grups monofilètics, que poden ésser quantificades i contrastades estadísticament. Les crítiques dirigides als seguidors de l’escola de la vicariància se centren en el paper secundari que aquests atribueixen a l’ecologia geogràfica i a la paleontologia dins la biogeografia històrica, i, sobretot, als seus atacs a Darwin i a llurs deixebles pel seu dogmatisme de considerar la dispersió com a diferent de la vicariància.
Finalment, els mètodes moderns també s’han beneficiat de la confirmació geològica de la teoria de la tectònica de plaques i de la deriva dels continents, que n’és la conseqüència directa. Segons aquesta teoria, en el Paleozoic, fa uns 225 milions d’anys, tots els continents en formaven un de sol, que hom anomena Pangea. Durant el Juràssic (fa 180 milions d’anys) aquest gran continent es va fragmentar i la porció nord o Lauràsia es va subdividir en els precursors dels actuals continents de l’Amèrica del Nord i Euràsia. En canvi, la porció sud o Gondwana va originar gradualment l’Amèrica del Sud, Àfrica, l’Antàrtida, l’índia, Austràlia i Nova Zelanda.
La taula següent mostra el nombre de famílies, gèneres i espècies de peixos vivents al món. Fan part de les espècies epicontinentals totes les de la divisió primària, la majoria de les de la divisió secundària i un nombre important de les perifèriques; per contra, s’han considerat les formes diàdromes com a típicament marines. El nombre d’espècies marines es pot deduir de les dues últimes columnes. En la sistemàtica utilitzada, l’autor estableix un total d’espècies de peixos inferior a la xifra de 20 000 citada en aquest mateix capítol. (La taula ha estat elaborada per l'autor amb dades de J.G. Nelson).
| Nombre d'espècies, gèneres i famílies de peixos al món | ||||
|---|---|---|---|---|
| ORDRE | FAMÍLIES | GÈNERES | NOMBRE D’ESPÈCIES | |
| EPICONTINENTALS | ||||
| Petromizontiformes | 1 | 9 | 31 | 24 |
| Mixiniformes | 1 | 5 | 32 | 0 |
| Heterodontiformes | 1 | 1 | 6 | 0 |
| Hexanquiformes | 2 | 4 | 6 | 0 |
| Lamniformes | 7 | 56 | 199 | 0 |
| Esqualiformes | 3 | 19 | 76 | 0 |
| Rajiformes | 8 | 49 | 315 | 10 |
| Quimeriformes | 3 | 6 | 25 | 0 |
| Ceratodiformes | 1 | 1 | 1 | 1 |
| Lepidosireniformes | 2 | 2 | 5 | 5 |
| Celacantiformes | 1 | 1 | 1 | 0 |
| Polipteriformes | 1 | 2 | 11 | 11 |
| Acipenseriformes | 2 | 6 | 25 | 15 |
| Semionotiformes | 1 | 1 | 7 | 7 |
| Amiformes | 1 | 1 | 1 | 1 |
| Osteoglossiformes | 4 | 9 | 15 | 15 |
| Mormiriformes | 2 | 11 | 101 | 101 |
| Clupeïformes | 4 | 72 | 292 | 25 |
| Elopiformes | 3 | 5 | 11 | 0 |
| Anguil·liformes | 22 | 133 | 603 | 0 |
| Notacantiformes | 3 | 6 | 24 | 0 |
| Salmoniformes | 24 | 145 | 508 | 80 |
| Gonorinquiformes | 4 | 7 | 16 | 14 |
| Cipriniformes | 26 | 634 | 3000 | 3000 |
| Siluriformes | 31 | 470 | 2000 | 1950 |
| Mictofiformes | 16 | 73 | 390 | 0 |
| Polimixiformes | 1 | 1 | 3 | 0 |
| Percopsiformes | 3 | 5 | 8 | 8 |
| Gadiformes | 10 | 168 | 684 | 5 |
| Batracoïdiformes | 1 | 18 | 55 | 2 |
| Lofiformes | 15 | 57 | 215 | 0 |
| Indostomiformes | 1 | 1 | 1 | 1 |
| Ateriniformes | 16 | 167 | 827 | 500 |
| Lampridiformes | 10 | 18 | 35 | 0 |
| Bericiformes | 15 | 39 | 143 | 0 |
| Zeïformes | 6 | 25 | 50 | 0 |
| Singnatiformes | 6 | 44 | 200 | 2 |
| Gasterosteïformes | 2 | 7 | 10 | 3 |
| Sinbranquiformes | 3 | 7 | 13 | 8 |
| Escorpeniformes | 21 | 260 | 1000 | 100 |
| Dactilopteriformes | 1 | 4 | 4 | 0 |
| Pegasiformes | 1 | 2 | 5 | 0 |
| Perciformes | 147 | 1257 | 6880 | 950 |
| Gobiesociformes | 3 | 42 | 144 | 2 |
| Pleuronectiformes | 6 | 117 | 520 | 3 |
| Tetraodontiformes | 8 | 65 | 320 | 8 |
| TOTAL | 450 | 4032 | 18818 | 6851 |
Les àrees biogeogràfiques dels peixos continentals
En l’àmbit mundial, s’han descrit unes 50 espècies d’àgnats (sense mandíbules), unes 550 espècies de peixos cartilaginosos (condrictis) i unes 20 000 espècies de peixos ossis (osteïctis); de tot aquest vast conjunt de peixos, n’hi ha uns 8275 que habitualment colonitzen les aigües epicontinentals i uns 11 650 que sempre viuen a la mar.
Dins dels peixos continentals, encara, poden distingir-se els que obligatòriament són d’aigua dolça i els que denoten diversos graus d’eurihalinitat, és a dir, una certa tolerància als medis salins. A fi de comprendre llur distribució i dispersió, se n’ha fet una classificació ecològica, que en considera tres grups principals: els peixos de la divisió primària, els peixos de la divisió secundària i els peixos de la divisió perifèrica. Els peixos de la divisió primària tenen poca tolerància salina, molt sovint gens, i, en conseqüència, són confinats a les aigües dolces en sentit estricte. Els peixos de la divisió secundària habitualment colonitzen les aigües epicontinentals, però la seva relativa eurihalinitat els permet de fer incursions a les aigües marines costaneres; cal esmentar que els avantpassats d’aquests peixos evolucionaren a la mar. Finalment, els peixos de la divisió perifèrica utilitzen la mar com a via de dispersió i ocupen les aigües continentals en alguna fase del seu cicle biològic; així, algunes de les espècies han quedat confinades a aquestes aigües continentals, mentre que d’altres són autèntics peixos marins que efectuen incursions als rius i als llacs.
Prop de la tercera part de les espècies de peixos ossis són ostariofisis (unes 6200 espècies), que juntament amb 450 altres espècies d’osteïctis formen la divisió primària. Si els afegim les aproximadament 1625 espècies de la divisió secundària, en resulta un total de 8275 espècies d’osteïctis que colonitzen les aigües epicontinentals. Això reflecteix la gran diversitat d’hàbitats possibles en les aigües no marines, tenint en compte que els oceans representen un 97% del volum de l’aigua mundial, mentre que les aigues dels rius i els llacs només arriben al 0,0093% (sense incloure l’aigua atmosfèrica, les aigües subterrànies, les masses de glaç, etc), i que, en conseqüència, un 41% dels peixos ossis del món ocupen menys del 0,01% del volum d’aigua de la Terra. Finalment, la divisó perifèrica inclou unes 115 espècies que habiten temporalment a la mar. Són peixos migradors diàdroms, que o bé es reprodueixen a les aigües dolces (anàdroms) o bé ho fan a la mar (catàdroms), però sempre la seva existència es troba molt lligada a les aigües dolces, fins al punt que diverses espècies de salmònids i de galàxids han establert definitivament llur residència als rius o als llacs, aïllats del medi marí.
W.L. Sclater i A.R. Wallace van definir, els anys 1858 i 1876 respectivament, els límits dels sis regnes o regions zoogeogràfiques continentals al món, basant-se en els conjunts de grups característics de les diferents parts de la Terra i en els nuclis d’endemisme. Actualment, encara conserven gran part de la seva validesa, però tanmateix, atenent als estudis més recents, sembla que convé definir unes regions ictiogeogràfiques lleugerament diferents a les considerades tradicionalment per a la resta de grups d’animals.
Les espècies del nostre país pertanyen a famílies característiques, sovint no exclusives, de la regió paleàrtica. Aquesta regió es caracteritza per una gran diversitat de ciprinoïdeus a l’Àsia meridional i per una escassesa d’espècies arcaiques o relictes, que es refereixen a Psephurus o Myxocyprinus, exclusivament. Es tracta d’un territori extens habitat per un nombre modest de grups de peixos: només 420 espècies i 36 famílies a l’URSS i Europa, enfront de les 550 espècies i les 48 famílies que viuen a Tailàndia, per posar un exemple. A més, el 50% de les espècies de les conques àrtiques són diàdromes i el 10% de les famílies són pròpiament marines. Les àrees de distribució de les famílies s’estenen latitudinalment sobre Àsia i Europa. Totes van rebre els efectes de les glaciacions amb el consegüent desplaçament de les faunes vers el S i l’extinció d’un elevat nombre d’espècies. Finalment, recordem que la regió paleàrtica té els seus límits meridionals més enllà de la Mediterrània i al N del Sàhara.
Segons les dades de la deriva dels continents, l’Amèrica del Nord i Euràsia es van separar definitivament a la darreria del Mesozoic. Això provocà que les ictiofaunes comunes dels dos continents evolucionessin independentment, amb la pertinent diferenciació de grups propis a cada banda de l’oceà Atlàntic, llavors recent. Aquest oceà actuava com una barrera infranquejable per als peixos de la divisió primària i molt sovint per als de la secundària. Les famílies pròpies de la regió neàrtica són els àmids, els ictalúrids, els ambiòpsids, els afredodèrids, els percòpsids i els centràrquids. Pel que fa a les famílies no endèmiques, la major part són compartides amb la regió paleàrtica, la qual cosa suggereix que ambdues regions formen una superregió holàrtica.
Hi ha tres regions tropicals: la neotròpica, l’etiòpica i l’oriental. Igual com les àrees boreals, aquestes regions tropicals tenen un origen comú en el primitiu continent de Pangea, però després dels desplaçaments continentals es modificaren notablement les seves afinitats faunístiques. La regió neotròpica (l’Amèrica del Sud i l’Amèrica Central) és la que té un nombre més elevat de famílies endèmiques de resultes del seu llarg aïllament, fins al punt que només comparteix algunes famílies primitives amb Àfrica i unes altres, de dispersió recent, amb l’Amèrica del Nord; es caracteritza pel fet de contenir conques hidrogràfiques enormes i una gran varietat de siluriformes i de caracins, que constitueixen la majoria de les 2000 espècies i les 46 famílies que la colonitzen. La regió etiòpica es distingeix perquè conserva diversos elements arcaics i presenta l’espectre filogenètic més complet de totes les regions, a més d’oferir la diversitat més alta de grups no ostariofisis (cipriniformes); es tracta d’un continent que disposa de rius molt antics i de grans llacs d’origen relativament recent. Finalment, la regió oriental és força heterogènia. Això és degut, d’una banda, a la invasió d’una fauna forània després de la col·lisió del continent indi amb Àsia. D’altra banda, també hi intervenen raons poc conegudes, com ara les relatives als seus límits a Malàisia. La regió oriental és dominada pels ciprínids, seguits dels bàgrids i els clàrids. Però, curiosament, no la colonitza cap gènere de ciprínid ni de silúrid, endèmics de l’índia, sinó que són compartits entre la resta de la región oriental i l’etiòpica. Pel que fa als elements no ostariofisis, es dona una situació força semblant.
Pel que fa a les famílies que ocupen la regió austral, només n’hi ha tres que pertanyin a les divisions primària i secundària: els ceratodòntids, els osteoglòssids i els melanotènids. Cal remarcar que aquesta regió comprèn, ultra la tradicional regió australiana, l’extrem sud de l’Amèrica del Sud a partir dels 38 graus de latitud S aproximadament, ocupat per famílies diàdromes. De fet tota la regió està molt lligada a la mar, car de les 18 famílies que hi viuen 15 corresponen a la divisió perifèrica.
La taula següent mostra el grau d’endemisme dels peixos epicontinentals a les regions biogeogràfiques, expressat com el percentatge de famílies endèmiques respecte del total de famílies que les colonitzen.
| Grau d'endemisme | |
|---|---|
| REGIÓ BIOGEOGRÀFICA | % DE FAMÍLIES ENDÈMIQUES |
| Neotròpica | 70 |
| Etiòpica | 47 |
| Oriental | 44 |
| Austral | 39 |
| Neàrtica | 30 |
| Paleàrtica | 10 |
Les àrees biogeogràfiques dels peixos marins
La biogeografia dels peixos marins és ben diferent de la dels peixos epicontinentals. Més que la immensitat dels oceans o el nombre d’espècies que hi habiten, és l’absència de barreres clarament definides per a la dispersió dels organismes, en un medi fluid i relativament homogeni com el marí, allò que dificulta la descripció dels patrons de distribució dels grups i la delimitació d’unes regions zoogeogràfiques.
La distribució de cada organisme depèn, a part les seves exigències ecològiques i la seva capacitat de dispersió, de factors estrictament oceanogràfics i geogràfics. En aquest sentit, els factors geogràfics es refereixen a la situació dels continents i a les característiques de les costes, però en general ofereixen poques barreres, atesa la continuïtat d’aquestes costes. En canvi, els factors oceanogràfics solen ésser determinants. El més important és el règim de temperatures anuals, però també hi intervenen els corrents marins i la salinitat, entre altres factors. A més, tenen efectes diferents, com a limitadors de les distribucions, segons les espècies considerades. Així, factors com un gradient de temperatura acusat o una distància important poden actuar com a filtre i deixar passar una espècie determinada (per exemple, de tipus pelàgic) i en canvi poden ser una autèntica barrera per a una altra (una de litoral, per exemple, o una de bentònica). D’altra banda, el cicle biològic de moltes espècies comprèn una fase larval pelàgica que pot permetre una dispersió a llarga distància per l’acció dels corrents marins; aquest factor tot sol pot arribar a explicar disjuncions notables sense haver de recórrer a la deriva dels continents.
La temperatura a la superfície marina oscil·la entre –2 °C, a les mars polars i, els 30 °C, a les aigües tropicals; tanmateix, entre zones pròximes les diferències són força inferiors, llevat de les superfícies de contacte entre corrents càlids i freds. Els peixos poden mostrar un cert grau de tolerància a diferents intervals de temperatura, segons el seu grau d’euritèrmia. Però a aquest component fisiològic se superposa el factor tròfic; les espècies filtradores depenen de les fluctuacions que sofreix la distribució del plàncton, condicionada per la temperatura, entre altres factors. Hi ha nombroses famílies que tenen representants septentrionals i representants meridionals, amb àrees de distribució més o menys paral·leles a l’equador o antiequatorials. L’exemple clàssic és constituït pels clupèids, cosmopolites però amb espècies exclusivament boreals (arengs, espadines), espècies d’aigües temperades (sardines) i espècies tropicals (sardinetes); són diverses les espècies de sardines que queden circumscrites entre les línies isotermes de 12 °C de la superfície marina a ambdós hemisferis.
D’altra banda, els corrents oceànics determinen certes migracions biològiques, com ara la de les anguiles de l’Atlàntic nord (Anguilla rostrata i A. anguilla): els adults de les anguiles migren a la mar dels Sargassos per a ferhi la fresa, a profunditats desconegudes; posteriorment, les larves d’A. Anguilla són transportades pel corrent del golf fins a les costes europees, on remunten els rius. També hi ha corrents submarins relacionats amb el relleu dels fons oceànics, de la plataforma i del talús continental. A més, n’hi ha d’altres d’abissals que comuniquen les enormes masses d’aigües fredes d’ambdós pols; la seva temperatura, relativament uniforme, en determinats casos facilita una dispersió d’espècies abissals en direcció latitudinal per sota les aigües superficials, a aigües més càlides.
A l’hemisferi nord es van donar importants migracions en direcció sud com a conseqüència de les glaciacions. Aquestes migracions foren especialment importants al llarg de les costes occidentals del Pacífic i de l’Atlàntic, on el gradient de temperatures és més marcat a causa del sentit de rotació de la Terra i, per tant, dels corrents.
Cal fer notar que l’existència d’espècies ecològicament equivalents a les regions polar, temperada o tropical d’ambdós costats de l’equador, sovint s’explica per l’evolució convergent de les espècies al llarg del seu procés adaptatiu en ambients similars però separats en l’espai; aquesta discontinuïtat equatorial es troba sobretot entre gèneres, tot i haver-n’hi pocs que es presentin fora de les latituds tropicals.
Altres discontinuïtats distribucionals que cal esmentar són les longitudinals. En aquest sentit hi ha espècies amfiboreals o antiboreals amb representants al nord del Pacífic i al nord de l’Atlàntic, però no pas a l’oceà Àrtic, que se situa entre els dos anteriors. Aquesta discontinuïtat probablement és fruit de les migracions d’espècies al llarg del litoral nord-asiàtic durant les èpoques càlides del Pliocè preglaciar i del Plistocè postglaciar.
Un altre aspecte interessant és el de les distribucions amfitropicals o antitropicals. Es tracta d’espècies idèntiques o molt pròximes que hi ha al Japó i en àrees tan allunyades com la Mediterrània, la costa occidental africana, el Carib i Califòrnia, i que tanmateix no es troben en les àrees intermèdies. És un cas de vicariància, com el de la distribució actual dels representants dels gènere Elops, una reminiscència de la primitiva mar de Tetis, situada entre Lauràsia i Gondwana, que constituí una gran via de dispersió de les faunes del Cenozoic.
Els peixos marins es poden classificar zoogeogràficament d’acord amb la zona de la mar on viuen. Segons aquest criteri, hom en pot distingir tres categories: la ictiofauna litoral, la ictiofauna pelàgica o de mar oberta i la ictiofauna abissal o d’aigües profundes. La ictiofauna litoral és la que viu a les costes, als escullals i en general a les zones que no ultrapassen els 200 m de profunditat; la immensa majoria de peixos marins pertanyen a aquest grup, com ara la família dels gòbids, amb unes 900 espècies, que és una de les més importants dins els peixos marins. La ictiofauna pelàgica o de mar oberta comprèn els peixos que viuen prop de la superfície, a menys de 200 m de fondària, però lluny de la costa en general; solament unes 250 espècies de peixos marins ocupen aquestes capes epipelàgiques il·luminades. La ictiofauna abissal o d’aigües profundes inclou els peixos marins que viuen per sota dels 200 m de profunditat (així i tot, es donen casos d’incursions en aigües més superficials); les 2300 espècies que conformen aquest grup poden ésser mesopelàgiques, batipelàgiques, abissopelàgiques, batíais, abissals o hadals. Pel que fa a les espècies mesopelàgiques, alguns autors les inclouen, amb les epipelàgiques, dins la ictiofauna pelàgica.
Pel que fa als peixos litorals, malgrat la mobilitat de moltes de les espècies i l’absència, en general, d’uns límits clars per a les distribucions marines, hom pot reconèixer quatre regions ictiogeogràfiques tropicals: la indo-pacífica i l’atlàntica oriental, l’atlàntica occidental i la pacífica oriental, totes quatre separades per continents o per grans extensions oceàniques gairebé infranquejables per a la gran majoria de les espècies litorals. La regió indo-pacífica és delimitada a l’W per la costa oriental d’Àfrica fins a la mar Roja, i a l’E, pel Japó, les illes Hawaii, la Polinèsia i la meitat nord d’Austràlia; els seus límits coincideixen amb els dels esculls coral·lins. La regió atlàntica oriental correspon a la costa occidental africana i té afinitats amb l’anterior, com també amb l’atlàntica occidental i la mediterrània. La regió atlàntica occidental s’estén des del sud de Florida fins a Rio de Janeiro (o bé, més al nord, a Bahia, segons els autors). Finalment, la regió pacífica oriental queda compresa entre el golf de Califòrnia i el nord del Perú. D’aquestes quatre regions tropicals, la indo-pacífica és la més rica d’espècies, sobretot pel que fa a l’àrea compresa entre les Filipines, Nova Guinea i Queensland (Austràlia); a continuació se situen la zona del Carib i la de Panamà al Pacífic, i, més lluny, l’oest africà, relativament pobre. La pacífica oriental mostra una tendència clara a diferenciar-se gradualment de l’atlàntica occidental des del tancament de Panamà, malgrat la barrera que representa la immensa extensió d’aigua oceànica profunda al Pacífic oriental.
Les regions àrtica i antàrtica són força empobrides. L’estabilitat de la convergència antàrtica fa que la regió del mateix nom sigui una de les més isolades; és notable que en cap d’aquestes dues no es trobin pràcticament espècies epipelàgiques. Per contra, els límits de les zones temperades boreals i australs fluctuen d’un any a l’altre, així com els dels corrents marins.
La Mediterrània és un bon exemple del caràcter difús de les fronteres entre regions. Aquesta mar forma part de la regió atlànticomediterrània, la qual comprèn les costes atlàntiques d’Europa i del nord d’Àfrica, com també la mar Càspia i la mar Negra. Malgrat el seu aïllament geogràfic, un 35% de les 450 espècies litorals i pelàgiques mediterrànies es troben també al nord del Cercle Àrtic a Europa.