Les característiques i les adaptacions antòmiques i fisiològiques
El tegument
La pell
La pell dels peixos és formada per dues capes, l’epidermis i el derma. La primera és constituida per diverses capes cel·lulars, entre 10 i 30; no obstant això, els signàtids, com el cavallet de mar (Hippocampus), presenten una epidermis extremament fina (2 o 3 capes de cèl·lules) i, contràriament, la de l’esturió (Acipenser sturio) pot assolir 3 mm de gruix. La capa exterior de l’epidermis sol ésser formada per cèl·lules esquamoses que constantment són substituïdes per d’altres originades a la zona més profunda, pròxima al derma. Aquesta capa externa és coberta d’un mucus de natura essencialment glicoproteica, que té una gran capacitat d’absorbir l’aigua i confereix a la pell el seu aspecte lliscós tan característic. Aquesta substància és secretada per les glàndules mucoses i la seva quantitat i composició química són variables segons els diferents grups de peixos; per exemple, en els àgnats (Myxine glutinosa), les glàndules secretores de mucus són especialment abundoses. El mucus dels peixos té una missió eminentment protectora contra els diversos tipus de malalties i contra el contacte amb determinats sòlids i soluts del medi aquàtic; a més, és important per al control de l’osmoregulació en algunes espècies, sobretot en les migratòries, que freqüentment alternen medis diferents, però també en les que no tenen escates, com les bavoses (Blennius). Alguns peixos, com ara els ciprínids, durant el període de reproducció desenvolupen una sèrie de tubercles nupcials o de protuberàncies que cobreixen diferents parts del cos, especialment la part anterior del rostre.
El derma és més gruixut que l’epidermis i d’estructura més complexa. És format per un estrat esponjós i un altre de compacte, el qual en realitat es tracta d’un teixit connectiu amb escassesa de cèl·lules. Al seu interior trobem cèl·lules pigmentàries (cromatòfors), vasos sanguinis, nervis, fotòfors i escates.
Les escates i les altres formacions externes
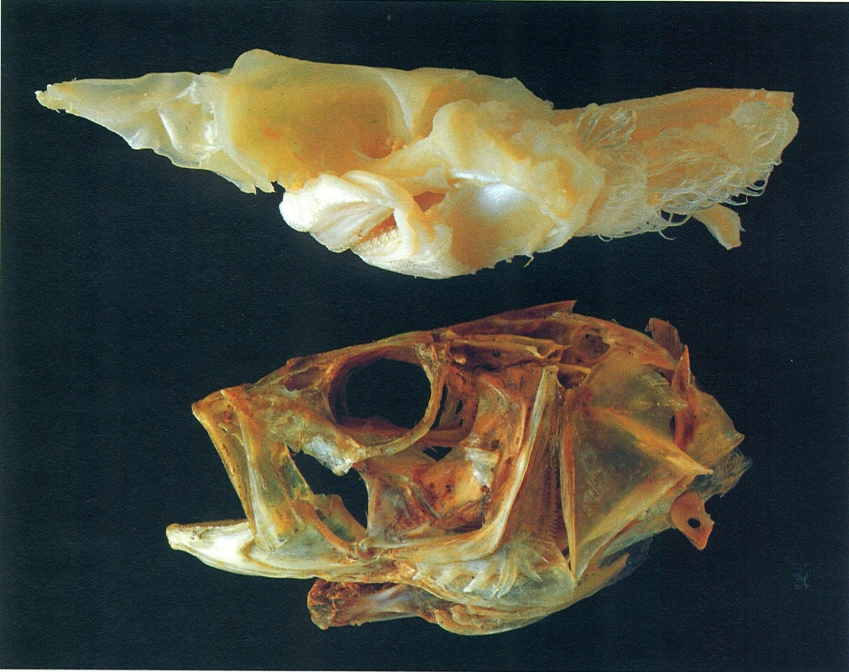
La immensa majoria d’osteïctis tenen el cos revestit d’escates d’origen dèrmic. No obstant això, algunes espècies les substitueixen per plaques o espines, com l’esturió (Acipenser sturio), l’espinós (Gasterosteus aculeatus) o el peix eriçó (Diodon hystrix). Per contra, n’hi ha que presenten la pell nua i llefiscosa, desproveïda d’escates, com els blènnids (bavoses).
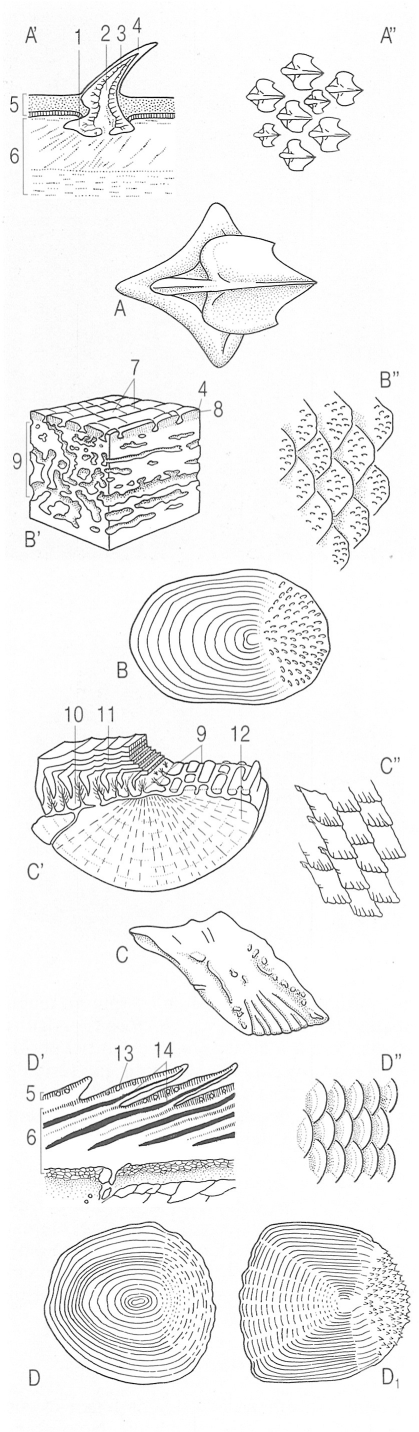
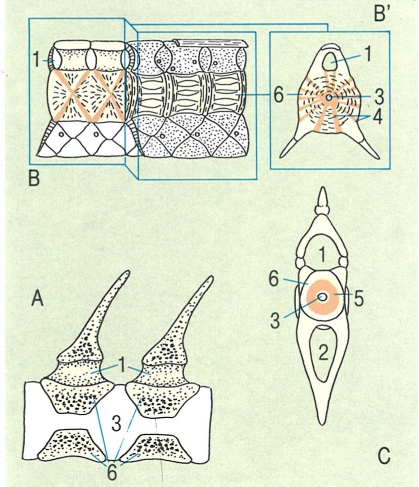
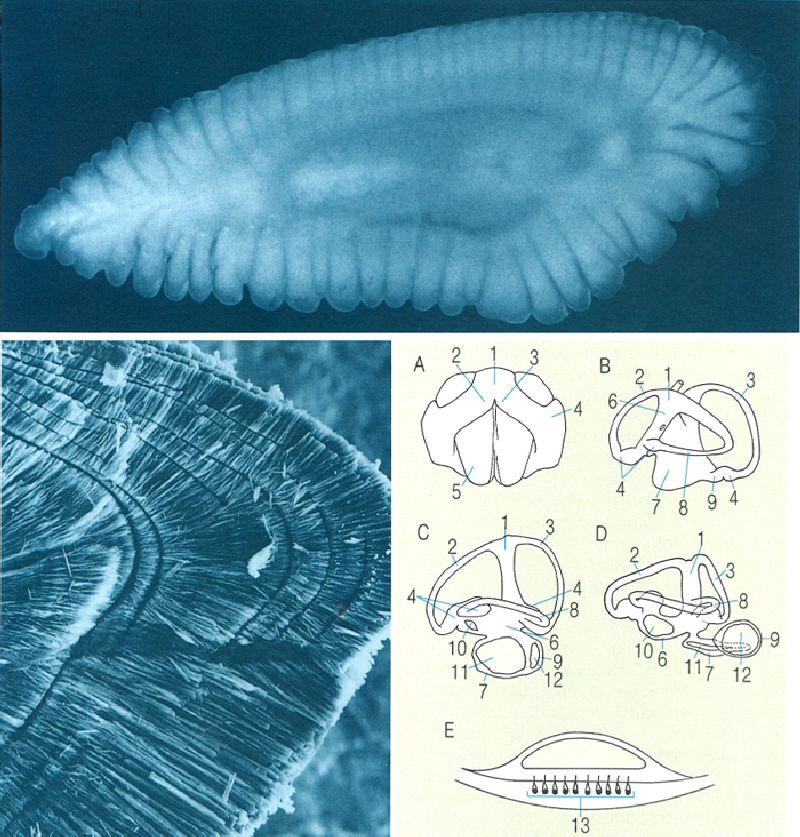
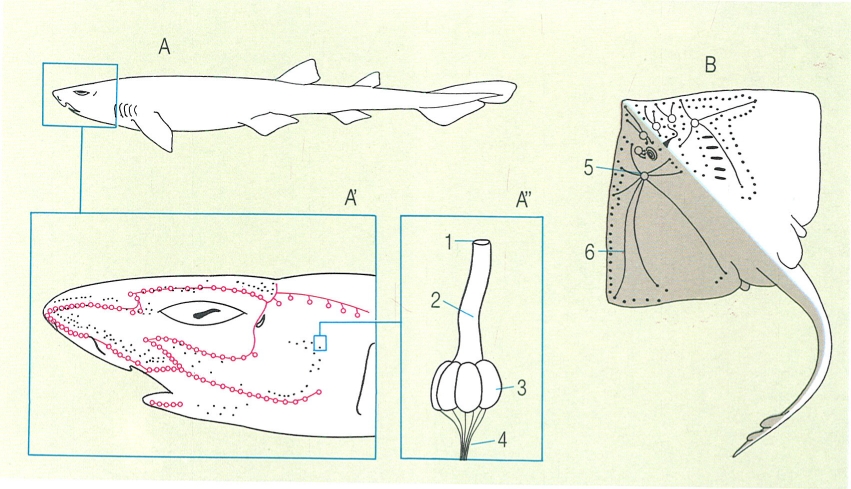
Diferents tipus d’escates en els peixos. A escata placoide o denticle dèrmic d’un peix cartilaginós i detall de la seva estructura en una secció sagital (A’) i de la seva disposició sobre la pell (A"). B escata cosmoide vista dorsalment, secció vertical d’una placa cosmoide d’un ostracoderm (B’) i visió general de la seva disposició en un actinisti, el celacant (B"). C escata ganoide d’un actinopterigi primitiu i detall de la seva estructura en una secció sagital (C’) i de la seva disposició en la pell (C"). D i D1 escates cicloide i ctenoide d’un teleosti, i detall de la seva estructura en una secció sagital (D’) i de la seva disposició en la pell (D"). 1 placa basal, 2 cavitat de la polpa, 3 vitrodentina, 4 esmalt, 5 epidermis, 6 dermis, 7 escates cosmoides, 8 cosmina, 9 os vascularitzat, 10 dentina, 11 ganoïdina (esmalt), 12 os laminar, 13 glàndula mucosa, 14 escata.
Amadeu Blasco, a partir de fonts diverses.
La majoria de peixos cartilaginosos (taurons i rajades) tenen el cos entapissat d’estructures exosquelètiques denominades denticles dèrmics o escates placoides, que ofereixen una arquitectura força peculiar: una placa basal, materialment enclosa en la pell, i una part externa, que bàsicament consisteix en una espina orientada vers la cua i que els dona un aspecte més aviat de dent que no pas d’escata; malgrat tot, la seva composició, a base de vitrodentina, revela que difereixen força de les dents dels mamífers. La morfologia d’aquestes estructures s’empra força com a caràcter sistemàtic dins aquest grup de peixos. Els denticles dèrmics tenen una funció protectora, però també són importants per a la natació del peix. A diferència de les escates dels peixos ossis, els denticles no augmenten de mida amb el creixement de l’individu. Llur forma, mida i distribució varia segons els grups i les espècies. Així, en gran part dels taurons revesteixen tot el cos, però en les rajades són escassos i es distribueixen d’una manera irregular. Un nombre ampli d’espècies els presenten forts i aguts, com per exemple el gatvaire (Scyliorhinus stellaris) i el porc marí (Oxynotus centrina), o bé poden ésser filiformes i flexibles, als agullats negres (Etmopterus), i fins poden mancar en els gèneres Torpedo (tremoloses), Dasyatis (escurçanes) i Myliobatis (milanes).
Plaques òssies de mida i forma variables, poden recobrir diferents parts del cos i fins i tot substituir les escates dèrmiques en determinats casos. En el dibuix s’han representat les de les espècies: 1 esturió (Acipenser sturio), 2 espinós (Gasterosteus aculeatus), 3 lluerna rafet (Aspitrigla cuculus).
Amadeu Blasco, a partir de fonts diverses.
Les escates dels peixos crossopterigis poden ésser, com en el cas del celacant (Latimeria chalumnae), de tipus cosmoide i d’estructura complexa; són formades per quatre capes compostes essencialment per una substància força semblant a la dentina: la cosmina.En les escates ganoides la capa de cosmina és substituïda per dentina i s’hi afegeix una capa inferior (isopedina), a més d’una capa superficial d’esmalt format per una substància denominada ganoidina.Els condrostis perden gran part de les escates i només els persisteixen algunes plaques al llarg del cos. A partir d’aquest model es troben diferents modificacions, com el cas dels holostis, en que les escates només tenen dues capes (Lepisosteus) o són transformades en discs ossis prims (Amia).
Les escates característiques dels peixos ossis evolucionats són les elasmoides. Són més fines i d’estructura més simple que les cosmoides i les ganoides: només són formades per capes òssies i per teixit connectiu; inserides al derma, se sobreposen i s’imbriquen entre elles de manera que només en queda una porció al descobert. Aquestes escates generalment creixen d’una manera més o menys proporcional a mesura que creix el peix. Això fa que es vagin dipositant capes òssies concèntriques a partir d’una zona central o focus, talment que, en ésser observades al microscopi, apareixen tot un seguit d’anells fins i concèntrics molt propers entre ells. Aquests anells, que augmenten de mida vers l’exterior, s’anomenen circuli. Hi ha encara altres marques, com els anells anuals (annuli), que consisteixen en unes bandes fosques de creixement lent, on els circuli són molt pròxims entre ells. Aquests anells anuals s’intercalen amb unes altres bandes més clares o zones de l’escata on els circuli són més separats. A més d’aquests anells també hi ha anells falsos i de reproducció, radis o estries, rugositats, denticulacions o ctenidis, etc.
Hi ha dos tipus d’escates elasmoides, les cicloides i les ctenoides. Les primeres són típiques dels peixos ossis no tan evolucionats i ofereixen una forma arrodonida o ovoide. La forma de les escates cicloides varia més ostensiblement i tant poden ésser circulars, en determinats ciprínids, com ovalades, en l’anguila (Anguilla). La mida de les escates també és sotmesa a un elevat grau de variació; si bé en l’anguila són menudes i estan encloses dins la pell, en la carpa (Cyprinus carpio) i la guerxa (Alosa alosa) són ben grans i força aparents. Algunes espècies presenten escates decídues que es desprenen fàcilment, com ara els clupèids (Sardina, Engraulis, etc.); tanmateix és més habitual que s’adhereixin fortament a la pell, com les de la truita (Salmo trutta). Les escates dels peixos ossis molt evolucionats són ctenoides, bàsicament. De morfologia més o menys quadrangular, aquest model d’escates es caracteritza essencialment per tenir unes fines denticulacions anomenades ctenoides, que apareixen a la porció de l’escata exposada a l’exterior i confereixen a la pell una certa aspror o adherència; el serrà ver (Serranus cabrilla) n’és un bon exemple.
La gran diversificació i especialització que hari sofert els teleostis es palesa en una notable variació dels tipus morfològics, que van des de les espècies mancades d’escates, com ara les bavoses (Blennius), fins a les que solament estan proveïdes d’escudets ossis, com l’espinós (Gasterosteus aculeatus). D’altres espècies combinen escates de mides i formes diferents, com per exemple a les lluernes (Trigla). Però també és freqüent de trobar un barreja d’estructures cicloides i ctenoides a diferents parts del cos dins d’una mateixa espècie, com s’observa en alguns burrets (gòbids) i en els pleuronectiformes o peixos plans.
És força comú d’utilitzar les escates com a caràcter sistemàtic i per això es comptabilitzen les que constitueixen la denominada línia lateral, que consisteix en un sistema sensorial localitzat a la zona mitjana de cada flanc. La línia lateral presenta un model d’escates foradades que poden apreciar-se a primer cop d’ull. D’altra banda, el nombre d’escates comptades en sentit transversal també té valor sistemàtic. En aquest cas, primer es comptabilitzen les escates del flanc superior, des de l’inici de la primera aleta dorsal fins a la línia lateral, i seguidament es compten les del flanc inferior, des del començament de l’aleta anal fins a la línia lateral, en sentit invers.
L’esquelet dels peixos
El condrocrani
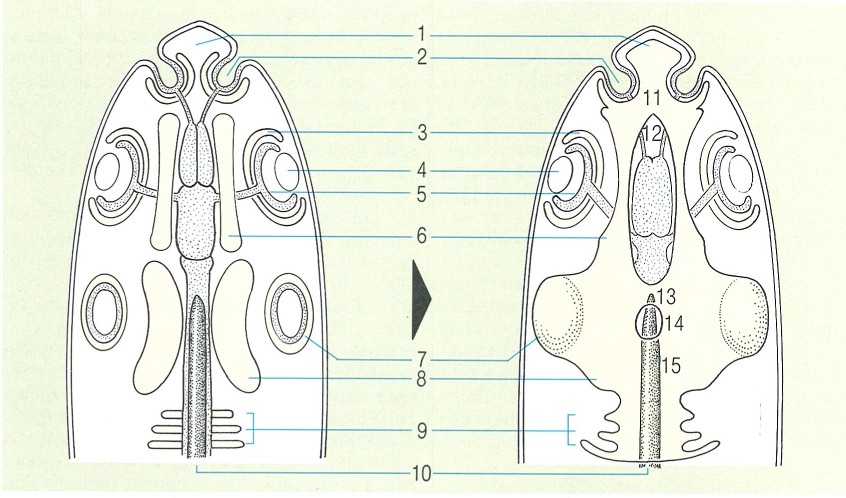
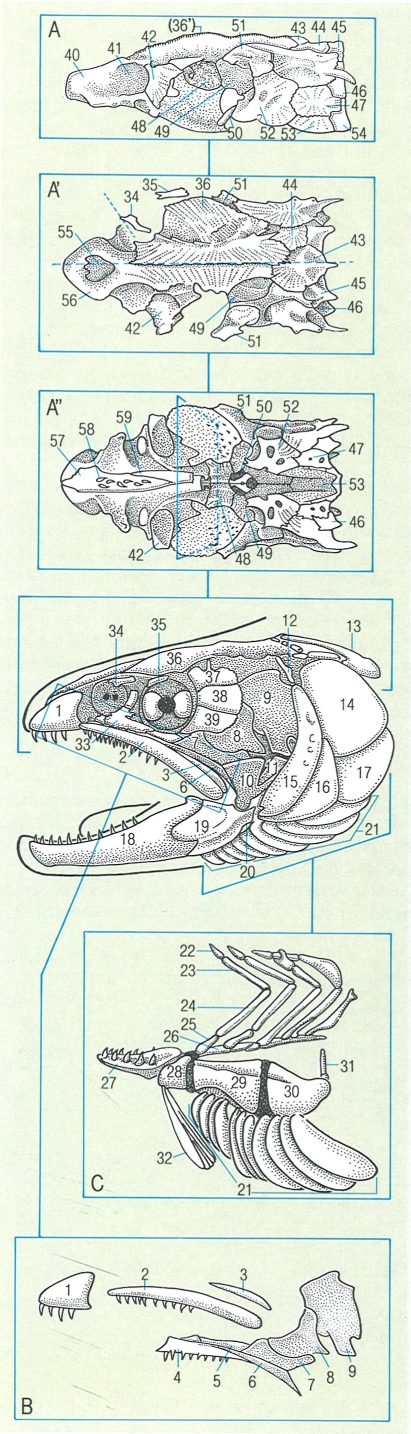
Esquema del desenvolupament del condrocrani o crani cartilaginós en els peixos. S’hi han indicat les parts: 1 rostre, 2 fosseta olfactòria, 3 càpsula òptica, 4 cristal·lí, 5 fosseta òptica, 6 cartílags precordals, 7 càpsula ótica, 8 placa paracordal, 9 vèrtebres occipitals, 10 notocordi, 11 placa etmoide, 12 finestra hipofissària, 13 barra acrocordal, 14 finestra basicranial, 15 placa basilar.
Amadeu Blasco, a partir de C. K. Weichert.
El desenvolupament del condrocrani, també anomenat crani cartilaginós, és força similar al dels diferents grups de vertebrats pel que fa referència a les primeres fases. S’origina a la part anterior del notocordi (o esquelet axial primitiu), poc després de la formació de l’encèfal durant el desenvolupament embrionari. Primer es formen un parell de cartílags o plaques paracordals que es disposen a cada banda del notocordi i s’expandeixen vers l’exterior fins a assolir les càpsules auditives; així, aquest parell de plaques paracordals acaben per constituir la denominada placa basal. D’altra banda, aquesta placa basal, que s’estén posteriorment fins a fusionar-se amb les vèrtebres que s’han soldat durant el procés evolutiu, al crani, constitueix la part posterior del neurocrani. Davant les plaques paracordals hi ha els cartílags precordals que, fusionats per l’extrem anterior, formen la placa etmoide; aquesta placa, al seu torn, es prolonga cap endavant i origina el rostre. En canvi, de la unió posterior d’aquests mateixos cartílags precordals amb la placa basal, en resulta la finestra hipofisària, que és una gran obertura que correspon a la hipòfisi. Simultàniament a aquest procés, també es desenvolupen les càpsules nasals, les òtiques i les oculars, que corresponen als òrgans de l’olfacte, l’oïda i la vista, respectivament. Aquestes estructures, unides a l’anterior, excepte les càpsules oculars que formen l’escleròtica, conformen el condrocrani.
En els pisciformes apareixen una sèrie d’arcs viscerals que envolten la regió faríngia i constitueixen el suport de les brànquies. Gran part d’aquests arcs viscerals es relacionen amb l’aparell branquial però també n’hi ha de modificats, que realitzen altres funcions específiques. El primer d’aquests arcs viscerals, denominat arc mandibular, es divideix en una porció dorsal i una altra de ventral: el cartílag palatoquadrat i el cartílag de Meckel, que respectivament contribueixen a la formació de la mandíbula superior i la inferior. El segon arc visceral dels peixos, l’arc hioïdal, es divideix en una secció dorsal o hiomandibular i una altra de ventral formada bàsicament pel ceratohial i el basihial. La hiomandibular es relaciona amb la suspensió o suport de les mandíbules, mentre que l’estructura ventral representa l’element de suport de la llengua. La resta dels arcs viscerals dels peixos sostenen les brànquies i és per això que reben la denominació d’arcs branquials. L’arc posterior pot trobar-se modificat en alguns casos perquè origina les dents faríngies pròpies d’alguns grups de peixos.
El crani dels àgnats
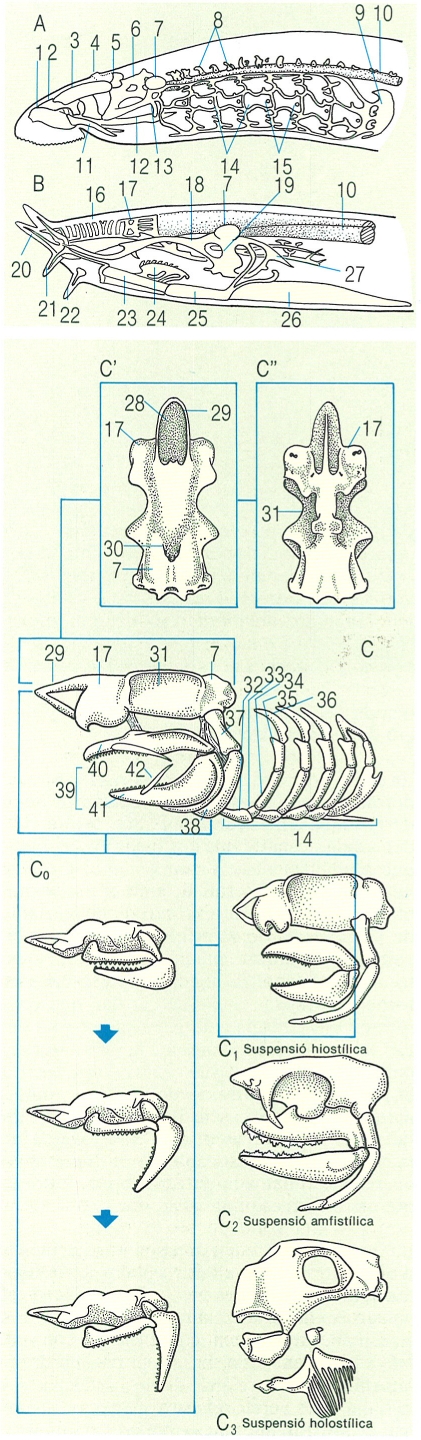
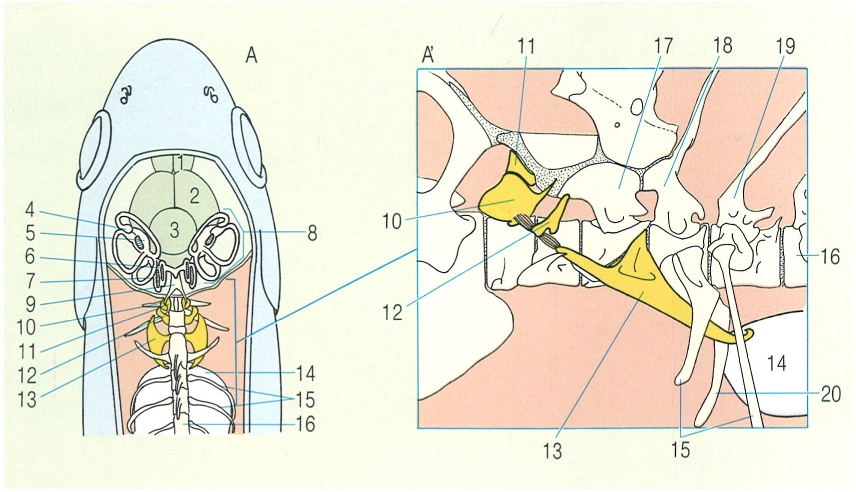
Esquema general del crani dels àgnats i dels elasmobranquis. A dalt: A vista lateral del crani d’un petromizontiforme, i B vista lateral del crani d’un mixiniforme. A baix: C visió lateral del crani d’un agullat (Squalus), detall de les parts dorsal (C’) i ventral (C"), i del tipus de suspensió mandibular (C1, C2 i C3); i C° esquema del moviment° d’obertura i tancament de la boca, que mostra l’avançament de la mandíbula superior i la rotació de l’os hiomandibular. 1 cartílag anular, 2 cartílag anterodorsal, 3 cartílag posterodorsal, 4 obertura naso-hipofisària, 5 càpsula olfactòria, 6 cartílag orbital, 7 càpsula òtica, 8 arcs neurals, 9 cartílag pericardial, 10 notocordi, 11 cartílag lateral anterior (estilet), 12 cartílag lingual, 13 cartílag estiloide (arc hioïdal), 14 arcs branquials, 15 obertures branquials, 16 cartílag dels tubs nasals, 17 càpsula olfactiva, 18 cartílag palatí, 19 arc hioïdal intern, 20 tentacle labial, 21 tentacle oral, 22 tentacles nasals, 23 cartílag lingual anterior, 24 cartílag dentari, 25 cartílag lingual mitjà, 26 cartílag lingual posterior, 27 cartílag intern posterior, 28 fossa precerebral, 29 rostre, 30 fossa endolimfàtica, 31 òrbita, 32 os basibranquial, 33 os hipobranquial, 34 os ceratobranquial, 35 os epibranquial, 36 os faringobranquial, 37 hiomandibular, 38 ceratohial, 39 arc mandibular, 40 palatoquadrat, 41 cartílag de Meckel, 42 cartílag labial.
Amadeu Blasco, a partir de fonts diverses.
El crani més primitiu que hom troba dins el grup dels àgnats és, sens dubte, el dels mixínids. Aquest crani presenta uns trets morfològics ben característics dels primers estadis del desenvolupament cranial dels pisciformes: una estructura cartilaginosa, que forma unes parets, i una volta de teixit conjuntiu. Les llamprees, però, tenen un crani una mica més evolucionat. El cervell és recobert per les expansions dels cartílags precordals que formen les parets i la volta de la regió cefàlica; la resta, és coberta per teixit conjuntiu. El conjunt del condrocrani (és a dir, els cartílags precordals i paracordals i les càpsules òtiques) és homòleg al dels vertebrats superiors. Hi ha uns altres cartílags que fan la funció de suport a la boca circular, la llengua, els òrgans olfactoris i la regió superior del musell (cartílags dorsals). La regió branquial és sostinguda per una estructura contínua que constitueix la denominada cistella branquial, que d’altra banda no és homòloga a l’esquelet visceral de la resta de vertebrats.
El crani dels condrictis
El crani d’un condricti, com el d’agullat ver (Squalus acanthias, a dalt), és format de cartílag i presenta una estructura més o menys simple. En els osteïctis, en aquest cas un serrà (Serranus, a baix), el crani és format per un gran nombre d’ossos, que poden tenir un grau variable d’ossificació; segons aquest grau parlem d’ossos de membrana o dèrmics i d’ossos de cartílag o endocondrals.
Jordi Vidal
Els condrictis, també coneguts pel nom de peixos cartilaginosos, com ara les rajades i els taurons, presenten un crani constituït per cartílag freqüentment calcificat. El condrocrani d’aquest gran grup de pisciformes és format per una sola peça, que no és sinó la reunió de tots els elements fusionats, i aquesta peça única és proveïda de tot un entrellat d’orificis que possibiliten l’entrada i la sortida dels vasos sanguinis i dels nervis cranials. La part anterior d’aquest condrocrani és força característica i constitueix l’anomenat rostre, la presència del qual obliga la boca a situar-se en una posició subterminal; en algunes espècies el rostre pot ésser de dimensions considerables. Les mandíbules dels elasmobranquis es formen a partir del primer arc visceral, que es divideix en dues parts: el cartílag palatoquadrat i el cartílag de Meckel, articulats entre ells a la part posterior. La suspensió d’aquests cartílags és possible gràcies a l’existència d’unes fibres lligamentoses que uneixen el palatoquadrat amb el cartílag hiomandibular; aquest darrer, que pertany al segon arc visceral anomenat arc hioïdal, alhora s’uneix amb la regió auditiva del crani. Aquest tipus de suspensió mandibular rep la denominació d’hiostílica i és la més comuna d’entre la majoria dels peixos vivents. Amb tot, un reduït grup de taurons tenen la suspensió amfistílica; en aquest cas al crani s’uneixen tant el cartílag palatoquadrat com l’hiomandibular. A la part inferior de l’arc hioïdal, a continuació del cartílag hiomandibular, hi trobem el ceratohial. A la part ventral es localitza el basihial, que connecta els ceratohials d’ambdós costats del peix. A continuació hi ha els arcs branquials, que en la majoria d’elasmobranquis són cinc, si bé en un reduït grup d’espècies n’hi ha un o dos d’addicionals. Cadascun d’aquests arcs branquials és constituït per tot un seguit d’elements cartilaginosos diferents: el basibranquial, l’hipobranquial, el ceratobranquial, l’epibranquial i, finalment, el faringobranquial.
El crani dels holocèfals, també denominats quimeres (Chimaera), que constitueixen un petit grup dins els condrictis, és diferent del que acabem de descriure. En aquests pisciformes, el cartílag palatoquadrat està completament fusionat al neurocrani i constitueix un tercer tipus de suspensió mandibular: la suspensió autostílica.
El crani dels peixos ossis
Esquema general del crani dels peixos ossis (el del dibuix correspon al salmó. Salmo salar), detalls de l’endocrani en visió lateral (A), dorsal (A’) i ventral (A"), i dels ossos que formen la mandíbula (B) i els arcs branquiostegals (C): 1 premaxil·lar, 2 maxil·lar, 3 supramaxil·lar, 4 palatí, 5 endopterigoides, 6 ectopterigoides. 7 pterígoides, 8 metapterigoides, 9 hiomandibular, 10 quadrat, 11 simplèctic, 12 supraopercular, 13 posttemporal, 14 opercular, 15 preopercular, 16 interopercular, 17 subopercular, 18 dentari, 19 angular, 20 retroarticular, 21 radis branquiostegals, 22 faringobranquial, 23 epibranquial, 24 ceratobranquial, 25 hipobranquial, 26 basibranquial, 27 basihial, 28 hipohial, 29 ceratohial, 30 epihial, 31 interhial, 32 urohial, 33 lacrimal, 34 nasal, 35 supraorbital, 36 parietal (36’ cresta), 37 intertemporal, 38 postorbital, 39 jugal, 40 rostre, 41 càpsula nasal, 42 etmoide lateral, 43 supraoccipital, 44, postparietal, 45 epiòtic, 46 intercalar, 47 exoccipital, 48 orbitosfenoide anterior, 49 orbitosfenoide posterior, 50 basisfenoide, 51 esfenòtic, 52 proòtic, 53 basioccipital, 54 vèrtebra, 55 sostre membranós de la cambra rostral, 56 endocrani cartilaginós, 57 vòmer, 58 dents vomerianes, 59 parasfenoide.
Amadeu Blasco, a partir de fonts diverses
El crani dels peixos ossis presenta una estructura esquelètica d’una gran varietat segons el grup concret que hom considera. Així, els peixos ossis primitius, com ara els esturions (condrostis), tenen un crani poc ossificat. Fins i tot entre els teleostis, els menys evolucionats (salmònids) encara tenen algunes estructures cartilaginoses, que en una gran part dels ordres de peixos han estat reemplaçades per teixit ossi pròpiament dit. El crani dels teleostis, en general, és constituït per una combinació d’ossos i d’estructures cartilaginoses; així, hi trobem ossos de membrana o dèrmics que procedeixen d’una ossificació directa del teixit conjuntiu, però també hi trobem ossos de cartílag o endocondrals, procedents del teixit cartilaginós ossificat. Per tant, es tracta d’un crani format per un complex d’estructures no comparables a les dels àgnats i els condrictis. Des d’aquesta base, el crani dels teleostis pot considerar-se com una estructura derivada del condrocrani cartilágines, del dermatocrani (compost per ossos de membrana) i de l’esquelet visceral o esplancnocrani, derivat dels arcs viscerals.
El neurocrani és format per una regió occipital, que inclou el foramen magnum i el còndil occipital còncau on s’insereix la primera vèrtebra. D’altra banda, aquesta estructura és constituïda per la regió de les càpsules òtiques o auditives, la regió orbital i, finalment, la regió de les càpsules olfactives.
La regió occipital, situada a la part posterior del neurocrani, és formada per quatre ossos de cartílag: el basioccipital, que forma part del còndil occipital; els dos ossos exoccipitals, que a ambdós costats conformen el foramen magnum i que, juntament amb el basioccipital, originen una estructura triple responsable de l’articulació de la primera vèrtebra; i, finalment, el supraoccipital, que forma la cresta occipital. D’altra banda, la regió òtica és formada per cinc ossos de cartílag que conformen una càpsula auditiva a cada costat del cap: el proòtic, que se situa a la part anterior, per davant de l’os basioccipital; l’esfenòtic, a la part posterior externa de l’òrbita i, juntament amb l’anterior, articulat a l’hiomandibular; el pteròtic, situat a la cara externa de la càpsula i projectat cap enrere; l’opistòtic o intercalar, situat a la part posterior, entre el pteròtic i l’exoccipital; i l’epiòtic.
La regió formada pels ossos cartilaginosos d’origen trabecular enclou els dos ossos alisfenoides o pterosfenoides, l’orbitosfenoides, el basisfenoides i el prefrontal o ectoetmoides. Els alisfenoides conformen la part superior de l’òrbita i estan en contacte amb el proòtic. L’orbitosfenoides, en canvi, és un os de posició mitjana, que en els teleostis menys evolucionats (clupeïformes, salmoniformes i cipriniformes) forma un septe inferior. El basisfenoides és un os imparell en forma de lletra Y localitzat a la part posterior de l’òrbita, i l’ectoetmoides es troba a la part anterior i constitueix la zona de contacte amb l’os lacrimal, d’origen dèrmic.
Els ossos dèrmics, o de membrana, en realitat formen el revestiment del condrocrani; estan situats a la volta i a la zona basal del crani. Els que ocupen la posició més posterior són els dos parietals, que recobreixen una part de la volta juntament amb l’os supraoccipital. Davant d’aquests parietals observem dos ossos de gran dimensió: els frontals. Els ossos frontals constitueixen el sostre del crani, incloent-hi la part superior de les òrbites. L’etmoides o dermetmoides, que és un os imparell, completa la part davantera del musell juntament amb els ossos nasals. A la part ventral més anterior de la cavitat bucal trobem el vòmer que, consegüentment, forma part de la boca i sovint és proveït de dents. A continuació hi ha el parasfenoides, un os llarg que posteriorment connecta amb el basioccipital i tanca completament la base del crani. A l’entorn de les òrbites hi ha un conjunt d’ossos anomenats circumorbitals.
Els ossos que constitueixen les mandíbules també presenten estructures d’origen doble, és a dir, ossificades i de cartílag. La mandíbula superior primària dels teleostis, el palatoquadrat, és homòleg al dels elasmobranquis, però en aquest cas és ossificat i format per diversos ossos: el palatí, amb dents a la part basal; l’endopterigoides, que revesteix la boca; l’ectopterigoides, que també és un os de revestiment i connecta el quadrat amb el palatí; el metapterigoides, situat sobre el quadrat; i, finalment, el quadrat, que presenta un còndil on s’articula la mandíbula inferior. Externament a aquesta estructura apareix la mandíbula funcional o secundària dels peixos ossis, essencialment formada per dos ossos de membrana premaxil·lars i maxil·lars, ambdós proveïts de dents. En alguns grups hi ha un petit os, el supramaxil·lar, a l’extrem posterior del mateix maxil·lar. La mandíbula inferior és constituïda per un os petit anomenat retroarticular, o suprangular, i per l’os angular, o articular, que s’articula amb el quadrat i es projecta cap endavant originant una regió sense ossificar o cartílag de Meckel. El dentari, proveït de dents, s’uneix a aquest cartílag.
L’arc hioïdal dels teleostis és ossificat en parts diferents. L’os més gran és l’hiomandibular, que s’articula a la regió òtica del crani i juntament amb el simplèctic actuen com un suspensor perquè es traven al metapterigoides i al quadrat respectivament. A la cara interna del segon arc apareix un os petit, l’interhial, que s’articula entre l’hiomandibular i el simplèctic i fa possible la suspensió de l’epihial, el ceratohial, els hipohials (superior i inferior) i el basihial, que suporta la llengua i en algunes espècies és proveït de dents (Salmo). Travats a l’arc hioïdal hi ha alguns ossos de membrana que formen la sèrie opercular i que en conjunt constitueixen l’opercle. L’estructura de l’opercle és decisiva durant la respiració de l’animal i és la responsable d’activar el flux d’aigua a través de les brànquies. Aquests ossos operculars són: el preopercular, l’opercular, el subopercular i l’interopercular. La cambra branquial dels peixos teleostis queda tancada gràcies a l’existència d’una sèrie de radis branquiostegals situats a cada flanc, a continuació de la sèrie opercular descrita; aquests radis branquiòstegs s’articulen sobre els ossos epihial i ceratohial. Finalment, cal esmentar la presència d’un os de membrana imparell, l’urohial, en contacte amb el basihial.
Els cinc arcs branquials dels teleostis consisteixen en una sèrie d’ossos de cartílag; els tres primers arcs consten dels segments faringobranquial i epibranquial a la part superior, i del ceratobranquial i l’hipobranquial, a la inferior. Al quart arc branquial l’os faringobranquial és unit al tercer arc i hi manca l’hipobranquial. Finalment, el cinquè arc branquial consisteix en un os únic. Els segments inferiors dels diferents arcs estan connectats per una sèrie d’ossos basibranquials.
La columna vertebral i les costelles
Morfologia de les vèrtebres dels peixos, de tipus amficèlic (A), i centres de calcificació (en color taronja): B calcificació radial (esterospondília) en una vèrtebra de tauró (Lamna), en secció transversal i en visió frontal (B’), i C calcificació en anell (ciclospondília). 1 canal neural, 2 canal hemal, 3 notocordi, 4 calcificació radial, 5 anell de calcificació, 6 centre.
Francesc J. de Sostoa, a partir de fonts diverses.
L’esquelet axial primitiu, o eix principal del cos, consisteix en el notocordi, inclòs en una beina de teixit conjuntiu fibròs que, gairebé en tots els vertebrats és substituït per la columna vertebral, si bé encara persisteix en els àgnats i en la fase adulta d’alguns peixos. La columna vertebral és formada per una sèrie de segments o vèrtebres que van des de la base del crani fins a l’aleta caudal. Les vèrtebres s’articulen entre elles i donen rigidesa al cos, però també el doten d’una certa flexibilitat. En la majoria de peixos les vèrtebres són constituïdes per teixit ossi, tot i que també poden ésser cartilaginoses. La vèrtebra típica és formada per una estructura cilíndrica, o centre, i per una sèrie d’arcs: l’arc neural, a la part superior i l’hemal a la inferior; això no obstant, també s’hi poden trobar d’altres elements, segons el grup de peixos que hom consideri.
Els àgnats tenen uns arcs cartilaginosos incomplets que són vestigis dels arcs neurals i hemals; aquestes vèrtebres no tenen centre. Els més primitius són els mixínids, que només tenen un notocordi inclòs en una beina cartilaginosa. Les llamprees presenten dos parells de rudiments d’arcs neurals per segment.
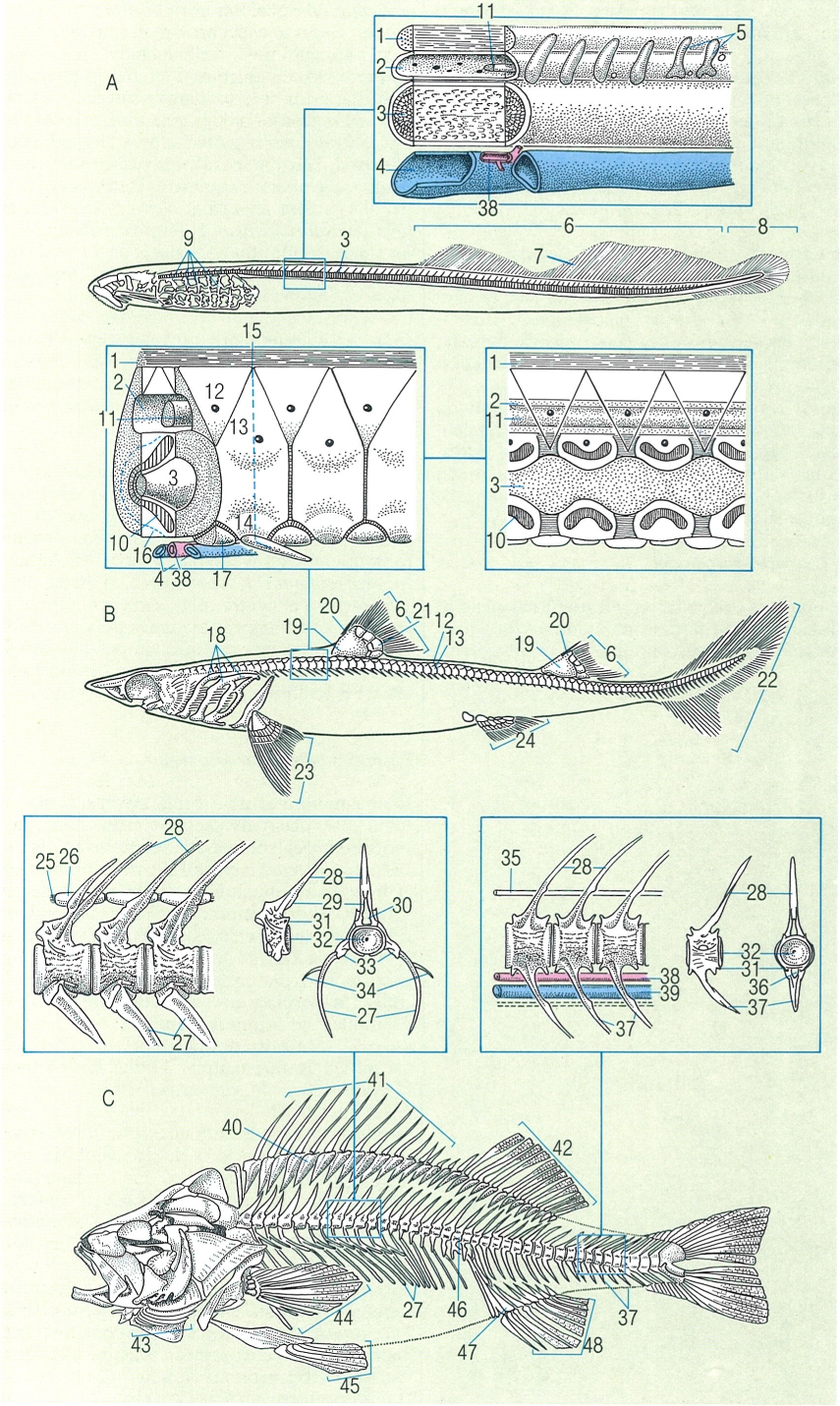
La columna vertebral i les costelles en els àgnats (A), en el cas concret de la llamprea de mar (Petromyzon marinus); en els condrictis (B), en el cas de l’agullat (Squalus); i en els osteïctis (C), en el cas de la perca (Perca), amb detalls de les seccions transversal i sagital en cada cas. 1 lligament espinal, 2 canal neural, 3 notocordi, 4 vena postcardinal, 5 arcs neurals, 6 aleta dorsal, 7 radis de l’aleta dorsal, 8 aleta caudal dificerca, 9 cistella branquial, 10 àrea calcificada, 11 medul·la espinal, 12 placa interdorsal, 13 placa basidorsal, 14 placa basiventral, 15 miosepte, 16 límit de la càpsula notocordal, 17 placa interventral, 18 arcs branquials, 19 cartílag basal, 20 espina, 21 cartílags radials, 22 aleta caudal heterocerca, 23 aleta pectoral, 24 aleta pelviana, 25 lligament neural, 26 supradorsal, 27 costella pleural, 28 espina neural, 29 arc neural, 30 canal neural, 31 zigapófisi, 32 centre, 33 basiapòfisi, 34 os intermuscular, 35 lligament interneural, 36 canal hemal, 37 espinal hemal, 38 aorta dorsal, 39 vena caudal, 40 pterigiòfor, 41 radis espinosos, 42 radis tous o segmentats, 43 radis branquiostegals, 44 radis de l’aleta pectoral, 45 radis de l’aleta pelviana, 46 primera vèrtebra caudal, 47 pterigiòfors de l’aleta anal, 48 radis de l’aleta anal.
Amadeu Blasco, a partir de fonts diverses.
Els elasmobranquis tenen una columna de tipus cartilaginós amb un centre amficèlic bicòncau que és travessat pel notocordi. Aquest notocordi presenta una constricció a la zona on travessa el centre de la vèrtebra i s’eixampla a l’espai comprès entre les cavitats de les vèrtebres contigües; aquesta particularitat li confereix una forma de rosari. A les parts anterior i posterior del centre, algunes espècies presenten una simple capa cilíndrica de calcificació (ciclospondília); en canvi, n’hi ha que tenen diverses capes cilíndriques (tectospondília), i d’altres amb una calcificació radial que, en adoptar una forma estrellada, reben la denominació d’esterospondília. A la part superior, per sobre del centre, hi apareixen els cartílags que formen l’arc neural i tanquen la medul·la espinal. Entre les plaques neurals s’observen les plaques o cartílags intercalars, i així s’origina una estructura contínua. A ambdós costats de la part ventral de les vèrtebres del tronc hi ha unes plaques cartilaginoses o processos transversos. En canvi, a les vèrtebres de la regió caudal aquestes estructures són substituïdes per l’espina hemal i els arcs hemals, que tanquen les artèries i les venes caudals.
En els teleostis, la columna vertebral és òssia i constituïda per dos models de vèrtebres, les del tronc i les caudals, mal que també es trobin diferències segons el grup de peixos. Gran part dels teleostis presenten vèrtebres amficèliques, amb restes de notocordi que, en forma d’un cordó estret, travessa la part central del cos vertebral i també reomple l’espai bicòncau que resta entre dues vèrtebres contigües. Els centres vertebrals s’articulen mitjançant unes petites estructures o zigapòfisis; aquestes són prolongacions de l’arc neural i se situen a les cares anterior i posterior de la vèrtebra. A la part superior de la vèrtebra hi ha l’arc neural que es prolonga en una llarga espina; a la inferior hi ha les basiapòfisis o parapòfisis, que s’articulen a les costelles. Els denominats ossos intermusculars es troben a la zona inferior, prop de les basiapòfisis (epipleurals) i a la base de l’arc neural (epineurals). A la regió caudal, les basiapòfisis es fusionen i originen l’arc hemal, que es projecta cap avall i forma l’espina hemal.
La zona posterior de la regió caudal mereix una especial atenció atesa la seva configuració ben apropiada per a sostenir l’aleta caudal. En la majoria de teleostis poc evolucionats, els centres de les vèrtebres són corbats, orientats cap amunt i formen una estructura denominada uròstil. A la part inferior d’aquest complex els hipurals substitueixen els arcs hemals i sostenen els radis de l’aleta caudal. Força sovint, en els teleostis poc evolucionats, els hipurals tendeixen a fusionar-se i a formar grans plaques que donen suport als lòbuls superior i inferior de l’aleta.
L’esquelet apendicular
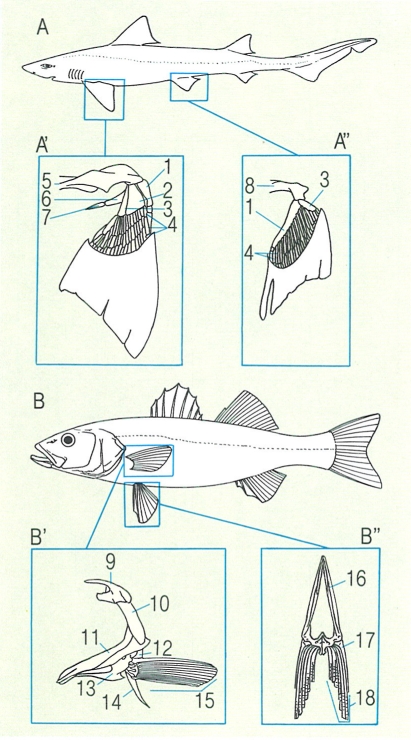
Esquelet apendicular en els elasmobranquis (A) i en els peixos ossis (B), amb detalls de les cintures escapular (A’, B’) i pelviana (A", B"). 1 protopterigi, 2 mesopterigi, 3 metapterigi, 4 cartílags radials, 5 barra coracoide, 6 escapular, 7 suprascapular, 8 cartílag pelvià, 9 posttemporal, 10 supracleitre, 11 cleitre, 12 escàpula, 13 coracoide, 14 postcleitre, 15 radis de l’aleta pectoral, 16 ossos pelvians, 17 radi espinós, 18 radis tous.
Francesc J. de Sostoa, a partir de fonts diverses.
La cintura escapular en els condrictis és sostinguda per un cartílag anomenat escàpulo-coracoides, constituït per una sola peça superior o escapular i per una altra d’inferior o coracoides. És en aquest cartílag on s’articulen els tres cartílags basals de l’aleta: el protopterígium, el mesopterígium i el metapterígium. Sobre aquests tres cartílags s’insereixen una sèrie de segments radials que donen suport als radis ceratòtrics o cartilaginosos.
En els peixos ossis, l’escàpula i el coracoides estan ossificats; la cintura és constituïda, bàsicament, per un os llarg anomenat cleitre o clèitrum, a més del supraclèitrum i del postclèitrum. En els teleostis primitius, les aletes pectorals ocupen una posició baixa, i per això disposen d’un os addicional, el mesocoracoide, que uneix el clèitrum amb el coracoides, com s’observa en els salmònids i els clupèids. Els ossos radials s’uneixen al marge ventral de l’escàpula; sobre d’aquests ossos s’insererixen els radis lepidòtrics, força característics dels peixos ossis.
La cintura pelviana és força més senzilla que l’escapular, pel que fa al seu disseny estructural. En els peixos cartilaginosos, aquesta cintura pelviana és constituïda pel cartílag pelvià, o isquiopúbic, on s’articulen els dos basals (protopterígium i metapterígium). A partir d’aquests, es connecten els segments radials i els radis ceratòtrics. La cintura pelviana dels osteïctis, en canvi, és formada per dos ossos plans, els basipterigis, als quals s’insereixen els radis lepidòtrics.
La musculatura i la locomoció dels peixos
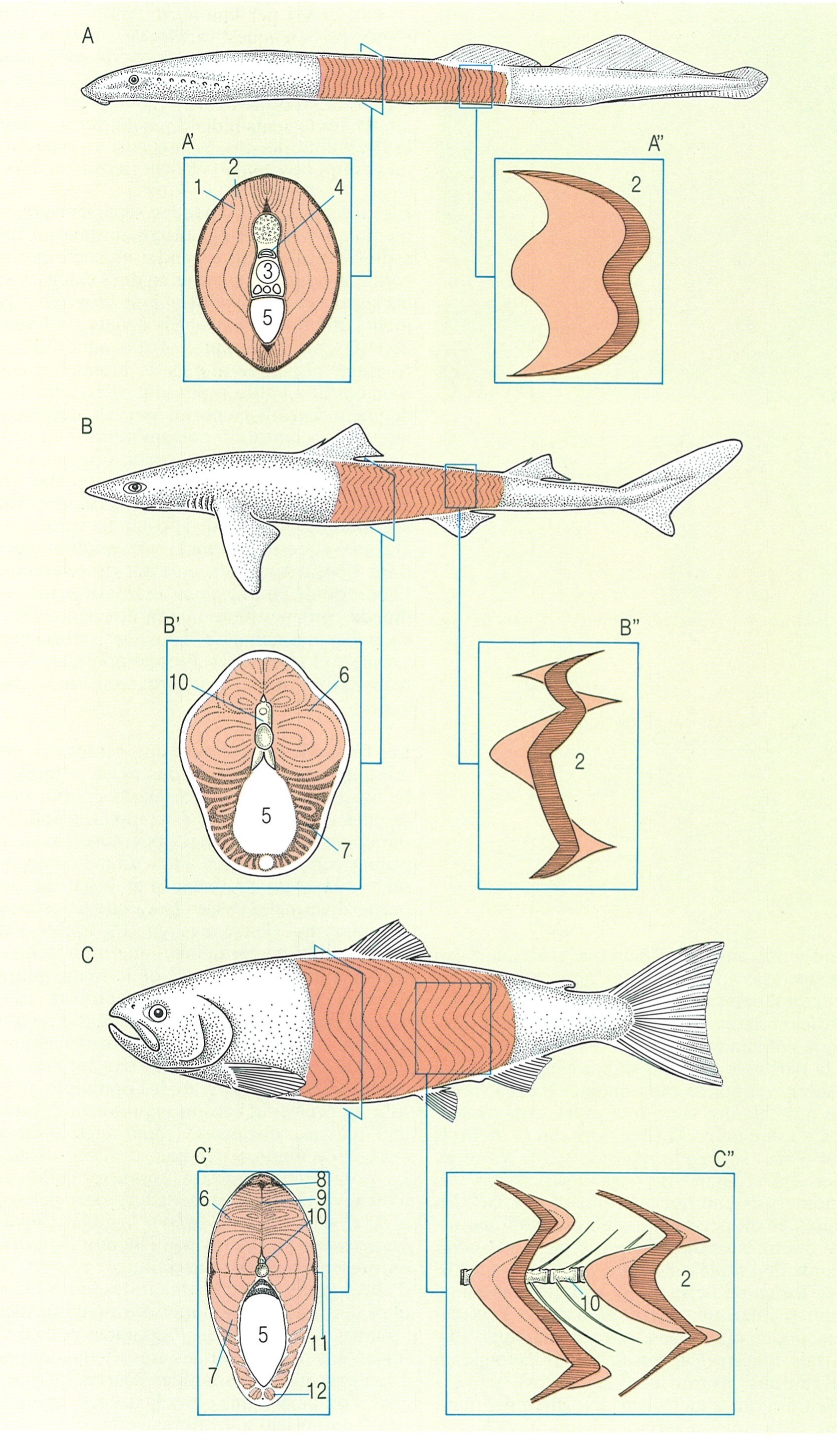
Musculatura del cos en els àgnats i els pisciformes. A llamprea (Petromyzon marinus), B agullat (Squalus) i C truita comuna (Salmo trutta). En cada cas s’ha representat una secció transversal del cos (A’, B’, C’) i un esquema de la forma del miòmer (A", B", C"). 1 miosepte, 2 miòtom, 3 notocordi, 4 canal neural, 5 cavitat del cos, 6 musculatura epiaxial, 7 musculatura hipoaxial, 8 musculatura supracarinal, 9 septe vertical, 10 vèrtebra, 11 musculatura vermella lateral, 12 musculatura infracarinal.
Amadeu Blasco, a partir de fonts diverses.
D’una manera general, hom pot dir que el sistema muscular dels peixos consta d’un conjunt de músculs llargs que recobreixen el cos i abasten des del cap fins al peduncle caudal. A més d’aquests, però, també cal considerar tota una sèrie complexa de músculs cranials relacionats amb les mandíbules, les brànquies i l’opercle. D’altra banda, els peixos també tenen una musculatura associada a les aletes parelles i imparelles que, en definitiva, són responsables del seu moviment.
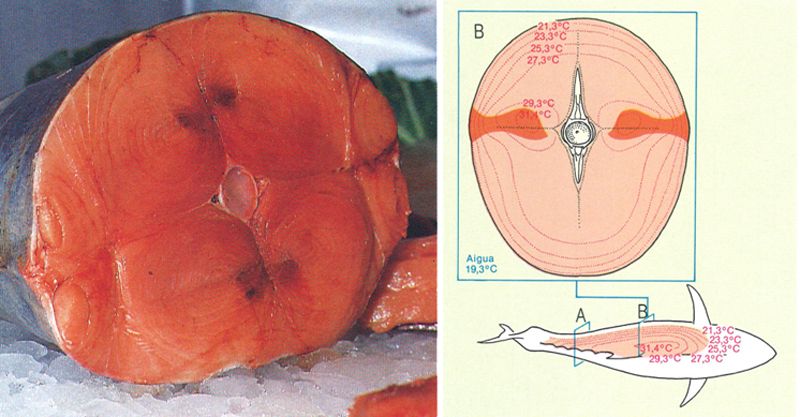
La proporció de musculatura vermella en el cos d’un peix és en relació amb la seva capacitat i velocitat de natació: és relativament abundant en els peixos grossos i grans nedadors, que avancen a una velocitat constant, i escassa en el peixos petits i capaços de fer moviments bruscs i curts. La fotografia, que correspon a la secció A del dibuix, ens mostra la carn de la tonyina, amb una alta proporció de musculatura blanca, que, de fet, en diverses espècies adopta una coloració rosada; la musculatura vermella, reduïda a no més d’un 18% de la massa muscular, apareix en aquesta secció posterior del cos en posició lateral, en forma d’unes petites taques de color roig fosc, a banda i banda de la columna vertebral. A baix, l’esquema representa una secció transversal (B) del cos de la tonyina. S’hi han indicat les temperatures del cos, que de vegades són superiors a les de l’ambient. La musculatura vermella s’ha representat de color més fosc.
Jordi Vidal; Amadeu Blasco.
La musculatura del tronc s’organitza en forma d’una sèrie de segments curts o miòmers. Els miòmers estan separats entre ells per unes parets transversals formades per teixit conjuntiu i per fibres de col·lagen; aquestes parets s’anomenen mioseptes. El conjunt d’aquesta estructura muscular no és sinó el fruit de la segmentació que experimenta la musculatura axial dels vertebrats.
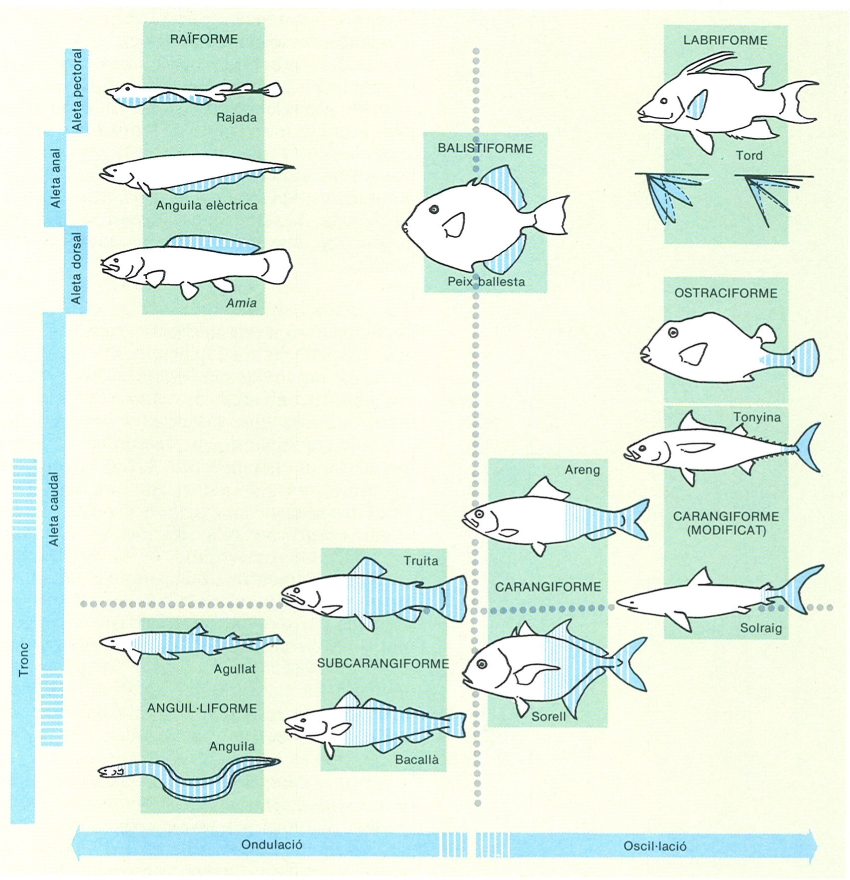
La locomoció en els peixos. A dalt, diferents tipus de locomoció en diferents espècies de condrictis i osteïctis: tipus raïforme, balistiforme, labriforme, ostraciforme, carangiforme, carangiforme modificat, subcarangiforme i anguil·liforme (els noms dels tipus no es refereixen als ordres); s’han ressaltat en el dibuix les aletes o parts del cos responsables del moviment de la natació. A baix, esquema de la manera com es torça el cos durant la locomoció (A) i els canvis que experimenten els miòmers de cada costat (A’). B locomoció dels peixos i forces que entren en joc durant la torsió lateral.
Román Montull, modificat a partir d’originals de Linsey (1978), Mc Farland et al (1979), Alexander (1967) i Walker (1979).
Les fibres musculars que conformen els miòmers es disposen d’una manera paral·lela a l’eix longitudinal del cos i s’insereixen entre cada parella de mioseptes. Això no obstant, cadascun dels miòmers se situa més o menys transversalment respecte a l’eix del cos i defineix uns angles oposats que s’orienten cap endavant i cap enrere; així, aquest disseny estructural dels miòmers recorda la lletra W. La posició relativa dels miòmers és diferent prop de la columna vertebral, on s’insereixen, que a la part més externa, sota la pell. D’aquesta manera, el pla de cada miòmer o cada secció de musculatura compresa entre dos mioseptes, s’estén entre 3 i 12 regions intervertebrals del tronc, segons la regió del cos i les espècies. En els àgnats la posició superficial dels miòmers és anterior a la zona d’inserció. En canvi, en els peixos ossis i els elasmobranquis, els miòtoms es disposen en espiga, a banda i banda de l’espina, és a dir, amb la característica forma de lletra W. D’aquesta manera, si hom realitza un tall transversal en l’eix del cos, poden observar-se diferents miòmers disposats concèntricament (com per exemple en els taurons, les tonyines i les truites); això explica el grau d’inclinació dels diferents plans que talla aquesta secció. Els miòmers s’insereixen en els centres de les vertebres, com també les seves espines hemals i neurals. Els mioseptes són incontràctils, però gaudeixen d’una certa capacitat de deformació; a més, aquestes estructures possibiliten els moviments laterals i també serveixen per a reduir la flexibilitat del cos, la qual cosa és especialment important en el cas dels grans nedadors.
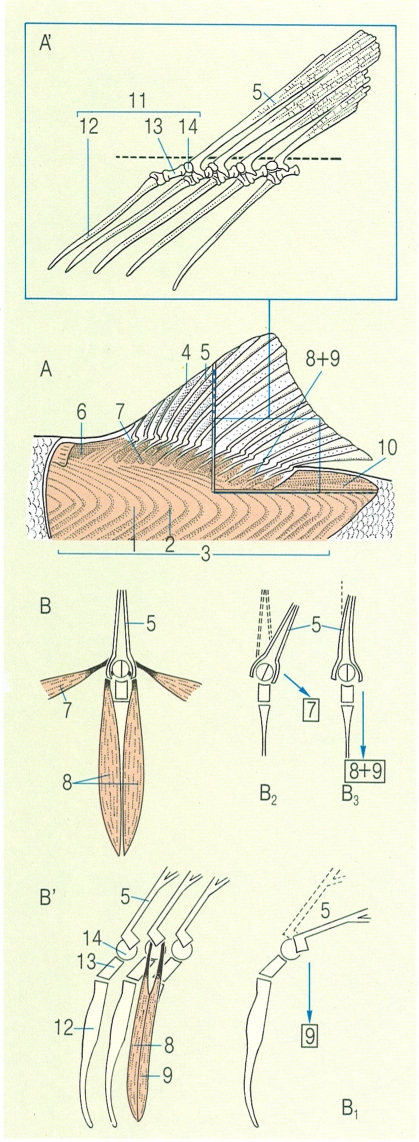
Musculatura superficial i profunda de l’aleta dorsal d’un peix (A) i detall dels pterigiòfors i els radis (A’) on s’insereixen els músculs principals. A baix, esquema dels músculs associats a cada radi (B en visió frontal i B’ en visió lateral); efectes dels músculs depressor (B1), i inclinador (B2) i efecte global dels músculs antagònics erector i depressor (B3). 1 miòmer, 2 miosepte, 3 musculatura epiaxial, 4 membrana de l’aleta, 5 radi segmentat o tou, 6 múscul protractor dorsal, 7 múscul inclinador dorsal, 8 múscul erector dorsal, 9 múscul depressor dorsal, 10 múscul retractor dorsal, 11 pterigiòfors o elements de suport de l’aleta dorsal, (12 proximal, 13 mitjà, 14 distal).
Amadeu Blasco, a partir de fonts diverses.
Els músculs del tronc queden separats en dues meitats a cada costat del cos mitjançant un septe vertical. D’altra banda, un septe horitzontal divideix els músculs en dues categories: els epaxials a la part dorsal i els hipaxials a la meitat inferior del cos. Els àgnats no tenen aquest septe horitzontal. Totes aquestes estructures permeten al peix de brandar el cos d’un costat a l’altre i, per tant, d’adquirir l’adequada força locomotriu per als desplaçaments. La contracció dels miòtoms d’un dels flancs fa que el cos es corbi vers un dels costats; consegüentment, cap i cua s’atansen gràcies a l’articulació de les vèrtebres, tot aprofitant la rigidesa de la columna vertebral. Durant aquesta operació els miòtoms del flanc oposat romanen en estat de relaxació. Una sèrie de contraccions alternatives i coordinades originen una ona en direcció cèfalocaudal que, fonamentada en la resistència que exerceix el medi en la direcció oposada, és la responsable que el peix progressi cap endavant.
Les fibres de musculatura que conformen els miòtoms poden ésser de dos tipus: vermelles (o fosques) i blanques. Aquesta divisió, que té unes connotacions més complexes que les purament morfològiques, pot observar-se a primer cop d’ull si es realitza un tall transversal per la regió postanal: la zona lateral superficial vermella és més fosca que la resta de les fibres que l’acompanyen. La forma d’aquestes fibres vermelles, que representen d’un 5 a un 18% del total de la musculatura del peix, respon a una funció aeròbica; és a dir, gaudeixen d’un elevat contingut d’hemoglobina, a més d’ésser profusament vascularitzades, i per això necessiten oxigen. Aquest model de fibres vermelles és l’òptim per a assolir una velocitat constant i prolongada, amb una màxima eficiència i amb una despesa energètica mínima per part de l’organisme. La proporció d’aquestes fibres vermelles és indicativa dels hàbits nedadors dels peixos; així, el salmó (Salmo salar), l’agullat (Squalus acanthias) o la tonyina (Thunnus thynnus) en presenten elevats percentatges respecte a d’altres grups. Contràriament, les fibres blanques dels miòtoms són menys vascularitzades i pobres en mitocondris. Preparades per a una funció anaeròbica, són les més adequades per a realitzar natacions ràpides i curtes, però en canvi comporten una elevada despesa energètica, en oposició a les fibres vermelles.
En alguns peixos, com ara els escòmbrids (les tonyines, entre d’altres), la temperatura corporal supera la del medi on viuen; les fibres vermelles són les responsables d’aquest fenomen per l’efecte de la contracció. Molts d’aquests peixos també utilitzen un mecanisme de contracorrent circulatori força enginyós basat en el fet que la sang calenta, provinent de les fibres vermelles, passa a través d’una xarxa de capil·lars propera als vasos que transporten sang freda i oxigenada; d’aquesta manera l’intercanvi gasós queda afavorit per la diferència tèrmica i es pot mantenir una temperatura constant en la zona corresponent. Aquests dos sistemes de fibres, vermelles i blanques, funcionen independentment tant en el cas dels àgnats com en el dels condrictis, i en el dels peixos ossis primitius; en tots aquests grups ambdós sistemes són utilitzats d’una manera alternativa. Amb tot, sembla que els peixos ossis evolucionats poden utilitzar-los indistintament tot tenint en compte les limitacions específiques de cada grup; en conseqüència, la fibra blanca també serveix per a practicar una natació continuada o de travessia.
Els peixos han adoptat diversos mètodes de locomoció; així, els diferents grups utilitzen, d’acord amb llurs adaptacions, les aletes parelles o imparelles per a produir moviments ondulatoris, aletejar o remar. Pel que fa, però, a la immensa majoria, la propulsió resulta de combinar dos processos bàsics: la generació d’ones transversals a través del cos i els moviments de l’aleta caudal. Quant a les ones transversals, s’originen a partir de les contraccions laterals del cos. En alguns peixos, com els escòmbrids (tonyines), la propulsió es produeix principalment pel moviment de l’aleta caudal; en aquest cas les flexions del cos són molt escasses. Això explica que l’aleta caudal d’aquests peixos presenti un disseny hidrodinàmic força característic de forma semilunar i amb un peduncle caudal estret però potent, per a poder realitzar cap a 10 flexions per segon.
Els peixos caracteritzats per una natació lenta presenten unes aletes pectorals desenvolupades i llargues per tal de reduir l’efecte de resistència de l’aigua; en canvi, els peixos de natació ràpida les tenen curtes i estretes amb la finalitat d’evitar turbulències. En aquest sentit també són importants la posició de les aletes parelles i l’existència de determinats mecanismes especials, com ara la secretació de certes substàncies, amb la missió de reduir la resistència a l’aigua, com s’observa al peix llima (Ruvettus pretiosus).
La velocitat que assoleix la natació dels peixos varia segons la mida de l’animal i, malgrat tot, hom estima que els més ràpids (com ara les tonyines, que fan 1 m de llargada aproximadament) poden atènyer velocitats de fins a 42 km/h. Hi ha una sèrie de músculs destinats a moure les aletes, com ara els supracardinals, que connecten amb els elements de suport dels radis o pterigiòfors de l’aleta dorsal, tant a l’extrem anterior com al posterior. Els anomenats infracardinals, són responsables del moviment de l’aleta anal. Aquests músculs actuen, respectivament, com a protractors i retractors de les aletes imparelles. Els radis de les aletes, a més, són proveïts de músculs inclinadors capaços de produir el balandreig d’aquests elements esquelètics. Les aletes parelles presenten músculs adductors i arectors connectats individualment amb cada radi. Els moviments dels radis de l’aleta dorsal produeixen una ondulació que desencadena un efecte propulsor. Amb tot, les aletes pectorals també poden ésser emprades per a avançar, a més d’ésser utilitzades per a realitzar moviments verticals o per a frenar la natació.
Els sistemes de relació i control dels peixos
El sistema nerviós central
El sistema nerviós assoleix el màxim grau de complexitat en els vertebrats, tot i que a títol comparatiu el dels àgnats resulta més senzill que no el dels pisciformes. Tradicionalment hom el divideix en un sistema nerviós central i un sistema nerviós perifèric. El central és format per l’encèfal i per la medul·la espinal; és el responsable de les funcions d’integració i de coordinació. El sistema nerviós perifèric, en canvi, consta, de nervis i de ganglis que connecten amb l’encèfal i amb la medul·la espinal.
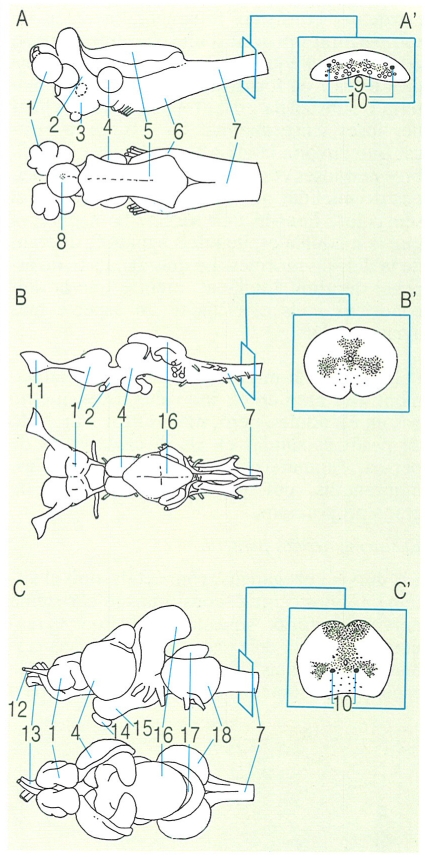
Esquema de l’estructura de l’encèfal en els àgnats (A), en el cas concret de la llamprea (Petromyzon marinus); en els elasmobranquis (B), en el cas de l’agullat (Squalus); i en els teleostis (C), en el cas de la carpa (Cyprinus carpió). S’han dibuixat també les respectives seccions transversals a nivell de la medul·la (A’ B’ i C’). 1 hemisferi cerebral, 2 tàlem, 3 hipotàlem, 4 lòbul òptic, 5 plexe coroïdal, 6 bulb raquidi, 7 medul·la oblonga, 8 òrgan pineal, 9 axons de Muller, 10 axons de Mauthner, 11 bulb olfactori, 12 tracte olfactori, 13 nervi òptic (II), 14 pituïtària, 15 lòbul inferior, 16 cerebel, 17 lòbul facial, 18 lòbul del vague.
Francesc J. de Sostoa, a partir de fonts diverses.
Durant el procés de desenvolupament dels peixos la part anterior del tub neural s’engruixeix i origina l’encèfal o cervell, en sentit ampli; la resta d’aquest tub neural acaba per constituir la medul·la espinal. A la part anterior es produeixen dues constriccions que delimiten tres engruiximents o vesícules: el cervell anterior o prosencèfal, el cervell mitjà o mesencèfal i el cervell posterior o romboencèfal.
El cervell anterior o prosencèfal es divideix en el telencèfal i el diencèfal; el telencèfal és parell i constitueix els anomenats hemisferis cerebrals a la part anterior dels quals apareixen els lòbuls olfactoris o rinencèfal. Aquests lòbuls olfactoris s’uneixen als bulbs olfactoris per mitjà de fibres nervioses i d’aquí parteixen els nervis olfactoris que s’estenen fins a l’òrgan olfactiu. Cal esmentar que aquests lòbuls olfactoris assoleixen unes dimensions considerables en els àgnats i que semblantment es troben força desenvolupats en els elasmobranquis, però que en canvi són de mida reduïda en la majoria dels peixos ossis. El diencèfal, l’altra part del cervell anterior, s’origina per una constricció del telencèfal i forma els òrgans parapineal i pineal dels àgnats, com també l’òrgan pineal dels pisciformes, sensible als canvis lluminosos. La part ventral del cervell anterior (prosencèfal) constitueix l’hipotàlem, que integra les funcions del sistema autònom perifèric. A la part superior del mesencèfal (cervell mitjà) s’origina un parell d’estructures prominents, els lòbuls òptics, la funció bàsica dels quals constitueix el sentit de la vista. Aquest mesencèfal també pot complir unes altres funcions, com ara les relacionades amb el sistema sensorial (o acusticolateral). Finalment, el romboencèfal o cervell posterior és format pel cerebel i per la medul·la oblonga, relacionada amb al medul·la espinal. El cerebel dels àgnats és poc desenvolupat (llamprees) i fins pot no haver-n’hi (mixínids). Tanmateix en els pisciformes és ben constituït tot i haver-hi marcades diferències entre els diversos grups. La funció primordial del cerebel se centra en la coordinació dels moviments, en el manteniment de l’equilibri i del to muscular. Amb tot, sembla que el cerebel també es relaciona amb la percepció dels impulsos elèctrics generats per alguns peixos com els mormírids africans. D’altra banda, la medul·la oblonga rep els impulsos motors de gran part dels nervis cranials (del nervi V al X).
La medul·la espinal és contínua, des de la medul·la oblonga fins a la regió caudal. Aquesta estructura consisteix fonamentalment en un tub perforat i proveït d’un solc central estret. Al voltant d’aquest solc s’observa una zona més fosca i de forma peculiar, que és la matèria o substància grisa, força rica en cèl·lules nervioses sense mielina. La substància blanca, que envolta la grisa, és constituïda per fibres nervioses conductores i proveïdes de beines de mielina, que són les responsables del seu color. Fent-hi una secció, hom observa que la medul·la espinal dels àgnats és diferent de la dels pisciformes. La dels àgnats té un aspecte deprimit i aplanat, mentre que la dels condrictis i els osteïctis té un aspecte més aviat circular.
Pel que fa a la mida de l’encèfal, és relativament més gran en els individus joves que no pas en els adults, però, malgrat tot, la relació de pesos és similar en els diversos grups de peixos. Tanmateix, cal exceptuar-ne els elasmobranquis, que presenten un encèfal de grans proporcions.
El sistema nerviós perifèric
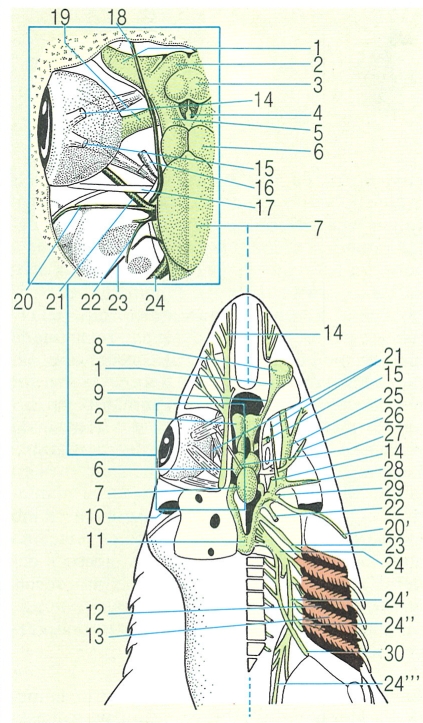
Sistema nerviós perifèric en els peixos, concretament en els elasmobranquis (l’ampliació de la zona pròxima al cervell és feta segons un pla diferent del que mostra la figura general). S’indiquen les parts: 1 bulb olfactorí, 2 hemisferi cerebral, 3 làmina terminal de l’encèfal, 4 epífisi, 5 tàlem, 6 lòbul òptic, 7 cerebel, 8 sac olfactori, 9 tracte olfactori, 10 espiracle, 11 medul·la oblonga, 12 cambra branquial i 13brànquia; els músculs: 14 oblic superior, 15 recte superior, 16 recte anterior i 17 recte posterior; i els nervis cranials següents: 18 oftàlmic superficial (V+VII), 19 òptic (II), 20 facial (VII) i branca hiomandibular (20’), 21 oftàlmic (V), 22 estatoacústic (VIII), 23 glosso-faringi (IX), 24 vague (X) i branques visceral (24’), branquial (24") intestinal (24"), 25 infraorbital (V+VII), 26 oculomotor (III), 27 troclear (IV), 28 mandibular (V), 29 palatí (V), 30 hipobranquial.
Amadeu Blasco, a partir de fonts diverses.
Els nervis i els ganglis, connectats amb el sistema nerviós central, constitueixen el sistema nerviós perifèric. Aquestes connexions es realitzen per mitjà dels nervis espinals i dels cranials; els espinals s’enllacen amb la medul·la espinal i els cranials ho fan amb l’encèfal i amb la medul·la espinal oblonga. Gran part dels nervis contenen dos tipus de fibres diferents: les unes s’encarreguen d’enviar missatges vers les distintes parts del cos i les altres els reben i alhora els canalitzen cap al sistema nerviós central. Les primeres, que consegüentment envien els impulsos a la musculatura, a les diverses glàndules i als òrgans, constitueixen les denominades fibres motores. Les altres són les fibres sensorials, que reben dels òrgans sensorials els esmentats impulsos per a transferir-los al cervell. Cadascun dels nervis espinals, que són parells, presenta una rel dorsal i una rel ventral que queden unides a la medul·la espinal, excepte a les llamprees. Molt a prop del punt de fusió d’ambdues rels apareixen tres branques: la dorsal, la ventral i la visceral. Les dues primeres innerven la pell i la musculatura axial; en canvi la branca visceral és la que en definitiva innerva els diferents òrgans.
Cal esmentar que els nervis cranials són força més especialitzats que els espinals; formen un conjunt d’onze nervis que connecten el cervell amb alguns òrgans sensorials i músculs, però, amb tot, gran part d’aquests nervis innerven regions del cap. Els nervis 0 i I, és a dir, el terminal i l’olfactiu, són associats al sentit de l’olfacte. El segon nervi (II), l’òptic (tracte òptic), és el responsable de transportar els impulsos visuals. En el cas dels àgnats cada nervi connecta directament amb l’ull corresponent de cada banda; en canvi, en els elasmobranquis són fusionats tot formant un quiasma òptic i en els peixos ossis es creuen per sota el cervell de manera que el que procedeix del lòbul dret innerva l’ull esquerre i viceversa. Els nervis oculomotor (III), troclear (IV) i abducent (VI), són motors. El trigaran (V) i el facial (VII) són nervis mixtos, amb una part de fibres sensorials i una altra de fibres motores. L’auditiu (VIII) és de tipus sensitiu i innerva l’oïda. El glosso-faringi (IX) és mixt i innerva la primera fenedura branquial. El vague (X) és un nervi mixt que té diverses branques i fa moltes funcions, com ara la de connectar amb el sistema de la línia lateral, a més d’innervar els òrgans interns i les brànquies.
Els òrgans dels sentits: l’ull i la vista
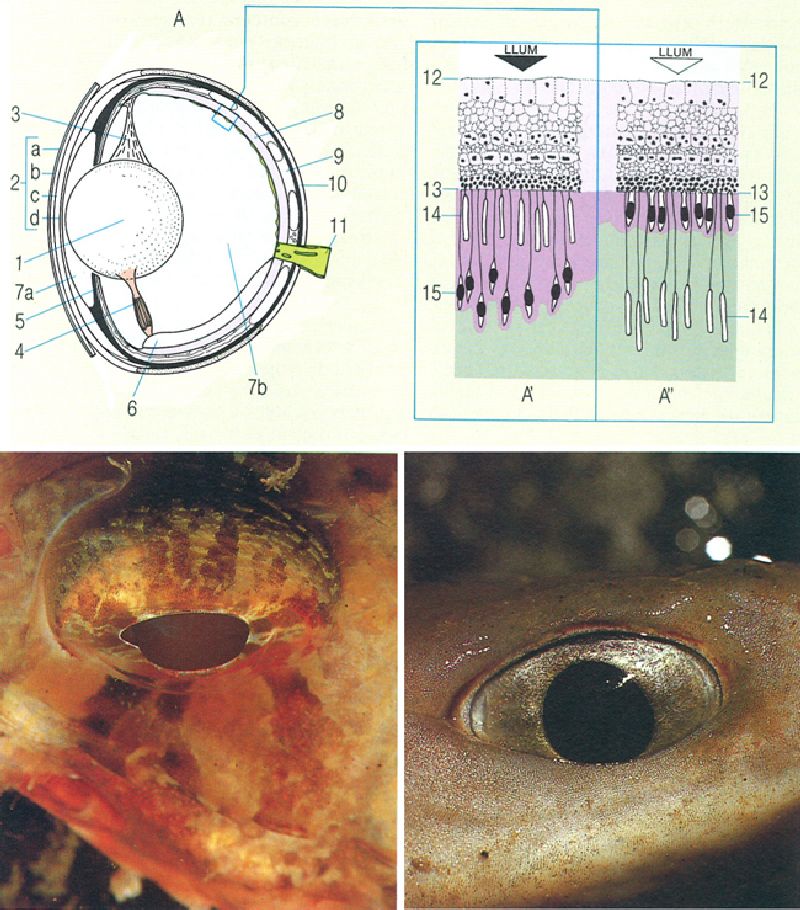
Estructura de l’ull dels peixos (A) i detall de la retina dels teleostis en el cas d’adaptació a la fosca (A’) i a la llum (A"). 1 lent o cristal·lí, 2 còrnia (a capa conjuntiva, b capa dèrmica, c capa escleròtica, d capa subescleròtica), 3 lligament suspensor de la lent, 4 múscul retractor de la lent, 5 iris o pupil·la, 6 procés falciforme, 7a cambra de l’humor aquós, 7b cambra de l’humor vitri, 8 retina, 9 coroide, 10 escleròtica, 11 nervi òptic, 12 membrana limitant interna, 13 membrana limitant externa, 14 bastons, 15 cons. A baix, les fotografies mostren, a la dreta, l’ull el·lipsoïdal d’un tauró (condricti), que es caracteritza per les grans dimensions del cristal·lí i l’escassa mobilitat del globus ocular; i, a l’esquerra, l’ull circular i notablement mòbil d’un peix ossi; l’angle superior des del qual s’ha fet aquesta foto permet de notar la posició del cristal·lí (de color fosc en la fotografia), que es projecta enfora de l’iris.
Francesc J. de Sostoa, a partir de fonts diverses; Xavier Safont/M. Alba Camprubí i Juan A. Moreno.
El fotoceptor primari de la visió dels peixos és, naturalment, l’ull, que presenta un gran nombre de modificacions; tanmateix, pot dir-se que l’estructura bàsica primordial és francament similar a la dels diferents grups de peixos.
A la part interior de l’ull hi ha la còrnia, formada per diverses capes de cèl·lules transparents (conjuntiva, dermal, escleròtica i subescleròtica). A continuació de la còrnia hom troba una cavitat o cambra anterior, plena d’un líquid transparent anomenat humor aquós. Aquesta cambra, per mitjà de l’iris o pupil·la, es comunica amb la cambra posterior. El cristal·lí és una estructura esfèrica que es troba al centre de l’iris i el travessa; en general se sosté per mitjà del lligament suspensor i pot desplaçar-se gràcies al múscul retractor, bé que hi ha diferències entre els diversos grups de peixos. Darrere el cristal·lí hi ha una cambra proveïda d’una substància semitransparent, l’humor vitri. A la paret interna del globus ocular hi ha la retina, que limita posteriorment la cambra de l’humor vitri. La retina és constituïda per dos tipus de cèl·lules diferents: els cons i els bastons. Aquestes cèl·lules són allargades i es disposen en tota la superfície amb els extrems dirigits vers la llum. Aquestes dues modalitats cel·lulars contenen pigments capaços d’absorbir la llum de la imatge enfocada; seguidament la informació és enviada al cervell per mitjà del nervi òptic. Aquest nervi surt de la part posterior de l’ull i travessa la retina, la coroide i l’escleròtica, que completen el globus ocular.
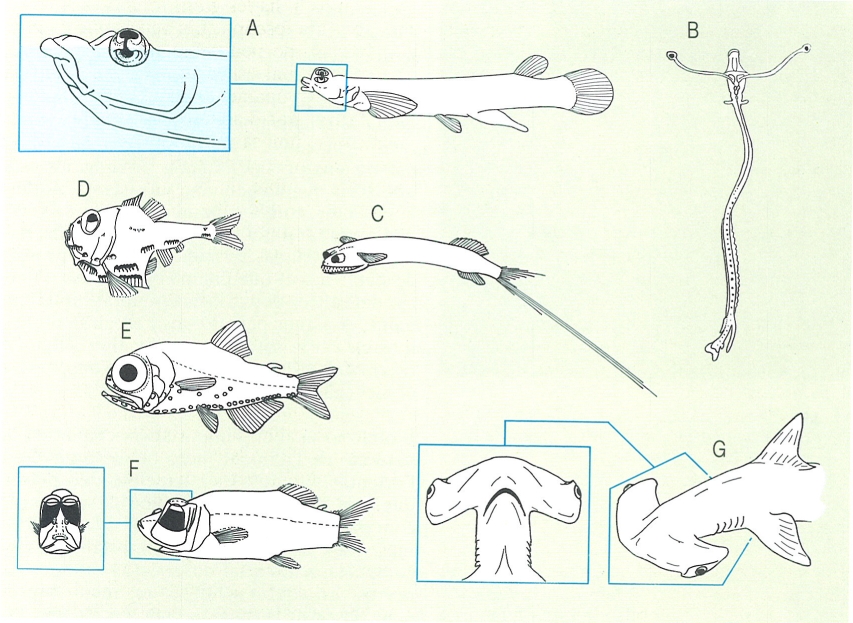
El tipus d’ulls que presenten els peixos és en relació amb els seus hàbitats. En general, els que viuen a les grans fondàries, on no arriba la claror, els tenen molt desenvolupats, a més de tenir un nombre molt elevat de bastons o de pigments fotosensibles (E, corresponent a Electrona rissoi); un cas curiós són les larves d’ldiacanthus fasciola (B), que els tenen situats a l’extrem d’un llarg peduncle. Els que viuen a la zona mesopelàgica acostumen a tenir-los tubulars i situats en unes protuberàncies, sovint mòbils, que poden orientar per a augmentar la capacitat de detecció de preses; s’han representat en els exemples C (Gigantura chuni), D (Argyropelecus) i F (Opisthoproctus). Hi ha també el cas (A), ben curiós, dels anomenats quatre-ulls (Anableps tetrophthalmus), que neden per la superfície de l’aigua i tenen la retina dividida en dues parts, de manera que tenen alhora visió aèria i aquàtica. El peix martell (G Sphyrna zygaena) té els ulls situats als extrems de les expansions laterals del cap.
Francesc J. de Sostoa, a partir de fonts diverses.
Entre els àgnats, l’ull dels mixínids és sensiblement diferent del de les llamprees. Els mixínids tenen uns ulls atrofiats i ni de bon tros funcionals; els de les llamprees, en canvi, són primitius però ben constituïts. L’ull de les llamprees no té parpella, i la còrnia no és fusionada a la pell; com que hi manca el lligament suspensor, la regulació del cristal·lí solament es realitza per la pressió de l’humor vitri. El sistema d’enfocament, força especial en aquests vertebrats, s’efectua per mitjà d’un múscul esfèric que en contraure’s deprimeix la còrnia; consegüentment, la pressió interna fa que el cristal·lí es desplaci cap enrere i possibiliti la localització d’objectes llunyans.
Els ulls dels elasmobranquis són de dimensions considerables. Les parpelles són simples membranes d’escassa mobilitat, les anomenades membranes nictitants. El globus ocular és el·lipsoïdal i, en canvi, el cristal·lí és esfèric i molt gros. La part posterior de la retina conté una capa de guanina que reflecteix la llum (tapetum lucidum), i per això els ulls dels elasmobranquis brillen quan són il·luminats; sembla que la missió d’aquesta capa és la d’incrementar la sensibilitat. El sistema d’enfocament consisteix en la contracció del múscul protractor, ja que aquests peixos també manquen de múscul retractor del cristal·lí.
D’altra banda, els peixos ossis estan mancats de parpelles. En aquest sentit, les úniques estructures que s’hi assemblen són les membranes adiposes d’alguns peixos nedadors, com ara la tonyina (Thunnus thynnus) i l’areng (Clupea harengus); això no obstant, aquestes membranes són immòbils i fan una funció exclusivament hidrodinàmica. El moviment del cristal·lí durant l’enfocament, en aquest cas, es realitza mitjançant el múscul retractor; pel que fa a la resta de l’estructura, pot considerar-se força similar a la dels elasmobranquis.
Els peixos estan adaptats a tota una gamma d’ambients lumínics molt diferents. Dins les aigües tèrboles d’alguns rius i llacs la visibilitat es limita a un o dos metres de distància; en canvi, les aigües litorals marines gaudeixen d’una transparència que permet la visibilitat fins a 30 m. A la mar, la llum solar penetra fins a 100 m quan les aigües són transparents, però des d’aquesta profunditat fins a 1100 m hi ha una zona de penombra, encara que a partir de 750 m la llum és pràcticament nul·la; més al fons hi ha les grans fondàries abissals, en l’absoluta foscúria. Els peixos que colonitzen la zona de penombra han de tenir una adaptació visual especial; així, presenten un gran desenvolupament dels ulls i del cristal·lí, amb la finalitat d’adquirir una superfície de captació lluminosa més gran. Altrament, en l’ull d’aquests peixos hom observa un augment del nombre de bastons o de pigments fotosensibles, que els fa més eficients, especialment pel que fa a l’espectre del blau. En d’altres casos les modificacions dels ulls condueixen a noves estructures més especialitzades, com ara els ulls tubulars d’alguns peixos.
Els pigments fotosensibles dels bastons i dels cons de la retina són ben diferents segons que es tracti de peixos marins (rodopsines) o d’aigua dolça (porfiropsines). Si bé els bastons són més sensibles a la baixa intensitat lluminosa, la funció dels cons és més relacionada amb els colors i, per això, només són fotosensibles en cas d’elevada il·luminació. Això explica que les proporcions entre cons i bastons siguin extraordinàriament variables entre les espècies diürnes i les nocturnes o les que viuen en zones de penombra. Naturalment, el nombre de bastons dels peixos d’hàbits nocturns és força elevat, perquè han de captar amb la màxima eficiència l’escassa llum ambiental.
És ben sabut que els peixos gaudeixen d’una bona visió. D’una banda, perquè l’índex de refracció de la còrnia és similar al de l’aigua. Altrament, el fet de tenir un cristal·lí esfèric redueix al màxim les aberracions visuals. A més, la posició lateral dels ulls, juntament amb d’altres característiques estructurals, els permet de tenir un angle de visió força més ampli i de tipus circular.
Algunes de les espècies que colonitzen la zona mesopelàgica (entre 200 m i 1000 m de profunditat), especialment els que viuen entre els 500 i els 700 m, presenten uns ulls de tipus tubular. Aquests ulls experimenten un allargament de la pupil·la que els confereix una morfologia de tub curt, semblants a uns binocles orientats cap amunt o cap endavant, segons les espècies. Aquesta modalitat d’ulls ofereix alguns avantatges, com ara el fet d’augmentar la capacitat de detecció de les preses en un ambient on els organismes utilitzen la tècnica del contrallum per tal de camuflar-se. Cal esmentar que en gran part dels casos es combinen diverses adaptacions, com ara una concentració més gran de rodopsina o l’existència d’una lent auxiliar que els permet una determinada visió lateral.
Hi ha unes espècies mexicanes, com l’Anableps, que tenen una retina dividida en dues parts adaptades a proporcionar simultàniament una visió aèria i aquàtica; d’aquí que se’ls anomeni «quatre-ulls». Tenint en compte la varietat de colors que ofereixen la majoria de peixos litorals, no és estrany de pensar que aquests peixos, a més dels taurons, gaudeixen d’una visió en color. De fet s’hi ha comprovat l’existència de tres models de cons diferents, especialment dissenyats per a aquest tipus de visió multicromàtica.
Tot i ésser escassos, també hi ha peixos d’aigua dolça cavernícoles o habituats a viure als rius i als llacs subterranis. Aquets peixos solen presentar uns ulls completament atrofiats que s’emplacen sota la pell i que manquen de funcionalitat. Ultra això, també n’hi ha que conserven uns ulls vestigials proveïts de nervis òptics rudimentaris i que, en conseqüència, són sensibles als canvis d’intensitat lluminosa. Cal afegir que, en el cas dels peixos cavernícoles, la detecció de l’aliment i, per tant, la subsistència queda assegurada pel desenvolupament considerable que experimenten els altres òrgans sensorials.
Els òrgans dels sentits: l’olfacte i el gust
Els òrgans olfactoris se situen a l’interior d’unes estructures en forma de sac que s’emplacen a la part anterior del cap, davant els ulls. A diferència del que hom observa en els altres vertebrats superiors, els sacs nasals gairebé mai no es comuniquen amb la faringe, fora dels mixínids, dels peixos pulmonats i els uranoscòpids. Així, és freqüent de trobar dos orificis externs a cada banda del cos, que connecten aquests sacs olfactoris cecs amb l’exterior. Per tal que l’animal obtingui una resposta als estímuls olfactoris, l’aigua, carregada de tot un seguit de substàncies químiques dissoltes, ha de circular en contacte amb l’epiteli olfactori (roseta), on hi ha els quimioceptors. L’aigua circula a través del sac olfactori per diversos mecanismes: en alguns peixos, per la presència dels cilis que recobreixen la cavitat de l’òrgan i que, amb llur moviment, activen la circulació; en d’altres, la circulació és deguda als moviments respiratoris de l’animal, que funcionen com una bomba activa; finalment, hi ha peixos que l’aconsegueixen per mitjà de la simple natació.
Els àgnats tenen un únic orifici nasal, situat a mig musell. El seu sac olfactori és força característic, proveït de nombrosos plecs i recobreixen d’epiteli olfactori, estructura que és envoltada al seu torn per una càpsula olfactòria de tipus cartilaginós. Del sac olfactori parteix un tub llarg que discorre en direcció ventral, anomenat conducte naso-faringi. En el cas de les llamprees aquest tub és tancat a l’extrem, tot i que en els mixínids es comunica amb la faringe. Per tal d’impulsar l’aigua les llamprees es valen de la musculatura branquial, que és la responsable de produir la pressió necessària per tal que l’aigua entri i surti del conducte naso-faringi i, finalment, s’introdueixi en l’òrgan olfactori.
Els òrgans olfactoris dels pisciformes són parells i, a més, presenten altres diferències respecte als dels àgnats. Les rajades i els taurons acostumen a exhibir apèndixs cutanis i estries que projecten externament els orificis nasals a fi de facilitar l’entrada d’aigua durant la natació. D’altra banda, entre els peixos ossis hi ha diversitat d’estructures dels òrgans olfactoris, però gran part de les espècies presenta dos orificis nasals a cada banda, l’anterior destinat a l’entrada d’aigua i el posterior, a la seva expulsió. Determinats peixos, com ara les anguiles, tenen uns orificis d’entrada tubiformes força separats dels posteriors i es valen dels cilis per fer-hi circular l’aigua. L’espinós (Gasterosteus aculeatus) disposa d’un únic orifici nasal per banda, que s’omple i es buida alternativament amb la mateixa respiració. En canvi, l’areng (Clupea harengus) mostra uns òrgans limfàtics pròxims als sacs olfactoris que actuen com un sistema hidràulic; és a dir, com una bomba de pressió. Tampoc no és rar el fet de trobar un sol orifici nasal a cada banda, de mida més gran i proveït de paret nasal, que compleix la mateixa funció dels altres dos orificis esmentats anteriorment. A alguns peixos ossis i als elasmobranquis els cal la natació per a fer fluir l’aigua a través dels sàculs olfactoris, com s’observa en el luci (Esox lucius).
L’olfacció és d’una importància crucial en la detecció de l’aliment, però també intervé en l’orientació durant les migracions dels peixos que han de reconèixer el lloc de naixença, com passa amb el salmó (Salmo salar). Endemés, juga un paper força important dins el comportament social de diverses espècies de peixos. En aquest sentit, és realment sorprenent observar la manera com, en algunes espècies, per exemple el «guppy» (Lebistes), la simple presència de determinades substàncies secretades pels mascles o per les femelles pot ésser captada per individus del sexe contrari i com això és capaç de desencadenar el comportament reproductor. Semblantment, el barb roig (Phoxinus phoxinus), quan és agredit, secreta un seguit de substàncies encarregades de donar l’alarma a la resta de la mola (feromones). Per aquest mateix procediment un determinat nombre de ciprínids, com també altres peixos d’aigua dolça, són capaços de detectar la presència d’unes altres espècies.
Són responsables del sentit del gust un altre tipus de quimioceptors: els botons gustatius. Aquestes estructures presenten una forma de nieló característica. Es troben sota la pell, al fons d’una petita depressió o porus gustatiu, recoberta de pèls gustatius. Els receptors gustatius poden ésser situats a diferents parts del cos, com a la cavitat bucal, la pell, les aletes i les barbes sensorials. Els receptors gustatius dels àgnats es localitzen a la regió faríngia i a la superfície cefàlica; els dels elasmobranquis, a la boca i la faringe. En canvi, en els peixos ossis tenen un emplaçament més ampli que abasta les diferents regions corporals, tot i que dominen especialment al cap, a la boca, als llavis (salmònids, cobítids, i ciprínids) i també a les barbes sensorials (ciprínids) o a l’extrem dels radis llargs. Alguns (Ictalurus) d’aigua dolça constitueixen un cas especial, perquè, desproveïts d’escates, presenten els receptors gustatius repartits per tota la pell del cos.
Els òrgans dels sentits: el sistema acusticolateral: l’oïda interna i la línia lateral
L’oïda interna dels peixos actua alhora com a òrgan de l’equilibri i com a òrgan acústic. En el dibuix (a baix, a la dreta) s’ha representat el laberint membranós d’una llamprea (A), un elasmobranqui (B), un salmònid (C) i un ciprínid (D), i una secció d’una màcula (E), on es mostren les cèl·lules sensorials que formen una capa per sota dels otòlits (representats en groc). A baix, a l’esquerra podem veure, en una fotografia feta al microscopi electrònic, un tall transversal d’un otòlit (sageta) de llobarro (Dicentrarchus labrax) que permet de veure els microcristalls d’aragonita disposats radialment, i les discontinuïtats causades per les variacions del creixement, que apareixen fent solcs concèntrics. A dalt, otòlit de lluç (Merluccius merluccius), on s’observen diversos anells de creixement formats amb periodicitat estacional i disposats al voltant del nucli, opac, format en el període juvenil. 1 creu, 2 canal vertical anterior, 3 canal vertical posterior, 4 ampul·la, 5 sac ciliat, 6 utricle, 7 sàcul, 8 canal horitzontal, 9 lagena, 10 lapil, 11 sagita, 12 asterisc, 13 cèl·lules sensorials.
Francesc J. de Sostoa; J. M. Fortuño / J. Biosca / Institut de Ciències del Mar.
La denominació de sistema acusticolateral al·ludeix a la múltiple funcionalitat d’aquest sistema sensorial bàsicament integrat per l’oïda interna i per la línia lateral, a més d’altres òrgans més especialitzats. La funció global del sistema consisteix a captar tota mena d’ones (acústiques i de pressió), i també a actuar com a òrgan de l’equilibri i de l’acceleració angular. L’oïda interna, l’única que tenen els peixos, és la responsable de mantenir l’equilibri, d’informar-los sobre el grau d’acceleració que assoleixen i de la percepció del so. D’altra banda, el complex de la línia lateral és un òrgan, específic dels peixos i dels amfibis de vida aquàtica o en fase larval, que s’encarrega de detectar les ones de pressió que arriben al cos. Aquesta, però no és pas l’única funció de la línia lateral, car també es relaciona amb la percepció tèrmica. A més a més, alguns peixos van proveïts d’electroceptors capaços de detectar camps elèctrics o bé impulsos elèctrics emesos per altres peixos.
L’oïda interna és formada per un conjunt d’ossos i per una estructura força delicada i complexa, anomenada laberint membranós, que consta d’una sèrie de tubs i de cavitats recobertes d’epiteli sensorial i és integrada per dues cambres o eixamplaments, l’utricle, o cambra superior, i el sàcul, o cambra inferior, que en el cas dels peixos se subdivideix tot formant la lagena.
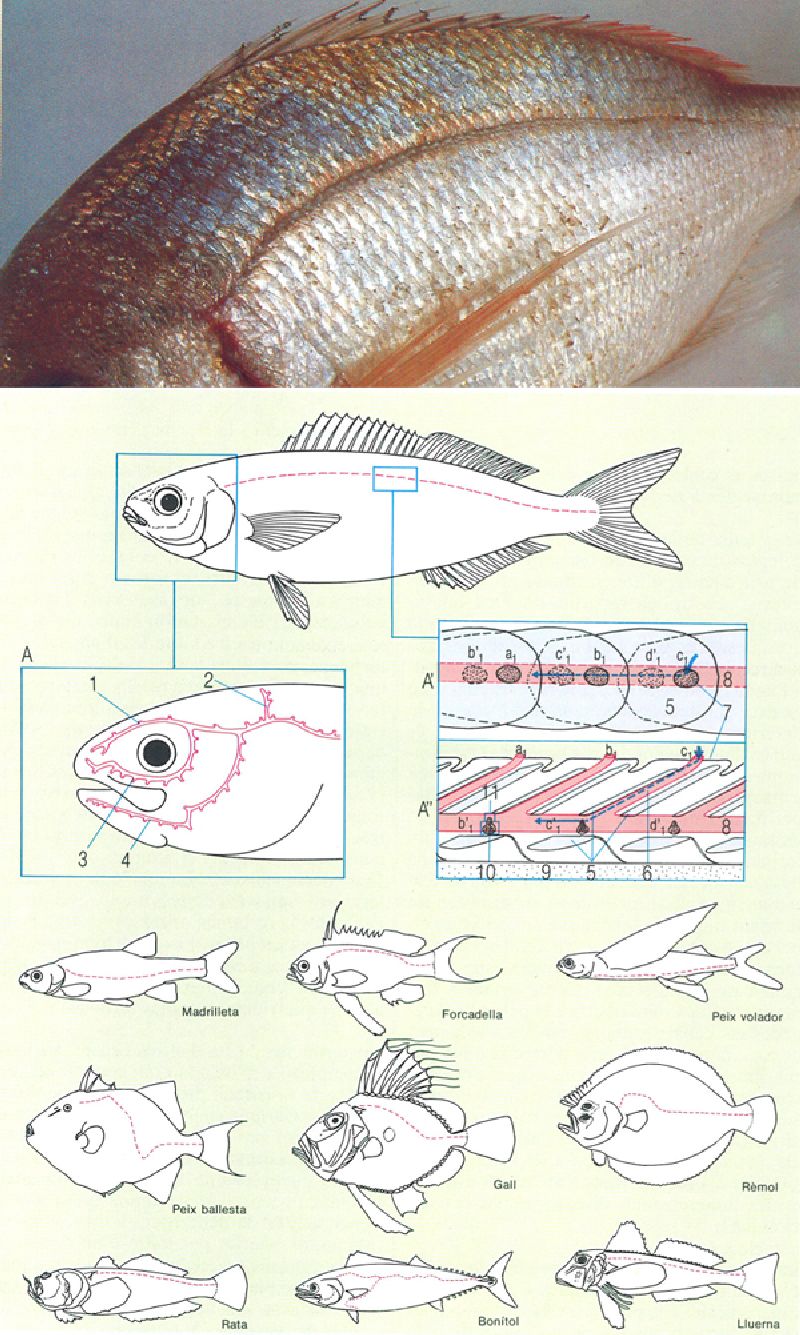
La percepció de les ones de pressió que es propaguen pel medi aquàtic es fa per mitjà dels neuromasts, receptors sensorials que es disposen seguint els nervis, al cap i a la línia lateral. En la fotografia s’observa la línia lateral d’un pagell (Pagellus erythrinus), formada per un seguit d’escates proveïdes d’uns petits porus o canals que comuniquen internament amb els neuromasts. En el dibuix, a baix, s’ha representat un model general de la posició de la línia lateral (en vermell), amb un detall (A) de la manera com es pot ramificar a la regió del cap (sovint, amb valor taxonòmic), i amb les seccions longitudinal (A’) i perpendicular (A") de la línia lateral, tallant les escates justament per la meitat del tub o canal de la línia lateral; en aquest dibuix s’han exagerat, per a fer-lo més comprensible, el gruix de l’escata i el de la línia lateral, i s’hi indiquen amb els símbols a1 a’1, b1, b’1, etc., els dos porus de cada escata (a’1 i b’1 queden amagats sota la part de l’escata que cavalca sobre la línia) que posen en comunicació els petits canals, i el trajecte que segueixen les ones de pressió. A la part de baix del dibuix s’han representat diferents tipus de línies laterals. 1 canal supraorbital, 2 canal supratemporal, 3 canal infraorbital, 4 canal mandibular, 5 escata, 6 tub que travessa l’escata, 7 porus, 8 canal de la línia lateral, 9 nervi, 10 neuromast, 11 cúpula.
Jordi Vidal; Francesc J. de Sostoa.
Encara hi ha un conjunt de tres tubs estrets i allargats, els canals semicirculars plens d’una substància densa, dita endolimfa. Aquests canals, que es disposen en plans perpendiculars (canals horitzontal, anterior i posterior), tanquen llurs circuits a la corresponent cambra de l’utricle; també es caracteritzen per un petit eixamplament a la part inferior (prop de l’utricle), que rep la denominació d’ampul·la. Els receptors sensorials pròpiament dits constitueixen les anomenades crestes de les ampolles, o les màcules o taques de les parets de les cambres. Així, a l’utricle es troba la màcula de l’utricle i la màcula neglecta; a les altres cambres hi ha la màcula del sàcul i la màcula de la lagena. Tant les crestes com les màcules són innervades per diverses branques del nervi auditiu. A l’interior de les tres cambres hi ha unes masses dures: els otòlits, que són unes estructures compostes essencialment de carbonat càlcic i proteïna. Aquests otòlits estan en contacte amb una membrana o cos gelatinós on penetren els filaments de les cèl·lules pilíferes sensorials. Les tres cambres de l’oïda interna intervenen com a òrgans de l’equilibri, especialment l’utricle; això no obstant també actuen com a òrgans auditius, sobretot el sàcul i la lagena. Les ones acústiques, quan es propaguen a través de l’aigua fan vibrar els otòlits, sempre que aquests tinguin una densitat considerable i, en conseqüència, estimulen les cèl·lules pilíferes de les màcules. El funcionament d’aquesta estructura com a òrgan de l’equilibri es basa en el fet que els moviments del cap de l’animal, en qualssevol dels plans, origina un desplaçament de l’endolimfa cap als canals semicirculars i, d’aquesta manera, estimula les cèl·lules pilíferes dels receptors sensorials o neuromasts de les crestes; aquests impulsos són transmesos al cervell d’una manera immediata per mitjà del nervi auditiu. Consegüentment, en aquest cas el funcionament de l’òrgan és directament relacionat amb l’avanç lineal del peix o bé amb el moviment de rotació del cos. Tanmateix la informació respecte als canvis de posició del cos i de l’acceleració procedeix d’una manera específica de les cèl·lules pilíferes de les màcules de les cambres, que són excitades per mitjà del desplaçament i per la pressió que efectuen els otòlits sobre aquestes àrees sensorials.
Esquema de l’aparell o òrgan de Weber, un conjunt de petits ossets (en groc) que posen la bufeta natatòria (en blanc) en comunicació amb el laberint membranós de l’oïda mitjana per tal d’amplificar els sons. A visió dorsal del cap d’un ciprínid i detall en visió lateral (A’), ampliat, per mostrar la posició dels canals semicirculars de l’oïda interna i la seva relació amb l’òrgan de Weber i la bufeta natatòria. 1 lòbul olfactori de l’encèfal, 2 lòbul òptic de l’encèfal, 3 cerebel, 4 ampul·la, 5 utricle, 6 sàcul, 7 lagena, 8 canals semicirculars, 9 si imparell, 10 escafium, 11 claustrum, 12 intercalarium, 13 tripus, 14 bufeta natatòria, 15 costella, 16 tronc de la vèrtebra, 17 tercer arc neural, 18 quart arc neural, 19 cinquè arc neural, 20 os suspensori.
Francesc J. de Sostoa, a partir de fonts diverses.
Pel que fa a les diferències entre els diversos grups d’àgnats i de peixos, l’estructura d’aquests òrgans varia sensiblement. En els àgnats (mixínids), el laberint consisteix en una estructura molt senzilla, formada per un sol anell en representació dels canals semicirculars anterior i posterior soldats. A la part inferior hi ha una cambra basal proveïda d’una màcula i d’una membrana o cos gelatinós amb inclusions de carbonat càlcic; aquestes inclusions actuen com un otòlit i exciten les cèl·lules sensitives. En canvi, les llamprees tenen un aparell més complex, amb dos canals semicirculars proveïts d’ampolles amb crestes sensorials. Endemés, disposen d’un utricle, un sàcul i una lagena, com també d’una màcula comuna; petits cristalls de carbonat càlcic inclosos en una mena de massa gelatinosa fan la funció dels otòlits. Hom considera que el sistema de l’equilibri de les llamprees és bastant desenvolupat, i que els permet de conèixer amb una relativa exactitud llur posició en l’aigua. El laberint dels elasmobranquis presenta algunes diferències respecte al dels àgnats que el fan més evolucionat. En primer terme, cal esmentar l’existència d’un utricle ben diferenciat, com també una lagena que es diferencia com una prolongació del sàcul. Hi apareixen, a més a més, els tres canals semicirculars que connecten amb l’utricle. Malgrat tot, aquests peixos encara estan mancats de veritables otòlits, la funció dels quals és substituïda per la presència d’unes membranes gelatinoses amb incrustacions calcàries.
Les ampul·les o canals de Lorenzini dels elasmobranquis són òrgans associats al sistema acústico-lateral, capaços de percebre canvis de temperatura, ones magnètiques i camps elèctrics. En el dibuix s’ha representat la seva distribució en el cap d’un tauró (A, A’, en negre les ampul·les i en vermell els porus de la línia lateral) i un detall d’una ampul·la de Lorenzini (A"); i en el cos d’una rajada (B), en visió dorsal (en gris) i ventral (en blanc), per mostrar la posició dels porus de les ampul·les i de les càpsules on desemboquen els canals de Lorenzini (se n’han representat només alguns). 1 porus extern, 2 tub, 3 ampul·la, 4 nervi, 5 càpsula, 6 canal.
Francesc J. de Sostoa, a partir d’originals diversos.
En els peixos ossis hom observa una disposició semblant pel que fa a les estructures del laberint, la qual, si més no, és més evolucionada quant al grau de diferenciació. Així, el laberint membranós consta clarament de tres cambres (l’utricle, el sàcul i la lagena) i de tres canals semicirculars. Tant l’utricle com els canals s’emplacen a la part superior, mentre que el sàcul i la lagena ocupen una posició inferior. Els otòlits dels peixos ossis són ben constituïts, bé que varien de forma i de mida segons les espècies. D’altra banda, cal esmentar que el gruix dels otòlits augmenta progressivament amb l’edat de l’individu, perquè durant el període de creixement s’hi dipositen, diàriament, fines capetes calcàries concèntriques; aquesta deposició de capes manté una determinada regularitat amb els períodes de creixement (que coincideixen amb els mesos temperats) i amb els de pausa, i consegüentment aquestes estructures poden emprar-se com un mètode per a determinar, amb exactitud, l’edat dels individus. Els peixos ossis més evolucionats disposen de tres tipus d’otòlits calcaris: l’anomenada sagitta o sageta, gran i localitzada al sàcul; el lapillus o lapil, que es troba a l’utricle; i l’asteriscus o asterisc, que es troba a la lagena.
Determinats peixos ossis, com ara els ostariofisis (amb unes 6200 espècies, gairebé totes d’aigua dolça), es caracteritzen per un conjunt d’ossets que constitueixen l’anomenat òrgan de Weber, que fan la funció de connectar la bufeta natatòria amb el laberint de l’oïda interna. Aquesta estructura millora la capacitat d’audició dels peixos: quan les ones sonores topen amb la bufeta natatòria fan que aquesta canviï de volum i transmeti les vibracions a través d’una cadena de petits ossets (claustrum, tripus, intercalarium i escafium) fins al sàcul, a la base de l’oïda. En d’altres casos, com en alguns clupèids, la bufeta natatòria s’estén en unes prolongacions acabades en forma de petites ampolles que penetren a l’utricle del laberint.
Els elasmobranquis, és a dir, els taurons i les rajades, sembla que estan dotats d’audició, però hom creu que aquesta capacitat es limita als sons de baixa freqüència. En aquests peixos el centre receptor dels sons és la màcula neglecta de l’utricle.
El sistema de la línia lateral consisteix en un complex de canals i de receptors sensorials o de neuromasts, d’origen similar als de l’oïda interna. Es troben repartits pel cap, on segueixen trajectòries peculiars segons cada grup i espècie, però també es distribueixen al llarg de la línia mitjana, és a dir a cada flanc, a banda i banda del cos. Aquest sistema, que és exclusiu dels àgnats, dels pisciformes i de determinats estadis dels amfibis aquàtics, és especialment adaptat per a percebre les vibracions o les ones de pressió que es propaguen a través de l’aigua, que o bé són originades pel medi físic o pels propis organismes. La unitat receptora típica és el neuromast, conformat per una cúpula o prolongació i per una base constituïda per cèl·lules pilíferes i de sosteniment. Aquests neuromasts es disposen tot formant fileres en la mateixa direcció que els nervis. Aquesta unitat estructural se situa al fons d’uns canals plens de mucus, que en alguns casos (holocèfals) poden arribar a formar veritables solcs oberts a la pell; altres vegades els canals romanen tancats i solament es comuniquen a l’exterior per mitjà d’uns porus minúsculs o d’uns orificis que travessen completament les escates de la línia lateral. Les ones de pressió, per unes vies o per les altres, arriben a les cúpules dels neuromasts, els quals les transfereixen a les cèl·lules sensitives i d’aquestes passen al cervell a través de les branques dels diversos nervis cranials (VII i X).
La línia lateral presenta modificacions d’acord amb les característiques i els hàbits de cada espècie. Els peixos ossis que viuen a les aigües corrents solen tenir els canals de la línia lateral tancats, com s’observa en el llop de riu (Noemacheilus barbatulus); en canvi, en d’altres peixos poden trobar-se parcialment o totalment oberts (determinats teleostis) i àdhuc en alguns, com el barb roig (Phoxinus phoxinus), es formen grups més o menys aïllats de cèl·lules sensorials. En el cas dels àgnats i els elasmobranquis, els òrgans sensorials romanen a l’interior d’uns solcs. La disposició de la línia lateral al llarg del cos, ja que les ramificacions d’aquesta al cap presenten uns patrons de distribució diferents fins i tot entre espècies, també varia força i sovint depèn de l’hàbitat que ocupen els peixos. Algunes vegades és corbada, com en els sorells (Trachurus), o dorsal, com en els peixos bentònics, per exemple la rata (Uranoscopus); altres vegades, ocupa una posició ventral en alguns peixos epipelàgics, com el peix volador (Exocoetus volitans). La funció de la línia lateral és la detecció de les preses, és a dir, de l’aliment, però també es relaciona amb la detecció d’altres peixos i la de determinats obstacles. Aquesta característica és especialment important en els peixos cavernícoles o en aquells que viuen en aigües tèrboles.
Nogensmenys hi ha d’altres òrgans associats al complex de la línia lateral, com ara les ampul·les de Lorenzini pròpies dels elasmobranquis. En els taurons aquestes estructures es localitzen al cap, especialment a la part davantera. En canvi, en les rajades consisteixen en uns tubs plens de mucus, amb una base globular proveïda de receptors sensitius que s’enclouen en la pell; aquests tubs, comunicats amb l’exterior per mitjà d’un porus, tenen la funció de detectar una sèrie d’impulsos elèctrics lliurats per altres peixos, tot i que d’una manera secundària també tenen la capacitat de percebre els canvis de temperatura de l’aigua. Les rajades i els taurons els empren per a orientar-se amb l’ajut dels camps magnètics o bé per a detectar la presència de les preses per mitjà dels canvis del potencial muscular, cosa que també s’ha observat en el gat (Scyliorhinus canicula). Els electroceptors dels peixos elèctrics d’aigua dolça, com ara l’anguila elèctrica (Electrophorus electricus), presenten una funció similar. Alguns elasmobranquis dotats de la facultat de produir electricitat, com la tremolosa (Torpedo), tenen un model diferent de receptor sensorial, que en aquest cas s’emplaça al cap: les vesícules de Savi.
L’aparell digestiu dels peixos
La boca
Fruit de les adaptacions a diferents formes d’alimentació, l’aparell bucal dels peixos mostra una gran diversificació, que no solament concerneix les proporcions i la disposició de la boca, sinó també la forma i la mida de les dents. La gran majoria de peixos mostren un dels models bàsics de disposició bucal: terminal, subterminal, ínfera i súpera. La més característica és la terminal, en què la boca se situa a la regió davantera del cap i en una posició mitjana. Aquesta morfologia bucal permet d’alímentar-se entre dues aigües, com també a la superfície o al substrat. Les espècies adaptades a la vida bentònica, que viuen i es nodreixen en zones properes al fons, solen tenir un tipus de boca infera o bé subterminal adaptada a aquests hàbits alimentaris; en aquests casos, la boca se situa a la part ventral del cap o orientada cap avall. Finalment, cal citar l’existència d’un altre grup de peixos que s’alimenten a la superfície o bé capturen les preses que els passen per sobre; consegüentment, tenen l’obertura bucal a la part superior del cap, és a dir, súpera.
Els ciclostòmats o àgnats difereixen dels altres vertebrats perquè tenen una boca circular en forma de ventosa, desproveïda de mandíbules i suportada per cartílags. Aquest tipus de boca succionadora permet que les formes paràsites puguin adherir-se al cos dels peixos, que perforen amb l’acció dels anomenats odontoides cornis. Aquests odontoides, que revesteixen l’embut bucal i la llengua, actuen d’una manera semblant a les dents, com passa en la llamprea de mar (Petromyzon marinus).
Entre els condrictis, la immensa majoria de taurons tenen una boca semilunar força característica, que ocupa una posició infera (llevat de l’espècie Chlamydoselachus anguineus, que la presenta terminal). Aquest emplaçament bucal els obliga a mossegar o capturar les preses tot efectuant desplaçaments laterals i inclinant el cos uns 90 graus. Malgrat tot, les rajades (Raja), que també presenten una boca infera, es nodreixen de peixos i d’invertebrats que capturen al fons. D’altres elasmobranquis hipotremats, com ara les mantes (Mobula, Manta), considerats uns grans nedadors pelàgics, tenen la boca situada a l’extrem del rostre; en aquest cas s’alimenten de petits peixos i invertebrats que condueixen fins a l’obertura bucal mitjançant unes expansions de les aletes pectorals.
Els peixos ossis normalment tenen la boca situada a l’extrem del cap, en posició terminal; així i tot, es troba tota mena d’especialitzacions que afecten la mida i posició de la boca. Els més primitius, com ara l’esturió (Acipenser sturio), tenen una boca menuda, circular i ínfera que precedeix un rostre prolongat; es nodreixen de petits invertebrats bentònics. Endemés, hi ha nombroses espècies d’hàbits bentònics que també presenten una boca subterminal, com ara alguns ciprínids: els barbs (Barbus) i les madrilles (Chondrostoma). En canvi ben pocs tenen una boca súpera com la gambúsia (Gambusia affinis), que captura insectes a la superfície de l’aigua. Però també hi ha alguns depredadors que habiten les grans fondàries marines de la plataforma continental, com ara el rap (Lophius piscatorius) i la rata (Uranoscopus scaber), que es camuflen i es valen d’una boca súpera per a sorprendre les seves preses quan els passen sobre el cap.
La mida de la boca, la seva posició i les característiques de les dents constitueixen uns bons indicadors dels hàbits alimentaris de cada espècie. Determinats depredadors, com ara els grans taurons (Prionace, Carcharodon, Isurus, etc.), l’espet (Sphyraena sphyraena) i el luci (Esox lucius), a més de comptar amb unes grans mandíbules, tenen dents tallants i de mida considerable. D’altra banda, el pelegrí (Cetorhinus maximus) i Polyodon spatula tenen una enorme obertura bucal que els serveix per a capturar petites preses o bé per a filtrar plàncton, perquè tenen dents molt rudimentàries, o no en tenen.
Entre els peixos amb hàbits alimentaris molt especialitzats, poden citar-se els de boca tubiforme, com el trompeter (Macroramphosus scolopax) i l’agulleta (Syngnathus acus): la boca s’obre a l’extrem d’un tub allargat que els permet d’engolir les preses per succió; n’hi ha d’altres que tenen mandíbules llargues i estilitzades, proveïdes de dents petites però agudes, com ara l’agulla (Belone belone). Determinades espècies disposen d’una boca protràctil que els serveix per a augmentar el volum de la cavitat bucal quan la projecten cap enfora. Pot citar-se l’esturió (Acipenser sturio) i el gall (Zeus faber), però també s’observa un efecte similar en alguns peixos abissals, com Chauliodus sloani, d’estructura bucal diferent. En alguns casos la forma de les mandíbules resulta francament grotesca, com passa en l’agulla de fons (Nemichthys scolopaceus), que les té corbades cap enfora i en sentit oposat. També n’hi ha d’espectaculars, com les del peix espasa (Xiphias gladius), la mandíbula superior del qual es prolonga en forma d’enorme i agut punxó.
La dentició
Les dents dels peixos poden revestir diferents ossos del cap. Normalment s’implanten al premaxil·lar, al maxil·lar i al dentari, però també poden fer-ho al vòmer i als palatins. En algunes espècies es poden fer extensives als ossos pterigoides, al parasfenoides, al basibranquial i al glossohial (a la llengua). Finalment, hi ha grups de peixos que tenen dents a la part superior o inferior del cinquè arc branquial (dents faríngies).
Dentició en els peixos ossis. A esquema d’una boca de peix oberta que mostra la posició de tots els ossos que poden portar dents en els peixos teleostis. 1 premaxil·lar, 2 vòmer, 3 maxil·lar, 4 palatí, 5 parasfenoide, 6 pterigoide, 7 basibranquial, 8 dentari, 9 glossohial, 10 dent faríngia. B visió ventral del cap d’una bagra (Leuciscus cephalus), per a mostrar la disposició dels arcs faringis, com també la forma, les dimensions i l’orientació de les dents. C dents caniniformes d’un làbrid (Halichoeres radiatus). D dents molariformes d’un espàrid (Calamus). E un altre tipus de disposició de les dents còniques, caniniformes, d’un peix depredador, en aquest cas un caràcid sud-americà.
Román Montull, a partir de fonts diverses.
Els àgnats, en lloc de veritables dents, presenten estructures de tipus cònic i de naturalesa còrnia (odontoides). Les autèntiques estructures dentàries comencen a aparèixer en els elasmobranquis (taurons i rajades).
En l’escala evolutiva, les dents dels taurons es consideren escates placoides que, a més de modificar la seva estructura i la seva funció, han canviat d’emplaçament. Aquestes dents són constituïdes essencialment per dentina coberta d’una capa d’esmalt. En examinar la mandíbula d’un selaci hom hi pot observar una sèrie de fileres de dents en diferents estadis de desenvolupament; a mesura que es desgasten són substituïdes contínuament per noves fileres. En general, solament és funcional la filera més exterior, fora de determinats taurons, com les mussoles (Mustelus), i de la majoria de rajades o d’hipotremats, que utilitzen diverses fileres alhora. Les dents dels elasmobranquis ofereixen una gran diversitat de formes i mides; pot succeir fins i tot que, dins la mateixa espècie (com en la família dels hexànquids), les dents de la mandíbula superior difereixin de la inferior. Generalment les dents dels taurons són subtriangulars, bé amb cúspides auxiliars o sense; així, el tauró blanc (Carcharodon carcharias) disposa d’una sola cúspide orlada de fines denticulacions i Eugomphodus taurus té les dents llises i formades per una cúspide principal, flanquejada per dues cúspides auxiliars. En els hipotremats, com ara les rajades (Raja), la milana (Myliobatis aquila) i els peixos guitarra (Rhinobatos), les dents es disposen en diverses fileres que recorden un mosaic de peces ròmbiques o hexagonals. D’altres peixos cartilaginosos, com les quimeres (holocèfals), mostren una dentició ben diferent, que es redueix a tres parelles de plaques dentàries llises, de funció eminentment trituradora.
Els peixos ossis o osteïctis, en comparació dels cartilaginosos o condrictis, de dentició exclusivament mandibular, presenten una dentició força més complexa i variada. Les dents s’implanten més fermament que les dels elasmobranquis i poden localitzar-se en diferents ossos de la boca, bé constituint-hi una o diverses fileres, bé estructurant-s’hi en forma de mosaic. La substitució dentària es realitza, a diferència dels anteriors, de forma irregular: les peces noves es formen sota les gastades o bé entre elles, però mai no es reemplacen fileres completes d’una manera simultània. Els peixos ossis depredadors, particularment els ictiòfags, com ara l’espet (Sphyraena sphyraena), Chauliodus sloani, el llobarro (Dicentrarchus labrax) i el luci (Esox lucius), entre d’altres, tenen unes dents fortes i agudes de morfologia diferent. Determinats peixos presenten algunes dents de forma i funció diferent a la resta, com les desenvolupades i corbades canines dels blènnids o bavoses (Blennius) i d’alguns gòbids o burrets (Gobius), les quals els permeten de capturar i d’immobilitzar les preses. És força extens el conjunt de peixos que tenen una alimentació mixta i que devoren diferents tipus d’invertebrats (cucs, crustacis, mol·luscs, etc); llur dentició concorda amb l’alimentació, com és el cas d’alguns espàrids com l’orada (Sparus auratd) i dels làbrids o tords (Labrus). Els peixos lloro (escàrids) tenen les dents soldades constituint un curiós aparell mastegador en forma de bec d’ocell; amb aquesta estructura poden desprendre i trencar bocins de corall del qual aprofiten la part orgànica i expulsen el carbonat càlcic (que conforma l’esquelet) a través del tub digestiu en la majoria de casos. Finalment, els ciprínids com el barb (Barbus) o la bagra (Leuciscus), habitants comuns de les nostres aigües dolces, no tenen dents a la part anterior de la boca i, en canvi, estan dotats d’un sistema triturador ben peculiar: les dents faríngies, que es disposen en una o diverses fileres i constitueixen un disseny característic de cada gènere.
El tub digestiu i els òrgans associats
Estructura del tracte digestiu en diferents tipus de peixos: A àgnats (llamprea). B elasmobranquis (tauró), C dipnous (Protopterus), D halecomorfs (Amia), E salmoniformes (Salmo), F perciformes (Diplodus). Cal notar la presència de conducte pneumàtic (16), que comunica el tracte digestiu amb la bufeta natatòria en els salmoniformes (fisòstoms), i que manca en els perciformes (fisoclists). 1 cavitat oral, 2 esòfag, 3 faringe (tub respiratori en les llamprees), 4 intestí, 5 vàlvula espiral, 6 cloaca, 7 estómac, 8 pílor, 9 conducte biliar, 10 anus, 11 glàndula digitiforme, 12 conducte pulmonar, 13 pulmó esquerre, 14 vàlvula pilòrica, 15 dents faríngies, 16 conducte pneumàtic, 17 vesícula biliar, 18 cecspilòrics.
Amadeu Blasco, a partir de fonts diverses.
El tracte digestiu varia considerablement al llarg de l’escaia evolutiva. Així, els grups primitius, desproveïts de mandíbules (àgnats), tenen un conducte poc diferenciat i ben diferent del dels condrictis i els osteïctis; així i tot, és constituït d’una manera general per l’esòfag, l’estómac i l’intestí.
A la cavitat bucal dels peixos hi ha un seguit de glàndules secretores d’un mucus que facilita el trànsit de l’aliment, ja que no tenen glàndules salivals. De la boca l’aliment passa per la faringe, tota perforada per les fenedures branquials, cap a l’esòfag, que sol ésser curt i dilatable, especialment en els depredadors que engoleixen preses grans; és en aquest tram que alguns peixos, com els mugílids o Hisses (Mugil, Liza), disposen de glàndules digestives.
Estructura general de l’aparell digestiu en els peixos: A àgnats (Ilamprea, Petromyzon marinus), B elasmobranquis (agullat, Squalus) i C teleostis flsòstoms (truita de rierol, Salvelinus fontmalis) i D teleostis fisoclists (perca americana, Micropterus salmoides). 1 obertura bucal, 2 faringe, 3 esòfag, 4 estómac, 5 cecs pilòrics, 6 pílor, 7 duodè, 8 intestí, 9 cloaca, 10 anus, 11 vàlvula espiral, 12 conducte urinari, 13 bufeta urinària.
Amadeu Blasco, a partir de fonts diverses.
En els peixos ossis menys evolucionats (fisòstoms), l’esòfag es connecta amb la bufeta natatòria. Després de l’esòfag l’aliment arriba a l’estómac, que és força modificat en alguns grups i indiferenciat en d’altres. Així, els àgnats, les quimeres i fins i tot algunes famílies de peixos ossis (com per exemple, els ciprínids), manquen d’un estómac pròpiament dit, talment que aquesta secció del tub digestiu no pot diferenciar-se de l’intestí. Malgrat tot, en la majoria dels peixos ossis l’estómac és ben diferenciat i fins varia de forma i de mida segons el règim alimentari de cada espècie: els ictiòfags, com ara el luci (Esox lucius) o el llobarro (Dicentrarchus labrax), el tenen de forma allargada, mentre que en els peixos de règim omnívor és de forma típicament corbada com una U. Són nombroses les modificacions de l’estómac que apareixen com a resultat de les adaptacions a règims alimentaris molt especialitzats, com l’estómac triturador de les Hisses (Mugil, Liza). I també és força comuna l’existència d’uns petits sacs o cecs pilòrics a la part posterior de l’estómac, que secreten enzims digestius. L’intestí dels peixos carnívors és més curt que el de les espècies herbívores; en aquest darrer cas pot excedir diverses vegades la longitud del cos. Però hi ha d’altres modificacions que diferencien els peixos primitius (condrictis i alguns osteïctis) dels més evolucionats. D’una banda, hi ha la vàlvula espiral, en substitució del veritable intestí que caracteritza la majoria d’osteïctis. D’altra banda, en les rajades, els taurons i en alguns peixos ossis primitius (dipnous), el recte desemboca a l’interior de la cloaca mentre que en la resta d’espècies desemboca a l’anus.
El fetge és un òrgan força desenvolupat en la majoria de peixos, però això s’acusa molt més en els elasmobranquis. Normalment és format per dos lòbuls, bé que en algunes espècies n’hi pot haver encara un altre de dimensions reduïdes i en d’altres, fins i tot, pot trobar-se’n un de sol, com s’esdevé en el salmó (Salmo).
El pàncrees dels àgnats és poc diferenciat, en contraposició al de les rajades i al dels taurons, ben constituït i amb dos lòbuls força característics, a més d’un conducte pancreàtic que connecta directament amb l’intestí; en els peixos ossis hi ha un pàncrees difós que es troba prop del fetge.
Sovint hom inclou la melsa entre els òrgans i les glàndules digestius, si bé la seva funció consisteix a controlar la producció de glòbuls vermells. Es tracta d’un òrgan relativament fàcil de reconèixer en els elasmobranquis, no solament pel color vermell que presenta sinó també per la seva forma piramidal i pel seu emplaçament proper a la regió pilòrica de l’estómac. En la majoria dels peixos ossis la melsa és força reduïda i de coloració vermella fosca.
L’aparell respiratori dels peixos
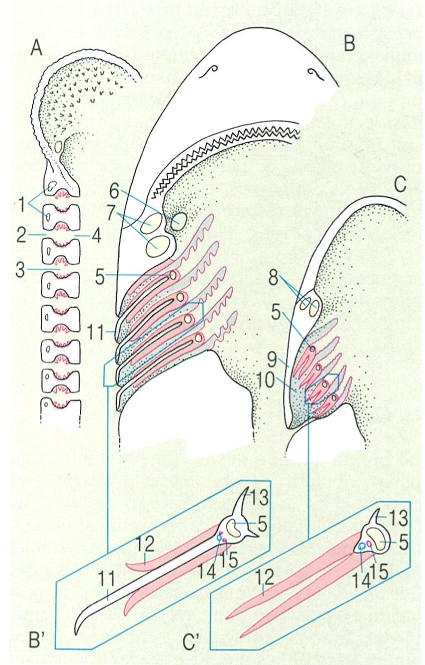
Configuració i nombre de cambres branquials (A àgnats, B elasmobranquis, C teleostis) i secció d’un arc branquial (B’ d’elasmobranqui, C’ de teleosti). 1 secció del cartílag de la cistella branquial, 2 fenedura branquial externa, 3 cambra branquial, 4 fenedura branquial interna, 5 arc branquial, 6 espiracle, 7 arc mandibular, 8 arc hioïdal, 9 opercle, 10 cambra opercular, 11 septe branquial, 12 filament branquial, 13 espina branquial, 14 artèria aferent, 15 artèria eferent.
Román Montull, a partir de fonts diverses.
El procés fisiològic de l’intercanvi de gasos, la captació d’oxigen mitjançant els eritròcits i l’eliminació del diòxid de carboni són força semblants en tots els vertebrats. La diferència bàsica entre els peixos i els tetràpodes consisteix en la presència de brànquies, que són substituïdes per pulmons en la resta de vertebrats. Les brànquies estan adaptades a la respiració en un medi mil vegades més dens que l’aire i amb un contingut considerablement menor d’oxigen. Són capaces de captar el 80% de l’oxigen que es troba dissolt en l’aigua que les travessa; en canvi, els mamífers no n’aprofiten més d’un 20-25%.
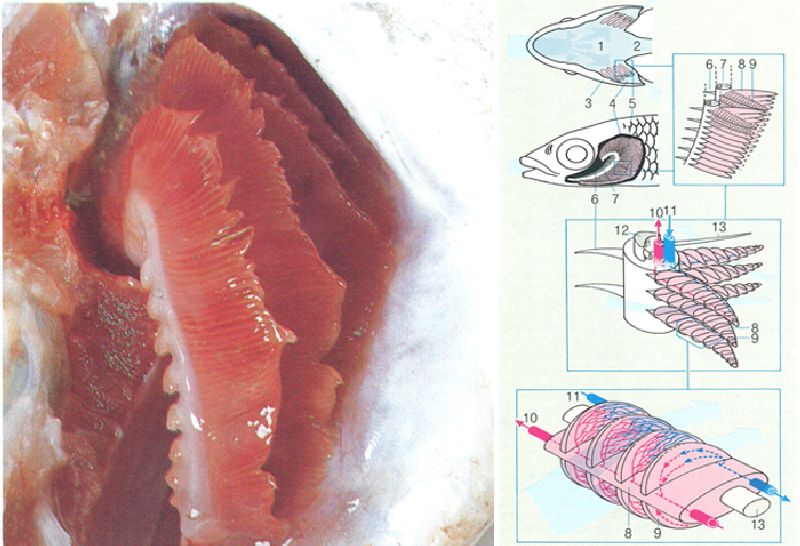
Les brànques dels peixos, que veiem en la fotografia (a l’esquerra), d’un peix ossi, concretament d’un pagell (Pagellus erythrinus) al qual s’ha arrencat I’opercle, són d’una estructura bastant uniforme en tots els grups. Cada brànquia consisteix (a la dreta) en un eix esquelètic arquejat, l’arc branquial (en groc), del qual pengen dues hemibrànquies, com dues ales de papallona tancades, cadascuna de les quals duu nombroses laminetes o làmines secundàries, molt ben irrigades. L’aigua entra per la boca del peix, es fica per entre aquestes laminetes perquè es produeixi l’intercanvi de gasos, i surt a l’exterior a través de l’opercle. 1 faringe, 2 esòfag, 3 opercle, 4 cambra opercular, 5 brànquies, 6 branquiospines, 7 arc branquial, 8 làmina primària o filament branquial, 9 làmina secundària, 10 artèria eferent, 11 artèria aferent, 12 esquelet branquial, 13 espina cartilaginosa de suport. Les fletxes de color blau clar indiquen la direcció dels corrents d’aigua.
Jordi Vidal; Román Montull, a partir de fonts diverses.
Les brànquies dels àgnats estan tancades dins d’uns sacs que es comuniquen alhora amb l’interior de la faringe i amb l’exterior, mitjançant les fenedures branquials, que s’obren independentment les unes de les altres.
L’estructura de les brànquies no és gaire diferent en les diverses classes de pisciformes. Essencialment consisteixen en una doble làmina sostinguda per un arc branquial. Aquestes làmines són constituïdes per fileres de filaments que s’implanten al llarg d’una estructura esquelètica o septe. La sèrie laminar d’una sola cara del septe s’anomena hemibrànquia i el conjunt d’ambdues làmines, holobrànquia. L’aparell branquial de les rajades i els taurons és format per un seguit de cambres branquials que s’obren aïlladament a l’exterior. El nombre d’aquestes fenedures branquials oscil·la entre cinc i set, segons les espècies. Les làmines branquials se separen per mitjà d’un septe ben desenvolupat. A més de les obertures branquials, els elasmobranquis presenten un espiracle a cada costat del pla de simetria, que no és sinó un orifici arrodonit que comunica la faringe amb el medi i que al seu interior conté la corresponent hemibrànquia o pseudobrànquia, sense funció respiratòria. D’altres peixos cartilaginosos, com ara les quimeres (holocèfals), consten de quatre arcs branquials recoberts per un replec cutani, que també rep la denominació de fals opercle, ja que recorda l’opercle dels peixos ossis. Els septes branquials dels peixos ossis manifesten una reducció progressiva i força considerable: en els més evolucionats es limiten a un ribet estret al llarg de l’arc branquial. En aquests peixos hi ha una tapa òssia o opercle que protegeix les brànquies i que s’articula amb l’arc hioïdal, de manera que l’opercle pot realitzar els moviments d’obertura i tancament que regulen el flux d’aigua circulant; també és típica la presència d’una pseudobrànquia a la cara interna de l’opercle.
Els peixos empren la pressió de dues bombes naturals per tal de fer circular l’aigua i, doncs, de respirar. D’una banda, en contreure’s la cavitat bucal, s’hi origina una pressió; seguidament, en obrir-se, la boca actua com una bomba suctora i fa entrar l’aigua. Posteriorment s’expandeix la cavitat branquial (bomba opercular), cosa que fa circular l’aigua a través de les làmines branquials on s’efectua l’intercanvi gasós. Finalment, es contreuen els opereles i l’aigua és expulsada a l’exterior. Aquest procés és força més simple en algunes espècies nedadores: el fet mateix d’avançar amb la boca oberta manté un flux continu d’aigua a través de les brànquies, com passa en els taurons i en alguns escòmbrids.
Els peixos bentònics han desenvolupat diferents adaptacions que els permeten de romandre inactius al fons i alhora continuar respirant. Així, alguns elasmobranquis, com les rajades (Raja) i les tremoloses (Torpedo), disposen d’un mecanisme de succió parabranquial que estableix un flux d’aigua a través dels espiracles, mentre romanen completament enterrats al substrat. Els peixos ossis bentònics com ara l’aranya (Trachinus araneus) i l’escórpora vera (Scorpaena porcus), resolen aquest problema augmentant el volum de les cambres branquials i el grau de desenvolupament dels radis branquiostegals.
Determinades espècies que viuen en hàbitats pobres d’oxigen –majoritàriament a l’Àsia, l’Àfrica i Amèrica del Sud–, estan dotades d’uns mecanismes especials per a captar-ne i pal·liar aquest defecte. Aquests mecanismes sovint consisteixen en un seguit de modificacions de l’aparell branquial, com ara els òrgans laberintiformes dels anabàntids (Anabas i Betta), el sistema de cambres branquials arborescents del siluriforme Clarias o les cambres aèries de Saccobranchus fossilis, que és una espècie originària de l’Índia. Altres espècies presenten adaptacions que només faciliten la respiració en determinades èpoques de l’any, quan el contingut d’oxigen dissolt a l’aigua és molt baix; així, els cobítids (Cobitis, Noemacheilus) tenen una porció de tub digestiu més vascularitzada i amb funció respiratòria que capta l’aire de la superfície. Alguns peixos també practiquen la respiració cutània, com ara l’anguila (Anguilla anguilla) i de l’anomenat saltador del fang (Periophthalmus), que viu en ambients tropicals.
La bufeta natatòria també pot actuar com un òrgan respiratori en els peixos dipnous, anomenats també «peixos pulmonats» perquè utilitzen aquest òrgan d’una manera semblant a un pulmó (Neoceratodus, Protopterus i Lepidosiren). Tant és així que la bufeta natatòria consta d’un o dos lòbuls compartimentats, proveïts alhora de petits sàculs que recorden els alvèols pulmonars. En alguns peixos fisòstoms la funció respiratòria de la bufeta natatòria és temporal o bé suplementària; aquest és el cas del sud-americà «pirarucu» (Arapaina gigas), un dels peixos d’aigua dolça més grans del món, i d’Amia i Lepisosteus, que colonitzen les aigües dolces de l’Amèrica del Nord.
La bufeta natatòria i l’equilibri hidrostàtic dels peixos
Estructura i disposició de la bufeta natatória en: A un peix fisòstom (bagra, Leuciscus cephalus), en el qual es comunica amb l’aparell digestiu per mitjà del conducte pneumàtic, i B en un peix fisoclist (esparrall, Diplodus annularis), en el qual aquell conducte ha desaparegut; en B’ s’ha dibuixat un detall, ampliat, de la bufeta, que mostra la glàndula del gas i la rete mirabilis; en B" s’ha representat l’òval, una petita cambra accessòria, molt vascularitzada, present en algunes famílies, que serveix per a reabsorbir els gasos. 1 bufeta natatoria, 2 conducte pneumàtic, 3 estómac, 4 intestí, 5 cecs pilòrics, 6 òval, 7 «rete mirabilis», 8 aorta dorsal, 9 artèria aliacomesentèrica, 10 artèria de la glándula del gas, 11 vena cardinal anterior, 12 vena cardinal posterior, 13 vena suprahepàtica, 14 vena porta, 15 vena de la glàndula del gas, 16 canal de Cuvier.
Román Montull, del natural i fonts diverses.
La bufeta natatòria és un sac ple d’aire, simple o lobulat i proveït d’una o de diverses cambres. Es localitza a la part superior del cos, sota la columna vertebral, encara que a les primeres etapes del desenvolupament hom ha observat que s’origina a partir de la paret del tub digestiu. Aquesta relació encara preval en els peixos anomenats fisòstoms, perquè hi ha una comunicació permanent entre aquest òrgan i l’esòfag o l’estómac mitjançant el conducte pneumàtic; en canvi, desapareix en els peixos teleostis més evolucionats (anomenats fisoclists) si més no en la fase adulta, i només s’observa a les primeres etapes del creixement larval, possiblement com una part del procés de formació de la mateixa bufeta natatòria. Tot i ésser un òrgan característic dels peixos ossis, no apareix en algunes espècies, sobretot en les bentòniques (gòbids i blènnids), a causa de llur adaptació a la vida del fons. També n’estan mancats els àgnats i els peixos cartilaginosos. Les funcions de la bufeta natatòria són diverses. A més de complir una funció hidrostàtica, es relaciona amb la producció i la percepció de sons, i amb la respiració.
La funció hidrostàtica és la més primordial i comuna a la majoria de peixos ossis (teleostis). La bufeta natatòria de les espècies d’aigua dolça és més voluminosa que la dels peixos marins, en relació amb la diferència de densitat entre aquests dos medis aquàtics. Això s’explica per la mateixa funció d’aquest òrgan, que és la de contrarestar el pes del peix dins l’aigua, és a dir, neutralitzar-lo i facilitar la flotabilitat sense necessitat de fer un esforç muscular; això evita la despesa energètica contínua que comportaria la manca de bufeta natatòria. Quan els peixos efectuen desplaçaments verticals i, consegüentment, canvien de profunditat, han de controlar la pressió dels gasos de la bufeta natatòria per tal de compensar constantment llur pes corporal en l’aigua. Per als peixos fisòstoms això no representa cap problema, perquè poden eliminar o, contràriament, engolir aire per la boca, i a través del conducte pneumàtic desinflar o inflar la bufeta. Els peixos fisoclists adopten un mecanisme més sofisticat i força efectiu: una densa xarxa de capil·lars sanguinis («retia mirabilia») al voltant de la bufeta natatòria hi facilita l’intercanvi de gasos (oxigen, nitrogen i diòxid de carboni), gràcies a les diferències que s’originen entre les pressions parcials dels gasos a ambdós compartiments. Finalment, la capacitat hidrostàtica que denoten els enormes peixos desproveïts de bufeta natatòria, com ara els elasmobranquis, s’assoleix mitjançant l’esforç muscular i mitjançant una morfologia corporal òptima: disposen d’unes aletes pectorals especialment dissenyades per a oferir una considerable resistència a l’aigua, i, a més, algunes espècies presenten un gran desenvolupament hepàtic amb un contingut ric en lípids i en esqualè (hidrocarbur), que contribueixen a la flotabilitat.
Gran part dels peixos fisòstoms colonitzen les aigües dolces, mentre que els fisoclists són els grans colonitzadors del medi marí. Per tal que la bufeta natatòria neutralitzi el pes del peix dins l’aigua, ha de tenir una capacitat d’un 7% respecte al volum corporal en els peixos d’aigua dolça i d’un 5% en els marins. D’altra banda, aquest òrgan sempre ha d’estar dilatat a fi de mantenir la seva capacitat hidrostàtica. D’aquesta manera està obligat a contrarestar contínuament la pressió del medi, que augmenta a raó d’una atmosfera per cada 10 m de profunditat. Consegüentment, quan el peix davalla a una profunditat de 10 m es troba sotmès a una pressió de dues atmosferes i la bufeta natatòria es comprimeix a la meitat del volum que tenia a la superfície. Per tal de mantenir la pròpia flotabilitat, el peix infla la bufeta i evita d’esdevenir més dens que l’aigua. Quan el peix pretén de tornar a la superfície es produeix una situació diferent; llavors li cal eliminar l’excés de gas perquè l’efecte contrari li duplica el volum del seu òrgan hidrostàtic en passar de dues a una atmosfera de pressió. En condicions naturals el peix realitza un ascens lent que li permet de reduir gradualment el volum sobrant, però resulta fatal si s’extreu sobtadament el peix de l’aigua, perquè li esclata la bufeta o bé s’infla com un globus. Això explica que durant l’activitat pesquera sovint s’observin exemplars que expulsen per la boca llur bufeta natatoria o d’altres òrgans, com ara l’aparell digestiu, impulsats per aquesta mateixa.
L’aparell circulatori dels peixos
La sang
Els peixos generalment presenten un escàs volum de sang al cos, enfront de la resta de vertebrats. Oscil·la entre 0,06 i 0,08 ml/g en els elasmobranquis, i encara és sensiblement més petit en els peixos ossis, entre 0,02 i 0,04 ml/g. Aquestes xifres equivalen a un 6,6% del pes corporal en el cas dels condrictis i a un 3% en el dels osteïctis. Aquest descens del volum sanguini dins l’escala filogenètica pot respondre al mateix desenvolupament del sistema vascular, que s’assoleix plenament en els peixos ossis més evolucionats i que fa innecessari un gran volum sanguini per a realitzar amb eficàcia el transport d’oxigen i de les altres substàncies que es troben al torrent circulatori dels peixos. La sang transporta ions i determinats compostos orgànics com ara les hormones, les vitamines i algunes proteïnes plasmàtiques (l’alfa, la beta i la gammaglobulina, l’albúmina, etc.). La concentració osmòtica de la sang varia segons l’hàbitat del peix i segons el seu grau d’adaptació. La sang porta diferents tipus de cèl·lules, com ara els glòbuls vermells o eritròcits i els glòbuls blancs o leucòcits. Del color vermell característic dels eritròcits n’és responsable l’hemoglobina, que compleix la funció de transportar oxigen en combinació reversible; aquesta cèl·lula sanguínia és, a diferència dels mamífers, nucleada i usualment en els peixos és de forma oval, però en els àgnats és circular. Hi ha diferències importants pel que fa a la mida dels eritròcits dels diversos grups d’àgnats i de pisciformes; aquests valors oscil·len entre els 9 microns de les llamprees, els 12-14 microns dels osteïctis i els 20-27 microns dels elasmobranquis; endemés hi ha una relació inversa entre la mida i el nombre de glòbuls vermells, cosa que explica que en els taurons hi hagi 0,5 × 106/mm3 d’eritròcits i en canvi n’hi hagi uns 2 × 106/mm3 en la majoria de peixos ossis. Finalment, cal afegir que aquestes cèl·lules s’originen al ronyó i a la melsa. Pel que fa als leucòcits, no tan nombrosos com els eritròcits, els pisciformes en tenen uns 0,15 × 106/mm3; aquesta xifra inclou els granulòcits, els trombòcits, els limfòcits i els monòcits; aquestes cèl·lules sanguínies es formen a l’òrgan de Leydig en el cas dels elasmobranquis i als ronyons dels teleostis.
El cor i el sistema vascular
L’aparell circulatori en els diferents grups de peixos. A, A’ funcionament de la circulació en els àgnats (una llamprea en el dibuix) i detall de l’estructura del sistema cardioaòrtic (A"). B, B’ funcionament de la circulació en els elasmobranquis (un tauró en el dibuix) i detall de l’estructura del cor (B"). C, C’ funcionament de la circulació en els peixos ossis i detall ampliat de l’estructura del cor (C"). 1 artèria carótida, 2 anell cefàlic, 3 aorta dorsal, 4 artèria branquial eferent, 5 brànquies, 6 aorta ventral esquerra, 7 aorta ventral dreta, 8 aorta ventral, 9 artèria branquial aferent, 10 artèria pseudobranquial, 11 espiracle, 12 artèria epibranquial, 13 artèria cardíaca, 14 artèria oftàlmica, 15 artèria caròtida, 16 circuit cefàlic, 17 artèria de la bufeta natatòria, 18 artèria celíaca, 19 artèria mesentèrica, 20 vas aferent, 21 vas eferent, 22 artèria hioïdal, 23 si venós 24 aurícula, 25 ventricle, 26 bulb o con aòrtic, 27 si venós, 28 con arterial, 29 bulb arterial.
Amadeu Blasco, a partir de fonts diverses.
El cor típic dels peixos presenta un esquema ben simple: bàsicament és format pel si venós, una aurícula, un ventricle i el con arterial. En realitat consisteix en una bomba impulsora que es localitza a la cavitat pericàrdica, és a dir, sota la faringe i entre les brànquies. Preparat per a realitzar una circulació simple, difereix força del cor dels mamífers, apropiat a una circulació doble i completa. La sang retorna al cor pel si venós, entra a l’aurícula i és impulsada pel ventricle cap a les brànquies a través del con arterial; el ventricle manté el ritme cardíac.
Els àgnats tenen un cor primitiu i no tan compacte com el de la resta de vertebrats. En les llamprees s’observa un si venós de morfologia allargada i tubular, a més d’una aurícula col·locada asimètricament a l’esquerra del ventricle, a la qual desemboca una vena cardinal proveïda d’un eixamplament. La sang passa al ventricle a través d’un orifici proveït d’una vàlvula i d’aquí a un esbós de con arterial; després es dirigeix a l’aorta ventral, de la qual parteixen vuit parelles d’artèries branquials aferents que la fan arribar a cadascuna de les set fenedures branquials i a les hemibrànquies, i finalment ateny l’aorta dorsal mitjançant un sistema eferent simètric. Pel que fa al sistema de vasos sanguinis, l’aorta ventral de les llamprees és simple fins a la tercera fenedura branquial, i entre les fenedures tercera i la quarta es divideix en dues branques (dreta i esquerra); l’aorta origina vuit parelles d’artèries branquials aferents, que penetren a les parets de les obertures branquials, mentre que l’aorta dorsal és simple i de disposició mitjana, tret del cap, on defineix una estructura circular anomenada anell cefàlic. La vena caudal es localitza sota l’artèria caudal, i es divideix tot formant les venes cardinals posteriors; aquestes venes són les que reben la sang dels ronyons, les gònades i les músculs dorsals.
En els elasmobranquis, tant el si venós com l’aurícula, el ventricle i el con arterial són ben desenvolupats; en els taurons, a més d’ésser l’aurícula més gran que el ventricle i d’haverhi un con arterial proveït de vàlvules, l’aorta ventral es prolonga quasi sempre en cinc artèries aferents i el sistema eferent és format per les artèries epibranquials, que desemboquen a l’aorta dorsal. El sistema de vasos sanguinis consta de cinc artèries aferents que parteixen d’una sola aorta ventral i penetren a cadascuna de les obertures branquials; d’aquesta manera la sang arriba a cada holobrànquia i a la pseudobrànquia de l’espiracle. L’aorta dorsal és simple i es converteix en l’artèria caudal a la part posterior del cos.
En els peixos ossis, en canvi, trobem un ventall ben ampli de modificacions, fruit del llarg procés evolutiu a què han estat sotmesos des dels dipnous, els condrostis (esturions) i els holostis (Amia i Lepisosteus) fins als teleostis primitius i, encara, d’aquests als teleostis més evolucionats. En els dipnous (peixos pulmonars), especialment el gènere Lepidosiren, l’aurícula és dividida en dues seccions per un septe vertical: la regió dreta rep la sang venosa del si i l’esquerra acull la sang oxigenada de la vena pulmonar; aquesta separació es fa extensiva al ventricle per un septe longitudinal incomplet. A continuació hi ha un con arterial espiraliforme i flexible, proveït d’unes vàlvules semilunars (vàlvula espiral) que el subdivideixen en una porció dorsal i una de ventral, de manera que la separació dels dos tipus de sang és incompleta: així, la sang venosa de la regió dreta del cor passa essencialment a la porció dreta del con arterial, mentre que la sang que torna dels pulmons ho fa a la porció ventral. El cor dels teleostis s’assembla força al dels elasmobranquis, però el con arterial és reduït a una estructura petita i dilatable, que adopta una coloració blanquinosa quan el peix és mort, i gaudeix d’una gran capacitat d’expansió; una única vàlvula impedeix el retrocés de la sang quan és impulsada a gran pressió pel ventricle. El sistema de vasos sanguinis és similar al dels elasmobranquis, però solament s’hi troben quatre artèries aferents perquè desapareix la branca corresponent a l’espiracle, exclusiva dels peixos cartilaginosos. L’aorta dorsal, situada a la regió anterior del cos, és doble en els teleostis, diferentment dels elasmobranquis; en canvi, la zona posterior d’aquesta aorta dorsal és simple en ambdós casos i constitueix l’artèria caudal a l’extrem del cos. Les artèries subministren sang oxigenada a les vísceres i a la musculatura, a partir de l’aorta i de l’artèria caudal. En els pisciformes la vena caudal penetra als ronyons per mitjà del sistema portarenal. D’altra banda, les venes postcardinals reben la sang dels ronyons, de les gònades i de la musculatura; finalment la condueixen fins al conducte de Cuvier on convergeixen les venes cardinals anteriors, les subclàvies i les jugulars. A partir d’aquest conducte de Cuvier la sang venosa entra al si venós, on rep, a més a més, la sang procedent del sistema portahepàtic.
D’una manera general hom pot dir que la sang dels pisciformes circula a través d’un sistema format pel cor i pels vasos sanguinis. El cor compleix la funció de bombà propulsora i impulsa la sang venosa a través de l’aorta ventral fins a les artèries branquials aferents. Les contraccions del ventricle generen una elevada pressió que obliga la sang venosa a passar pels canals interlaminars i d’aquesta manera la fan arribar, ja oxigenada, a les artèries eferents; des d’aquestes, la sang passa a l’aorta dorsal i es distribueix per tot el cos. Quan la sang travessa les brànquies se satura entre un 85 i un 95% d’oxigen i es transforma en sang arterial. D’altra banda, la sang venosa procedent dels òrgans i de la musculatura retorna al cor i desemboca al si venós on s’inicia novament el cicle.
L’aparell excretor i l’excreció dels peixos
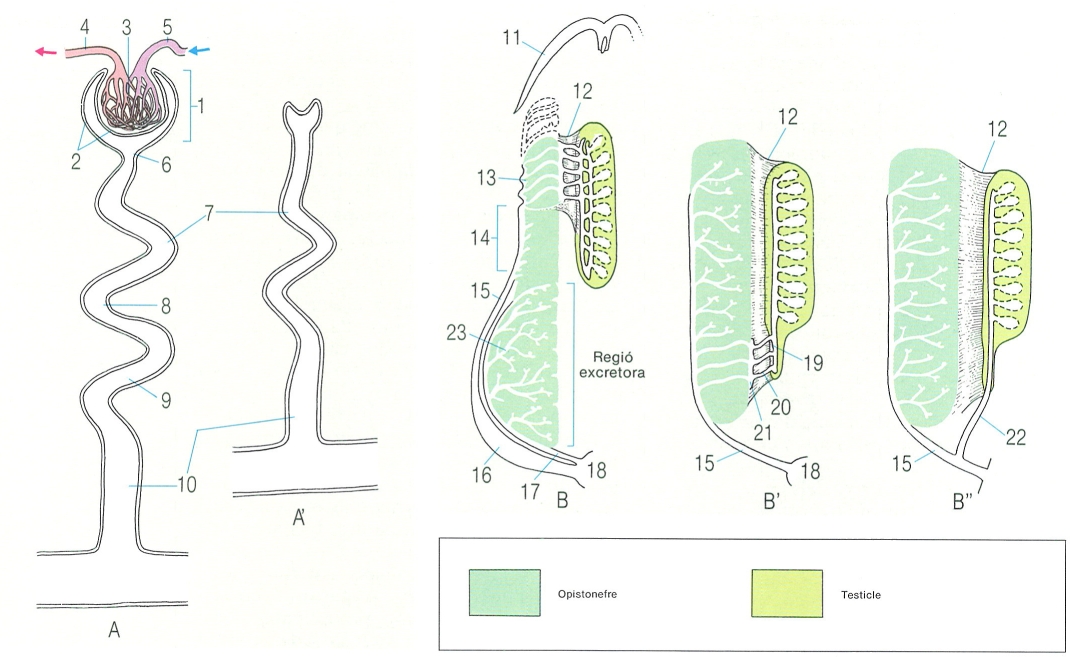
L’aparell excretor en els pisciformes. A l’esquerra, tipus de nefrons dels peixos ossis: A nefró glomerular, típic dels peixos d’aigua dolça, i A’ nefró aglomerular, típic dels marins. A la dreta, tipus de ronyons: B ronyó d’elasmobranqui, B’ ronyó d’un dipnou (Lepidosiren), i B" ronyó de teleosti. 1 corpuscle renal, 2 càpsula de Bowman, 3 glomèrul, 4 artèria eferent, 5 artèria aferent, 6 coll, 7 segment proximal, 8 segment intermedi, 9 segment distal, 10 túbul col·lector 11 oviducte vestigial, 12 mesorquium, 13 conducte de l’epidídim, 14 òrgan de Leydig, 15 conducte arquinèfric, 16 vesícula seminal, 17 conducte urinari principal, 18 cloaca, 19 canal testicular, 20 conducte eferent, 21 canal renal lateral, 22 espermiducte, 23 conducte urinari secundari.
Román Montull, a partir de fonts diverses.
L’estructura dels ronyons és força variada i complexa, segons les peculiaritats de cada grup de pisciformes i les adaptacions fisiològiques a la vida en ambients tan diferents com són el marí i l’aigua dolça. En els grups més primitius de vertebrats, com ara els àgnats, els condrictis i alguns osteïctis (condrostis i holostis), l’aparell reproductor és associat a 1’excretor. Malgrat tot, ambdues funcions queden ben separades en gran part dels peixos ossis.
Els ronyons dels àgnats i dels pisciformes són parells, generalment allargats, prims i de coloració vermella fosca. Són emplaçats a la regió dorsal i pròxims entre ells, per sobre de la columna vertebral i sota el peritoneu. Els ronyons compleixen la missió d’eliminar determinats productes d’excreció que són tòxics per a l’organisme. Aquestes toxines s’alliberen a l’exterior en forma d’orina (o bé dissolts en ella) mitjançant els conductes urinaris. La funció excretora es concentra bàsicament a la porció posterior dels ronyons, on poden presentar-se determinades modificacions, com sol passar amb els grups primitius esmentats anteriorment; és a dir, en aquesta part de l’òrgan pot persistir una funció reproductora. En altres peixos la porció anterior del ronyó actua com a òrgan hematopoètic; això vol dir que té la capacitat de produir glòbuls vermells, per bé que també pot presentar funcions endocrines.
La unitat estructural bàsica de l’aparell excretor és el nefró, constituït pel corpuscle de Malpighi i per una aglomeració de capil·lars sanguinis que rep la denominació de glomèrul. Al seu torn, el glomèrul és envoltat d’una estructura de paret doble o càpsula de Bowman. La sang que arriba al glomèrul torna al cor a través de les venes postcardinals. Les substàncies excretades per l’organisme passen del glomèrul a l’interior de la càpsula de Bowman i d’aquesta al túbul renal, el qual es diferencia en una porció terminal i una altra de distal.
La regió anterior del ronyó, també anomenada regió paranèfrica, és una estructura transitòria que solament perdura en l’estat adult, i a més modificada, en els àgnats (mixínids) i en un reduït grup de pisciformes (mictòfids, zoàrcids i gobiesòcids). Les llamprees, els condrictis i la resta d’osteïctis tenen un ronyó de tipus opistonèfric, que té un emplaçament posterior i disposa de glomèruls interns. Entre els àgnats, els mixínids encara conserven a més a més el ronyó pronèfric. Són ronyons parells de mida petita i situats en posició dorsal, prop del cor. Cadascun d’aquests ronyons presenta un gran corpuscle format per la unió de tres glomèruls. La funció d’aquesta estructura és força complexa en els individus en estat adult, i tan aviat pot actuar com un òrgan hematopoètic com tenir una funció limfàtica. Els ronyons opistonèfrics dels àgnats, de morfologia prima i allargada, s’estenen al llarg del cos de l’animal i estan proveïts de més de 30 corpuscles a cada banda. En aquesta varietat de ronyons els túbuls renals són simples (arquinèfrics) i van a parar a un conducte que desemboca en un si uro-genital, el qual es connecta amb l’exterior mitjançant una obertura situada a l’extrem de la papil·la uro-genital. Les llamprees només tenen ronyons opistonèfrics emplaçats al llarg de la regió dorsal i envoltats de teixit adipós. Els túbuls renals són ciliats i van a parar en un conducte que desemboca en un si uro-genital; la sortida a l’exterior es fa de la mateixa manera que en els mixínids.
Els elasmobranquis tenen un ronyó aplatat i sovint de morfologia lobulada. En els mascles, la part anterior d’aquest òrgan forma part de l’aparell reproductor i constitueix l’òrgan de Leydig; en canvi, aquesta regió és força reduïda en les femelles. Els ronyons de la femella adopten una coloració vermella fosca i es localitzen sota el peritoneu, rere una aparença d’òrgan segmentat. Endemés tenen conductes urinaris que s’eixamplen de la zona anterior a la posterior fins a constituir a cada costat els anomenats sins urinaris; aquests, units entre ells, conformen el si urinari mitjà que desemboca a l’exterior per la papil·la urinària. En canvi, cadascun dels ronyons del mascle presenta cinc conductes urinaris secundaris que es reuneixen en un conducte urinari principal, els quals desemboquen al si uro-genital. La concentració d’urea en la sang d’aquests pisciformes és força elevada.
Finalment, si bé el ronyó dels peixos ossis ofereix una gran diversitat morfològica, també pot dir-se que presenta una estructura força uniforme. Tanmateix, en els més primitius, com ara els condrostis (esturions), els mascles encara tenen ronyons amb una funció doble, és a dir, reproductora i excretora. En canvi, els teleostis han evolucionat fins al punt de presentar diferents estructures renals segons el medi que colonitzen. Així, els d’aigua dolça gaudeixen d’una estructura de nefró típic, que consta del glomèrul, la càpsula i el túbul renal. Les espècies marines, en canvi, disposen d’unitats renals modificades, amb uns glomèruls força reduïts o bé sense glomèruls. Hom pot dir que en els teleostis l’evolució ha portat a una considerable diversitat de formes que afecten el volum dels glomèruls, la forma dels túbuls renals i, finalment, tant la posició com les característiques dels ronyons.
Els sistemes d’osmoregulació dels peixos
L’osmoregulació i l’excreció en els peixos ossis. S’indiquen les diferències en l’intercanvi de sals, amoni, clorur i sodi entre un peix d’aigua dolça, a dalt (barb, Barbus), i un peix marí, a baix (sardina, Sardina) i els respectius medis on viuen.
Román Montull, a partir de fonts diverses.
Els peixos han de mantenir un equilibri constant entre la concentració de sals dels seus teixits i la del medi que els envolta. Això fa que s’hagin hagut d’adaptar als diferents ambients a base de regular i mantenir el contingut d’aigua, de sals (sodi, potassi i clor) i d’altres soluts en llurs cèl·lules. La regulació de l’equilibri osmòtic constitueix una de les principals missions de l’aparell excretor, si bé en aquest procés també intervenen les brànquies i, indirectament, molts altres mecanismes. Els àgnats presenten els fluids corporals amb el mateix contingut de sals que l’aigua marina, i per això s’anomenen isosmòtics, com els mixínids. En canvi, els peixos marins tenen una concentració osmòtica inferior a la del medí i per això són hiposmòtics; tendeixen a perdre aigua i a guanyar ions. Finalment, els peixos d’aigua dolça són hiperosmòtics, car elsseus líquids corporals són més concentrats que els de l’aigua on viuen; tendeixen a ingressar aigua i a perdre ions de l’organisme.
Entre els àgnats, els mixínids, d’origen netament marí, són isosmòtics o en tot cas lleugerament hipertònics respecte de l’aigua de mar. No obstant això, les llamprees, que consten de diverses espècies d’aigua dolça o que si més no passen una part del seu cicle biològic als rius, són força diferents dels mixínids pel que fa a l’aspecte de la regulació osmòtica. Així, quan viuen en aigües dolces, la pell i les brànquies són permeables a l’entrada d’aigua; la llamprea de riu (Lampetra fluviatilis) ingressa diàriament fins a un terç del seu propi pes d’aigua; les sals poden ésser excretades per una abundositat d’orina i a través de la superfície del cos, i la compensació d’aquestes sals eliminades s’efectua per mitjà de les brànquies i per l’alimentació, perquè després aquestes substàncies són seleccionades pel mateix ronyó. Quan viuen a mar, les llamprees posen en funcionament el mecanisme contrari; la sang és més diluïda que l’aigua de mar (hiposmòtica) i llavors tendeixen a expulsar aigua per les brànquies, per la pell i per l’orina. En aquest cas, per tal de compensar el balanç de sals poden ingressar aigua marina o captar-ne els ions selectivament mitjançant les anomenades cèl·lules de clorur. Com que en aquest cas la pèrdua d’aigua pot realitzar-se per diverses vies, l’orina és escassa.
En els elasmobranquis i els holocèfals, que colonitzen la mar d’una forma exclusiva, s’observa que la concentració d’ions a la sang s’assembla força a la dels peixos marins, que són hiposmòtics. Nogensmenys, l’osmolaritat d’aquests peixos és similar o superior a la de la mar, però en aquest cas especial és important l’elevada concentració d’urea i d’altres compostos nitrogenats en la sang, que d’alguna manera també contribueixen a mantenir l’equilibri osmòtic de l’organisme. Quan l’urea, com a producte metabòlic, passa pel ronyó se’n reabsorbeix entre un 70 i un 90% en uns segments especials; això significa que pot arribar a representar entre un 2 i un 2,5% del plasma sanguini. Com que les brànquies d’aquests peixos són molt permeables, l’ingrés d’aigua és important, però l’excés s’excreta en forma d’orina. D’altra banda, l’entrada de sals es realitza per difusió a través de les brànquies i per via digestiva, perquè la concentració d’aquestes sals és inferior als teixits amb relació al medi on viu l’animal. Finalment, el ronyó i la glàndula rectal constitueixen la principal via de sortida dels ions.
Els peixos ossis marins (teleostis) són hiposmòtics, amb una concentració d’un terç o un quart respecte al medi en el qual viuen. Per tal d’equilibrar la pressió osmòtica ingereixen aigua marina per via oral; en aquest sentit, la quantitat d’aigua ingerida varia segons la concentració del medi. L’excés d’alguns ions, com ara el sodi o el clor, originat per la ingestió d’aigua marina és excretat selectivament a través de les brànquies gràcies a l’existència de les cèl·lules de clorur; d’altres, ions són eliminats pel ronyó. Alguns teleostis presenten unes unitats renals de tipus aglomerular, és a dir, sense glomèrul, que no constitueixen sinó un mecanisme per a evitar les pèrdues d’aigua.
Els teleostis que colonitzen les aigües dolces són hiperosmòtics, en contraposició als marins: els fluids corporals tenen una pressió osmòtica que supera la del medi. Consegüentment tendeixen a ingressar aigua del medi, la qual cosa es realitza per mitjà de qualsevol superfície permeable; així, reïxen a diluir l’elevada concentració de sals al seu cos. Alguns ions, com ara el sodi i el clor, són excretats d’una forma passiva (per simple difusió) a través de les brànquies i de l’epiteli. El ronyó elimina l’excés d’aigua i això es tradueix en una gran producció d’orina. El fet que la principal pèrdua d’aigua es realitzi a través de l’orina comporta una reducció de les sals dissoltes, que ha d’ésser compensada per via digestiva, és a dir, per mitjà de l’alimentació. Això no obstant, també cal esmentar l’existència d’altres mecanismes actius que, selectivament i a través de les cèl·lules de clorur, els permet de recuperar els soluts eliminats.
Entre els peixos, com en altres organismes aquàtics, hom troba algunes espècies que gaudeixen d’una gran tolerància a les diverses concentracions salines; aquests organismes reben les denominacions d’eurihalins. Contràriament, n’hi ha que són molt estrictes i fets a les aigües on viuen; aquests organismes que mostren una tolerància reduïda a un estret marge de variació s’anomenen estenohalins. Els peixos marins poden ésser eurihalins i tolerar, en conseqüència, aigües salabroses que presenten entre un 18 i un 0,5‰ de sals dissoltes; àdhuc n’hi ha que són capaços de viure en aigüa dolça, amb valors inferiors a un 0,5‰ de sals dissoltes. Així mateix els peixos d’aigua dolça poden ésser eurihalins i, en contraposició als peixos marins, mostrar una gran tolerància a l’augment de concentració de sals del medi. Hom observa que algunes espècies tan aviat poden viure en aigüa dolça i salabrosa com en aigua de mar. D’una manera generalitzada, però, aquests canvis comporten per a l’organisme una gran despesa energètica, imprescindible per tal de controlar el seu equilibri osmòtic en un medi de condicions tan diferents. Hi ha diverses categories de peixos eurihalins: els diàdroms (anàdroms si viuen a la mar i catàdroms en cas invers), els estuarians i els migradors estacionals. Els peixos diàdroms, que realitzen part del seu cicle biològic en les aigües marines, abans no passen d’un medi a l’altre mostren uns canvis progressius de llur fisiologia. De fet, quan romanen en el medi marí la concentració de sals del seu organisme és superior a la que tenien quan vivien en aigua dolça. Per tal d’adaptar-se a la vida marina les espècies catàdromes han d’engolir aigua del medi, com ara l’anguila (Anguilla), que durant la seva migració a la mar es veu obligada a ingerir diàriament una quantitat d’aigua equivalent al 4-10% del seu pes corporal; en canvi, quan viu en aigües dolces la ingestió d’aigua és menyspreable. Determinades espècies modifiquen la funció renal; en aquest sentit disminueixen la filtració i simultàniament incrementen la reabsorció d’aigua. Les espècies estacionals i les estuarianes són les que han de suportar canvis més bruscos, pel fet de viure en àrees d’estuari on les variacions ambientals succeeixen d’una forma ràpida i al llarg d’un àmbit força ampli. Aquest és el cas de la llissa llobarrera (Mugil cephalus) i del fartet (Aphanius iberus), per citar-ne alguns exemples.
L’aparell reproductor dels peixos
Les gònades i l’aparell reproductor en els peixos condrictis (Scyliorhinus canicula) i osteïctis (Leuciscus cephalus). 1 ovari, 2 obertura de l’oviducte, 3 glàndula nidamental o secretora de la càpsula de l’ou, 4 oviducte, 5 ronyó, 6 conducte urinari, 7 si uro-genital, 8 testicle, 9 conducte eferent, 10 epidídim, 11 espermiducte o conducte de Wolff, 12 vesícula seminal, 13 sac espermàtic, 14 papil·la urogenital, 15 conductes urinaris secundaris, 16 conducte urinari principal, 17 bufeta natatòria.
Amadeu Blasco, a partir de materials diversos.
Les gònades són unes estructures de morfologia més o menys allargada emplaçades a la zona dorsal de la cavitat abdominal del cos. Ja hem comentat l’estreta relació d’aquestes estructures amb els ronyons, sobretot pel que fa a determinats grups d’àgnats i de pisciformes poc evolucionats.
La gònada dels àgnats és ben simple i imparella; en aquest cas és suspesa del mesenteri i se situa al mateix centre de la cavitat del cos. Concretament en els mixínids el testicle és irregular, lobulat i mancat d’espermiducte, i l’ovari tampoc no té oviducte; consegüentment, els productes genitals (esperma i òvuls) són lliurats dins la pròpia cavitat abdominal i han de sortir a l’exterior per mitjà d’un porus abdominal. Les llamprees també presenten una gònada imparella que en l’estat adult gairebé ocupa tota la cavitat abdominal fins al punt que pot arribar a abolir la funcionalitat de l’aparell digestiu; el sistema de reproducció és similar, però la dimensió dels ous és més reduïda i la sortida a l’exterior es realitza a través d’un parell de porus abdominals situats a l’extrem d’una papil·la uro-genital.
En els elasmobranquis la situació és ben diferent, perquè hi ha un aparell reproductor ben constituït. El dels mascles consta d’un parell de testicles ben desenvolupats que ocupen la part anterior de la cavitat abdominal i s’uneixen a la paret dorsal mitjançant unes bandes de teixit. Alhora, els testicles es comuniquen amb la part anterior del ronyó, anomenada també òrgan de Leydig (de funció reproductora), per mitjà dels conductes eferents. A la primera porció, aquests conductes eferents constitueixen l’epidídim i a la part posterior s’engruixeixen per a originar els conductes deferents, situats a la part ventral. La porció posterior s’eixampla i forma la vesícula seminal, que presenta un sac espermàtic adherit en posició dorsal; és en aquest darrer que s’acumula l’esperma. Les vesícules i els sacs espermàtics desemboquen a la cloaca a través de la papil·la uro-genital; des d’aquesta cavitat l’esperma passa als pterigopodis o apèndixs copuladors originats per una modificació de les aletes pelvianes. Els ovaris de les femelles són dobles, si bé l’esquerre pot ésser de dimensions més reduïdes i l’emplaçament és força similar al dels testicles. Pel que fa a les espècies ovípares, els òvuls es desprenen a la cavitat abdominal i es dirigeixen vers la glàndula nidamental (secretora de la càpsula de l’ou) a través del vestíbul de l’oviducte, on es produeix la fecundació; a continuació, l’ou, encapsulat i proveït d’una coberta còrnia, circula pels respectius oviductes i surt a l’exterior per una obertura comuna. En les espècies vivípares s’observa que els nous individus es desenvolupen a la part posterior de l’oviducte.
Els holocèfals (quimeres) mostren determinades modificacions que afecten els conductes eferents, certament molt nombrosos. A més, l’estructura de la vesícula seminal apareix compartimentada. D’altra banda, les femelles presenten dos oviductes que s’obren aïlladament a l’exterior, fet que constitueix una veritable excepció dins els pisciformes.
En els peixos ossis els testicles són unes masses allargades més o menys cintiformes i de coloració blanquinosa que s’emplacen a ambdós costats de la bufeta natatòria. En les espècies més evolucionades no hi ha cap connexió entre l’aparell reproductor i el sistema excretor (bé que, en algunes, els espermiductes es reuneixen amb els conductes urinaris en un si uro-genital). Els ovaris d’aquests peixos més evolucionats també ofereixen un aspecte allargat i cilíndric; els oviductes, que són curts, s’hi inclouen a l’extrem posterior. De vegades els oviductes d’ambdós ovaris es fusionen i s’obren a l’exterior a través d’una obertura comuna, també anomenada papil·la. No cal dir que hi ha tota una gradació de modificacions, fruit dels diferents estadis evolutius. En els peixos ossis més primitius els ous es disposen a la cavitat abdominal i els oviductes no hi són inclosos; en els teleostis, en canvi, l’ovari constitueix una estructura ben integrada i tancada.