La reproducció dels peixos
Potser una de les característiques més notables dels pisciformes és que gaudeixen d’una elevada fecunditat, cosa que es posa de manifest en una massiva producció d’ous i d’embrions. Nogensmenys la fecunditat oscil·la d’uns grups als altres; l’àmbit de variació se situa entre uns quants ous, en el cas de Latimeria, i diversos milions, com s’observa en els teleostis marins, per exemple, el bot (Mola mola). Els peixos ofereixen diferents estratègies reproductives, cadascuna caracteritzada per determinades adaptacions morfològiques, per processos fisiològies, pel comportament i l’hàbitat. Totes aquestes estratègies són orientades a preservar l’espècie, bé produint una gran quantitat d’ous i emprant diversos mecanismes de protecció de la posta o de les cries, o bé assegurant la supervivència dels nous individus per mitjà d’altres mètodes.
El període de reproducció és limitat temporalment, però pot oscil·lar entre uns pocs dies i alguns mesos, segons les espècies. Gran part dels peixos es reprodueixen a la primavera i a l’estiu, però tampoc no manquen les espècies que ho fan en altres èpoques de l’any. L’inici de la reproducció apareix com una resposta hormonal per part de l’organisme enfront de diversos mecanismes ambientals, principalment la temperatura de l’aigua i el fotoperíode o, dit d’una altra manera, la durada relativa del dia. Aquests mecanismes són els que actuen com a desencadenants del procés; nogensmenys, en determinats casos hi intervenen d’altres factors, com ara la intensitat del flux d’aigua (per exemple als rius) i el grau de salinització.
El procés fisiològic de la reproducció s’origina a l’hipotàlem, que activa la segregació d’hormones gonadòtropes. Aquestes hormones són les responsables del desenvolupament i del control de l’activitat ovàrica i testicular, tot estimulant la producció d’ovòcits i d’esperma. En canvi, els ovaris i els testicles s’encarreguen de la secreció d’hormones sexuals femenines (estrògens) i masculines (andrògens), respectivament. Així mateix, controlen la regulació dels caràcters sexuals secundaris que fan un paper força important dins el comportament reproductiu de les espècies.
Els tipus de reproducció
Hom descriu tres models bàsics de reproducció: la bisexual o gonocòrica, l’hermafrodita i la gimnogenètica.
La reproducció gonocòrica és la més comuna entre els àgnats i els pisciformes; aquest tipus de reproducció parteix de l’existència de dos sexes diferenciats, i del concurs d’ambdós gàmetes (òvul i espermatozoide) per a la formació del zigot o òvul fecundat, el desenvolupament del qual dona l’embrió.
L’hermafroditisme rep la denominació de sincrònic quan una porció de la gònada actua com a ovari i una altra com a testicle d’una manera simultània malgrat que sol haver-hi la participació d’un altre individu i l’intercanvi dels seus productes sexuals; en canvi, l’hermafroditisme seqüencial és proteràndric quan primer maduren les gònades masculines i proterogínic quan s’inverteix l’ordre de maduració. Si bé l’hermafroditisme no és usual en algun grup de peixos ossis, com ara els ostariofisis d’aigua dolça, és freqüent en els peixos marins (actinopterigis). L’estratègia reproductiva sincrònica resulta avantatjosa per a les espècies caracteritzades per la dispersió, és a dir, amb una baixa densitat poblacional o amb dificultats d’encontre durant el període de reproducció. Per això és freqüent entre els peixos abissals i mesopelàgics (alepisàurids, cloroftàlmids i estomiàtids); tanmateix hom també la troba en algunes famílies de peixos demersals i litorals, com ara els serrànids i els làbrids. L’hermafroditisme seqüencial és una estratègia que permet d’assegurar una adequada proporció de sexes; consegüentment constitueix una manera de garantir l’èxit reproductiu i també és important dins el comportament sexual d’algunes espècies que es reprodueixen en grup. Aquest sistema de reproducció és típic de les espècies batipelàgiques (estòmids) o litorals (serrànids, espàrids i làbrids).
La gimnogènesi, a diferència dels altres mecanismes de reproducció, consisteix en la formació del zigot i de l’embrió sense el concurs del gàmeta masculí; per aquest sistema únicament s’originen generacions de femelles. Malgrat tot, es tracta d’una modalitat poc usual entre els peixos i només ha estat descrita en determinats grups, com ara els ciprinodòntids i els ciprínids, per exemple el carpí vermell (Carassius auratus).
Una immensa majoria de peixos són ovípars; això significa que produeixen ous que són expulsats a l’exterior durant la fresa o la posta. Contràriament, en d’altres l’embrió es desenvolupa a l’interior del cos i surt al medi en forma d’aleví, com és el cas de les espècies ovovivípares i vivípares.
L’oviparisme

L’oviparisme és la modalitat de reproducció més freqüent en els peixos ossis. La fotografia ens mostra una parella de truita comuna (Salmo trutta) en plena reproducció. Excaven un petit forat a la grava del fons d’un riu de muntanya i la femella (al davant, en la foto) i el mascle es col·loquen de costat, de manera que quan ella comença a expulsar els òvuls, sovint ajudada del mascle, que li pressiona la cavitat abdominal, ell els va fecundant; acabada la posta, la cobreixen fins al naixement dels petits.
Peter Parks / Oxford Scientific Films - Firo-Foto.
L’oviparisme, la modalitat més generalitzada de reproducció, és alhora extraordinàriament variable. Sense anar gaire més lluny, entre els àgnats hi ha diverses estratègies reproductives. Per exemple, els mixínids produeixen una escassa quantitat d’ous però de mida considerable, mentre que les llamprees paràsites en produeixen molts (entre 24 000 i 240 000 a Petromyzon marinus), tots de mida reduïda; això no obstant, les formes lliures, més petites, ponen entre 400 i 9000 ous. Entre les llamprees la mortalitat és molt elevada durant les primeres fases del desenvolupament i solament un 7,8% (com a màxim) d’ous arriben a la desclosa. Els elasmobranquis i els holocèfals presenten espècies ovípares i ovo-vivípares. Les ovípares ponen ous grossos i proveïts d’una càpsula còrnia que els protegeix.com s’esdevé amb les quimeres (holocèfals), les rajades (raids) i algunes famílies de selacis (taurons). Entre els peixos ossis es presenten totes les varietats de reproducció, però hom pot dir que hi dominen clarament les formes ovípares. La grandor i el nombre d’ous que ponen aquests peixos també és força variable; de fet això constitueix un reflex fidel de les innombrables adaptacions i estratègies d’aquest grup de peixos tan diversificat.
Hom pot agrupar les espècies ovípares d’acord amb les seves característiques ecomorfològiques i etològiques o comportamentals. Així, és possible d’observar diverses adaptacions dels ous i les larves en ambients ben diferents, com també tot un seguit d’adaptacions referides a la parada nupcial i a la cura parental de la posta i de les cries.
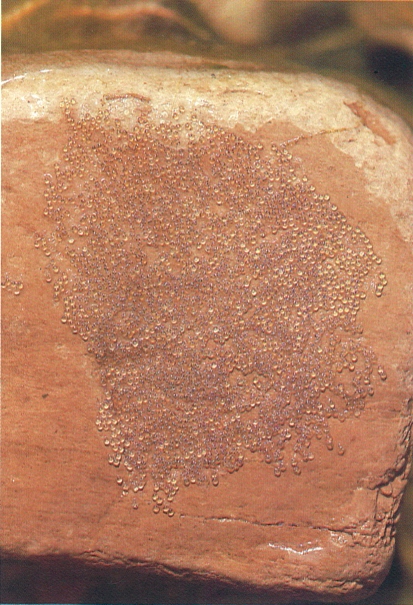
Posta de bavosa de riu (Blennius fluviatilis). Aquesta espècie té el costum d’excavar una petita cova sota una pedra i adherir-hi els ous per la cara interna. Després, el mascle vigila el niu durant tot el període d’incubació.
Adolf de Sostoa.
La majoria de peixos no protegeixen els ous ni la descendència i es limiten a deslliurar els ous a l’aigua, abandonats a llur sort. Algunes d’aquestes espècies efectuen la posta entre dues aigües o a la superfície, com fan els escòmbrids, com per exemple, la tonyina (Thunnus thynnus), i els clupèids, entre ells, la sardina (Sardina pilchardus). Els ous d’aquests peixos es mantenen en suspensió a l’aigua perquè contenen unes petites gotes lipídiques en el seu interior i és gràcies a aquest mecanisme que gaudeixen d’una gran capacitat de dispersió. En aquest cas les postes són constituïdes per una gran quantitat d’ous i la fecundació es realitza en massa: es barregen els ous i l’esperma lliurat al medi en grans quantitats. Una altra modalitat consisteix a dipositar els ous sobre diferents substrats de sorra, grava, pedres i roques, o bé sobre la vegetació aquàtica; la fresa es realitza en grup, i els ous bé romanen al fons o s’adhereixen al mateix substrat valent-se de determinades substàncies enganxoses, com s’observa en l’areng (Clupea harengus) i els ciprínids, com per exemple els barbs (Barbus) i les madrilles (Chondrostoma). Això no obstant, també hi ha peixos que es limiten a ocultar els ous, sense exercir cap mena de vigilància. Aquests solen excavar forats a la sorra o a la grava del substrat i hi enterren els ous; en són bons exemples el salmó (Salmo salar) i la truita (Salmo trutta). Altres espècies els dipositen en coves o bé a l’interior d’alguns invertebrats, com ara el ciprínid Rhodeus, que amb aquesta finalitat aprofita els mol·luscs bivalves d’aigua dolça.

Alguns dels sistemes de protecció de la posta són realment sofisticats, com el del cavallet de mar (Hippocampus ramulosus), en què el mascle la transporta durant tot el període d’incubació dins una bossa incubadora. La fotografia és feta just en el moment de néixer un aleví, que es pot veure encara amb la cua enrotllada.
Giuseppe Mazza.
El comportament d’alguns peixos durant l’època de reproducció és destinat a protegir els ous, els alevins o bé ambdues coses. N’hi ha que s’esmercen en la construcció de refugis o de nius més o menys perfeccionats. A aquests efectes utilitzen diferents tipus de substrats erosionats (sorra, grava, pedres, roques) o plantes aquàtiques. Els blènnids s’estimen més fer la fresa en cavitats o orificis i adhereixen els ous sobre el mateix substrat. Alguns peixos es valen de determinades substàncies per tal d’agregar els materials vegetals que conformen el niu, com ho fa el conegut espinós (Gasterosteus aculeatus). D’entre els que gaudeixen d’algun tipus de cura parental, n’hi ha que no construeixen un niu pròpiament dit, sinó que simplement adhereixen els ous a les roques o a les pedres (per exemple els gòbids o cabots); d’altres, els dipositen sobre plantes aquàtiques. Àdhuc n’hi ha que poden arribar a formar una massa flotant d’ous a la superfície de l’aigua, com els peixos asiàtics Anabas i Betta.
Dins d’aquest ampli espectre de modalitats reproductives, cal fer esment de les espècies que incuben els ous i els transporten en diferents parts del cos. Possiblement els més espectaculars són els incubadors bucals, que protegeixen els ous i les cries a l’interior de la boca. Així mateix, dins d’aquesta categoria també hi ha peixos que incuben els ous a les cambres branquials i d’altres, com els cavallets de mar (Hippocampus), que els duen (els mascles) a l’interior de la bossa incubadora, estructura d’origen cutani que té aquesta funció.
L’ovo-viviparisme i el viviparisme
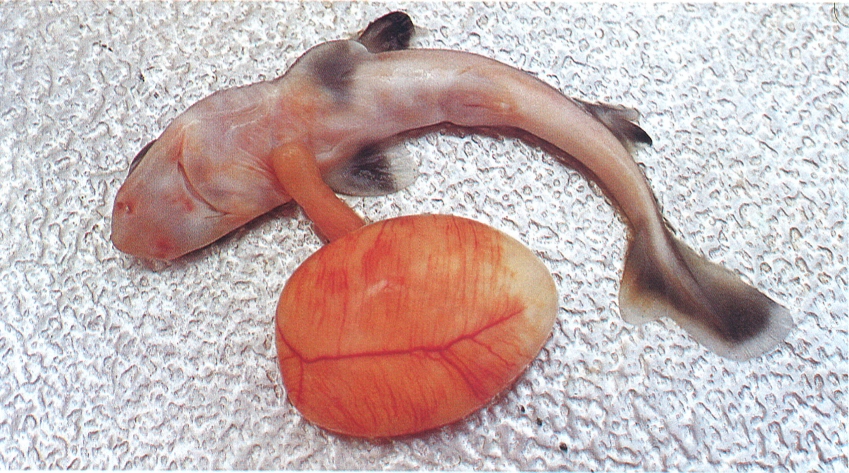
L’oviparisme i l’ovoviviparisme són modalitats de reproducció que presenten molts elasmobranquis. Durant el desenvolupament, aquestes espècies depenen de les reserves vitel·lines i, encara que alguns poden practicar l’oofàgia, d’altres es nodreixen a costa d’un gran sac vitel·lí, com el cas que recull la fotografia, corresponent a l’agullat xato (Squalus blainvillei).
Juan A. Moreno.
Un mecanisme eficaç de protecció de la filiada també consisteix a independitzar, si més no relativament, els ous i els embrions del medi extern i preservar-los, en conseqüència, dels canvis ambientals desfavorables, dels predadors, els paràsits i les malalties, entre d’altres. Endemés, aquestes estratègies reproductives permeten una reducció del nombre d’ous, fet que representa un estalvi energètic important per a l’organisme; d’altra banda aquests mecanismes garanteixen la fertilització, ja que la fecundació és interna, en contraposició al que succeeix a la majoria d’ovípars, que expulsen l’esperma en l’aigua i sobre els ous. Amb tot, els ovo-vivípars constitueixen una excepció dins el sistema; així, les quimeres, alguns selacis ovípars i determinats peixos ossis ponen ous però disposen d’un aparell copulador i la fecundació és interna. D’altra banda els anomenats ovo-vivípars incuben els ous dins la cavitat interna del cos i alliberen els alevins en el medi quan estan ben desenvolupats. Malgrat tot, a diferència dels vivípars, la mare no subministra cap mena d’aliment durant el desenvolupament embrionari. Aquesta forma de reproducció és força usual entre els selacis o taurons, com els bocadolços (Hexanchus) o els agullats (Squalus), i també en el celacant (Latimeria chalumnae).
Els peixos vivípars són aquells que produeixen embrions que neixen vius i que durant el període d’incubació es nodreixen, parcialment o totalment, de la mare. Els vivípars presenten múltiples adaptacions entre les quals cal destacar l’existència d’una pseudoplacenta; és a dir, l’alimentació suplementària per mitjà d’unes secrecions produïdes per la pròpia mare. Hom coneix diversos exemples de selacis vivípars com la tintorera (Prionace glauca) i la mussola vera (Mustelus mustelus) i d’entre els peixos ossis potser un dels més populars és el de la gambúsia (Gambusia affinis), que durant el període de reproducció arriba a originar entre 40 i 200 alevins cada vint i tants dies.
El dimorfisme sexual

El dimorfisme sexual pel color (dicromatisme) es manifesta d’una manera molt espectacular en alguns grups de peixos, per exemple en els blènnids, els ciprinodòntids i els làbrids. Les diferències entre mascles i femelles poden ésser permanents, com en el cas de l’espècie de làbrid que il·lustren les fotografies, Ctenolabrus rupestris, en la qual el mascle (a dalt) és més vistent que la femella (a baix). Però moltes vegades mascles i femelles són pràcticament iguals tot l’any excepte durant l’època de la reproducció, moment en què els mascles desenvolupen uns determinats caràcters externs destinats a cridar l’atenció de la femella.
Antonio Cruz.
És ben conegut el fet que la reproducció comporta l’aparició d’estructures internes diferents en cadascun dels sexes, tot i que des de l’òptica de la morfologia externa sovint resulta difícil de distingir-ho. Tanmateix, a vegades les diferències sexuals es palesen d’una forma aparent i constitueixen l’anomenat dimorfisme sexual, que pren el nom de dicromatisme quan es refereix a les diferències de coloració entre ambdós sexes. Aquestes diferències externes poden presentar-se d’una manera bé transitòria o estacional; en aquest cas només es manifesten durant el període de reproducció i, consegüentment, formen part del comportament reproductor. Però també poden ésser el resultat de determinades adaptacions morfològiques permanents, en gran part dels casos destinades a facilitar la còpula, l’oviposició o fresa i la incubació. D’altra banda, ocasionalment els caràcters sexuals secundaris constitueixen una forma d’identificació sexual o bé estan integrats dins el comportament nupcial de l’espècie.
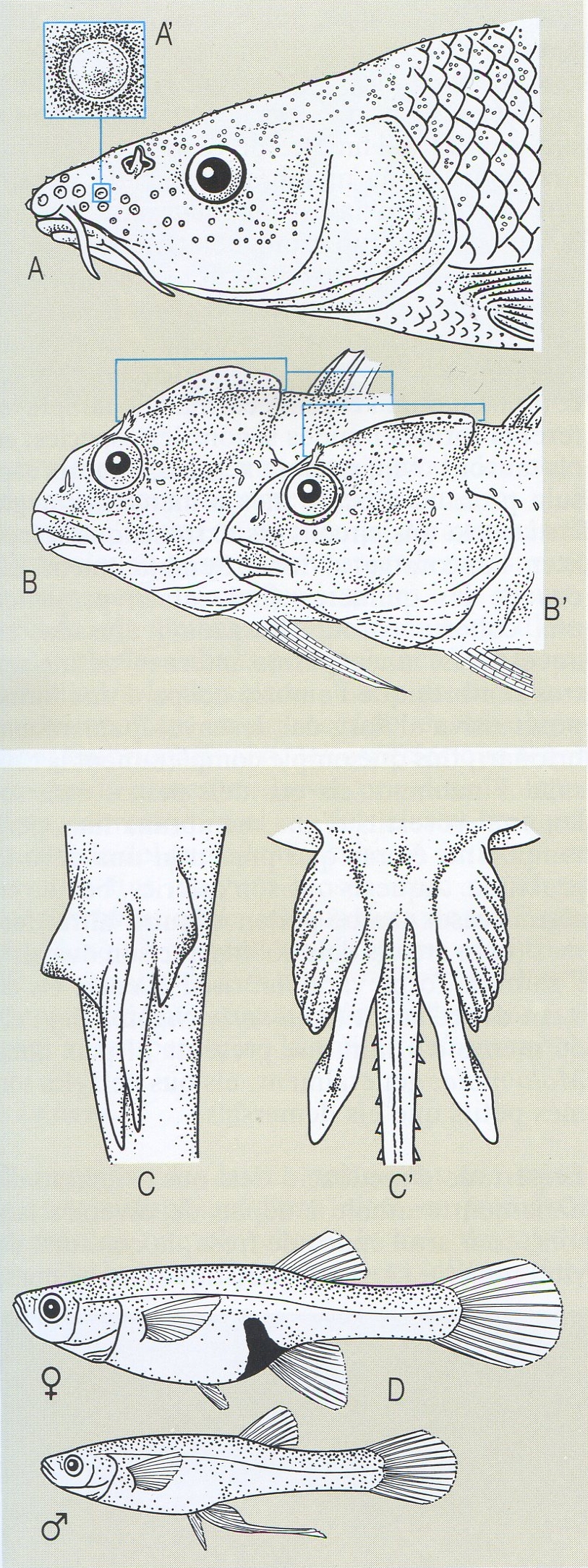
Dimorfisme sexual secundari (a dalt) i òrgans copuladors (a baix) en els peixos. A tubercles nupcials al cap dels barbs (Barbus), i detall d’un tubercle (A’). B mascle de bavosa de riu (Blennius fluviatilis) en l’època nupcial (B’ en l’època de repòs), que mostra el gran desenvolupament de la cresta. C òrgans copuladors o pterigopodis de tauró (Scyliorhinus); C’ organs copuladors o pterigopodis de rajada (Raja). D parella de gambúsies (Gambusia affinis); es pot observar la peculiaritat de l’aleta anal del mascle, transformada en un òrgan copulador o gonopodi.
Román Montull, del natural i altres fonts
Els caràcters dimòrfics transitoris ofereixen un ventall força ampli de variacions dins els peixos. D’una banda consisteixen en canvis de coloració associats al període reproductor; en aquest cas gairebé sempre és el mascle que llueix unes coloracions llampants i brillants per tal d’atreure la femella, dissuadir altres mascles o bé marcar el territori (així s’observa en els blènnids i els tripterígids). Els canvis morfològics també són força diversos; en alguns peixos, determinades parts del cos augmenten de mida (per exemple les aletes) i en d’altres adopten formes diferents, com ara les mandíbules del salmó (Salmo salar), que s’arquegen i es revesteixen de dents fortes d’una forma espectacular. Entre els ciprínids és força usual l’aparició dels denominats tubercles nupcials, que no són sinó unes excrescències cutànies que cobreixen diferents parts del cos. En d’altres casos es desenvolupen apèndixs o estructures especials, com ara la cresta nupcial de la bavosa de riu (Blennius fluviatilis) o la bossa incubadora dels singnàtids, que és una estructura d’origen dèrmic.
És força freqüent que els individus d’ambdós sexes siguin de dimensions diferents. Generalment s’observa que les femelles són més grosses que els mascles, fet que resulta avantatjós des del punt de vista funcional, perquè els proporciona una capacitat més gran de produir ous. Hi ha adaptacions relacionades amb la còpula i amb la fecundació interna, com els pterigopodis dels elasmobranquis i el gonopodi dels peixos ossis com ara la gambúsia (Gambusia affinis). Aquests òrgans copuladors s’originen per una modificació de les aletes pelvianes, en el cas dels elasmobranquis; el gonopodi, en canvi, procedeix d’una adaptació de l’aleta anal. Tampoc no és inusual de trobar situacions en què el dimorfisme sexual afecti la forma i la mida de les aletes.
El desenvolupament
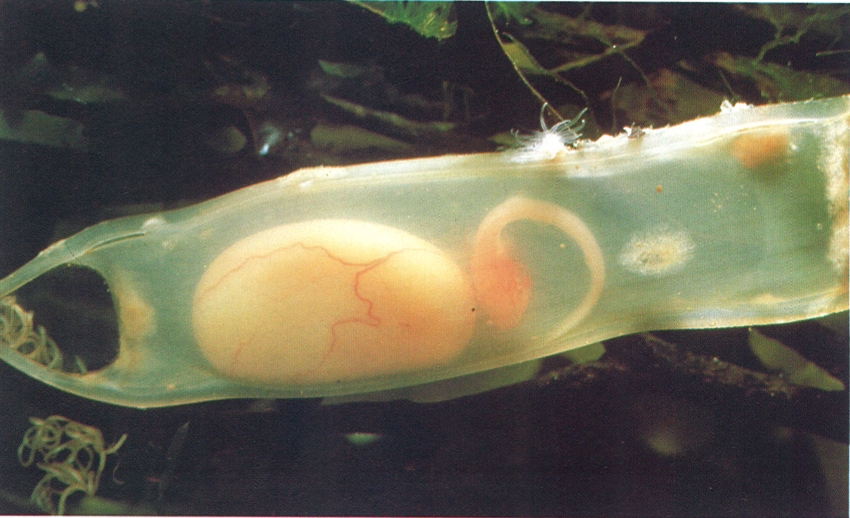
Els ous dels elasmobranquis són protegits per una càpsula còrnia. En el cas del gat (Scyliorhinus), són de forma allargada i presenten als extrems uns llargs circells que serveixen per a fixar-se al substrat. En la fotografia es distingeix, per transparència, l’embrió unit a un sac vitel·lí enorme.
G.I. Bernard/Oxford Scientific Films - Firo-Foto.
Hom entén el desenvolupament com el procés que s’esdevé des del moment que es forma l’ou o zigot, com a resultat de la fecundació de l’òvul per un espermatozoide, fins que l’individu assoleix la maduresa sexual, és a dir, fins que es converteix en adult. El procés de la fecundació es realitza en l’aigua o en la cavitat corporal, segons si es tracta d’espècies ovo-vivípares o vivípares, respectivament. Els espermatozoides, que són mòbils i força més petits que l’òvul, s’hi situen a l’entorn però solament n’hi ha un que reïx a penetrar-hi per un orifici minúscul anomenat micròpil, emplaçat en un dels pols. En realitat els espermatozoides que es troben en estat de repòs dins la gònada s’activen en contacte amb l’aigua i inicien una frenètica carrera que clourà amb la fecundació de l’òvul. L’activitat dels espermatozoides té una durada molt breu perquè disposen de poques reserves energètiques, però oscil·la entre alguns segons (30 segons en la truita, Salmo trutta) i escassos minuts. En aquest espai tan curt de temps han de realitzar la fecundació, perquè endemés l’òvul també s’activa amb l’aigua i transcorreguts alguns minuts es tanca el micròpil; en aquest sentit, doncs, no és estrany que els peixos fabriquin elevades quantitats de productes sexuals.
Els òvuls dels peixos són telolecítics, és a dir, presenten el vitel més concentrat en un dels pols. La quantitat de vitel és variable entre els diferents grups d’àgnats i de pisciformes; a més, aquest factor condiciona el tipus de divisió o, més ben dit, de segmentació de l’ou. La segmentació holoblàstica o total és pròpia dels ous proveïts d’un elevat contingut de vitel i en què el zigot es divideix íntegrament; aquest tipus de segmentació s’observa en les llamprees, els esturions i els peixos pulmonats. Tanmateix, gran part de les espècies (mixínids, condrictis i osteïctis) la presenten del tipus meroblàstic o parcial, que es limita a la segmentació d’un dels pols (pol animal) i deixa una gran part de l’ou sense segmentar. Nogensmenys, algunes espècies resulten difícils d’enquadrar en un d’aquests models i presenten una forma de segmentació intermèdia.
L’ou
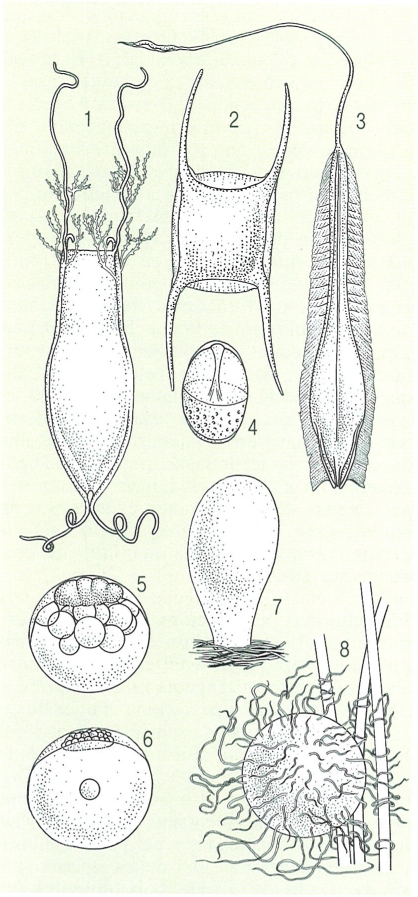
Tipus d’ous de peixos, 1 càpsula amb ou de tauró (Scyliorhinus), 2 càpsula amb ou de rajada (Raja), 3 ou de quimeriforme (Hydrolagus), 4 ou de seitó (Engraulis encrasicolus), 5 ou de moll o roger (Mullus), 6 ous de pedàs (Bothus), 7 ou de gòbit (Gobius), 8 ou de peix teleosti amb filament per a adherir-se a la vegetació.
Román Montull, a partir de fonts diverses.
Immediatament després de la fecundació s’inicia el procés de la segmentació, durant el qual l’ou primer es divideix en dues cèl·lules, després en quatre, després en vuit i així successivament fins a originar la blàstula, que, posteriorment i mitjançant el procés de la gastrulació, dona lloc a la gàstrula. Durant la gastrulació s’origina una estructura amb dues capes de cèl·lules, l’externa o epiblast i la interna o hipoblast. Posteriorment la gàstrula es va allargant i s’hi van diferenciant les distintes àrees que originaran els diversos teixits i òrgans de l’embrió; en aquest darrer cas el procés s’anomena organogènesi, que és el que, en definitiva, completa l’individu.
Els ous dels àgnats són força diferents si es comparen els de les llamprees (cefalaspidomorfs) amb els dels mixínids (pteraspidormofs): en el primer cas són molt petits (només fan un mil·límetre de diàmetre), esfèrics i revestits d’una membrana delicada, mentre que en els mixínids són allargats, fan uns 30 mm de longitud i romanen tancats dins una càpsula còrnia proveïda de filaments amb ganxos que els permeten d’ancorar-se entre ells o bé al substrat. Pel que fa als elasmobranquis, un cop els òvuls són fecundats es recobreixen d’una càpsula còrnia que adopta diferents formes i mides segons les espècies; en els taurons ovípars, per exemple, aquesta càpsula pren un aspecte més o menys rectangular i porta uns circells als extrems que li faciliten l’ancoratge al fons. Semblantment, els ous de les quimeres (holocèfals) presenten una coberta còrnia allargada i afusada; en aquest cas, unes parets internes defineixen tres cambres que l’embrió ocupa d’una forma progressiva al llarg del desenvolupament embrionari, fins que omple completament la càpsula. Finalment, els ous dels peixos ossis solen anar revestits d’una membrana fina, però tampoc no és rar que presentin una coberta gruixuda. Aquests ous són esfèrics, bé que en alguns casos també poden adquirir altres formes (com ara els dels gòbids que són ovalats), i de mida força variable: oscil·la entre els de 9 cm del celacant (Latimeria chalumnae) i els de menys d’1 mm que presenta el peix lluna Mola mola; generalment, els ous pelàgics són més petits que els demersals.
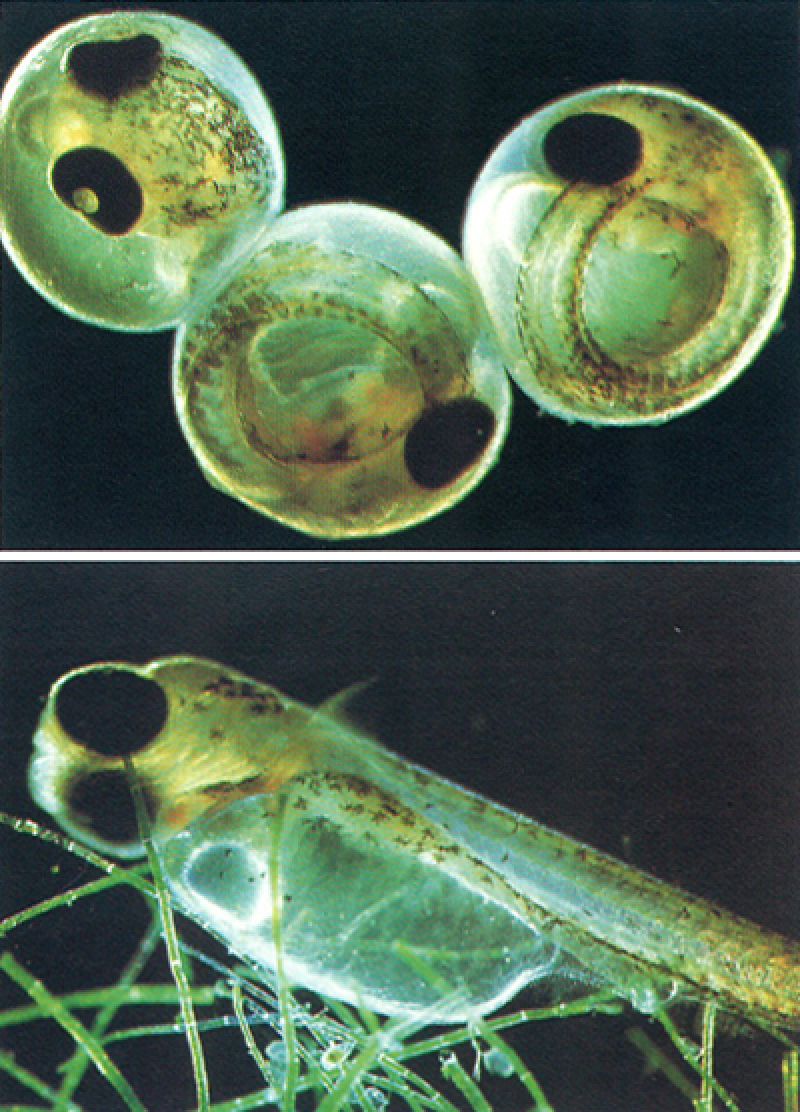
Els ous dels peixos ossis són generalment esfèrics i translúcids, com s’observa en la fotografia de dalt, en què es mostra una sèrie d’ous d’espinós (Gasterosteus aculeatus) en un estadi molt avançat de desenvolupament, amb els ulls i el cos perfectament conformats. A baix veiem una larva acabada de néixer de la mateixa espècie, que en les primeres etapes de la seva vida es nodrirà a partir del sac vitel·lí, cosa que limitarà en gran manera la seva mobilitat: ha de restar al fons o sobre la vegetació fins a atènyer un grau més avançat de desenvolupament.
David Thompson / Oxford Scientific Films - Firo-Fito.
El període d’incubació dels ous és extraordinàriament variable i depèn de diversos factors, com ara l’època de fresa, la quantitat de vitel nutritiu i les característiques ambientals, entre d’altres. En una gran part de les espècies, el desenvolupament embrionari és molt complet i al cap de poc temps d’haver nascut els alevins ja s’assemblen força als pares, però en altres casos, com les anguiles i els pleuronèctids, els alevins sofreixen una sèrie d’etapes posteriors abans no assoleixen una forma semblant a la dels pares. La mida de l’ou és força important en el procés del desenvolupament embrionari; així, les espècies que tenen ous petits presenten larves poc desenvolupades i viceversa. En el cas dels elasmobranquis la durada del període d’incubació és llarga, de vegades fins d’alguns mesos, com en el gat (Scyliorhinus canícula) que dura de 6 a 7 mesos. Això no obstant, en els peixos ossis l’àmbit de variació és més ampli: entre 2 i 4 dies la carpa (Cyprinus carpio) i de 2 a 4 mesos (segons la temperatura) la truita (Salmo trutta).
La larva
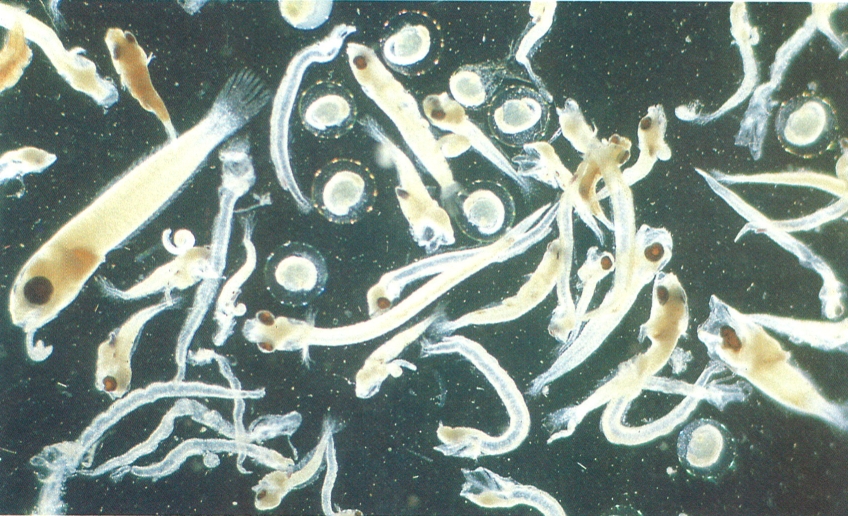
La varietat de tipus d’ous i de larves que viuen en suspensió en la massa d’aigua oceànica és considerable, com s’observa en aquesta fotografia, d’una mostra d’ictioplàncton de la mar catalana. L’ictioplàncton es captura amb una xarxa de plàncton de malla de 300 a 500 microns, i fent pesques ascendents (verticals o obliques) a una velocitat de 10 a 20 m/min.
Anna Sabatés.
Normalment les larves pelàgiques són transparents i disposen d’una aleta en forma de membrana que els recorre la línia dorsal del cos. De bon principi la boca no sol ésser funcional, i les larves han de nodrir-se del sac vitel·lí, que apareix com una expansió ventral. Al llarg d’aquesta primera etapa es va conformant l’obertura bucal, que a causa de la seva mida reduïda limita l’alimentació de la larva a preses o partícules de dimensions també molt petites. Moltes d’aquestes larves gaudeixen d’unes determinades característiques que els garanteixen la flotabilitat i que alhora els permeten de derivar amb els corrents marins: per exemple, petites gotes d’oli, un elevat contingut d’aigua en els teixits (com ara els leptocèfals de les anguiles) o encara unes determinades adaptacions morfològiques; efectivament, algunes larves tenen unes estructures força espectaculars, que desapareixen en l’estat adult, com ara les llargues aletes i els filaments de la larva del rap (Lophius piscatorius) o les espines i els apèndixs de les dels tetraodontiformes, com el bot (Mola mola).
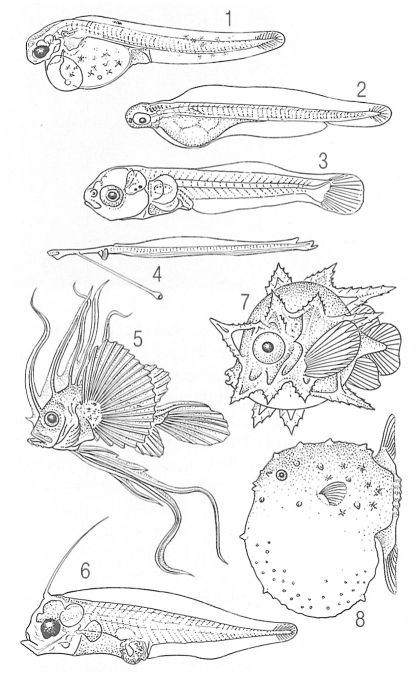
Tipus de larves de peixos ossis, per il·lustrar la gran varietat de formes que poden presentar Corresponen a les espècies següents: 1 Stizostedion vitreum, 2 seitó (Engraulis encrasicolus), 3 roger o moll (Mullus barbatus), 4 Idiacanthus fasciola, 5 rap (Lophius piscatorius), 6 pedàs (Bothus), 7 Ranzanía laevis, 8 bot (Mola mola).
Román Montull, a partir de fonts diverses.
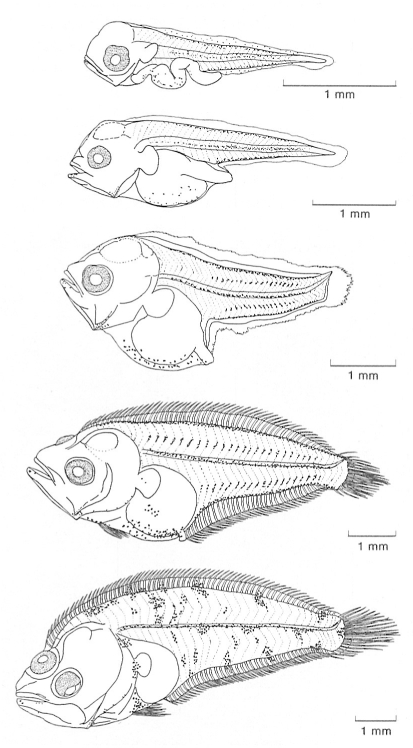
Desenvolupament larval d’un peix pla, la palaia vera (Citharus linguatula). S’han representat els diferents estadis per observar la progressiva migració de l’ull, en aquest cas de la dreta, cap a un costat del cos, l’anomenat zenital o pigmentat, fenomen que caracteritza tots els pleuronectiformes.
Anna Sabatés.
Alguns ictiòlegs parlen d’un estadi ulterior en el desenvolupament larval dels peixos, anomenat estadi postlarval, durant el qual desapareix completament el sac vitel·lí i la larva es troba en un avançat estadi de desenvolupament, si bé aquest estadi és força difícil de precisar en algunes espècies. Finalment el desenvolupament, i segons els casos la metamorfosi de la larva, acaba quan aquesta adquireix la condició de juvenil, moment a partir del qual ja ha adoptat l’aspecte extern de l’adult, s’inicia el període de creixement i s’acaba per assolir la maduresa sexual.
El creixement
Una de les característiques biològiques dels peixos que els fa ben diferents de la resta de vertebrats és la seva capacitat de creixement continu al llarg de tota la vida; aquest creixement solament se supedita a la disponibilitat d’una bona i adequada font d’aliment. Consegüentment, els individus d’una espècie concreta que tenen la mateixa edat comparteixen una mida similar i, d’altra banda, les diferències importants de mida s’observen entre individus d’edats i sexes diferents. Això fa que el creixement constitueixi un dels factors més emprats com a indicador del grau de salut de les poblacions; en aquest sentit l’estudi del creixement permet de conèixer moltes de les peculiaritats que s’han esdevingut durant la vida d’un individu.
Hom pot dir, d’una manera senzilla, que una taxa ràpida de creixement indica abundor d’aliment i condicions de vida favorables. Contràriament, un creixement lent fa pensar en una manca de recursos o en l’escassa salut d’una població, ocasionada per d’altres factors. Tanmateix, el ritme de creixement d’una espècie no és constant ni de bon tros, sinó que al llarg d’un mateix cicle anual manifesta períodes favorables i períodes desfavorables, estretament relacionats amb la temperatura de l’aigua, ja que, com a animals poiquiloterms o de sang freda, la temperatura exterior els condiciona l’activitat metabòlica. D’altra banda, la taxa de creixement baixa a mesura que transcorre la vida de l’individu.
L’edat dels peixos és relativament fàcil de determinar, si més no pel que fa als de les regions fredes i temperades, on les diferències entre períodes freds i càlids es manifesten en una sèrie de marques o de franges que es visualitzen en determinades estructures externes esquelètiques: les escates, els otòlits, els radis de les aletes i determinats ossos. Així, de primavera a tardor el peix creix d’una manera ràpida i, en conseqüència, apareix una franja ampla en les esmentades estructures; contràriament, durant l’hivern aquesta franja o anell es fa estreta i apareix com una marca fosca. D’aquesta manera, mitjançant el simple comptatge d’aquestes franges més fosques es pot determinar l’edat d’un exemplar. Endemés, per aquest procediment hom pot reconstruir la història d’un peix i deduir el creixement que va experimentar en èpoques anteriors, com també d’altres aspectes relacionats amb el seu cicle biològic, com ara la reproducció i les migracions, per citar-ne alguns exemples. La longevitat màxima dels peixos és difícil de conèixer en estat natural però sembla que algunes espècies, com la carpa (Cyprinus carpio), poden assolir, en condicions naturals, 25 anys, i més de 50 en captivitat. En general les espècies que viuen més anys també assoleixen mides relativament més grans i pesos més elevats que les de vida curta. D’altra banda l’inici de la maduresa sexual també té relació amb la longevitat. Així, les espècies de vida més efímera són més precoces i es reprodueixen entre 6 mesos, o menys, i dos anys de vida. Entre sexes també s’observen diferències importants; les femelles són més longeves i acostumen a assolir mides i pesos més grans, fet que suposa un avantatge adaptatiu que comporta una producció més gran d’ous.
El creixement dels peixos és alterat per determinats factors biòtics i ambientals. Entre els biòtics cal citar els que afecten les relacions interspecífiques i les intraspecífiques; ho és la competència pels recursos alimentaris, però també ho són les relacions socials entre els individus d’algunes espècies concretes. Entre els factors ambientals, el més important és, indubtablement, la temperatura; de fet, per a cada espècie hi ha un àmbit de variació òptim de temperatura dins el qual el creixement és màxim, i fora del qual, per sota o per sobre, el creixement disminueix. Semblantment, alguns factors dependents de la temperatura també afecten el creixement; per exemple el contingut d’oxigen de l’aigua, que és força important en el cas de la truita (Salmo trutta). En aquest mateix sentit alguns productes d’excreció, com ara l’amoni, poden resultar nocius a determinades concentracions, especialment quan es troben en forma de nitrits. Un altre dels factors que pot modificar el creixement dels peixos són els canvis de salinitat, perquè afecten l’equilibri osmòtic de les espècies estenohalines.
El comportament i les relacions socials

Les relacions entre diferents espècies de peixos inclouen l’anomenat comportament netejador (desparasitador), que practiquen diverses espècies marines, sobretot a les mars tropicals, però també a la Mediterrània: el tord cua-negre (Symphodus melanocercus), que veiem en la fotografia desparasitant una salpa (Sarpa salpa), és un bon exemple d’aquest comportament, beneficiós per a ambdós.
Jean-Georges Harmelin.
El comportament dels peixos pot definir-se com una resposta a les interaccions que s’originen entre ells mateixos o amb relació als estímuls ambientals. Tanmateix, aquesta resposta depèn de les característiques particulars de cada espècie i concerneix un ventall ben ampli d’aspectes diferents. Davant la impossibilitat d’efectuar una revisió completa de totes les facetes del comportament dels peixos, ens cenyirem als aspectes més fonamentals, sobretot als que fan referència al comportament social, alimentari, reproductor i locomotor.
Per relacionar-se amb els individus de la seva mateixa espècie o bé amb els d’altres espècies, els peixos es valen de diversos senyals, marques i coloracions; això significa que el sentit de la vista és d’una importància crucial a l’hora de mantenir aquest comportament. Aquests senyals o semàfors, si bé poden ésser de caràcter permanent, també poden aparèixer només en determinades èpoques de l’any i formar part del comportament reproductor. D’altra banda, els colors i les taques que adopten els peixos poden ésser més o menys estables; així, hi ha la possibilitat que canvïin d’una manera ràpida i momentània i que s’origini un aclariment o un enfosquiment de l’individu de resultes d’una agressió, d’una motivació sexual o del comportament alimentari. A més d’exhibir diverses coloracions, que en alguns casos es presenten en unes combinacions realment espectaculars, els peixos també solen presentar taques de formes molt característiques: ocels, franges, línies i punteigs, entre d’altres. Tota aquesta ornamentació sol anar acompanyada d’una sèrie de moviments i de positures, particularment importants durant la parada nupcial. Tanmateix, hi ha peixos que es valen d’altres sentits per entaular relació amb els altres individus: n’hi ha que generen sons o vibracions, però també n’hi ha que, a aquests mateixos efectes, produeixen impulsos elèctrics (els gimnòtids i els mormírids, entre d’altres). Paral·lelament s’ha detectat l’existència de determinats senyals químics en el comportament social, com ara les feromones o d’altres substàncies advertidores, que possibiliten el mutu reconeixement entre els individus d’una mateixa espècie, o que, d’altra banda, permeten detectar estats d’alarma, com és el cas de la substància de Schreckstoff secretada pel barb roig (Phoxinus phoxinus).
Els recursos de comunicació i defensa
La coloració
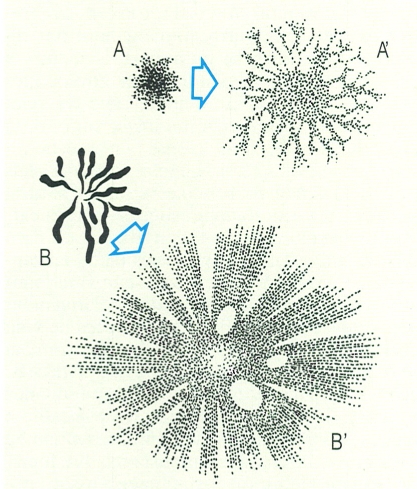
Els colors dels peixos són conseqüència de dos efectes diferents. Els tons argentats són produïts per efectes òptics sobre unes cèl·lules determinades, mentre que els altres colors es produeixen gràcies a l’existència de cèl·lules proveïdes de pigments. En el dibuix s’han representat dues modalitats de cromatòfors: de tipus dendrític, amb el pigment agregat (A) o dispers (A’), i de tipus radial, amb el pigment agregat (B) o dispers (B’).
Román Montull, a partir de C.E. Bond; David Thompson i Chris Catton
La immensa varietat de colors que poden arribar a exhibir els peixos i la gran riquesa de gammes, potser només comparable a la dels ocells (dins els vertebrats), és un dels aspectes que han cridat més l’atenció d’experts i profans. L’exuberància de tonalitats i de formes fa difícil una sistematització completa de totes les possibles combinacions i variacions, que abasten des dels ocels fins a les franges i àdhuc tota una sèrie de taques i dissenys senzills o bigarrats. Els patrons de coloració poden relacionar-se amb l’hàbitat del peix; en aquest sentit constitueixen mecanismes de camuflatge o de cripsi, de mimetisme i d’advertiment. Però, així mateix, la coloració també pot formar part del comportament social i reproductor d’una espècie.
La majoria de peixos tenen alguna mena de pigmentació a la pell, malgrat que no sempre correspon a un patró conspicu. Els pigments es disposen en cèl·lules especials que reben la denominació del pigment o del color que porten. Dos tipus de cèl·lules són responsables de la coloració dels peixos: les unes, els cromatòfors, són les que contenen els pigments (biòcromes); les altres, els esquematòcits o iridiòcits, presenten colors estructurals físics. En aquest darrer cas el color és el producte de reflexió de la llum per determinades estructures argentades que actuen com un mirall, a causa de les substàncies que contenen les cèl·lules dels teixits. Els cromatòfors s’emplacen sota la pell, especialment al derma, però també poden trobar-se a l’epidermis o bé en determinats òrgans i teixits (com ara els ulls, el peritoneu, etc). D’altra banda, els components orgànics responsables del color són diversos i complexos, com la melanina, la flavina, la porfirina, els carotens, les pterines i la purina, per citar-ne alguns exemples. Entre els cromatòfors hi ha els melanòfors, és a dir, els que contenen pigment negre o melanina, que són els més generalitzats; els eritròfors presenten pigments vermells i ataronjats, a causa dels carotens i altres compostos; els xantòfors són de color groc gràcies a la presència de carotens grocs; els leucòfors, de color blanc, contenen purines i guanina. Tanmateix, sovint hi ha cèl·lules amb més d’un pigment, i en una gran part dels casos la coloració resulta de la combinació de les diferents cèl·lules pigmentàries o cromatòfors. Pel que fa als esquematòcits o iridiòcits, contenen cristalls de guanina i de purina, i constitueixen l’estrat argentí, car reflecteixen un 80% de la llum i són els que proporcionen l’aparença de coloració arjentada als peixos.

Els mecanismes de defensa d’algunes espècies arriben a un grau de sofisticació tan gran com el que il·lustra la fotografia: un cavallet de mar australià (Phyllopteryx foliatus) que combina la coloració i la forma per tal de confondre’s amb les algues del seu ambient.
AGE Fotostock.
Els peixos tenen la capacitat de modificar llur coloració d’una manera ràpida o també per períodes més llargs de temps. Els pigments dels cromatòfors es disposen en forma de grànuls que poden modificar llur distribució espacial a l’interior de la cèl·lula; aquesta particularitat es deu a l’existència d’un procés fisiològic a partir del qual aquestes cèl·lules es concentren formant un punt o bé s’expandeixen tot adoptant una forma ramificada; d’aquesta manera, quan els pigments es concentren a la porció central de la cèl·lula, el peix adopta una coloració pàl·lida, i, en canvi, quan els grànuls s’expandeixen, l’àrea acolorida augmenta de mida i la coloració es fa més intensa. Per mitjà de la combinació de diferents tipus de cromatòfors, aquest procés permet de modificar la coloració, a més de la intensitat. El mecanisme de control dels cromatòfors, que és el responsable dels canvis de coloració, és hormonal però amb la intervenció del sistema nerviós; en aquest sentit, cal assenyalar que l’adrenalina regula la concentració o l’expansió dels melanòfors.
La funció primordial de la coloració dels peixos és la comunicació amb els individus de la mateixa espècie, o amb els d’altres espècies, per tal d’establir-hi unes relacions socials. D’altra banda, però, també pot formar part dels caràcters sexuals secundaris durant l’època de reproducció, o, també, pot constituir un mecanisme de protecció enfront dels depredadors; en aquest darrer cas la coloració pot ésser una forma d’advertir el depredador de la verinositat pròpia o bé pot ésser una disfressa (mimetisme). Tanmateix, la coloració té un significat adaptatiu amb relació a la protecció, com passa amb la pigmentació que hi ha al voltant de la medul·la espinal de les larves transparents dels peixos, o respecte a la seva funció de la termoregulació.
Força sovint la coloració es relaciona amb l’hàbitat de l’animal. Així, trobem que gran part de les espècies que viuen entre dues aigües exhibeixen un contraombrejat aparent, que consisteix a tenir la part dorsal fosca i la ventral argentina; aquest fet s’observa clarament en la tintorera (Prionace glauca), el verat (Scomber scombrus) i l’agulla (Belone belone), per exemple. D’aquesta manera, mirat des de dalt, el color del peix és fosc i es confon amb el del fons; contràriament, quan el miren els peixos que neden per sota, el seu ventre argentí es confon amb el reflex de la llum que ve de la superfície. Aquesta característica els permet de passar desapercebuts, tant per a ocultar-se davant dels seus depredadors com per aproximar-se a les seves preses. Les espècies bentòniques poden adoptar coloracions críptiques per tal d’assemblar-se al substrat o a la vegetació: així s’observa en els pleuronèctids o peixos plans, les escórpores (Scorpaena), els blènnids o bavoses, els gòbids o burrets i els singnàtids o agulletes; en una gran part dels casos, els canvis de coloració es produeixen d’una manera ràpida i la similitud amb el medi és realment sorprenent.

La cripsi és un mecanisme que permet a un gran nombre d’espècies de peixos bentònics de passar desapercebudes en fer-se confusibles amb el substrat o amb la vegetació. En la fotografía superior només un ull denuncia la presència d’una agulleta (Syngnathus typhle) perfectament dissimulada, perquè el contorn del seu cos es confon amb les fulles de posidònia, i n’imita fins i tot els epibionts. En la fotografia central s’intueix el cos d’una escórpora fosca (Scorpaena porcus), que resta immòbil al fons i adopta la mateixa coloració i aspecte que les algues fotòfiles. En la fotografia inferior, un pedàs (Bothus podas) mostra una homocromia absoluta amb el substrat sorrenc.
Adolf de Sostoa i Antonio Cruz.
Una altra manera de camuflar-se és adoptar una coloració disruptiva; és a dir, fer que la silueta del peix es confongui amb el fons o amb l’ambient. Això s’assoleix amb la presència de franges i de taques molt contrastades, que tendeixen a desfigurar la imatge i el perfil de l’animal. Gràcies a aquest mecanisme, molts peixos litorals reïxen a camuflar-se entre les branques dels coralls o entre les estructures d’altres organismes. En determinats casos la presència d’ocels o falsos ulls fa desviar l’atenció del depredador vers altres zones del cos menys vitals que els ulls veritables. D’altres vegades, els ulls queden dissimulats per mitjà de línies o de franges horitzontals o verticals que els travessen i els oculten.
La coloració advertidora o conspicua representa també un mecanisme per a donar a conèixer el sexe de l’individu durant el període de reproducció, com és ben típic de l’espinós (Gasterosteus aculeatus), i també una tècnica d’atracció per a escollir la parella com passa amb els tripterígids i els làbrids. De vegades les franges i les taques tenen un significat especial en la formació de les moles de peixos, car els ajuda a orientar-se i a coordinar-se els uns amb els altres; així s’observa en el joell o peix sense sang (Atherina boyeri) i en la madrilla (Chondrostoma toxostoma).
Determinades coloracions brillants o conspícues adverteixen també sobre un perill determinat i això és freqüent en els peixos verinosos, com el peix escorpí (Pterois volitans), que viu als esculls coral·lins; aquest mecanisme, però, també pot denotar un caràcter tòxic o no comestible.
La combinació de coloració i forma d’una determinada disfressa fa que el depredador confongui l’animal amb qualsevol altre objecte sense interès; potser aquest és un dels sistemes més eficaços de protecció i un dels que gaudeix de més avantatges adaptatius. Per exemple, pot consistir a adoptar forma de fulla (Monocirrhus polycanthus, de l’Amèrica del Sud) o d’alga, com ara l’espectacular cavallet de mar australià (Phyllopteryx foliatus), que s’hi confon perfectament tant pel que fa a la forma com pel color. Finalment, hi ha un altre mètode que fa confondre els altres peixos: el mimetisme. En aquest cas l’animal adopta la forma i el color d’altres espècies considerades perilloses o verinoses; d’aquesta manera s’aprofita de la mala fama. Però també pot tractar-se d’un mimetisme agressiu com el que manifesta el blènnid Aspidontus taeniatus, que adopta el mateix aspecte i comportament del làbrid netejador (Labroides dimidiatus), que s’alimenta d’ectoparàsits d’altres peixos i aconsegueix d’aquesta manera mossegar i arrencar trossos de pell als peixos que, confiats, els confonen.
La bioluminescència
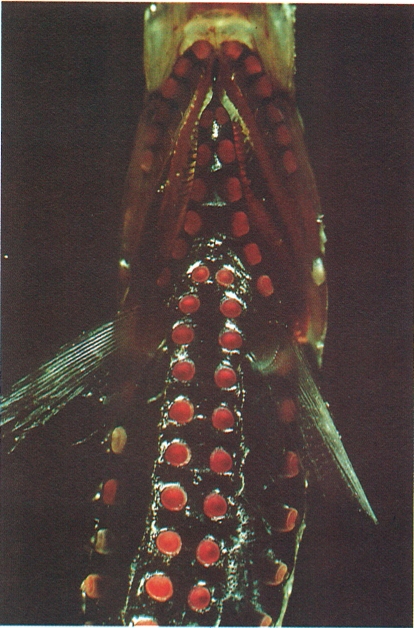
Els fotòfors dels peixos tenen la missió de produir llum en els ambients on no arriben les radiacions lumíniques solars. Són molt freqüents en moltes de les espècies mesopelàgiques, com en la destral d’argent (Argyropelecus hemigymnus), en què apareixen una mena de botons vermells distribuïts per tota la regió ventral.
Peter Parks / Oxford Scientific Films - Firo-Foto.
Alguns peixos, com molts altres organismes, tenen la capacitat de produir llum. Ho fan per dues vies diferents. La primera, de tipus químic, es produeix per la interacció de dos productes, l’enzim luciferasa i la substància luciferina, la combinació dels quals desencadena una reacció lluminosa a l’interior d’uns òrgans especials que s’anomenen fotòfors. Un altre mètode consisteix a utilitzar bacteris que viuen en simbiosi amb el peix; aquests bacteris infecten l’individu en la fase larval i viuen a l’interior d’una estructura en forma de glàndula. Gran part dels peixos que disposen d’un mecanisme bioluminescent viuen a les grans profunditats marines de la zona mesopelàgica, entre els 300 i els 1000 m, és a dir, a la zona de penombra. Aquesta facultat tan peculiar és compartida per un grup relativament nombrós d’espècies, que pertanyen a 42 famílies i que comprenen del 10 al 15% dels gèneres de peixos marins, entre els quals dominen els teleostis; en canvi, només hi ha dos gèneres d’elasmobranquis capaços de produir llum. Els més representatius entre els teleostis, cal cercar-los en els ordres dels salmoniformes, els mictofiformes, els bericiformes, els perciformes i els lofiformes.
Els fotòfors amb fotòcits, de reacció química, són freqüents en els mictofiformes; aquests fotòfors es distribueixen pel cos i pel cap dels peixos seguint una sèrie de línies, malgrat que també poden presentar-se d’una manera aïllada. Generalment es disposen a la regió ventral i tant el seu nombre com la seva disposició són emprats com a caràcter sistemàtic. Un fotòfor típic consta d’una lent rudimentària, de la glàndula o fotòcit i d’una capa interna reflectora (generalment de guanina). Entorn d’aquest patró estructural hi ha una certa variabilitat morfològica que depèn del lloc d’emplaçament; així, els ventrals tenen forma de tub i proporcionen una il·luminació més puntiforme i, en canvi, els dorsals adopten una disposició d’àmplies cambres dotades de determinades finestres per on es projecta la llum vers l’exterior. El color de la llum abasta des del blau verdós fins al vermell, passant pel groc, segons el tipus de pigment que presenten i la coloració de la lent, que en el cas dels peixos destral (Argyropelecus) és d’un color vermellós, molt semblant al de la llum a baixa intensitat.
Els fotòfors amb bacteris simbionts solen produir una lluminositat espectacular, com ara els anomalòpids de la mar Roja (Photoblepharon) a causa de l’enorme quantitat de bacteris que s’hi arriben a allotjar. Aquest tipus d’estructures es troba associat a l’anus, a la regió ventral de l’aparell digestiu o bé al cap; per exemple, sovint apareixen a la regió anal d’algunes espècies de macrúrids, gàdids i traquíctids. En d’altres casos són situats a l’esòfag i projecten la llum envers la bufeta natatoria o la musculatura, de manera que la claror es translluu a través dels teixits. Determinats peixos, com ara els anomalòpids (propis d’aigües litorals tropicals), presenten grans fotòfors sota els ulls i poden modificar la il·luminació tot i amagar l’òrgan lluminós, fent-lo rodar dins una fosseta pigmentada de negre, la qual impedeix la sortida de la llum.
La funció dels fotòfors és la comunicació social entre els individus de la mateixa espècie en un hàbitat sovint caracteritzat per una escassa lluminositat ambiental, bé per a reconèixer-los o amb finalitat sexual, ja que en diversos casos mascles i femelles tenen uns patrons de distribució dels fotòfors ben diferents. Altres vegades representa una forma d’eludir els depredadors, com per exemple quan els fotòfors, situats ventralment, projecten la llum cap avall i produeixen el mateix efecte que els iridiòcits, és a dir, eviten el contraombrejat des de sota i es fan, en certa manera, invisibles. També tenen una funció advertidora per tal de posar a l’aguait els possibles depredadors del perill que corren, per exemple simulant la possessió d’òrgans verinosos. Finalment, poden emprar-se per a atreure les preses (en aquest cas es tracta de fotòfors disposats prop la boca o a les barbes sensorials), o tan sols per a poder-les localitzar en la penombra, si els fotòfors es troben al cap.
L’emissió i la captació de sons
Els peixos tenen la capacitat d’emetre diferents tipus de sons, per mitjà d’òrgans o estructures diversos, com els radis i les espines, les dents, la boca, la bufeta natatòria i d’altres estructures annexes, a més de l’aparell digestiu. L’aigua constitueix un bon medi per a la transmissió del so, que s’hi propaga a velocitats de fins 1500 m/s; en conseqüència, no és estrany que un considerable nombre d’espècies utilitzi normalment aquesta peculiaritat com un mètode de comunicació i que, alhora, aquests organismes hagin desenvolupat determinades adaptacions per tal de detectarhi els sons. La producció de so també afecta, des d’una òptica àmplia, determinades activitats rutinàries, com ara la natació, els moviments de salt, l’alimentació i la respiració. Això no obstant, únicament farem referència als aspectes que tenen un significat adaptatiu.
Són força importants els sons produïts per estridulació de les peces dentals o per les dents faríngies, amplificats en aquest cas per la bufeta natatòria. Així mateix, cal considerar els sons originats pels radis de les aletes, com passa per exemple, amb l’espinós (Gasterosteus aculeatus), o bé per altres parts de l’esquelet, com ho fa el cavallet de mar (Hippocampus ramulosus). En diverses espècies la bufeta natatòria actua com un òrgan de ressonància; les vibracions (de fins 400 Hz de freqüència) es produeixen gràcies a una sèrie de músculs especials que s’insereixen a la seva paret, com és el cas de les lluernes (Trigla) i del gall (Zeus faber). Aquests músculs també es poden localitzar en algun dels extrems de la bufeta natatòria o bé poden actuar d’una manera indirecta per l’efecte del moviment dels radis o d’estructures associades a les primeres vèrtebres, que romanen en contacte amb la paret de la bufeta.
Sembla que l’emissió de sons té una funció relacionada amb la defensa del territori, si bé també pot constituir un so d’alarma en resposta a l’atac d’un depredador. Amb tot, també té un paper important dins el comportament social i reproductor d’algunes espècies de peixos; per exemple, el cavallet de mar (Hippocampus) i els gòbids o burrets.
L’electricitat
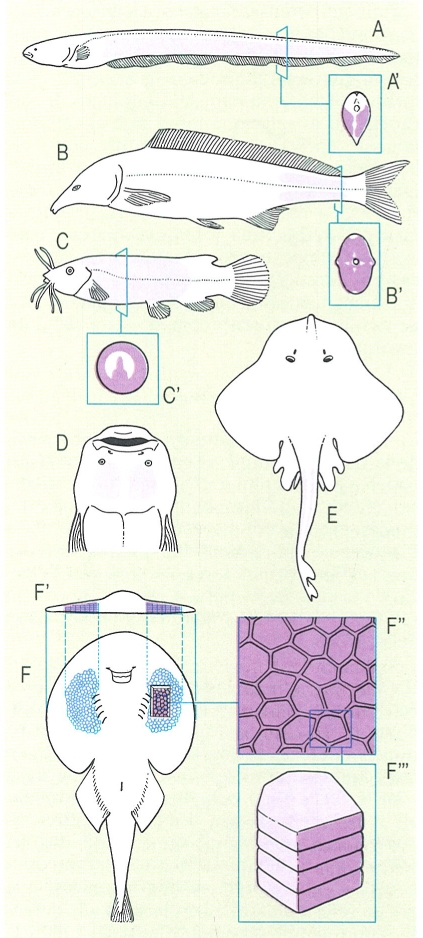
La possessió d’òrgans elèctrics, no gaire generalitzada entre els peixos de les nostres latituds, és un altre sistema de comunicar-se i relacionar-se amb l’ambient i els altres organismes. En el dibuix s’ha representat la posició dels òrgans elèctrics en diversos pisciformes: A anguila elèctrica (Electrophorus electricus), amb una secció del cos per mostrar la disposició dels òrgans elèctrics (A’), B i B’ mormírid (Ghathonemus), C i C’ peix gat elèctric (Malapterurus electricus), D rata de mar exòtica (Uranoscopus), E rajada (Raja), i F tremolosa negrosa (Torpedo nobiliaria), amb detalls de la disposició dels òrgans elèctrics en secció transversal (F’), i de la forma dels electròcits, vistos per sobre (F") i de costat (F’").
Román Montull, a partir de fonts diverses.
La producció d’electricitat suposa l’existència d’uns òrgans elèctrics que deriven de les fibres nervioses o bé són fibres musculars modificades, o plaques motores. Aquests òrgans són característics d’alguns elasmobranquis (tremoloses i rajades), i també d’alguns peixos ossis (l’anguila elèctrica, el peix gat elèctric, alguns uranoscòpids exòtics i els peixos elefant d’Àfrica).
Les tremoloses (Torpedo), que colonitzen els fons de la plataforma continental, tenen els òrgans elèctrics situats a ambdós costats del cap; els òrgans elèctrics d’aquests peixos són integrats per una sèrie de plaques elèctriques (electròcits) apilades les unes sobre les altres, i formen unes columnes prismàtiques paral·leles, separades entre elles per teixit conjuntiu. Cadascuna d’aquestes plaques, constituïda per diferents capes de cèl·lules, s’innerva per la part externa (capa elèctrica). Els òrgans elèctrics de les tremoloses poden originar descàrregues de fins 220 V, en direcció dorsiventral, com per exemple la tremolosa negrosa (Torpedo nobiliaria). D’altres elasmobranquis, com les rajades (Raja), poden arribar a produir descàrregues elèctriques, bé que de baixa potència. Una de les espècies més temibles és l’anguila elèctrica (Electrophorus electricus), que viu a la conca amazònica i pot arribar a originar descàrregues de fins a 650 V, malgrat que les que genera normalment se situen entre els 300 i els 400 V. En aquesta espècie les plaques elèctriques formen columnes longitudinals al llarg de la part ventral del cos i la direcció del corrent elèctric discorre des de la cua fins al cap. Alguns peixos presenten òrgans elèctrics a la regió caudal del cos, com ara els peixos elefant d’Àfrica (mormírids). Però tampoc no manquen espècies, com alguns uranoscòpids (Astroscopus), que els tenen emplaçats al cap, darrere els ulls. Finalment, els òrgans elèctrics del peix gat elèctric (Malapterurus electricus) es localitzen a la regió mitjana del cos i són capaços d’originar potencials considerables.
Una de les funcions dels òrgans elèctrics és de servir de sistema d’electrolocació, és a dir, per a la localització de tota mena d’objectes durant la nit o en aigües tèrboles, que és on habiten els peixos elèctrics d’aigua dolça (mormírids, gimnàrquids, gimnòtids i electrofòrids), per mitjà de la producció de camps elèctrics. Aquest mecanisme també és emprat per a detectar els altres individus de la mateixa espècie, ja que la freqüència d’ona és característica de cada espècie (per exemple en les rajades). Finalment, les descàrregues elèctriques poden constituir un sistema de defensa o bé ésser utilitzades en la captura de preses.
La producció de verí
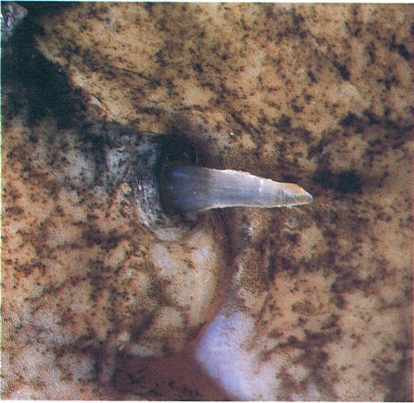
Les espines verinoses són freqüents en diverses famílies d’elasmobranquis i d’osteïctis. En molts casos hi ha glàndules de verí secretores de toxines, capaces de produir transtorns greus en els humans; en d’altres, les toxines són produïdes per un epiteli glandular. En la fotografia veiem l’espina opercular de la rata (Uranoscopus scaber), que produeix un cert efecte tòxic, no comparable al d’altres famílies, com els escorpènids (les escórpores) o els traquínids (les aranyes).
Antonio Cruz.
Gran part de les glàndules verinoses que presenten els peixos deriven de l’epidermis, com una modificació de les glàndules mucoses. Aquests òrgans són capaços de provocar transtorns greus o intoxicacions, en cas que el peix sigui ingerit per un altre organisme, fins i tot per l’home. Quan les toxines són injectades poden produir dolors seriosos i fins i tot la mort. La funció d’aquests òrgans pot ésser tant ofensiva com defensiva.
Els elasmobranquis exhibeixen tota mena d’espines verinoses proveïdes d’un epiteli glandular tòxic; aquest és el cas de les espines dorsals de taurons com l’agullat ver (Squalus acanthias), o de les espines caudals de les escurçanes (Dasyatis), de les milanes (Myliobatis) i de les quimeres (Chimaera). Alguns peixos ossis, com ara els escorpènids (Scorpaena), tenen a les aletes dorsal, anal i pelviana unes estries per on discorren les toxines produïdes per les glàndules verinoses. Les aranyes (Trachinus) tenen glàndules verinoses associades a les espines operculars i als radis espinosos de l’aleta dorsal; pel que fa als darrers, el verí és força potent i desencadena uns efectes tant neurotòxics com hemotòxics.
Hi ha un tipus d’intoxicació, que rep el nom de «ciguatera», produïda per més de tres-centes espècies de peixos marins tropicals o subtropicals. Aquest fenomen és imprevisible, ja que la seva toxicitat depèn del lloc i de l’estació de l’any. Altrament algunes espècies produeixen toxines a les gònades durant el període de reproducció, però tampoc no manquen peixos que les acumulen al fetge, l’intestí i la pell, independentment del seu estat sexual.
Jerarquia i territorialitat
Un aspecte força destacable del comportament d’alguns peixos és la possessió d’un territori o d’una àrea que és ocupat i defensat per un o per diversos individus d’una mateixa espècie; aquest territori és emprat exclusivament com a zona de cria, d’alimentació o de refugi. Els fenòmens de territorialitat comporten l’establiment d’una jerarquia segons la qual determinats individus exerceixen un paper de domini i uns altres de submissió. La jerarquia i la territorialitat també poden establir-se entre individus joves, com s’observa en els salmons del Pacífic; en aquest cas, hom observa diferències de creixement entre els individus nascuts al mateix any en diferents zones de la conca hidrogràfica. En alguns rius s’estableix una competència entre els mateixos joves que malden per ocupar determinades posicions favorables, per exemple als llocs on es canalitza el flux d’aigua i on, en conseqüència, resulta més fàcil de capturar els insectes que baixen arrossegats pel corrent (deriva). En alguns casos, fins i tot pot haver-hi un clar domini dels individus més forts i agressius, que ocupen uns territoris determinats i condemnen la resta de la població a d’altres llocs menys favorables, on forçosament el seu creixement s’endarrereix. Generalment tots aquests fenòmens poden tenir incidència en la regulació d’algunes poblacions de peixos. Hi ha espècies en les quals el comportament defensiu solament es desencadena contra els individus d’una altra espècie; però també n’hi ha d’altres que es mostren agressives a qualsevol espècie que gosi entrar a llur territori. Una gran part dels fenòmens de territorialitat es relaciona força amb el comportament reproductor (per exemple entre els blènnids i els gasterostèids, entre d’altres). Determinades famílies de peixos són territorialistes, però això no comporta, forçosament, d’establir una jerarquia aparent, com ara els pecílids, els centràrquids i els làbrids, entre d’altres. D’altra banda, s’ha comprovat que en alguns casos l’agressió s’origina per tal de defensar l’aliment, com si es tractés d’un territori.
El comportament reproductor

En el comportament reproductor de l’espinós (Gasterosteus aculeatus), el mascle construeix el niu, amb algues, sorra o fang, i hi duu la femella després d’efectuar una dansa nupcial estereotipada, per fer-hi la posta. En la fotografia superior, la femella és a l’interior del niu, fent-hi la posta, mentre que el mascle, a fora, lluu la seva característica lliurea nupcial, de tons vermellosos. A baix s’observa la posta, posada a l’interior del niu, que és vigilada pel mascle fins que els ous es desclouen.
David Thompson / Oxford Scientific Films - Firo-Foto.
La reproducció constitueix una de les facetes de la biologia en què el comportament té un paper preponderant i es manifesta de diverses maneres, àdhuc en diferents etapes del procés. Alguns dels aspectes més destacables d’aquest comportament fan referència a les migracions reproductores, la recerca i l’elecció de la parella, la parada nupcial, la còpula i la fresa, com també la defensa de la posta i dels alevins.
En algunes espècies la reproducció es realitza en col·lectivitat i, per tant, no hi ha elecció d’una parella concreta. Aquests peixos acostumen a efectuar migracions: s’agrupen en unes àrees determinades on fan tant la fresa com la fecundació dels ous, i posteriorment els abandonen a la sort. En canvi, d’altres espècies necessiten reconèixer els individus del sexe contrari i a aquest efecte observen unes pautes de comportament ben marcades, que poden referir-se tant a la selecció i a la defensa d’un territori apte per a la fresa, com a la construcció de nius o bé a d’altres mecanismes relacionats amb la fresa i amb les etapes posteriors.
Durant el període de reproducció la territorialitat es manifesta d’una manera molt acusada, sobretot en aquelles espècies que construeixen nius o que dipositen els ous en algun refugi, a redós dels depredadors. La majoria de les vegades el mascle és el responsable de la cura de la posta. Aquest comportament és força característic dels blènnids i d’altres grups de peixos que solen lluir coloracions brillants; aquests peixos realitzen tot un ritual per tal de dissuadir llurs enemics, a base d’adoptar diverses positures i d’efectuar moviments intimidatoris. Es tracta d’espècies guarnides amb tota mena de senyals visuals al cap i al cos, com ara ratlles, taques o ocels, i dotades de coloracions espectaculars que llueixen en desplegar les aletes bruscament. Alguns peixos es mostren més agressius i es llancen sobre les mandíbules dels seus adversaris, que sovint són d’altres mascles de la mateixa espècie, per tal d’engrescar-los en una lluita força peculiar. En el cas dels salmons, els combats entre els mascles poden arribar a ésser força violents; d’altra banda, aquests peixos presenten determinades modificacions, com ara la curvatura de les mandíbules i la presència de dents afilades i fortes, que els permeten de mossegar amb més facilitat tots aquells que gosen competir pel seu territori de fresa. Entre els anomenats peixos combatents (Betta splendens) la lluita pot arribar a comportar la mort del perdedor.
L’elecció de la parella pot dur-se a terme per diversos procediments. En alguns casos el reconeixement es produeix per la visualització de determinats senyals constituïts per colors conspicus, taques al cos i a les aletes o fins i tot pel llambreig dels fotòfors, la qual cosa és pròpia d’alguns peixos marins. Així mateix, en determinades espècies són importants els estímuls químics; a aquests efectes secreten unes substàncies les olors de les quals són reconegudes pels individus del sexe contrari. D’altres, es valen dels sons o els camps elèctrics.
La parada nupcial, que precedeix la fresa, pot consistir en el fet d’atreure la femella; en aquest sentit, el mascle adopta diferents posituras i empra senyals visuals. D’altres vegades consisteix en una autèntica dansa de passes estereotipades, com per exemple passa amb l’espinós (Gasterosteus aculeatus). La parada nupcial pot anar acompanyada de tota mena d’aproximacions, de contactes, de positures i de desplegaments, àdhuc de sons i d’olors que incitin a la còpula o la fresa.
La cura dels ous i dels alevins pels progenitors és un comportament força comú en bastants espècies de peixos. D’una banda pot consistir simplement en el fet de protegir o de cobrir els ous amb grava o pedres, com és corrent entre els salmònids i la llamprea de mar (Petromyzon marinus). En d’altres ocasions també comporta la vigilància i la protecció dels ous (els blènnids i l’espinós) o bé el fet de dur els ous a sobre, com fan els cavallets de mar (Hippocampus) i les agulletes de mar (Syngnathus). L’elecció del lloc de fresa i la construcció del niu és una activitat que realitza només el mascle o mascle i femella conjuntament. Després de la parada nupcial i de la construcció del niu, el mascle incita una o diverses femelles, tal com s’observa en els blènnids, per tal de conduir-les a l’interior del seu niu, on fan la fresa i es fecunden els ous.
D’una banda, el comportament reproductor proporciona l’avantatge de poder seleccionar els individus més aptes de cada població. De l’altra també protegeix la integritat de l’espècie, perquè les seves pautes de comportament són peculiars i, consegüentment, això redueix la probabilitat d’hibridació o d’encreuament, que portaria a la procreació d’individus estèrils. Endemés, els mecanismes de protecció de la posta i de les cries redueixen la mortalitat de la població. Finalment, cal esmentar que determinats mecanismes de comportament tendeixen a allunyar els adults dels llocs de fresa amb l’objecte de reduir el risc de predació.
Les relacions interspecífiques
Pel que fa a les relacions interspecífiques dels peixos, cal considerar que són força nombroses i a vegades s’estableixen entre individus corresponents a tipus biològics diferents.
Són bastant comuns els fenòmens de desparasitació de la boca, les brànquies i la pell; aquesta activitat, especialment freqüent en els peixos marins d’aigües tropicals (làbrids i gòbids), fa que, tant al moment de la neteja com després, els netejadors siguin reconeguts. Això fa que els peixos que practiquen aquesta activitat de desparasitació arribin a adquirir una certa immunitat enfront dels seus clients, que sovint són depredadors potencials. Aquestes tasques de neteja generalment solen realitzar-se en uns territoris fixos i comporten tot un codi de positures de l’individu que vol ésser desparasitat, com per exemple situar-se estàticament amb les aletes desplegades o bé amb la boca o els opercles oberts; tanmateix, aquest ofici no deixa d’ésser arriscat. Aquests peixos netejadors actuen en solitari o bé en petits grups. Com sol succeir en el cas de determinats mecanismes de simbiosi o de benefici mutu, sempre hi ha aquelles espècies mimètiques que malden per a aprofitar-se de la immunitat de les altres i que normalment són agredides pels veritables netejadors. En aquest sentit, és freqüent d’observar una determinada defensa del territori on poden exercir aquest ofici, de la mateixa manera que d’altres peixos defensen l’aliment.

Les rèmores (Remora i Echeneis) viatgen adherides al cos d’elasmobranquis, en aquest cas, una manta (Mobula mobular), i’ d’altres peixos pelàgics, gràcies a l’efecte d’una potent ventosa que tenen a la part dorsal del cos.
Neville Zell / Oxford Scientific Films - Firo-Foto.
És força conegut el cas de relació interspecífica de les rèmores (Remora); aquests peixos presenten l’aleta dorsal modificada en un òrgan de fixació que els permet d’adherir-se el cos d’altres peixos, taurons o d’altres organismes aquàtics. Aquest comportament, si més no, els ofereix la possibilitat de beneficiar-se dels trossos d’aliment que deixen els depredadors mentre mengen. D’altres, com el peix pilot o vairó (Naucrates ductor), aprofiten la protecció que els faciliten d’altres peixos més grans o els taurons.
En determinades ocasions s’associen espècies de peixos de diferent règim alimentari; així, hi ha serrànids que es col·loquen prop d’alguns peixos herbívors i es nodreixen a base dels crustacis i dels peixos que s’escapen. Hom coneix diversos tipus de relació que s’estableixen entre peixos i d’altres organismes aquàtics; en aquest sentit cal esmentar els casos de simbiosi entre crustacis i peixos, que els permet de compartir un mateix refugi (per exemple, entre algunes espècies de gòbids i de crustacis decàpodes). Així mateix és possible de trobar determinats crustacis que exerceixen l’ofici de netejadors d’alguns peixos.
El comportament locomotor i la formació de moles
Els peixos responen a diversos estímuls per orientar-se, efectuen diferents moviments i desplaçaments o bé s’agreguen. Entre aquests estímuls cal citar el fototactisme, o resposta a la llum, que és molt important en el cas dels peixos pelàgics diürns; aquests hi responen positivament i això explica que puguin sentir-se atrets per una font lluminosa durant la nit. Aquesta atracció és utilitzada pels pescadors professionals, per exemple en la pesca de la sardina (Sardina pilchardus) i del seitó (Engraulis encrasicolus), que reïxen a concentrar els peixos valent-se de llums artificials per tal d’envoltar la mola mitjançant les xarxes de cèrcol. Tanmateix, alguns peixos ofereixen una resposta negativa a la llum, especialment els d’hàbits nocturns, com el cavilat (Cottus gobio). La posició dels peixos en l’aigua és controlada per l’òrgan de l’equilibri, el laberint, però també hi intervé la il·luminació vertical sobre la regió dorsal del cos (geotactisme); d’aquesta manera, si es canvia l’orientació de la llum, el cos del peix s’inclina en la mateixa direcció. Un altre aspecte força interessant del comportament locomotor dels peixos és llur resposta quan són sotmesos a un camp elèctric, que fa que s’orientin i es desplacin vers el pol positiu del camp; aquesta peculiaritat ha induït al desenvolupament i la pràctica d’una tècnica de captura científica denominada pesca elèctrica, que constitueix un dels millors mètodes per a estudiar i quantificar les poblacions de peixos continentals. Determinades espècies, sobretot les bentòniques, tendeixen a sentir-se atretes per alguns substrats o ambients. D’altra banda, n’hi ha que necessiten la presència i el contacte d’altres individus de la mateixa espècie i s’atreuen entre ells. És ben coneguda la reacció d’alguns peixos als corrents d’aigua (reotactisme). Aquest estímul és particularment important en el cas de les espècies anàdromes i catàdromes, com ara el salmó (Salmo salar) i l’anguila (Anguilla anguilla), que s’orienten a contracorrent quan migren aigües amunt (en el primer dels casos els adults i en el segon els juvenils). Els estímuls químics (quimiotactisme) tan aviat poden ésser emprats en la localització de l’aliment com en la detecció dels individus de la mateixa espècie. Aquests estímuls també serveixen de sistema d’alarma (alguns ciprínids) i en algunes espècies migratòries són utilitzats per a orientar-se i per a reconèixer el lloc de procedència, com passa amb els salmons.

La formació de moles constitueix un comportament característic de moltes espècies pelàgiques de petites dimensions, com els clupèids, que s’hi disposen d’una manera ordenada, segons la mateixa orientació (moles polaritzades). Aquestes agrupacions responen a determinats estímuls o perills (per exemple, l’atac d’un depredador) d’una manera coordinada, com un tot.
Xavier Safont / M. Alba Camprubí.
Un dels aspectes del comportament social que hom pot observar fàcilment tant en aigües dolces com marines és, indubtablement, la formació de moles o grups de peixos. Els individus que constitueixen una mola o banc de peixos són regits per una atracció mútua que els fa agrupar tot formant una massa. En algunes espècies, com ara els clupèids, aquestes masses poden ésser de milions d’individus; els individus que integren la mola solen ésser de la mateixa mida i força sovint són joves. No s’ha de confondre aquest tipus de formacions amb les agrupacions que experimenten algunes espècies quan són atretes per un ambient determinat; en aquest sentit, a l’entorn dels esculls coral·lins es concentra una gran quantitat de peixos a la recerca de protecció i aliment.
Hom pot dir que hi ha dos tipus de moles, les polaritzades i les no polaritzades. Les moles polaritzades es caracteritzen pel fet que tots els individus que les conformen mantenen la mateixa orientació i una distribució ordenada, com també una velocitat de marxa uniforme i sincronitzada dins del grup. En canvi, en les moles no polaritzades els peixos es distribueixen d’una manera més o menys aleatòria i no mantenen una formació estricta. Algunes espècies observen unes pautes de comportament tan marcades que això les obliga a viure en moles, si més no a les primeres fases de vida, com s’observa en els clupèids, els engràulids, els escòmbrids, els aterínids i els mugílids, entre d’altres. La forma i la mida de les moles és variable. Pel que fa a la forma, sovint depèn de les activitats que realitzen els peixos i sol adoptar dissenys allargats, ovalats o quadrats; quan es tracta de peixos que efectuen moviments ràpids, les moles solen presentar forma de tascó. La mida del banc canvia en cada espècie; els més nombrosos són els d’areng (Clupea harengus) i d’altres clupèids com la sardina (Sardina püchardus) però tampoc no són menyspreables els de les llisses (Mugil i Liza). Els menys nombrosos potser són els que formen algunes espècies d’aigua dolça.
D’entre els òrgans dels sentits, sembla que la vista constitueix l’estímul més important i el que en definitiva permet als peixos de mantenir la formació estructurada de la mola. D’altra banda, hi ha d’altres característiques importants que els faciliten l’orientació: la forma del cos, la coloració o la presència de determinades marques visuals, com ara les franges fosques dels flancs, que, per cert, constitueixen uns excel·lents senyals de referència per als peixos. Els avantatges adaptatius de les moles són diversos, però es fonamenten en un fet tan senzill com cercar la companyia dels individus de la seva mateixa espècie. Un dels avantatges més importants és la disminució del risc de l’individu d’ésser atacat per espècies depredadores. Així mateix, el fet de viure en mola també facilita la recerca d’una font d’aliment apropiada, i no cal dir que representa una estratègia força encertada com a forma de reproducció, com és el cas dels peixos pelàgics. Darrerament hom ha pogut comprovar que aquest tipus de formacions també permeten d’augmentar l’eficiència hidrodinàmica.

L’efectivitat de les moles com a unitat estructural es fonamenta en la cohesió dels individus que les integren. Es tracta de constituir una agrupació harmoniosa, en la qual cada individu mantingui una relació proporcional respecte als que l’envolten. A les mars tropicals és relativament corrent d’observar prop del litoral formacions com la de la fotografia, compostes fonamentalment d’apogònids o aterínids.
Josep M. Gili.
Una mola de peixos es presenta, enfront dels possibles depredadors, com un gran organisme que resulta de l’adicció de tots els individus que la conformen. Aquest efecte, si més no, aconsegueix desconcertar el depredador, perquè se li fa difícil arribar a distingir els ulls o el cap d’un individu en concret i, en conseqüència, poder-lo atacar. D’altra banda, una bona coordinació de moviments també permet a la mola una resposta immediata des de qualsevol dels flancs i realitzar, per tant, tota mena de maniobres que poden ésser de fugida, envoltants o estratègiques per tal d’impedir o de dificultar l’atac del depredador. Però, amb tot, els peixos que formen moles són les preses preferides dels depredadors pelàgics que, per a capturar-los, es valen de diverses estratègies. Així, tant les tonyines (Thunnus) com els sorells (Trachurus) i altres caràngids ataquen les moles en grup, els produeixen pànic i aconsegueixen de disgregar-les parcialment per atacar-ne els components. Una altra de les estratègies consisteix a atacar-los quan decau la llum del dia i quan, en conseqüència, la cohesió del banc també minva. Un altre mètode de captura efectiu consisteix a realitzar salts o moviments bruscos que desconcertin la mola i aprofitar la confusió regnant.
Les moles de peixos són força eficients pel que fa a la recerca d’aliment; de fet, és més fàcil de localitzar-lo en grup, i sovint, mentre es nodreixen, adopten una formació circular. D’altra banda, la formació de moles facilita la reproducció dels peixos pelàgics: mascles i femelles es disposen en estrats i, en lliurar a l’aigua els seus productes sexuals, aquests es barregen i es troben més fàcilment. S’ha comprovat que aquest sistema de reproducció és d’una enorme eficiència a l’hora d’assegurar un èxit reproductiu més gran. Entre els avantatges adaptatius que ofereix aquesta forma tan peculiar d’agrupar-se, cal citar l’efecte hidrodinàmic, que es basa d’una banda en una disposició harmoniosa del peix, talment que formen diagonals i se separen entre ells d’una manera proporcional a la de llur cos. Amb aquest tipus de formació gairebé marcial, es reïx a originar un determinat corrent físic d’atracció que arrossega el peix que ve al darrere i que, d’alguna manera, produeix un efecte contrari al de la turbulència. A més, també disminueix l’efecte de fricció de l’aigua a causa de la segregació de determinades substàncies, com ara el mucus, que en aquest cas s’estén pel grup sencer. L’orientació també és més efectiva i permet una natació més ràpida de tot el conjunt. A més també disminueix la turbulència individual perquè, des d’un punt de vista hidrodinàmic, la mola es comporta com un únic cos amb propietats plàstiques i d’emmotllament. Normalment cal atribuir el canvi de direcció durant la marxa de la mola a l’acció concertada de tots els membres que la integren i sol iniciar-se al mateix centre de la mola.
A part les moles hi ha d’altres tipus d’agregacions en resposta a diverses circumstàncies que, si bé sovint es relacionen amb la recerca d’aliment, també poden referir-se a concentracions d’individus durant el període de fresa. Encara, en determinades èpoques de l’any poden produir-se acumulacions d’individus, com s’observa en les madrilles (Chondrostoma toxostoma), que constitueixen moles de centenars d’exemplars als rius mediterranis durant l’hivern. Aquestes moles solen romandre estàtiques, bé al fons dels tolls, sota pedres de grans dimensions o a l’interior de les coves formades a les vores del riu; conjuntament, s’hi solen trobar nombrosos barbs (Barbus) o bagres (Leuciscus).
Les migracions dels peixos

Les migracions dels salmònids, com el salmó (Salmo salar), tenen una periodicitat anual. Els salmons són espècies anàdromes, que passen gran part de la seva vida a la mar, però que han d’anar a fer la posta a les aigües dolces. Durant la tardor remunten els rius on van néixer, salvant tota mena d’obstacles, fins arribar als fons de sorra o grava adequats per a fer-hi la posta.
Zardoya Press.
Alguns peixos realitzen determinats moviments en massa, o migracions, que poden tenir un caràcter anual, estacional o periòdic. Cal diferenciar, però, entre les migracions a petita escala, com les que realitzen algunes espècies fluvials durant el període de fresa (ciprínids i salmònids), i les grans i espectaculars migracions d’alguns taurons, de les tonyines, els salmons i les anguiles, en què recorren milers de quilòmetres.

La migració de les espècies catàdromes, com l’anguila (Anguilla anguilla), es fa en sentit invers a la dels salmons (anàdroms). Les anguiles van a reproduir-se a la mar dels Sargassos i, en tornar, després de passar per canvis notables al llarg del viatge, entren en els rius en forma d’angules translúcides; després, emprenen una migració ascendent pels rius, que els obliga a superar tota mena d’obstacles, com assuts i preses.
Marcel·la Chinchilla.
Hi ha múltiples maneres de classificar les espècies migratòries; potser la més encertada és de dividir-les en tres grans grups: les diàdromes, les oceanòdromes i les potamòdromes. D’una banda les espècies diàdromes són aquelles que efectuen migracions entre la mar i les aigües dolces o continentals. Entre aquestes, hi ha les anàdromes, que passen una gran part de la seva vida a la mar i cerquen les aigües dolces per a reproduir-s’hi, les catàdromes, que realitzen el cicle invers, i les amfídromes, que alternen ambdós medis, sense una clara motivació de caràcter reproductiu. El segon gran grup d’espècies migradores, les oceanòdromes, són aquelles que, d’una manera exclusiva, només efectuen migracions a diversos indrets del medi marí. Finalment les potamòdromes sempre romanen a les aigües continentals, però realitzen desplaçaments massius, gairebé sempre amb finalitats reproductores.
La causa de les migracions i els mecanismes que utilitzen els peixos per tal d’arribar a determinats indrets d’encontre o per a orientar-se i trobar el camí de tornada és un dels aspectes de la vida dels peixos que suscita més gran interès. De fet, el motiu d’aquests desplaçaments té diversos orígens. D’una banda, pot relacionar-se amb la recerca d’una àrea adient per a reproduir-se, és a dir, la recerca d’individus sexualment madurs en l’indret i el període de l’any adequats; en el cas específic de les migracions reproductores, aquest comportament implica la recerca de zones adients per al desenvolupament dels ous i de les larves (és el cas de la truita, del salmó i d’altres peixos fluvials). D’altra banda, també pot referir-se a la localització de zones caracteritzades per una alta productivitat on troben aliment més abundant; sembla que, en alguns casos, com els peixos mesopelàgics, són freqüents les migracions verticals nocturnes que tenen com a objecte la recerca d’aliment. Pel que fa a les espècies pelàgiques, les migracions minven a més la possibilitat de competir pels recursos alimentaris entre els individus de la mateixa mida i edat, i també redueixen el canibalisme. Finalment, un altre motiu per a migrar és la recerca de zones adients de refugi i cria.
No cal dir que les migracions comporten també determinats desavantatges. Per exemple, requereixen una gran reserva energètica, factor que s’agreuja pel fet que mentre els peixos fan el trajecte de migració deixen d’alimentar-se. Així, per exemple, l’anguila (Anguilla anguilla) ha d’acumular una gran quantitat de greix, que després és emprat com a font d’energia durant el viatge i en la producció dels productes sexuals. Les migracions comporten encara el risc d’una gran pèrdua d’ous i larves per l’efecte d’alguns corrents oceànics, i també una exposició més gran als depredadors.
Les espècies anàdromes passen gran part de la seva etapa de creixement en aigües marines i penetren als rius per realitzar-hi la fresa; això s’observa en el salmó (Salmo salar), la truita marina (Salmo trutta trutta), la guerxa (Alosa alosa), la llamprea de mar (Petromyzon marinus) i l’esturió (Acipenser sturio). En canvi, les catàdromes, com ara l’anguila (Anguilla anguilla), viuen als rius i d’octubre a gener migren a la mar dels Sargassos on es reprodueixen. Les larves de les anguiles, també anomenades leptocèfals, sofreixen diverses transformacions fins a convertir-se en les delicades angules que, a la tardor i a l’hivern, després de tres anys de viatge, reïxen a remuntar els rius.
Entre els peixos que realitzen migracions marines, d’una banda cal citar els que només efectuen desplaçaments locals, com ara els que van de les zones profundes a les àrees més superficials. Els mictòfids o els gonostomàtids es mouen verticalment, uns 500 m o més, durant la nit; van des de la zona mesopelàgica, on habiten de dia, fins a la zona superficial, tot seguint els mateixos ritmes migratoris de les masses de zooplàncton de les quals s’alimenten. Altres peixos realitzen desplaçaments horitzontals, des del fons de la plataforma continental fins a la zona litoral, per a cercar-hi àrees de fresa; aquest és el cas del llobarro (Dicentrarchus labrax). Alguns peixos recorren distàncies més grans, com l’orada (Sparus aurata) o la sardina (Sardina pilchardus), i n’hi ha que fins recorren milers de quilòmetres, com diverses espècies de selacis o com les tonyines (Thunnus thynnus), per tal de cercar zones d’alimentació o de reproducció. L’areng (Clupea harengus) efectua migracions espectaculars a la recerca d’aigües més càlides on abundi el plàncton. Les espècies potamòdromes realitzen migracions a les aigües dolces, especialment per a cercar-hi un lloc que reuneixi els requeriments adequats per a la fresa; entre aquests peixos, n’hi ha que fan desplaçaments curts (alguns ciprínids i salmònids) i n’hi ha que remunten els llargs rius tropicals, com ara el «bocachico» (Prochilodus) de la regió neotropical.
Per a activar el comportament migratori, els peixos requereixen determinats estímuls ambientals, com és ara la temperatura, la llum, els corrents o la salinitat. En altres casos, com en les migracions reproductores, hi intervenen factors de tipus fisiològic, malgrat que també hi influeixin els estímuls ambientals o el comportament social. En llur periple migratori els peixos poden emprar diversos mecanismes de navegació, com l’orientació per mitjà de quimioceptors, és a dir, ensumant i reconeixent els llocs de procedència, com ho fan els salmons, que fins i tot són capaços de reconèixer l’aigua del riu on van néixer. Però també n’hi ha d’altres que tenen la capacitat de reconèixer la topografia del fons o bé de guiar-se pels canvis de temperatura i de salinitat. D’altres s’orienten amb el sol i fins n’hi ha que, amb la mateixa finalitat, es regeixen pels camps magnètics i elèctrics.
Les relacions tròfiques
L’alimentació

Els peixos herbívors s’han especialitzat a menjar vegetals aquàtics. Les madrilles (Chondrostoma toxostoma), que veiem en la fotografia, són un bon exemple d’aquesta especialització: per a raspar les algues que recobreixen les pedres del fons, disposen d’un llavi inferior especialitzat, corni, que deixa sobre les pedres unes marques característiques, d’un o dos centímetres de llargada.
Adolf de Sostoal.
L’alimentació dels peixos és molt diversa, i depèn en gran part del grau d’especialització. Els peixos poden ocupar diferents nivells tròfics en els diversos ecosistemes; per això és possible de classificar-los en herbívors, carnívors, detritívors i omnívors, segons llurs hàbits alimentaris. Tanmateix, el règim alimentari de les larves i dels joves sol diferir sensiblement del dels adults. Així, la majoria de larves es nodreixen de zooplàncton, especialment de copèpodes, cladòcers i rotífers, segons l’abundància amb què es presenten cada medi i hàbitat. A les aigües dolces algunes espècies, com ara els ciprínids, s’alimenten de diferents tipus d’organismes petits, entre els quals s’inclouen els primers estadis larvals d’insectes bentònics. Les larves de les llamprees (Petromyzon, Lampetra) constitueixen un cas especial: presenten un règim típicament micròfag i es nodreixen de la matèria orgànica que filtren del fons.

Els peixos filtradors s’alimenten dels organismes que viuen suspesos en l’aigua. Alguns elasmobranquis, com el pelegrí (Cetorhinus maximus), mantenen la boca completament oberta mentre neden i tota l’aigua que els entra és filtrada a les brànquies, on queden retinguts tota mena d’organismes i partícules d’aliment, que posteriorment passen a l’esòfag.
Tony Crabtree / Oxford Scientific Films - Firo-Foto.
En realitat, els peixos veritablement herbívors, que s’alimenten directament de vegetació aquàtica, són ben pocs, tant a les aigües dolces com a les marines: la majoria consumeixen plàncton, algues filamentoses o diatomees, com ara les madrilles (Chondrostoma toxostoma) que bàsicament mengen diatomees. En determinats ambients tropicals, on les aigües dels rius inunden periòdicament grans extensions de terra ferma, hi ha algunes espècies de peixos que aprofiten alguns dels recursos vegetals, com ara fulles, flors i fruits (per exemple el gènere Puntius d’Indondèsia i Colossoma de l’Amèrica del Sud). Hi ha peixos, com la carpa xinesa (Ctenopharyngodori) que s’alimenten de vegetació aquàtica. Si bé en el contingut gàstric de molts ciprínids europeus, com els barbs (Barbus), s’observa una gran quantitat de restes vegetals, de fet aquests peixos pràcticament només aprofiten els organismes epibionts que viuen associats a la vegetació, com ara diversos estadis larvals d’insectes. A les aigües marines també hi trobem algunes espècies amb hàbits herbívors, que principalment mengen algues calcàries, i si bé són més abundants a les mars tropicals, tampoc no en manquen exemples a la Mediterrània, com ho són algunes espècies de làbrids i de blènnids.

Entre els peixos depredadors n’hi ha que s’especialitzen a alimentar-se de peixos, com el lucí (Esox lucius), que s’amaga entre la vegetació i caça a l’aguait tota mena de peixos, i de vegades també altres vertebrats. Juntament amb les anguiles i les truites grosses, és l’espècie més voraç de les nostres aigües dolces.
Colin Milkins / Oxford Scientific Films - Firo-Foto.
Els peixos planctívors (per exemple els clupèids, els engràulids, els coregònids i els centràrquids) tenen una mandíbula protràctil i gaudeixen, a més, d’un sistema força eficient per a filtrar l’aigua i retenir-ne els organismes. Aquesta adaptació consisteix en una sèrie d’espines en forma de pinta (branquiospines) que se situen a la cara interna dels arcs branquials. Alguns elasmobranquis de mida gran, com el pelegrí (Cetorhinus maximus), es nodreixen de tots els organismes que els entren a la boca per filtració (peixos, cefalòpodes i crustacis, entre d’altres). Hi ha una gran varietat de peixos microdepredadors que s’alimenten de petits animalons (anèl·lids, crustacis, insectes, mol·luscs i d’altres) que capturen al fons, entre dues aigües o a la superfícies (com ara els blènnids, els gòbids, els gasterostèids, els ciprínids, els singnàtids, etc). N’hi ha, encara, que no capturen l’aliment de manera directa, sinó que ho solen fer en companyia d’algues o plantes (com els ciprínids i els silúrids, entre d’altres). Un determinat nombre d’espècies carnívores es nodreixen a base de capturar preses de mida mitjana; la majoria prefereixen menjar peixos, com fan per exemple el luci (Esox lucius) i la tonyina (Thunnus thynnus), però tampoc no manquen aquells peixos capaços d’ingerir amfibis, rèptils i àdhuc ocells o mamífers. En aquest sentit alguns elasmobranquis constitueixen un cas extrem, com ara el tauró blanc (Carcharodon carcharias), perquè poden atacar diversos tipus de mamífers marins i, fins i tot, són capaços d’atacar l’home. Hi ha una gran varietat de peixos micròfags que tan aviat es nodreixen d’organismes com de matèria orgànica o dels detrits que es troben al sediment; en aquest cas l’aliment s’obté filtrant l’estrat semilíquid del fons, sia ingerint el fang directament o succionant la superfície de les pedres i de les plantes. També hi ha peixos, com la carpa (Cyprinus carpio), que tenen un règim omnívor: es nodreixen d’invertebrats, però també ingereixen algues i detrits.

La relació entre l’estructura de l’aparell bucal i l’alimentació es pot exemplificar amb aquestes dues fotografies. A dalt, veiem les característiques de la boca d’una espècie típicament carnívora, que s’alimenta bàsicament de peixos, el lluç (Merluccius merluccius), dotat de dents llargues i esmolades, capaces d’immobilitzar qualsevol presa. A baix, la boca protràctil d’un peix planctívor, l’alatxa (Sardinella aurita), que canalitza els organismes planctònics, gràcies al seu embut bucal, i els reté a les branquiospines dels seus arcs branquials.
Jean-Georges Harmelin.
Gran part dels peixos són eurífags o generalistes, és a dir, que s’alimenten de diferents tipus d’animals o preses, que capturen de manera oportunista. Això explica que el seu règim alimentari es modifiqui al llarg de l’any segons l’abundància d’unes o altres preses. Sovint s’observa que determinats grups sistemàtics de peixos que conviuen en un mateix hàbitat tenen un règim alimentari molt similar. Si bé aquest fenomen no arriba a constituir cap problema quan l’aliment és abundant, sí que pot desencadenar una dura competència quan hi ha escassesa de recursos. En canvi, els denominats peixos estenòfags o selectius són aquells que escullen un aliment determinat i capturen específicament algun tipus de preses concretes.
Els peixos també poden representar una bona font d’aliment per a diferents grups d’organismes. A la mar tant els peixos joves com algunes espècies de peixos de petites dimensions són ingerits sovint per diferents menes d’invertebrats marins, com ara els cefalòpodes, les anemones o les meduses, entre d’altres. Però els peixos adults també són presa de molts ocells, com els corbs marins (Phalacrocorax), les baldrigues (Puffinus), els cabussons (Podiceps), els xatracs (Sterna) i els gavots (Alca). També poden ésser devorats per diversos mamífers marins, com ara les foques (Monachus) i els cetacis. D’altra banda, a les aigües dolces les larves dels peixos poden ésser capturades per alguns insectes aquàtics de bones dimensions, com les larves dels odonats i els ditíscids. Són relativament freqüents les espècies de vertebrats que es nodreixen de peixos; cal citar entre d’altres la serp d’aigua (Natrix maura) i la tortuga d’aigua ibèrica (Mauremys caspica), que durant els mesos càlids capturen una quantitat de peixos considerable. A més dels rèptils, hi ha alguns ocells i mamífers que també consumeixen peixos d’aigua dolça, per exemple el blauet (Alcedo atthis), i també la llúdriga (Lutra lutra).
El comportament alimentari
L’alimentació dels peixos està relacionada tant amb els aspectes morfològics com amb els biològics i, també, amb el comportament. És a dir, els peixos que des d’un punt de vista morfològic són molt especialitzats, també ho són a nivell del seu comportament alimentari. Així i tot, es fa difícil predir l’alimentació d’una espècie a partir de les seves característiques, perquè hi intervenen diversos factors, d’entre els quals cal citar l’abundància i la disponibilitat d’aliment. Gran part dels peixos varien la seva alimentació segons els recursos que hi ha més abundants a cada moment. En aquest sentit, no és estrany de trobar espècies, com ara la truita (Salmo trutta), que a determinades èpoques de l’any capturen insectes bentònics i en altres períodes es nodreixen, principalment, dels animals que deriven pel corrent a la superfície de l’aigua.
L’estructura de l’aparell bucal, com és lògic, està estretament relacionada amb l’alimentació, però també ho estan els òrgans dels sentits: la majoria dels peixos depenen de la vista per a reconèixer i capturar les preses o, d’una forma més generalitzada, detectar l’aliment; també n’hi ha, però, que utilitzen l’olfacte, el sistema acústicolateral o bé els camps elèctrics.
La recerca de l’aliment depèn molt del tipus d’espècie i d’hàbitat. La majoria de peixos són d’hàbits diürns i s’alimenten durant les hores solars, però també n’hi ha que practiquen l’activitat alimentària durant la nit; i encara n’hi ha moltes que aprofiten les hores crepusculars per a nodrir-se. La localització de l’aliment és força variable i depèn de diversos factors. En alguns casos la font d’aliment és a prop del lloc de residència dels peixos, que, en conseqüència, a penes es mouen d’una àrea reduïda. En canvi, altres vegades la recerca de l’aliment comporta haver de recórrer llargues distàncies, cosa ben normal en les espècies pelàgiques migratòries. Daltra banda, l’activitat de cercar l’aliment pot realitzar-se en solitari, en grups força reduïts o bé en massa.
Algunes espècies seleccionen l’aliment, i és freqüent que aquesta elecció estigui força més lligada a la mida de les preses o de les partícules nutritives, que no pas a la seva forma. En determinades circumstàncies, però, la selecció del menjar es relaciona força amb la seva olor, el gust, la textura o fins i tot amb el color. Els peixos tendeixen a una eficiència òptima en les captures, de manera que la qualitat de l’aliment sempre sigui superior a l’esforç invertit en la recerca, en la captura i en la mateixa ingestió de l’aliment.
La captura de les preses és una activitat que es pot realitzar en solitari o en grup, i això depèn de cada espècie concreta. Així, les tonyines (Thunnus thynnus) es reuneixen per a perseguir les moles de preses, mentre que els lucis (Esox lucius) prefereixen de sotjar-les en solitari. Sovint es troben diversos tipus d’activitat social que comporten la cooperació de dos o més individus amb l’objecte de desenterrar o d’extreure determinades preses, com ara els mol·luscs i els anèl·lids; fins i tot, com passa amb els taurons, aquesta tasca col·lectiva pot basar-se en el fet de trossejar o d’esmicolar una presa.