
Els ascomicets comprenen unes 30 000 espècies de fongs, 16 000 de les quals es troben en forma de liquen, associades amb algues. Això vol dir que, si hi afegim també els deuteromicets (ascomicets sense reproducció sexual), de cada deu espècies de fong, sis són ascomicets. La fotografia mostra un exemple ben típ3ic d’ascomicet no liquenificat i pluricel·lular, el de les cassoletes, i concretament, la cassoleta vermella (Sarcoscypha coccinea). S’hi veuen diverses fructificacions cupuliformes. La part externa d’aquestes fructificacions és l’excípul, I la interna, de color carmí més viu, és entapissada per l’himeni, on hi ha els ascs, productors d’espores. També hi ha ascomicets unicel·lulars, d’organització molt simple, però igualment formadors d’ases: són els llevats, d’una gran importància industrial.
Joaquim Carbó / SCM
Aquest és el grup més ampli i diversificat de fongs. Les estimacions més recents (1988) n’accepten unes 30 000 espècies, de les quals més de 16 000 corresponen a ascomicets liquenificats. Si hom considera que els 17 000 deuteromicets coneguts són en realitat ascomicets que han perdut la reproducció sexual, es veu com la importància d’aquest grup en el conjunt dels fongs és enorme (un 60%).
Es tracta bàsicament de fongs terrestres, que només secundàriament i en nombre relativament petit s’han adaptat a viure a l’aigua dolça o a la mar. En general són sapròfits, però se’n coneixen molts de paràsits. El miceli és format per hifes molt ramificades, septades, però amb septes no complets sinó perforats per un porus simple, que pot anar associat a cossos de Woronin, unes masses arrodonides que els poden obturar, sobretot quan l’hifa sofreix alguna alteració o envelleix. La paret conté una bona proporció de quitina. Alguns grups molt especialitzats presenten l’aparell vegetatiu en forma de llevat, format per cèl·lules disperses que es divideixen per gemmació; l’especialització els ha portat a la pèrdua parcial i fins i tot total de la quitina de la paret. La reproducció sexual es produeix per contacte gametangial, gametangiogàmia, espermatització o fusió de cèl·lules vegetatives, i condueix, de forma immediata o després d’una dicariofase (hifes dicariòtiques), a l’aparició d’esporangis en forma de sac, on, prèvia meiosi, es produeixen espores a l’interior. Es tracta, doncs, d’endosporangis, i s’anomenen ascs (del grec ascós, bot, sac). Les espores produïdes pels ascs, anomenades ascòspores, queden lliures per trencament o lisi de la paret o, més sovint, són expulsades amb força a través d’un aparell apical més o menys complicat.
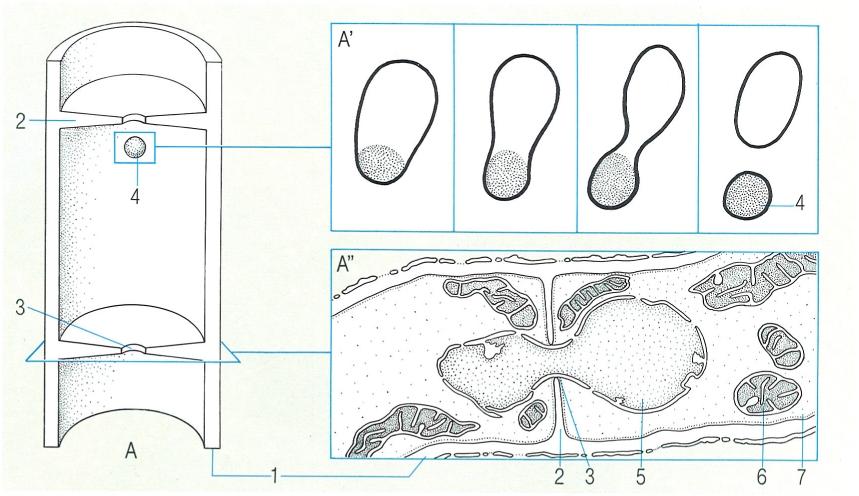
La característica més destacada del miceli dels ascomicets és el fet que les seves hifes, ramificades, tenen septes foradats per un porus, el qual pot ésser obturat pels anomenats cossos de Woronin. El dibuix mostra una secció longitudinal d’una hifa (A) per a fer visibles els septes perforats, i un detall de la formació d’un cos de Woronin a partir d’un microcòs (A’) i del pas d’un nucli a través d’un septe (A"), en una hifa de Neurospora crassa. Hom hi ha indicat: 1 paret de la hifa, 2 septe, 3 porus, 4 cos de Woronin, 5 nucli, 6 mitocondri, 7 citoplasma.
Biopunt, a partir de fonts diverses
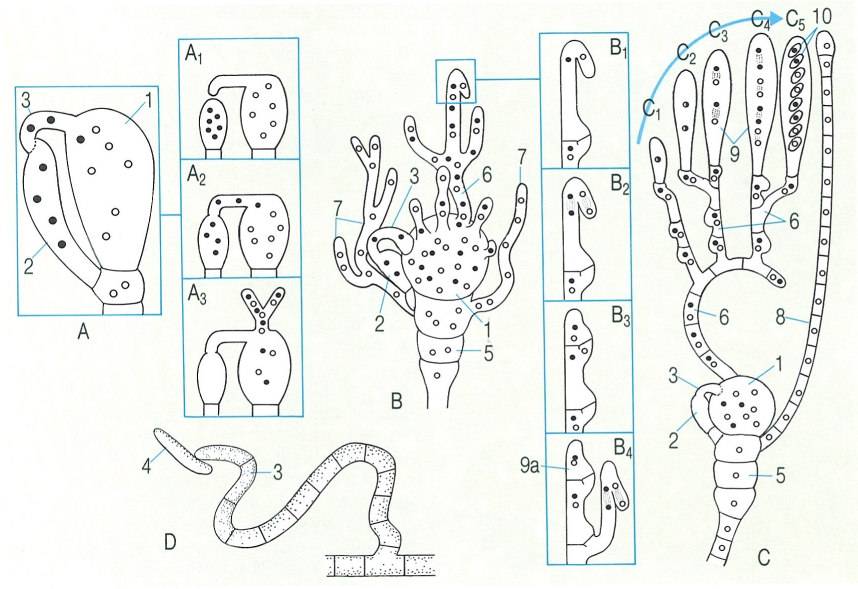
La fecundació dels ascomicets que conduirà al desenvolupament dels ascs es pot produir per un contacte gametangial (A) o bé per espermatització (D). A Contacte gametangial. amb la intervenció de la tricògina, en Pyronema domesticum: A1 creixement de la tricògina vers l’anteridi; A2soldadura i penetració dels nuclis masculins a l’ascogoni; A3 inici del creixement de les hifes ascògenes, encara sense septes. B Creixement de les hifes ascògenes (fase posterior a A3), juntament amb hifes estèrils monocariòtiques, i fases (B1-B4) del creixement, mitjançant la formació d’uncínuls, de les hifes ascògenes (B1 inici de la formació d’un uncínul en una hifa ascògena; B2 divisió conjugada dels dos nuclis; B4 fusió de l’uncínul amb la cèl·lula posterior i pas d’un dels nuclis; B4 formació d’un uncínul en una ramificació i cariogàmia a la cèl·lula terminal). C Formació d’ases a partir de les hifes ascògenes, i de paràfisis a partir de les hifes monocariòtiques (d’esquerra a dreta, fases de la maduració de l’asc: C1 zigot; C2, C3 dues fases de la meiosi; C4 mitosi; C5 maduració de les ascòspores). D Soldadura d’un espermaci damunt de la tricògina d’un ascogoni. Hom hi ha indicat: 1 ascogoni, 2 anteridi, 3 tricògina, 4 espermaci, 5 cèl·lules basals, 6 hifa ascògena, 7 hifa haploide (monocariòtica), 8 paràfisis, 9 asc (9a cèl·lula apical que, per cariogàmia, es pot transformar en el zigot C1), 10 ascòspora.
Biopunt, a partir de fonts diverses
La taxonomia dels ascomicets s’havia basat tradicionalment en el tipus de fructificacions (ascocarps o ascomes). Actualment, s’ha vist que molts grups diferents poden donar fructificacions o diferenciacions vegetatives que s’assemblen.
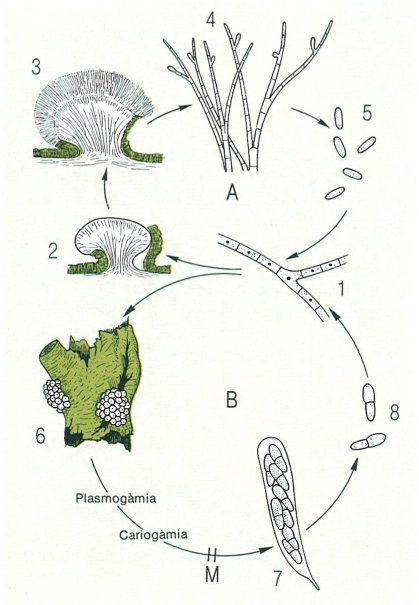
Un model de cicle vital dels ascomicets és el de Nectria cinnabarina. En el cicle asexual (A), el miceli (1) forma un estroma (2), que constituirà la base de l’esporodoqui (3), a la superfície del qual creixeran els conidiòfors (4), formadors de conidis (5), els quals, després de germinar, donen lloc a nous micelis, monocariòtics. En el cicle sexual (B), el miceli desenvolupa gametangis (anteridis i ascogonis), i de la fecundació se’n deriven hifes ascògenes dicariòtiques, que es ramifiquen dins els peritecis (6), i originen ascs (7), prèvia cariogàmia i meiosi. Els ascs donaran ascòspores (8). Ouan aquestes germinen, donen lloc a un nou miceli, monocariòtic. M meiosi.
Biopunt, modificat, a partir de C. J. Alexopoulos
Molts òrgans aparentment simples i primitius són el resultat de fenòmens de simplificació evolutiva, sovint a causa de la interrupció del desenvolupament en una fase juvenil (paidomorfosi). Una correcta interpretació de les afinitats naturals s’ha de basar en l’estudi d’estructures molt estables, com els ascs i, en concret, els aparells apicals i les formes de dehiscència (alliberament de les espores) i en la ontogènia (etapes del desenvolupament) dels cossos fructífers. Els micòlegs actualment es dediquen a aquest camp d’estudi, però encara no disposen de dades referents a tots els grups implicats. Hom creu que els ascomicets procedeixen de zigomicets, probablement d’entomoftorals. Però l’absència de miceli dicariòtic o la diferència en la maduració dels ascs ha fet pensar que les endomicètides (Endomycetidae) i les tafrinomicètides (Taphrinomycetidae), que no formen fructificacions, poden haver evolucionat com a línies separades de la resta dels ascomicets, des de molt antic. Per la seva gran especialització i per l’absència d’hifes dicariòtiques, hom situarà també les laboulbeniomicètides (Laboulbeniomycetidae) en una subclasse independent. Però el gran gruix dels ascomicets, els que tenen fructificacions i hifes dicariòtiques, correspon a la subclasse de les ascomicètides (Ascomycetidae). A l’hora d’exposar les generalitats d’aquest darrer grup, hom s’ocuparà dels aspectes generals característics de la majoria dels ascomicets, que cal conèixer quan es volen estudiar i identificar.
Les endomicètides: llevats esporògens
Aquesta subclasse comprèn els ascomicets amb aspecte de llevats. De fet, en l’evolució dels fongs diverses vegades han aparegut fases de desenvolupament del tipus llevat, caracteritzades per un aparell vegetatiu unicel·lular, amb reproducció per escissió o per gemmació. La distinció entre els llevats i els fongs miceliars és subjectiva, i es basa gairebé sempre en consideracions de tipus pràctic. Molts formen, en cultiu pur, colònies pigmentades (vermelles, ataronjades, grogues, etc.).
De fet, els llevats corresponen a un grup heterogeni d’organismes adaptats a viure en condicions d’una gran riquesa d’aliments, sovint en els medis líquids. A part les fases levuriformes del cicle vital de diversos fongs (tafrinomicètides, exobasidiàcies, ustilaginàcies), estudiats en els seus grups respectius, hi ha tres grups principals de llevats: les esporobolomicetals, que deriven de basidiomicets; les blastomicetals o llevats no esporògens, que no presenten formes sexuals ni bal·listòspores; i les endomicetals (Endomycetales), que, juntament amb dos petits ordres, protomicetals (Protomycetales) i ascosferals (Ascosphaerales), integren les endomicètides.
Les tafrinomicètides
Es tracta d’un grup de fongs paràsits caracteritzat perquè els ascs no es troben formant part d’aparells fructífers sinó disposats en capes que recobreixen la superfície dels teixits de plantes superiors o de falgueres, i perquè les parets de les hifes no contenen quitina. La classe conté l’únic ordre de les tafrinals (Taphrinales), amb una sola família, les tafrinàcies (Taphrinaceae).
Les laboulmeniomicètides
Són ascomicets molt especialitzats, caracteritzats pel fet de tenir peritecis i per l’absència d’un veritable miceli. Gairebé tots són paràsits obligats d’artròpodes, i s’inclouen a l’ordre de les laboulbenials (Laboulbeniales), però n’hi ha alguns amb peritecis més complexos, que viuen paràsits sobre algues, i s’inclouen dins l’ordre de les espatulosporals (Spathulosporales).
Les ascomicètides
Aquesta subclasse, força homogènia i versemblantment monofilètica, comprèn la majoria dels ascomicets. En tots els seus representants, després de la plasmogàmia es formen hifes amb dos nuclis per compartiment (dicariòtiques), que es ramifiquen (en general dins una fructificació formada per hifes monocariòtiques) fins a donar, als extrems, ascs. Per això se’n diuen hifes ascògenes. És una forma d’aconseguir que, després de la plasmogàmia, es produeixin no un sinó molts zigots, ja que la cariogàmia es produeix en unir-se els dos nuclis de cada cèl·lula terminal de les hifes ascògenes. Cada cèl·lula terminal es convertirà doncs en un zigot diploide, el qual, de seguida, sofrirà la meiosi i donarà 4 o (si s’hi afegeix una mitosi) 8 nuclis. La cèl·lula terminal de què hom parla s’haurà convertit en un asc i, al voltant de cadascun dels nuclis es formaran 4 o 8 ascòspores (a vegades moltes més). En total, una fecundació condueix a molts centenars o milers d’espores.
L’oogoni (anomenat ascogoní) té una prolongació filamentosa, unicel·lular o pluricel·lular, la tricògina, on es realitza la fecundació, ja sia per espermatització, amb la intervenció d’espermacis —que són gàmetes masculins amb aspecte d’espora, que es produeixen en criptes anomenades espermatangis—, o per unió amb un anteridi (contacte gametangial o gametangiogàmia). No sempre es produeix una fecundació normal, ja que hi ha casos, no pas rars, d’aparellament espontani de nuclis en l’ascogoni, amb aparició d’hifes ascògenes sense prèvia fecundació (partenogàmia), encara que arribin a diferenciar-se anteridis. També es poden produir directament hifes ascògenes a partir d’hifes somàtiques (autogàmia). A l’interior de l’ascogoni fecundat, els nuclis es divideixen i s’aparellen sense unir-se (ja que només s’ha produït la plasmogàmia). Després hom veu aparèixer uns diverticles on se situen les parelles de nuclis, que es van dividint de manera conjugada (tots dos alhora). Els diverticles es van allargant i aviat es converteixen en hifes, amb septes que separen els dicàrions (compartiments amb dos nuclis). Tot això acostuma a passar en el si de la fructificació, formada a partir de les hifes monocariòtiques que envoltaven l’ascogoni i hom pot veure com, per dins, les hifes dicariòtiques creixen, es ramifiquen i s’infiltren. En tot cas, les hifes dicariòtiques destinades a produir els ascs, les hifes ascògenes, creixen d’una manera molt particular, ja que l’àpex s’incurva cap enrere i forma l’uncínul, semblant a un ganxo, a la part apical del qual se situen els dos nuclis que, mentrestant, comencen a dividir-se. Una de les còpies d’un dels nuclis passa a la punta del ganxo, la qual toca l’hifa i s’hi solda. En el punt de soldadura s’obre un porus pel qual aquell nucli fill passa a l’hifa, alhora que es forma un septe transversal que separa els dos nuclis apicals (una còpia de cadascun dels precedents) dels dos subapicals (les dues còpies restants). El que queda de l’uncínul es veu en cada septe, en forma de bony lateral. També es pot veure força bé a la base de cada asc. D’aquesta manera, on hi havia una cèl·lula amb dos nuclis diferents (una còpia del nucli «pare», i una còpia del nucli «mare»), hi ha ara dos dicàrions, cadascun amb una còpia de cada un d’aquests nuclis, com a resultat del procés d’uncinulació.
Els ascs
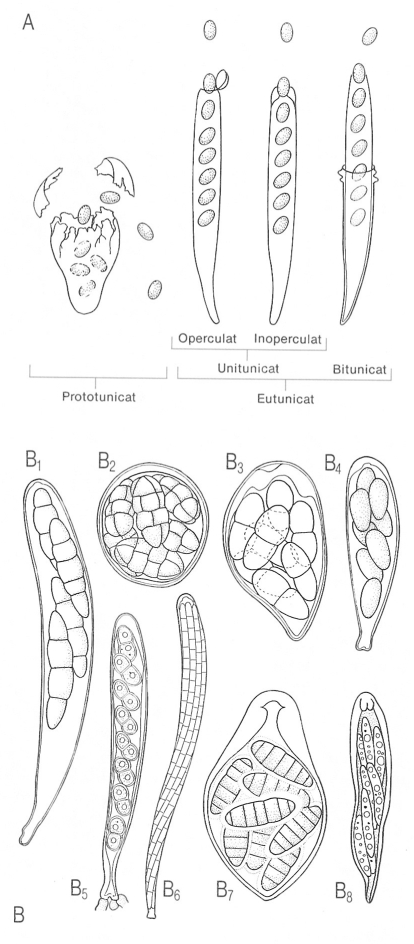
Tipus d’ascs segons l’estructura de la paret i els mecanismes de dehiscència (A), i diversos tipus d’ascs (B) per a mostrar la seva diversitat morfològica. B1 Asc claviforme, tetraspòric; B2 globós, octospòric; B3 piriforme; B4 curtament claviforme; B5 cilíndric, amb espores uniseriades; B6 cilíndric, amb espores filamentoses; B7 lageniforme o inversament claviforme, B8 claviforme, amb espores pluriseriades.
Biopunt, a partir de fonts diverses.
L’asc jove és una cèl·lula terminal de les hifes ascògenes i, per tant, conté dos nuclis haploides. Aquests, aviat s’uneixen (cariogàmia), completant la reproducció sexual que s’havia iniciat amb la plasmogàmia, a l’ascogoni. El nucli originat és el nucli del zigot, i és l’únic diploide del cicle vital. Sense interrupció, sofreix una meiosi seguida d’una mitosi, i s’originen 8 nuclis haploides al voltant dels quals es formaran sengles ascòspores. Els ascs poden tenir forma esfèrica, ovoide, de porra (claviformes) o cilíndrica. La forma i les dimensions poden servir per a separar gèneres i espècies. Hom suposa que degueren existir ascs amb una paret prima i simple, semblant a la de les endomicètides (ascs prototunicats), que alliberaven les espores per ruptura o lisi de la paret. Els que trobem actualment amb aquesta mena de paret s’interpreten, però, com a producte de la simplificació d’ases de paret més complexa, per un procés de reducció (ascs pseudoprototunicats). A la majoria dels ascs, la paret consta d’una endotúnica, interna, i d’una exotúnica, externa, però en la majoria, aquestes capes estan soldades i no es desenganxen ni en el moment d’expulsar les espores (ascs unitunicats). Aquests ascs s’obren per un mecanisme d’obertura (opercle) o per un porus, i poden ésser operculats, rostrats (formen una protrusió cònica en obrirse), de dehiscència explosiva, etc. En altres casos, l’endotúnica és elàstica (formada per fibres disposades en acordió) i poc unida a l’exotúnica. Aquesta, en els ascs madurs, s’obre o es trenca i deixa que l’endotúnica s’infli i s’estiri, fins que el seu àpex està en posició d’ejacular les espores. Parlem llavors d’ases bitunicats, i el mecanisme d’obertura pot ésser com el descrit («jack-in-the-box» o en capsa de sorpreses, també dit fissitunicat) o d’una altra manera (rostrat, semifissitunicat, bilabiat).
El nombre normal d’espores és de 8 per asc, ja que després de la meiosi els nuclis acostumen a sofrir una sola mitosi. Però hi ha ascs amb 4, 2 i 1 espores, i ascs que les contenen en un nombre molt elevat (superior a 100).
En la taxonomia moderna hom atorga molta importància a les diferenciacions apicals (aparell apical), relacionades amb l’expulsió de les espores, que són molt variades. En uns casos, la part apical s’obre mitjançant un opercle o tapadoreta. Aquests ascs s’anomenen operculats. Per oposició, hom parla d’ases inoperculats quan hi ha un porus apical envoltat de diferenciacions més o menys complexes de l’endotúnica (per exemple, anells) que, apicalment, acostuma a ésser més gruixuda. També hi ha ascs que no tenen dehiscència apical i alliberen les espores per lisi o estripament de la paret. També té importància, en la taxonomia, el color (per exemple, blau) que prenen les diferents parts de l’asc quan se les tracta amb reactius iodats.
Les espores
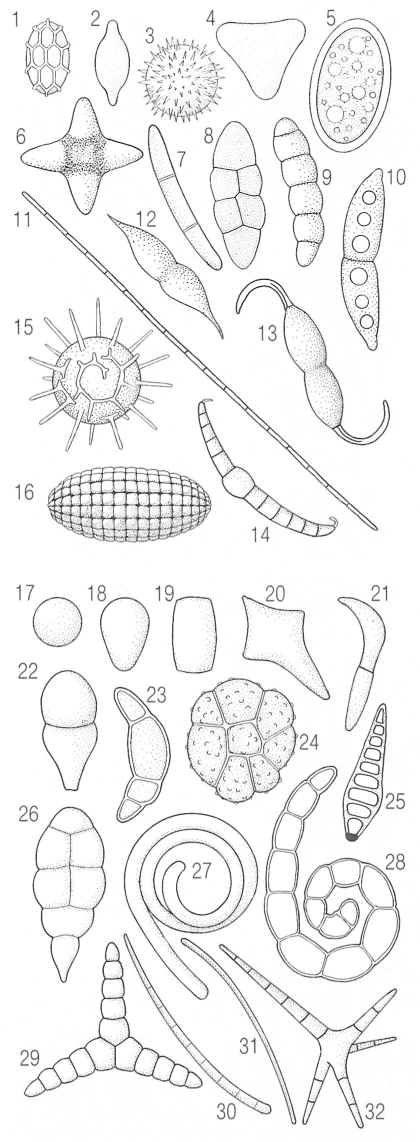
Exemples de la diversitat de les ascòspores o espores sexuals (a dalt), originades per ascs, i de les conidiòspores o espores asexuals (a baix), originades per cèl·lules conidiògenes. Poden ésser hialines (transparents, per exemple 1-5) o brunes (fuliginoses, per exemple, 8, 9, 16, 23, 24), i poden presentar septes transversals (transseptes, en 7, 9, 10, 11, 14, 22, 25) i longitudinals (longiseptes, en 8, 16). La superfície pot ésser llisa (per exemple, en 2, 5, 11) o ornamentada (per exemple, en 1, 3, 15, 24). En general, les conidiòspores o conidis són més petites que les ascòspores, i es produeixen en gran quantitat.
Biopunt, a partir de fonts diverses.
Un cop formats els nuclis de l’asc, hom veu aparèixer, en les ascomicètides, un sac aplanat, de membrana unitària, derivat del reticle endoplasmàtic: la vesícula ascal (o membrana ascosporal), que va creixent i estenent-se per la part perifèrica del citoplasma de l’asc, fins a envoltar gairebé del tot els nuclis. A continuació es va escanyant entre els nuclis, fins a delimitar un espai de citoplasma al voltant de cadascun. Després es formarà per dins la paret esporal, que té diverses capes protectores. La part de citoplasma que queda fora és el periplasma, que pot intervenir en l’ornamentació esporal i en les reaccions que condueixen a l’expulsió de les espores, ja que, quan aquestes són madures, els glúcids insolubles que conté sofreixen hidròlisi. Això fa pujar la pressió osmòtica, que fa que penetri aigua en l’asc fins que aquest esclata per la part apical, expulsant amb força les espores.
Les espores, molt variades pel que fa a llur mida, forma, color i ornamentació de la paret, poden ésser unicel·lulars o pluricel·lulars. Les pluricel·lulars sofreixen una ulterior divisió del nucli durant la maduració, i hom hi veu aparèixer diversos septes. Així, podem tenir espores bi, tricel·lulars, etc. També s’hi poden afegir septes longitudinals, que condueixen a la formació d’espores murals. En general, les espores germinen i produeixen tants tubs de germinació com cèl·lules tenen. La forma és sovint el·lipsoïdal (d’una el·lipse que girés entorn de l’eix llarg), però n’hi ha d’allargades, cilíndriques i fins filamentoses. Pel que fa al color, poden ésser incolores (hialines) o bé brunes (fuliginoses). La paret pot ésser llisa o variadament ornamentada. Sovint contenen una o més grans gútules lipídiques. Es tracta de les reserves que necessiten per a mantenir-se vives fins que poden germinar.
Els cossos fructífers
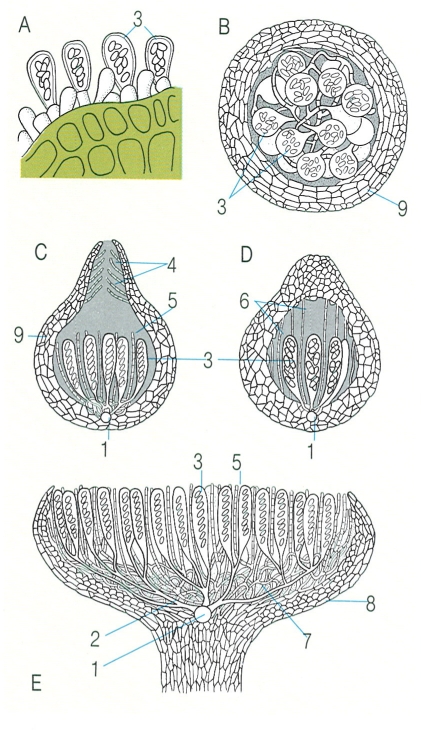
Tipus d’ascomes i de desenvolupament: A ascs nus, sense formar ascocarp; B clistoteci, totalment tancat; C periteci (d’origen ascohimenial); D pseudoteci (d’origen ascolocular), abans de la formació de l’ostíol. E apoteci. Hom hi ha indicat: 1 centre, amb restes de l’ascogoni, 2 hifes ascògenes, 3 asc, 4 perífisis, 5 paràsifis, 6 pseudoparàsifis, 7 hipoteci, 8 excípul, 9 paret de l’ascocarp.
Biopunt, a partir de fonts diverses.
S’anomenen ascomes o ascocarps, i són formats per hifes estèrils, que protegeixen les hifes ascògenes i fan més eficaç la maduració i dispersió d’espores per part dels ases. Poden ésser des de molt petits (menys de 0,2 mm) fins a bastant grossos (20 cm d’alçada en algunes Morchella, o 10 cm de diàmetre en algunes Peziza). Hom en distingeix actualment quatre grans grups morfològics. En primer lloc, els clistotecis, que acostumen a ésser petits i es caracteritzen perquè són completament tancats i perquè les hifes estèrils envolten del tot la zona on es formen, sense ordre, els ascs. Per tant, les espores només podran quedar lliures per disgregació de la paret del clistoteci. Actualment s’ha observat que molts grups sense relació evolutiva directa poden produir clistotecis. En segon lloc tenim els peritecis, més o menys tancats, no tan petits (0,2 - 2 mm), amb una paret preformada, el peridi o pireni, que adopta una certa forma d’ampolleta, ja que la part apical (l’ostíol) és una perforació que comunica amb l’exterior. En tercer lloc, els apotecis, que poden ésser des de petits (0,5 mm) fins a atènyer les mides més grosses del grup, i són oberts, amb forma de plat, de cassoleta, de copa o amb pezu i formes més complexes, com la de les múrgules o la de les orelles de gat; els ascs hi estan sempre en contacte amb l’exterior. També hi ha ascomicets que només formen ascs nus, no pertanyents a cap cos fructífer.
Queda per explicar el quart tipus, però abans cal dir que hi ha dues grans maneres de formació dels ascocarps: l’ascohimenial i l’ascolocular. En la primera, envoltant l’ascogoni, les hifes monocariòtiques comencen a organitzar el que seran els teixits de l’ascocarp, i formen una cavitat propera al carpocentre, on hi ha hagut l’ascogoni i la fecundació. Les hifes ascògenes creixen i s’organitzen al mateix temps, dins la cavitat, normalment formant un himeni, on els ascs i les hifes protectores (hamateci) es disposen en una capa. En canvi, en els ascocarps ascoloculars, primer es produeix una massa d’hifes compacta, l’estroma, i, des de dins, les hifes ascògenes, sovint formant un feix, van provocant la lisi de les hifes de l’estroma. Aquesta activitat disgregadora obre progressivament una cavitat (anomenada lòcul) que és ocupada al mateix temps pels ascs joves. Es forma així una fructificadó, l’ascostroma, delimitada pel que resta de l’estroma, que quan té aspecte de periteci (amb un origen diferent, però,) s’anomena pseudoteci. És el quart tipus de fructificació. Quan és allargat, es parla d’histeroteci, i quan és aplanat i protegit per un escutel d’estructura més o menys radial, hom parla d’un tirioteci.
L’hamateci
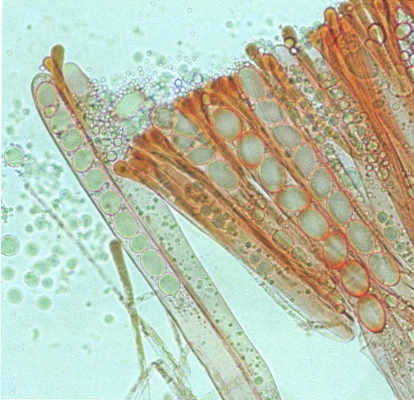
Secció de l’himeni (capa fèrtil) del bocabadat bru (Helvella acetabulum), tractada amb lugol. Hi veiem alguns ascs madurs, amb 8 ascòspores el·lipsoïdals seriades, al costat d’ases immadurs i de paràfisis (filaments acabats en una dilatació apical, protectora de l’himeni).
Oriol Martí i Jesús Gabaldon.
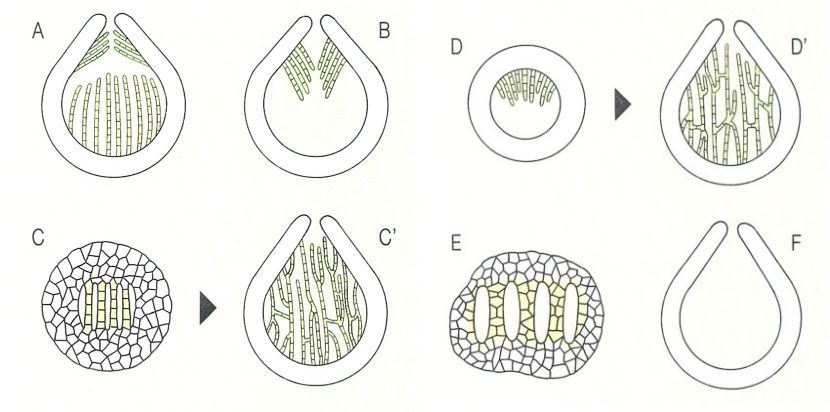
Tipus d’hamatecis en els ascomicets (en groc, en el dibuix): A hamateci format perparàfisis que surten del fons de la cavitat de l’ascoma i perífisis que neixen vora l’ostíol; B hamateci sense paràfisis i només amb perifisoides, descendents; C hamateci amb parafisoides, restes dels teixits de l’ascoma jove, que s’expansionen després, i originen una xarxa de filaments (C’); D hamatecis amb pseudoparàfisis, que apareixen al sostre de l’ascoma jove, s’anastomitzen i van a unir-se a la part basal del lòcul (D’); E hamatecis amb ascs situats en lòculs individuals, dispersos per l’estroma; F sense hamateci de cap mena.
Biopunt, a partir d’O. Eriksson.
Els ascs apareixen sovint acompanyats d’hifes estèrils que constitueixen l’hamateci (del grec hama, alhora, conjuntament). L’himeni és doncs el conjunt dels ascs i l’hamateci. La taxonomia moderna atorga molta importància al tipus d’hamateci dels ascocarps. Entre els principals, cal esmentar els següents: les paràfisis, hifes que s’originen a la base de l’himeni (és a dir, en el fons de la cavitat de l’ascoparp), des d’on s’allarguen fins a l’àpex dels ases, on sovint es dil·laten (paràfisis capitades); els parafisoides, que s’originen a partir de les restes del teixit situat entre els ascs, que, en separar-se, prenen un aspecte de filaments estrets, anastomitzats i amb septes distants; les pseudoparàfisis (característiques dels ascomicets bitunicats), hifes originades a la part superior de l’ascocarp, sobre el nivell dels ascs, que penetren després entre aquests i baixen fins a fixar-se a la base de la cavitat, i que sovint són gruixudes, ramificades i anastomitzades; les perífisis, hifes del canal de l’ostíol, no ramificades ni anastomitzades; i les perifisoides, hifes originades a la part superior de l’ascocarp, sobre la capa dels ascs, però tan curtes que no arriben a introduir-se a l’himeni.
Els conidis
A més del cicle sexual, moltes ascomicètides tenen un cicle asexual, normalment per producció de mitòspores que es dispersen i germinen aviat, els conidis, que es formen sobre hifes especialitzades anomenades conidiòfors. Els conidiòfors poden aparèixer lliures o reunits en fructificacions, els conidiomes (picnidis o acèrvuls). La fase conidiògena, dita anamòrfica, pot tenir molta o poca importància, segons l’espècie. El tipus d’anamorf és actualment un caràcter important en la descripció dels gèneres d’ascomicets. En alguns casos, agafa tanta importància que la fase sexual o telomòrfica tendeix a desaparèixer. Quan ens és desconeguda, aquests fongs s’estudien entre els deuteromicets.
Sistemàtica
Seguint la suggerència d’Eriksson (1988), hom començarà estudiant els ordres amb ascs bàsicament unitunicats, amb un primer grup d’ordres amb apotecis, que corresponen a l’antic grup dels discomicets (s’assenyalen amb un asterisc els ordres de fongs que tenen representants liquenificats). Són les pezizals* (Pezizales), les leocials* (Leotiales), les ritismatals (Rhytismatales), les ostropals* (Ostropales) i les patel·larials* (Patellariales), aquestes darreres, amb ascs bitunicats. A continuació hom tractarà un segon grup d’ordres amb peritecis, que corresponen aproximadament a l’antic grup dels pirenomicets. Són les hipocreals (Hypocreales), les clavicipitals (Clavicipitales), les diatripals (Diatrypales), les xilarials* (Xylariales), les sordarials (Sordariales), les diaportals (Diaporthales) i les fil·lacorals (Phyllachorales). Hom seguirà amb els ordres amb ascs pseudoprototunicats (secundàriament simplificats), on s’inclouran els ordres que tenen predominantment clistotecis, que corresponen grosso modo a l’antic grup dels plectomicets. S’hi han inclòs les eurotials (Eurotiales), les halosferials (Halosphaeriales), les onigenals (Onygenales), les elafomicetals (Elaphomycetales), les microascals (Microascales), les ofiostomatals (Ophiostomatales) i les ascosferals (Ascosphaerales).
Els fongs amb ascs fissitunicats, que corresponen a l’antic grup dels loculomicets, no són encara prou ben coneguts per a separar-los de forma estable en ordres. Per això s’han inclòs tots en un sol gran ordre, el de les dotideals* (Dothideales).
Resta un grup residual d’ordres amb els ascs atípics, que encara no es poden situar enlloc amb seguretat. Són les erisifals (Erysiphales), les meliolals (Meliolales) i les corinelials (Coryneliales).