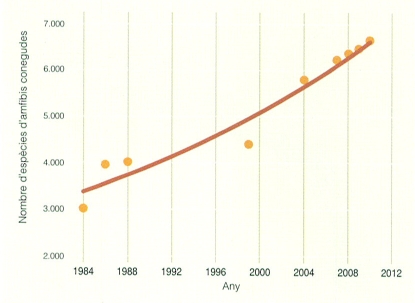
Més del 50% de les espècies d’amfibis conegudes s’han descobert durant els darrers tretze anys. El nombre de tàxons d’aquest grup de vertebrats també s’ha anat incrementant, ja que cada vegada hi ha més investigadors que es dediquen a l’herpetologia i es disposa de tècniques d’estudi més perfeccionades, fet que possibilita la publicació d’un nombre d’estudis més elevat.
Dades dels autors.
El coneixement dels amfibis i els rèptils dels Països Catalans ha experimentat grans canvis, com també el d’aquest grup de vertebrats a escala mundial. Ho reflecteix una dada molt significativa: si l’any 1986 es coneixien un total de 3.952 espècies d’amfibis i rèptils, al febrer del 2010 se n’havien descrit 6.629. Això representa un augment del 67%. Aquest increment és fruit, sens dubte, d’un augment de la investigació herpetològica a nivell mundial; l’ús de noves tècniques moleculars, que han revolucionat el coneixement evolutiu de molts grups; la globalització de les comunicacions, que permet treballar amb equips supranacionals, i l’alt nivell de formació dels científics. Tot això es tradueix en un increment notable de les publicacions, tant a nivell nacional com internacional. Però no solament ha aparegut una nombrosa bibliografia de temàtica herpetològica, sinó que universitats, centres de recerca i museus d’arreu del món mantenen equips d’investigació dedicats a l’estudi i la conservació de diverses espècies d’amfibis i rèptils.
Aquest augment de la recerca herpetològica arreu del món també s’ha donat en l’àmbit del nostre territori. Pràcticament tota la informació herpetològica referent als Països Catalans de què es disposava el 1987 es fonamentava en l’ingent treball en solitari realitzat per Maria Victòria Vives-Balmaña sobre Catalunya, en els articles de Joan Mayol, Joan Antoni Alcover i Maria Antònia Cirer sobre les Balears i en alguns estudis sobre altres indrets del territori. En canvi, actualment es disposa d’una bibliografia abundant, tant pel que fa a la totalitat dels Països Catalans com a regions concretes. Cal destacar la publicació de l’Atlas dels amfibis i rèptils de Catalunya i Andorra (1995), Peces continentales, anfibios y reptiles de la Comunidad Valenciana (2002) i Rèptils i amfibis de les Balears (1985 i 2003). Durant tot aquest temps també han aparegut nombrosos llibres d’abast espanyol que inclouen informació herpetològica valuosa sobre el nostre territori, com Distribución y biogeografía de los anfibios y reptiles en España y Portugal (1997), Atlas y Libro Rojo de los anfibios y reptiles de España (2002), a més de nombroses guies d’amfibis i rèptils que seria llarg d’enumerar.
Un altre fet fonamental ha estat l’aparició de nombroses monografies d’abast comarcal o dedicades a espècies concretes. Les unes i les altres tenen com a objectiu donar a conèixer les comunitats herpetològiques locals i, juntament amb l’aparició de molts grups dedicats a l’herpetologia, són una mostra evident de l’interès creixent per la fauna d’amfibis i rèptils. Entre les monografies d’abast comarcal es poden esmentar l’Atlas de distribution des reptiles et amphibiens du Languedoc-Roussillon (1987), Els amfibis i els rèptils. Banyoles: fauna comarcal (1993), Anfibios y reptiles de la región de Murcia (1993), Artesa de Lleida. Fauna: peixos, amfibis, rèptils, ocells i mamífers (1995), La lagartija de las islas Columbretes. Biología y conservación (1996), La fauna vertebrada d’Osona (1998), Animals del Montseny. Mamífers, ocells, rèptils i amfibis (2000), Los anfibios del Alto Aragón (2000), The Biology of Lacertid Lizards. Evolutionary and Ecological Perspectives (2004), Enciclopèdia de Menorca, volum 2, Peixos, amfibis i rèptils (2005), Anfibios y reptiles en la provincia de Teruel (2006), Atles dels amfibis i rèptils del Parc Natural de la Serra de Montsant (2007), Introducció a la fauna vertebrada de la Safor (1988), Fauna vertebrada terrestre del municipio de Alicante (1990), Anfibios y reptiles del término natural de Pinoso (2002), Anfibios y reptiles de Teulada (2008), etc.
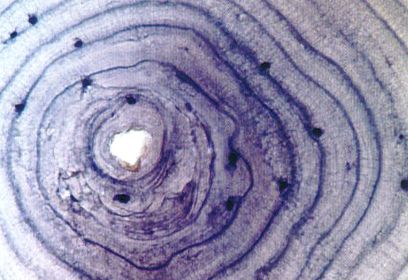
En un tall histològic dels ossos d’un amfibi es poden observar les pauses del creixement corresponents a la diapausa hivernal (en blau més fosc). Per tant, comptant els anells de creixement ossis, de manera semblant a com es compten els d’un arbre, es pot estimar l’edat d’un exemplar d’amfibi. Aquestes tècniques d’estudi reben el nom d’esqueletocronologia.
Albert Montori.
Si l’any 1987 l’objectiu fonamental de la Història Natural dels Països Catalans era donar a conèixer les espècies presents al nostre territori, actualment, i sense deixar de banda la diversitat herpetològica, es vol posar l’accent en l’estructura i el funcionament de les comunitats herpetològiques, analitzar els perills que les amenacen dins el context de la crisi de biodiversitat que afecta el conjunt del planeta i avaluar l’estat de les poblacions d’amfibis i rèptils dels Països Catalans. Indubtablement, en els vint-i-tres anys transcorreguts des de la primera edició, les tècniques d’estudi dels amfibis i els rèptils han avançat molt i han permès acumular nova informació herpetològica. Molts avenços tècnics cabdals han tingut lloc en el camp de la genètica, que ha esdevingut una ciència molt important per a l’estudi de la filogènia i l’ecologia de les poblacions. Això ha permès donar llum, unes vegades més que d’altres, a algunes de les problemàtiques herpetològiques actuals. D’altra banda, el desenvolupament d’un maquinari cada cop més potent ha permès desenvolupar eines de treball (sistemes d’informació geogràfica, internet, etc.) amb les quals és possible dur a terme estudis acurats a escales geogràfiques molt grans i manipular alhora un gran nombre de dades ambientals. Tot això s’explica amb detall als capítols següents, en els quals s’actualitzen els coneixements sobre l’herpetofauna dels Països Catalans.
Els estudis de distribució i biogeografia dels amfibis
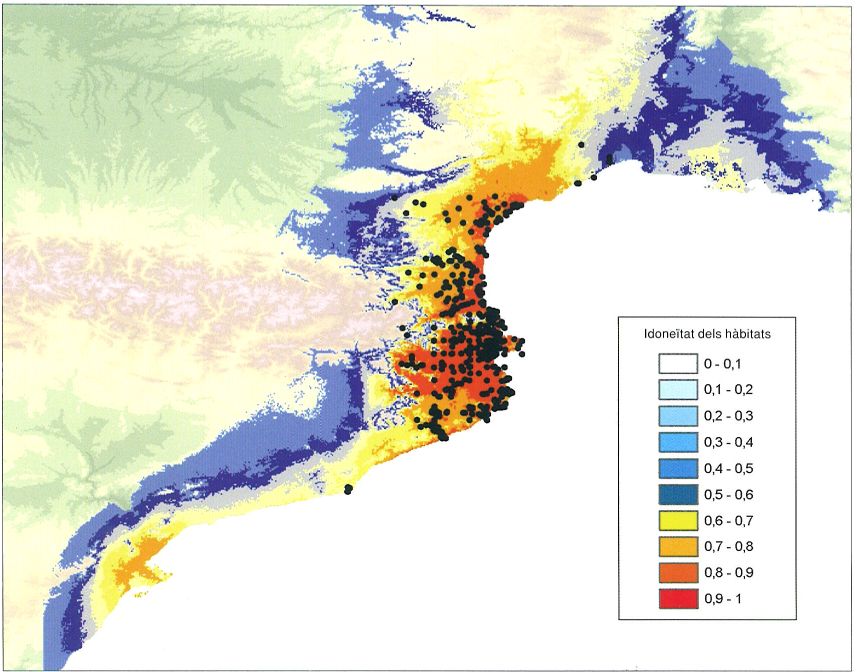
Àrea de distribució potencial del gripau pintat (Discoglossus pictus, taques de color) obtingut amb un SIG i localització real dels exemplars d’aquesta espècie (punts negres). El gripau pintat va ser introduït a Banyuls de la Marenda (Roselló) al final del segle XIX, i des d’aleshores s’ha anat expandint progressivament en totes direccions, però especialment cap al sud i el nord. Recentment ha colonitzat la conca mitjana i baixa de la Tordera i continua avançant litoral avall i remuntant el riu. També ha estat introduït recentment al delta del Llobregat.
Daniel Villero.
La distribució geogràfica dels amfibis als Països Catalans es coneix molt millor que ara fa vint-itres anys, no solament perquè durant aquest temps s’han anat fent nous estudis, sinó també perquè la metodologia i les tècniques de treball han fet un salt qualitatiu que permet analitzar millor no tan sols aquesta distribució sinó també els requeriments i els condicionaments ambientals filogeogràfics i, fins i tot, la conservació. Però allò que ha afavorit més l’aprofundiment en el coneixement de la distribució dels amfibis ha estat el creixent nombre d’herpetòlegs i el seu repartiment pel territori. Així, en l’elaboració de l’Atlas dels amfibis i rèptils de Catalunya i Andorra (1995) i de l’Atlas de los anfibios y reptiles de la Comunidad Valenciana (1999), es va dedicar un gran esforç a omplir els buits d’informació existents. En el cas dels amfibis, es van localitzar i visitar moltes basses i punts d’aigua per a fer rendible l’esforç de prospecció. Aquest treball, que s’ha anat ampliant per tots els Països Catalans, ha permès descriure més detalladament la distribució de les espècies, cosa que s’ha reflectit en els mapes de distribució. Mentre que en els primers es dibuixaven simplement taques sobre l’àrea de distribució coneguda de cadascuna de les espècies, a poc a poc es va anar dividint el territori en quadrícules de 10 × 10 km, d’acord amb un sistema de coordenades que es basa en la projecció cartogràfica anomenada UTM (Universal Transverse Mercator), que, a diferència del sistema de coordenades geogràfiques, s’expressa en metres i no en graus. Recentment, l’aparició d’eines que, mitjançant l’ús de satèl·lits, permeten determinar la localització geogràfica exacta dels exemplars (GPS, Global Positioning System) ha fet possible precisar encara més la distribució de les espècies, fins al punt que actualment els mapes de distribució ja no es basen en les quadrícules UTM, sinó en la posició exacta dels exemplars observats.
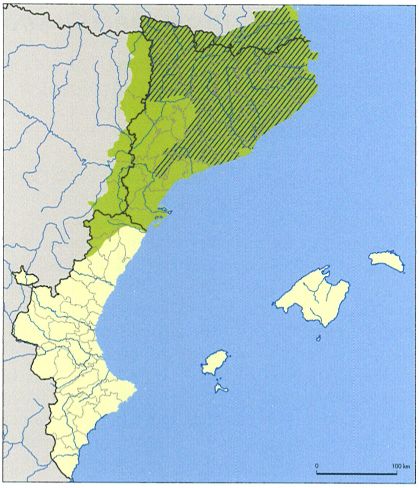
Àrea de distribució coneguda de la salamandra (Salamandra salamandra) l’any 1987 (en verd i ratllat verd) i l’any 2010 (en ratllat verd). El coneixement de la distribució de les diverses espècies d’amfibis als Països Catalans ha avançat de manera substancial. De moltes espècies, no tan sols se’n coneix més bé la distribució, sinó que també se n’han pogut determinar els requeriments ecològics i localitzar-ne amb precisió les poblacions.
IDEM, a partir de dades dels autors.
En general es pot dir que els canvis més importants que es reflecteixen en els mapes de distribució es deuen, simplement, a un coneixement més exacte del territori, tot i que també n’hi ha hagut de taxonòmics que, lògicament, han modificat la distribució. En pocs casos, però, s’ha produït un augment real significatiu de l’àrea de distribució d’una espècie. Entre les novetats taxonòmiques i faunístiques més destacables cal esmentar especialment el fet que l’augment del nombre de dades, cada cop més exactes, sobre la distribució dels amfibis, ha coincidit amb dues circumstàncies rellevants: d’una banda, la unificació de les bases de dades i, de l’altra, la integració d’aquestes bases en entorns web. Per exemple, la base de dades de l’Atlas y Libro Rojo de los anfibios y reptiles de España tenia més de 200.000 registres, nombre actualment ultrapassat des que es pot introduir més informació a través d’adreces electròniques especialitzades. La centralització de la informació ha permès elaborar mapes més complets i unificar l’esforç de prospecció del territori i, alhora, ha fet possible integrar aquesta informació amb altres dades ambientals mitjançant els sistemes d’informació geogràfica (SIG). D’aquesta manera s’han pogut elaborar mapes de distribució potencial de cadascuna de les espècies i esbrinar quins són els condicionaments que en limiten la distribució. Aquestes noves tècniques permeten fer representacions espacials molt precises i són una eina molt adient per a avaluar com la transformació del paisatge i els canvis globals (per exemple, l’escalfament del planeta) poden modificar l’àrea de distribució de cada espècie.
Els estudis ecològics

La presència i el manteniment de punts d’aigua és vital per a la supervivència de les poblacions d’amfibis. A la regió mediterrània tenen una especial importància les basses temporals, ja que permeten el desenvolupament de comunitats d’amfibis d’una gran biodiversitat. Per aquest motiu, la Directiva Hàbitats de la Unió Europea considera hàbitats prioritaris els sistemes aquàtics temporals de la regió mediterrània.
Albert Montori
Aquests darrers anys, l’estudi dels amfibis ha experimentat una evolució notable. Si abans els treballs se centraven en la composició de les comunitats d’amfibis, actualment l’objectiu és comprendre el funcionament d’aquestes comunitats, determinar les interaccions entre els seus components i analitzar la resposta de les espècies a les diferents situacions a què estan sotmeses. A causa del seu peculiar cicle biològic, els amfibis són organismes especialment susceptibles a les alteracions del medi. Les basses i altres nuclis d’aigua els serveixen per a reproduir-se i constitueixen l’hàbitat de les larves des que neixen fins que completen el desenvolupament, mentre que els ambients terrestres són els hàbitats dels juvenils i els adults. Aquest cicle tan complex implica migracions anuals entre ambdós hàbitats, tant dels adults com dels juvenils. Tenint en compte aquesta migració d’un hàbitat a un altre i la inestabilitat temporal de molts dels ambients aquàtics utilitzats per a la reproducció, es pot dir que els amfibis funcionen com un sistema de metapoblacions en el qual la distribució de les espècies canvia freqüentment d’un any a l’altre, fins i tot en ambients no alterats. Això és el que s’ha observat, per exemple, en un estudi dut a terme durant quatre anys consecutius als parcs naturals de Collserola i el Garraf. Aquest treball demostra que totes les espècies experimenten extincions locals però, alhora, colonitzen nous punts d’aigua. Aquestes taxes d’extinció-colonització varien en funció del tipus d’ambient que utilitza cada espècie per a reproduir-se. Les espècies d’ambients més inestables, com el gripau corredor (Bufo calamita) i el gripauet (Pelodytes punctatus), són les que mostren uns índexs d’extinció-colonització més elevats.

Molt lligat a les basses naturals, l’ofegabous o tritó meridional (Pleurodeles waltl) ha vist com els darrers anys la seva àrea de distribució s’anava fragmentant, especialment a Alacant i a València, cosa que posa en perill la viabilitat de moltes de les poblacions d’aquesta espècie.
Vicent Sancho
En ambients fragmentats, aquesta dinàmica no es pot dur a terme, perquè el medi és poc permeable a la migració, i aleshores les àrees on es produeixen extincions locals no tornen a ser recolonitzades posteriorment. Com a conseqüència, a llarg termini les poblacions van quedant aïllades les unes de les altres i es tornen més susceptibles a l’extinció. Aquest procés és el que molt possiblement ha tingut lloc a la serra de Collserola, on les dues principals xarxes viàries que travessen el parc natural han deixat aïllada una zona central. Però és obvi que aquest fenomen també es dóna en molts altres indrets dels Països Catalans on el creixement humà ha fragmentat les poblacions i les ha abocat sense remei a l’extinció. El tritó meridional, o ofegabous (Pleurodeles waltl), n’és un exemple.
A València, l’excessiva fragmentació de les seves poblacions ha ocasionat pràcticament la desaparició de l’espècie a la franja litoral. El fet que totes les espècies mostrin un patró de distribució semblant permet sostenir la hipòtesi que la presència de carreteres i altres zones impermeables als amfibis dificulta la recolonització dels ambients aquàtics existents a les zones isolades.
Un altre aspecte molt interessant de la dinàmica de les comunitats d’amfibis és el fet que, en contra del que caldria esperar, les espècies amb una taxa i una capacitat de dispersió més grans són les més afectades per la fragmentació. Les espècies que s’allunyen poc dels seus centres reproductors i s’hi mantenen fidels tenen menys probabilitats de desaparèixer quan el paisatge es fragmenta, en varia la permeabilitat i les distàncies entre les localitats habitades s’incrementa. Si es té en compte això, els bufònids (gripaus típics) s’haurien de considerar el grup més sensible a aquesta alteració del paisatge, ja que són organismes amb una gran mobilitat anual. No és una coincidència que els gripaus siguin els amfibis que més atropellaments pateixen a les carreteres.
Actualment les poblacions d’amfibis ja es gestionen tenint en compte aquest funcionament de metapoblacions, i també la importància de les xarxes de basses i dels ambients adequats. D’aquesta manera és més fàcil assegurar la continuïtat i el manteniment de les biocenosis d’amfibis. Per a poder gestionar les comunitats de batracis, en primer lloc cal saber on es localitzen els punts d’aigua i les característiques de cadascun (naturalesa, temporalitat, etc.). Cal mantenir l’heterogeneïtat dels ambients aquàtics, ja que cada espècie té unes preferències diferents. La major part de les espècies tenen prou plasticitat per a poder desenvoluparse en ambients variats, tot i que les taxes de supervivència i l’eficàcia del creixement i el desenvolupament varien segons l’indret. Per exemple, hi ha espècies típiques de basses efímeres i temporals, com el gripau corredor i el gripauet, que exploten amb gran eficàcia aquests ambients. En canvi, altres espècies d’amfibis prefereixen basses permanents, com el gripau comú (Bufo bufo) i la granota verda (Pelophylax perezi [=Rana perezi]), i encara d’altres colonitzen les aigües oligotròfiques de pous o fonts, com la salamandra (Salamandra salamandra) i el tòtil (Alytes obstetricans). La pèrdua de l’heterogeneïtat ambiental pot alterar les interaccions entre espècies i, a mitjà termini, podria modificar l’estructura de les comunitats. És per això que els inventaris d’ambients aquàtics han d’incloure els ambients temporals, com les zones inundables i les basses temporals, generalment ignorades per les lleis de protecció i les administracions. Aquests ambients no solament ajuden a reduir la distància entre basses permanents i afavoreixen la connexió entre poblacions, sinó que són l’hàbitat principal de diverses espècies.
Però és evident que també cal conèixer les característiques de les diferents espècies que constitueixen la comunitat i les limitacions que els imposen els diversos ambients. Un cop coneguda la xarxa d’ambients aquàtics disponibles, cal establir les característiques que presenten. En un espai determinat hi ha un seguit de punts d’aigua que es poden classificar en gradients. Per exemple, si es considera la permanència d’aigua, es poden classificar els ambients aquàtics en efímers, temporals o permanents. No es tracta pas d’una categorització rígida, ja que la distribució i l’abundància dels diferents medis aquàtics poden variar d’un any a un altre en funció de la climatologia. És evident que la temporalitat del medi aquàtic és un factor limitant molt important per a les espècies d’amfibis, ja que obliga els capgrossos a completar la metamorfosi en un temps determinat. Lògicament, la biologia i la història evolutiva de cada espècie de la comunitat d’amfibis determinaran si és capaç o no de colonitzar els medis efímers o temporals. Aleshores, per què no es decanten pels medis permanents, més estables, on la continuïtat de l’aigua està assegurada? Els medis efímers són clarament més impredictibles, però, d’altra banda, presenten un factor avantatjós respecte dels medis permanents: l’absència de comunitats de predadors. Els medis permanents mantenen sempre comunitats aquàtiques molt més madures i, en conseqüència, hi viuen comunitats de depredadors estructurades i de mida més gran. Els medis permanents acostumen a mantenir comunitats de peixos i de macroinvertebrats que no poden viure en medis temporals i efímers. Per això, cada espècie segueix una o altra estratègia reproductora en funció de les característiques biològiques que li són pròpies. Quan s’estudien les comunitats, s’observa com les espècies es distribueixen en gradients, i la major riquesa específica correspon als valors intermedis del gradient, on podria semblar que aquestes pressions són més tènues. No obstant això, a les zones centrals dels gradients apareix un factor nou: la competència interespecífica. En resum, si en un extrem del gradient tenim un major risc de dessecació i un menor risc de mort per depredació, a l’altre extrem del gradient hi ha un menor risc de dessecació i un major risc de depredació, mentre que a la zona intermèdia del gradient augmenta la competència interespecífica.
Les diferents espècies d’amfibis presenten, també, estratègies reproductores ben diferenciades, que no es poden deslligar dels aspectes que hem comentat. Alguns anurs tenen períodes reproductors extremament dilatats i es poden reproduir al llarg de tot l’any. Aquest és el cas del gripauet, que pot pondre ous tot l’any i amb aquesta estratègia augmenta la probabilitat de supervivència. Però també hi ha espècies amb un període reproductor molt curt, com la granota roja (Rana temporaria) i el gripau comú, que realitzen una única posta a l’any formada per nombrosos ous. Per tant, les variades estratègies de reproducció dels amfibis són determinades per tres factors que interactuen entre ells: el risc de dessecació, el risc de depredació i la competència interespecífica i intraespecífica durant els estadis larvals.
Els estudis sobre conservació

El tòtil (Alytes obstetricans) és una de les espècies d’amfibis més afectades pel fong Batrachochytrium dendrobatidis, que a Peñalara (Madrid) i als Pirineus ja ha causat extincions en massa. Una altra amenaça que també l’afecta més que a altres amfibis és la contaminació i la pèrdua d’hàbitats aquàtics, ja que, en tenir un període larval molt llarg, els capgrossos es veuen obligats a passar l’hivern a l’aigua i fer la metamorfosi a la primavera següent.
Albert Montori.
L’any 1987 la legislació existent en matèria de medi ambient era tot just incipient i als Països Catalans encara no es donava una importància especial a la conservació de les espècies d’amfibis ni als seus hàbitats. La prioritat fonamental era conèixer la fauna de batracis present al territori, i tot just es començaven a entreveure els greus problemes ambientals que l’afecten. A més, les administracions eren molt poc sensibles a aquests problemes. Actualment la situació ha canviat molt, i els riscos a què estan sotmesos els amfibis s’han diversificat i incrementat de forma gairebé exponencial. Els darrers cinquanta anys, i de manera més accentuada les dues darreres dècades, la biodiversitat general s’ha reduït molt arreu del món i això, en el cas dels vertebrats, ha significat el declivi i la desaparició d’espècies i de poblacions naturals. Entre tots els vertebrats, els amfibis són el grup més afectat per aquest declivi. Això és perquè, si es comparen amb altres grups de vertebrats, els amfibis tenen taxes de dispersió i colonització menors. A més, presenten limitacions ecològiques i fisiològiques importants, ja que, en dependre tant del medi terrestre com del medi aquàtic per a completar el cicle vital, són molt sensibles a l’alteració de l’ambient.
Per a explicar el declivi de les poblacions d’amfibis s’han proposat diferents causes: les malalties emergents, la contaminació, la major incidència de la radiació ultraviolada, la pèrdua d’hàbitats, la introducció d’espècies exòtiques i, fins i tot, el canvi climàtic. La Unió Internacional per a la Conservació de la Natura (UICN) ha avaluat recentment l’estat de conservació de totes les espècies d’amfibis del món i ha arribat a la conclusió que el 32,5% estan en perill d’extinció.

En set petits cursos d’aigua verticals i rocallosos del Montseny, on a l’estiu sovint l’aigua només hi circula subterràniament, hi viu una espècie de tritó descoberta recentment: el tritó del Montseny (Calotriton arnoldi). Com que el període larval d’aquesta espècie s’allarga més enllà d’un any, l’extracció d’aigua del Massís per a consum humà en pot posar en risc la supervivència. A més, alguns d’aquests rierols són força accessibles i podria produir-se una extracció il·legal d’exemplars de tritó del Montseny.
Albert Montori.
Els països que voregen la mar Mediterrània alberguen al voltant de 400 milions de persones, i d’aquestes, 135 milions viuen a la costa mediterrània. Això exerceix una pressió notable sobre l’entorn costaner i causa una pèrdua generalitzada de biodiversitat. La pèrdua d’hàbitats, ja sigui perquè s’han destruït o perquè han quedat alterats, és la causa principal de la desaparició d’espècies i poblacions d’amfibis, especialment a les zones periurbanes i costaneres. Per això s’han redactat normes legislatives que intenten invertir aquesta tendència generalitzada. La llei de conservació de la natura vigent l’any 1987 era el Reial Decret 3181/1980, pel qual es protegien totes les espècies d’amfibis menys el gripau comú, la granota verda i la salamandra. Més endavant, el Reial Decret 439/1990 va incloure una nova espècie d’amfibi, el ferreret (Alytes muletensis), que no hi constava a causa de la seva recent descoberta, i també li va conferir la categoria d’espècie protegida. A més, es va publicar el primer Catàlogo Nacional de Especies Amenazadas (CNEA). L’any 1989, el Parlament espanyol va aprovar la Llei 4/1989 de conservació dels espais naturals, que corregia les deficiències de la Llei de parcs naturals i l’ajustava a la Constitució del 1978. Aquest canvi de perspectiva va ser important, ja que, per primer cop, es parlava explícitament de la conservació dels hàbitats. Cal tenir en compte que el 1987 (any en què es va editar el volum de la Història Natural dedicat als amfibis, els rèptils i els mamífers) a l’Estat espanyol tot just acabava d’entrar en vigor el conveni de Berna (1 de setembre de 1986). Aquest conveni va ser el primer intent de regular la conservació de les espècies i els hàbitats a Europa. Sí que és veritat que ja existia el conveni CITES, conegut també com a conveni de Washington (1973), que intentava controlar el mercat internacional d’espècies salvatges i la seva explotació comercial –cosa que aconseguia amb poc o molt d’èxit–. Un pas endavant fonamental en la conservació dels hàbitats i les espècies el constitueix la Directiva Hàbitats 92/43/CEE, que incorpora fins a cinc annexos de conservació on s’inclouen els hàbitats naturals i la fauna i la flora silvestres protegides dins l’àmbit de la Unió Europea. Aquesta Directiva s’adequa a la normativa espanyola amb el Reial Decret 1997/1995. Actualment, a l’Estat espanyol, la llei vigent que han de complir totes les comunitats autònomes és la Llei 42/2007 del patrimoni natural i de la biodiversitat. Amb aquesta llei es modifica el CNEA, que tan sols inclou les espècies en perill i vulnerables. El CNEA s’incorpora al Listado de especies silvestres en régimen de protección especial, que inclou les espècies que abans es catalogaven com d’interès especial, insuficientment conegudes i gairebé amenaçades. Aquesta llei ha representat un salt qualitatiu, atès que fa prevaler la protecció de la natura, els hàbitats i les espècies per sobre els usos del territori. A més, dóna protecció legal a totes les zones incloses dins la Xarxa Natura 2000. Totes les comunitats autònomes tenen transferides les competències en matèria de conservació i estan obligades a complir les directives estatals.

La realització de grans obres d’infraestructura sovint comporta, entre altres coses, grans moviments de terres, que tenen un impacte directe sobre la fauna en general i sobre les poblacions d’amfibis en particular.
Marc Franch.
A Catalunya, la llei vigent és el Decret legislatiu 2/2008, pel qual s’aprova el text refós de la Llei de protecció dels animals. Aquest decret manté fora de qualsevol tipus de protecció la granota verda i el gripau pintat (Discoglossus pictus), tot i que aquest darrer és una espècie d’introducció antiga. Al País Valencià, el decret en vigor és el Decret 32/2004 del Consell de la Generalitat, amb el qual es crea el Catàleg valencià d’espècies de fauna amenaçades. Aquest catàleg no considera cap amfibi en perill d’extinció i només inclou dins la categoria de vulnerables el tritó meridional i el gripau pintat meridional (Discoglossus jeanneae), mentre que classifica com a espècies protegides el gripau comú i la granota verda. A les Balears, el decret vigent en matèria de protecció dels amfibis és el Decret 75/2005, pel qual es crea el Catàleg balear d’espècies amenaçades i d’especial protecció. Aquest catàleg incorpora elements diferenciadors amb relació a altres catàlegs en el sentit que, tot i que l’Estratègia Europea d’Espècies Introduïdes (EEEI) considera improcedent aplicar mesures de conservació sobre les espècies potencialment invasores, el Govern Balear diferencia entre les espècies d’introducció antiga –que fins i tot poden mostrar una diferenciació genètica important– i les espècies d’introducció recent. Per això, el calàpet o gripau verd (Bufo balearicus [=B. viridis balearicus]) i la reineta (Hyla meridionalis) consten en aquesta relació com a espècies d’interès especial, tot i tractarse d’espècies introduïdes. A la Franja, és el Decret 48/1995 el que regula el Catàleg d’espècies amenaçades d’Aragó, i al Rosselló-Vallespir ho fa l’Arrèté du 19 de novembre de 2007, llei d’abast nacional que fixa la llista de les espècies d’amfibis i rèptils protegides a tot el territori francès i determina les diverses modalitats de protecció. Cal destacar els anomenats llibres vermells, que especifiquen el nivell d’amenaça de cada espècie. Bé que no tenen valor legal, són utilitzats normalment com a obres de referència, ja que han estat elaborats per especialistes.

Els ous i les larves dels amfibis són molt sensibles a la contaminació química de l’aigua. Concentracions de nitrats, nitrits i fosfats que són tolerades per l’espècie humana resulten letals per a la major part d’amfibis i sovint són responsables de la desaparició de les poblacions de batracis de les zones agrícoles i industrials. La posta de la fotografia és de gripauet (Pelodytes punctatus).
Albert Montori.
Un altre aspecte important en el camp de la conservació són els plans de gestió i recuperació d’hàbitats i espècies. Durant els darrers vint-i-tres anys, als Països Catalans s’han desenvolupat o executat importants projectes de gestió de les poblacions d’amfibis. Un dels més coneguts és el dedicat al ferreret, que ha esdevingut un bon exemple de pla de conservació per la seva eficàcia. Des del 1981, any que es va descobrir el ferreret, s’han desenvolupat uns quants programes destinats a preservar i recuperar aquesta espècie: s’han protegit les localitats on era present (emparades en la Xarxa Natura 2000 de la UE), s’han eliminat selectivament les espècies introduïdes que el depredaven (especialment la serp d’aigua i la granota verda), i, sobretot, s’ha criat en captivitat per poder reforçar les poblacions existents i introduir-lo en noves localitats. A més, s’ha intentat conscienciar la societat sobre el valor patrimonial d’aquesta espècie. La població total de larves de ferreret s’ha incrementat: de les 14.915 existents el 1991 es va passar a les 24.365 del 2006 (amb un recompte màxim el 2004 de 30.052 larves). Això ha estat possible, bàsicament, gràcies a la introducció del ferreret en localitats noves. L’àrea d’ocupació d’aquest anur s’ha incrementat un 100%. La UICN ha reflectit aquest èxit: si abans considerava l’espècie en perill, a la darrera edició del Red Data Book (2008) la qualifica de vulnerable. Malauradament, l’espècie està amenaçada per un nou factor negatiu: algunes de les seves poblacions han estat infestades pel fong Batrachochytrium dendrobatidis, provinent d’installacions de cria d’amfibis. Per sort, aquest fong, que ocasiona mortalitats molt elevades entre les poblacions de tòtil, sembla que no afecta gaire el ferreret. Actualment s’està desenvolupant el segon pla de recuperació de l’espècie, que està previst finalitzar el 2012.
També a les Balears, s’estan executant plans de gestió per a afavorir l’expansió del calàpet o gripau verd. Amb aquest objectiu s’ha dut a terme el projecte Life Basses. Conservació i Gestió de Basses Temporals a Menorca, que té com a finalitat crear les eines de gestió adequades per a la conservació de vint-i-sis basses d’aquesta illa.
A Catalunya, des de l’any 2006 s’està duent a terme el projecte de conservació del tritó del Montseny (Calotriton arnoldi), que té com a finalitat garantir la supervivència d’aquest urodel. L’objectiu principal és la reproducció en captivitat, per tal que, en cas de desaparició de les poblacions lliures, sigui possible crear noves poblacions de tritó, un cop eliminades les causes que les van fer desaparèixer. De moment s’ha aconseguit la reproducció en captivitat de l’espècie i es té intenció d’alliberar exemplars al medi, però no està clar si les causes del declivi de les poblacions lliures s’han corregit.
A Tarragona s’està portant a terme un pla de seguiment de les poblacions de tritó meridional, que té com a objectiu determinar l’estat actual de les poblacions i detectar les causes de rarefacció de l’espècie per tal d’estudiar la manera d’eliminar-les.
Al País Valencià, el projecte de conservació d’amfibis més important realitzat fins ara ha estat l’anomenat LIFE. Restauració d’Hàbitats Prioritaris per als Amfibis, cofinançat amb fons europeus i del Govern valencià. El projecte ha restaurat vora un centenar de punts d’aigua d’interès per als amfibis inclosos dins la Xarxa Natura 2000, ha desenvolupat plans de conservació d’amfibis amenaçats i ha engegat una àmplia campanya d’educació ambiental.
D’altra banda, als Països Catalans s’estan duent a terme diversos projectes que es fonamenten en el voluntariat (anomenats també citizen science). Entre aquests projectes cal destacar el projecte SAC (Seguiment d’Amfibis de Catalunya) i el projecte SARE (Seguiment d’Amfibis i Rèptils d’Espanya). Aquesta mena de projectes recolzen, fonamentalment, gràcies al desenvolupament d’internet, que permet als participants comunicar-se i interactuar ràpidament. En la mateixa línia, el gran nombre de continguts herpetològics apareguts a la xarxa han permès acostar els amfibis a la població, amb el benefici consegüent per a la seva conservació.
Són molts els elements que actuen negativament sobre les poblacions d’amfibis i que en produeixen el declivi. La pèrdua de l’hàbitat, juntament amb la seva fragmentació, és considerada una de les principals causes del decreixement de la biodiversitat. La pèrdua d’hàbitats naturals i l’aïllament dels que resten (per explotació forestal, creació d’àrees agrícoles o prats, construccions urbanístiques, xarxes viàries i ferroviàries, línies elèctriques, etc.) augmenta el risc d’extinció perquè redueix la mida de les poblacions a nivells en què el nombre d’individus no és prou alt per a mantenir-les.
Els incendis forestals són una de les pertorbacions que comporten un canvi més dràstic i ràpid del medi natural, el qual situen en una fase molt inicial de la successió ecològica. Actualment el 90% dels incendis forestals que es declaren a la regió mediterrània estan relacionats amb l’activitat humana. Cada any, al conjunt de la conca mediterrània es produeixen uns 50.000 incendis forestals que cremen un milió d’hectàrees aproximadament.
La pèrdua de cobertura vegetal comporta una disminució de la humitat del sòl, cosa que perjudica els amfibis, que depenen molt de la humitat. Les espècies més terrestres, com el gripau corredor i el gripau d’esperons (Pelobates cultripes), i les espècies forestals, com el gripau comú i la salamandra, poden veure’s molt afectades per la pèrdua de cobertura vegetal. La reineta és l’espècie més vulnerable als incendis forestals. Com que aquest amfibi habita freqüentment entre la vegetació i és una espècie termòfila i activa durant el dia, presenta una mortalitat directa pel foc molt més alta que la de les altres espècies d’amfibis. La granota verda i el gripauet són les espècies menys afectades pels incendis forestals, probablement gràcies als hàbits essencialment aquàtics de la primera i cavernícoles de la segona.
L’activitat agrícola té diversos efectes negatius. En primer lloc, homogeneïtza els hàbitats i, per tant, disminueix la diversitat vegetal; en segon lloc, sovint contamina greument els aqüífers amb plaguicides i fertilitzants, cosa que fa impossible la supervivència dels amfibis. La pèrdua d’hàbitat és especialment greu en les monocultures extensives, i es reflecteix, també, en una pèrdua de riquesa d’espècies. Aquest és el cas de les grans extensions dedicades als cereals i als fruiters de Lleida, la Franja de Ponent i València, on moltes de les espècies d’amfibis que les habitaven pràcticament han desaparegut. Per altra part, l’acidificació i la contaminació de l’aigua augmenten la mortalitat dels amfibis, especialment durant les fases aquàtiques. El nitrat d’amoni, un dels productes més utilitzats com a fertilitzant arreu del món, fa augmentar els nivells ambientals de ions nitrogen en masses d’aigua properes als punts d’aplicació. Les concentracions ambientals anormalment altes de nitrat d’amoni poden provocar una elevada mortalitat larval o reduir les taxes de creixement dels embrions i de les larves de diverses espècies d’amfibis. Aquest és un dels factors que podrien explicar la desaparició de la major part dels amfibis dels deltes del Llobregat i de l’Ebre. A més, cal tenir en compte que els nivells màxims de nitrats i nitrits acceptats per a l’aigua depurada són també letals per a les larves de moltes espècies d’amfibis.
Cal no oblidar, però, que si bé l’agricultura i la ramaderia tradicionals van afavorir la presència dels amfibis en crear basses de rec, abeuradors i canals, el canvi d’usos i costums ha produït la progressiva desaparició o degradació dels punts d’aigua. Durant els darrers vint anys, als Països Catalans s’ha pogut detectar una alteració progressiva i una pèrdua constant de molts punts d’aigua. Aquest efecte va lligat a la millora de l’eficàcia del reg (substitució del reg per inundació o de tesa pel de degoteig) i a la reducció de l’activitat ramadera i agrícola tradicional. Però també és evident que els abocaments, la modificació del medi, la dessecació de punts d’aigua per sobreexplotació dels aqüífers i les activitats d’oci hi han tingut molt a veure.

Les vies de comunicació actuen com a barreres que impedeixen el pas dels amfibis, i fragmenten i aïllen les poblacions. Les conseqüències són diverses. D’una banda, molts amfibis moren atropellats quan intenten arribar als punts de reproducció. De l’altra, la fragmentació poblacional redueix el nombre d’individus i impedeix el flux gènic, la qual cosa provoca extincions.
Albert Montori.
D’altra banda, les estructures viàries, que són un element cada vegada més comú i abundant al nostre entorn, tenen una influència cabdal sobre les poblacions i els ecosistemes. Per a poder analitzar els efectes de la xarxa viària en l’entorn natural és important que es conegui l’impacte ecològic de les carreteres. En primer lloc, causen la mort directa de molts exemplars d’amfibis per atropellament, com també una mortalitat indirecta per la destrucció de l’hàbitat de les diferents espècies, que no troben llocs per a reproduir-se ni alimentar-se. En segon lloc, tenen un efecte barrera, ja que divideixen les àrees de distribució dels organismes en zones més petites que queden separades per estructures sovint infranquejables. Això impedeix l’accés a l’aliment i a les zones de reproducció i impossibilita el flux gènic entre poblacions. Estudis realitzats a Catalunya per a avaluar l’impacte de la xarxa viària secundària sobre els amfibis han posat de manifest que les espècies que pateixen més els efectes dels vehicles són el gripau comú, la salamandra, el gripau corredor i la reineta.
Un altre factor responsable del declivi de les poblacions d’amfibis són les anomenades malalties emergents. Es tracta de malalties que han aparegut recentment o bé de malalties antigues el nivell d’incidència de les quals ha augmentat de manera dràstica en poc temps. En el cas dels amfibis, els agents causals d’aquestes malalties són determinats tipus de virus i fongs. El 1997 es va detectar a Espanya el primer cas de quitridiomicosi, una malaltia fúngica produïda per Batrachochytrium dendrobatidis que actualment ja és present en set països europeus. A Espanya, on s’ha detectat pràcticament a totes les comunitats autònomes, la quitridiomicosi ha causat mortalitats molt elevades de tòtil a Astúries, Lleó, Zamora, Osca i Madrid. També s’han registrat mortalitats elevades de gripau comú, salamandra i granota verda.
Un altre problema important són les introduccions i les translocacions d’espècies. L’any 2007 es va detectar a Menorca una població de tòtil, introduïda accidentalment en un viver de plantes ornamentals. La presència de l’espècie a l’illa va ser valorada negativament, ja que hi era previst reintroduir-hi el ferreret menorquí (Alytes talaioticus, conegut com a subfòssil d’època romana i considerat avui sinònim d’A. muletensis). La detecció precoç d’aquesta població introduïda va permetre eliminar-la, segons sembla, amb èxit. A Catalunya s’ha detectat recentment la granota africana d’ungles (Xenopus laevis) a dues localitats dels voltants de Barcelona. Una de les poblacions d’aquesta granota probablement ha estat eradicada, però l’altra segurament no. A més, es té constància de la presència d’exemplars aïllats de granota toro (Aquarana catesbeiana) i tritó de ventre de foc (Cynops pyrrhogaster) a Collserola. També són conegudes les translocacions de tritó jaspiat o verd (Triturus marmoratus) i tritó palmat (Lissotriton helveticus [=Triturus helveticus]) a Collserola, i les de gripau pintat a Collserola i al delta del Llobregat.
La taula següent presenta la llista patró dels amfibis presents als Països Catalans i estat de conservació de cadascun. Hi figuren els noms científics aprovats en la Lista patrón de los anfibios y reptiles de España i, entre parèntesis, els reconeguts amb anterioritat. A absent, CR en perill crític, DD dades insuficients, DE d’especial protecció, DIE d’interès especial, EN en perill, LC preocupació menor, NE no avaluada, NT gairebé amenaçada, P protegida, VU vulnerable. (*) i (−) indica que no estan protegides o són absents al territori corresponent. UICN Unió Internacional per a la Conservació de la Natura, CNEA Catálogo Nacional de Especies Amenazadas.
Amfibis presents als Països Catalans
| Espècie | Nom comú | Catalunya | llles Balears | País Valencià | La Franja | Rosselló / Vallespir | UICN (2008) | CNEA (2006) | Libro Rojo (2002) |
|---|---|---|---|---|---|---|---|---|---|
| Calotriton arnoldi (Euproctus asper) | Tritó del Montseny | EN | A | A | A | A | CR | NE | NT |
| Calotriton asper (Euproctus asper) | Tritó pirinenc | DIE | A | A | DIE | NT | NT | DIE | NT |
| Salamandra salamandra | Salamandra | DIE | A | A | DIE | LC | LC | NE | VU |
| Triturus marmoratus | Tritó jaspiat o verd | DIE | A | A | DIE | LC | LC | DIE | LC |
| Lissotriton helveticus (Triturus helveticus) | Tritó palmat | DIE | A | A | DIE | LC | LC | DIE | LC |
| Pleurodeles waltl | Ofegabous o tritó meridional | VU | A | VU | DIE | A | NT | DIE | NT |
| Discoglossus pictus | Gripau pintat septentrional | NE * | A | A | A | DD | LC | NE | LC |
| Discoglossus jeanneae | Gripau pintat meridional | A | A | VU | DIE | A | NT | DIE | NT |
| Alytes muletensis | Ferreret | A | EN | A | A | A | VU | EN | CR |
| Atytes obstetricans | Tòtil | DIE | A | DIE | DIE | LC | LC | DIE | NT |
| Alytes dickhyllenii (Alytes obstetricans) | Tòtil | A | A | DIE | A | A | VU | NE | VU |
| Pelobates cultripes | Gripau d’esperons | VU | A | DIE | DIE | VU | NT | DIE | NT |
| Pelodytes punctatus | Gripauet | DIE | A | DIE | DIE | LC | LC | DIE | LC |
| Bufo bufo | Gripau comú | NE | A | P | DIE | LC | LC | NE | LC |
| Bufo calamita | Gripau corredor | DIE | A | DIE | DIE | LC | LC | DIE | LC |
| Bufo balearicus (Bufo viridis) | Gripau verd o calàpet | A | DIE | A | A | A | LC | DIE | VU |
| Hyla meridionalis | Reineta | DIE | DIE | A | DIE | LC | LC | DIE | NT |
| Rana temporaria | Granota roja | DIE | A | A | DIE | LC | LC | DIE | LC |
| Pelophylax perezi (Rana perezi) | Granota verda | NE * | NE * | P | DIE | NT | LC | NE | LC |
| Pelophyiax kl. grafi (Rana perezi) | Granota verda hibridogenètica | NE * | A? | A | A? | DD | NT | NE | − |
L’actualització taxonòmica dels amfibis
El coneixement dels batracis dels Països Catalans, i d’arreu del món, presenta moltes novetats, que s’aniran comentant espècie per espècie.
Pel que fa a la salamandra, algunes de les 14 subespècies que se n’havien descrit fins ara han estat elevades a nivell d’espècie (Salamandra algira, S. inframaculata i S. corsica). D’altres continuen considerant-se subespècies però la seva àrea de distribució s’ha pogut precisar més clarament, com en el cas de S. s. bejarae. Aquesta subespècie es creia present a Catalunya, però actualment es considera que se circumscriu als massissos muntanyosos del centre de la península Ibèrica. La subespècie de salamandra present als Països Catalans és S. s. europaea, bé que es discuteix encara la presència de S. s. fastuosa, ja que aquesta subespècie és vivípara, i a la Vall d’Aran s’han observat larves de salamandra a l’aigua. Als Països Catalans S. s. europaea és present a Andorra, Catalunya, el Llenguadoc-Rosselló i el nord de la Franja. Falta a tota la Depressió Central Catalana i al sud de les muntanyes de Prades.

El tritó pirinenc (Calotriton asper) presenta granulacions en abundància per tot el cos, tret que no s’observa en el tritó del Montseny (Calotriton arnoldi). Aquesta i altres diferències morfològiques i genètiques són les que van motivar els investigadors a considerar el tritó del Montseny i el tritó pirinenc dues espècies diferents, bé que prou properes per a classificar-les dins el mateix gènere.
Albert Montori.
El tritó pirinenc, abans inclòs dins el gènere Euproctus, actualment s’ha reclassificat dins el gènere Calotriton i s’ha subdividit en dues espècies diferents: C. asper, per a la qual es manté la denominació popular de tritó pirinenc, i C. arnoldi, batejat com a tritó del Montseny. Aquestes dues espècies segurament es van diferenciar en temps recents, fa entre un milió i un milió i mig d’anys. Se suggereix que l’espècie antecessora d’aquests dos tritons de muntanya ibèrics es va refugiar durant algun període glacial indeterminat en una àrea favorable situada al sud dels Pirineus, àrea que incloïa el Montseny. Aproximadament fa un milió i mig d’anys, va aparèixer una barrera que va impedir el flux gènic entre les poblacions del Montseny i les de la resta de l’àrea de distribució de l’espècie. Aquests dos nuclis van evolucionar per separat fins a donar lloc a les dues espècies del gènere Calotriton reconegudes actualment. Finalment, a les acaballes del Plistocè, fa uns 250.000 anys, una nova barrera va interrompre el flux gènic entre les poblacions situades a ambdues bandes de la Tordera. Això podria explicar les diferències observades entre les subpoblacions de C. arnoldi. Les anàlisis de tres gens mitocondrials indiquen que el grau de diferenciació entre el tritó pirinenc i el tritó del Montseny és del 3,1%, prou elevat per considerar-los dues espècies diferents. Aquesta diferenciació és confirmada per l’anàlisi multivariant de 16 caràcters morfològics i per les diferències osteològiques existents entre ambdues espècies. Les característiques descrites en les altres pàgines de la Història Natural per a Euproctus asper són vàlides per a C. asper. Finalment, cal esmentar la descoberta a les Corberes, a França, d’un grup de poblacions hipogees i epigees de C. asper. Aquest nucli es troba aïllat i, ara per ara, es desconeix si es diferencia genèticament i taxonòmicament de la resta de poblacions de C. arnoldi.
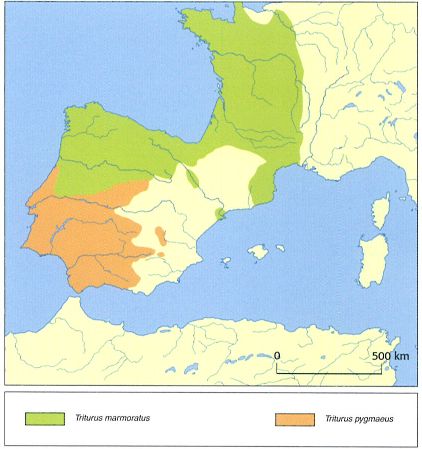
Àrea de distribució de les dues espècies de tritons del gènere Triturus presents a la península Ibèrica: el tritó jaspiat o verd (T. Marmoratus, en verd) i el tritó pigmeu (T. Pygmaeus, en marró). Tot i que existeix un cert grau d’intercanvi gènic, s’ha confirmat que es tracta de dues espècies separades.
IDEM, a partir de dades dels autors.
El tritó jaspiat o verd es considerava dividit en dues subespècies: Triturus marmoratus i T. pygmaeus, que actualment es consideren espècies separades. Segons les dades moleculars disponibles, aquestes dues espècies es van separar fa uns 3,2 milions d’anys, durant el Pliocè, a banda i banda de la vall del Guadalquivir. L’extinció posterior de les poblacions de T. marmoratus del límit meridional i l’expansió cap al nord de T. pygmaeus podria explicar les àrees de distribució actuals. La divergència genètica mitocondrial entre aquestes dues espècies de tritons és del 4,5%, molt semblant a la que s’ha observat entre T. dobrogicus i T. karelinii, que es consideren espècies clarament diferenciades. Als Països Catalans només hi ha T. marmoratus, que presenta dues poblacions ben separades. La més gran, clarament relacionada amb les poblacions del sud de França, s’estén pel Rosselló, les comarques gironines, part de les comarques del Berguedà, Osona, el Bages, el Vallès Oriental i l’Occidental i el Maresme. La segona, relacionada amb les poblacions aragoneses, es troba al Baix Ebre i el Montsià, bé que manca a la plana del delta de l’Ebre. Aquesta distribució suggereix l’existència d’una doble colonització als Països Catalans: una onada que va baixar per la vall de l’Ebre i una altra que provenia del sud-est de França. Cal indicar que existeixen poblacions introduïdes a la serra de Collserola.
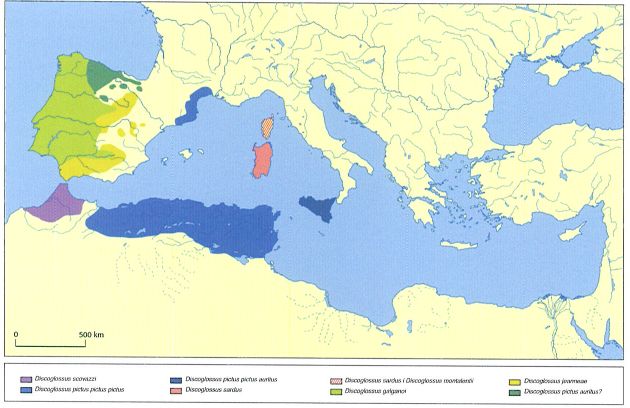
Àrea de distribució del gènere Discoglossus a la regió iberomagriblna i a les illes de la Mediterrània occidental.
IDEM, a partir de dades dels autors.

El gripau pintat septentrional (Discoglossus pictus) és una espècie naturalitzada que arribà als Països Catalans fa més de cent anys procedent del nord d’Àfrica, probablement des d’Algèria.
Albert Montori i Sandra Pareja.
L’any 1987 es reconeixia la presència del gripau pintat als Països Catalans, i s’apuntava que les poblacions del País Valencià podrien correspondre a Discoglossus galganoi, espècie descrita feia poc. Es desconeixia, però, l’origen i l’assignació taxonòmica real de les dues poblacions del gènere Discoglossus presents als Països Catalans. Les poblacions de la part occidental de la península Ibèrica es descrivien com a D. galganoi, una espècie clarament diferent de les espècies africanes. Aleshores els estudis no concretaven quina era la situació taxonòmica de les poblacions valencianes, que quedaven suposadament incloses dins l’espècie D. galganoi, com tampoc la de les poblacions de Catalunya i el Rosselló, que es consideraven D. pictus. L’any 1986, D. galganoi es va escindir en dues espècies, la nominal i D. jeanneae. Malgrat que aquesta darrera espècie fou força discutida, avui dia és acceptada. Més tard, estudis moleculars van demostrar que les poblacions presents a Catalunya i el Rosselló provenien d’Algèria, i s’assignaren a la subespècie D. p. auritus, encara que la validesa de les subespècies D. p. pictus i D. p. auritus també ha estat posada recentment en dubte. Segons aquests mateixos estudis, l’espècie present al País Valencià correspon a D. jeanneae, mentre que l’espècie present a Catalunya i al Rosselló és D. pictus. Per tant, D. pictus és una espècie introduïda, tal com ho plantejava una de les hipòtesi plausibles l’any 1987. El cas del gripau pintat als Països Catalans és, doncs, prou interessant. Totes les dades disponibles indiquen que aquesta espècie va ser introduïda al final del segle XIX o al principi del XX a Banyuls de la Marenda. La primera citació correspon a M. Wintrebert i data de l’any 1906. El 1948, l’espècie ja s’havia estès cap al nord fins a Perpinyà, però no va ser fins el 1957 que se’n va fer la primera observació a Catalunya, concretament a la població de Portbou, a escassament 8 km de Banyuls. Pocs anys després es va localitzar a Girona, Figueres i Empúries, cosa que confirma la colonització del nord-est peninsular per aquesta espècie. Des que es va introduir, el gripau pintat s’ha anat expandint cap al nord-est pel departament dels Pirineus Orientals, i actualment arriba fins al riu Orb, situat a uns 130 km de Banyuls de la Marenda, com també a altres localitats dels departaments de l’Aude i de l’Erau. El riu Orb és, ara per ara, el límit nord-est natural de l’àrea de distribució del gripau pintat, malgrat que recentment s’han trobat poblacions introduïdes aïllades a Lattes i a Vailhauquès, ambdues relativament a prop de Montpeller. Al Rosselló i al Vallespir el gripau pintat ocupa tota la franja litoral i ja s’està expandint cap a l’interior seguint la Tet i el Tec. Cap al sud, i d’acord amb les darreres dades publicades, aquest gripau ocupa la major part de la província de Girona. L’expansió de l’espècie cap a terres meridionals és molt més accentuada i ràpida que cap a l’interior. Actualment s’està expandint a una velocitat mitjana d’1,5 km l’any, de manera que la seva àrea de distribució és cada vegada més gran. Recentment s’ha descobert que ha entrat a la província de Barcelona seguint, d’una banda, la conca de la Tordera i, de l’altra, el litoral de la Selva. Les citacions antigues de l’espècie a Montserrat i algunes localitats del Bages han de ser desestimades. Noves dades obtingudes recentment indiquen que l’espècie també ha estat introduïda al delta del Llobregat, on es troba en expansió, i a Collserola, on s’ha pogut comprovar que s’hí reprodueix. Com qualsevol espècie introduïda, competeix amb les espècies autòctones. Per exemple, s’ha comprovat que les larves del gripau pintat competeixen amb les del gripau corredor i amb les del gripauet, i que els hàbitats de reproducció dels adults s’encavalquen força amb els del gripau corredor.
El tòtil és una altra espècie que també ha experimentat canvis taxonòmics importants. Fa uns quants anys, el gènere Alytes present a la península Ibèrica i a les Balears es considerava subdividit en tres espècies: A. obstetricans, A. cisternasii i A. muletensis. Actualment, estudis moleculars han demostrat les divergències existents entre les poblacions del sud de la Península, que s’han assignat a l’espècie nova A. dickhyllenii, i les poblacions del nord d’Àfrica, que es descriuen com a A. maurus. Aquests estudis també han dividit A. obstetricans en quatre subespècies: A. o. obstetricans, A. o. boscai, A. o. almogavarii i A. o. pertinax. Tan sols les dues darreres són presents als Països Catalans, encara que no es coneix exactament el límit geogràfic que separa una de l’altra. A. o. almogavarii ocupa tot Catalunya, el Rosselló i el Vallespir, possiblement tot Aragó i una àmplia franja de Castella-la Manxa que arriba a Guadalajara i possiblement al nord de Madrid. A. o. pertinax ocupa la costa de llevant, i limita al nord amb l’Ebre i les zones properes de Tarragona, i al sud, amb Múrcia i Albacete. Cap a l’interior s’estén pel sud del Sistema Ibèric i per algunes serres de Castella-la Manxa, i arriba a l’est de Toledo, el sud de Guadalajara i el sud-est de Madrid.
Pel que fa al ferreret, actualment es considera que inclou la forma fòssil menorquina descrita com a Alytes talaioticus. Per tant, aquesta darrera forma és sinònim de l’espècie vivent.
L’any 2000 es va descriure Pelodytes ibericus, una nova espècie del gènere Pelodytes que es troba a la part sud-occidental de la península Ibèrica. Aquesta nova descripció no afecta l’espècie present als Països Catalans, però sí que modifica la distribució peninsular del gripauet. Sembla que les dues espècies es van diferenciar per vicariança durant el Miocè, a ambdues bandes del canal marítim beticorifeny. Estudis realitzats sobre poblacions catalanes de gripauet han determinat que es tracta d’una espècie molt oportunista, amb una gran plasticitat fenotípica i amb un elevat potencial reproductor, ja que és capaç de reproduir-se tot l’any.
També aquests darrers anys, la família dels bufònids o gripaus típics, una de les famílies d’amfibis més àmpliament esteses arreu del món, ha experimentat nombroses revisions, per tal de clarificar-ne la filogènia i la taxonomia. Pel que fa als Països Catalans, s’ha proposat canviar el nom genèric del gripau corredor (Bufo) pel d’Epidalea (E. calamita). Atès que aquest canvi ha estat proposat com una solució temporal i no l’accepten tots els autors, el comitè de taxonomia de l’Asociación Herpetológica Española, que elabora i actualitza una llista patró dels amfibis i els rèptils d’Espanya, ha decidit, de moment, no acceptar-lo.

El gripau verd o calàpet (Bufo balearicus), propi de les Balears, és una de les espècies d’amfibis reconegudes recentment. Fins fa poc les poblacions baleàriques d’aquest gripau s’assignaven a l’espècie Bufo viridis, la mateixa que habita a l’Europa continental. Estudis moleculars han demostrat, però, que les diferències amb les poblacions continentals són prou grans per a considerar-lo una espècie diferent.
Jordi Muntaner.
Passa una cosa semblant amb el gripau verd o calàpet (Bufo viridis). S’ha proposat el canvi de B. viridis per Pseudepidalea viridis. Aquest canvi tampoc no ha estat acceptat per tothom, per motius semblants als adduïts precedentment. Estudis moleculars recents han conclòs que l’espècie de gripau verd present a l’arxipèlag balear és diferent de la resta dels gripaus verds europeus, i ha passat a anomenar-se B. balearicus. Aquests estudis també han dividit B. viridis de l’Europa circummediterrània en cinc espècies diferents. El gripau verd present a les Balears és, en conseqüència, una espècie diferent de l’europea que, a més de les Balears, habita a Còrsega, Sardenya, la península Itàlica i l’extrem nord de l’illa de Sicília, on és simpàtric amb B. siculus. La manca de restes fòssils a les Balears (on el registre paleontològic quaternari és força complet) indica que l’espècie hi va ser introduïda des de les illes de la mar Tirrena.
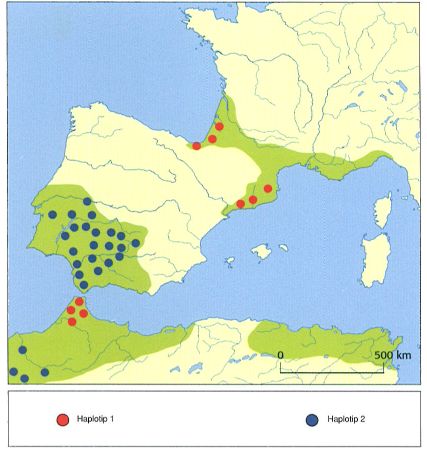
Àrea de distribució dels haplotips de la reineta a la península Ibèrica i al Marroc, que confirma l’existència de dos llinatges d’aquesta espècie. D’altra banda, la poca variabilitat dels haplotips europeus indica que les poblacions de reineta presents a Europa procedeixen del Marroc.
IDEM, a partir de dades dels autors.
Estudis recents sobre la reineta (Hyla meridionalis) demostren que les poblacions presents als Països Catalans són introduïdes. Segons dades obtingudes mitjançant anàlisis moleculars, les reinetes presents a Catalunya, el Rosselló i el Vallespir són semblants a les nord del Marroc i les illes Canàries. A causa de la baixa variabilitat genètica existent en les poblacions del nord-est peninsular, que només disposen d’un sol haplotip, se suposa que es tracta d’una colonització molt recent procedent del nord del Marroc o del nord-oest d’Algèria. Aquesta introducció possiblement es remunta a l’època dels fenicis, i segurament va començar al sud-est de l’actual França, de manera semblant al que va passar amb el gripau pintat al segle XX. La reineta també ha estat introduïda a l’illa de Menorca, on viu des del segle XIX. En canvi, se’n descarta la presència al País Valencià, encara que s’ha citat en localitats del sud de Tarragona molt properes a Castelló. La situació és semblat a Aragó, amb citacions molt pròximes al límit administratiu d’aquesta comunitat, però sense que encara se’n pugui confirmar la presència.
En un altre pàgina de la Història Natural es descriu la reineta septentrional i s’afirma que aquest amfibi s’escampa, de manera dispersa, per la meitat septentrional de Catalunya i el sud-est del Rosselló.
Els autors d’aquesta actualització creiem que aquest amfibi mai no ha estat present als Països Catalans i que les citacions adscrites a aquesta espècie són fruit d’errors d’identificació, atès que s’assembla molt a la reineta comuna o meridional. Tot i que en el proper catàleg d’espècies amenaçades de Catalunya probablement aparegui com a extingida, creiem que no hi ha estat mai present i que els pocs exemplars de les col·leccions existents, mal etiquetats, no procedeixen dels Països Catalans. D’altra banda, una suposada extinció per competència seria molt estranya, ja que en altres indrets de la península Ibèrica les dues reinetes són simpàtriques i es mantenen mecanismes d’exclusió reproductora que minimitzen la hibridació. A més a més, la reineta septentrional podria haver sobreviscut en indrets de caràcter més centreuropeu on la reineta comuna o meridional no és capaç de sobreviure.
Les granotes del gènere Rana també han experimentat canvis importants. Per un cantó, les anomenades granotes verdes europees han passat a pertànyer al gènere Pelophylax, considerat abans com un subgènere, de manera que només les granotes brunes conserven el nom genèric de Rana. La inconsistència del gènere Rana, considerat un grup parafilètic, ha estat la causa d’aquests canvis. Per tant, la granota verda ha passat a anomenar-se Pelophylax perezi. D’altra banda, es considera que aquesta espècie no és present al nord d’Àfrica, ja que s’ha comprovat que les granotes verdes d’aquella zona són morfològicament i genèticament diferents de les de la Península (per això s’han classificat com a P. saharicus).
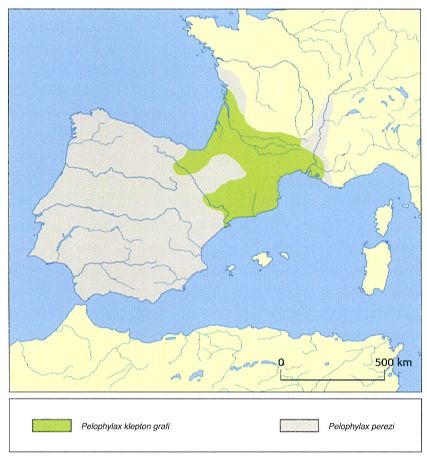
Àrea de distribució de la granota verda (Pelophylax perezi, en gris) a la península Ibèrica i de les poblacions hibridogenètiques d’aquesta espècie amb P. ridibundus, conegudes com a Pelophylax klepton grafi (en verd). Les dades disponibles indiquen que les granotes hibridogenètiques no s’allunyen gaires quilòmetres de l’Ebre, però es tracta de dades antigues que caldria actualitzar. També s’hauria de comprovar si s’han introduït altres espècies críptiques als Països Catalans per mitjà de les granges de cria de granotes.
IDEM, a partir de dades dels autors.
Pelophylax perezi, com altres granotes verdes europees, presenta un procés evolutiu, poc comú entre els vertebrats, que és regulat per un mecanisme reproductor denominat hibridogènesi. Aquest procés es caracteritza perquè dues espècies es reprodueixen i donen lloc a híbrids que, en general, no tenen descendència viable si es reprodueixen entre ells. Tanmateix, aquests híbrids poden retroencreuar-se amb una de les espècies parentals i donar lloc a nous híbrids. Aquests nous híbrids es denominen hibridogenètics, ja que procedeixen de l’encreuament entre un exemplar d’una de les dues espècies parentals i l’híbrid. Els exemplars hibridogenètics presenten una meiosi particular. Durant la gametogènesi, o formació dels gàmetes, una de les dotacions cromosòmiques parentals es perd i, per tant, cada gàmeta conté un sol genoma parental pur, que es transmet clònicament. El genoma exclòs correspon al de l’espècie simpàtrica. El conjunt de les espècies parentals i l’híbrid rep el nom de klepton. Les formes hibridogenètiques procedents de la hibridació de P. ridibundus amb P. perezi es coneixen amb el nom de P. klepton grafi. Aquest klepton està inclòs en l’annex III del conveni de Berna, i la UICN el considera gairebé amenaçat, ja que la seva àrea de distribució s’ha reduït un 30% els darrers deu anys, a conseqüència de la competència amb P. ridibundus, espècie invasora que ha estat introduïda en diversos indrets de França. P. kl. grafi és pràcticament indistingible morfològicament de P. perezi; només se n’ha dit que pot assolir mides més grans, que té les membranes interdigitals una mica més desenvolupades i el tubercle metatarsià també més gran.
Pelophylax klepton grafi es distribueix pel terç inferior de França i el quadrant nord-est de la península Ibèrica. Als Països Catalans es troba pràcticament a tot Catalunya, el Rosselló, el Vallespir i la Franja, i només manca als indrets on P. perezi no és present, com per exemple, a la Vall d’Aran. S’han detectat altres poblacions hibridogenètiques a Saragossa (vall de l’Ebre), a Navarra (Altsasu) i al País Basc (Arrona, Irun), i s’ha comprovat que a la part navarresa de l’àrea de distribució, l’híbrid ha creuat l’Ebre cap al sud. És probable que l’entrada de l’híbrid a la Península s’hagi realitzat per les parts baixes dels Pirineus orientals i occidentals i per les valls que s’obren a França (Cerdanya). Sembla que P. kl. grafii s’ha anat estenent seguint la vall de l’Ebre. El fet que no s’estengui més cap al sud es pot atribuir a una menor eficiència del genoma de P. ridibundus en climes més temperats.
Cal desestimar la presència de la granota àgil (Rana dalmatina) als Països Catalans, ja que les citacions d’aquesta espècie han estat sempre dubtoses, mai no s’han pogut confirmar i generalment es feien en ambients poc favorables per a l’espècie. Passa el mateix amb la granota pirinenca (R. pyrenaica). Les prospeccions realitzades aquests darrers tres anys a la Franja i al Principat a la recerca de poblacions d’aquesta granota no han donat cap resultat positiu. Actualment les poblacions de granota pirinenca més properes als Països Catalans es localitzen a uns 30-40 km cap a l’oest de la Franja de Ponent. L’absència d’aquesta espècie a Catalunya podria explicar-se per la colonització molt recent des d’un refugi glacial situat al sud dels Pirineus, probablement a cavall entre Navarra i Aragó, de manera que la granota pirinenca encara no ha tingut temps d’expandir-se cap a la part més oriental dels Pirineus.