Concepte d’artròpode i de parartròpode

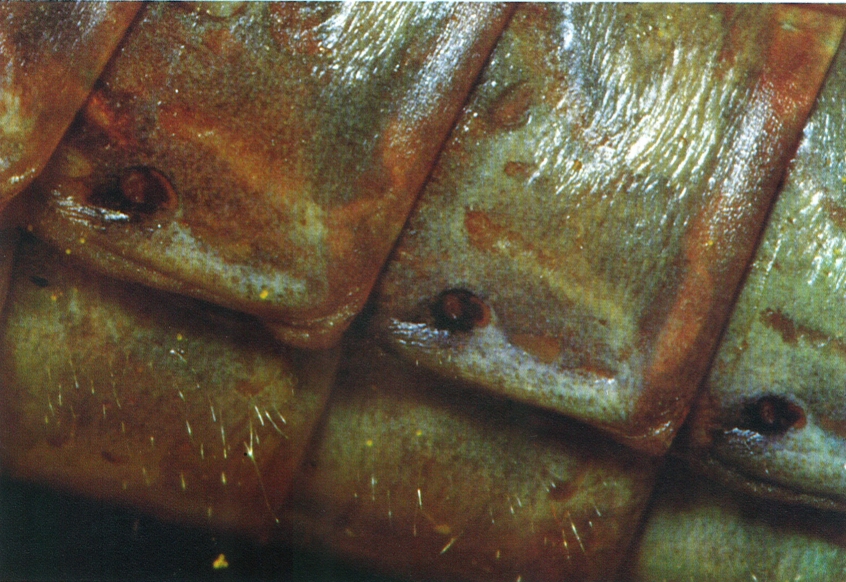
Els artròpodes, malgrat la seva gran diversitat morfològica, tenen en comú la metamerització del cos i la seva divisió en regions, els apèndixs articulats i la presència al damunt del cos d’un exosquelet fet en gran part de quitina, a més d’altres caràcters d’ordre intern. A la fotografia (un insecte odonat, Calopterix virgo), hom pot apreciar-hi les tres regions del cos pròpies dels insectes (cap, tòrax i abdomen), la situació dels apèndixs en els segments toràcics i la presència dels dos parells d’ales que els caracteritza.
Enric Curto.
Malgrat la diversitat aparent de l’organització dels diferents grups d’artròpodes, tots corresponen a un pla estructural d’una gran homogeneïtat. Els artròpodes es poden reconèixer externament per la presència d’unes determinades característiques. Una de les més clares és la cutícula endurida o exosquelet, fet principalment de quitina, que els recobreix tot el cos. D’altra banda, aquest és metameritzat o segmentat, és a dir, format per una sèrie d’unitats estructurals —els metàmers o segments— que s’articulen entre ells, i que s’agrupen de manera que es poden distingir regions o tagmes diferents. Un altre caràcter típic, la presència d’un parell d’apèndixs articulats a cada metàmer, és el que els ha donat el nom. Hi ha, però, grups d’organismes que mostren només algunes d’aquestes característiques, mentre que alhora, per altres caràcters, s’aproximen a altres grups d’invertebrats. Aquests organismes, petits animalons, els uns paràsits, els altres habitants d’ambients humits o de vida aquàtica, són, doncs, afins als artròpodes, però d’origen i significació encara no prou aclarits. Per això, els tractarem separadament sota la denominació de parartròpodes, terme molt indicat per a expressar llur característica comuna de grups afins als artròpodes.
La cutícula externa o exosquelet dels artròpodes és una capa impermeable i endurida, d’origen epidèrmic, que protegeix el cos de les deformacions mecàniques i alhora independitza els òrgans i teixits interns del medi, tot impedint-ne l’assecament. Això ha contribuït a les possibilitats de colonització d’ambients que han estat prohibitius per a d’altres grups d’invertebrats, i fins i tot de cordats. La presència d’un exosquelet rígid obliga a un tipus de creixement discontinu. Això s’aconsegueix per mitjà de la muda, és a dir, per la substitució periòdica de la carcassa vella, petita, per una de nova, més gran; aquest procés es regula per la secreció de glàndules endocrines.
La mateixa rigidesa del cos fa necessari un tipus especial de locomoció per mitjà de parells d’apèndixs articulats, és a dir, fets de diferents artells articulats, que en permeten la mobilitat. En el cas dels insectes, les ales, bé que no corresponen a veritables apèndixs locomotors, col·laboren a un tipus especial de desplaçament, el vol. En principi, cada metàmer del cos porta un parell d’apèndixs, però aquesta disposició queda alterada pel procés de tagmosi. En les formes primitives, tots els apèndixs són semblants i serveixen per a la locomoció, mentre que en les formes més evolucionades els apèndixs de les diferents regions o tagmes s’especialitzen per fer determinades funcions o, fins i tot, desapareixen.
Per bé que la presència d’un exosquelet fet de quitina, i alhora la metamerització heterònoma (és a dir, la divisió del cos en metàmers no tots iguals, agrupats en tagmes) i la presència d’apèndixs articulats metamèrics, són suficients per definir els artròpodes, hi ha també caràcters de la seva anatomia interna que són homogenis en tot el grup, i que, per tant, també tenen un valor diagnòstic: la cavitat general del cos de tipus esquizocèlic, l’aparell circulatori obert i lacunar, el tipus de respiració (fonamentalment branquial en les formes aquàtiques, traqueal en les terrestres i excepcionalment cutània), l’estructura dels òrgans excretors (la presència de tubs de Malpighi en les formes terrestres), la manca de ciliació als epitelis interns, la musculatura feta de fibres estriades disposades sense fer una capa contínua, i la presència d’un sistema nerviós ganglionar en posició ventral.
En general els artròpodes són dioics, és a dir, que a cada espècie hi ha mascles i femelles, sovint amb un dimorfisme sexual accentuat. N’hi ha, però, d’hermafrodites. La reproducció acostuma a ser sexual, bé que també hi ha casos nombrosos de partenogènesi. Són ovípars i, excepcionalment, ovovivípars i vivípars. Els ous, en general, són rics en vitel·lus i centrolecítics, de segmentació parcial i superficial. Excepcionalment, hi ha ous amb poc vitel·lus, i, en aquest cas, la segmentació és espiral i total. Hi ha, però, nombrosos casos intermedis.
El desenvolupament és típicament indirecte, és a dir, que al llarg del desenvolupament postembrionari se succeeixen tota una sèrie de fases juvenils o larvàries, que en alguns casos experimenten canvis importants en el pas d’una a l’altra, fins a entrar en la fase d’adult. Aquest procés, en conjunt, rep el nom de metamorfosi.
El tipus d’organització dels artròpodes s’ha revelat com el més capacitat per a l’evolució, raó per la qual aquest és el grup numèricament més important de tot el regne animal, tant pel que fa al nombre d’espècies com d’individus.
Els plans d’organització
L’estructura segmentada del cos dels artròpodes que hom observa externament correspon a tot un pla d’organització general. Secundàriament, però, aquesta organització segmentada del cos es pot perdre, tant pel que fa a l’estructura externa, com a la interna. A la fotografia hom pot veure clarament l’existència de plaques dorsals (tergites) i ventrals (esternites) a cada metàmer, i la situació dels estigmes respiratoris als segments de l’abdomen d’un saltamartí (insecte ortòpter); en aquest cas, les plaques laterals o pleurites són reduïdes.
Enric Curto.
Els artròpodes representen el grau més alt d’èxit evolutiu dins el regne animal, del qual és el grup més diversificat i més abundant: sobrepassen un milió d’espècies, que ve a ser les 3/4 parts de totes les espècies animals. D’altra banda, cal afegir el desconeixement absolut de la fauna d’artròpodes de diverses regions del planeta que, pel fet d’haver restat inexplorades fins ara, rendeixen centenars d’espècies noves cada any, que van engruixint contínuament el seu nombre total.
Els artròpodes són, també, nombrosos dins el registre fòssil. Els més antics, datats del Cambrià, a l’Era Primària, constituïren una fracció important de la fauna aquàtica dominant. Posteriorment, iniciaren la colonització del medi terrestre i aconseguiren adaptar-s’hi i atènyer-hi una gran importància numèrica. Actualment dominen el medi aquàtic, especialment representats pel grup dels crustacis, i el medi terrestre, amb els aràcnids, els miriàpodes i els insectes, essent aquests darrers els únics invertebrats capaços de volar, cosa que ha afavorit en gran manera la seva dispersió.
La seva gran versatilitat evolutiva els ha permès d’adaptar-se als medis més diversos, des de les profunditats abissals oceàniques fins a les condicions de l’alta muntanya. D’altra banda, han aconseguit, també, aprofitar per a la seva alimentació materials que són indigeribles per a d’altres invertebrats i fins i tot per als cordats, fet que ha contribuït al seu èxit numèric. Pel que fa a les formes de vida, la majoria són de vida lliure, però també n’hi ha que són paràsits de vegetals, d’altres animals i fins i tot de l’home. Són molts els grups d’artròpodes que, amb la seva activitat, proporcionen beneficis i avantatges a l’home i el seu entorn, com per exemple els que actuen de pol·linitzadors, els que contribueixen a la formació dels sòls, els que fabriquen substàncies útils (per exemple, mel i cera), o els que controlen les poblacions d’espècies que minen els conreus i els boscos.
La morfologia externa dels artròpodes és molt variada i és el grup on es manifesta amb més plenitud allò que hom ha anomenat «el barroc de la Naturalesa». És entre els artròpodes on es troben els invertebrats més vistosos i atraients, de vegades fins i tot extravagants i monstruosos. La seva bellesa, en molts casos, únicament pot ésser comparada a la d’algunes aus i a la de certs peixos tropicals. Les seves dimensions són igualment variables: hi ha artròpodes de mides inferiors a 1 mm (microartròpodes) i d’altres de fins a 3 m de llargada, com és el cas excepcional del crustaci Macrocheira kaempferi, propi de les mars del Japó, que constitueix la mida màxima dins dels artròpodes. Entre aquests límits, hi ha, però, la majoria de les espècies, que són de mida mitjana i petita.
Dins dels artròpodes s’inclouen tres grans grups ben diferents: el dels trilobitomorfs, el dels quelicerats i el dels mandibulats o antenats.
Els trilobitomorfs constitueixen un grup fòssil, que tingué el seu màxim apogeu a les mars del Paleozoic, i s’extingiren al Permià.
El grup dels quelicerats reuneix els merostomats i els picnogònids, propis del medi marí, i els aràcnids, fonamentalment terrestres. Els merostomats compten amb un gran nombre d’espècies fòssils, i les espècies actuals viuen a les costes del Pacífic. Els picnogònids, generalment de dimensions reduïdes, es troben amplament distribuïts pel medi marí. Finalment, els aràcnids constitueixen el grup més nombrós i diversificat dels quelicerats. Els seus representants viuen principalment al sòl, sigui a la superfície o a l’interior, del qual moltes vegades, com és el cas dels àcars, constitueixen una fracció important de la biomassa. Generalment actuen com a importants depredadors de certs organismes, mentre que d’altres col·laboren a la formació de l’humus. També n’hi ha (determinats àcars) que poden constituir flagells importants per als conreus, o que transmeten i provoquen malalties.
Els mandibulats o antenats inclouen els crustacis, d’hàbits aquàtics, i els miriàpodes i insectes, eminentment terrestres. Els crustacis són molt abundants tant en el medi marí com a les aigües salabroses i continentals. Es tracta d’un grup molt diversificat, d’una gran importància econòmica pel fet de formar part d’un dels primers graons de les cadenes tròfiques, el dels consumidors primaris. Els crustacis representen en el medi aquàtic allò que els insectes representen en el medi terrestre pel que fa a la seva diversitat i la seva abundància. Generalment són de vida lliure, però també n’hi ha de paràsits, interns i externs, que poden arribar a provocar greus malalties als seus hostes, fins i tot la mort. El grup dels miriàpodes, mancat de tota significació taxonòmica, reuneix quatre grups diferents: el dels quilòpodes (els anomenats centpeus o centcames), el dels diplòpodes (els milpeus), els símfils i els pauròpodes. Generalment viuen al sòl, i són depredadors o detritòfags. Alguns poden constituir greus flagells per als conreus. Els insectes constitueixen el grup més diversificat i numèricament més important de tots els artròpodes i de tot el regne animal. La majoria són terrestres i capaços de volar, però n’hi ha que, secundàriament, s’han adaptat al medi aquàtic i d’altres que no tenen ales. El grup dels insectes té un gran interès econòmic, sigui pels beneficis o pels perjudicis que causen a l’home i al seu entorn.
Les regions del cos
Una de les característiques que defineixen els artròpodes és el fet de tenir el cos segmentat, és a dir, constituït per una successió de segments o metàmers que, alhora, s’agrupen formant diferents regions anomenades tagmes; altrament, aquests metàmers solen estar proveïts d’un parell d’apèndixs cada un.
La metamerització
Els metàmers tenen una estructura bàsica fixa, bé que la seva morfologia pot variar molt d’uns grups als altres, igual que el seu nombre. Cada metàmer és revestit de cutícula, però aquest revestiment no és continu, sinó que fa quatre plaques esquelètiques diferents: una de dorsal o tergita, una de ventral o esternita i dues de laterals, les pleurites. Al límit entre les pleurites i les esternites s’articulen els apèndixs, en principi, un parell a cada metàmer. Els diferents metàmers s’articulen entre ells gràcies a l’existència d’una membrana articular.
L’estructura interna dels metàmers també respon a un model general. En les fases embrionàries, cada metàmer presenta esbossos mesodèrmics de dues cavitats celomàtiques, de les quals els individus adults tenen únicament les restes. Ventralment són travessats per la doble cadena del sistema nerviós, que a cada metàmer diferencia un parell de ganglis, dels quals neixen nervis que van als diferents òrgans i estructures metamèrics. Els dos ganglis de cada metàmer reben el nom de neuròmers. En posició central els travessa el tub digestiu i, per sobre d’aquest, el vas dorsal o cor, que, responent també a la metamerització, diferencia un parell d’orificis (els ostíols) a cada metàmer. Cada metàmer té una musculatura pròpia constituïda, en principi, per quatre feixos longitudinals de fibres estriades, dues de dorsals i dues de ventrals, i per musculatura dorsiventral, disposada simètricament.
La tagmatització
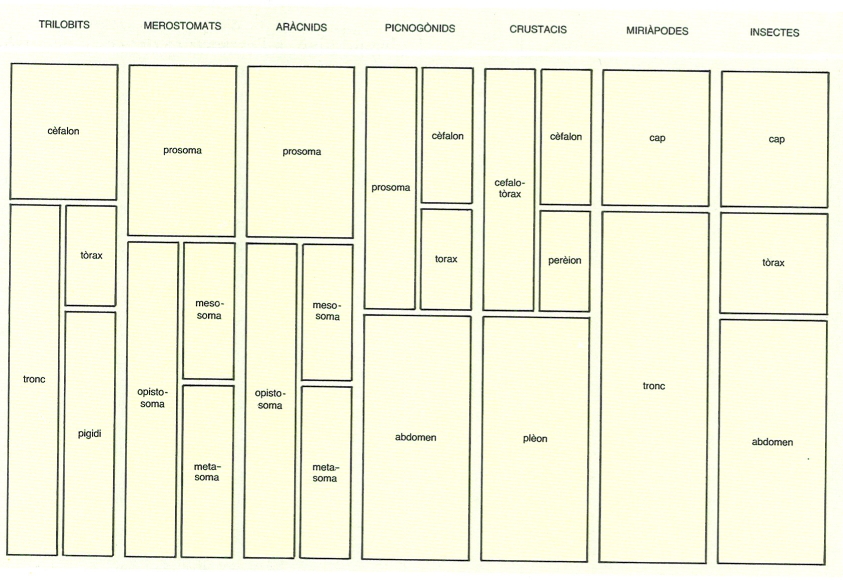
Esquema de la tagmatització o divisió del cos en regions en els diversos grups d’artròpodes. Noteu que en els artròpodes és general la tendència a la fusió de les diferents regions del cos en combinacions distintes, les quals prenen noms especials a cada cas. Cal dir que els noms que són comuns a diferents grups no sempre són donats a regions homòlogues, és a dir, dotades d’un nombre igual de segments, i que tampoc no és igual el nombre total de segments del cos.
Maber, original dels autors.
La metamerització primitiva (metamerització homònoma), per la qual tots els metàmers i tots els apèndixs són equivalents, queda alterada pel fenomen de la tagmatització. La tagmatització consisteix en la diferenciació de regions del cos o tagmes, formades per sèries o conjunts de metàmers (el concepte es correspon amb el de metamerització heterònoma). La unió de diversos metàmers per a formar un tagma pot arribar a ser total, de manera que, externament, els diferents segments o metàmers deixen de diferenciar-se. En aquest procés, els apèndixs corresponents a cada metàmer poden modificar-se, i fins i tot desaparèixer.
La tagmatització és la responsable de la gran diversitat aparent de l’estructura del cos dels artròpodes, que es manifesta entre els diferents grups i a vegades, fins i tot, dins d’un mateix grup. Els diferents tagmes varien no només per la morfologia dels metàmers que els integren, sinó també pel seu nombre, la qual cosa moltes vegades en dificulta l’homologació i la dels apèndixs respectius. No obstant això, a l’estructura del cos dels artròpodes hi és constant la diferenciació d’una regió anterior o cefàlica, resultat del procés de cefalització i clarament separada de la resta del cos.

La possessió d’ales és un caràcter exclusiu dels insectes, dins del gran grup dels artròpodes. És ben coneguda la gran varietat de models d’ales que aquests ostenten, tant pel que fa a la seva estructura i coloració com pel seu nombre i el seu grau de modificació. En principi, però, el nombre d’ales original dels insectes és de dos parells i sempre s’articulen en els dos segments posteriors del tòrax. Les plaques que cobreixen els segments són subdividides en d’altres plaques menors, que es veuen a la fotografia, corresponent a un espiadimonis (insecte odonat). El primer segment del tòrax, desproveït d’ales, pren un desenvolupament quasi dorsal, i dóna un aspecte característic al tòrax.
Enric Curto.
La regió cefàlica es caracteritza per la desaparició de la metameria primitiva i per la modificació dels apèndixs corresponents a cada metàmer, els quals prenen una funció tàctil (les antenes), prènsil (els quelícers, els pedipalps, etc.) o mastegadora (les mandíbules i les maxil·les). El segment apical de la regió cefàlica, anomenat àcron, i el segment terminal del cos, anomenat tèlson, no es consideren metàmers, pel fet que no tenen esbossos mesodèrmics a les fases embrionàries. Els tagmes posteriors a la regió cefàlica varien molt segons els diferents grups i prenen noms característics.
En els trilobits, considerats com els artròpodes més arcaics, el cos és dividit en dues regions, una d’anterior o cèfalon, i una de posterior, el tronc, el qual pot subdividir-se en tòrax i pigidi. Els apèndixs de tots els metàmers són semblants, exceptuant les antenes i els cercs (quan n’hi ha).
En els merostomats hi ha dues regions, una anterior o prosoma, on se situen els quelícers i cinc parells de potes, i una de posterior o opistosoma, que també es pot subdividir en dues regions, anomenades mesosoma i metasoma. L’opistosoma porta apèndixs i el tèlson és força desenvolupat.
En els aràcnids la tagmatització és molt semblant a la dels merostomats. A la regió anterior o prosoma hi ha els quelícers, els pedipalps i quatre parells de potes locomotores, mentre que a la regió posterior o opistosoma no hi ha apèndixs locomotors. En alguns grups, com per exemple el dels escorpins, l’opistosoma té dues regions diferents: una d’anterior o mesosoma i una de posterior o metasoma.
En els pienogònids la tagmatització del cos és peculiar. El prosoma o regió anterior es diferencia en dues zones: el cèfalon, on hi ha els quelícers o quelífors, els pedipalps i uns apèndixs especials anomenats ovígers, i el tòrax, format per un nombre de metàmers que oscil·la entre 4 i 6, tots proveïts d’apèndixs locomotors, i el primer soldat al cèfalon. L’opistosoma, en posició terminal, és molt reduït i no porta apèndixs.
En els crustacis el cos és dividit en tres regions: el cèfalon, el perèion i el plèon. El cèfalon o regió anterior, porta dos parells d’antenes i els apèndixs bucals (un parell de mandíbules i dos parells de maxil·les). El perèion, de posició intermèdia, i el plèon, o regió terminal, són formats per un nombre de metàmers variable segons els grups, i els apèndixs corresponents s’anomenen pereiopodis i pleopodis, respectivament. Al final del cos, pot haver-hi encara una furca caudal, que s’interpreta com a tèlson. És freqüent que en aquest grup hi hagi fusió parcial o total del cèfalon i el perèion, els quals es recobreixen per una closca comuna. En aquest cas, es parla de cefalotòrax, si bé no hi ha un acord total sobre l’ús d’aquest terme: alguns autors el reserven per als casos de fusió d’alguns dels metàmers del perèion al cèfalon i, en aquest cas, la resta de segments del perèion s’anomena tòrax; d’altres, només el fan servir per indicar la fusió total del cèfalon i el perèion. També hi ha qui anomena abdomen al plèon. D’altra banda, per a molts grups de crustacis hi ha una terminologia pròpia per descriure les diferents parts del cos i els apèndixs corresponents, com és el cas dels copèpodes, als quals es considera formats per un cefalosoma, un metasoma i un urosoma.
Els miriàpodes (quilòpodes, diplòpodes, símfils i pauròpodes) tenen dues regions clarament diferenciades: el cap, proveït d’un parell d’antenes i de diferents peces bucals (un parell de mandíbules i un o dos parells de maxil·les), i el tronc, format per un nombre de metàmers elevat, tots proveïts d’apèndixs locomotors.
En els insectes les regions del cos són ben diferenciades. El cap porta un parell d’antenes i les peces bucals (un parell de mandíbules i dos parells de maxil·les). El tòrax és format per tres metàmers, cadascun proveït d’un parell d’apèndixs locomotors o potes i dos parells d’ales, situades en els dos metàmers darrers. L’abdomen generalment va desproveït d’apèndixs locomotors, però porta els apèndixs relacionats amb la reproducció, i al seu extrem, els cercs terminals.
Els apèndixs
Els artròpódes presenten, típicament, un parell d’apèndixs a cada metàmer, en posició lateroventral. Per modificacions posteriors, però, poden reduir-se o fins i tot desaparèixer de determinats metàmers o de tagmes sencers.
Estructura
La característica principal d’aquests apèndixs, que dóna nom al grup, és que són formats per un seguit de peces o artells, units per punts d’articulació que en permeten el moviment. Els apèndixs s’originen a partir d’evaginacions de la paret del cos, en les quals penetren diversos elements: expansions mesodèrmiques, que després degeneren; músculs derivats de la musculatura corporal, però independitzats d’ella; i nervis, originats a partir dels ganglis segmentaris corresponents. Exteriorment són recoberts de cutícula, amb un grau d’enduriment diferent: en algunes de les espècies aquàtiques, la cutícula, prima i flexible, permet els canvis respiratoris a través seu; als crustacis bentònics i a la majoria dels artròpodes terrestres, en canvi, la cutícula és gruixuda i dura.
Els artells són anells buits, units per una membrana articular elàstica, que permet llur moviment. Per dins, s’hi insereixen els músculs, que són estriats i capaços de realitzar moviments ràpids i potents. Generalment hi ha músculs antagònics, és a dir, flexors i extensors, a cada artell, però en alguns grups n’hi ha únicament de flexors, i en aquest cas l’extensió, la fan gràcies a canvis de la pressió corporal interna. El punt d’articulació de dos artells consecutius permet el seu moviment al voltant d’un eix, com una frontissa. Els eixos de rotació dels diferents artells no solen coincidir, de manera que això permet al conjunt de l’apèndix fer moviments complexos. A les zones articulars es diferencien còndils (1 o 2) i les cavitats a les quals s’articulen, responsables de la rotació.
Tipus morfològics bàsics
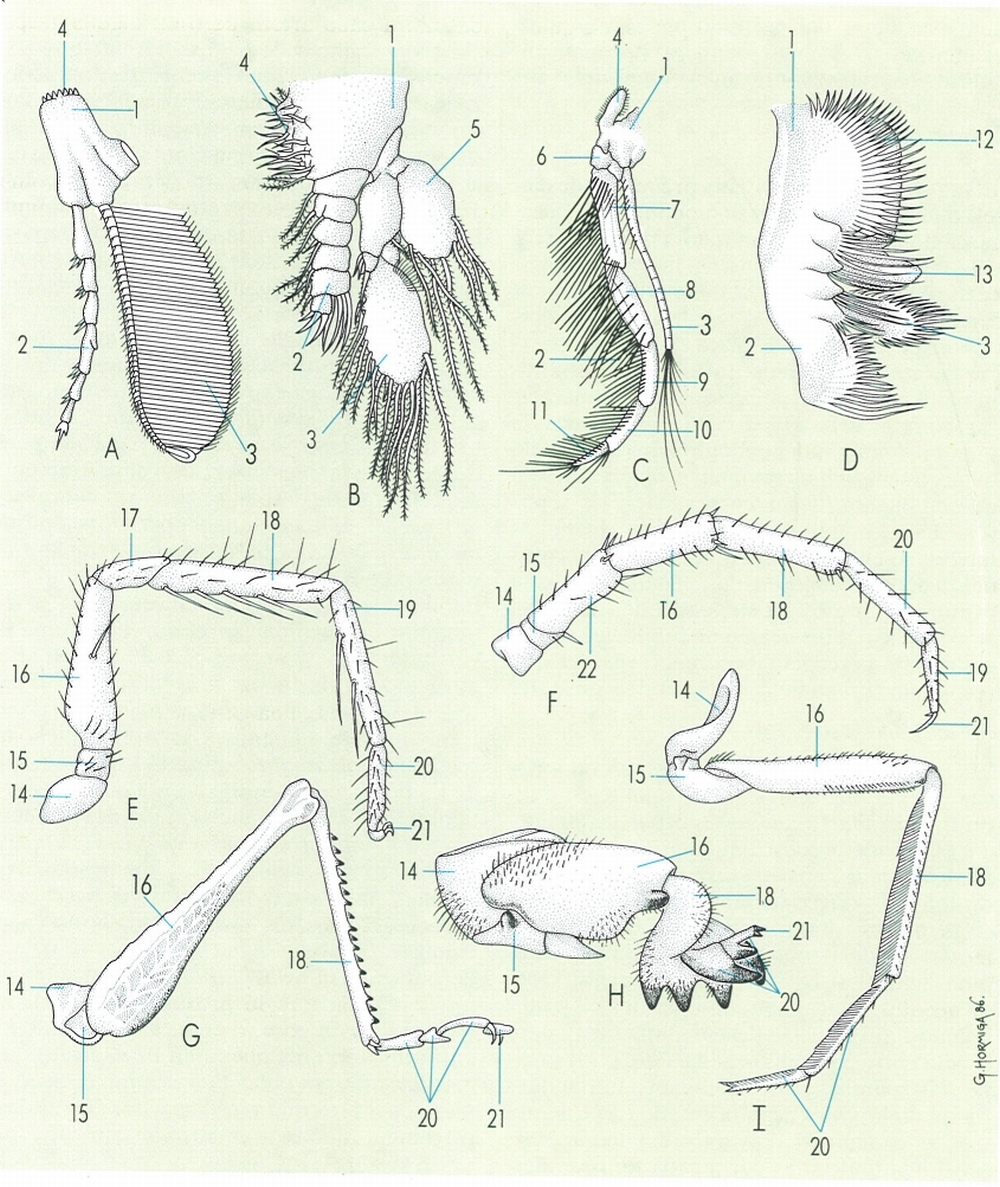
Models morfològics d’apèndixs de diferents grups d’artròpodes. Birramis: A trilobit, B crustaci cefalocàride, C crustaci decàpode, D crustaci branquiòpode. Unirramis: E aràcnid aranèid, F miriàpode quilòpode, G insecte ortòpter saltador, H insecte ortòpter excavador, I insecte coleòpter nedador. Hom hi aprecia: 1 protopodi o simpodi, 2 endopodi, 3 exopodi, 4 endita, 5 pseudoepipodi, 6 basis, 7 isqui, 8 meros, 9 carp, 10 propodi, 11 dàctil, 12 exita, 13 epipodi, 14 coxa, 15 trocànter, 16 fèmur, 17 patel·la, 18 tíbia, 19 metatars, 20 tars, 21 ungla, 22 prefèmur.
Gustavo Hormiga, a partir de fonts diverses
La morfologia dels apèndixs dels artròpodes és molt variada. Malgrat això, tenint en compte la dels grups actuals i la dels grups fòssils, es poden establir dos tipus diferents bàsics d’apèndixs: els unirramis i els birramis. A tots dos tipus hi ha sempre una regió anterior o proximal, que els connecta amb el cos, i una regió posterior o distal, que en principi té una funció locomotora.
Els apèndixs unirramis tenen l’aspecte d’un eix simple, amb una zona proximal o coxopodi i una regió distal o telopodi, segmentades i constituïdes per un nombre d’artells variable. En el coxopodi, sovint, s’hi diferencien petites protuberàncies, que s’anomenen exites o endites, segons que se situïn a la part externa o interna de l’apèndix. Els apèndixs unirramis són típics dels artròpodes terrestres: els aràcnids, els miriàpodes i els insectes.
Els apèndixs birramis es caracteritzen per la presència d’una regió proximal, anomenada protopodi o simpodi, damunt de la qual s’insereixen dues branques, una de principal interna, dita endopodi, i una altra d’externa, l’exopodi. Totes dues branques són segmentades i, en conjunt, constitueixen la part distal de l’apèndix. El protopodi és format per un o més artells, i pot tenir una o més expansions laterals, anomenades epipodis, de funció respiratòria, i també exites i endites, com a la regió proximal dels apèndixs unirramis. Els apèndixs birramis són característics dels trilobitomorfs i dels crustacis. En alguns casos, aquests apèndixs perden la independència dels artells i esdevenen laminars.
Origen i evolució
És impossible de precisar amb certesa quina fou l’estructura de l’apèndix original, a partir del qual s’han originat, per transformació o per reducció, tots els tipus actuals. Hi ha, però, diverses teories que volen explicar l’origen i l’evolució dels apèndixs dels artròpodes, que es poden reunir en tres grans grups. El primer grup inclouria les teories que consideren l’apèndix primitiu unirrami. De totes, la més consistent és la que proposa que els apèndixs dels onicòfors, anomenats lobopodis, haurien pogut ésser els precursors de l’apèndix unirrami típic, a partir del qual s’ha originat l’apèndix birrami. Els lobopodis són apèndixs parells metamèrics no articulats però extensibles. Així, per allargament i fragmentació en artells i per enduriment del tegument i ajust de la musculatura, podrien haver donat origen a un apèndix unirrami típic, el qual, per un desenvolupament exagerat d’una exita, hauria originat l’apèndix birrami típic. El segon grup inclouria les teories que consideren l’apèndix primitiu birrami, teories que es fonamenten en l’existència d’apèndixs birramis en els grups més antics d’artròpodes, com per exemple els trilobits. La pèrdua de l’exopodi hauria pogut donar origen a un apèndix unirrami i, en aquest cas, l’endopodi dels apèndixs birramis seria comparable al telopodi dels unirramis. D’aquestes teories, la més antiga fa derivar els apèndixs unirramis dels birramis dels trilobits, els quals, però, no corresponen a l’estructura dels apèndixs birramis dels crustacis. La més moderna postula que no només els apèndixs unirramis, sinó també els birramis dels trilobits, poden haver derivat dels apèndixs d’un grup particular de crustacis, el dels cefalocàrides, l’organització dels quals demostra la seva antiguitat: els seus apèndixs tenen aspecte de trirramis, i són formats per un protopodi bisegmentat, del qual neixen un pseudoepipodi i una sèrie de lòbuls i espines, un exopodi de dos artells i un endopodi de sis artells. El tercer grup de teories considera que els apèndixs unirramis i els birramis tenen un origen diferent. Aquestes teories són mantingudes pels autors partidaris del polifiletisme i del difiletisme, per a qui els grups amb apèndixs diferents haurien tingut orígens diferents.
Tipus funcionals
Els apèndixs articulats han permès el desenvolupament de diferents modalitats de desplaçament i de tota una gamma d’especialitzacions i modificacions. Alhora, suposen certs avantatges, com la possibilitat de mantenir el cos separat del substrat, suspès dels apèndixs, cosa que desplaça cap avall el centre de gravetat del cos, tot donant-li una estabilitat màxima. La presència de musculatura pròpia dels apèndixs fa que aquests es puguin moure independentment del cos (la qual cosa li evita moviments innecessaris) i que es puguin plegar en la posició de repòs (els apèndixs plegats donen més estabilitat que els apèndixs estesos). Les modificacions dels apèndixs suposen l’especialització en la captura de preses, la subjecció de l’aliment, la masticació, l’intercanvi de productes reproductors, la percepció d’estímuls sensorials, la respiració (brànquies), la posta dels ous, etc.
Els apèndixs locomotors

El tipus de moviment dels apèndixs locomotors varia en funció de la llargada i el nombre d’apèndixs. Un cas típic és el dels miriàpodes, com el milpeus (Ommatoiulus sabulosus) de la fotografia, que il·lustra el moviment metacrònic.
Antoni Serra.
Els apèndixs locomotors permeten el desplaçament pel medi terrestre i l’aquàtic en diferents modalitats (caminar, córrer, saltar, nedar) i, a més, construir galeries i grimpar. D’altra banda, una mateixa espècie és capaç de realitzar diferents modalitats de locomoció. En el cas dels insectes, proveïts d’ales, el moviment dels apèndixs locomotors es combina amb el vol. El nombre d’apèndixs dedicats al desplaçament varia segons els grups i també al llarg del desenvolupament, però oscil·la entre 3 parells (cas dels insectes) i més de 100 parells, com en alguns miriàpodes (certs diplòpodes i quilòpodes). L’estructura i el funcionament dels apèndixs locomotors són força uniformes per a cada tipus de desplaçament.
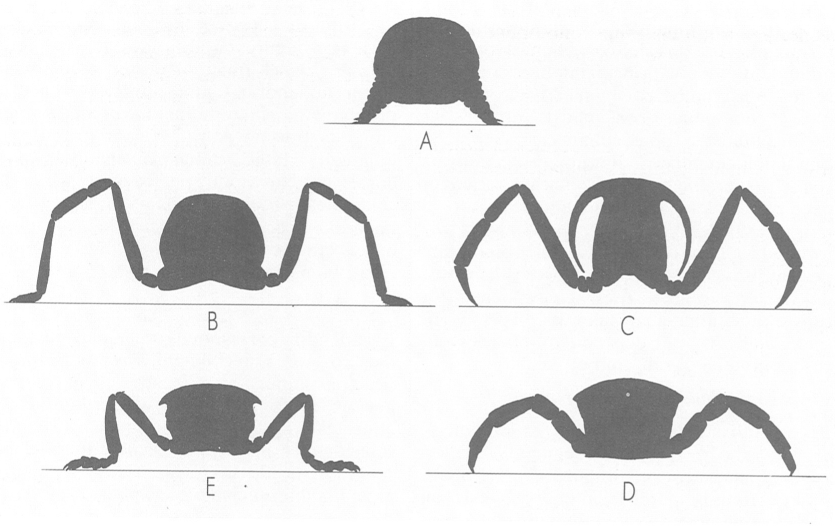
El tipus de suspensió del cos condiciona les característiques de la locomoció en els diferents grups d’artròpodes, que són representats a la il·lustració per: A onicòfors (Peripatus), B aràcnids (aranya), C crustacis decàpodes (Astacus), D miriàpodes quilòpodes (Lithobius) i E insectes (Forficula). En els onicòfors, les potes o lobopodis no tenen articulacions i recolzen en el substrat únicament la seva zona apical. Als aràcnids, la suspensió del cos és característica per la peculiaritat de tenir dos punts d’articulació principals a les potes, gràcies a la presència d’un segment suplementari, la patel·la, i perquè recolzen en el substrat tot el darrer artell. Els crustacis en recolzen només la punta, com fan també els miriàpodes. Els insectes, en canvi, recolzen el darrer artell de la pota i la seva única articulació principal els dóna una suspensió particular del cos.
Servei de Fotografia/Román Montull, a partir de fonts diverses
Els apèndixs caminadors i corredors són els més ben representats dins dels artròpodes. En el caminar i els moviments derivats, com el de córrer, el moviment de cada apèndix depèn de la seva pròpia musculatura i de la del cos, però el moviment regular i ordenat de tot el conjunt dels apèndixs depèn de mecanismes de coordinació entre els metàmers. Aquesta coordinació és especialment important en les espècies proveïdes d’un nombre elevat d’apèndixs locomotors, ja que significa el control del nombre de potes en contacte amb el substrat. Els diferents grups taxonòmics presenten peculiaritats en aquest moviment. En alguns miriàpodes, proveïts d’un gran nombre d’apèndixs locomotors, el nombre de potes que es mouen a cada moment depèn de la velocitat de desplaçament. Aquest tipus de desplaçament s’anomena metacrònic, i és característic perquè aparentment les potes fan un moviment semblant al d’una ona, en el qual cada pota es mou desfasada respecte a les seves veïnes; el valor d’aquest desfasament és el que determina la velocitat de desplaçament de l’individu. Els miriàpodes i els crustacis, en desplaçar-se sobre el substrat, hi recolzen únicament l’extrem apical dels apèndixs, mentre que els insectes i els aràcnids hi posen el darrer artell de les potes, cosa que els proporciona més estabilitat. D’aquesta manera, els dos primers grups es comporten com a ungulats, i els dos darrers com a plantígrads o digitígrads. Els insectes, amb només tres parells de potes, són capaços de moure’s a velocitats diferents i de combinar el seu moviment amb el de vol, el de natació i el de salt. La seva capacitat de moure’s endavant, endarrere i de costat demostra l’elevat grau de coordinació que hi ha entre les diferents potes. Hi ha espècies que empren totes sis potes per a caminar, però el més general és que n’utilitzin tres per al desplaçament mentre mantenen les altres tres fixes al substrat. En aquest cas, els apèndixs de cada metàmer tenen acció antagònica: quan caminen endavant, aixequen del substrat les potes de l’esquerra del primer i el tercer metàmers toràcics i la pota dreta del segon metàmer, mentre les tres potes restants mantenen el contacte amb el substrat. Aquesta posició dóna una estabilitat màxima. Quan les potes que s’havien aixecat prenen contacte una altra vegada amb el substrat, són les altres tres les que inicien el seu desplaçament endavant. Alguns insectes d’aigua, com els sabaters (Gerris), els col·lèmbols, i alguns dípters (algunes mosques i mosquits), caminen per la superfície de l’aigua, gràcies a la presència d’uns pèls hidròfugs al darrer artell de les potes, que faciliten el desplaçament pel damunt de la pel·lícula superficial de l’aigua. Hi ha insectes depredadors, com els pregadéus, que es desplacen amb els dos darrers parells de potes i fan servir el primer principalment per a capturar i subjectar les preses.

L’adequació dels apèndixs a les diferents estratègies de desplaçament pot il·lustrar-se amb el cas dels insectes que viuen a l’aigua, com les notonectes (Notonecta, a la fotografia), que es pengen de la pel·lícula superficial de l’aigua de les basses mitjançant els petits ganxets que tenen als dos parells de potes anteriors i els pèls del final del darrer segment abdominal. Per a completar l’eficàcia del sistema disposen d’unes estructures ventrals, que consisteixen en sèries de pèls distribuïdes deixant canals centrals, els quals s’omplen d’aire i fan que el centre de gravetat de l’animal es desplaci cap al dors; així, aquest té la seva posició estable panxa enlaire.
Enric Curto.
Els apèndixs nedadors són especialment freqüents al grup dels crustacis, especialment els petits, i també en alguns insectes. En els crustacis sovint són amples i laminats. Alguns grups, com el dels branquiòpodes, tenen pràcticament tots els apèndixs del cos d’aquest tipus, i els mouen d’una manera metacrònica. D’altres, com els copèpodes, únicament fan servir per a nedar alguns dels apèndixs del cos, mentre s’ajuden amb la furca caudal. Els crustacis grossos, com els decàpodes, neden lentament tot combinant el moviment d’uns certs apèndixs amb moviments bruscs de l’extrem posterior del cos; alguns caminen pel substrat del fons. Hi ha insectes que són aquàtics en estat adult, com alguns heteròpters (especialment els notonèctids i els coríxids) i alguns coleòpters (ditíscids, girínids i hidrofílids), i són capaços de desplaçar-se per la superfície de l’aigua i de nedar-hi lliurement. Generalment, utilitzen el darrer parell de potes, on porten determinades estructures especialitzades. Hi ha d’altres insectes aquàtics que es desplacen caminant pel fons o bé deixant-se portar pel corrent.
Els apèndixs minadors i excavadors generalment són apèndixs locomotors especialitzats i, en alguns casos, apèndixs bucals modificats. Alguns artròpodes els utilitzen per a construir tubs i galeries en el medi on viuen, com certs escarabats (principalment escarabèids), homòpters (larves de cigales), etc. Els cadells (Gryllotalpa gryllotalpa), els insectes més ben adaptats a l’excavació, tenen el primer parell d’apèndixs en forma de pala, amb els quals obren galeries a terra i tallen arrels. En d’altres casos (crustacis amfípodes, algunes aranyes colonials), hi ha uns apèndixs modificats per a excavar les galeries mentre que d’altres els serveixen per expulsar a l’exterior la terra sobrera. Entre els insectes hi ha grups i espècies característics per la seva capacitat de fabricar-se refugis molt perfeccionats, per exemple: les formigues, les vespes colonials, determinades abelles i els tèrmits. D’altra banda, les espècies que es nodreixen de fusta (anomenades xilòfagues) fan servir les peces bucals modificades per a triturar la fusta i per a obrir-s’hi pas, com és el cas d’alguns miriàpodes (els diplòpodes), alguns crustacis (especialment els isòpodes) i molts insectes (sobretot, coleòpters).
Els apèndixs saltadors generalment són utilitzats com a complement dels apèndixs caminadors. S’adeqüen al salt els darrers parells de potes i, en alguns casos, s’ajuden de determinades regions del cos. Aquest tipus de moviment es troba principalment entre els insectes, però també en alguns crustacis i aràcnids (d’una manera ocasional). Entre els insectes saltadors, són peculiars els col·lèmbols, que tenen apèndixs abdominals especialitzats i que generalment es desplacen caminant però que poden saltar bruscament a l’aire, i també les puces (sifonàpters), els llagosts (ortòpters) i alguns homòpters i coleòpters, que per mitjà de la contracció i extensió brusca del darrer parell de potes aconsegueixen fer salts d’una llargada considerable. Tots els insectes indicats, tret dels col·lèmbols i les puces, combinen el salt amb el vol.
Els apèndixs bucals, tàctils i prensors

La presència d’apèndixs prensors en les espècies caçadores pot donar lloc a curiosos fenòmens de convergència, com és el cas de les espècies il·lustrades, el cranc roquer (Pachygrapsus marmoratus, a dalt) i una aranya (Xysticus, a baix), que adopten una estampa comparable per la posició dels apèndixs funcionalment equivalents.
Ramon Torres i Enric Curto.
Els apèndixs relacionats amb l’alimentació, en general corresponen als segments cefàlics; però hi ha excepcions importants. La quantitat i el tipus de parells d’apèndixs bucals són caràcters que es presenten d’una manera molt homogènia a cada grup i que han servit, juntament amb altres caràcters, per a separar els grans grups d’artròpodes. Per exemple, els quelícers són uns apèndixs característics que responen sempre a un mateix model morfològic, i la presència dels quals és suficient per a identificar les espècies com a artròpodes quelicerats (aràcnids, merostomats i picnogònids) o no posseïdors de quelícers (insectes, crustacis, etc.). Les mandíbules, maxil·les i maxíl·lules dels crustacis corresponen també als segments cefàlics i tenen peces serrades per a tallar l’aliment en les espècies macròfagues, i més modificades encara en les filtradores, micròfagues. En el cas dels insectes, la diversitat de les peces bucals i la complexitat de les estructures modificades ateny un grau màxim. En les funcions de les peces bucals poden col·laborar també altres apèndixs, no pròpiament cefàlics ni tampoc específicament alimentaris, sinó com a òrgans prensors de l’aliment, o també com a mastegadors; aquest darrer cas correspon, per exemple, al de les coxes dels pedipalps dels aràcnids o d’altres apèndixs més allunyats encara de la boca, que en els crustacis fan alhora la funció locomotora i la masticatòria (gnatopodis).
Altres funcions i especialitzacions
Els apèndixs corresponents a la part posterior dels cos són especialment afectats de modificacions importants, dirigides a capacitar-los per a fer funcions molt concretes. Entre aquestes transformacions, hi ha estructures relacionades amb la respiració, com és el cas, segurament, de les fil·lotràquees dels aràcnids, i la diferenciació en expansions foliàcies de la branca externa (exopodi) dels apèndixs abdominals dels crustacis.
Una de les funcions especials que solen rebre els apèndixs del final del cos és la relacionada amb la reproducció: diferenciació de cambres incubadores, òrgans copuladors, etc. Hi ha, encara, modificacions per a altres funcions, molt diverses; així, es consideren derivats dels esbossos apendiculars estructures tan diferents com les pintes dels escorpins, les fileres de les aranyes, etc. Les modificacions morfològiques de determinades parts d’alguns apèndixs poden donar lloc a estructures que funcionen com a òrgans estriduladors.