La cavitat general del cos
L’estructura general del cos i la localització i formació dels diferents òrgans, aparells i sistemes vénen determinades pel desenvolupament embrionari, en particular l’evolució de les fulles blastodèrmiques i la formació del celoma i els seus derivats. En efecte, un caràcter propi dels artròpodes és la reducció del celoma en l’estat adult, malgrat la seva presència a la fase embrionària en forma de parells de vesícules disposades metamèricament. Aquestes són, d’altra banda, les responsables de la metamerització del cos. Simultàniament al procés de reducció de les cavitats celomàtiques durant l’organogènesi, s’esdevé la formació secundària de la cavitat general del cos o hemocel, que és de tipus esquizocèlic i podria ser el resultat de la fusió del blastocel i les cavitats celomàtiques primitives. L’hemocel és ple d’un líquid circulatori anomenat hemolimfa, que fa simultàniament el paper que fan la sang i la limfa en altres grups d’animals.
Les restes del celoma primitiu poden mantenir-se en els adults, formant les regions internes dels òrgans excretors d’origen metamèric, les gònades i els gonoductes parells. Entre els òrgans excretors d’origen metamèric hi ha les glàndules antenals i maxil·lars dels crustacis, les glàndules coxals dels quelicerats, els ronyons labials, els ronyons cefàlics i les glàndules labials, entre d’altres, d’alguns miriàpodes, i dels insectes apterigots. També són derivats mesodèrmics la musculatura, el teixit conjuntiu i l’aparell circulatori (el cor i l’hemolimfa).
L’hemocel dels adults es pot dividir en diverses cavitats o sins gràcies a l’aparició de membranes o envans que s’estenen longitudinalment al cos. En primer lloc apareix un envà per sobre del tub digestiu, que delimita dos sins, un de dorsal (anomenat si pericàrdic perquè allotja el cor) i un de ventral. En molts casos apareix un segon envà per sota del tub digestiu, que també delimita dos sins, un de central (o si perivisceral), que envolta les vísceres, especialment l’aparell digestiu, i un de ventral (o si perineural), que envolta la cadena ventral del sistema nerviós. El desenvolupament d’aquests sins és diferent a cada grup d’artròpodes i, a més, encara poden aparèixer altres envans, cosa que complica notablement la compartimentació de l’hemocel en sins.
El tegument
El tegument, és a dir, la paret del cos, té una estructura fonamental homogènia en tots els artròpodes. És format per tres elements: una membrana basal, interna, una capa de cèl·lules epitelials o epidermis i, al damunt, la cutícula, constituïda fonamentalment de quitina i altres substàncies químiques que li confereixen duresa, i que constitueix la capa més externa.
L’estructura tegumentària
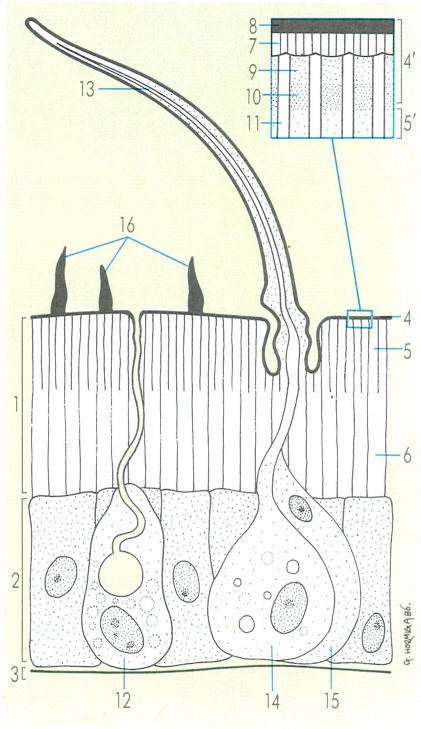
Capes i formacions del tegument dels artròpodes: 1 cutícula, 2 epidermis, 3 làmina basal, 4 epicutícula (4’ detall), 5 exocutícula (5’ detall), 6 endocutícula, 7 ceres, 8 ciment, 9 epicutícula externa, 10 epicutícula interna, 11 canals porosos, 12 cèl·lules glandulars, 13 pèl o macrotric, 14 cèl·lula tricògena, 15 cèl·lula tormògena, 16 microtrics.
Gustavo Hormiga, a partir de fonts diverses
Els tres elements tegumentaris són, com ha estat dit, la membrana basal, l’epidermis i la cutícula, els dos primers interns, i el tercer, que és el més destacable, extern.
Les capes internes
La membrana basal és una capa contínua, molt prima, que separa l’epidermis de la cavitat general del cos o hemocel. Té una estructura amorfa o granular. L’epidermis és constituïda per una capa de cèl·lules monostratificades, principalment poligonals, cúbiques o cilíndriques. Aquestes cèl·lules, a més de secretar la cutícula, produeixen el líquid exuvial, que dissol la part interna de la cutícula vella abans de cada muda, i n’absorbeixen els productes residuals. Entre les cèl·lules epidèrmiques hi ha cèl·lules especialitzades, com les que formen les glàndules tegumentàries (unicel·lulars o pluricel·lulars), les responsables de la formació de determinats òrgans sensorials i les que donen origen a elements cuticulars diversos, com pèls, esquames, esperons, etc. D’altra banda, aquesta capa és travessada per les terminacions musculars que van a inserir-se a la capa més interna de la cutícula. En els insectes, hi ha també unes cèl·lules especials, els enòcits, relacionades amb la secreció de lípids.
La cutícula

El fenomen de la muda es presenta a tots els artròpodes, ja que per a créixer els és imprescindible desprendre’s periòdicament de la protecció quitinosa que els recobreix. A les fotografies veiem una exúvia —closca quitinosa abandonada després de la muda— de tortugueta (Triops cancriformis, a dalt) i una altra d’escorpí groc (Buthus occitanus, a baix).
Josep M. Barres i Xavier Parellada.
La cutícula és una formació rígida, acel·lular, i d’estructura complexa, en la qual es produeixen importants processos bioquímics, alguns regulats per enzims. Constitueix el revestiment més extern del cos i dels apèndixs, i, localment, s’invagina per a formar l’endosquelet. També revesteix el sistema traqueal, les regions anterior (estomodeu) i posterior (proctodeu) del tub digestiu, els canals excretors d’algunes glàndules i determinades parts de l’aparell reproductor, és a dir, totes les formacions d’origen ectodèrmic. Ultra la funció protectora mecànica, la cutícula determina la forma del cos, redueix la transpiració i proporciona una base ferma per a la inserció dels músculs. La cutícula pot servir, també, d’aliment de reserva, ja que, en general, després de la muda, els joves individus es mengen la cutícula vella despresa.
Les conseqüències de la presència d’una cutícula són: l’absència total de cilis vibràtils a la superfície dels epitelis, el moviment relatiu del cos i dels apèndixs i els seus artells, que es fan gràcies a l’elasticitat i flexibilitat de les membranes articulars; la formació d’apodemes, sobre els quals s’insereixen els músculs; la substitució de la capa muscular contínua present a d’altres invertebrats per feixos de músculs estriats, disposats metamèricament; i el creixement discontinu i el fenomen associat de la muda.
El gruix de la cutícula és molt diferent a cada grup d’artròpodes i també a les diferents regions del cos. Pot ser extremament prima, fent com una delicada membrana al voltant del cos, com és el cas de la cutícula d’algunes larves de mosquit, de menys d’un micró de gruix; o prou gruixuda per a constituir una autèntica cuirassa, com en el cas del límul o cassola de les Filipines (Limulus), de més de tres mil·límetres de gruix. En els insectes, els aràcnids i la majoria dels miriàpodes, la cutícula és poc o no gens calcificada. En canvi, en els crustacis i alguns miriàpodes (els diplòpodes), té un grau de calcificació important, cosa que n’afavoreix la rigidesa. En general, presenta ornamentacions externes i porta diverses formacions tegumentàries superficials, com pèls i esquames. També es troba en relació amb les glàndules i òrgans sensorials de funció i estructura diversa. Quan la cutícula és acabada de formar, és flexible i elàstica, i bé que en alguns casos persisteix en aquest estat, en general experimenta un procés d’esclerificació, pel qual s’endureix i s’enfosqueix; alhora que es delimiten diferents esclerites rígides, separades per zones membranoses on la cutícula es manté flexible (membranes articulars) i permet l’articulació de les esclerites. Aquesta disposició, que compagina rigidesa i flexibilitat, es manté en les exúvies que els artròpodes abandonen en mudar.
La cutícula és formada per dues capes diferents, una d’externa i prima, anomenada epicutícula, i una altra d’interna i més gruixuda, la procutícula.
L’epicutícula o cutícula externa és de composició i ultrastructura molt complexa i variable segons els grups. En la seva composició intervenen ceres i lipoproteïnes (especialment, cuticulina), però mai quitina. La seva funció és impermeabilitzar el revestiment del cos per evitar la pèrdua d’aigua. S’estructura en diferents capes (un mínim de dues i un màxim de quatre), les dues més internes secretades per l’epidermis i les dues més externes produïdes per les glàndules dèrmiques.
La procutícula forma la gran part del gruix de la cutícula i és la capa responsable de la seva rigidesa. A la seva composició hi intervé la quitina i diverses proteïnes (escleroproteïnes). La quitina (un hidrat de carboni) és una substància elàstica i flexible, permeable a l’aigua i a diverses substàncies dissoltes, i incolora, que té una estructura fibril·lar. Representa entre 1/4 i 1/2 del pes sec de la cutícula. Les escleroproteïnes representen aproximadament 1/4 del pes sec total de la cutícula i són les principals responsables de la rigidesa de la cutícula; entre aquestes proteïnes hi ha l’artropodina, l’esclerotina i la resilina; aquesta, descoberta recentment en els insectes, és una proteïna elàstica que es localitza principalment a les zones articulars. La resta de components de la procutícula en representen un tant per cent del pes sec total molt petit, si bé tenen una gran importància fisiològica. Moltes vegades, la duresa de la cutícula es deu a l’acció combinada de les escleroproteïnes i de dipòsits de sals minerals, fonamentalment carbonat càlcic i fosfat càlcic, que poden arribar a representar un tant per cent del pes sec total de la cutícula molt elevat, com és el cas de molts crustacis i molts diplòpodes.
La procutícula també s’estructura en capes diferents: l’exocutícula, la més externa, i l’endocutícula, sovint separades per la mesocutícula. Per sota de l’endocutícula, i en contacte amb l’epidermis, pot haver-hi encara la capa subcuticular. L’exocutícula és una capa homogènia, dura i acolorida, atès que sol contenir melanines i altres pigments, disposats de maneres diverses; és constituïda principalment per una escleroproteïna (l’esclerotina) i quitina, en una proporció baixa. L’endocutícula és una capa elàstica, incolora i no endurida, que representa el gruix principal de tota la cutícula; té estructura laminar i a la seva composició hi intervé d’una manera majoritària la quitina, bé que també hi ha una proteïna (l’artropodina) i mucopolisacàrids; és en aquesta capa on es produeixen els dipòsits de sals càlciques. La mesocutícula, present en molts crustacis, insectes i aràcnids, és una capa acolorida, feta de quitina i proteïnes. La capa subcuticular o subcutícula basal, té estructura granular i és formada per microfibril·les cuticulars, semblants a les que formen l’endocutícula, però disposades sense un ordre concret.
Els colors cuticulars
En els artròpodes, el color extern del cos es deu a la presència de pigments englobats a la cutícula, a l’epidermis o bé en els teixits subepidèrmics (colors químics), a l’estructura física de la cutícula (colors físics) o bé a irisacions dels pèls i esquames que recobreixen la seva superfície i la dels apèndixs. Generalment, però, el color resulta de la combinació de tots tres tipus.
A la majoria dels artròpodes, el color es manté estable al llarg de la vida, però en alguns pot experimentar canvis importants. De vegades (a molts crustacis decàpodes) el color extern experimenta un cicle de variació cromàtica en relació amb la intensitat de llum que arriba al medi. En d’altres casos el color trasllueix diferents estratègies dels organismes, sigui perquè tendeix a confondre’s amb el substrat o bé, al contrari, perquè per la seva brillantor tendeix a destacar notablement del medi.
Els pigments són de naturalesa diversa: carotens, pterines, pigments sanguinis (hemoglobina i altres) i, especialment, melanina i ommocroms. En general, els colors químics són menys estables que els físics, però quan se situen a la cutícula, es mantenen pràcticament sense cap alteració fins i tot després de la mort de l’individu. Els colors físics es mantenen inalterats mentre no es modifiqui l’estructura de la cutícula i en general són els responsables dels tons metàl·lics i les irisacions, que varien de tonalitat segons l’orientació de l’organisme respecte de la claror.
Les formacions cuticulars
La superfície de la cutícula pot presentar diferents formacions externes que, pel seu origen, corresponen a engruiximents de la capa superficial de la cutícula, o bé a formacions originades a partir de cèl·lules epidèrmiques especialitzades.
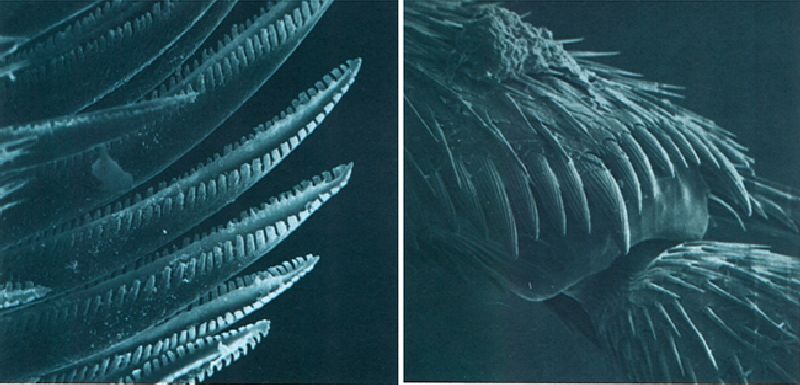
Els pèls, els esperons i les apófisis són abundants a la superfície cuticular dels artròpodes. Els pèls espinosos de la mandíbula d’un quilòpode (a l’esquerra) són molt quitinitzats, ja que col·laboren en la trituració dels aliments (× 650). Els esperons de la regió apical de la tíbia d’un coleòpter catòpid (a la dreta) són articulats i segregats per diverses cèl·lules epidèrmiques (× 660).
Antoni Serra i Marina Blas.
Els engruiximents locals de la cutícula donen origen a formacions d’aspecte molt variable que constitueixen el relleu de la cutícula: grans, estries, costelles, rugositats, etc. De vegades, aquestes formacions sobresurten clarament de la cutícula, bé que s’hi mantenen rígidament unides i sense cap articulació.
Aquestes formacions s’anomenen apòfisis. Els principals tipus d’apòfisis són els microtrics o pèls fixos, semblants als pèls de revestiment i les espines. Determinats grups o espècies d’artròpodes tenen formacions particularment i característicament desenvolupades, com ara banyes i protuberàncies (a molts insectes), tubercles especials (en alguns aràcnids), etc.
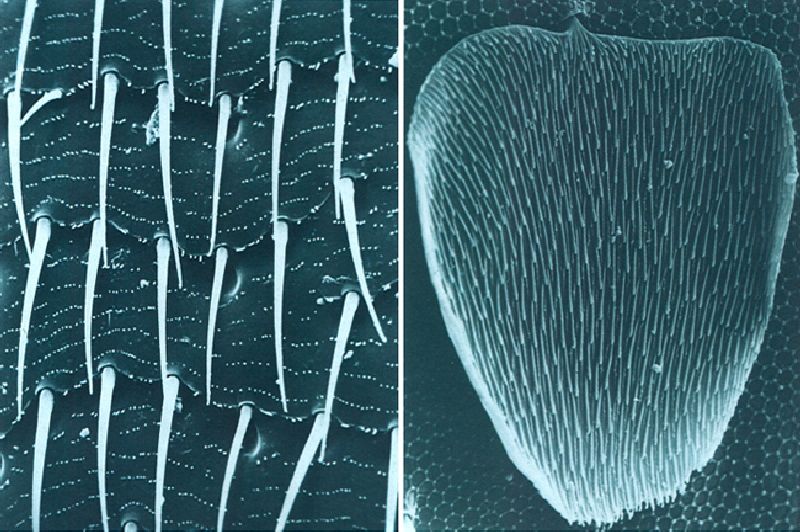
Les formacions cuticulars de la superfície del cos dels artròpodes sovint només són apreciables amb l’ajut del microscopi electrònic d’escandallatge. A les fotografies hom pot veure, a l’esquerra, pèls de revestiment o macrotrics, microtrics i estries transversals dels èlitres d’un coleòpter cavernícola (× 400). A la dreta, una esquama, que no és més que un pèl de revestiment modificat, damunt d’un fons fet d’altres elements ornamentals del tegument d’un col·lèmbol (× 5000).
Marina Blas i Eduardo Mateos.
Les formacions cuticulars originades a partir de cèl·lules epidèrmiques especialitzades són mòbils o articulades. Bàsicament, n’hi ha de dos tipus: les sedes, pèls o macrotrics, i els esperons. Els primers s’insereixen dins una fosseta o alvèol enfonsada en la cutícula o bé elevada sobre un tubercle, on connecten amb la cutícula a través d’una membrana articular. Els pèls són produïts per una sola cèl·lula tricògena, i la membrana articular per una o diverses cèl·lules tormògenes. A cada muda, els pèls són eliminats amb la cutícula vella, i se’n formen de nous a la nova cutícula, a partir de les cèl·lules tricògenes corresponents. La disposició i la forma dels pèls o sedes pot servir per a classificar alguns grups d’artròpodes. N’hi ha de diferents tipus: pèls de revestiment, que recobreixen la superfície externa del cos i dels apèndixs i poden ser plomosos, pectinats, etc.; quan són molt robusts s’anomenen cerres; esquames, que són pèls de revestiment molt modificats; sedes i esquames glandulars, que donen sortida als productes segregats per les glàndules epidèrmiques; sedes o pèls sensorials, que són modificats per percebre i transmetre als centres nerviosos els estímuls externs o interns i estan en contacte amb cèl·lules sensorials (n’hi ha de molt especialitzats, com és el cas de les tricobòtries). Els esperons corresponen a formacions epidèrmiques pluricel·lulars, és a dir, secretades per diverses cèl·lules epidèrmiques. Se situen a diferents regions del cos, especialment als apèndixs.
L’endosquelet
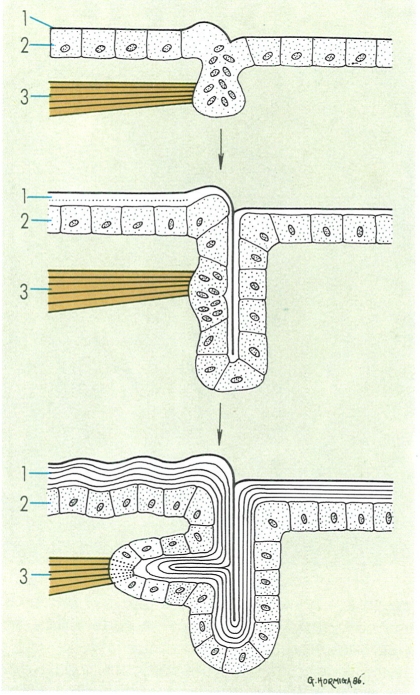
La formació d’apodemes s’inicia quan la cutícula comença a engruixir les seves diferents capes per sobre de l’epidermis. Els músculs, que de primer s’insereixen damunt d’aquesta, progressivament van penetrant en el tegument fins a arrelar a la cutícula. Precisament aquesta és una de les funcions principals dels apodemes, és a dir, augmentar i enfortir la superfície d’inserció dels músculs. Hom remarca: 1 cutícula, 2 epidermis, 3 musculatura.
Gustavo Hormiga, a partir de fonts diverses
L’endosquelet o esquelet intern és el conjunt d’invaginacions del tegument o apodemes a les quals s’insereixen els músculs. Els apodemes poden ser buits o compactes i es disposen metamèricament. La seva morfologia varia segons quina sigui la seva funció mecànica; n’hi ha en forma de cresta, de placa, de branca, etc. Els apodemes en forma de cresta reforcen la resistència de l’exosquelet, fent-lo més rígid i compacte a les zones del cos sotmeses a forces de tracció. Els que tenen forma de branca, generalment serveixen per a augmentar la superfície d’inserció dels músculs. Hi ha apodemes molt prims, anomenats tendons, que serveixen de punt d’inserció a una única fibra muscular.
La musculatura
Els músculs, en els artròpodes, no es disposen formant una capa contínua, sinó en feixos musculars individuals. Són de tipus estriat.
Les fibres musculars, en principi, s’insereixen en la cara interna de l’epidermis, però, al llarg del desenvolupament, acaben inserint-se directament en la cutícula, gràcies a la formació de tonofibril·les a l’interior de les cèl·lules epidèrmiques, que vénen a ser com prolongacions de les fibres musculars, i que penetren fins a l’interior de la cutícula. Segons la seva localització, hi ha musculatura de dos tipus: esquelètica i visceral.
Els músculs esquelètics s’insereixen a banda i banda de les articulacions i permeten el moviment lleu dels diferents metàmers del cos, el moviment dels artells dels apèndixs i el dels apèndixs respecte al cos. Són formats per fibres paral·leles o disposades en ventall. Els músculs que mouen les ales i l’armadura genital també són d’aquest tipus. Els músculs esquelètics es disposen per parells, metamèricament.
Tall transversal del cos d’un artròpode per a mostrar, dins d’un metàmer típic, la distribució dels diferents feixos musculars i la de la resta d’elements anatòmics: 1 diafragma, 2 si dorsal o pericàrdic, 3 cor, 4 si central o perivisceral, 5 tub digestiu, 6 si ventral o perineural, 7 sistema nerviós, 8 músculs longitudinals dorsals, 9 músculs longitudinals ventrals, 10 músculs dorsiventrals, 11 músculs dels apèndixs.
Amadeu Blasco, a partir de fonts diverses
Els músculs viscerals envolten les vísceres, i en alguns casos poden constituir un revestiment més o menys regular, fet per una capa de fibres longitudinals i una altra de fibres circulars, disposició especialment característica de la part mitjana del tub digestiu, anomenada mesodeu. Els esfínters corresponen a engruiximents locals de la capa de fibres circulars. En altres punts del cos, com en determinades àrees del tub digestiu, dels òrgans genitals, dels tubs de Malpighi, etc., els músculs viscerals formen una xarxa discontínua de fibres ramificades que asseguren el peristaltisme dels òrgans, els moviments de deglució, el tancament dels estigmes, el moviment dels diafragmes, etc. En aquest cas es tracta de músculs molt modificats, al voltant de la naturalesa dels quals no hi ha, actualment, un acord total.
El moviment dels músculs en general és controlat pel sistema nerviós: les fibres es contrauen rítmicament per impulsos nerviosos i necessiten un període de recuperació per a poder-se estimular novament. Aquest tipus de músculs s’anomenen síncrons o sincrònics, i el seu moviment és relativament lent. Els músculs que mouen les ales en la majoria dels insectes, en canvi, tenen un ritme de contracció que no es regula per impulsos nerviosos; aquest tipus de músculs s’anomenen asíncrons o asincrònics, i es contrauen molt més ràpidament que els anteriors. És il·lustrativa dels dos tipus de contracció la comparança entre els músculs sincrònics de les ales de les papallones, amb una freqüència de 10 a 20 contraccions per segon, i els de les ales de la majoria de la resta dels insectes, asincrònics, amb una freqüència de contracció generalment variable entre 50 i 220 contraccions per segon, amb màxims de fins a 1000 contraccions per segon (alguns dípters).
Entre els insectes hi ha la complexitat muscular més gran de tots els artròpodes. Com a exemple, les erugues d’algunes papallones poden tenir fins a 3000 músculs disposats metamèricament, bé que externament aquesta disposició queda emmascarada per l’aparició dels tagmes o regions del cos.
Els sistemes de relació i control
El sistema nerviós constitueix un nexe d’unió entre els òrgans sensorials, que són excitats per diversos estímuls externs i interns, i els òrgans efectors, com els músculs, les glàndules, etc. Per mitjà dels òrgans efectors els organismes reaccionen als estímuls i modifiquen la seva conducta. El grau de coordinació és en relació amb la complexitat del sistema nerviós.
D’una manera simultània, hi ha cèl·lules nervioses que actuen com a cèl·lules secretores de neurohormones. Aquestes substàncies resulten imprescindibles per a la integració i la coordinació del funcionament de les glàndules endocrines, les secrecions de les quals controlen diferents processos fisiològics d’importància fonamental per a la vida dels organismes.
El sistema nerviós

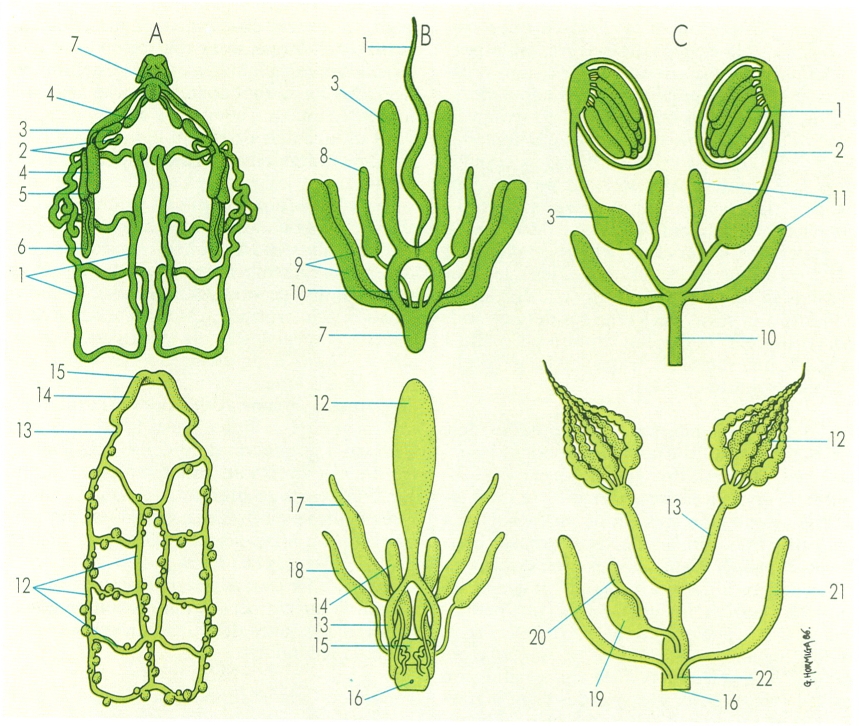
En l’organització del sistema nerviós dels artròpodes hom pot detectar una relació entre el grau d’evolució assolit i el grau de concentració dels diferents elements ganglionars. Aquest fenomen es manifesta dins les diverses línies evolutives, cosa que il·lustra el dibuix en la comparació de dos models corresponents a aràcnids, el de l’escorpí (A) i el d’un aranèid (B); a crustacis, el d’un fil·locàride (C) i el d’un decàpode (D), i a insectes coleòpters, el d’un lícid (E) i el d’un escarabèid (F). En els miriàpodes (G), la cadena ventral és poc concentrada i s’estén al llarg de tot el cos.
Servei de Fotografia/Román Montull, a partir de fonts diverses
El sistema nerviós dels artròpodes es caracteritza pel fet d’ésser ganglionar i hiponeure, és a dir, constituït per parells de ganglis units per comissures i connectius, tots en posició ventral respecte al tub digestiu, tret dels ganglis que constitueixen el cervell o massa ganglionar supraesofàgica. No hi ha els plexes subcutanis que es troben en altres grups d’invertebrats, i la seva estructura general correspon a la dels anèl·lids, bé que amb molta més complexitat. És format per una massa ganglionar supraesofàgica o cervell, en posició dorsal, per una cadena ganglionar ventral, que presenta un parell de ganglis a cada metàmer, i per una anella periesofàgica que uneix ambdues regions. En general, els primers ganglis de la cadena ventral es troben fusionats i constitueixen la massa ganglionar infraesofàgica, la qual es manté diferenciada de la resta de la cadena ventral. En relació amb el sistema nerviós central hi ha un sistema nerviós simpàtic.
El cervell o massa ganglionar supraesofàgica és el resultat de la fusió de tres parells de ganglis que formen el protocervell, el deutocervell i el tritocervell. Si bé tots tres se situen en posició preoral, només els dos primers ocupen aquesta posició primitivament, ja que el tritocervell, en un principi, ocupava una posició postoral. En alguns crustacis, com els branquiòpodes, el tritocervell es manté independent dels altres dos ganglis cerebrals. D’altra banda, en els quelicerats no hi ha deutocervell, de manera que el cervell és constituït únicament pel protocervell i el tritocervell.
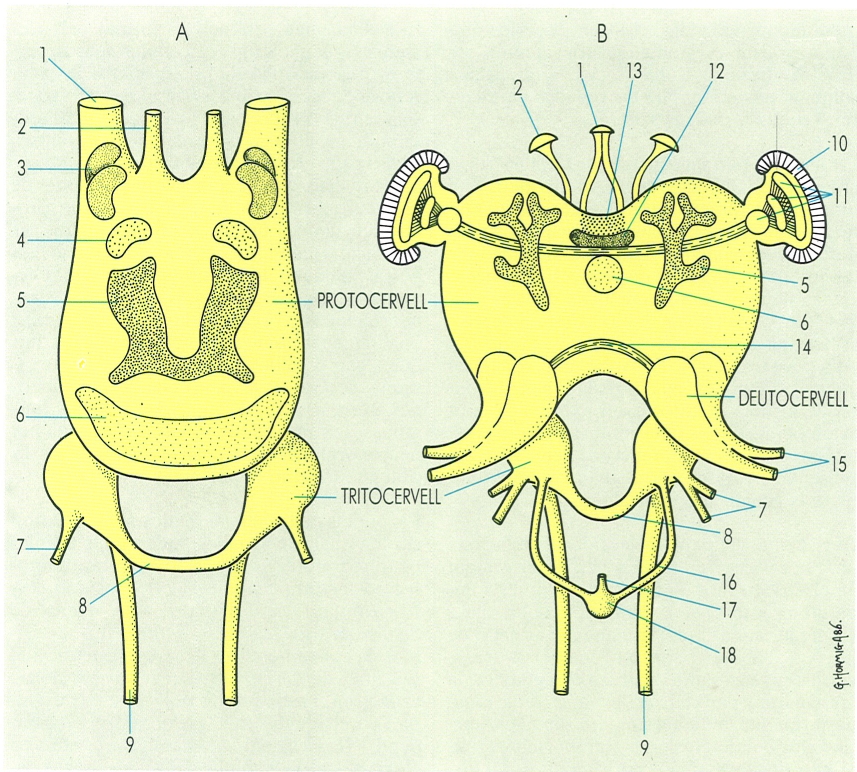
Parts principals del cervell dels artròpodes aràcnids (A) i insectes (B): 1 i 2 nervis dels ocel·les, 3 i 4 centres visuals primaris i secundaris, 5 cossos pedunculats, 6 cos central, 7 nervis del tritocervell, 8 comissura tritocerebral, 9 inici del collar periesofàgic, 10 ulls compostos, 11 làmines ganglionars del lòbul òptic, 12 part cerebral, 13 part intercerebral amb cèl·lules neurosecretores, 14 comissura deutocerebral, 15 nervis del deutocervell, 16 sistema nerviós simpàtic anterior, 17 nervi recurrent, 18 gangli frontal. Hom pot observar en el dibuix les diferències i les semblances entre ambdós cervells, especialment la manca de deutocervell i la major simplificació general dels centres nerviosos en el dels aràcnids.
Gustavo Hormiga, a partir de fonts diverses
El protocervell és format per dos lòbuls cerebrals separats per la "pars intercerebralis" (part intercerebral), de funció neurosecretora. D’aquests lòbuls parteixen nervis que van a innervar els ulls compostos i/o els ulls simples o ocel·les, ja que al protocervell se situen els centres òptics i ocel·lars. Aquesta regió cerebral és la més desenvolupada en tots els artròpodes, perquè inclou els centres d’integració i coordinació, molt desenvolupats en els insectes socials. El protocervell presenta diferents zones o regions, amb un desenvolupament més o menys gran segons els grups, i una sèrie de lòbuls laterals, els més importants dels quals són els lòbuls òptics. Als lòbuls òptics se situen els centres òptics, que són centres d’associació de les impressions visuals. El protocervell innerva l’àcron i l’anomenat metàmer preantenal dels insectes, miriàpodes i crustacis, i la regió acronal dels quelicerats, tots portadors dels òrgans visuals.
El deutocervell o cervell mitjà és format per dues masses nervioses reunides per una comissura supraesofàgica. Inclouen els centres antenals, dels quals parteixen els nervis corresponents. Innerva les antenes dels insectes i els miriàpodes, el primer parell d’antenes dels crustacis (les antènules) i el metàmer on se situen. Es considera extremament reduït en el grup dels quelicerats, grup en el qual manquen les antenes.
El tritocervell o cervell posterior és constituït per dues masses nervioses que porten dos centres nerviosos simètrics. D’aquests centres neixen les fibres que formen la comissura subesofàgica, donat que la posició primitiva del tritocervell és postoral; pel procés de cefalització i pel gran desenvolupament dels connectius que el relacionen amb la cadena ventral i que formen l’anomenada anella periesofàgica, posteriorment passa a una posició preoral. El tritocervell innerva els quelícers i el seu metàmer corresponent en el grup dels quelicerats, el segon parell d’antenes dels crustacis, i l’anomenat metàmer premandibular dels insectes i els miriàpodes. D’altra banda, és en relació amb el sistema nerviós simpàtic anterior, que innerva la regió anterior del tub digestiu.
La relació entre els diferents lòbuls cerebrals és assegurada per una gran quantitat de fibres i de nervis, i en particular pels cossos pedunculats, que són una diferenciació del protocervell, i que coordinen el funcionament de tots els centres nerviosos.
La cadena ventral és constituïda per un nombre determinat de parells de ganglis, similar al nombre de metàmers del cos, no innervats pel cervell. Això no obstant, poques vegades aquestes parelles de ganglis es mantenen independents, sinó que hi ha una tendència general a la fusió d’unes determinades parelles de ganglis de la cadena ventral.
Una primera mostra d’aquesta tendència és la diferenciació de la massa ganglionar infraesofàgica pel procés de cefalització. Simultàniament, els darrers parells de ganglis de la cadena ventral tendeixen a fusionar-se i, en alguns casos extrems, són tots els ganglis de la cadena ventral que es concentren formant una sola massa ganglionar.
La massa ganglionar infraesofàgica correspon a la concentració dels ganglis de la regió anterior de la cadena ventral, i el nombre de ganglis que s’hi concentra depèn de cada grup. En els aràcnids és feta de 5 parells de ganglis i innerva 5 parells d’apèndixs del prosoma (els pedipalps i 4 parells de potes caminadores) i els metàmers corresponents. En els mandibulats, és a dir, els insectes, els crustacis i els miriàpodes, la massa ganglionar infraesofàgica generalment és constituïda per tres parells de ganglis, i en conjunt rep el nom de gnatocervell, perquè innerva les peces bucals i els seus metàmers. De cada gangli de la cadena ventral, en surten parells de nervis que innerven els metàmers corresponents i les estructures relacionades amb ells. Aquests ganglis, a més, porten un o diversos tipus de cèl·lules neurosecretores, de disposició característica en els diferents grups. Aquesta regió del sistema nerviós central es relaciona amb el sistema nerviós simpàtic posterior o visceral.
El sistema simpàtic
El sistema nerviós simpàtic es relaciona amb el sistema nerviós central i innerva les vísceres. S’hi poden veure dues regions perfectament independitzades: el sistema nerviós simpàtic anterior o estomatogàstric, que en general es troba ben diferenciat, i el sistema simpàtic posterior, que de vegades no es diferencia de la cadena ventral.
El sistema estomatogàstric innerva la regió anterior del tub digestiu o estomodeu, i té una relació directa amb el tritocervell. Generalment porta un gangli principal ben diferenciat (el gangli frontal dels insectes i els diplòpodes, el gangli esofàgic dels crustacis...), que pot anar acompanyat d’altres ganglis secundaris (el gangli hipocerebral dels insectes i els diplòpodes, els ganglis estomacals dels crustacis, etc.). Es troba molt reduït en alguns miriàpodes, en els quals el gangli frontal es fusiona al tritocervell i forma el pont estomodial. En els quelicerats aquest sistema tampoc no és ben diferenciat, bé que hi és representat pel pont estomodial d’Hamstrom i per uns ganglis innominats.
El sistema simpàtic posterior es relaciona amb la cadena ventral. És ben desenvolupat en els insectes, on és format per un nervi imparell o central, situat entre el parell de ganglis de la cadena ventral, i anomenat nervi de Leydig; d’aquest parteixen dos nervis transversals simètrics que innerven el tegument, la musculatura parietal, els espiracles, la regió posterior del tub digestiu, les gònades, etc. En relació amb els nervis transversals parells hi ha situats els òrgans perisimpàtics. La disposició dels nervis transversals i dels òrgans perisimpàtics varia segons el grau de concentració dels ganglis de la cadena ventral. El sistema simpàtic posterior també es troba representat en els crustacis superiors, en els quals els ganglis corresponents s’han incorporat a la cadena ventral. En els miriàpodes és tot el sistema el que s’hi ha incorporat. En els quelicerats (arenèids) és representat per un nervi imparell, els ganglis del qual queden incorporats a la cadena ventral, que és molt concentrada.
El sistema endocrí
La majoria de les funcions vitals dels artròpodes són regulades per hormones segregades per glàndules de secreció interna l’acció de les quals és controlada per neurohormones, segregades al seu torn per cèl·lules nervioses especialitzades (neurosecretores). Simultàniament, la secreció de les neurohormones és controlada per estímuls nerviosos, com passa en altres grups d’animals. Dins dels artròpodes aquest sistema de coordinació química és força constant pel que fa als seus components. El grau de desenvolupament i la localització concreta dels diferents elements que l’integren, però, és molt variable segons cada grup.
El sistema neuroendocrí és format pels elements següents: grups de cèl·lules neurosecretores, responsables de la secreció de les neurohormones, i localitzats en el cervell (especialment el protocervell) i a la cadena ventral, sempre en llocs molt concrets i constants a cada grup; grups d’àxons disposats centrífugament, que actuen com a transportadors de les neurohormones des del lloc on es produeixen fins als òrgans neurohemals que les emmagatzemen (en altres casos, aquests àxons aboquen les neurosecrecions a l’hemolimfa, que les transporta fins als òrgans neurohemals); òrgans o àrees neurohemals, on s’emmagatzemen les neurohormones fins que s’han d’alliberar, cosa que requereix estímuls nerviosos; diverses glàndules endocrines, responsables de la secreció de les hormones, la funció de les quals és regulada per neurohormones que controlen fenòmens fisiològics de gran importància.
Els òrgans neurohemals, a més de la seva funció habitual, de vegades poden tenir cèl·lules neurosecretores, capaces d’elaborar neurosecrecions pròpies; aquest és el cas dels cossos cardíacs dels insectes, els òrgans de Schneider dels aràcnids i d’altres. La seva situació, el seu nombre, la seva estructura i el seu grau de desenvolupament varien molt a cada grup, i reben noms especials; per exemple, en els quelicerats, els òrgans de Schneider I i II, el gangli estomatogàstric, etc.; en els crustacis, la glàndula del si i els òrgans pericàrdics, etc.; en els miriàpodes, l’òrgan de Gabe, l’òrgan cerebral neurohemal, etc.; en els insectes, els cossos cardíacs i els òrgans parasimpàtics. De vegades els òrgans neurohemals són difosos. Les glàndules endocrines varien igualment segons els grups, i algunes de les denominacions especials són: la glàndula de la muda (o òrgan Y) i les glàndules androgèniques dels crustacis; les glàndules de la muda dels miriàpodes; les glàndules de la muda (o protoràciques) i els cossos alats dels insectes. No s’ha detectat la presència de glàndules endocrines en els quelicerats, però s’ha comprovat que la muda és induïda per una hormona, l’ecdisona, de composició semblant a l’hormona corresponent dels crustacis, els insectes i els miriàpodes, que fa la mateixa funció. El funcionament del sistema neuroendocrí en els artròpodes difereix notablement d’uns grups als altres.
En conjunt, el sistema neuroendocrí regula fenòmens tan importants com la muda, el creixement, el desenvolupament embrionari i especialment la metamorfosi, el procés de desenvolupament de les gònades i d’alguns caràcters sexuals secundaris, certes funcions metabòliques i fins i tot el canvi de color del cos segons un ritme diari, com en el cas d’alguns crustacis.
Els òrgans dels sentits
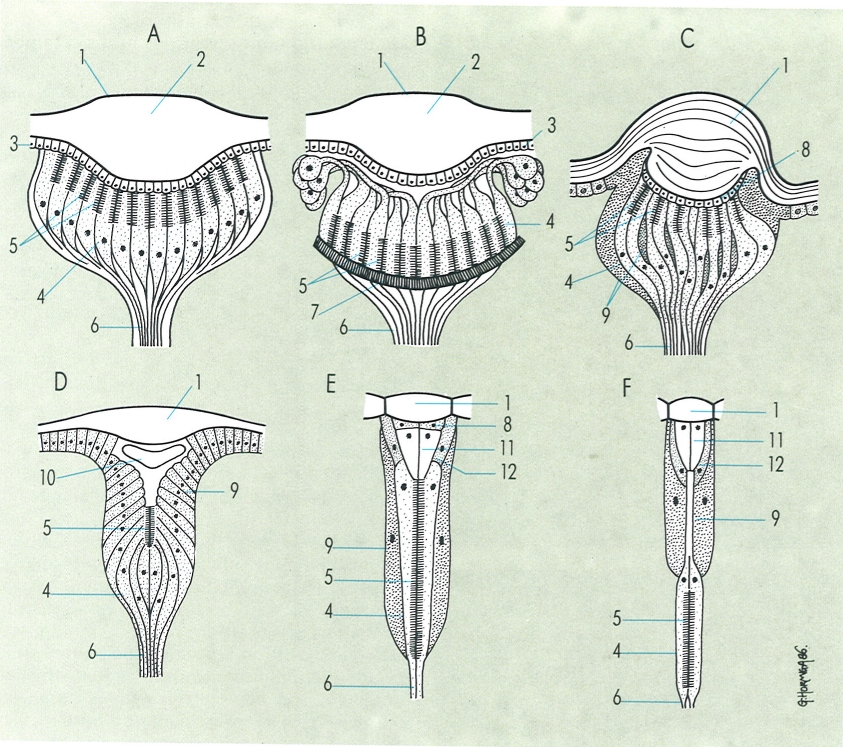
Diferents models d’òrgans visuals en els artròpodes: A ocel·le de visió directa d’aràcnid, B ocel·le de visió indirecta d’aràcnid, C ocel·le d’insecte, D estemmata d’insecte, E ommatidi d’aposició d’un ull compost d’insecte, F ommatidi de juxtaposició d’un ull compost d’insecte. Hom hi distingeix: 1 còrnia, 2 cristal·lí, 3 cos vitri (cornèola o capa de cèl·lules corneanes), 4 cèl·lules retinals, 5 rabdoma, 6 fibres nervioses que, en conjunt, formen el nervi òptic, 7 tapetum, 8 capa de cèl·lules corneanes, vítries o cristal·lines, 9 cèl·lules pigmentàries, 10 cos cristal·lí, 11 con cristal·lí, 12 cèl·lules del cristal·lí.
Gustavo Hormiga, a partir de fonts diverses
Els òrgans dels sentits es troben molt desenvolupats i diversificats en els artròpodes, i alguns arriben a atènyer un grau elevat de complexitat. El seu gran desenvolupament ha intervingut d’una manera decisiva en l’adaptació dels artròpodes a ambients molt variats i en l’adquisició d’una conducta complexa, especialment en els insectes superiors.
Malgrat la diversitat aparent, els òrgans sensorials mostren una mateixa organització general. Són formats per tres tipus d’elements cel·lulars, sempre de naturalesa ectodèrmica: una o diverses cèl·lules sensorials, que corresponen a cèl·lules epidèrmiques especialitzades, relacionades per mitjà de fibres sensitives amb un centre nerviós, que constitueixen el punt de partida d’un arc reflex, el qual comprèn una neurona d’associació i una neurona motora; formacions cuticulars, de morfologia i propietats variables, que asseguren la transmissió de l’excitació produïda per factors externs o interns a la cèl·lula sensorial; i elements annexos, representats per una o més cèl·lules epidèrmiques associades a la cèl·lula sensorial, la funció dels quals és secretar els elements cuticulars o aïllar la cèl·lula sensorial de l’ambient.

Els ulls capaços de formar imatges han aparegut a diferents moments i en grups independents al llarg de l’evolució dels artròpodes. És per això que la seva estructura no és uniforme, sinó que respon a diverses estratègies. Una d’elles és la de mantenir una lent única voltada de cèl·lules sensorials cobertes d’una capa de pigment i proveïda de músculs que permeten l’enfocament de les imatges; és el cas de les aranyes (a dalt, un saltícid). Una altra, la dels crustacis i els insectes (al mig, un dípter) consisteix a mantenir múltiples facetes. Els extingits trilobits posseïen ja un sistema visual facetat, com demostra l’estructura fossilitzada que veiem a la fotografia (a baix).
Ramon Torres i Enric Curto.

Diferents formacions sensorials dels artròpodes, vistes al microscopi electrònic d’escandallatge. A dalt, part apical de les antenes d’un diplur (campodèid), on s’observa la fosseta olfactòria dins la qual es localitzen els òrgans sensorials cupuliformes, de funció eminentment quimioceptora, voltats de diversos pèls de revestiment; també s’hi veuen formacions en roseta, situades entre els pèls i proveïdes d’un por central, que probablement correspon a una glàndula tegumentària, ja que es troba comunicat amb una petita cavitat on se situa una vesícula piriforme (× 1400). Al mig, pèls plomosos als quals, a causa de llur situació, hom atribueix una funció quimioceptora (× 320). A baix, una tricobòtria situada a les antenes d’un diplur (japídid), de funció mecanoceptora, és a dir, capaç de percebre vibracions (× 1080).
Marina Blas i Antoni Serra.
El conjunt dels elements esmentats constitueix una sensília. Les sensílies són, doncs, òrgans sensorials simples i aïllats, o bé cadascuna de les unitats funcionals que constitueixen un òrgan sensorial compost. Les sensílies més simples són els pèls sensorials, en els quals és senzill d’apreciar els diferents components. Les sensílies, i per tant els òrgans sensorials, es relacionen sempre amb el tegument, atès que són d’origen ectodèrmic. Per això, se situen a la cutícula del cos i als apèndixs, a l’estomodeu i al mesodeu, a les invaginacions traqueals, etc. Capten estímuls procedents del medi (òrgans exteroceptors) o interns (òrgans propioceptors o interoceptors). Hi ha diferents tipus de sensílies i diverses formes d’especialització. Segons la naturalesa de l’estímul que reben, hom distingeix diversos tipus d’òrgans sensorials en els diferents grups d’artròpodes, que tenen estructura i desenvolupament diferent. Poden ésser classificats en mecanoceptors, quimioceptors i fotoceptors, i de cadascun d’aquests tipus n’hi ha diverses variacions.
Els òrgans mecanoceptors més senzills corresponen als pèls o sedes tàctils, que es reparteixen per tot el tegument. Són estimulats pels desplaçaments del pèl respecte de la base articular, i actuen com a exteroceptors o com a propioceptors, segons quina sigui la seva localització al cos de l’animal. A més de percebre sensacions tàctils, es troben implicats en l’orientació dels organismes respecte a la gravetat, en l’orientació de la posició del cos, en la recepció de les vibracions del substrat (en medi sòlid o líquid) i en les que procedeixen del medi aeri. Reben noms particulars a cada grup. Els òrgans propioceptors també són mecanoceptors, en aquest cas estimulats per les pressions i tensions que s’originen a l’interior del cos.
Un tipus especialitzat de mecanoceptor és el format pels òrgans sensorials termoceptors, sensibles a la temperatura. Pel que sembla, però, en la percepció d’aquest tipus d’estímul també hi intervenen els òrgans visuals, especialment en la reacció dels organismes envers la font emissora de calor.
Els òrgans quimioceptors reaccionen a estímuls de naturalesa química. En les formes aquàtiques, aquestes substàncies són captades a partir del medi aquós, on es troben dissoltes; al medi terrestre poden ésser detectades a distància o bé per contacte. Les que es detecten a distància són gasos, i els òrgans que els capten s’anomenen olfactoris; les que es detecten per contacte són líquides o sòlides, i els òrgans receptors s’anomenen gustatius. En alguns casos, una mateixa àrea sensitiva respon a tots dos tipus d’estímul. Un tipus particular d’òrgan quimioceptor són els higroceptors, sensibles a la quantitat de vapor d’aigua de l’atmosfera, i típics de les espècies terrestres. Els òrgans quimioceptors són d’estructura simple, constituïts per una sola sensília, en forma de pèl o de papil·la. Se situen fonamentalment als apèndixs de la regió cefàlica, o també als altres apèndixs i a diferents regions del cos. Són d’una importància vital per a la coordinació de la conducta de les espècies. En general, els òrgans olfactoris són específics per a substàncies com les feromones (hormones produïdes per una espècie i que actuen sobre la mateixa espècie), les quals poden ser d’agregació, d’alarma, sexuals, morfogenètiques, etc. A les espècies paràsites són específics per a les substàncies que emeten els hostes, de manera que els faciliten la seva localització. Precisament, l’especificitat de molts paràsits respecte d’un hoste concret (sigui animal o vegetal) es deu a aquest fenomen. En altres casos, els òrgans olfactoris capten les olors procedents de les fonts d’aliment. En els insectes és on es presenten aquests tipus d’òrgans sensorials amb més perfeccionaments.
Els òrgans fotoceptors són estimulats per l’energia lluminosa i la sensació que produeixen és la captació, més o menys difusa, de la llum que arriba al medi. En els casos més evolucionats, aquests òrgans permeten reproduir l’ambient per mitjà de la formació d’imatges, i reben el nom d’òrgans visuals o ulls. En els artròpodes, la localització, l’organització i el nombre varia molt segons els grups, i també el tipus d’imatge que produeixen. Això no obstant, hom en pot reconèixer dos tipus principals: els ulls simples o ocel·les i els ulls compostos. A més, en alguns grups (larves i nimfes d’alguns insectes, adults d’alguns apterigots i alguns miriàpodes) hi ha un tipus d’ulls anomenat estemmata, que participen alhora de les característiques dels ocel·les i dels ulls compostos. Tot i que únicament hi ha dos tipus bàsics d’ulls, aquests no són homòlegs en els diferents grups d’artròpodes: els ocel·les dels aràcnids són diferents dels dels crustacis, els miriàpodes i els insectes, i els ulls compostos dels merostomats són diferents dels dels crustacis i els insectes.
Els ocel·les dels aràcnids i els picnogònids se situen, en nombre variable segons els grups, a la regió anterior del prosoma. La seva estructura varia segons la localització exacta en el prosoma i segons els grups que es consideren. En general, consten d’un aparell diòptric i d’un aparell sensorial o retina. L’aparell diòptric és format per la còrnia, el cristal·lí (que és la lent de l’ull) i el cos vitri, aquest constituït per una capa de cèl·lules (en alguns casos absent) que rep també el nom de corneola o capa de cèl·lules corneanes. La retina agrupa les cèl·lules visuals, que tenen, en un extrem, unes diferenciacions semblants a uns bastonets, els rabdòmers, que contenen els pigments fotosensibles. Les cèl·lules visuals poden restar aïllades o associar-se formant retínules. El conjunt de rabdòmers constitueix el rabdoma. De les cèl·lules retinals neixen les fibres nervioses que constitueixen el nervi òptic, el qual relaciona els ulls amb els centres ocel·lars del protocervell. Les cèl·lules retinals queden cobertes per dues capes de cèl·lules, la membrana pre-retinal i la post-retinal, que en els ulls de visió indirecta és fortament pigmentada i rep el nom de tapetum; generalment darrere de la membrana post-retinal hi ha una capa de cèl·lules pigmentàries. En els aràcnids hi ha, però, dos tipus fonamentals d’ocel·les, els de visió directa, característics de les espècies diürnes, i els de visió indirecta, típics de les nocturnes. En els ocel·les de visió directa, el rabdoma s’orienta cap a la font de claror, és a dir, en contacte amb l’aparell diòptric, mentre que en els ocel·les de visió indirecta, s’orienta en sentit contrari de manera que els raigs de llum han de travessar tota la capa retinal per a estimular el rabdoma. En aquests casos, en compensació, la membrana post-retinal presenta una capa cristal·lina brillant que reflecteix la llum, la qual és captada pels diferents rabdòmers. Hi ha nombroses variacions respecte als models generals d’ocel·les dels aràcnids. En molts casos, coexisteixen ambdós tipus d’ocel·les en un mateix individu.
Els ocel·les dels miriàpodes se situen en posició lateral al cap. La seva estructura fonamental és força semblant a la dels ocel·les dels crustacis i dels insectes, tret del cas dels escutigeromorfs (grup dels quilòpodes), els ulls dels quals són considerats per alguns autors com a ulls compostos modificats; d’altres autors consideren més correcte emprar el terme estemmata.
Els ocel·les dels crustacis i dels insectes se situen dorsalment al cap. Consten d’una còrnia, més engruixida al centre que als voltants, que actua com una lent biconvexa, la qual és segregada per una capa de cèl·lules anomenades corneanes, vítries o cristal·lines, capa que pot tenir un desenvolupament molt variable, i fins i tot pot mancar. Directament relacionada amb la còrnia o amb la capa de cèl·lules corneanes hi ha la retina, formada per cèl·lules visuals reunides en retínules i separades per cèl·lules pigmentàries. Al centre de cada retínula hi ha el rabdoma. Tot el conjunt d’elements que formen l’ocel·le és voltat d’una capa de cèl·lules pigmentàries que constitueixen una capa refractària: la membrana postretinal o membrana basal. De les cèl·lules retinals en surten les fibres que formen el nervi ocel·lar, que relaciona els ulls amb el centre ocel·lar del protocervell.
Els ulls laterals de larves, nimfes o adults d’alguns insectes substitueixen, per la seva situació, els ulls compostos. Per això, hom pot pensar que es tracta d’ulls compostos reduïts, fets de pocs ommatidis, amb cristal·lí absent o rudimentari i mancats de capa de cèl·lules corneanes. Bé que són formacions larvàries, de transició, hi ha insectes que els tenen en l’estat d’adult (col·lèmbols, sifonàpters etc., i també certs miriàpodes). Els estemmates són innervats, com els ulls compostos, pel lòbul òptic del protocervell.
Els ulls compostos són característics dels crustacis, els insectes i els xifosurs (merostomats) actuals, i també dels trilobits i merostomats fòssils. Són formats per la juxtaposició d’unitats funcionals (sensílies) anomenades ommatidis. Cada ommatidi porta el seu aparell diòptric independent, tret d’algunes excepcions (els merostomats xifosurs), als quals és comú. La cara externa de l’ommatidi, hexagonal en insectes i quadrangular en els crustacis, es renova periòdicament a cada muda, i rep el nom de faceta. Cada ommatidi es compon d’una faceta o còrnia (elaborada per cèl·lules corneanes laterals, sovint pigmentades) per sota la qual hi ha el cristal·lí o con cristal·lí, format en general per la juxtaposició de quatre peces, secretades per les cèl·lules cristal·lines. El con cristal·lí concentra i transmet la llum cap al rabdoma. En relació amb el cristal·lí se situa la retínula, formada per un nombre variable de cèl·lules visuals, les quals porten, a la seva cara interna, els rabdòmers, que constitueixen el rabdoma. Aquesta és la regió fotosensible de l’ommatidi. Cada ommatidi queda envoltat de cèl·lules pigmentàries, que l’aïllen dels ommatidis veïns. A partir de la regió distal de les cèl·lules retinals es diferencien les fibres nervioses que formen el nervi òptic, el qual relaciona els ommatidis amb el lòbul òptic del protocervell. Hi ha nombroses variacions d’aquest model general d’ommatidi.
Segons l’estructura i la disposició de les cèl·lules pigmentàries, hi ha dos tipus principals d’ulls i de visió: els ulls d’aposició o de visió diürna, i els de superposició o de visió nocturna. En els ulls d’aposició, les cèl·lules pigmentàries se situen envoltant completament cada ommatidi, impedint així que un mateix raig de llum en travessi més d’un. D’altra banda, capten únicament els raigs que incideixen perpendicularment en l’aparell diòptric. Cada ommatidi forma una imatge i, per tant, es produeixen tantes imatges com ommatidis hi ha, nítides però poc lluminoses, atès que aprofiten únicament una petita fracció dels raigs lluminosos que arriben als ulls. Per això, són característics de les espècies diürnes. En els ulls de superposició, les cèl·lules pigmentàries es concentren d’una manera discontínua a la regió proximal i distal de l’ommatidi, de manera que la regió intermèdia queda oberta als raigs lluminosos, els quals poden travessar diversos ommatidis. D’aquesta manera es produeix la superposició de les imatges respectives, cosa que fa que aquestes siguin més lluminoses (perquè aprofiten tots els raigs de llum que incideixen sobre els ulls), però menys nítides. Aquest tipus de visió és característic de les espècies nocturnes. En alguns casos, els ulls i la visió poden ser alternativament per aposició i per superposició. Això és degut al fet que el pigment dels ommatidis pot experimentar migracions en funció de la quantitat de llum que hi ha al medi. Aquest fenomen s’aprecia en alguns grups de crustacis i insectes.
Els diferents tipus d’ulls tenen graus i característiques de percepció diferents. Els ocel·les perceben els canvis d’intensitat lluminosa del medi i intervenen d’una manera decisiva en l’orientació direccional dels organismes. Els ulls compostos reprodueixen l’ambient per mitjà de la formació d’imatges, en alguns casos amb percepció de colors, i també són sensibles als canvis d’intensitat lluminosa.
Els òrgans visuals són molt importants per a la vida dels organismes donat que la quantitat de llum que hi ha a l’entorn és un factor que actua sobre la seva activitat. No obstant, això, l’equilibri perfecte de l’organisme en el seu medi és el resultat del funcionament coordinat de tots els seus receptors sensorials (mecanoceptors, quimioceptors i fotoceptors).
L’aparell digestiu i l’alimentació
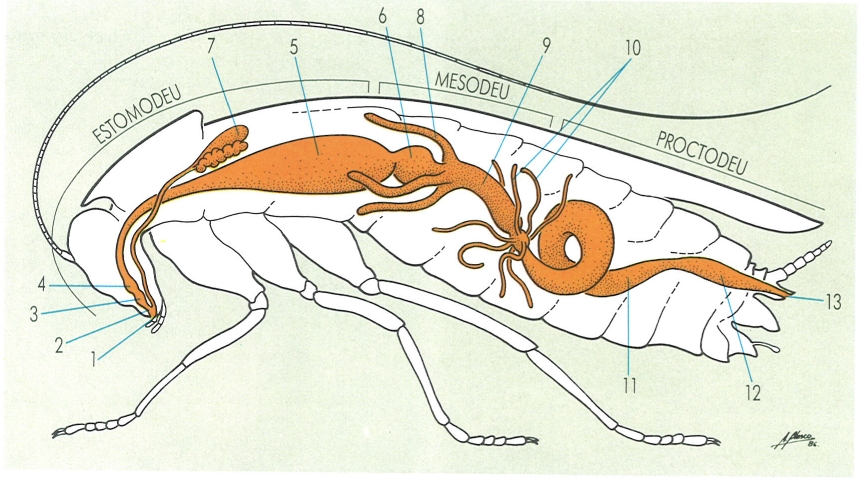
L’aparell digestiu dels artròpodes, en aquest cas exemplificat i simplificat en una panerola (insecte blàtid), consta de tres trams ben diferenciats i d’unes estructures bàsiques, que varien segons l’especialització alimentària. Els elements principals de l’aparell digestiu i altres estructures relacionades, presents a la majoria dels artròpodes, s’indiquen a continuació: 1 boca, 2 cavitat bucal, 3 faringe, 4 esòfag, 5 pap, 6 proventricle, 7 glàndules salivals, 8 cecs digestius, 9 intestí glandular, 10 tubs de Malpighi, 11 ili, 12 recte, 13 anus.
Amadeu Blasco, a partir de fonts diverses
En els artròpodes, l’aparell digestiu mostra una gran varietat de formes, però, en general, és rectilini i més o menys llarg en funció del règim alimentari. S’hi distingeixen tres regions, una d’anterior o estomodeu, una d’intermèdia o mesodeu, i una de posterior o proctodeu, a cadascuna de les quals s’hi diferencien regions amb funcions diferents, que reben noms especials.
L’estomodeu s’inicia a la boca, prop de la qual se situen els apèndixs bucals, quan n’hi ha, en nombre i importància molt variable. Ací es fa la trituració de l’aliment i se n’inicia la digestió, gràcies a l’acció de glàndules annexes, com les glàndules salivals, que hi desemboquen. És d’origen ectodèrmic, i, per tant, és revestit de cutícula. En molts casos l’estomodeu va proveït de denticles i formacions quitinoses que intervenen en la trituració de l’aliment. Comprèn diferents zones, com la cavitat bucal, la faringe, l’esòfag, el pap, el pedrer, el molinet gàstric, etc., diferents a cada grup i relacionades amb el tipus d’aliment (sòlid o líquid, tou o dur, etc.).
El mesodeu és d’origen endodèrmic i també s’anomena estómac glandular perquè és la regió del tub digestiu on té lloc la digestió enzimàtica de l’aliment que ha estat prèviament triturat a l’estomodeu. També s’hi fa l’absorció dels productes digerits. Generalment aquest tram del tub digestiu és tubular, i de vegades s’anomena intestí. Pot presentar cecs digestius, com en el cas dels quelicerats. El nombre de glàndules annexes, la seva forma i la disposició són molt variables segons els grups. Són glàndules que, amb les seves secrecions, participen en la digestió, l’excreció, l’absorció i l’acumulació de substàncies de reserva.
El proctodeu és la regió terminal del tub digestiu i també rep el nom d’intestí posterior. Desemboca a l’exterior a través de l’anus. És d’origen ectodèrmic, per la qual cosa va revestit de cutícula, que es renova a cada muda. És en aquesta zona on es formen els excrements, per mitjà de la reabsorció d’aigua, i d’aquesta manera contribueix a l’osmoregulació interna. En relació amb el proctodeu pot haver-hi glàndules rectals, de funcions diverses.
Entre l’estomodeu i el mesodeu, i entre el mesodeu i el proctodeu, hi ha unes vàlvules o esfínters que regulen el pas de l’aliment. En els artròpodes terrestres (aràcnids, miriàpodes i insectes), a la zona límit entre el mesodeu i el proctodeu hi ha uns òrgans excretors especials, els tubs de Malpighi, que aboquen al tub digestiu diversos productes que cal eliminar del cos.

Els hàbits alimentaris formen part d’una estratègia particular de cada espècie, la qual condiciona, no només la seva conducta, sinó també la seva morfologia i la seva distribució. A les fotografies hom vol il·lustrar tres tipus d’alimentació: la dels carnívors (a dalt, a l’esquerra), dels quals veiem la cranca de nas llarg (Pisa armata, colonitzada al dors per colònies de briozous), la dels paràsits (a baix), com Lernanthropus kroyerii, copèpode que viu a les brànquies dels peixos, i la dels que s’alimenten de productes vegetals (a dalt, a la dreta), amb múltiples especialitzacions, com la del lepidòpter libador de la fotografia (gènere Hemaris), que amb la seva llarga trompa, xucla el nèctar de les flors.
Xavier Palaus, Mercè Durfort i Enric Curto.
La paret del tub digestiu, especialment en el mesodeu, presenta músculs que asseguren els moviments peristàltics, tot afavorint el desplaçament de l’aliment. En alguns grups (especialment els insectes, i també els crustacis copèpodes i decàpodes), quan l’aliment arriba al mesodeu, queda embolicat dins d’una membrana especial, la membrana peritròfica, que protegeix el delicat epiteli intestinal de l’acció mecànica de l’aliment. Aquesta membrana es va formant a mesura que els aliments arriben al mesodeu, i s’elimina amb els excrements, de manera que es renova constantment.
La digestió dels aliments en general és interna, és a dir, que es fa a l’interior del tub digestiu. En els grups que no presenten peces bucals diferenciades, com els aràcnids, és externa: aboquen els sucs digestius a l’exterior, al damunt de les preses capturades, i posteriorment absorbeixen els líquids resultants de la digestió parcial de l’aliment i els acaben de digerir en el mesodeu.
L’èxit numèric d’un determinat grup animal és en gran part funció de la seva versatilitat alimentària, és a dir, de la gamma d’aliments que pugui utilitzar, i dels mitjans de què disposi per a aconseguir-los. Els insectes ens proporcionen un bon exemple d’aquesta versatilitat: són capaços d’alimentar-se de materials durs (llavors, grans, pol·len, etc.), de líquids (sang, saba, nèctar, etc.), de preses vives que capturen ells mateixos, de matèria orgànica en descomposició (cadàvers, excrements, restes vegetals, etc.) i de materials com fusta, pell, plomes, suro, tabac, etc., que solen constituir l’aliment pràcticament exclusiu d’algunes espècies.
Segons les fonts d’aliment, els artròpodes es poden classificar, a grans trets, en omnívors (polífags), carnívors (zoòfags), herbívors (fitòfags) i sapròfags. Els omnívors presenten una dieta alimentària àmplia i variada, composta de matèria orgànica de qualsevol procedència. Els carnívors es nodreixen de carn o dels fluids interns d’altres animals vius. Els herbívors utilitzen com a font d’aliment les plantes o els seus líquids interns (saba, nèctar, etc.). Els sapròfags s’alimenten de matèria orgànica (animal o vegetal) en descomposició. Dins de cadascuna d’aquestes categories hi ha nombroses especialitzacions, i cada tipus de dieta rep un nom específic. Els artròpodes, en general, actuen d’una manera activa per localitzar i capturar els seus aliments. Com a excepció hi ha el cas dels endoparàsits, que s’alimenten, d’una manera passiva, de les molècules del medi que difonen directament al seu interior.
Segons la grandària de l’aliment, i independentment de la seva naturalesa, els artròpodes es poden classificar en macròfags i micròfags, dels quals se separen també els que s’alimenten de fluids. La macrofàgia és característica d’un gran nombre d’espècies, totes proveïdes d’apèndixs bucals o formacions especials per a triturar l’aliment. L’aliment és de naturalesa diversa: petits animalets o algunes de les seves parts, i matèria vegetal, com fulles, fusta, fruits, etc. Són macròfags els merostomats, que trituren les algues amb la base dels apèndixs del prosoma, molts crustacis, els miriàpodes i un gran nombre d’insectes, tots proveïts d’apèndixs bucals (mandíbules i un o dos parells de maxil·les, i en alguns casos, altres apèndixs accesoris). Les espècies micròfagues s’alimenten de partícules petites, que es troben en suspensió en el medi. Per això, el règim micròfag és característic de les espècies aquàtiques i intersticials. Per a aconseguir l’aliment, es requereixen apèndixs especialitzats, capaços de produir corrents d’aigua en direcció a la boca, o capaços també d’actuar com a filtres per a la selecció i concentració de les partícules en suspensió. Es comporten com a micròfags un gran nombre de crustacis i algunes larves aquàtiques d’insectes. Els crustacis micròfags, en general provoquen corrents d’una manera activa, per mitjà del moviment de determinats apèndixs o regions del cos. Les larves i nimfes d’insecte, en canvi, acostumen a presentar estructures de filtració i, en lloc de produir corrents d’aigua, se situen elles mateixes de manera que puguin aprofitar els corrents naturals de l’aigua; generalment viuen a les aigües continentals (rabeigs de rius, llacs, llacunes, etc).
Hi ha un gran nombre d’espècies que s’alimenten de fluids, sigui d’origen animal (sang, humors dels teixits, secrecions, etc.) o vegetal (saba, nèctar, secrecions, etc.). Presenten mecanismes per a fixar-se al substrat sobre el qual s’alimenten, i per a perforar la paret del cos i els teixits, i també aparells especialitzats per a xuclar-ne o llepar-ne els líquids nutritius. Les espècies endoparàsites tenen mecanismes que els permeten d’introduir-se a l’interior dels seus hostes, dels quals s’alimenten posteriorment. L’alimentació a base de fluids és característica dels artròpodes paràsits d’animals i vegetals, com certs insectes i crustacis i alguns aràcnids (especialment, àcars). També la majoria dels aràcnids s’alimenten de fluids, donat que, tot i ésser grans depredadors i capturar preses, s’alimenten dels seus sucs interns; no tenen peces trituradores a la boca i fan la digestió externament al seu cos.
L’aparell respiratori i la respiració
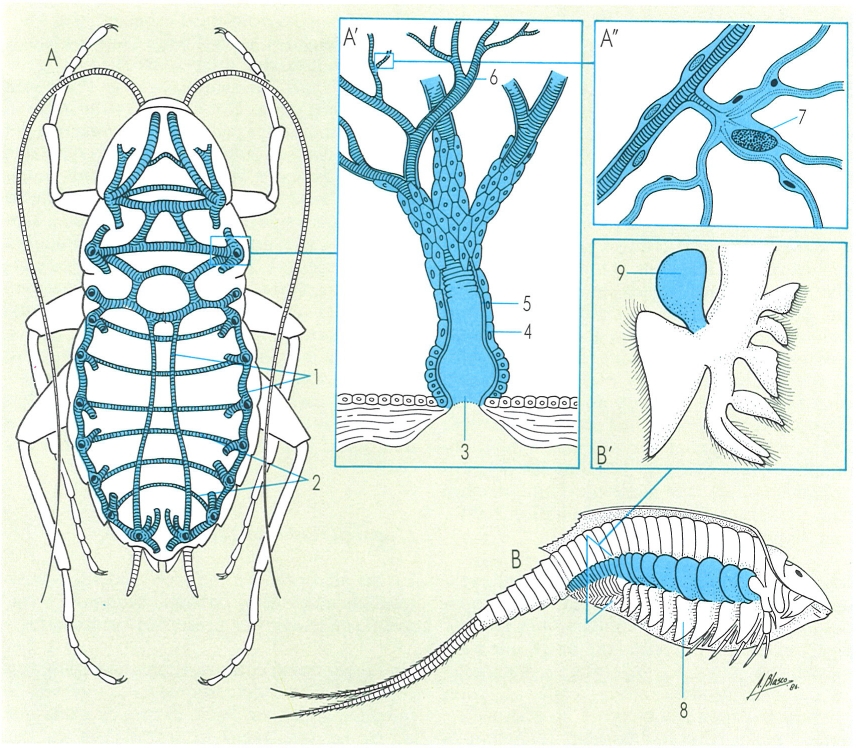
Els sistemes respiratoris dels artròpodes consten de dispositius destinats a augmentar la superfície d’intercanvi de gasos i a transportar-los a tot l’organisme. El dibuix il·lustra el sistema traqueal, típic dels insectes (A aspecte general, corresponent a la cara ventral del cos, A’ detall d’una tràquea, A" detall de les traquèoles), i el sistema branquial dels crustacis (B aspecte general, B’ detall). Hom hi distingeix: 1 tubs o troncs traqueals longitudinals, 2 troncs traqueals transversals, 3 estigma, 4 epidermis, 5 cutícula, 6 traquèoles, 7 cèl·lules traqueals o traqueoblasts, 8 brànquies, 9 epipodi.
Amadeu Blasco, a partir de fonts diverses
En els artròpodes hi ha tres modalitats bàsiques de respiració: cutània, branquial i traqueal, que sovint es presenten combinades.
En la respiració cutània, l’intercanvi de gasos (oxigen i diòxid de carboni) es fa a través de la cutícula que recobreix el cos, és a dir, que no hi ha una regió localitzada on es faci aquest intercanvi. Aquest és el tipus de respiració més primitiu, donat que no comporta l’existència d’òrgans especialitzats; entre els artròpodes actuals es troba en les formes aquàtiques i petites, especialment de crustacis (copèpodes, alguns ostràcodes, cirrípedes i fins i tot determinats decàpodes), en els picnogònids, i en algunes formes terrestres, com els pauròpodes i alguns insectes (certs collèmbols i proturs), en els aràcnids (certs àcars) i excepcionalment en altres grups. Els organismes terrestres de respiració cutània han de viure en medis saturats d’humitat per tal d’evitar la deshidratació, i fonamentalment es distribueixen pel medi edàfic.
Al llarg de l’evolució, en augmentar les dimensions corporals i impermeabilitzar-se el tegument, la majoria dels artròpodes, siguin aquàtics o terrestres, han anat desenvolupant òrgans respiratoris localitzats en determinades zones del cos i dels apèndixs, brànquies o tràquees, segons els grups.
La respiració branquial és característica dels grups aquàtics (crustacis, merostomats i trilobitomorfs). L’intercanvi de gasos es fa en uns òrgans especials, les brànquies, que es localitzen en els apèndixs. El prototipus de les brànquies dels crustacis correspon a un lòbul pla (l’epipodi) que té el tegument molt prim i molt vascularitzat. A partir d’aquest model se n’han originat d’altres més complexos, en els quals hi ha un eix central amb ramificacions laterals, laminades, arborescents o filamentoses. Als crustacis, les brànquies es localitzen principalment en els pereiopodis, però en els isòpodes i els hoplocàrides es relacionen amb els pleopodis. El nombre de brànquies i les seves dimensions varia molt en relació a l’hàbitat i la conducta de les diferents espècies. En general, les brànquies són lliures, però en els decàpodes se situen a l’interior d’unes cambres especials, les cambres branquials, que s’originen a partir d’unes expansions del cefalotòrax. En els merostomats (els xifosurs), les brànquies se situen als cinc darrers parells d’apèndixs de l’opistosoma. Aquests apèndixs, plans i molt vascularitzats, presenten a la seva superfície interna nombrosos plecs molt fins, disposats com els fulls d’un llibre, a través dels quals es fa l’intercanvi de gasos. En el grup fòssil dels trilobits, les brànquies se situaven a l’exopodi de tots els apèndixs del cos, que presentava un aspecte filiforme. Independentment de la seva localització, la ventilació de les brànquies s’aconsegueix per mitjà del moviment rítmic de tots o d’alguns dels apèndixs del cos, i fins i tot del de les diferents regions del cos i dels metàmers que les integren.
A més de les brànquies típiques, hi ha altres tipus de brànquies especialitzades al medi terrestre, com és el cas de les brànquies d’alguns crustacis isòpodes, anomenades pseudotràquees, que corresponen a brànquies modificades. En d’altres casos, com en alguns cirrípedes (els bàlans) i en certs isòpodes, s’han desenvolupat estructures especials que protegeixen les brànquies de la dessecació quan els animals resten fora de l’aigua.
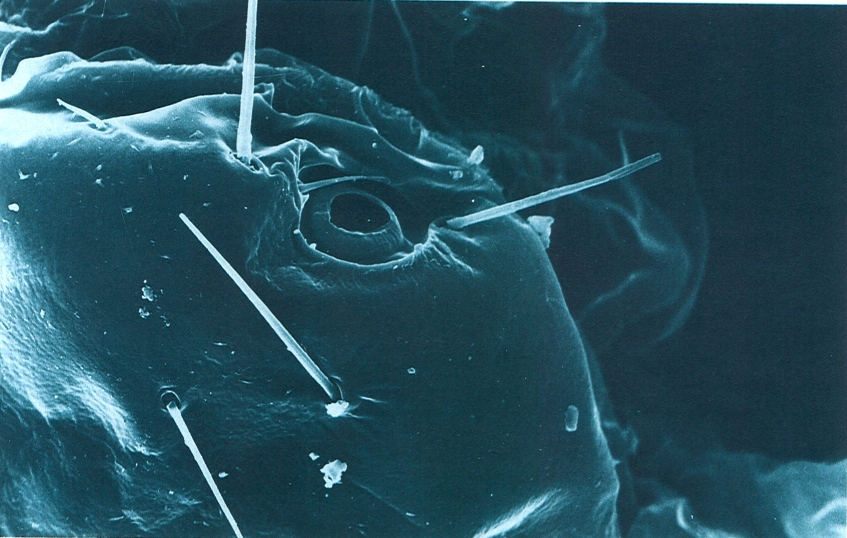
Estigma respiratori d’un insecte protur, situat dorsilateralment al mesotòrax, vist al microscopi electrònic d’escandallatge (× 2480). Hom hi distingeix, ultra l’obertura estigmàtica, les sedes protectores que l’envolten i uns pèls llargs i rígids, particulars de l’espècie fotografiada.
Marina Blas.
La respiració traqueal és una conseqüència de l’adaptació al medi terrestre, i és característica de bona part dels artròpodes terrestres (aràcnids, miriàpodes i insectes). En aquest cas, l’intercanvi de gasos es fa a través d’unes invaginacions del tegument, anomenades tràquees, per les quals circulen els gasos relacionats amb la respiració. Hi ha dos tipus principals de tràquees: les fil·lotràquees i les dendrotràquees.
Les fil·lotràquees o pulmons dels aràcnids consisteixen en unes invaginacions globoses del tegument que tenen la paret interna recoberta de làmines amples i primes, les quals es mantenen separades per mitjà d’unes formacions especials. L’estructura d’aquests pulmons recorda la de les brànquies dels merostomats. Els espais que queden entre les làmines dels pulmons sempre és ple d’aire, la qual cosa permet l’intercanvi de gasos amb l’hemolimfa, que les irriga profusament. Els pulmons comuniquen amb l’exterior a través d’unes escletxes, generalment ovalades, que se situen a la superfície ventral de l’opistosoma. En alguns aràcnids, com els escorpins, hi ha únicament pulmons, mentre que en d’altres, com els àcars, hi ha exclusivament dendrotràquees, i en d’altres encara, com els aranèids, coexisteixen tràquees d’ambdós tipus.
Les dendrotràquees o tràquees típiques, són invaginacions tubulars del tegument, més o menys ramificades, que es fiquen per la cavitat general del cos, tot envoltant els diferents òrgans i teixits, i penetrant-los. Comuniquen amb l’exterior a través d’uns orificis parells anomenats estigmes. En molts casos hi ha mecanismes d’oclusió que regulen l’obertura i el tancament dels estigmes, i, per tant, l’entrada i la sortida de l’aire que circula per l’interior de tot el sistema. Originalment, les tràquees tenen una disposició metamèrica; en els grups més primitius, n’hi ha un parell a cada metàmer, independents l’una de l’altra, però el cas més corrent és que les tràquees dels diferents metàmers es comuniquin i tinguin troncs longitudinals comuns. Moltes vegades, això porta també a la reducció del nombre d’estigmes, i llavors aquests es localitzen únicament a determinades regions del cos. Les ramificacions traqueals es van fent cada vegada més primes alhora que augmenta la permeabilitat de les seves parets. Totes les ramificacions són permeables als gasos respiratoris, però el pas dels líquids es restringeix a les traquèoles, és a dir, les ramificacions més primes. Les traquèoles porten, a l’extrem terminal, unes cèl·lules estrellades, dites cèl·lules traqueolars, que amb les seves prolongacions envolten les cèl·lules dels teixits i arriben a penetrar-hi. És en aquest punt on es fa, principalment, l’intercanvi respiratori. La capacitat respiratòria dels diferents trams de les tràquees es deu a l’estructura diferencial de la cutícula. Pels tubs traqueals circula l’aire, captat directament del medi, el qual és difós cap a les cèl·lules. En aquest transport no hi intervé l’hemolimfa. La circulació dels gasos pot ser passiva o activa. Hi ha diversos mecanismes que regulen l’entrada i la sortida de l’aire de les tràquees, sigui mecanismes directes, que únicament impliquen les tràquees, o indirectes, que impliquen el moviment de determinades regions del cos, com és el cas dels grills i les llagostes (els ortòpters), els quals, amb els moviments abdominals, regulen la inspiració i l’expiració dels gasos. Malgrat l’avantatge que representa la respiració traqueal en els artròpodes terrestres, suposa alhora un perill constant de deshidratació, que obliga a l’existència de mecanismes especials d’oclusió dels estigmes.
Hi ha encara un altre tipus d’òrgan respiratori, les traqueobrànquies, propi d’alguns artròpodes terrestres que secundàriament han tornat al medi aquàtic. Les traqueobrànquies permeten captar l’oxigen dissolt en l’aigua, i són característiques de molts insectes aquàtics. També hi ha casos d’organismes aquàtics que únicament són capaços de captar els gasos de l’aire, especialment entre els insectes. En aquests casos, hi ha tot un conjunt d’adaptacions que permeten prendre una reserva d’aire i emmagatzemar-la mentre l’insecte es troba submergit en l’aigua.
La principal diferència entre els diversos tipus de respiració és que, en la cutània i la branquial, l’hemolimfa, proveïda de pigments respiratoris, transporta els gasos per tot el cos, de manera que els sistemes respiratori i circulatori es troben estretament relacionats. En el cas de la respiració traqueal, en canvi, ambdós sistemes són independents, atès que els gasos circulen exclusivament a través de les tràquees, i l’hemolimfa generalment no porta pigments respiratoris.
L’aparell circulatori i la circulació
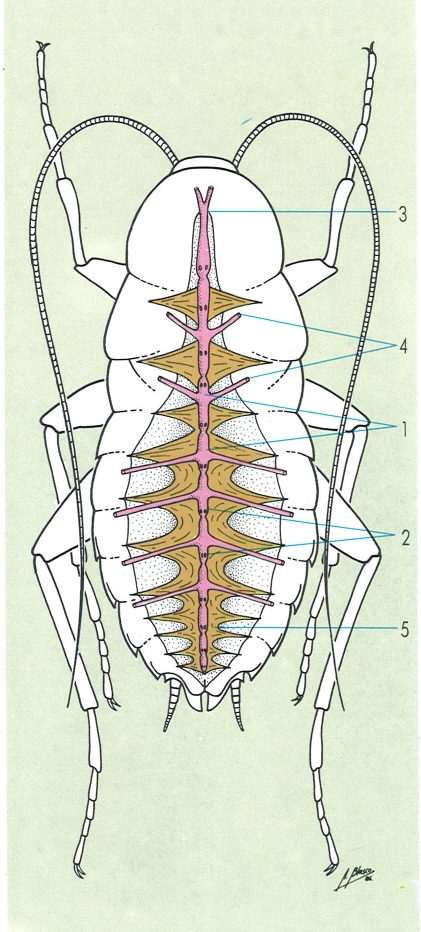
L’aparell circulatori dels artròpodes és format per un cor dorsal, diversos vasos i uns sins on es recull tota la sang abans que torni al cor. El cor no és més que un vas dorsal al qual es formen cambres o ventricles metamèrics, cadascun proveït d’un parell d’ostíols per on entra la sang. Els músculs alars mantenen el cor al seu lloc i col·laboren als moviments de contracció. Hom distingeix a la il·lustració: 1 ventricles, 2 ostíols, 3 artèria anterior, 4 artèries laterals, 5 músculs alars.
Amadeu Blasco, a partir de fonts diverses
L’aparell circulatori dels artròpodes és obert i lacunar: l’hemolimfa banya directament les cèl·lules que formen els òrgans i els aparells. Es caracteritza per la presència d’un òrgan propulsor contràctil, el cor, i un sistema de vasos.
El cor se situa en posició dorsal, inclòs dins del si pericàrdic. Es compartimentat en una sèrie de cambres separades per vàlvules que impedeixen el retrocés de l’hemolimfa. Cada cambra porta un parell d’orificis, els ostíols, pels quals entra l’hemolimfa.
El sistema arterial té un desenvolupament molt divers, segons els grups. De l’extrem anterior del cor parteix una artèria, l’aorta anterior, que posteriorment es pot ramificar. De l’extrem posterior pot sortir-ne l’aorta posterior, i de vegades artèries laterals. Quan el cor es contrau (moviment de sístole), l’hemolimfa surt del cor per les artèries i va a parar als sins i llacunes de la cavitat general del cos, i posteriorment al si pericàrdic. Durant el moviment de diàstole, un moviment de dilatació del cor fa que s’obrin els ostíols, i l’hemolimfa torna a entrar al cor. L’hemolimfa, doncs, circula alternativament pels vasos, les llacunes i els sins, i és per això que la circulació es diu oberta i lacunar. El sistema arterial descrit només es troba ben desenvolupat en els grups de respiració branquial, com la gran part dels crustacis, els merostomats i els aràcnids de respiració pulmonar. En la majoria dels artròpodes de respiració traqueal, especialment en els insectes, aquest sistema es troba força reduït, perquè les tràquees substitueixen fisiològicament, en molts aspectes, l’aparell circulatori.
El sistema venós generalment és poc desenvolupat. Al llarg de l’evolució hi ha una tendència general a la reducció de la grandària del cor i de l’aparell circulatori en general. Alguns artròpodes de petites dimensions, com els crustacis copèpodes, els pauròpodes (miriàpodes), els proturs (insectes) i d’altres, no tenen cor ni vasos diferenciats, i l’hemolimfa els circula exclusivament gràcies a moviments musculars.
L’hemolimfa dels artròpodes és un teixit líquid, pel qual circulen metabòlits intermediaris dels processos químics. Es compon d’un fluid que porta diverses substàncies dissoltes i cèl·lules sanguínies o hemòcits en suspensió. Alguns grups porten els pigments respiratoris dissolts al plasma, en lloc de concentrar-los a les cèl·lules sanguínies. Els pigments respiratoris són, fonamentalment, hemoglobines (en els crustacis entomostracis i les larves d’alguns insectes aquàtics) i hemocianines (en els crustacis malacostracis, els xifosurs, els escorpins, etc.), i predominen en els grups de respiració branquial.
Els òrgans excretors i l’excreció
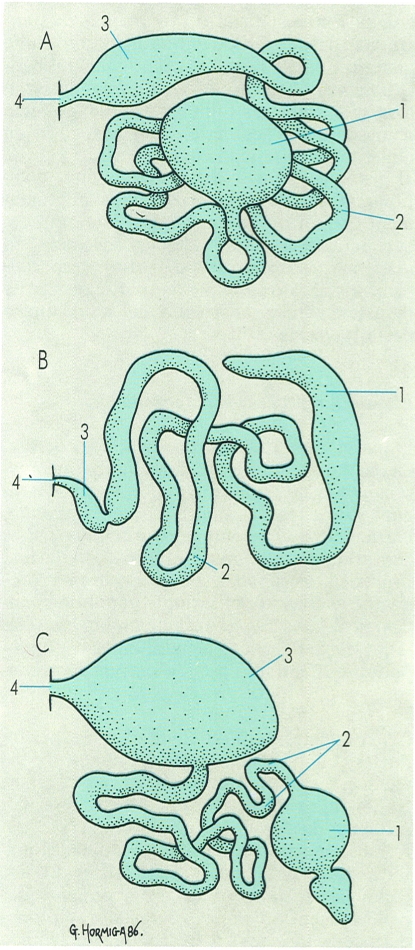
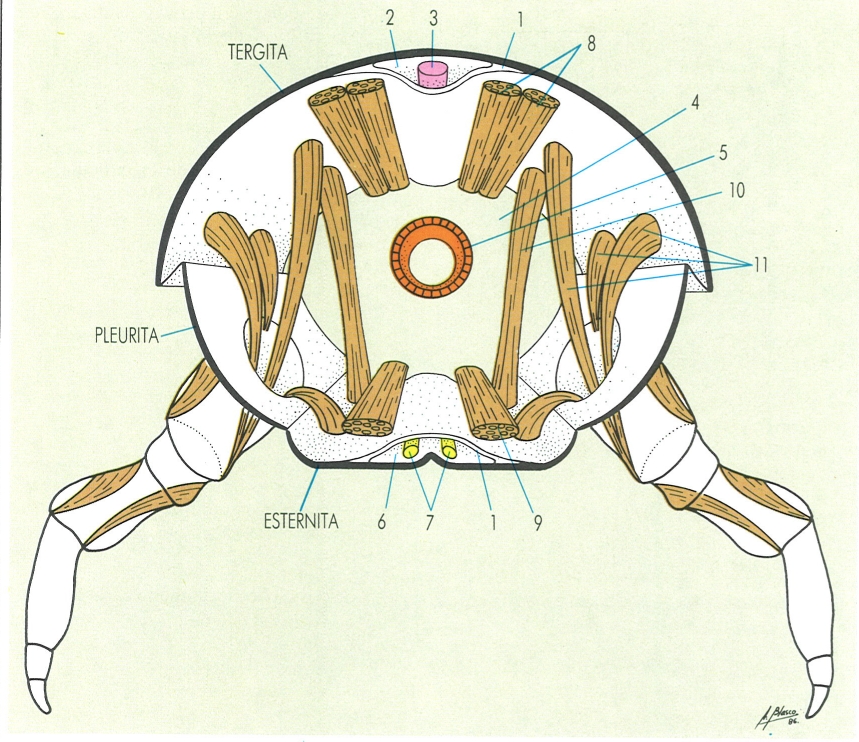
Col·laboren a l’excreció, en els artròpodes, determinades glàndules d’origen metamèric, és a dir, situades originalment a cada segment. Generalment es relacionen amb els apèndixs corresponents, cosa que explica que, malgrat la seva funció excretora, prenguin noms relatius a apèndixs que fan funcions diverses. Així, en els aràcnids, hi ha les glàndules coxals (A), i en els crustacis, les glàndules antenals (B) i maxil·lars (C). En els insectes adopten diversos noms, com els de ronyons cefàlics, glàndules labials, maxil·lars, etc. Totes, però, tenen una estructura similar. A la il·lustració, hom distingeix: 1 sac, 2 nefroducte, 3 bufeta, 4 nefropor.
Gustavo Hormiga, a partir de fonts diverses
Els artròpodes presenten dos tipus bàsics d’òrgans excretors: metamèrics, és a dir, d’origen mesodèrmic, i no metamèrics, en aquest cas, d’origen discutit, però certament no mesodèrmic.
Els òrgans excretors metamèrics consten d’una bossa terminal i un tub excretor o celomiducte, de llargada variable, en el qual es pot diferenciar una petita dilatació, anomenada bufeta, prop de l’orifici excretor o nefropor. El tram final del conducte excretor pot ser d’origen ectodèrmic. Els orificis excretors, sempre parells, se situen en relació amb determinats apèndixs del cos, cosa que demostra el seu origen metamèric. Aquests tipus d’òrgans excretors són característics dels crustacis, els xifosurs (merostomats) i els aràcnids, bé que també poden presentar-se en alguns miriàpodes i insectes. Als crustacis prenen el nom de glàndules antenals o maxil·lars, segons si els orificis se situen a la base del segon parell d’antenes o del segon parell de maxil·les; en general, tret del cas dels ostràcodes, no coexisteixen ambdós tipus, però, en canvi, sí que poden succeir-se al llarg del desenvolupament post-embrionari. Als xifosurs i als aràcnids reben el nom de glàndules coxals, perquè l’orifici se situa a la base de les coxes dels apèndixs del prosoma i el seu nombre oscil·la entre 4 parells (en els xifosurs) i un sol parell (en alguns aràcnids); en alguns casos poden faltar, i llavors l’excreció es fa per mitjà dels tubs de Malpighi, que poden coexistir amb les glàndules coxals, especialment en els aràcnids. En alguns miriàpodes i certs insectes apterigots, aquests òrgans excretors metamèrics es relacionen amb les peces bucals, i fins i tot amb les antenes (en els pauròpodes) i reben el nom de ronyons cefàlics, labials o maxil·lars; no obstant això, en aquests grups, els tipus d’òrgans excretors predominants són els tubs de Malpighi. Finalment, les glàndules salivals també s’han volgut interpretar com a òrgans excretors metamèrics especialitzats.
Els òrgans excretors no metamèrics s’anomenen tubs de Malpighi. Es troben a la majoria dels insectes (tret dels col·lèmbols i àfids, entre altres), en els miriàpodes i en alguns aràcnids. Consisteixen en una sèrie de tubs prims i cecs per un extrem, que floten lliurement en la cavitat general del cos i desemboquen al tub digestiu, a la zona límit entre el mesodeu i el proctodeu. En algunos casos (certs insectes) l’extrem lliure es pot unir al tub digestiu. No hi ha acord sobre si l’origen dels tubs de Malpighi és endodèrmic (és a dir, derivats del mesodeu) o ectodèrmic (derivats del proctodeu). El nombre de tubs de Malpighi és variable. La seva funció és captar els productes residuals que porta l’hemolimfa per mitjà de la filtració, i després abocar-los al tub digestiu, des d’on són expulsats a l’exterior juntament amb els productes no digeribles. En alguns casos, participen en l’excreció unes cèl·lules especialitzades, com els nefròcits dels aràcnids i els crustacis, i les cèl·lules pericardials dels insectes.
Els tubs de Malpighi filtren l’hemolimfa i en recullen els productes residuals per tal d’abocarlos al darrer tram del tub digestiu. Per això, generalment tenen l’extrem lliure, surant a la cavitat general del cos (a dalt), però a vegades presenten l’extrem adossat al final del recte (criptonefridisme) per tal de reabsorbir l’aigua (a baix). Hom diferencia: 1 mesodeu, 2 proctodeu, 3 tubs de Malpighi llargs, 4 tubs curts, 5 tubs criptonefridiats, 6 regió criptonefridiada.
Gustavo Hormiga, a partir de fonts diverses
Excreció significa, per definició, separació dels productes residuals dels fluids interns i expulsió a l’exterior. Aquests productes són el resultat del metabolisme dels organismes, i la seva naturalesa és funció de la naturalesa del material que es degrada o s’assimila metabòlicament. Tots aquests compostos s’han de transportar dissolts o en suspensió, des de les zones on es produeixen fins a les àrees on s’eliminen, i els mecanismes d’excreció depenen de la quantitat d’aigua disponible per als organismes. D’altra banda, la dieta és un altre factor que determina la naturalesa dels productes excretats. L’excreció també es pot alterar per diferents processos fisiològics inherents al cicle biològic de les espècies.
Els productes excretats principals són: el diòxid de carboni i l’aigua, productes finals del metabolisme oxidatiu, i els compostos nitrogenats, dels quals destaquen la urea, l’amoníac i l’àcid úric. També s’eliminen altres substàncies, com són àcids orgànics, derivats d’àcids nucleics, aminoàcids, sals, etc.
L’excreció dels productes nitrogenats únicament es fa en forma d’amoníac en les espècies que disposen d’una gran quantitat d’aigua, perquè l’amoníac resulta molt tòxic per al propi organisme, si no és molt diluït. Això fa que aquest tipus d’excreció sigui característic dels crustacis. L’excreció dels productes nitrogenats en forma d’àcid úric es relaciona amb els organismes de vida terrestre, i és característic dels insectes i els miriàpodes; l’àcid úric és molt poc soluble a l’aigua i no excessivament tòxic en les concentracions en les quals es pot trobar dissolt. Els aràcnids, tot i ésser de vida terrestre, eliminen els productes nitrogenats en forma de guanina, un derivat dels àcids nucleics que en d’altres grups experimenten un grau més alt de degradació. Finalment, la urea, producte molt soluble a l’aigua i no tan tòxic com l’amoníac, és excretada pels organismes que disposen d’una quantitat d’aigua moderada; generalment l’eliminació de productes nitrogenats en forma d’urea es combina amb l’excreció d’amoníac o d’àcid úric, i sempre es fa en quantitats proporcionalment petites.
L’eliminació de l’aigua es regula indirectament i en funció dels productes excretats, especialment els de naturalesa nitrogenada, de manera que els òrgans excretors (especialment els tubs de Malpighi), participen en la regulació iònica i osmòtica del medi intern dels organismes.
L’aparell reproductor i la reproducció
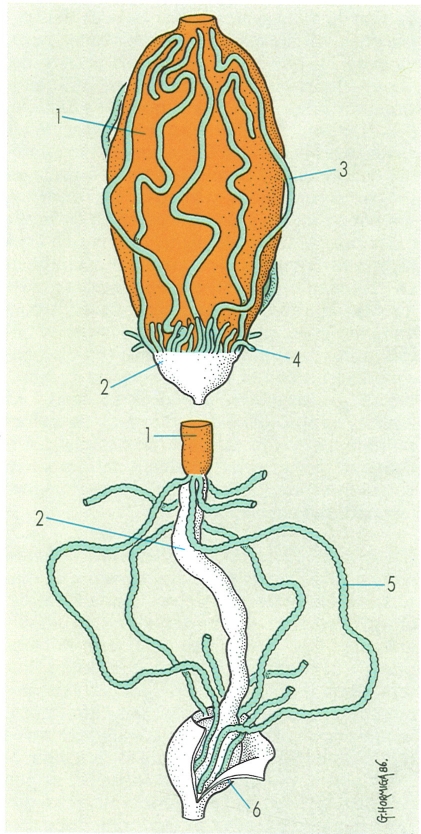
Models diferents d’aparells reproductors masculins (a dalt) i femenins (a baix), presents en els artròpodes: A model amb les gònades metamèriques, corresponent als escorpins, B model amb les gònades imparelles, corresponent als quilòpodes, C model amb les gònades parelles, corresponent als insectes i el més generalitzat en el conjunt dels artròpodes. Hom hi distingeix: 1 testicles, 2 espermiducte, 3 vesícula seminal, 4 receptacle seminal, 5 glàndula cilíndrica, 6 òrgan paraxial, 7 atri genital, 8 glàndules dorsals, 9 glàndules ventrals, 10 canal ejaculador, 11 glàndules annexes, 12 ovaris, 13 oviductes, 14 receptacle seminal, 15 atri genital, 16 orifici genital, 17 glàndula dorsal, 18 glàndula ventral, 19 espermateca o receptacle seminal, 20 glàndula de l’espermateca, 21 glàndules annexes.
Gustavo Hormiga, a partir de fonts diverses
L’aparell reproductor dels artròpodes deriva, en part, de les cavitats celomàtiques que apareixen a la fase embrionària. Per això, en alguns grups (picnogònids, escorpins, diplurs, entre d’altres), les gònades (ovaris i testicles) conserven la metameria primitiva. A la majoria dels artròpodes, però, l’aparell reproductor és format per un únic parell de gònades, que a vegades es fusiona en una de sola. Aquest és el cas més general entre els diplòpodes, quilòpodes i pseudoscorpins, entre altres.
Els conductes genitals (oviductes i espermiductes) també tenen origen metamèric, però aquesta disposició únicament es manté en els picnogònids, als quals cada gònada té el seu propi conducte genital que desemboca a l’exterior independentment dels altres. A la majoria dels artròpodes hi ha únicament dos conductes genitals, que desemboquen independentment a l’exterior (crustacis, diplòpodes, certs insectes) o bé conflueixen en un conducte únic, que desemboca en un únic porus genital (sínfïls, quilòpodes, gairebé tots els insectes i aràcnids); en aquests casos, la part terminal de l’aparell genital (el conducte imparell) és d’origen ectodèrmic. Els gonoductes poden presentar diferenciacions diverses al llarg del seu recorregut. En general hi ha glàndules annexes, de naturalesa i funció variades, que participen a la formació dels espermatòfors, les ooteques, etc.
Els orificis genitals o gonopors se situen a diferents punts del cos segons els grups: al límit entre el prosoma i l’opistosoma en els aràcnids i els meròstoms, al límit entre el perèion i el plèon en els crustacis, a la part anterior del cos en els artròpodes pregoneats (pauròpodes, sínfils, diplòpodes i alguns quilòpodes) i al seu extrem terminal en els opistogoneats (certs quilòpodes i els insectes). En els picnogònids hi ha nombrosos parells d’orificis genitals, que desemboquen a la base de les potes. Els orificis múltiples per a la còpula i la posta, presents a les femelles d’alguns insectes, també representen restes de la primitiva metamerització de l’aparell genital. En relació amb els orificis genitals, moltes vegades hi ha estructures relacionades amb la reproducció, de nombre i funció molt diversa.
Els apèndixs relacionats amb la reproducció són d’origen i morfologia diversos. En els mascles s’anomenen òrgans copuladors i es poden haver originat a partir d’expansions i evaginacions dels marges de l’orifici o l’atri genital, a partir d’apèndixs especialitzats relacionats amb l’orifici genital (a la majoria dels insectes) o bé a partir d’apèndixs modificats, no relacionats amb l’orifici genital (als aranèids, els diplòpodes, alguns insectes). En uns casos serveixen per a introduir els espermatozoides a les vies genitals de la femella, tot actuant com a veritables òrgans copuladors, però també poden actuar indirectament, dipositant l’esperma al substrat, o simplement subjectant la femella durant la còpula. En les femelles, els apèndixs genitals estan relacionats amb l’orifici genital; moltes vegades constitueixen un òrgan especialitzat, per a afavorir la posta, que rep noms diferents segons els grups (oviscapte, ovopositor, etc.). En alguns grups, les femelles porten apèndixs o estructures especialitzades per a transportar i protegir els ous fins al naixement de les larves o els juvenils, i en d’altres, aquests transport i protecció continuen després de l’eclosió dels ous, fins que els petits es poden valer per ells mateixos.
Els artròpodes, en general, són gonocòrics, és a dir, unisexuats, i moltes vegades mostren un dimorfisme sexual acusat. Hi ha, però, alguns casos d’hermafroditisme, principalment entre els crustacis, i més excepcionalment entre els insectes i els àcars (aràcnids). La reproducció és fonamentalment amfigònica: requereix la fecundació dels gàmetes (espermatozoides i òvuls) produïts pel mascle i la femella. Hi ha, també, nombrosos casos de partenogènesi, en diferents modalitats, especialment entre els insectes, però també en els crustacis, els miriàpodes i els aràcnids.
La fecundació és, generalment, interna, però en els grups aquàtics és majoritàriament externa. Entre els quelicerats, els merostomats actuals (xifosurs) tenen fecundació externa; la femella diposita els òvuls en forats fets a la sorra de la platja i posteriorment el mascle hi deixa els espermatozoides al damunt. En els picnogònids, hi ha una pseudocòpula: el mascle i la femella fan coincidir els seus orificis genitals, i després la femella expulsa els òvuls i el mascle hi diposita els espermatozoides al damunt; posteriorment és el mascle qui recull la posta i se la carrega als ovígers, on la portarà fins a l’eclosió dels ous. En alguns crustacis, els mascles no tenen òrgans copuladors, i la fecundació dels ous es fa per aproximació dels orificis genitals del mascle i la femella. Entre els crustacis proveïts d’òrgans copuladors la fecundació pot ser tant interna com externa. El cranc de riu, per exemple, té fecundació externa, perquè a l’acte de la còpula el mascle no fa servir els òrgans copuladors per a introduir els espermatozoides a les vies genitals de la femella, sinó per a deixar-los adherits a la seva regió esternal del perèion; posteriorment, la femella expulsa els òvuls, i amb ells una substància capaç de fer desprendre els espermatozoides.

La transferència directa de l’esperma del mascle a les vies genitals de la femella implica l’existència d’uns òrgans copuladors especials i, sovint, també, de formacions particulars del mascle destinades a retenir la femella durant la còpula, si bé això es troba igualment en organismes als quals la transferència de l’esperma és indirecta. A la fotografia s’il·lustra l’aparellament de dues típules (Tipula maxima).
Enric Curto.
La fecundació interna, característica dels grups terrestres (aràcnids, miriàpodes i insectes) pot implicar una transferència de l’esperma o els espermatozoides directa o bé indirecta, i una gran varietat de modalitats. En els casos més simples de transferència indirecta, el mascle diposita gotes d’esperma o espermatòfors al substrat, siguin lliures o bé subjectes a diferents estructures (fils, peduncles, etc.), i posteriorment, la femella, sigui d’una manera accidental o bé incitada pel mascle, se’ls introdueix a les vies genitals, on es fa la fecundació. De vegades és el mateix mascle el que recull l’espermatòfor i l’introdueix al gonoducte femení, per mitjà d’alguns apèndixs (les antenes, les peces bucals, les potes, etc.). Aquestes i d’altres modalitats són pròpies de nombrosos aràcnids, molts miriàpodes i insectes apterigots, etc. En d’altres casos, el mascle no diposita l’esperma en el substrat, sinó que se la transfereix a òrgans copuladors especialitzats que no estan en relació directa amb l’aparell genital masculí. Aquest és un cas freqüent en els aràcnids, els quals utilitzen els pedipalps com a òrgans copuladors, i amb ells recullen l’esperma i la introdueixen a les vies genitals femenines durant la còpula; aquest tipus de fecundació s’ha observat també entre els diplòpodes i alguns insectes alats, com les libèl·lules (odonats). En general, la fecundació interna indirecta, sigui amb apèndixs especialitzats o sense, és característica dels aràcnids, els miriàpodes i certs insectes (especialment els apterigots). Hi ha, també, el cas d’alguns decàpodes especialitzats, en els quals els mascles transfereixen l’esperma a òrgans copuladors separats dels orificis genitals, i amb aquests la introdueixen a l’aparell genital de la femella. La fecundació interna directa és característica de la majoria dels insectes pterigots (alats). Es caracteritza perquè els òrgans copuladors dels mascles estan en relació directa amb el seu aparell genital. Després del festeig, que pot ser més o menys sofisticat, els mascles introdueixen, durant la còpula, l’esperma o els espermatòfors a les vies genitals de la femella, on es produeix la fecundació.
En general els artròpodes són ovípars, però hi ha alguns casos d’ovoviviparisme i de viviparisme. En els ovípars hi ha casos que les femelles deixen els ous, aïllats o en grups, sense cap protecció, directament damunt o molt a prop d’allò que serà la font d’aliment dels petits. Moltes vegades, però, les femelles dipositen els ous a l’interior d’estructures o nius que fabriquen prèviament. Entre les estructures protectores de la posta secretades per glàndules annexes de l’aparell genital de la femella, són característiques les ooteques de pregadéus i paneroles, els capolls de seda, etc. En altres casos, les femelles tenen cura de la posta fins al moment de l’eclosió dels ous, i fins i tot n’hi ha que protegeixen i vigilen els descendents fins que assoleixen un estat avançat de desenvolupament. En alguns grups, principalment dels crustacis, les femelles porten dispositius especials per a transportar la posta al damunt del seu cos. En casos excepcionals, com el dels picnogònids, són els mascles els que s’ocupen de la posta.
En els casos d’ovoviviparisme, les femelles incuben els ous dins del cos. Tot i que els embrions reben aliments de la mare, no hi ha estructures especials que els posin en relació amb l’úter matern. Es comporten com a ovovivípars molts aràcnids (escorpins, pseudoscorpins, alguns àcars, etc.) i alguns casos aïllats dins d’altres grups.
El viviparisme, si bé no és un fenomen freqüent, es troba força estès entre alguns grups d’insectes (homòpters, dípters, dermàpters, entre d’altres). En aquest cas, els embrions es relacionen amb l’úter matern per mitjà d’una mena de placenta, que pot ser difosa o ben delimitada, a través de la qual reben l’aliment de la mare.
El desenvolupament embrionari i postembrionari
Els ous
Diferents tipus de posta d’artròpodes. En el cas de les aranyes, la massa d’ous sol quedar recoberta de fils de protecció, que li donen un aspecte característic; a la fotografia (posta d’Aculopeira ceropegia, a dalt) aquests fils han estat parcialment eliminats per a fer visibles els ous. En un altre cas, el dels crustacis decàpodes (al mig), la femella guarda els ous al final del cos, entre els apèndixs abdominals fins al moment de l’eclosió, cosa que és només un dels exemples de les múltiples especialitzacions en aquest sentit. D’altra banda, és corrent que les femelles dels insectes (a baix) deixin la posta damunt d’un substrat capaç de proveir d’aliment les larves que neixen dels ous.
Jordi Berthold, Enric Curto i Josep M. Moraleja.
Els ous són de morfologia i dimensions molt variables. El cas més general és que siguin centrolecítics: el zigot, resultat de la fusió de l’òvul i l’espermatozoide, se situa al centre de l’ou, envoltat d’una gran quantitat de vitel·lus. Aquest tipus d’ou és característic de les espècies ovípares. En alguns casos, però, hi ha ous pobres en vitel·lus, especialment en formes paràsites i vivípares. Tant en un cas com a l’altre, els ous van revestits d’una o més membranes, una de les quals s’anomena còrion.
La segmentació dels ous es fa d’una manera o d’una altra segons la quantitat i la distribució de vitel·lus al seu interior. Els ous pobres en vitel·lus presenten una segmentació espiral i total, més o menys típica, mentre que als ous centrolecítics, rics en vitel·lus, la segmentació és parcial i superficial. En alguns casos, la segmentació és total a les primeres fases, i es va transformant d’una manera progressiva en superficial i parcial. També s’ha observat un altre tipus de segmentació, la discoidal, en els ous d’alguns escorpins, la qual és una modificació de la segmentació superficial.
La formació i el desenvolupament de l’embrió
El desenvolupament embrionari s’inicia, als ous centrolecítics, amb mitosis successives del nucli; no es formen, però, les membranes corresponents i el vitel·lus es manté indivís. A mesura que es van succeint les divisions, els nuclis van emigrant gradualment cap a la perifèria, on es formen les membranes cel·lulars, que no es prolonguen cap a l’interior del vitel·lus; les cèl·lules així formades, se situen, doncs, al voltant del vitel·lus, constituint la fase anomenada de periblàstula. El desenvolupament continua amb la formació del disc germinatiu (o estria germinativa) a partir d’un determinat grup de blastòmers, és a dir, a partir d’una zona determinada de la periblàstula. Aquesta zona constitueix l’àrea embrionària, que es diferencia per sobre el vitel·lus, per engruiximent i diferenciació dels blastòmers que la componen. Posteriorment es produeix una proliferació cel·lular i es diferencien els fulls blastodèrmics més interns: l’endoderma i el mesoderma. A continuació, l’estria germinativa que origina l’embrió es metameritza i, per excavació al mesoderma, es formen les vesícules celomàtiques, que més endavant desapareixeran. L’estria germinativa primer, i l’embrió després, envolten externament el vitel·lus, mentre aquest últim el va consumint. Quan el nou organisme s’alimenta a expenses del vitel·lus de l’ou, o dels productes elaborats per la mare en els casos d’ovoviviparisme i viviparisme, s’anomena embrió.
Els estadis larvaris

El desenvolupament postembrionari, dins d’un grup tan extens com el dels artròpodes, ateny una àmplia gamma de modalitats, des de casos senzills, als quals les cries neixen gairebé amb el mateix aspecte que els adults, fins a la coneguda complexitat dels cicles de molts insectes, als quals les cries passen per diversos estadis de morfologia absolutament diferent a la de l’adult. La taràntula de la fotografia (Lycosa fasciventris), que carrega les cries al dors, il·lustra el primer tipus, el de desenvolupament directe, típic de pràcticament tots els aràcnids. La de baix, d’una eruga de papallona (Papilio machaon), netament diferent de l’adult, correspon al tipus de desenvolupament complex conegut amb el nom de metamorfosi.
Xavier Perellada i Enric Curto.
El desenvolupament postembrionari comprèn tots els passos de desenvolupament que experimenta l’organisme des que emergeix de l’ou fins que es converteix en un individu adult. A la majoria dels artròpodes, els organismes acabats de sortir de l’ou són força diferents dels adults, i reben el nom de larves, nimfes, juvenils, etc., segons la fase de desenvolupament en què es troben i segons els diferents grups. La transformació de larva a adult es pot fer d’una manera progressiva, per l’adquisició successiva dels diferents caràcters d’aquest, o bé a través de canvis bruscs i profunds, que en conjunt es coneixen amb el nom de metamorfosi.
L’aspecte de les larves dels artròpodes és molt variat: cada grup té un o diferents estadis larvaris típics, que reben noms especials. El control de la metamorfosi i del desenvolupament postembrionari en general es fa per hormones, produïdes per glàndules endocrines; aquestes són regulades per neurohormones, produïdes per cèl·lules neurosecretores. La metamorfosi assoleix la seva màxima complexitat en els insectes holometàbols, als quals comporta fenòmens d’històlisi (destrucció dels òrgans i teixits larvaris), histogènesi (formació d’òrgans i teixits típics de l’adult a partir d’uns grups de cèl·lules indiferenciades a les fases larvàries: els discs imaginals) i de conservació de teixits i òrgans larvaris modificats. En alguns casos, els individus que emergeixen de l’ou difereixen únicament de l’adult per les seves dimensions i l’absència dels òrgans genitals; en aquest cas el desenvolupament es considera directe, bé que hi ha diferents estadis juvenils abans d’assolir la maduresa sexual.
En els artròpodes és característic que les successives larves o formes larvàries tinguin el mateix aspecte i tipus de vida, encara que poden no ser iguals que els dels adults. En alguns casos, però, les larves que se succeeixen tenen característiques diferents. En aquest cas es parla d’hipermetamorfosi. La hipermetamorfosi sol relacionar-se amb organismes de vida parasitària, cas que es dóna en alguns crustacis (certs copèpodes i isòpodes paràsits), aràcnids (determinats àcars paràsits), i en molts insectes, com els coleòpters (meloèids, alguns estafilínids i caràbids), estresípters, diversos dípters (bombílids, círtids, nemèstrids, etc.) i nombrosos himenòpters entomòfags. Aquest tipus de desenvolupament es caracteritza per l’existència d’una successió de larves amb característiques morfològiques, fisiològiques i ecològiques diferents, que poden passar de fer una vida lliure i mòbil a, per exemple, un estadi larvari immòbil i paràsit.
Independentment dels tipus larvaris, hi ha dos tipus bàsics de desenvolupament: anamorf i epimorf. En el desenvolupament anamorf, la larva que emergeix de l’ou té el cos dividit en un nombre de metàmers inferior al característic de l’adult i l’adquisició del nombre definitiu es va fent al llarg del desenvolupament postembrionari; aquest és el tipus de desenvolupament dels trilobits, els meròstoms, els picnogònids, els crustacis, els miriàpodes progoneats (símfils, diplòpodes i pauròpodes), els quilòpodes anamorfs (els litobiomorfs i els escutigeromorfs) i alguns insectes (els proturs). En el tipus de desenvolupament epimorf, característic dels aràcnids, alguns quilòpodes (geofilomorfs i escolopendromorfs) i la majoria dels insectes, la larva que emergeix de l’ou ja té el nombre de metàmers definitiu; pot ser, però, que l’aspecte de la larva sigui molt diferent del de l’adult, com passa en els insectes holometàbols, és a dir, tots aquells que tenen metamorfosi complicada.
Els artròpodes, independentment del tipus de cicle biològic, poden presentar el fenomen de la diapausa, és a dir, una disminució temporal del metabolisme que permet de mantenir un estat de vida latent o no activa durant un cert temps. La diapausa sol ésser desencadenada per factors ambientals desfavorables, com per exemple temperatures extremes, manca de recursos alimentaris, durada inadequada del fotoperíode, etc., però és controlada hormonalmente. En algunes espècies, la diapausa és un fenomen obligat, mentre que en altres és facultatiu. En conjunt representa un procés clarament adaptatiu: quan les condicions ambientals es tornen favorables, es reprèn la vida activa a partir de la fase estacionària. És freqüent que es produeixi en la fase d’ou, cas en què s’interrompen els processos de segmentació, embriogènesi i organogènesi; però també pot produir-se diapausa en altres moments del cicle vital, en estat de larva, prenimfa o nimfa, amb la interrupció consegüent del desenvolupament postembrionari, o fins i tot en estat d’adult.