No hi ha dubte que, de tots els grups d’artròpodes, el dels insectes és el més conegut i, alhora, el més diversificat i el més important numèricament de tot el regne animal. Podem afirmar que són organismes d’origen antic, ja que els primers fòssils d’insectes que es coneixen daten del Devonià, bé que fou a partir del Permià que començaren a adquirir importància numèrica, gràcies a la regressió dels oceans i a la diversificació de les plantes. Aquestes formes fòssils s’assemblaven ja a les formes actuals, cosa que no permet d’establir d’una manera clara l’evolució dels seus diferents grups. Actualment hom pot dir que els insectes dominen la Terra ja que hi ha prop d’un milió d’espècies conegudes, xifra que probablement encara és molt inferior a la real.
Malgrat la diversitat dels insectes, en conjunt conformen un dels grups en el qual es manté més estable la tagmatització del cos i, per tant, el seu aspecte general. D’altra banda, cadascuna de les regions del cos és constituïda sempre per un nombre fix de segments, fenomen que no es manifesta en la resta de grups d’artròpodes mandibulats.
Dins dels insectes trobem els invertebrats més vistents i acolorits, i també d’altres de tons foscos que poden passar completament desapercebuts. Malgrat la constància del nombre de tagmes del cos, el seu aspecte és molt variable: els uns són fràgils i delicats, mentre que els altres són d’aparença robusta i basta, i d’altres encara mostren una àmplia gamma de característiques que els situen entre uns i altres. Sovint es presenta, a més, un diformisme sexual acusat, que es manifesta en la forma, la mida i els colors del cos. La mida ateny una gran diversitat: oscil·la entre menys d’un mil·límetre (0,25 mm, en el cas d’alguns ptílids i alguns himenòpters paràsits) i 28 cm (en el cas de certes espècies de papallones tropicals), entre les espècies actuals; pel que fa referència als fòssils, inclouen l’espècie gegant dels insectes (una libèl·lula, Meganeura monyi, que feia 70 cm d’envergadura alar); de manera que hi ha insectes més petits que certs protozous i d’altres més grans que alguns cordats.
La gran importància numèrica dels insectes es deu, en part, a la seva gran facilitat de dispersió, atès que són els únics invertebrats capaços de volar (malgrat l’existència d’algunes formes sense ales); d’altra banda, alguns es dispersen d’una manera passiva, tot aprofitant els corrents d’aire, l’aigua o el desplaçament d’altres organismes (forèsia). En general, combinen la locomoció, la natació, el salt, etc. amb el vol, cosa que encara els proporciona més avantatges en la dispersió. Alguns fan migracions d’una manera regular.
Els insectes són molt versàtils i són capaços d’aprofitar nínxols ecològics i aliment que refusen d’altres invertebrats o, fins i tot, els cordats. Són capaços d’utilitzar com a aliments substàncies tan insòlites com el petroli, la fusta, banyes de vertebrats, pells, espècies, tabac, etc., i substàncies utilitzades també per altres animals, com excrements, cadàvers, matèria orgànica en descomposició, etc.; d’altres són depredadors, fitòfags o paràsits. Hi ha espècies que poden aguantar temperatures extremes, molt fredes (d’uns quants graus sota zero, als pols) o molt altes (a les aigües termals o als deserts). La facilitat de dispersió i la versatilitat ecològica són, al costat d’una elevada capacitat reproductora, els factors fonamentals que han determinat la seva importància actual.
Els insectes tenen també importància econòmica, ja que mantenen estretes relacions amb l’activitat de l’home i el seu entorn. Hi ha molts insectes que són beneficiosos per a l’home perquè actuen com a pol·linitzadors, són depredadors d’altres organismes nocius o perjudicials per a l’home o per als seus conreus o el bestiar, o bé perquè li proporcionen substàncies útils, com ara cera, mel, seda, laca, etc. Intervenen d’una manera decisiva en la formació del sòl i en la degradació de la matèria orgànica (cadàvers, excrements, etc.), i serveixen d’aliment a ocells, peixos i d’altres animals insectívors, útils a l’home. Hi ha, també, insectes que perjudiquen l’home, sigui perquè són causa de greus flagells per als conreus o les masses forestals, sigui perquè són paràsits de determinats animals o de l’home mateix —al qual poden produir molèsties o, fins i tot, poden transmetre malalties—, o sigui, també, perquè ataquen productes emmagatzemats (llegums, llavors, fruita, tabac, etc.) que l’home utilitza com a aliments o com a productes de comerç.
La distribució dels insectes comprèn tot el planeta i els biòtops més diversos. Moltes espècies són cosmopolites, especialment les antropòfiles, és a dir, les que viuen en relació amb l’home, ja sigui en els seus habitatges o bé en els productes de comerç que són transportats d’un lloc a l’altre. L’home trasllada voluntàriament les espècies que li són profitoses, especialment quan es tracta d’espècies que controlen les poblacions d’altres organismes que provoquen flagells, i fins i tot les cria en condicions de laboratori per tal d’alliberaries després. Entre els insectes també es dona el cas contrari: espècies de distribució molt restringida que constitueixen endemismes locals. De fet, la distribució actual dels insectes correspon als vestigis de llur distribució primitiva, de manera que l’estudi d’aquestes àrees ens dona informació sobre els canvis ocorreguts en els mars i els continents, alhora que ens permet de valorar la importància dels canvis climàtics esdevinguts al llarg dels diferents períodes.
Característiques morfològiques
Els insectes es poden caracteritzar fàcilment pel fet de presentar el cos dividit en tres regions ben diferenciades: cap, tòrax i abdomen. Al cap hi ha un parell d’ulls compostos i, de vegades, també ocel·les, a més d’un parell d’antenes, i els apèndixs bucals. El tòrax és constituït sempre per tres metàmers, cadascun amb un parell d’apèndixs o potes; en el tòrax s’articulen també les ales (dos parells), corresponents als dos darrers metàmers, però pot haver-n’hi un sol parell o cap. L’abdomen, constituït per onze metàmers, no porta apèndixs pregenitals (excepte en els apterígots) però té apèndixs relacionats amb la reproducció (genitàlia o terminàlia) i, de vegades, cercs terminals. També caracteritza els insectes la respiració de tipus traqueal, l’excreció per tubs de Malpighi (tret del cas d’alguns apterigots, que tenen òrgans excretors metamèrics) i la posició de l’orifici genital a la part posterior del cos (son opistogoneats). La resta dels caràcters d’anatomia interna són semblants als generals de tots els artròpodes. En general són dioics, i en molts casos els mascles i les femelles tenen caràcters externs que els diferencien; algunes espècies són hermafrodites. El tipus de reproducció més general és la sexual, però hi ha alguns casos de partenogènesi. Són ovípars, però alguns poden ser ovovivípars o, fins i tot, vivípars. El desenvolupament postembrionari és indirecte, ja que es fa a través de diverses formes juvenils o larvals que precedeixen l’estat d’adult; en la majoria dels casos, el pas d’un estadi larval al següent comporta canvis importants, coneguts en conjunt amb el nom de metamorfosi.
Ultra les consideracions específiques que es fan tot seguit, vegeu també les genèriques de qualsevol artròpode.
Les regions del cos
Tal com ja hem dit, hom sol diferenciar tres regions en el cos dels insectes: cap, tòrax i abdomen.
El cap
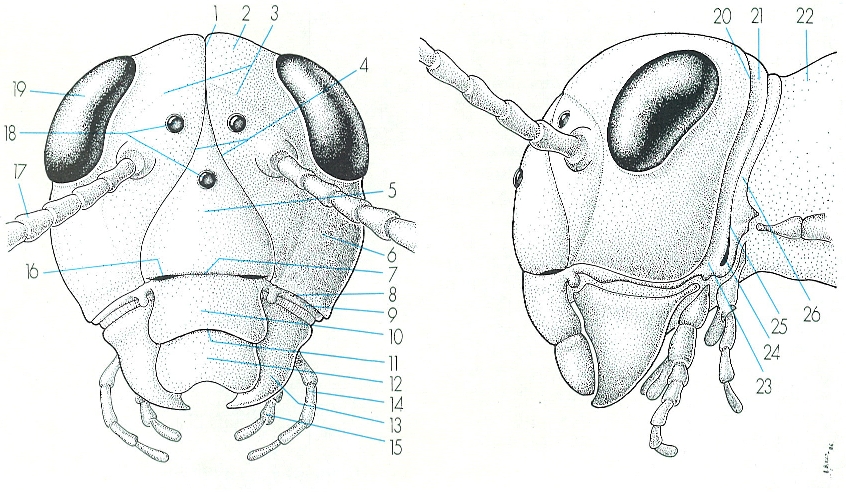
Estructura externa del cap d’un insecte ectògnat i posició de les sutures (i les esclerites que delimiten), dels ulls i dels apèndixs (antenes i peces bucals, en aquest cas mastegadores). Hom hi consigna: 1 sutura coronal, 2 vèrtex, 3 sutura postfrontal, 4 sutura frontal (que forma, juntament amb la sutura coronal, l’anomenada sutura epicranial), 5 front, 6 gena, 7 sutura frontoclipeal, 8 sutura subgenal, 9 subgena, 10 clipi, 11 sutura clipeolabral, 12 labre, 13 mandíbula, 14 maxilles, 15 llavi, 16 fosseta pretentorial, 17 antena, 18 ocelles, 19 ull compost, 20 sutura occipital, 21 occipuci, 22 cèrvix o coll, 23 postgena, 24 fosseta posttentorial, 25 sutura postoccipital, 26 postoccipuci.
Amadeu Blasco, original de Snodgrass.
Generalment es considera que el cap dels insectes és format per sis metàmers, a més de l’àcron, que es troben soldats com a resultat del procés de cefalització i que, en conjunt, formen la càpsula cefàlica, endurida i solcada per unes estries en posició característica. Aquests solcs, anomenats sutures, no responen a la metamerització primitiva (és a dir, als punts d’unió dels sis metàmers que formen el cap) sinó que són engruiximents interns de la cutícula. En conjunt, les sutures delimiten al cap una sèrie d’esclerites, regions o àrees diferents, que solen ésser força constants en el model primitiu de cap d’insecte i que corresponen als tipus ortògnat o hipògnat.
A la superfície dorsal del cap sol haver-hi una sutura en forma de lletra Y, anomenada epicranial, la branca imparella de la qual constitueix la sutura coronal, mentre que les branques parelles conformen la sutura frontal; en conjunt, la sutura epicraneal permet distingir dues zones al cap: una zona anterior o frontal, i una altra superior, la posterodorsal i lateral.
La zona anterior o frontal sol ésser limitada lateralment per la sutura frontogenal, i s’hi distingeixen diverses esclerites separades per sutures: el front, el clipi i el labre (aquest també anomenat llavi superior, perquè protegeix les peces bucals, tot i no ésser un apèndix), bé que pot haver-hi variacions, ja que secundàriament poden desaparèixer algunes de les sutures o aparèixer-ne de noves. La superfície interna del labre i del clipi és, en una gran part, membranosa, i constitueix l’epifaringe o paladar de la cavitat preoral; sol presentar lòbuls sortints, esclerificacions, ornaments cuticulars i sensílies, de les quals són especialment importants les esclerificacions anomenades tormes, de desenvolupament molt desigual en els diferents grups.
A la regió posterodorsal i lateral del cap hi ha diverses esclerites parelles, també separades per sutures. La sutura coronal separa dues esclerites parietals, que en conjunt formen el vèrtex, per sota del qual se situen les genes. A la regió posterior del cap hi ha dues sutures, l’occipital i la postoccipital, que delimiten l’occipuci o esclerita occipital (dorsal) i les postgenes (laterals); el postoccipuci en general queda reduït a una banda estreta i contínua, que constitueix el marge posterior del cap i que envolta un forat occipital.
En general, el quadre bucal queda separat de la gena i la postgena per una banda esclerificada, la subgena, que se separa de les anteriors per una sutura (la sutura subgenal) que corre entre les invaginacions anteriors (pretentorines) i posteriors (metatentorines) del tentori; les maxil·les s’articulen a l’occipuci i el llavi al postoccipuci.
Ultra les sutures esmentades ací, poden haver-n’hi d’altres de secundàries que varien a cada grup d’insectes i que delimiten esclerites especials (antenals, oculars, mandibulars, etc.).
Segons quina sigui la direcció de l’eix principal del cap i la de les peces bucals, hi ha diferents tipus estructurals, i nombroses formes intermèdies. En el cas del cap hipògnat o ortògnat que ja hem esmentat, l’eix principal del cap és vertical i les peces bucals es dirigeixen vers el substrat, de manera que el forat occipital se situa prop del pla transversal o en ell mateix. Aquest és el tipus de cap que hom sol emprar per a la descripció general del cap dels insectes, i és el característic de molts insectes fitòfags o depredadors, que s’ajuden de les potes per a subjectar l’aliment (com els ortòpters, els màntids, els neuròpters adults, etc.). En el tipus de cap prògnat l’eix principal és horitzontal o lleugerament inclinat cap avall, mentre que les peces bucals es dirigeixen endavant i moltes vegades el forat occipital queda inclinat respecte de la posició transversal, o bé conserva aquesta posició gràcies a l’allargament de la regió ventral del cap, fenomen que es deu a l’aparició d’unes esclerites especials: la gula o el pont epistomial; aquest és el tipus de cap característic dels coleòpters i de moltes larves. En el cas del cap opistògnat, l’eix principal és horitzontal: la regió apical es dirigeix enrere i les peces bucals s’insereixen ventralment i també dirigides enrere; aquest tipus de cap és característic, sobretot, dels insectes proveïts de peces bucals picadores (heteròpters, homòpters, dípters, etc.).
A la part distal del cap hi ha un constrenyiment més o menys marcat, que delimita el coll, és a dir, la zona on el cap s’articula al protòrax. Aquest coll o cèrvix és una zona flexible membranosa on es diferencien esclerites constituïdes per plaques parelles laterals i ventrals. Pel que sembla, aquesta regió intermèdia entre el cap i el tòrax deriva en part del protòrax, i en part del metàmer labial del cap, i és més desenvolupada en els grups primitius.
En general, els insectes tenen un parell d’ulls compostos en posició lateral, que de vegades coexisteixen amb tres ocel·les dorsals. També hi ha, però, casos nombrosos d’insectes sense ulls, que en aquest cas són anomenats anoftalms.
Al cap també hi ha un parell d’antenes, molt mòbils, formades per un nombre variable d’artells articulats, que a la base es vertebren amb la regió anterolateral de la càpsula cefàlica, a prop dels ulls compostos. La seva funció és, principalment, sensorial, ja que porten una gran quantitat de tricobòtries, sensílies, pèls sensorials, etc., que actuen, sobretot, d’òrgans tàctils i olfactius, i també com a òrgans de l’equilibri.
En alguns grups, però, les antenes són adaptades per a subjectar les preses, i en els mascles també poden servir per a subjectar la femella durant la còpula. El grau de desenvolupament i la forma de les antenes és molt variable, i, com hem vist, en els mascles sovint són diferents que en les femelles; de vegades són molt reduïdes i, fins i tot, en un grup, el dels proturs, poden ésser completament absents.
L’estructura de les antenes respon a un tipus bàsic: un primer artell anomenat escapus, un segon artell anomenat pedicel·le, i tota una sèrie d’artells articulats apicals que en conjunt constitueixen el flagel. El flagel varia de forma en els diferents grups; moltes vegades hi ha una regió basal diferenciada, anomenada funicle, i una regió distal anomenada clava. En el segon artell antenal solen situar-se uns òrgans cordotonals especials, exclusius dels insectes, els òrgans de Jonhston. Segons el tipus de musculatura hom considera dos tipus diferents d’antenes: el tipus segmentat i el tipus anellat. En el tipus segmentat, cada artell té músculs propis que li permeten un moviment relatiu, i és característic dels col·lèmbols i dels diplurs. En el tipus anellat, només té músculs propis l’escapus, i per mitjà d’aquests músculs es mouen el pedicel·le i el flagel, l’últim d’una manera passiva; aquest tipus és el que presenten tota la resta d’insectes. En conjunt, tota l’antena és accionada per músculs extrínsecs elevadors i depressors.
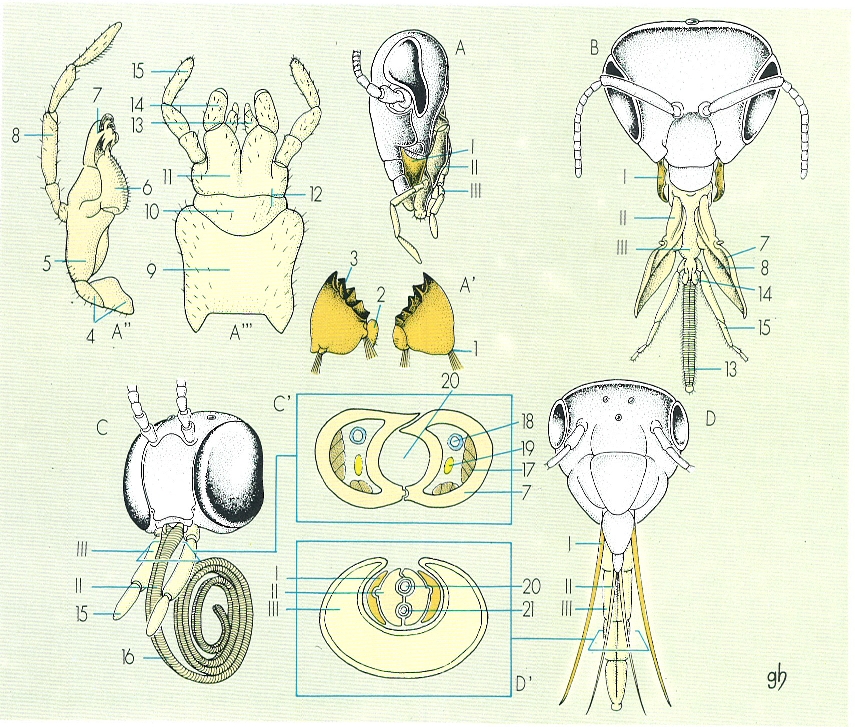
Tipus de peces, aparells bucals dels insectes. A l’aparell bucal de tipus mastegador, dibuixat en visió lateral (A), hom hi ha representat les peces per separat: A’ mandíbules, A" maxilles, A’" llavi, en el cas d’un llagost (ortòpter). L’aparell bucal de tipus mastegadorllepador, en visió frontal (B), correspon al cas d’una abella (himenòpter). L’aparell de tipus llepador-xuclador, en visió frontal (C), correspon a una papallona (lepidòpter), i en el tall transversal (C’) s’expressa l’origen maxil·lar de l’espiritrompa, formada a partir de les galees, les quals són molt allargades i delimiten un canal suctor. En l’aparell de tipus picador-xuclador, corresponenent a una xinxa (heteròpter), s’observa la transformació de mandíbules i maxil·les en estilets (desplegats en D, per fer-los més visibles), que queden recollits dins d’una mena d’estoig format pel llavi o rostre (D’). 1 còndil articular, 2 regió molar, 3 marge incisiu, 4 card, 5 estípit, 6 lacinia, 7 galea, 8 palp maxil·lar, 9 prementó, 10 mentó, 11 submentó, 12 postmentó (mentó i submentó), 13 glossa, 14 paraglossa, 15 palp labial, 16 espiritrompa, 17 músculs, 18 tràquea, 19 nervi, 20 canal alimentari xuclador, 21 conducte salival.
Gustavo Hormiga, a partir de fonts diverses
Les peces bucals dels insectes en general són lliures i visibles externament, per la qual cosa hom diu que els insectes són ectòtrofs. No obstant això, n’hi ha alguns (els col·lèmbols, els proturs i els diplurs) que són entòtrofs, és a dir, que tenen les peces bucals, en posició de repòs, allotjades en unes cambres internes de la regió cefàlica. Les peces bucals constitueixen la part més variable del cap dels insectes i comprenen tres parells d’apèndixs: un parell de mandíbules, un parell de maxil·les i el llavi inferior, que és resultat de la fusió del segon parell de maxil·les. En relació amb aquests apèndixs hi ha el labre o llavi superior i la hipofaringe o prefaringe, que formen part de la cavitat bucal (que no són apèndixs).
El desenvolupament i la forma de les peces bucals són en relació amb el tipus d’alimentació; en alguns casos arriben a atrofiar-se i deixen de ser funcionals, com és el cas dels efemeròpters i d’alguns lepidòpters i dípters que no s’alimenten en estat adult. Hi ha tres tipus bàsics d’aparells bucals i nombrosos casos intermedis i modificacions, que comentarem a continuació.
L’aparell bucal de tipus mastegador correspon al més primitiu i més característic dels insectes que es nodreixen d’aliments sòlids els quals han de triturar abans d’introduir-los a la cavitat bucal. Hi ha una gran varietat de modificacions respecte del model principal. El model típic té el labre ben desenvolupat i esclerotitzat dorsalment, mòbil i proveït d’una musculatura potent, ja que col·labora en la subjecció dels aliments i alhora protegeix les peces bucals. Les mandíbules, el primer parell d’apèndixs bucals, són triangulars i adaptades per a tallar i triturar l’aliment, a més de tenir sovint funció defensiva. El seu grau de desenvolupament i la forma varia d’uns grups als altres; generalment tenen dos punts d’articulació amb la càpsula cefàlica (mandíbules dicondíliques), tret d’alguns insectes apterigots, que en tenen un de sol (mandíbules monocondíliques). A les mandíbules es diferencien diverses regions: el marge incisiu i la zona molar, aquesta amb crestes internes formades per denticles, amb els quals es tritura l’aliment; poden tenir també sedes especials i formacions particulars, com la prosteca, el retinacle i la dent premolar, presents a diferents grups. El primer parell de maxil·les constitueix el segon parell d’apèndixs bucals i tenen una regió basal, formada en principi per dos artells (card i estípit), i una regió distal, composta de dos lòbuls, un d’extern (la galea) i un altre d’intern (la lacínia, en general proveïda d’espines sortints al marge intern); a l’estípit s’articula el palp maxil·lar, que és format per un nombre variable d’artells. El llavi és una peça única, que és resultat de la fusió d’un segon parell de maxil·les, i per tant, correspon al tercer parell d’apèndixs bucals; és constituït per una regió basal, formada de dues peces (el postmentó i el prementó, el primer sovint dividit també en submentó i mentó), a la qual s’articulen els palps labials, formats per un nombre variable d’artells i per una regió distal, constituïda per dos parells de lòbuls (dos d’interns o glosses, i dos d’externs o paraglosses) que en conjunt formen la lígula. En relació amb les peces bucals hi ha la hipofaringe, que és una estructura semblant a una llengua, a la base de la qual s’obre el conducte salival. En alguns grups la hipofaringe és trilobulada gràcies a l’existència de dos lòbuls laterals accessoris, anomenats superllengües. La hipofaringe divideix la cavitat bucal en dues cavitats: una de més gran, superior, anomenada cibari, i una d’inferior o salivari. En alguns grups, la hipofaringe es transforma en una mena d’estilet, per l’interior del qual circula el canal salival, mentre que en d’altres casos pot actuar, fins i tot, com a òrgan picador.
L’aparell bucal de tipus mastegador-llepador es caracteritza pel fet de tenir les mandíbules desenvolupades, tot i ésser propi d’insectes que s’alimenten de substàncies líquides (nèctar, per exemple); el fet s’explica perquè no serveixen per a l’alimentació, sinó especialment per a treballar i pastar substàncies sòlides, com la cera. Les maxil·les i el llavi són allargats i transformats en una mena de trompa que serveix per a libar el nèctar. Aquest tipus d’aparell bucal és típic dels himenòpters superiors, és a dir, les abelles i les vespes, entre altres.
L’aparell bucal de tipus picador-xuclador és relacionat amb els insectes que s’alimenten de fluids, siguin d’origen animal o vegetal. Implica la modificació de determinades estructures del cap i, lògicament, de les peces bucals i la hipofaringe, que en general solen transformar-se en estilets. Les mandíbules en alguns casos arriben a desaparèixer. Aquest tipus d’aparell bucal és típic dels proturs, alguns col·lèmbols, heteròpters, homòpters, tisanòpters, anoplurs, molts dípters i els sifonàpters. A cada grup, però, l’aparell bucal té característiques especials. En els heteròpters i els homòpters, les mandíbules i les maxil·les, desproveïdes de palps, són transformades en llargs estilets que corren per un solc intern del llavi format per diferents artells i anomenat rostre. El labre o llavi superior és allargat i, en posició de repòs, cobreix per sobre les peces bucals. En certs casos (alguns còccids), els estilets són més llargs que el labre, i llavors aquests es retrauen i es cargolen dins d’una cavitat del cap que, de vegades, es continua per dins del tòrax (la crumena). D’altra banda, les maxil·les delimiten dos canals: un de dorsal, suctor, i un de ventral, salival. La musculatura de la faringe actua com una poderosa bomba aspiradora que permet injectar la saliva en els animals o plantes que fan de font d’aliment, i posteriorment aspirar-ne els líquids interns. A partir d’aquest model bàsic, hi ha tota una àmplia gamma de variacions.
L’aparell bucal de tipus llepador-xuclador és propi d’insectes que s’alimenten de líquids i té una estructura variable segons cada grup. La trompa aspiradora en aquest cas es pot formar a expenses de les maxil·les i el llavi (en els tricòpters, dípters superiors, etc.) o bé a partir exclusivament de les maxil·les (en els lepidòpters). En els lepidòpters l’aparell bucal és molt especialitzat, ja que les mandíbules i el llavi són reduïts o vestigials, i només alguns (els micropterígids) tenen encara mandíbules. El llavi generalment només hi és representat pels palps i el labre també sol ésser reduït. El complex suctor es forma, com hem dit, exclusivament a partir del primer parell de maxil·les, les quals són transformades en una trompa anomenada espiritrompa; les peces basals d’aquests apèndixs són poc desenvolupades, i les galees, exageradament llargues, queden adossades i delimiten un conducte suctor. L’espiritrompa, en posició de repòs, resta cargolada en espiral. Els lepidòpters s’alimenten de nèctar, el qual succionen a través de l’espiritrompa per mitjà d’una potent bomba aspiradora.
El tòrax
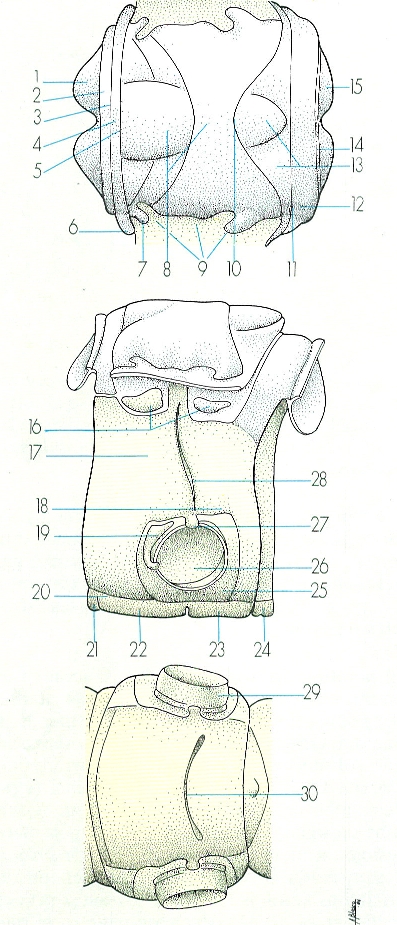
Estructura generalitzada d’un segment del tòrax dels insectes, en visió dorsal (A), que coincideix amb la tergita; visió lateral (B), en què veiem la tergita, la pleurita i l’esternita; i visió ventral (C), que coincideix amb l’esternita. Hom hi consigna: 1 fragma anterior, 2 acrotergita, 3 sutura anterocostal o antecosta II, 4 prescut, 5 sutura prescutal-escutal, 6 braç prealar, 7 esclerita subalar, 8 escut, 9 processos alars, 10 sutura escutal-escutellar, 11 sutura postnotal, 12 postnot, 13 escutel·la, 14 sutura anterocostal o antecosta III, 15 fragma posterior, 16 esclerites alars, 17 episternita, 18 sutura pleural, 19 epimerita, 20 sutura pleurosternal, 21 presternita, 22 basisternita, 23 esternella, 24 espinasternita, 25 membrana articular, 26 cavitat articular o cavitat coxal, 27 trocantí, 28 procés coxal pleural, 29 coxa, 30 sutura esternocostal.
Amadeu Blasco, original de Duporte i altres.
El tòrax és format per tres metàmers que, en principi, porten un parell de potes cadascun. La majoria dels insectes, en estat adult, tenen, a més, dos parells d’ales, que s’articulen en el mesotòrax i el metatòrax. També hi ha, però, insectes sense ales, cosa que pot representar un caràcter primitiu (cas dels apterigots) o bé un caràcter secundari (cas dels pterigots àpters, com els anoplurs, els mal·lòfags, els sifonàpters, etc.), és a dir, que les ales hagin desaparegut per reducció.
En principi, tots tres metàmers presenten un grau de desenvolupament semblant, però pot haver-hi diferències a causa de les especialitzacions. En alguns casos, la regió dorsal del protòrax (pronot) és molt més desenvolupada que en la resta de metàmers i forma com una mena d’escut (en els ortòpters, coleòpters, dictiòpters, etc.) i, en d’altres, és reduïda a una estreta banda (odonats, efemeròpters, etc.). Pel que fa al mesotòrax i al metatòrax (segments que, en conjunt, constitueixen el pterotòrax), en els insectes que tenen els dos parells d’ales semblants, aquests dos metàmers són equivalents, mentre que en els que tenen les ales anteriors més desenvolupades que les posteriors hi ha també un desenvolupament més gran del segment corresponent, és a dir, del mesotòrax, i viceversa. En alguns casos, com en els himenòpters, el primer metàmer de l’abdomen es troba incorporat al tòrax (propodeu o epinot).
Cada metàmer és format per quatre plaques o esclerites: una dorsal, anomenada tergita o not, una ventral, anomenada esternita o estèrnum i dues laterals, les pleures o pleurites.
A les formes primitives, les tergites eren constituïdes per una única esclerita, i se separaven les unes de les altres per petites esclerites intersegmentàries; són aquestes les que, per evolució, s’han associat a la tergita anterior o a la posterior i han constituït l’anomenat postnot en el primer cas i l’acrotergita en el segon; l’acrotergita queda separada de l’esclerita tergal per una sutura (anterocostal), on se situa el fragma tergal de l’endosquelet. D’altra banda, la tergita primitiva es pot subdividir en diverses esclerites: el prescut, l’escut i l’escutel·la, de desenvolupament desigual als diferents grups.
Les pleurites o pleures diferencien, en els pterigots, dues esclerites: una anterior, anomenada episternita o epistèrnum, i una de posterior, l’epimerita o epimeró; ambdues esclerites queden separades per una sutura, la sutura pleural. Aquestes esclerites poden presentar divisions secundàries i, fins i tot, en alguns casos es poden soldar totalment a les tergites.
Les esternites o estèrnum consten, en principi, d’una sola placa, i se separen per mitjà d’una esclerita intersegmentària, que s’associa a l’esternita immediata anterior. La placa esternal primitiva pot ésser subdividida en tres esclerites: la presternita, la basisternita i l’esternel·la. En general, la basisternita se separa de l’esternel·la per una invaginació cuticular, a la qual se situa la furca (endosquelet). D’altra banda, la zona equivalent a l’esternel·la és subdividida en diverses esclerites. La disposició de les esclerites esternals és diferent a cada grup d’insectes i el seu grau de desenvolupament es relaciona amb la proximitat de les coxes. A banda i banda de l’estèrnum es poden diferenciar altres esclerites especials.
L’abdomen
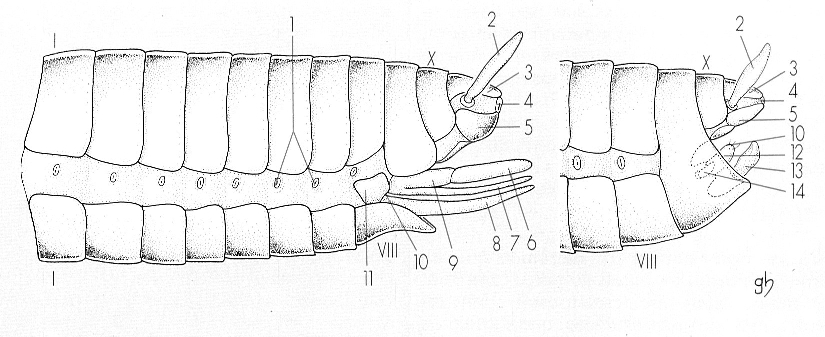
Esquema generalitzat de l’abdomen dels insectes, en una femella (A) i un mascle (B), on s’aprecia la disposició de les plaques tergal (dorsal) i esternal (ventral) de cada segment, separades per la membrana pleural, i l’estructura de la genitalia, en posició terminal al cos. Hom hi consigna: 1 estigmes respiratoris, 2 cerc, 3 epiprocte o placa supraanal, 4 anus, 5 paraprocte o placa perianal, 6 valva interna o posterior (segment IX), 7 valva dorsal o lateral (segment IX), 8 valva ventral o anterior (segment VIII), 9 valvifer (segment IX), 10 orifici genital, 11 valvifer (segment VIII), 12 edeagus, 13 valva, 14 paràmers.
Gustavo Hormiga, original de Snodgrass.
L’abdomen és constituït en la majoria dels insectes per onze metàmers més el tèlson, però en general no són tots visibles. Només els proturs i els col·lèmbols són excepció, ja que els primers tenen l’abdomen fet de dotze segments (si bé el darrer aparent correspon al tèlson, que te forma de metàmer), i els segons només de sis. L’abdomen pot unir-se al tòrax en tota la seva amplada (abdomen sèssil) o bé pot fer una mena d’estrangulament (abdomen pedunculat).
L’estructura dels metàmers abdominals és més senzilla que no la dels toràcics; tenen una tergita (dorsal) i una esternita (ventral), generalment indivises, i dues pleurites (laterals) en general membranoses, amb diverses esclerites diferenciades.
El nombre d’onze metàmers a l’abdomen es manté en els embrions i en alguns adults, però generalment aquests en tenen menys ja que hi ha un procés de reducció i modificació que sol afectar els primers i els darrers metàmers. El primer metàmer abdominal (i especialment la seva esternita) sol ésser reduït o bé vestigial; en els himenòpters apòcrits es fusiona al metatòrax i rep el nom de propodeu, epinot o segment mitjà, però, per contra, en alguns acrídids (ortòpters), aquest primer metàmer és el més desenvolupat i és el que porta els òrgans timpànics. A la regió posterior de l’abdomen el tèlson sol ésser vestigial i és representat per la membrana perianal. L’onzè metàmer també sol ésser modificat, i és representat, en els ordres inferiors, per una placa dorsal, l’epiprocte o placa supraanal, i dues peces esternals, les plaques perianals o paraproctes. D’altres vegades, com passa en molts insectes endopterigots, els darrers metàmers abdominals s’atenuen i arriben a telescopitzar-se i a formar un tub retràctil, fenomen que és manifest en alguns coleòpters, en dípters i en altres grups. La reducció d’uns metàmers i la telescopització d’uns altres fa que, de vegades, únicament restin com a metàmers aparents uns quatre o cinc dels onze originals, com és el cas d’alguns dípters. A l’abdomen, en posició terminal, se situa l’anus. L’orifici genital desemboca, generalment, entre el vuitè i el novè segments en el cas de les femelles i en el novè en el dels mascles.
La majoria dels insectes en fase embrionària porten esbossos apendiculars a tots els metàmers abdominals; en general, aquests apèndixs desapareixen de la regió pregenital durant l’ontogènia, però es mantenen, bé que molt modificats, a la regió genital (on formen part de l’anomenada genitàlia o terminàlia) i a la part terminal de l’abdomen, on formen els cercs, de funció sensorial o defensiva. Els insectes adults pterigots no tenen apèndixs pregenitals, mentre que els apterigots en porten a tots o alguns dels metàmers corresponents: els col·lèmbols en porten tres parells, molt típics, adaptats per a saltar, i tota la resta (els proturs, els diplurs, els microcorifis i els zigentomes) tenen les plaques coxals més o menys desenvolupades, amb estils i vesícules coxals semblants a les dels símfils (miriàpodes), en nombre i estructura variable segons els grups. En les formes larvals de pterigots, hom considera com a apèndixs abdominals les traqueobrànquies de les espècies aquàtiques i els propodis o pseudopodis de les erugues dels lepidòpters, himenòpters i altres. A la regió posterior de l’abdomen, al segment onzè, de vegades hi ha cercs, de forma i dimensions variables: llargs i multiarticulats o curts i formats de pocs artells o d’un de sol; els microcorifis, els zigentomes i els efemeròpters, a més dels ceres parells tenen un filament imparell, central, anomenat paracerc.
La regió genital és formada, a la majoria dels insectes, pels metàmers vuitè i novè, ja que és en aquests on es localitzen els orificis genitals i també els apèndixs modificats relacionats amb la reproducció. La genitàlia femenina consta típicament de tres parells de valves, que en conjunt constitueixen l’ovipositor o òrgan de posta, el grau de desenvolupament i de coadaptació del qual varia d’uns grups als altres, i fins i tot pot mancar; en alguns casos, l’ovipositor és modificat per a perforar i picar, i es relaciona amb glàndules de verí (en alguns himenòpters). El model general d’ovipositor consta d’un parell de valves centrals o anteriors que deriven del vuitè metàmer, un parell de valves dorsals o laterals, i un parell de valves internes o posteriors, que deriven del novè metàmer. Aquest tipus d’ovipositor és el típic dels microcorifis, els zigentomes i molts pterigots. La genitàlia masculina o edeagus deriva, almenys en part, del novè metàmer i presenta una gran diversitat de formes en el conjunt dels insectes, per la qual cosa es fa difícil d’homologar les peces que l’integren. D’altra banda, la seva estructura té un gran interès a nivell específic i genèric. Els mascles dels zigentomes i microcorifis presenten la genitàlia semblant a la de les femelles, però en difereixen pel complex fàl·lic. En els mascles dels pterigots, malgrat que hi ha diferents tipus de genitàlia, hi ha sempre uns òrgans per a la transferència de l’esperma que es troben en relació amb el canal ejaculador, i diferents peces per a subjectar la femella durant la còpula. Totes aquestes formacions acostumen a trobar-se retretes dins de la cambra genital, reforçada per esclerites.
Els òrgans de desplaçament: apèndixs i ales
Les potes s’articulen en posició ventral, entre les pleures i l’estèrnum a cadascun dels metàmers toràcics. El seu grau de desenvolupament i la forma es relacionen amb el tipus de locomoció (marxa, salt, natació, etc.), però sempre responen a un mateix pla estructural. A més d’actuar com a òrgans locomotors, les potes poden servir per a capturar aliments (per exemple, les potes prènsils dels màntids o les potes recol·lectores dels himenòpters), o poden presentar diferenciacions especials per a netejar-se les antenes, per a fixar-se als hostes (en el cas d’espècies paràsites), etc. En alguns casos, les potes presenten caràcters sexuals secundaris (dilatació dels tarsos en el mascle, diferent grau de desenvolupament, etc.) i fins i tot poden reduirse per diferents processos de regressió. El moviment de les potes és molt coordinat, ja que es poden desplaçar endavant, endarrere i de costat, i donen molta estabilitat al cos gràcies al fet de recolzar a terra tot el tars, i de presentar, sovint, formacions adherents.
Les potes són apèndixs unirramis, formats per cinc artells principals: coxa, trocànter, fèmur, tíbia i tars, tots articulats per mitjà de membranes articulars. La coxa és l’artell basal i és la que, de fet, articula l’apèndix al cos. El punt on la coxa s’articula al cos (pel procés coxal de les pleures i pel trocantí, quan n’hi ha), pot presentar una estructura complicada a causa de la diferenciació d’esclerites i del desenvolupament variable de les mateixes coxes. El trocànter, en general, és poc desenvolupat i es troba íntimament unit al fèmur; de vegades és subdividit (en alguns himenòpters paràsits i en odonats). El fèmur i la tíbia són els artells més llargs de la pota i en general, la tíbia porta esperons a la seva part distal. El tars consta, primitivament, d’un sol artell, però generalment es troba subdividit, i cadascun d’aquests artells que el componen s’anomena tarsòmer; el nombre màxim de tarsòmers és de cinc. A l’àpex del tars se situa el pretars, que en els casos més senzills és representat per una ungla (en col·lèmbols, proturs, etc.), encara que en general n’hi ha dues; a més, pot presentar formacions especials, com l’aroli i el pulvil·le que, coberts de pèls glandulars, actuen com a petites ventoses, i l’empodi.
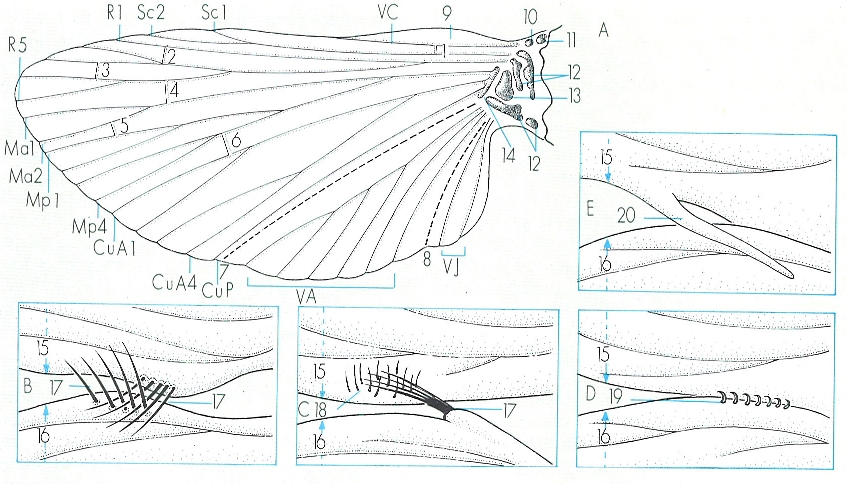
Esquema generalitzat d’una ala d’insecte, en el qual s’han representat les venes longitudinals i algunes de les transversals, els plecs principals i les esclerites de la base (A), i diferents tipus d’estructures alars, que permeten de mantenir les ales anterior i posterior de cada costat del cos unides durant el vol, corresponents a un mecòpter (B), molts lepidòpters heteròcers (C), himenòpters (D) i alguns lepidòpters (E). Hom hi consigna: VC vena costal, Sc subcostal, R radial, MA vena mediana anterior, MP mediana posterior, CuA cubital anterior, CuP cubital posterior, VA anal, VJ joval, 1 humeral, 2 transversa radial, 3 transversa del sector radial, 4 transversa medio-radial, 5 transversa mediana, 6 medio-cubital, 7 plec anal, 8 plec joval, 9 engruiximent precostal, 10 placa humeral, 11 tègula, 12 esclerites axil·lars, 13 placa mitjana posterior, 14 placa mitjana anterior, 15 ala anterior, 16 ala posterior, 17 fre, 18 retinacle, 19 hàmula, 20 jou.
Amadeu Blasco, a partir de fonts diverses
Les ales dels insectes són unes formacions especials, presents només en els pterigots, amb l’excepció d’alguns grups paràsits (anoplurs, sifonàpters i mal·lòfags), en els quals han desaparegut secundàriament, i d’altres espècies que pertanyen a grups alats, però que manquen d’ales a causa del seu tipus de vida peculiar; d’altra banda, en els insectes socials, algunes de les castes no tenen ales (tèrmits, formigues, etc.), i en d’altres (molts homòpters, per exemple), dins del cicle biològic s’alternen generacions alades i generacions àpteres; fins i tot, en algunes espècies, se succeeixen formes macròpteres, micròpteres i àpteres. En algunes espècies la presència d’ales és una manifestació del dimorfisme sexual: els mascles són alats i les femelles són micròpteres o, fins i tot, àpteres. D’altra banda, els apterigots no tenen ales, però en aquest cas es tracta d’un caràcter primitiu, ja que no és que al llarg de l’evolució les hagin perdudes sinó que mai no n’han arribat a tenir.
En principi, les ales són membranoses i tots dos parells tenen un desenvolupament semblant. No obstant això, en molts casos hom observa un enduriment de les ales del davant (que passen a anomenar-se èlitres, tegmines o hemèlitres) i, alhora, un desenvolupament desigual dels dos parells, o també la transformació del segon parell en uns òrgans especials, els balancins o halteris, que no serveixen per al vol, cosa que trobem en els dípters; encara, poden ésser les ales anteriors les que es redueixin, mentre que les posteriors són les responsables del vol en els mascles i les femelles són àpteres, fenomen ben conegut en els estrepsípters. Les ales són formacions tegumentàries recobertes de pèls, esquames, microtrics, etc., i de vegades presenten els marges plens de pèls llargs que els donen un aspecte ciliat; aquests trets serveixen per a caracteritzar els grans grups. Cada ala s’articula amb el metàmer toràcic corresponent per mitjà dels diversos processos tergals i per un de pleural. Alhora, a la base de l’ala es distingeixen diverses esclerites, en un nombre variable segons cada grup, que es diferencien a partir dels nervis longitudinals; les anomenades tègules, el procés humeral i els processos axil·lars són algunes d’aquestes esclerítes, que afavoreixen el moviment de les ales respecte del cos.
Les ales són constituïdes per dues capes de tegument que queden separades per un espai hemocèlic estret, recorregut en el seu interior per un conjunt de conductes esclerosats (nervis) pels quals circula hemolimfa, i que, per aquesta raó, s’anomenen també venes; aquest mateix espai de dins de les ales és recorregut, igualment, pel sistema de ramificacions de les tràquees.
La nervació alar és molt variada i caracteritza els diferents grups d’insectes. Hi ha nervacions longitudinals, que poden ramificar-se i donen consistència a l’ala alhora que impedeixen que es trenqui durant el vol, i nervacions transversals, que reben noms diferents segons la seva posició. En un model ideal d’ala d’insecte distingirem, des del marge anterior al posterior, els nervis o venes següents: costal, subcostal, radial, mediana, cubital i anal, alguns ramificats. A més, en els neòpters hi ha, almenys a les ales posteriors, altres nervis jovals. Totes aquestes nervacions poden fusionar-se en un grau variable a la base de l’ala. Entre els nervis longitudinals se situen nervis transversals que delimiten espais tancats i que hom anomena cel·les o cèl·lules alars; la nomenclatura específica d’aquests nervis depèn de la dels nervis longitudinals entre els quals se situen. De vegades, al marge anterior de l’ala, algunes cèl·lules s’engruixeixen, s’enfosqueixen de color i prenen el nom de pterostigma. Els nervis longitudinals delimiten tres regions a l’ala: el remigi és la regió més desenvolupada i la que participa d’una manera més activa al vol, i correspon a la zona que delimiten els nervis costal i cubital; el ventall o regió anal és una àrea petita ocupada pels nervis anals; i el jou o neala té un desenvolupament molt variable i només es diferencia quan hi ha nervis jovals (cas dels neòpters). En alguns grups que tenen les ales anteriors modificades en estructures protectores, les ales posteriors, que són les responsables del vol, es troben plegades sota les anteriors, i els plecs solen situar-se en el límit entre les regions ara indicades.
L’origen de les ales és un tema molt controvertit, per bé que la teoria més acceptada és la que defensa la seva derivació a partir d’unes evaginacions o expansions localitzades entre la regió tergal i la pleural, que són en relació amb la cavitat general del cos o hemocel; en principi, les ales eren fixes i posteriorment adquiriren les articulacions. Una altra teoria, també força acceptada, defensa que les ales deriven de formacions equivalents a les traqueobrànquies d’algunes larves aquàtiques. Des del punt de vista evolutiu hi ha una tendència a la simplificació de la nervació alar; per això, els grups considerats més primitius mostren una nervació més complexa, mentre que els més evolucionats únicament mantenen els nervis indispensables per a la dinàmica de vol, i fins i tot poden arribar a desaparèixer en algunes espècies, per exemple, d’heteròpters, himenòpters i dípters. L’ala s’estreny considerablement a la seva base, i els nervis longitudinals se solden entre ells i formen una mena de peduncle que permet una articulació sòlida i prima amb el tòrax.
Quan tots dos parells d’ales participen en el vol, el moviment de cada parell pot ésser independent, com passa en els paleòpters (odonats i efemeròpters) i en alguns neòpters (ortopteroïdeus), o bé poden haver-hi sistemes d’acoblament que faciliten el moviment coordinat dels dos parells, que es mouen sincrònicament. Hi ha diversos sistemes d’acoblament alar: el jou, el fre, l’hàmula, etc., que són peculiars de cada grup d’insectes. La funció d’aquests dispositius és de permetre que les ales romanguin unides durant el vol i que les de cada banda del cos es moguin d’una manera simultània, és a dir, segons un mecanisme sincrònic.
L’organització interna dels insectes
El tegument i l’endosquelet
L’estructura del tegument dels insectes no difereix del model general dels artròpodes, bé que és en ells on adquireix més complexitat. El tegument és la paret del cos i és format per tres capes que, de fora cap endins, són la cutícula, l’epidermis i la membrana basal.
La cutícula és una capa acel·lular, rígida, que protegeix externament el cos i els apèndixs de les deformacions mecàniques i de la transpiració. Actua com una mena d’esquelet extern (exosquelet). El seu component principal és la quitina i, en proporció menor, diverses escleroproteïnes (esclerotina, artropodina i resilina, aquesta darrera, exclusiva dels insectes); altres components de la cutícula són mucopolisacàrids i ceres, les quals són responsables de la impermeabilització. La cutícula és segregada per l’epidermis, que és una capa de cèl·lules monostratificades, i presenta ornamentacions diverses, que poden ésser resultat d’engruiximents o bé elements articulats segregats per cèl·lules epidèrmiques especialitzades. Quan és poc endurida, és pràcticament incolora, i va enfosquint-se a mesura que s’endureix; els pigments més freqüents són melanines (que produeixen colors molt diversos, des del groc al negre), pterines, característiques de les papallones (fluorescents i de colors variables entre el blanc i el vermell brillant, passant pel groc i el taronja), i carotenoides, que procedeixen de les plantes que formen part del seu règim alimentari i que de vegades, en metabolitzar-se, passen a formar part del tegument (donen colors grocs, vermells i ataronjats); n’hi ha molts d’altres, i de la combinació de tots plegats resulta l’àmplia gamma de colors que pot prendre el tegument dels insectes. En alguns casos, els grans de pigment, situats en cèl·lules especials, poden contraure’s o dilatar-se per acció hormonal, cosa que permet els canvis de color del cos, com és el cas dels insectes bastó, que imiten el color dels arbusts sobre els quals viuen. Quan els pigments es localitzen a la cutícula, els colors es mantenen inalterats després que l’insecte mor. A més dels colors químics, produïts per pigments, hi ha també colors físics, responsables de les tonalitats metàl·liques (verdoses, blavoses, argentades, etc.), que varien segons quina sigui l’orientació de l’insecte vers la font de claror; aquests colors es deuen a la mateixa estructura de la cutícula.
La duresa i la consistència de la cutícula obliga els insectes, com la resta d’artròpodes, a un creixement discontinu: periòdicament, els insectes canvien la cutícula vella per una de nova, més gran, pel procés de la muda, que és controlat per hormones. A més de revestir el cos i els apèndixs, la cutícula també revesteix el sistema traqueal, la regió anterior i posterior del tub digestiu, els canals excretors d’algunes glàndules i determinades parts de l’aparell reproductor; quan l’animal muda, també es renova la cutícula que recobreix totes aquestes estructures. La cutícula proporciona, alhora, una base sòlida per a la inserció dels músculs, i presenta invaginacions locals que constitueixen l’endosquelet dels insectes.
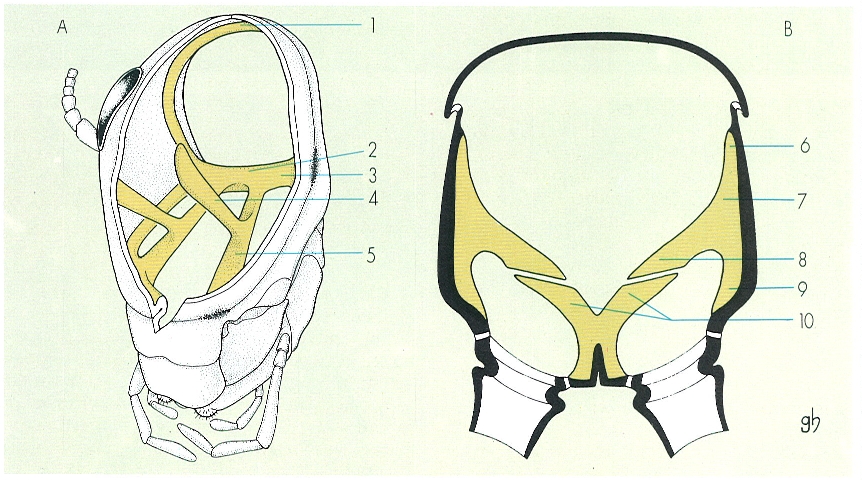
Esquema de l’endosquelet del cap (A) d’un insecte (parcialment seccionat per tal de fer-lo més visible) i del tòrax (B), en un tall transversal. Hom hi consigna: 1 fragma o cresta postoccipital, 2 cos tentarial 3 braços posteriors del tentori o metatentori, 4 braços dorsals del tentori o supratentori, 5 braços anteriors del tentori o pretentori, 6 procés alar, 7 cresta pleural, 8 braç pleural, 9, procés costal, 10 forca (correspon a les endosternites).
Gustavo Hormiga, a partir d’originals de Snodgrass.
L’endosquelet és el conjunt d’invaginacions cuticulars o apodemes sobre els quals s’insereixen els músculs que mouen els apèndixs i els que relacionen els diversos metàmers. En els insectes és més desenvolupat al cap i al tòrax, regions on es formen estructures complexes a causa de la coalescència de diferents apodemes.
Al cap hi ha el fragma postoccipital i el tentori. El fragma postoccipital correspon a una cresta interna del solc postoccipital, i al damunt seu s’insereixen els músculs que mouen les peces bucals (al davant) i els músculs protoràcics, motors del cap (al darrere). El tentori és una estructura molt característica damunt de la qual s’insereixen la majoria dels músculs cefàlics i que dona rigidesa a la càpsula cefàlica, subjecta l’encèfal i la regió anterior del tub digestiu, i alhora reforça els punts d’articulació de les peces bucals; en els apterigots entòtrofs és poc desenvolupat i format per diverses peces independents (pseudotentori), mentre que en els pterigots és format per una sola estructura, amb dos braços anteriors (pretentori), dos braços posteriors (metatentori) i dos de dorsals.
Al tòrax, l’endosquelet o endotòrax és format per invaginacions de les pleurites, tergites i esternites de cada metàmer i és particularment ben desenvolupat en els metàmers que porten les ales, és a dir, al mesotòrax i al metatòrax. Les invaginacions de les tergites, anomenades endotergites o fragmes, se situen a la regió anterior de cada metàmer toràcic, i les sutures corresponents les divideixen en dues esclerites desiguals, una d’anterior i estreta, i una altra de voluminosa i posterior; corresponen a plecs interns de les esclerites intersegmentàries i serveixen de punt d’inserció dels músculs dorsals longitudinals. Les invaginacions pleurals, anomenades endopleurites o apodemes laterals, sorgeixen de la part interna de les pleures. Cada metàmer portador d’ales presenta típicament un plec intern, anomenat cresta pleural, que se situa al llarg del solc pleural i acaba en el procés alar, dirigit cap amunt, i en el procés coxal, dirigit cap avall; i encara, sovint hi ha, a més, un braç pleural dirigit cap endins. Les invaginacions esternals o endosternites són formades en general per un parell d’apòfisis que, en conjunt, prenen forma de lletra Y i s’anomenen furca; els braços externs d’aquesta apòfisi es poden fusionar amb els braços pleurals o bé connectar-s’hi per mitjà de músculs. En molts insectes també hi ha un apodema mitjà, no ramificat, anomenat espina. D’aquest model bàsic d’endosquelet hi ha nombroses modificacions. A l’endotòrax s’insereixen els músculs que relacionen els diferents metàmers i els que mouen ales i potes.
A l’abdomen l’endosquelet és poc desenvolupat. La majoria dels metàmers tenen crestes internes dorsals o fragmes, semblants als del tòrax, als quals s’insereixen els músculs dorsals; en posició ventral sol haver-hi apòfisis esternals. Alguns metàmers poden presentar formacions especials de connexió amb els òrgans copuladors en el mascle, i amb l’ovipositor en les femelles.
La musculatura
La musculatura dels insectes, com passa a la resta dels grups d’artròpodes, és formada per feixos de fibres estriades que es disposen metamèricament. Hi ha dos tipus de músculs: els esquelètics i els viscerals. Els músculs esquelètics són molt desenvolupats i són els responsables del moviment de les antenes i les peces bucals, de les potes i de les ales, i també d’altres formacions, com els òrgans copuladors; alhora, s’encarreguen del moviment relatiu dels metàmers del cos i dels tagmes en què es divideix. S’insereixen en les formacions endosquelètiques (apodemes, tentori, etc.) o bé directament a la part interna de la cutícula i són en relació amb les peces o regions que s’han de moure. La nomenclatura dels músculs és variada i complexa; en alguns casos, com algunes erugues de papallona, hi ha més de mil músculs diferents. Els músculs viscerals es presenten aïllats o en grups, de vegades organitzats fent una mena de trama a l’entorn de les gònades i els seus conductes, el cor, la paret del tub digestiu, el diafragma, etc., i també formen els esfínters que hi ha a diferents punts del tub digestiu amb la missió de regular el pas de l’aliment. Les seves propietats, en alguns aspectes, són intermèdies entre les dels músculs esquelètics i els músculs llisos d’altres grups d’invertebrats; d’altra banda, segons el ritme de contracció, els músculs poden ésser sincrònics o asincrònics, aquests darrers localitzats només a les ales.
Els sistemes de relació i control
Els sistemes de relació i de control, en els insectes, responen al model general de tots els artròpodes, si bé en alguns grups arriben a atènyer un desenvolupament i una complexitat molt més grans, especialment en els insectes socials.
El sistema nerviós central
El sistema nerviós central dels insectes és ganglionar i hiponeure. Consisteix en una massa ganglionar supraesofàgica (és a dir, un cervell) i una cadena ventral proveïda d’un parell de ganglis per cada metàmer, bé que generalment no són tots visibles a causa de fusions secundàries; el cervell queda unit a la cadena ventral per mitjà del collar periesofàgic.
El cervell és constituït per tres parells de ganglis, que conformen tres parts diferenciades: el protocervell, el deutocervell i el tritocervell.
El protocervell és molt desenvolupat ja que és on resideixen els centres òptics i ocel·lars, a més dels centres d’integració i coordinació. És format per dos lòbuls cerebrals que queden separats per l’anomenada part intercerebral (pars intercerebralis), on hi ha les cèl·lules secretores de neurohormones que regulen el funcionament de les glàndules endocrines. Les diferents àrees del protocervell varien de desenvolupament segons els grups d’insectes. Una d’elles, l’anomenat cos central (pons cerebralis) és un centre d’associació important, responsable de les relacions dels lòbuls òptics simètrics i de la coordinació de les impressions visuals, i es relaciona amb altres zones del cervell; els cossos ventrals estableixen connexions amb els dos lòbuls del protocervell i es relacionen amb el cos central, els cossos pedunculats, els lòbuls òptics i altres parts; els cossos pedunculats se situen a banda i banda de la part intercerebral i són centres d’associació sensorial molt diferenciats en els insectes socials, que estableixen relació amb la resta de centres nerviosos i permeten la integració i la codificació dels estímuls rebuts, de manera que adeqüen la resposta motriu a la seva intensitat; dels cossos laterals, els més importants són els lòbuls òptics, als quals se situen els centres òptics, i solen ésser molt desenvolupats en relació amb la complexitat dels ulls compostos. El protocervell innerva la regió acronal i el metàmer preantenal, on se situen els ulls.
El deutocervell és format per dues masses nervioses reunides a través d’una comissura supraesofàgica, i en ell se situen els centres antenals, dels quals parteixen els nervis corresponents; innerva, per tant, les antenes i el metàmer corresponent.
El tritocervell és poc desenvolupat perquè el metàmer que innerva no porta apèndixs; és format per dues masses nervioses que contenen dos centres nerviosos. Es connecta amb el sistema nerviós simpàtic anterior, que innerva la regió anterior del tub digestiu. Els connectius que el relacionen amb la cadena ventral constitueixen el collar periesofàgic.
La cadena ventral és formada per tants parells de ganglis com metàmers té el cos, descomptant, és clar, els que són innervats directament pel cervell. No obstant això, en general hi ha ganglis fusionats, de manera que el nombre de parells de ganglis sol ésser inferior: els tres primers ganglis fusionats constitueixen la massa ganglionar infraesofàgica o gnatocervell, que innerva les peces bucals i els metàmers corresponents (el mandibular, el maxil·lar i el labial); els darrers ganglis també tendeixen a fusionar-se. D’altra banda, hi ha una propensió evolutiva generalitzada cap a la concentració del sistema nerviós, de manera que en els grups considerats més evolucionats la concentració és més gran, fins arribar al cas extrem de la fusió de tots els ganglis toràcics i abdominals fent una única massa nerviosa, que engloba, fins i tot, la infraesofàgica; hi ha tota una àmplia gamma de variació entre el model considerat més primitiu i el més evolucionat. De cada gangli de la cadena ventral parteixen nervis que innerven els metàmers corresponents, amb els seus apèndixs i formacions especials, quan n’hi ha; aquests ganglis contenen cèl·lules neurosecretores. En relació amb la cadena ventral hi ha el sistema nerviós simpàtic posterior o visceral, que és ben diferenciat.
El sistema nerviós simpàtic
El sistema nerviós simpàtic innerva les vísceres, regula les seves funcions i tot el funcionament automàtic de l’aparell reproductor, com és l’oviposició i la deposició de l’esperma. Té dues regions diferenciades, que prenen el nom de sistema simpàtic anterior o estomatogàstric i simpàtic posterior, caudal o genital.
El sistema estomatogàstric innerva la part anterior del tub digestiu, inclosa la cavitat oral. Se situa en posició dorsal respecte del tub digestiu i es compon d’un gangli principal, el gangli frontal, que és en relació amb el tritocervell, i d’un de secundari, el gangli hipocerebral, que es relaciona amb el primer per mitjà del nervi recurrent i també es pot relacionar amb altres ganglis (ingluvials, postventriculars, etc.). D’altra banda, els cossos cardíacs es connecten amb el gangli hipocerebral i amb el protocervell i, alhora, amb els cossos alats. Aquest model general queda sotmès a múltiples variacions en els diferents grups.
El sistema nerviós simpàtic posterior es compon d’un nervi imparell, l’anomenat nervi de Leydig, que discorre per sobre de la cadena ventral amb la qual es relaciona. Innerva el proctodeu, els òrgans genitals, els estigmes, el tegument i la musculatura. D’aquest nervi en neixen d’altres que són simètrics, a nivell de cada parell de ganglis de la cadena ventral, i en relació amb aquests se situen els ganglis perisimpàtics.
Els òrgans sensorials: els òrgans mecanoceptors
Els òrgans sensorials es troben molt desenvolupats en els insectes i perceben una gran varietat d’estímuls, externs i interns. En els casos més simples, un òrgan sensorial és constituït per una sola sensília, mentre que en els més complexos se’n reuneixen diverses, de manera que la sensília esdevé la seva unitat funcional. Hi ha diferents tipus de sensílies i sempre es disposen en relació amb formacions tegumentàries, atès que són d’origen ectodèrmic. Malgrat la gran diversitat dels òrgans sensorials, hom pot considerar-ne de tres tipus principals, segons el tipus d’estímul que poden rebre: mecanoceptors, quimioceptors i fotoceptors, cadascun dels quals pot presentar nombroses especialitzacions.
Els òrgans mecanoceptors són excitats per estímuls que impliquen la deformació mecànica d’alguna part del receptor (sensília), deformació que pot ésser deguda al contacte amb objectes sòlids o amb corrents d’aigua o d’aire; també poden respondre a tensions mecàniques internes, i en aquest cas actuen com a òrgans propioceptors; encara, hi ha casos en què són estimulats per canvis de la pressió de l’aire, i llavors actuen com a receptors de so. Un tipus especialitzat de mecanoceptor el constitueixen els òrgans termoceptors, sensibles a la temperatura. En conjunt, els òrgans mecanoceptors faciliten l’orientació dels organismes en el seu medi i alhora els permeten de reconèixer-ne les característiques; el control de la postura del cos, de la locomoció i del vol depèn d’aquest tipus d’òrgans sensorials. Els d’estructura més senzilla i alhora els més estesos en els insectes són els pèls tàctils, que es troben distribuïts per tota la superfície del cos i dels apèndixs, i que són estimulats quan es desplacen respecte de la seva base, és a dir, del punt on s’articulen.
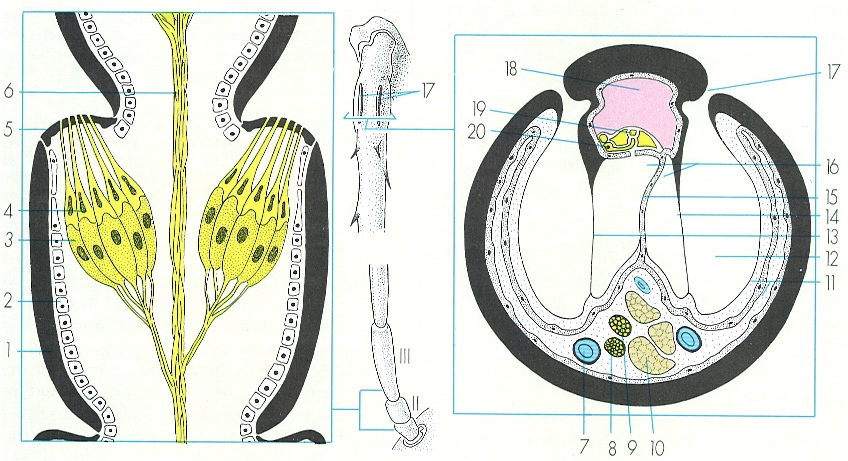
Òrgans cordotonals dels insectes: A òrgan de Jonhston, simplificat, que se situa en el segon artell antenal (A’) dels pterigots i els zigentomes, constituït per un o dos grups d’escolopidis; de funció mecanoceptora, intervé en el control de la positura i en la coordinació de moviments en el vol. B òrgan timpànic, simplificat, que correspon a un òrgan cordotonal molt especialitzat, capaç de percebre sons; en el cas que representa el dibuix —un ortòpter ensífer— se situa a la zona proximal de les tíbies (B’), i té les cèl·lules sensitives en contacte, a través del seu pol sensible, amb una membrana, la membrana timpànica, que s’allotja dins d’una cavitat. El dibuix mostra les parts següents: 1 cutícula, 2 epidermis, 3 cèl·lula sensorial, 4 cèl·lules escolopals o escolopidis, 5 membrana articular, situada entre el segon i el tercer artells antenals, 6 nervi antenal, 7 tràquees, 8 nervi tarsal, 9 nervi tibial, 10 músculs, 11 operele del timpà, 12 cavitat timpànica, 13 timpà anterior, 14 timpà posterior, 15 envans centrals de les tràquees timpàniques, 16 tràquees timpàniques, 17 cresta acústica, 20 cèl·lula sensorial.
Amadeu Blasco, a partir de fonts diverses
Els òrgans mecanoceptors sensibles als canvis de pressió són capaços de detectar les vibracions procedents del medi, ja sigui sòlid, líquid o aeri, i es troben molt especialitzats. Reben un nom diferent segons la seva funció. Els anomenats òrgans cordotonals poden ésser formats per un nombre molt variable de sensílies (entre una i milions) especialitzades, anomenades escolopals o escolopidis, que es localitzen a les zones articulars del cos i dels apèndixs, des d’on detecten els canvis de pressió per la flexió o distensió que aquestes experimenten amb la qual cosa adquireixen noció de la postura del cos; també capten les vibracions procedents del medi. L’òrgan cordotonal més generalitzat entre els insectes és l’òrgan de Jonhston, que es localitza en el segon artell de les antenes en els insectes alats (pterigots) i en els zigentomes i que, pel que sembla, intervé en el control del vol i de la natació, així com, en general, en el control de la postura del cos; en alguns dípters, també poden detectar sons. Hi ha, encara, altres tipus d’òrgans cordotonals, com les tricobòtries.
D’entre els òrgans mecanoceptors, són interessants també els òrgans auditius, que són estimulats pels canvis de pressió de l’aire i que han estat detectats exclusivament en els insectes; en els casos més simples es tracta de pèls auditius o acústics, i en els més complexos, d’òrgans timpànics. Els primers són òrgans cordotonals modificats mentre que els òrgans timpànics consten d’una membrana cuticular o timpànica, situada prop d’un sac traqueal i per sobre d’un grup d’òrgans cordotonals que reben el nom d’òrgan de Müller; de vegades, la membrana timpànica es troba allotjada dins d’una cavitat, la cavitat timpànica, que n’augmenta la ressonància. Els òrgans timpànics poden adoptar un aspecte molt variable i es localitzen a diferents parts del cos; se sap de la seva existència en els 11agosts (ortòpters), les cigales (homòpters cicàdids) i en algunes papallones (lepidòpters), entre altres.
En els insectes, la recepció de sons es combina amb l’emissió de sons, fenomen important per a la comunicació en moltes espècies. L’emissió de sons pot ésser induïda per estímuls externs, interns o sexuals. Els insectes poden emetre sons de maneres diferents: colpejant el terra amb alguna part del cos, fregant les estructures dels anomenats òrgans estriduladors, fent vibrar les ales durant el vol, o per mitjà d’òrgans especials, com fan les cigales, els ortòpters i altres insectes. Els sons emesos pels òrgans estriduladors i pels òrgans especials són molt més intensos que no els produïts per altres mitjans. Molts d’aquests sons no semblen tenir cap significat biològic; tanmateix, en el cas dels llagosts (els ortòpters), que tenen òrgans estriduladors i òrgans timpànics, no hi ha dubte que actuen com a reclams sexuals; en el cas de les cigales, influeixen en el gregarisme.
Entre els mecanoceptors propioceptors, és a dir, sensibles als estímuls interns de l’organisme, hem d’esmentar els receptors de tensió muscular, que es localitzen als músculs i els tendons, i els estatocists o òrgans de l’equilibri. D’òrgans de l’equilibri només se’n coneixen en els efemeròpters (l’òrgan de Palmer), i excepcionalment en algun himenòpter i algun isòpter.
Els òrgans sensorials: els òrgans quimioceptors
Els òrgans quimioceptors són sensibles a estímuls de natura química. En les espècies aquàtiques són estimulats per substàncies dissoltes a l’aigua i, en les terrestres, l’estímul pot captar-se a distància, a través dels òrgans olfactius, o bé per contacte, a través dels òrgans gustatius. L’estructura dels òrgans quimioceptors és senzilla. En general són formats per una sola sensília, que té aspecte de pèl o de papil·la, i es disposen a diferents regions del cos i dels apèndixs. Els òrgans olfactius, estimulats per gasos, són molt importants en el comportament de les espècies, atès que detecten a distància les feromones (d’alarma, d’agregació, sexuals, etc.) emeses per altres individus de la mateixa espècie i, també, els permeten de localitzar el lloc més adient per a deixar la posta; també afavoreixen la recerca de l’aliment i dels hostes en el cas d’espècies paràsites (precisament, és a aquests òrgans que es deu l’especificitat de molts paràsits). Els òrgans olfactius es localitzen preferentment a les antenes i en els palps maxil·lars i labials.
Els òrgans gustatius són estimulats per substàncies sòlides o líquides, i són molt importants en el reconeixement dels aliments. Se situen principalment a la cavitat oral, a les peces bucals, a la hipofaringe i als tarsos de les potes. Un tipus especial d’òrgan quimioceptor són els higroceptors, sensibles a la humitat relativa de l’ambient.
Els òrgans sensorials: els òrgans fotoceptors i fotoemissors
Els òrgans fotoceptors són estimulats per l’energia lluminosa i permeten als insectes de percebre la llum ambiental i, fins i tot, de reproduir el medi a través de la formació d’imatges. Els insectes també són capaços de percebre els colors, encara que l’extensió del seu espectre visible és diferent a la de l’home. D’altra banda, moltes espècies són especialment sensibles a les radiacions ultraviolades (de baixa longitud d’ona); segons sembla, les abelles són sensibles a aquest tipus de llum i la utilitzen per a orientar-se i per a localitzar l’aliment. La percepció de les formes en els insectes és menys perfecta que en l’home i la seva agudesa visual també és més petita.
Hi ha dos tipus bàsics d’òrgans visuals: els ocel·les o ulls simples i els ulls compostos. Algunes larves tenen uns ulls laterals anomenats estemmates o estemmatidis, d’estructura intermèdia entre els ocel·les i els ulls compostos; són característics d’algunes larves d’holometàbols i es localitzen al mateix lloc del cap on, després, els adults portaran els ulls compostos. Són formats per un nombre variable de sensílies i la seva estructura és molt diversa.
Els ocel·les se situen al cap, en posició dorsal i, generalment, es troben en nombre de tres, bé que en alguns casos pot no haver-n’hi cap. Són capaços de percebre la claror, de manera que intervenen en l’orientació dels organismes.
Els ulls compostos se situen en posició lateral al cap i, en general, atenyen un gran desenvolupament. Hi ha alguns insectes apterigots (els entòtrofs) que no en tenen mai, mentre que a d’altres, que originalment en tenen, se’ls poden anar reduint i, fins i tot, se’ls poden atrofiar completament. Són òrgans sensorials complexos, formats per un nombre de sensílies molt variable i sovint molt elevat (fins a milers), cadascuna de les quals rep el nom d’ommatidi. Segons quina sigui l’estructura del cristal·lí, hi ha quatre tipus d’ommatidis: eucons, acons, pseudocons i exocons. Independentment de quin sigui el tipus d’ommatidi, hi ha entre els ulls compostos dos tipus de visió: per aposició i per superposició. En la visió per aposició cada ommatidi forma la seva pròpia imatge, de manera que es formen tantes imatges com ommatidis componen l’ull; aquest tipus de visió és característic de les espècies diürnes. En la visió per superposició no hi ha un aïllament total entre els diferents ommatidis, i els raigs de llum poden travessar-ne més d’un, de manera que es formen diverses imatges superposades; aquest tipus de visió és característic de les espècies nocturnes. En alguns casos, els ulls poden actuar com a ulls d’aposició o de superposició segons quines siguin les condicions de llum ambiental.
Alguns insectes són capaços d’emetre llum. Aquesta facultat s’interpreta, en alguns casos, com un reclam sexual. La llum emesa pels insectes pot ésser originada per bacteris simbionts fosforescents o bé pot ésser produïda pel mateix metabolisme de l’insecte. Els òrgans lluminosos es localitzen a diferents parts del cos i la lluminositat es deu a la presència de dues substàncies: la luciferina i la luciferasa. La luciferina, en presència de la luciferasa, s’oxida i es transforma en oxiluciferasa, i alhora desprèn energia lluminosa. La llum que s’emet en aquest cas és freda i d’intensitat variable. Entre els insectes que tenen òrgans lluminosos hi ha alguns coleòpters (lampírids, elatèrids i cantàrids, especialment) i algunes larves de dípters (Arachnocampa luminosa); el cas d’aquesta darrera espècie, neozelandesa, és particular, ja que els seus òrgans lluminosos deriven dels tubs de Malpighi, i utilitza la llum emesa per a atraure les seves preses.
El sistema secretor
Controlat pel sistema nerviós, el sistema secretor s’encarrega de proporcionar a l’organisme substàncies que regulen funcions d’índole diversa, sigui fisiològica, conductual, reproductora, etc. En els insectes, les glàndules secretores són formades per una cèl·lula o un grup de cèl·lules que produeixen diferents substàncies. Hom pot distingir dos tipus de glàndules: les exocrines, proveïdes d’una obertura o conducte ben definit, pel qual descarreguen els seus productes a l’exterior del cos o a determinades vísceres, i les endocrines, que no tenen conductes especials i aboquen els seus productes (hormones) generalment a l’hemolimfa, que és qui les transporta per tot el cos.
El sistema endocrí
El sistema endocrí, també anomenat retrocerebral, és format pel conjunt de glàndules endocrines. El funcionament d’aquestes glàndules és regulat per les neurohormones, que són segregades per cèl·lules neurosecretores, les quals es localitzen especialment al protocervell (a la part intercerebral) i a la cadena ventral (als òrgans perisimpàtics). Les cèl·lules neurosecretores són cèl·lules nervioses modificades, i la seva secreció de neurohormones és, al seu torn, controlada pel sistema nerviós, de manera que en conjunt, aconsegueix una perfecta coordinació química i nerviosa de l’organisme. En els insectes, el sistema endocrí es troba ben diferenciat i la seva composició és força constant a tots els grups, bé que pot variar el grau de desenvolupament i la situació de les diverses glàndules. Hom diferencia bàsicament tres glàndules distintes: els cossos cardíacs, els cossos alats i les glàndules protoràciques.
Els cossos cardíacs són dos òrgans simètrics que es troben situats a les parets laterals de l’aorta, de vegades fusionats i fent una sola massa. Actuen com a òrgans neurohemals i com a glàndules endocrines: d’una banda, fan de magatzem de les neurohormones que es produeixen al protocervell i a d’altres regions cerebrals, i de l’altra, segreguen hormones pròpies. Tenen connexió amb el cervell a través de diversos nervis (els nervis cardíacs) i amb el gangli hipocerebral del sistema estomatogàstric i els cossos alats. Els cossos cardíacs regulen una infinitat de processos fisiològies (el ritme cardíac, el peristaltisme intestinal, el creixement, etc.), però es fa difícil saber quines funcions són regulades per les secrecions pròpies i quines per les secrecions exògenes.
Els cossos alats són dues glàndules simètriques que se situen a banda i banda de l’esòfag. Es connecten amb els cossos cardíacs i amb el gangli infraesofàgic. Aquestes glàndules endocrines segreguen l’hormona juvenil, responsable del manteniment dels caràcters larvals mentre dura el desenvolupament postembrionari; en la vida adulta assegura la vitel·logènesi en les femelles i l’activitat de les glàndules annexes a l’aparell genital.
Les glàndules protoràciques se situen prop del sistema nerviós simpàtic anterior, però es mantenen independents i tenen un aspecte molt variable segons els grups. Segreguen l’hormona de la muda, anomenada ecdisona, i en general degeneren en l’estat adult, ja que la majoria dels insectes deixen de créixer i, per tant, de mudar quan adquireixen els caràcters d’adult. Es connecten amb diferents regions del sistema nerviós (la massa ganglionar infraesofàgica, els ganglis protoràcics), segons quina sigui la seva situació al cos.
La fisiologia de les glàndules endocrines és força complexa. Les neurohormones que es fabriquen al cervell s’acumulen als cossos cardíacs, i passen a través d’aquests a l’hemolimfa. L’hormona del cervell o protoracotropa indueix l’elaboració de l’hormona de la muda, que és segregada per les glàndules protoràciques; també regula la diapausa en els estadis juvenils. D’altra banda, l’activitat dels cossos alats també és regulada per una neurohormona segregada pel cervell, que quan és present a l’hemolimfa indueix la secreció de l’hormona juvenil; l’acció de l’hormona juvenil sobre la metamorfosi és molt important, ja que si la muda es fa en presència d’una concentració elevada d’hormona juvenil es mantenen els caràcters larvals, mentre que si la concentració d’aquella hormona és baixa, l’organisme adquireix els caràcters de l’adult. Així, el creixement i el desenvolupament dels insectes és controlat i regulat per l’activitat neurosecretora del cervell.
El sistema exocrí
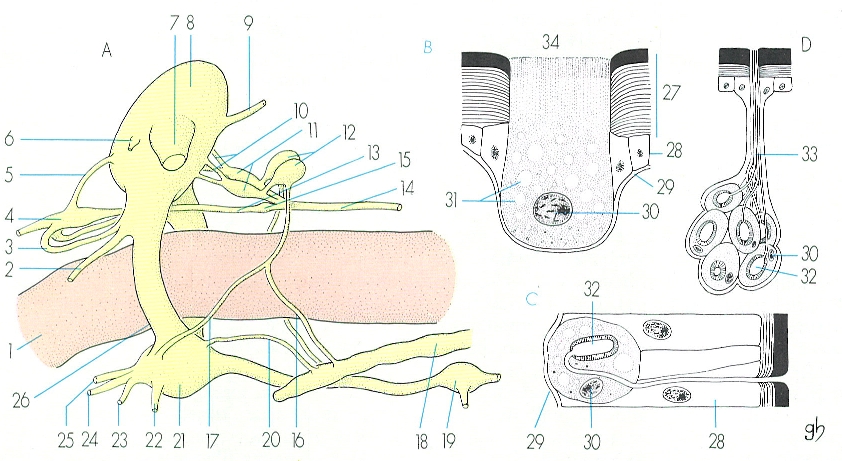
Esquema de diferents estructures del sistema secretor dels insectes: A part anterior del sistema nerviós central, del sistema nerviós simpàtic anterior o estomatogàstric i del sistema endocri o retrocerebral, B glàndula exocrina unicel·lular senzilla, que verteix la secreció per difusió a través de la cutícula, C glàndula exocrina unicel·lular, segons un model més evolucionat, que presenta un reservori dels productes secretats i un conducte eferent, ben delimitat, D glàndula exocrina pluricel·lular, en la qual cada cèl·lula presenta les mateixes característiques que les glàndules exocrines unicel·lulars representades a l’esquema C. Hom hi ha representat: 1 tub digestiu, 2 nervi del labre, 3 connectiu frontal, 4 gangli frontal, 5 connectiu, 6 nervi antenal, 7 nervi òptic, 8 cervell, 9 nervi tegumentari, 10 nervis dels cossos cardíacs, 11 cossos cardíacs, 12 cossos alats, 13 nervi recurrent, 14 nervi esofàgic, 15 gangli hipocerebral, 16 branca protoràcica del nervi dels cossos alats, 17 branca infraesofàgica del nervi dels cossos alats, 18 glàndula protoràcica, 19 gangli protoràcic, 20 nervi infraesofàgico-glàndula protoràcica, 21 massa ganglionar infraesofàgica, 22 nervi labial, 23 nervi maxil·lar, 24 nervi hipofaringi, 25 nervi mandibular, 26 collar periesofàgic, 27 cutícula, 28 epidermis, 29 membrana basal, 30 nucli d’una cèl·lula glandular, 31 secrecions, 32 reservori, 33 conducte eferent, 34 superfície d’excreció.
Gustavo Hormiga, a partir de fonts diverses
El sistema exocrí és constituït fonamentalment per glàndules tegumentàries, que poden ésser unicel·lulars o pluricel·lulars. La natura dels productes elaborats és molt variada, i dona lloc a diferents tipus de glàndules, dels quals comentarem els principals. Les glàndules sericígenes produeixen seda, generalment en forma de filets molt prims, que s’entreteixeixen i formen els embolcalls de larves i capolls. Les glàndules cerígenes produeixen cera, que s’utilitza per a recobrir-se el cos, per a protegir els ous i per a construir nius. Les glàndules lacíferes segreguen laca i són característiques d’un grup d’homòpters, els laccifèrids; d’aquest producte l’home n’obté la laca comercial, que és una substància resinosa. Les glàndules cefàliques, que se situen al cap, tenen secrecions variades; a més de les glàndules frontals, defensives, dels tèrmits (isòpters) i de les glàndules antenals d’alguns altres grups d’insectes, la resta se situa en relació amb els apèndixs bucals i reben el nom de glàndules mandibulars, maxil·lars o labials, segons el lloc on se situïn. De totes aquestes, les glàndules labials, també anomenades glàndules salivals, són les que segreguen substàncies més variades; a més de la seva funció digestiva en els insectes que mengen vegetals (fitòfags), poden tenir un efecte tòxic per a les plantes i també, de vegades, poden secretar seda. Un altre tipus de glàndules molt diferent són les glàndules de verí, que es troben especialment ben desenvolupades en els insectes himenòpters apòcrits, i que es troben en relació amb l’aparell reproductor de les femelles, les quals tenen l’oviscapte transformat en un fibló que es connecta directament a la glàndula. Les glàndules urticants són un altre tipus de glàndula verinosa, generalment connectada a pèls i sedes buits de dins i fràgils; són corrents en moltes erugues de papallona (per exemple, en la processionària del pi). Hi ha també glàndules repugnatòries, repel·lents o defensives, les secrecions de les quals desprenen una olor pudent i tenen propietats repel·lents, de manera que fan una funció defensiva important.
Les glàndules odoríferes, productores de feromones, segreguen substàncies oloroses que poden ésser reconegudes pels individus de la mateixa espècie i que actuen de maneres diverses entre ells, tot provocant-los una reacció específica. Solen associar-se a pèls o esquames i per tal que les seves secrecions siguin efectives cal que hi hagi òrgans sensorials (quimioceptors) capaços de reconèixer-les per l’olor. Aquestes secrecions poden ésser de diferents tipus: feromones marcadores, d’agregació, d’alarma i sexuals. Les feromones marcadores de pistes són corrents en els insectes socials i en algunes espècies gregàries i serveixen per a marcar el terreny per on circulen, per tal de trobar després el camí de tornada. Les feromones d’agregació en els insectes socials col·laboren a mantenir la unitat de la colònia, amb la corresponent separació de castes; en els insectes solitaris són emprades per a concentrar els individus en determinades zones (gregarisme), amb finalitats diverses: sigui per a la posta, la hivernada, etc. Les feromones d’alarma són freqüents en els insectes socials i serveixen per a informar la resta dels membres de la colònia de l’existència d’algun perill; aquestes feromones, com les marcadores i les d’agregació, no són gaire específiques i sempre actuen només a distàncies curtes. Les feromones sexuals són les responsables de l’atracció sexual i del reconeixement dels sexes, i ajuden a la localització de la parella i faciliten la còpula. Aquestes sí que són molt específiques i poden actuar a grans distàncies. En molts casos, són les femelles les qui emeten les feromones, i els mascles les capten i es desplacen fins a trobaries, però en d’altres passa el cas contrari. Les feromones sexuals són de natura molt variada.
L’aparell digestiu
Com a tots els artròpodes, l’aparell digestiu dels insectes mostra tres regions diferenciades: una d’anterior o estomodeu, una d’intermèdia o mesodeu, també anomenada estómac glandular, i una altra de posterior, el proctodeu. Les regions anterior i posterior són d’origen ectodèrmic i van revestides de cutícula, i, per tant, es renoven periòdicament amb la muda. El mesodeu, en canvi, és endodèrmic i és el lloc on es fa la digestió enzimàtica dels aliments. El tub digestiu és revestit de músculs viscerals, que faciliten la progressió de l’aliment.
L’estomodeu s’inicia a la boca, que és situada a la regió anterior del cos. En els casos més simples es tracta d’un tub rectilini, però generalment té diferents regions. A continuació de la boca hi ha la cavitat bucal (preoral), que és delimitada per les peces bucals i el labre, i en general presenta dues regions diferents, separades per la hipofaringe: el cibari (anterior o dorsal) i el salivari (posterior o ventral). Prop d’aquest darrer desemboquen les glàndules salivals.
A l’estomodeu es poden diferenciar també altres regions: la faringe, l’esòfag, el pap o ingluvi i el pedrer o proventricle. La importància de cadascun d’aquests trams depèn del règim alimentari. La faringe sempre és molt musculosa, i de vegades és tan desenvolupada que constitueix una bomba aspiradora especialment potent en els insectes que s’alimenten de fluids (xucladors). L’esòfag acostuma a ser un tub curt i estret, amb nombrosos plecs a les parets internes. El pap o ingluvi s’allotja al protòrax i actua com a magatzem d’aliment. El pedrer o proventricle és un estómac triturador i, a més de tenir una musculatura molt potent, va revestit internament de nombroses crestes, dents i espines que afavoreixen la trituració dels aliments sòlids. En el punt d’unió de l’estomodeu amb el mesodeu hi ha una vàlvula, la vàlvula cardíaca, que regula el pas dels aliments d’una regió a l’altra del tub digestiu, i és en relació amb un esfínter. La funció de l’estomodeu és la de participar en la captura i la trituració dels aliments sòlids i iniciar-ne la digestió enzimàtica per mitjà de la saliva.
La regió intermèdia, és a dir, el mesodeu o estómac glandular en general té forma de tub més o menys llarg, bé que en alguns casos pot tenir forma de sac. En alguns grups d’insectes s’hi diferencien regions amb funcions especialitzades; altres vegades, té diversos cecs gàstrics que, en general, es localitzen a la regió anterior, i vellositats intestinals, la funció de les quals és augmentar la superfície d’absorció. La funció del mesodeu és la digestió enzimàtica dels aliments i l’absorció dels productes ja digerits. En aquest nivell del tub digestiu els aliments s’envolten de l’anomenada membrana peritròfica, que impedeix que toquin directament l’epiteli intestinal i alhora n’afavoreix l’absorció; aquesta membrana es forma d’una manera contínua cada vegada que arriba aliment al mesodeu i s’elimina amb els excrements. Sol mancar en les espècies que s’alimenten de líquids.
La regió posterior o proctodeu mostra una estructura semblant a la de l’estomodeu. El seu inici queda marcat per la vàlvula pilòrica i l’esfínter corresponent, i pel punt d’inserció dels tubs de Malpighi (òrgans excretors). A la majoria dels insectes, el proctodeu té tres regions, l’ili, el còlon i el recte, i acaba en l’anus, a la part terminal del cos. El tram corresponent al recte pot presentar papil·les i glàndules rectals, la secreció de les quals té funcions diverses (defensives, atractives, etc.) i participa en la reabsorció de l’aigua i de certs ions, de manera que col·labora en la regulació hídrica de l’organisme i en l’eliminació dels productes residuals procedents del mesodeu; al marge d’aquestes funcions que el recte fa típicament, pot actuar també com a òrgan respiratori (les traqueobrànquies rectals d’alguns odonats i les ampolles rectals) i pot allotjar simbionts de natura diversa.
El tub digestiu pot presentar una infinitat de modificacions en funció de quin sigui el règim alimentari. En general, els insectes que s’alimenten de líquids tenen dispositius per a concentrar l’aliment; un cas extrem és l’estómac filtrador de molts homòpters. Altres modificacions es relacionen amb l’existència de micetomes a diferents regions; també en són el pap i el pedrer especialitzats dels himenòpters productors de mel, etc. Alguns insectes, com les larves de formiga lleó (neuròpter), i les de molts himenòpters endoparàsits, només eliminen els excrements quan muden, ja que tenen el mesodeu i el proctodeu incomunicats.
L’aparell respiratori
En els insectes, la respiració és traqueal tret d’alguns casos rars, en què és cutània. El sistema traqueal, però, pot presentar modificacions diverses, en relació amb el tipus de vida i el medi, que afecten principalment les espècies de vida aquàtica.
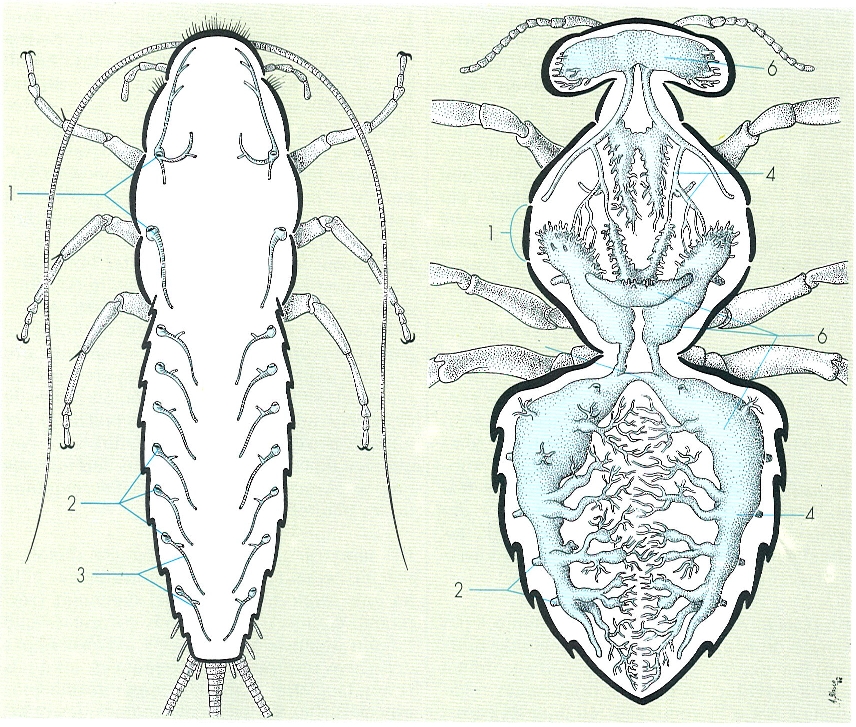
Aparell respiratori dels insectes, en un model primitiu, corresponent a un insecte apterigot (un maquílid, gènere Machilis), a l’esquerra, i a un model evolucionat, d’una abella obrera, a la dreta. Les diferències entre l’un i l’altre afecten el nombre d’estigmes i ta seva posició, l’existència de connectius transversals i longitudinals, i l’aparició de sacs aeris. 1 estigmes toràcics, 2 estigmes abdominals, 3 tràquees metamèriques independents, ramificades, 4 troncs traqueals longitudinals, 5 troncs traqueals transversals, 6 sacs aeris traqueals.
Amadeu Blasco, a partir de fonts diverses
Les tràquees són invaginacions del tegument que es ramifiquen dins del cos; per tant, van recobertes de cutícula, i aquesta cutícula com la de la resta del cos, es renova periòdicament amb la muda. Les tràquees comuniquen amb l’exterior a través dels estigmes o espiracles, que es disposen metamèricament en el tòrax i l’abdomen, i, cap a dins del cos, les seves ramificacions més primes, anomenades traquèoles, són les que penetren fins als teixits i les seves cèl·lules.
Els estigmes o espiracles en els casos més senzills (els apterigots) s’obren directament a les tràquees, però el més corrent és que s’obrin a una cambra, l’atri o cavitat substigmàtica, que en general és esclerificada i és on se situen diferents dispositius (sedes, pèls hidròfugs, etc.) dedicats a evitar la pèrdua d’aigua i l’entrada de partícules. També tenen diferents i variats sistemes de tancament. En general els estigmes són arrodonits i ovals i tenen al seu voltant una anella esclerificada, anomenada peritrema. El nombre d’estigmes és variable: en els pterigots oscil·la des de 10 parells (dos de toràcics i vuit d’abdominals) a cap (quan la respiració és cutània); en els apterigots, des d’un sol parell, situat en el protòrax, a la regió del coll (alguns col·lèmbols), fins a deu parells (en els zigentomes), disposats com en els pterigots; d’altra banda, alguns apterigots tenen respiració cutània.
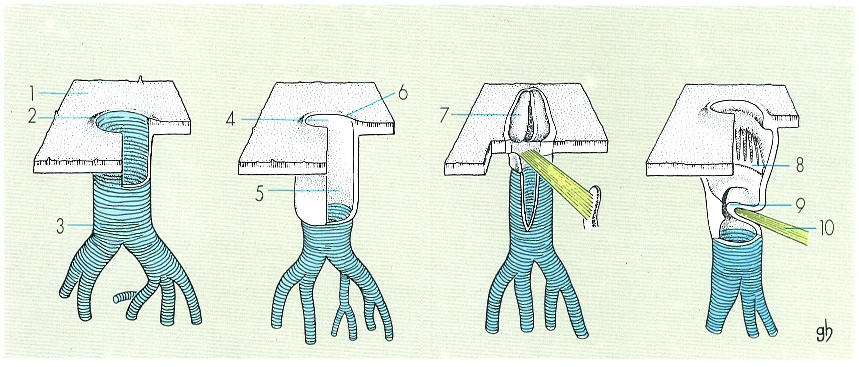
Diferents tipus d’estigmes respiratoris parcialment tallats per tal de fer-ne visible l’estructura interna, i ordenats per ordre de complexitat. 1 paret del cos, 2 orifici traqueal, 3 tràquea, 4 orifici atrial, 5 atri, 6 peritrema, 7 llavis de tancament de l’atri, 8 dispositiu de filtratge d’aire, 9 valva d’oclusió, 10 músculs.
Gustavo Hormiga, a partir d’un original de Snodgrass
Les tràquees, en principi, es disposen per parells, un a cada metàmer, i es mantenen independents. En general presenten tres branques: una de dorsal que va cap al cor i l’aorta, una de visceral que es ramifica entre el tub digestiu, l’aparell reproductor i la resta dels òrgans, i finalment una altra que es dirigeix cap al sistema nerviós. El sistema traqueal, però, augmenta de complexitat des dels insectes apterigots, no alats, fins als pterigots, alats; la disposició metamèrica de les tràquees només es manté en alguns insectes apterigots (proturs, alguns diplurs, etc.), mentre que a la majoria dels insectes les tràquees dels diferents metàmers s’ajunten per mitjà de dos troncs longitudinals dorsals i dos de ventrals que es relacionen amb les branques traqueals procedents de cada metàmer; de vegades hi ha troncs traqueals transversos, que relacionen les branques longitudinals dorsals i les ventrals. Els troncs traqueals, a més d’anar revestits de cutícula, presenten uns engruiximents espiralats, els ctenidis, que impedeixen que s’aixafin. Les tràquees són permeables als gasos (oxigen i diòxid de carboni) en tots els trams, mentre que als líquids només són permeables en els trams més fins (les traquèoles). Per les tràquees circulen els gasos respiratoris. En relació amb les tràquees pot haver-hi unes formacions especials anomenades sacs aeris, que són cambres molt dilatables, que faciliten el vol per tal com actuen com a òrgan hidrostàtic (quan es dilaten fan baixar el pes relatiu de l’organisme); també augmenten la quantitat d’aire que es renova periòdicament i alhora poden actuar com a òrgans amplificadors dels òrgans timpànics.
Les traquèoles, és a dir, les branques més primes de tot el sistema traqueal, poden contenir líquids i aire, i tenen les parets molt fines; en alguns casos són cegues i en d’altres presenten anastomosi. Són formacions intracel·lulars que es desenvolupen, gairebé sempre, a partir d’unes cèl·lules estrellades anomenades traqueoblasts. S’uneixen a les tràquees per mitjà d’una anella de ciment. L’extrem de les traquèoles penetra dins de les cèl·lules dels teixits i és ací que es produeix l’intercanvi de gasos.
En principi, l’aire circula per les tràquees per simple difusió, però en els insectes de mida mitjana o gran, i també en els que tenen molta activitat, hi ha uns mecanismes addicionals per a la ventilació; en aquests casos, hi intervenen els sacs aeris i la musculatura corporal. En la respiració traqueal el transport de gasos respiratoris és totalment independent de l’aparell circulatori.
Els insectes aquàtics presenten modificacions diverses respecte del sistema traqueal ara descrit, que és el més típic dels insectes. En alguns casos, només són capaços de respirar l’oxigen gasós, i per tant, han de sortir periòdicament a l’exterior per a respirar, mentre que en d’altres, poden captar l’oxigen dissolt a l’aigua i tenen dispositius especials per a fer-ho. En el primer cas, tenen dispositius en els estigmes dedicats a evitar que entri aigua dins de l’aparell traqueal, i també mecanismes especials que els permeten guardar bombolles d’aire mentre es mantenen submergits. Molts escarabats d’aigua (coleòpters) i heteròpters respiren per mitjà de l’anomenada brànquia física o brànquia de bombolla, i guarden l’aire sota els èlitres i els hemèlitres, respectivament. Els sifons que mostren moltes larves de dípters és un altre tipus d’especialització per a respirar oxigen atmosfèric i viure sota l’aigua; igualment enginyós és el cas dels insectes que respiren l’aire contingut a l’aerènquima de les plantes submergides.
Els insectes que respiren l’oxigen dissolt a l’aigua presenten diferents estratègies. N’hi ha que respiren a través del tegument (respiració cutània), però el més corrent és que tinguin òrgans especials per a afavorir el pas dels gasos respiratoris a les tràquees, les quals resten tancades. Aquests òrgans o formacions especials són: les traqueobrànquies, les brànquies espiraculars i el plastró. Les traqueobrànquies són extensions digitades o laminars, d’origen tegumentari, que porten a dins les tràquees ramificades i es disposen a diverses regions del cos; són pròpies de les nimfes i larves de molts insectes aquàtics (odonats, efemeròpters, plecòpters, megalòpters, alguns coleòpters i tricòpters), i aconsegueixen la ventilació pel mateix moviment del cos o de les estructures de la traqueobrànquia. Les brànquies espiraculars són una mena de bosses o sacs que se situen als estigmes; són típiques de les pupes de molts dípters i alguns coleòpters. El plastró consisteix en una fina pel·lícula d’aire que es manté recobrint unes determinades parts del cos, especialment els estigmes funcionals, la qual és retinguda per pèls hidròfobs i actua com una superfície d’intercanvi gasós amb l’aigua; el plastró és permanent, de manera que l’insecte que el posseeix no ha de pujar a la superfície.
Els insectes endoparàsits tenen estratègies respiratòries especials, que en alguns casos s’assemblen a les de les espècies aquàtiques. La respiració pot ser cutània, de manera que l’insecte paràsit capta l’oxigen de dins del cos del seu hoste. En altres casos, l’insecte té unes formacions digitades distals, en les quals es fa l’intercanvi de gasos. D’altres fan aquest intercanvi en el proctodeu. Finalment, algunes larves i pupes capten l’oxigen aeri per mitjà de formacions capaces de perforar la paret del cos de l’hoste, o fins i tot les tràquees de l’hoste quan aquest és un artròpode traqueat.
L’aparell circulatori
La circulació dels insectes, com la de la resta d’artròpodes, és oberta i lacunar. El líquid circulatori, l’hemolimfa, omple la cavitat general del cos o hemocel, la qual sol trobar-se dividida tot al llarg en tres sins, separats per dos diafragmes: el si pericàrdic, en posició dorsal, el perivisceral, en posició central, i el perineural, en posició ventral.
L’aparell circulatori dels insectes és simplificat. Es compon d’un cor dorsal i un sistema de vasos. El cor en les formes més primitives és llarg i s’estén per tot el tòrax i l’abdomen, però el més corrent és que sigui curt i ocupi només una part d’aquest. El cor queda situat dins del si dorsal o pericàrdic i té una vàlvula interna i una constricció externa a cada metàmer; així es delimiten unes cavitats metamèriques: els ventricles. Cada ventricle té un parell de forats, els ostíols, per on l’hemolimfa entra al cor. D’altra banda, hi ha els músculs alars, també metamèrics, que són responsables dels moviments del cor. El cor en general és tancat pel darrere mentre que pel davant es perllonga en una artèria, l’aorta, que es pot ramificar i que dirigeix l’hemolimfa vers la regió cefàlica. En alguns casos també hi ha uns vasos laterals parells que surten dels ventricles. Quan el cor es contrau (sístole) l’hemolimfa en surt per l’aorta, i va a parar a l’hemocel; després, retorna al si pericàrdic i al cor a través dels ostíols quan aquest es dilata (diàstole). En alguns casos, el cor té uns pors exhalants pels quals l’hemolimfa en surt durant la sístole. També pot haver-hi òrgans pulsàtils accessoris, en forma de sac i situats a diferents regions del cos, que actuen com a cors pulsàtils; es contrauen independentment del cor i la seva funció és la d’assegurar la circulació de l’hemolimfa a les ales i els apèndixs (les potes i les antenes).
L’hemolimfa pot ésser de colors diferents o, fins i tot, incolora. Els pigments que porta generalment procedeixen de l’alimentació o dels processos metabòlics, i no solen tenir cap significat biològic, ja que el transport dels gasos respiratoris és independent de l’aparell circulatori. És per això que l’hemolimfa dels insectes no porta pigments respiratoris tret d’alguns casos excepcionals, com el cas dels insectes que tenen respiració cutània. L’hemolimfa porta cèl·lules sanguínies o hemòcits de diferents tipus i funcions.
Els òrgans excretors
Els insectes excreten a través dels tubs de Malpighi, però poden haver-hi altres formacions que complementin aquesta funció. Els tubs de Malpighi es localitzen en el límit entre el mesodeu i el proctodeu, al tub digestiu, i tenen forma de tubs tancats per l’extrem, que suren a l’hemocel. El nombre de tubs de Malpighi oscil·la entre quatre i sis, nombre típic de la majoria dels insectes oligoneòpters i paraneòpters, fins a unes quantes desenes i més d’un centenar, com passa en els microcorifis, els paleòpters, molts polineòpters i els himenòpters; en alguns casos (com alguns apterigots i els àfids, per exemple) no n’hi ha, i llavors l’excreció es fa per mitjà d’altres òrgans. La funció dels tubs de Malpighi és captar els productes residuals que porta l’hemolimfa i abocar-los al tub digestiu, des d’on són expulsats a l’exterior del cos; d’altra banda, participen en la regulació osmòtica i iònica per mitjà de la reabsorció d’aigua i de ions i la concentració dels productes que cal excretar.
Els insectes, la majoria terrestres, en general disposen de poca aigua i, per tant, eliminen els productes nitrogenats en forma d’àcid úric principalment. Una de les especialitzacions dels tubs de Malpighi, que té relació amb la forma d’excreció, és el criptonefridisme, pel qual els extrems cecs dels tubs són adossats a les parets del recte i recoberts per una membrana perinèfrica; això afavoreix la reabsorció d’aigua, i per això es considera una adaptació als ambients o als aliments molt secs. Els tubs de Malpighi poden fer altres funcions no excretores: en alguns casos acumulen substàncies determinades, o actuen com a glàndules productores de seda (en algunes larves) o, fins i tot, fan d’òrgans productors de llum.
Alguns insectes tenen òrgans excretors independents del tub digestiu; són els anomenats ronyons cefàlics o glàndules labials o maxil·lars, segons la seva situació al cos. Són d’origen metamèric i consten d’un sac i un conducte mitjà (d’origen mesodèrmic) i d’un conducte excretor final (d’origen ectodèrmic), que desemboca a la base del llavi o de les maxil·les. Alguns autors interpreten les glàndules salivals dels insectes pterigots com a òrgans excretors modificats.
També hi ha diversos tipus de ronyons d’acumulació, com els cossos pericàrdics, situats a banda i banda del cor, i els nefròcits, que es troben dispersos per l’hemocel i acumulen productes residuals captats per l’hemolimfa. Aquests productes també poden ésser acumulats a l’epidermis (en els oenòcits) i a determinades cèl·lules del mesodeu. L’anomenat cos adipós té com a funció principal acumular productes de reserva, especialment glucogen i proteïnes, en els adipòcits o trofòcits, i productes residuals a les cèl·lules de l’urat. El cos adipós actua, doncs, com un ronyó d’acumulació; també conté els micetòcits o els bacteròcits, és a dir, cèl·lules que porten fongs o bacteris simbionts, que de vegades s’agrupen i formen els micetomes o els bacteriomes, de funció molt discutida; pot ésser que facilitin la digestió de determinats aliments, com per exemple la fusta, o bé que siguin sintetitzadors de vitamines i d’altres elements indispensables que els insectes són incapaços de fabricar i d’obtenir a partir dels aliments. Moltes vegades aquests simbionts es transmeten regularment als descendents per via genital (amb la posta).
L’aparell reproductor
Tret d’alguns casos excepcionals, els insectes són unisexuats, és a dir, hi ha mascles i femelles, i moltes vegades mostren un dimorfisme sexual accentuat, que es manifesta en el desenvolupament i la forma del cos i dels apèndixs, o en els colors i la grandària del cos; aquest dimorfisme també pot afectar el comportament ecològic o alimentari.
L’aparell reproductor es compon d’un parell de gònades (testicles en els mascles i ovaris en les femelles) i un parell de conductes (espermiductes i oviductes respectivament) de natura mesodèrmica, que conflueixen en un conducte imparell d’origen ectodèrmic (el canal ejaculador en els mascles i l’oviducte imparell i la vagina en les femelles); hi ha, a més, diverses glàndules annexes. En relació amb aquest aparell descrit, que és intern, els insectes tenen allò que podríem anomenar aparell genital extern o genitàlia, de natura apendicular, que serveix per a assegurar la transferència de l’esperma i la subjecció de la femella durant la còpula, en el cas dels mascles, i en el de les femelles afavoreix la posta dels ous a través d’estructures adients (ovipositor, oviscapte, etc.).
L’orifici genital se situa en el novè metàmer de l’abdomen en els mascles, i entre el vuitè i el novè en les femelles. Com que les gònades són d’origen metamèric, en alguns casos (alguns apterigots) encara es conserven restes d’aquesta metameria primitiva, i hi ha diversos parells de gònades que desemboquen a conductes genitals parells. En d’altres (per exemple, els efemeròpters), no hi ha conducte genital imparell, de manera que presenten dos orificis genitals. Algunes femelles de lepidòpters tenen dos orificis genitals, un per a la còpula i l’altre per a la posta, encara que el més corrent és que tant el mascle com la femella en tinguin un de sol.
Els testicles generalment són ovoides i són formats per un nombre variable de lòbuls o fol·licles, recoberts d’una membrana peritoneal. La forma dels fol·licles és variable i cadascun es connecta a l’espermiducte a través d’un canal curt o vas eferent, o bé per un tub testicular únic (l’estructura dels testicles en els apterigots difereix del model que hem comentat). En els fol·licles és on es formen els espermatozoides a partir de les espermatogònies o cèl·lules germinals primordials. Els espermiductes o vasos deferents parells varien molt de llargada a cada grup d’insectes; generalment cada espermiducte porta un eixamplament al llarg del seu recorregut per a formar la vesícula seminal, que de vegades és d’estructura molt complexa. A la vesícula seminal s’acumulen els espermatozoides, mòbils, des que es formen fins que són transferits a la femella. Finalment, els espermiductes es reuneixen, tret d’algunes excepcions, en un únic canal imparell, que constitueix el canal ejaculador, al final del qual hi ha la genitàlia externa o òrgan copulador. En relació amb les vies genitals hi ha d’un a tres parells de glàndules accessòries, les secrecions de les quals es barregen amb els espermatozoides i es relacionen amb la formació dels espermatòfors, i encara, en algun cas, poden actuar sobre la femella reduint la seva receptivitat en posteriors còpules o estimulant l’oviposició.
Els ovaris, en general, són dues masses compactes, formades per la reunió de diversos tubs ovàrics o ovarioles independents, que desemboquen a l’oviducte. Tot el conjunt queda embolicat per una membrana. En alguns casos les ovarioles no es diferencien sinó que els ovaris tenen forma de sac. D’altra banda, en alguns apterigots (per exemple, Japyx), les ovarioles conserven la seva disposició metamèrica primitiva. A les ovarioles se situen els òvuls en període de formació i les cèl·lules vitel·lines, quan n’hi ha. A cada ovariola es distingeixen les regions següents: el filament terminal, que és un cordó que uneix l’ovariola a la paret del cos i al diafragma dorsal (el conjunt de filaments terminals forma el lligament suspensor); el germari, situat a l’àpex, on es localitzen les cèl·lules germinals primordials i, en molts casos, les cèl·lules nutritives; el vitel·lari, que és la part principal de l’ovariola, on se situen els ovòcits en fase de vitel·logènesi, cadascun envoltat d’una coberta que delimita la cambra ovular o fol·licle; i finalment, el pedicel·le o peduncle, que connecta l’ovariola a l’oviducte. Les ovarioles poden ésser de dos tipus: panoístic i meroístic. A les de tipus panoístic totes les cèl·lules germinals es transformen en ovòcits; el vitel·lus penetra a l’òvul a través de les cèl·lules fol·liculars, que el capten de l’hemolimfa. El tipus meroístic es caracteritza perquè una part de les cèl·lules germinals es transformen en cèl·lules abortives que actuen com a trofòcits o cèl·lules nutritives. Segons quina sigui la situació de les cèl·lules vitel·lines, hi ha dos tipus d’ovarioles meroístiques: telotròfiques i politròfiques; en les ovarioles telotròfiques, també anomenades acrotròfiques, les cèl·lules nutritives se situen en el germari i estenen ponts citoplasmàtics o cordons tròfics amb els ovòcits; en les ovarioles politròfiques els trofòcits es desplacen amb l’ovòcit en formació, ja que tots dos són inclosos dins del mateix fol·licle.
El procés de la vitel·logènesi és regulat hormonalment. Quan el desenvolupament de l’òvul queda completat, aquest es cobreix amb l’embolcall vitel·lí i del còrion, que és format per diverses capes (entre dues i set), la més externa de les quals presenta ornamentacions i consistència variables; aquesta coberta evita la pèrdua d’aigua, permet el pas dels gasos respiratoris i la fecundació de l’òvul, al pol anterior, on és més prima i pot presentar petits orificis (micròpils) adients. Els ous madurs són alliberats de l’ovari i s’acumulen temporalment als conductes eferents abans no es produeix la posta.
Els conductes genitals parells o oviductes parteixen dels ovaris i generalment acaben confluint a la part final, on formen l’oviducte imparell i la vagina. En alguns casos, a la vagina hi ha la bossa copuladora, que pot comunicar directament a l’exterior, cosa que passa en algunes papallones; en alguns xilòfags (anòbids) hi ha bosses relacionades amb la vagina, on se situen fongs simbionts (micetomes) que es transmeten a la posta; en altres casos hi ha una o dues espermateques, que són reservoris on s’acumulen els espermatozoides després de la còpula, en espera que madurin els òvuls. A la part terminal de la vagina solen desembocar diverses glàndules, que participen en la formació dels embolcalls protectors dels ous i dels estoigs especials o ooteques, o bé segreguen substàncies mucoses, gelatinoses, que en contacte amb l’aire se solidifiquen i permeten la fixació de la posta.
La genitàlia externa de les femelles pren la forma d’òrgans de posta (ovipositor, oviscapte, etc.), la forma i el desenvolupament dels quals és en funció del tipus de substrat on facin la posta. Moltes vegades, però, la genitàlia és poc desenvolupada.
L’estratègia biològica dels insectes
La reproducció i el desenvolupament
Cicles biològics dels insectes, de tipus ametàbol (a dalt), en el cas d’un protur, de tipus heterometàbol (al mig), en el cas d’un dictiòpter, i de tipus holometàbol (a baix), en el cas d’un dípter. En el cas dels proturs, el desenvolupament és anamorf: van adquirint el nombre de segments del cos típic dels adults d’una manera progressiva. Hom ha indicat en groc les fases immatures i en gris els adults. 1 ous, 1’ ooteca, 2 nimfa, 3 larva, 4 pupa.
Servei de Fotografia/Román Montull, a partir de fonts diverses
Tot i que en general els insectes, com ja hem dit, són unisexuats, hi ha alguns casos d’espècies hermafrodites. L’hermafroditisme pot ésser funcional: és el cas de la cotxinilla Icerya purchasi, espècie a la qual s’ha observat l’autofecundació; en altres casos l’hermafroditisme no és funcional, com passa a Perla marginata, els mascles de la qual tenen un ovari connectat als testicles però els òvuls no arriben a formar-s’hi totalment.
La reproducció generalment és sexual i per oviparisme, és a dir, amb producció d’ous. No obstant això, hi ha determinades espècies que es poden reproduir partenogenèticament, i també trobem exemples d’ovoviviparisme i fins i tot de viviparisme. Així mateix hi ha casos entre els insectes de pedogènesi i de poliembrionia.
La partenogènesi és un tipus de reproducció per la qual els ous no fecundats experimenten l’embriogènesi i donen lloc a nous individus. Pot ésser de dos tipus: arrenotòcica i telitòcica. La partenogènesi arrenotòcica (arrenotòcia) constitueix un mecanisme determinant del sexe; quan les femelles ponen ous fecundats (per tant, diploides), aquests ous donen lloc a femelles, i quan els ous no han estat fecundats (són haploides), es desenvolupen partenogenèticament i donen lloc a mascles. En la partenogènesi telitòcica (telitòcia), només s’originen femelles. La partenogènesi pot ésser accidental o facultativa, de manera que s’alterna amb generacions sexuals, o bé obligada, ja que no hi ha mascles; d’altra banda, pot tractar-se de partenogènesi geogràfica.
La pedogènesi o paidogènesi és la capacitat d’adquirir la facultat reproductora en l’estat larval. En els insectes únicament es coneixen femelles pedogenètiques, que a més es reprodueixen partenogenèticament, i que, en molts casos, també són vivípares.
La poliembrionia és el fenomen pel qual a partir d’un ou s’originen diversos embrions, de manera que augmenta el potencial reproductor de les espècies. És conegut en molts insectes paràsits i parasitoides, i de vegades, va lligat a la pedogènesi.
L’ovoviviparisme es dona quan els ous es desenvolupen dins del cos de la mare, però a expenses del vitel·lus que contenen ells mateixos. Aquest procés de vegades ha estat confós amb el viviparisme, procés pel qual els embrions es desenvolupen a partir de l’aliment que els subministra la mare. Hi ha diferents tipus de viviparisme (adenotròfic, hemocèlic i pseudoplacentari) en funció del lloc on es localitzen els embrions i de la manera com reben l’aportació d’aliment matern.
La fecundació
La reproducció sexual requereix la fecundació dels òvuls i els espermatozoides. La fecundació sempre és interna en els insectes i es fa a les vies genitals femenines, bé que la transferència de l’esperma pot ésser indirecta o directa. En el primer cas, el mascle diposita l’esperma o els espermatòfors en el substrat i, posteriorment, la femella se’ls introdueix a la vagina, de vegades induïda pel mascle; en les espècies més evolucionades, el mascle només diposita espermatòfors en presència de la femella i, fins i tot, pot haver-hi un festeig previ; és el cas típic dels insectes apterigots. En la transferència directa cal que el mascle i la femella es trobin i que tingui lloc l’aparellament o còpula; en aquest cas, el mascle té els òrgans copuladors connectats a les vies genitals i els fa servir per a introduir l’esperma a la vagina de la femella durant la còpula. Aquest tipus de transferència és el típic dels insectes alats (pterigots), si bé també hi ha excepcions, com en els odonats, en què els mascles tenen els òrgans copuladors independents de l’orifici genital i els utilitzen per a recollir l’esperma que introdueixen posteriorment a la femella. Moltes vegades abans de la còpula hi ha un festeig, que pot arribar a ésser molt sofisticat. La missió del festeig és augmentar la receptivitat de la femella i calmar els seus instints agressius, i de vegades els mascles porten regals a la femella per a evitar ésser devorats després de la còpula.
Els estímuls que permeten que el mascle i la femella es trobin i es reconeguin són olfactius (feromones), fònies (sons) i visuals. Els més estesos són els olfactius; solen ésser les femelles les qui produeixen les feromones que atreuen els mascles des de grans distàncies, bé que també hi ha alguns casos en què els mascles emeten feromones sexuals. Els estímuls audibles són menys estesos i consisteixen en l’emissió de sons que són reconeguts pels individus del sexe contrari. Els estímuls visuals poden consistir a emetre llum, que és reconeguda pels congèneres, o en el reconeixement del color i la forma, tant la de l’insecte com la de determinats relleus de terreny que atreuen mascles i femelles i per tant, faciliten la seva coincidència a determinats punts.
Els ous que es formen per la fusió de l’òvul i l’espermatozoide, amb l’acúmul del vitel·lus corresponent, són embolicats per diverses capes de les quals la més externa presenta ornamentacions diverses. La forma dels ous és molt variable, i de vegades tenen diferenciacions especials, com l’opercle, que és el punt per on en surten les formes juvenils. El desenvolupament embrionari pot començar després de la posta o bé abans. Els ous dels insectes en general són rics en vitel·lus i de tipus centrolecític, amb segmentació parcial i superficial; alguns són pobres en vitel·lus i llavors la seva segmentació és total, i hi ha nombrosos casos de transició entre l’un i l’altre tipus.
La posta
La posta dels ous pot ésser immediata a la còpula o produir-se al cap d’un temps. Per tal que s’inicïi calen unes determinades condicions fisiològiques i ambientals: una dieta alimentària especial, condicions de temperatura, humitat i claror, etc. De vegades les femelles dipositen els ous aïlladament damunt del substrat, d’altres els deixen agrupats i els fixen per mitjà d’un peduncle o de substàncies adherents. En general, la femella escull el substrat que sigui adequat per a afavorir el posterior desenvolupament de les formes juvenils. El nombre de postes que pot fer una femella és variable, des d’una —cas més normal— fins als valors màxims que es donen entre els tèrmits, als quals les femelles ponen ous durant tota la vida, que acostuma a durar uns quants anys. El nombre d’ous de cada posta també és variable, amb màxims, també corresponents als tèrmits, de fins a 7000 ous diaris.
Hi ha molts insectes que protegeixen la posta, tot situant-la dins d’estructures especials, les anomenades ooteques; així mateix es considera una protecció el fet d’untar els ous amb substàncies pudents o mucilaginoses, i també la seva duresa. Les postes endofítiques (que es deixen dins dels vegetals) i les que es fan en cambres construïdes pels progenitors, dins de les quals han col·locat aliment, també es consideren mecanismes de protecció de la posta. De vegades els pares, o bé només la mare, tenen cura dels ous fins al moment d’emergència dels joves, i, fins i tot, els vigilen fins que arriben a un estat més avançat de desenvolupament; aquest fenomen és conegut en alguns insectes gregaris i en tots els insectes socials. Finalment, també hi ha casos en què la femella pon els ous a l’atzar, fins i tot durant el vol.
El estats juvenils
L’insecte que emergeix de l’ou quan ja ha acomplert el seu desenvolupament embrionari en general difereix de l’adult no solament en les dimensions del cos i per la seva incapacitat reproductora, sinó també en molts altres caràcters, tant morfològics com referents als seus hàbits ecològics i alimentaris. El temps que passa entre el naixement dels joves i l’adquisició de la capacitat reproductora (adults) comprèn allò que es coneix amb el nom de desenvolupament postembrionari. En general, al llarg del desenvolupament postembrionari els organismes muden diverses vegades per a créixer i experimenten canvis successius, que en conjunt es coneixen amb el nom de metamorfosi. Excepcionalment, alguns insectes (apterigots) tenen desenvolupament directe però tota la resta tenen metamorfosi, sigui simple o complicada.
Formes juvenils dels insectes. En general hom aplica el nom de nimfa a les fases juvenils dels insectes amb metamorfosi simple, i de arva i pupa a les fases mòbils i immòbils dels cicles amb metamorfosi complicada. La figura recull les formes juvenils següents: A nimfa d’ortòpter; B larva protopòdica d’himenòpter, molt modificada com a consegüència de dur una vida endoparàsita; C larva polipódica de lepidòpter, l’anomenada larva eruciforme o eruga, característica pels tres parells de potes toràciques i les falses potes o pseudopotes abdominals; D larva oligopòdica de tipus melolontiforme o escarabeïforme, mancada de potes abdominals, d’un coleòpter; E larva oligopòdica de tipus campodeïforme, també sense potes abdominals, d’un coleòpter; F larva oligopòdica àpoda i eucèfala d’un dípter aquàtic (mosquit), mancada de potes toràciques i amb el cap ben desenvolupat; G larva oligopódica àpoda i acèfala, d’un dípter (mosca), amb el cap reduït; H pupa exarata, amb els esbossos de les ales visibles i els apèndixs totalment independents del cos, d’un coleòpter; I pupa obtecta, amb les ales i els apèndixs soldats al cos i recoberta d’una closca protectora o pupari, que correspon a la darrera exúvia larval modificada, d’un dípter.
Servei de Fotografia/Román Montull, a partir de fonts diverses
La classificació més senzilla dels tipus de desenvolupament postembrionari distingeix els insectes ametàbols—amb formes juvenils molt semblants a l’adult i amb la possibilitat de mudar en aquesta darrera fase— dels insectes heterometàbols—amb fases de nimfa i adult o imago— i els insectes holometàbols—amb fase de larva, pupa i adult o imago. Hi ha altres classificacions més modernes i més completes, les quals, però, resulten més difícils d’aplicar.
En el cas dels heterometàbols, hi ha dos tipus de desenvolupament, segons els quals hom parla d’insectes hemimetàbols, quan les nimfes i els adults tenen hàbitats diferents, o paurometàbols, quan tenen el mateix hàbitat. En aquest cas, tots els estadis preimaginals són semblants, tant pel seu aspecte com pels seus hàbits ecològics, i reben el nom de nimfes. Les nimfes van adquirint els caràcters dels adults d’una manera progressiva.
En els insectes holometàbols se succeeixen dos tipus d’estadis preimaginals: el de larva i el de pupa, que és un estadi intermedi entre la fase de larva i la d’adult. Les larves són estadis preimaginals molt diferents de l’adult, tant pel que fa al seu aspecte com als seus hàbits. El pas de larva a adult requereix tota una sèrie de canvis importants que s’esdevenen en la fase de pupa, la qual és immòbil i àfaga. En aquests canvis es combinen fenòmens de formació dels òrgans de l’adult a partir de cèl·lules indiferenciades que formen els discs imaginals (histogènesi), de degeneració dels òrgans típicament larvals (històlisi), i de conservació, amb algunes modificacions, de determinats òrgans larvals. Un tipus especial d’holometabolia és la hipermetabolia, que es caracteritza per la successió de diversos tipus de larves amb característiques diferents abans d’arribar a la fase de pupa. La hipermetabolia no és molt freqüent; es presenta en els estrepsípters i en alguns coleòpters i himenòpters, entre altres.
Les nimfes dels insectes heterometàbols generalment s’assemblen força als adults, bé que en les primeres fases no tenen encara els esbossos de les ales; en els hemimetàbols, però, com ja hem dit, difereixen molt en els hàbits, ja que les nimfes solen ésser aquàtiques i tenen tota mena d’adaptacions a aquest medi de vida, mentre que els adults són terrestres. En els insectes apterigots, no alats, cap de les formes juvenils no té ales i s’assemblen força als adults.
Les larves dels insectes holometàbols difereixen en molts aspectes dels adults. Segons el nombre d’apèndixs que tenen, en distingim tres tipus principals: larves protopòdiques, larves polipòdiques i larves oligopòdiques, i dins de cada tipus hi ha encara nombroses variacions.
Les larves protopòdiques són característiques de molts himenòpters endoparàsits i d’alguns dípters, principalment; neixen en un estadi precoç de desenvolupament i només tenen els esbossos dels apèndixs del cap i del tòrax, els manquen les tràquees i gairebé les vísceres. Aquestes larves, també anomenades ciclopodiformes, eclosionen a l’interior de l’animal o planta parasitat o en medis nutricis especials.
Les larves polipòdiques tenen tres parells de potes toràciques i un nombre variable de potes abdominals; generalment són molt actives, i corresponen al model típic de les erugues de papallona i d’alguns himenòpters, també anomenades larves eruciformes, que sempre tenen el cos cilíndric i allargat.
Les larves oligopòdiques no tenen apèndixs abdominals, si bé algunes poden tenir cercs al final del cos i també traqueobrànquies, que són apèndixs respiratoris; en funció de l’estructura del cap i de les potes, i de la seva capacitat de moviment, en distingim tres tipus diferents: les campodeïformes, les melolontiformes i les àpodes. Les larves campodeïformes tenen el cos allargat i deprimit i tres parells de potes toràciques desenvolupades; són molt actives i es mouen ràpidament. Tenen peces bucals mastegadores, ja que són depredadores, i generalment porten cercs abdominals i tenen el tegument esclerotitzat. Aquest tipus de larves és típic de la majoria dels coleòpters i de molts neuropteroïdeus entre altres. Les larves melolontiformes o escarabeïformes són robustes, de cos corbat i tegument més aviat tou, amb el cap ben diferenciat i les peces bucals mastegadores; tenen potes toràciques però els són molt poc útils per a desplaçar-se; amb l’abdomen molt voluminós, solen viure enterrades i són vegetarianes. Són característiques d’alguns coleòpters i de certs himenòpters. Les larves ápodes o vermiformes no tenen apèndixs toràcics i pertanyen a diferents famílies de coleòpters, dípters i alguns himenòpters; segons quin sigui el grau de desenvolupament del cap, n’hi ha de tres tipus: larves eucèfales, que tenen el cap esclerotitzat i les peces bucals poc reduïdes; larves hemicèfales, amb la càpsula cefàlica i els apèndixs corresponents reduïts; i larves acèfales, amb la càpsula cefàlica totalment reduïda, les restes de la qual formen, juntament amb les peces bucals, un aparell bucofaringi que intervé en l’alimentació.
Les pupes són exclusives dels insectes holometàbols i poden ésser de dos tipus; en un cas, els apèndixs i els esbossos alars queden separats del cos, i reben el nom de pupes exarates; en l’altre, el de les pupes obtectes, els apèndixs i els esbossos de les ales són enganxats al cos per una secreció que s’aboca durant la muda de la pupa. Un altre caràcter que hom fa servir per a classificar les pupes és la seva capacitat de moure les mandíbules (pupes dèdiques) o la seva incapacitat de ferho (pupes adèctiques). Les pupes amb els apèndixs i els esbossos alars separats del cos i amb les mandíbules mòbils corresponen als rafidiòpters, planipennis, mecòpters, tricòpters i alguns lepidòpters, entre altres. Les pupes, també amb apèndixs i esbossos alars lliures però amb mandíbules immòbils, són característiques d’alguns dípters, sifonàpters i de molts coleòpters i himenòpters; finalment, les pupes amb els apèndixs i els esbossos alars enganxats al cos i les mandíbules immòbils són característiques de molts dípters i coleòpters i d’alguns planipennis i himenòpters. Les pupes generalment són immòbils completament, i no s’alimenten en absolut. És per aquesta raó que la majoria van protegides amb diferents cobertes, ja que són molt vulnerables als seus depredadors; únicament algunes pupes exarates i dèctiques poden fer petits moviments amb el cos i el primer parell de potes. Generalment les pupes se situen en càpsules subterrànies, dins de capolls de seda o d’altres substàncies que la mateixa larva fabrica en el seu darrer estadi, o també a l’interior de cel·les, d’excrements, de cadàvers o en el cos d’un hoste, en el cas de les espècies paràsites. En altres casos, les pupes adquireixen aspectes homòcroms o críptics.
Finalment, alguns dípters tenen pupari, que correspon a l’exúvia (cutícula o exosquelet que es desprèn a la muda) endurida de la darrera fase larval, que no s’arriba a desprendre, i dins de la qual es desenvolupa la pupa. El terme de prepupa no es refereix a aquesta fase sinó a la darrera fase larval d’alguns insectes, que és lleugerament diferent a la resta de fases i que és quiescent.
Les fases de repòs
En els insectes es pot produir un atur del creixement i el desenvolupament vital, fenomen que es coneix amb els noms de quiescència i diapausa. La quiescència és una detenció del creixement a causa de condicions ambientals desfavorables, que es produeix sobretot en els adults; si no es donen aquestes males condicions no es produeix la quiescència, i en el moment en què les condicions es tornen favorables, els organismes retornen a la seva vida activa. La diapausa és l’atur del desenvolupament a causa de factors interns, i és un fenomen regulat per hormones, que es pot produir en la fase d’embrió, en diferents estadis juvenils (de larva, nimfa o pupa) o en l’estat d’adult; moltes vegades el factor que desencadena la diapausa és ambiental, i els organismes tornen a la vida activa després de passar un període prestablert.
El polimorfisme
Molts insectes són polimòrfics, o sigui, una mateixa espècie pot presentar aspectes diferents. El polimorfisme més habitual és el corresponent al dimorfisme sexual, és a dir, el que afecta els mascles i les femelles de la mateixa espècie, que de vegades és molt accentuat. Un cas extrem és el dels lampírids o cuques de llum (coleòpters), en els quals els mascles tenen l’aparença normal de coleòpters, mentre que les femelles tenen aspecte de larva (neotènia).
També solen presentar polimorfisme les espècies que tenen diverses generacions anuals, de manera que hom distingeix entre les generacions de primavera, les d’estiu, etc., per la mida, el color o altres caràcters dels individus. Aquest tipus de polimorfisme es presenta en alguns lepidòpters i altres insectes.
Hi ha un altre tipus de polimorfisme que afecta el desenvolupament de les ales, que pot ser diferent en els diversos individus; així, hom distingeix entre individus àpters o no alats, micròpters o individus d’ales petites, i macròpters o individus d’ales grans. Aquest tipus de polimorfisme és present en moltes espècies d’heteròpters i homòpters, entre molts altres grups.
En els insectes socials és on el polimorfisme és més accentuat i més constant. Hom distingeix, dins d’una mateixa espècie, distintes castes, és a dir, diversos tipus d’individus que tenen aspectes diferents i que fan funcions especials a la colònia.
El vol
L’aerodinàmica del vol en els insectes és molt complicada. Depèn de la creació d’una zona de baixa pressió per davant i per sobre de l’insecte i d’una altra zona d’alta pressió per darrere i per sota. Aquests efectes s’aconsegueixen per mitjà del moviment de les ales. Alhora, és el resultat de les forces d’empenyiment i de gravetat i de la resistència de l’aire.
La disposició dels nervis longitudinals fa que la part anterior de les ales sigui rígida i dura mentre la posterior es manté tova i flexible; la rigidesa d’aquesta zona anterior és indispensable per a vèncer la resistència de l’aire. D’altra banda, abans d’iniciar el vol, els músculs que mouen les ales han d’atènyer una determinada temperatura i, per tal d’aconseguir-la, n’hi ha que les fan vibrar abans, encara parats damunt del substrat, mentre que d’altres simplement resten immòbils una estona al sol, per tal d’augmentar la temperatura corporal.
En general, durant el vol, mouen les ales d’un mateix costat del cos de manera que aquestes descriuen una figura en forma de 8, fent girar la superfície articular i batent-les periòdicament. Els insectes de vol més perfeccionat són capaços de volar cap endavant i cap enrere. A més, la diferència d’amplitud entre els batecs de les ales d’una banda i de l’altra és la que els permet girar de costat.
Els moviments de les ales es deuen a l’existència d’una musculatura toràcica potent. Els músculs són de dos tipus: indirectes i directes. Els indirectes són músculs longitudinals dorsals i músculs parells dorsiventrals; mai no s’insereixen a les ales, sinó que aquestes es mouen per les contraccions i dilatacions del metàmer. Els músculs directes s’insereixen a la regió axil·lar de les ales i a la regió esternal del metàmer corresponent i n’hi ha dos parells: els externs i els interns; la seva contracció fa que les ales pugin i baixin, respectivament. A més hi ha encara altres músculs que permeten el moviment horitzontal de les ales cap enrere, i fan que en la posició de repòs les ales anteriors cobreixin les posteriors, i fins i tot n’hi ha d’altres que permeten que les posteriors es pleguin sota les anteriors.
La velocitat de vol és variable i és en relació amb la forma de les ales i, especialment, amb l’anomenada elongació alar, que és el quocient entre la llargada alar màxima i la superfície alar; quan aquest valor és baix, el vol és lent i més aviat pesant (per exemple, en les llagostes, moltes papallones i altres grups d’insectes) i quan és alt, el vol és ràpid (en els mosquits, les abelles i vespes, les libèl·lules, etc.). També influeix en la velocitat de vol la freqüència dels batecs que, al seu torn, depèn de l’estructura del pterotòrax i del tipus de músculs que el regulen. La freqüència pot oscil·lar entre u i més de mil cicles per segon, bé que generalment se situa entre els vint i els dos-cents. A les ales que tenen freqüències de batec baixes els músculs responsables són sincrònics, és a dir, com els de la resta del cos; quan les freqüències de batec són altes, per contra, els músculs de vol són asincrònics, és a dir, que es contrauen amb una freqüència més alta que la de conducció nerviosa dels estímuls. En conjunt, la velocitat de vol a què poden arribar els insectes pot oscil·lar entre uns pocs quilòmetres per hora i fins als 60 o més quilòmetres per hora. La capacitat de vol sovint s’aparella a una gran capacitat de resistència, que permet a les espècies migradores córrer distàncies molt grans. Hi ha insectes que volen planant amb les ales immòbils, mentre que la majoria les mouen activament i poden mantenir el cos totalment immòbil a l’espai. Quan el primer parell d’ales perd la facultat de volar, pot prendre una funció estabilitzadora, que és el cas dels èlitres dels escarabats (coleòpters). En els dípters, el segon parell d’ales és transformat en balancins, que actuen com a òrgans d’equilibri. L’orientació durant el vol és deguda a estímuls visuals, però també a òrgans sensorials que porten a les antenes i als balancins (aquests, només presents en dípters i estrepsípters).
Tot i que la funció principal de les ales és la de volar, el seu moviment rítmic pot ajudar, a més, a augmentar la temperatura corporal. D’altra banda, les ales poden jugar també un paper determinant en relació amb la reproducció, atès que moltes vegades el seu moviment produeix un so que actua de reclam per als individus del sexe oposat.
Les relacions tròfiques
Els insectes, com tota la resta d’animals, estableixen relacions amb els organismes que viuen en el seu mateix ecosistema. De totes, les relacions tròfiques o alimentàries són les més conegudes.
Les espècies herbívores o fitòfagues s’alimenten de vegetals, mentre que les carnívores exploten altres espècies animals. En qualsevol cas hi ha especialitzacions al tipus d’alimentació i successions més o menys clares. Les espècies sapròfagues o descomponedores s’alimenten de matèria orgànica animal o vegetal en descomposició; de vegades utilitzen aquesta matèria com a aliment, però en molts casos s’alimenten dels organismes que viuen a dins, de manera que també hi ha unes successions establertes.
Les relacions tròfiques primàries
El règim alimentari dels insectes és molt variat, tant que es fa difícil d’establir una classificació. A grans trets, hom diferencia entre els omnívors, els fitòfags, els zoòfags i els sapròfags, però hi ha encara un gran nombre d’especialitzacions, com els que s’alimenten de cera, de pells, de banyes, de plomes, de teles o, fins i tot, de petroli. En molts casos no es pot parlar d’un règim estricte i s’observa el pas d’un a l’altre: el canibalisme d’algunes espècies fitòfagues és l’inici d’un règim sapròfag o carnívor. D’altra banda, de vegades el règim alimentari varia en els dos sexes; en els mosquits, els mascles són fitòfags i les femelles són hematòfagues i fitòfagues. Altres vegades, són les larves les qui tenen una dieta diferent de la dels adults. I, independentment del tipus de dieta, hi ha insectes que són molt estrictes en els seus requeriments alimentaris (monòfags), com per exemple els fitòfags que sempre s’alimenten de la mateixa planta, els paràsits que sempre volen el mateix hoste, etc., i d’altres (polífags) que en són poc, com els fitòfags que s’alimenten de qualsevol planta, els carnívors, etc.
Els insectes omnívors són capaços d’utilitzar com a aliment tota mena de substàncies animals i vegetals. Els fitòfags s’alimenten de productes vegetals i moltes vegades provoquen flagells. Exploten tota la gamma de productes: alguns s’alimenten de les fulles, d’altres viuen a l’interior dels teixits, d’altres aprofiten la fusta (els xilòfags, corticícoles o lignícoles), on excaven galeries; també n’hi ha de granívors, que mengen granes, i de carpòfils, que mengen pol·len; d’altres prefereixen el nèctar de les flors, i en aquest cas tenen peces bucals especialitzades per a recol·lectarlo (els himenòpters i els lepidòpters); uns altres grups s’alimenten de fruits i d’arrels, i n’hi ha molts que utilitzen la saba com a aliment i tenen peces bucals picadores; trobem, encara, un cas més especialitzat, que és el dels micetòfags, que s’alimenten de fongs. Els insectes zoòfags poden alimentar-se de preses vives que ells mateixos capturen (en aquest cas són carnívors depredadors), o bé dels sucs interns, és a dir, de la sang (són hematòfags); també poden comportar-se com endoparàsits, que viuen a l’interior dels seus hostes i es desenvolupen a costa d’ells. Els insectes sapròfags s’alimenten de matèria orgànica, animal o vegetal, en descomposició; la majoria dels insectes aparentment sapròfags en realitat s’alimenten de fongs i bacteris que viuen en aquests medis, i de larves d’altres organismes que sí que són sapròfags. Com a insectes sapròfags podem esmentar els necròfags, que descomponen els cadàvers, els saproxilòfags, que alteren els vegetals, i els copròfags, que viuen als excrements d’altres organismes.
Les relacions tròfiques col·laterals
A més de les relacions tròfiques primàries que hem esmentat hi ha les relacions tròfiques col·laterals, a les quals hi ha unes espècies que s’associen per tal d’obtenir un benefici, sigui mutu o no; d’aquests tipus de relacions comentarem el comensalisme, el mutualisme, la simbiosi i el parasitisme.
Els insectes comensals s’aprofiten de l’aliment sobrer de l’hoste i de les seves descamacions, mudes, excrements, etc. No perjudiquen l’hoste, però aquest no sol rebre tampoc cap benefici d’aquesta associació i, en tot cas, li poden resultar molestos. Hi ha un nombre molt elevat d’insectes comensals i són especialment abundants en els formiguers i termiters, on generalment s’alimenten del menjar emmagatzemat; de vegades els insectes mirmecòfils i termitòfils actuen com a depredadors dels seus hostes i, fins i tot, com a paràsits. Un altre grup important d’insectes comensals són els foleòfils i els nidícoles, que viuen als caus de mamífers i als nius d’ocells, respectivament. A causa de les mateixes característiques d’aquestes relacions, sovint es fa difícil d’establir els límits entre el que és comensalisme i el que és parasitisme; també es consideren relacions de comensalisme la forèsia i la tanatocrèsia, bé que en aquests casos el benefici no és alimentari: en la forèsia, el comensal es deixa transportar per l’hoste, amb el benefici d’augmentar la seva capacitat de dispersió; en la tanatocrèsia el comensal utilitza cadàvers, excrements, peces esquelètiques, secrecions, etc. de l’hoste amb finalitats diverses; n’és un exemple el cas de les larves de tricòpter, que aprofiten les closques de cargols, trossos de plantes i altres materials per a construir els estoigs típics dins dels quals viuen.
El mutualisme és l’associació de dues espècies per la qual ambdues reben algun benefici; de vegades és difícil diferenciar els casos de mutualisme dels de comensalisme o simbiosi. Un bon exemple de mutualisme és el de dues espècies que comparteixen el menjar que aconsegueixen conjuntament.
La simbiosi és la relació entre dos organismes per la qual tots dos reben algun benefici. El simbiont es refugia dins de l’hoste i viu a compte d’ell; l’hoste obté beneficis importants dels seus simbionts, fins al punt que no pot viure sense ells. Hi ha insectes que tenen bacteris i fongs simbionts al tub digestiu, als tubs de Malpighi, a les gònades, a l’hemocel, etc., sigui escampats formant els bacteriòcits i micetòcits o bé reunits en formacions especials anomenades bacteriomes i micetomes, respectivament. Aquest tipus de simbionts són molt corrents en els insectes; els faciliten la digestió de determinats aliments (cel·lulosa, sang, etc.) i els proporcionen substàncies indispensables per al seu desenvolupament que ells son incapaços d’obtenir dels aliments, com és el cas d’algunes vitamines. Els insectes també poden tenir protozous (especialment flagel·lats) i virus simbionts; en aquest cas, en general els insectes fan de vectors o transmissors dels virus i poden transmetre’ls a altres organismes, sigui plantes o animals. Un altre tipus de simbiosi és la que es produeix entre els insectes pol·linitzadors i les plantes entomòfiles, que permet que es produeixi la fecundació i la formació de fruits i llavors en aquestes plantes, i alhora proporciona nèctar i pol·len als insectes: en molts casos hi ha una coevolució molt marcada entre les plantes i els insectes pol·linitzadors. Hi ha un altre cas interessant, i és el d’uns insectes que s’associen amb fongs que viuen als llocs on aquests insectes solen alimentar-se; els fongs els proporcionen substàncies imprescindibles per al seu desenvolupament, que no formen part de la seva dieta, i hom ha observat que els insectes que mostren aquest tipus de simbiosi tenen menys simbionts interns que els altres. Els escolítids (coleòpters xilòfags) infecten les parets de les seves galeries de fongs, els quals els són indispensables per al desenvolupament de les larves; en aquests casos, els adults tenen uns òrgans especials per a transportar els fongs o bé els passen directament a la posta. Moltes vegades, els insectes xilòfags perjudiquen més pel fet de transmetre micosis a la fusta que no pas per la seva acció minadora. Algunes formigues i tèrmits crien fongs en determinats indrets dels seus nius per tal de menjar-se’ls, fenomen que també es pot considerar com un cas de simbiosi externa.
El parasitisme és una relació per la qual una de les dues espècies associades, el paràsit, obté beneficis a costa de les molèsties o perjudicis que causa a l’altra espècie, l’hoste. El paràsit es pot considerar com un depredador molt especialitzat que no arriba a matar l’hoste, o almenys no ho fa d’una manera immediata; de fet, l’extinció de l’hoste només és possible quan el paràsit en disposa d’altres, ja que la seva supervivència depèn de la supervivència del seu hoste, que li és la font d’aliment. Els insectes paràsits poden ésser ectoparàsits o endoparàsits. Els ectoparàsits viuen fora del cos de l’hoste, generalment són hematòfags i dermatòfags, i mostren diferents graus d’especificitat respecte de l’hoste; hi ha grups sencers d’insectes que són ectoparàsits, com les puces, els polls, les xinxes, i també hi ha espècies paràsites en grups de vida lliure, com passa en els dípters, les xinxes de camp, els escarabats, etc. Entre els paràsits externs hi ha els paràsits socials; moltes espècies d’himenòpters socials que no tenen obrers es fan adoptar per membres d’altres espècies colonials a les quals hi ha totes les castes, per la qual cosa aquest cas també es considera d’inquilinisme. Els endoparàsits viuen dins del cos de l’hoste, on s’alimenten dels òrgans i els líquids interns. L’endoparasitisme implica una coadaptació molt estreta dels dos individus associats. És rar en els insectes adults, però no deixa d’ésser corrent entre les larves; per exemple, en són les larves de certs dípters i de molts himenòpters, i també les d’alguns coleòpters i estrepsípters; com a excepció, en són també les femelles adultes d’alguns estrepsípters (els estilòpids). Les relacions de parasitisme es poden estendre en cadenes més o menys llargues (hiperparasitisme) i tenen una gran importància en la regulació de certes poblacions; l’hiperparasitisme és freqüent entre els himenòpters que tenen larves endoparàsites, les quals es desenvolupen a l’interior de larves d’altres insectes paràsits.
La generació de cecidis
Un bon nombre d’insectes fitòfags són capaços de produir unes deformacions especials en les plantes, les anomenades gal·les o cecidis. Les gal·les corresponen a hipertròfies o neoformacions que apareixen en les plantes com a resposta a determinats agents o accions d’un insecte. Aquest fenomen pot ésser degut simplement a l’acció mecànica de l’insecte, però el més corrent és que la picada de l’insecte s’acompanyi de la introducció de substàncies tòxiques (saliva, posta). Hi ha insectes, especialment himenòpters, que deixen la posta dins de les plantes, i això indueix la planta a formar les gal·les que, alhora, constitueixen un refugi per a la posta i els joves de l’insecte.
El tipus de gal·la és constant per a cada tipus de relació, però depèn tant de l’insecte com de la planta; és a dir, una mateixa planta pot formar diversos tipus de gal·les per a diferents insectes, però per a cada espècie d’insecte la fa sempre igual. Produeixen gal·les molts tisanòpters i homòpters, alguns heteròpters, coleòpters, lepidòpters, i molts himenòpters i dípters.
L’activitat pol·linitzadora
Són nombrosos els insectes que es comporten com a florícoles, és a dir, que visiten les flors. En alguns casos la seva presència sobre les flors és accidental, ja que hi cerquen només amagatalls o un lloc de cacera o d’aparellament, però d’altres hi van d’una manera regular i, fins i tot, han desenvolupat tota una sèrie de caràcters morfològics i de comportament que són clarament relacionats amb els seus costums florícoles o antòfils, i llavors els insectes viuen de les flors, ja que s’alimenten de pol·len i nèctar. D’altra banda, mentre recullen aquest aliment, s’enduen adherits als pèls del cos nombrosos grans de pol·len que d’aquesta manera seran transportats fins a una altra flor: són els insectes que anomenem pol·linitzadors.
L’evolució de l’antofília
L’origen dels insectes com a agents dispersadors d’espores es remunta al Carbonífer, abans que no hi hagués la gran radiació de les plantes amb flor (angiospermes). Més endavant, en el Permotriàsic, quan les plantes amb llavor (espermatòfits) començaren a adquirir una certa importància, alguns escarabats (coleòpters) s’alimentaven dels primordis seminals i del pol·len, de manera que podia ser que actuessin de pol·linitzadors. Poc a poc les plantes començaren a desenvolupar mecanismes per a protegir els òvuls de l’acció dels insectes, alhora que el pol·len esdevenia el principal agent atractiu que els vegetals els oferien. Paral·lelament s’anà extenent l’hermafroditisme dels vegetals, cosa que afavoria la pol·linització per part d’insectes antòfils, ja que podien alhora recollir i dipositar pol·len en una mateixa flor. Finalment, durant l’Eocè, el paper dels coleòpters, els dípters, els himenòpters i els lepidòpters en la pol·linització ja era ben establert.
L’evolució dels insectes antòfils ha estat estretament lligada i condicionada per l’evolució de les plantes amb flors, i viceversa. Totes aquestes adaptacions tenen com a funció l’optimització del procés de recol·lecció de nèctar i pol·len per part dels insectes i el desenvolupament de determinades estratègies reproductores en les plantes entomòfiles, destinades a aprofitar la mobilitat i la capacitat pol·linitzadora dels insectes. En aquest cas, insecte i planta representen un exemple ben clar de mutualisme, ja que han evolucionat fins a aconseguir un grau sorprenent d’adaptació mútua i, fins i tot, d’interdependència. S’arriba al cas d’alguns insectes que només recullen el pol·len d’una espècie vegetal (o de molt poques), i alhora plantes que només poden ésser pol·linitzades per determinades espècies o grups d’insectes.
Les adequacions a l’antofília
Els insectes pol·linitzadors han desenvolupat així unes característiques morfològiques i un comportament que permet que els òrgans reproductors de les plantes es posin en contacte amb el cos dels insectes. Per exemple, no actuen com a pol·linitzadores nombroses espècies de formigues que arriben als nectaris de les flors, però que a causa de les seves petites dimensions, no toquen els estams. Tampoc l’abella fustera (Xylocopa violacea), que extreu el nèctar de les flors dels conillets (Antirrhinum) des de fora, fent un foradet a la base de la flor, amb les mandíbules, de manera que no s’impregna de pol·len i, per tant, ha d’ésser considerada com un paràsit del sistema mutualista pol·linitzadors/plantes, no pas com un insecte pol·linitzador, cosa que li ha valgut nom de «lladre de pol·len».
L’eficàcia dels insectes com a pol·linitzadors també depèn de la seva mobilitat. Així, els insectes que visiten moltes flors per unitat de temps, com les abelles i els abellots, fan més pol·linitzacions que no els escarabats, que solen estar-se molta estona damunt de cada flor. En el cas de les plantes autoincompatibles, són més eficaços els insectes que canvien sovint de planta que no aquells que van de flor en flor però no es mouen d’una planta, ja que en el primer cas es produeixen moltes més pol·linitzacions creuades i en el segon moltes més autopol·linitzacions.
Finalment, és especialment important la constància de les espècies d’insectes respecte de les espècies vegetals per tal d’evitar que en visites successives es produeixin pol·linitzacions heterospecífiques, que representen una despesa inútil per part de la planta productora de pol·len i un inconvenient per a la planta receptora, ja que el bloqueig del seu estigma amb aquell pol·len impedeix o dificulta l’arribada (i per tant la germinació) del pol·len conspecífic. En general, els insectes florícoles presenten un grau de constància força elevat, ja que la seva adaptació mútua els facilita la manipulació de les flors, cosa que redueix el temps de visita a cada flor i optimitza la seva recol·lecció.
Els grups d’insectes antòfils
D’entre els grups d’insectes que actuen com a pol·linitzadors esmentarem, per la seva importància, els coleòpters, els dípters, els lepidòpters i, especialment, els himenòpters, si bé hi ha moltes espècies d’altres ordres que també actuen com a pol·linitzadors.
Els coleòpters són considerats els insectes antòfils més primitius ja que no presenten gaires adaptacions a aquest tipus de vida. De la nostra fauna, es comporten com a antòfils els mordèl·lids, edemèrids, cerambícids, clèrids, escarabèids, elatèrids, nitidúlids, melírids, cantàrids, brúquids, etc., encara que en molts casos no en són d’una manera exclusiva. Normalment els coleòpters només visiten les flors quan són adults i s’alimenten del pol·len i del nèctar i també dels primordis seminals, els pètals i els estams, de manera que de vegades fan més mal que bé a les plantes. El paper pol·linitzador dels coleòpters és molt més important en els ecosistemes tropicals, encara que en els climes temperats poden resultar força eficaços en flors de corol·la oberta, com les apiàcies i les rosàcies.
Dels dípters, els bombílids (braquícers), els sírfids i els conòpids (muscomorfs) són els més adaptats a la vida florícola, però també hi ha representants de les famílies dels antòmids, taquínids, múscids i cal·lifòrids, entre d’altres. Les larves de tots aquests grups de dípters són sapròfagues o paràsites i no viuen a les flors. Els adults tenen peces bucals de tipus llepador-xuclador que els faciliten l’extracció del nèctar. D’altra banda, per a recollir el pol·len primer el mullen amb saliva i després l’aspiren. Són més mòbils que no pas els coleòpters i són capaços de fer moviments més precisos. Poden actuar com a pol·linitzadors importants en les zones on els himenòpters són poc abundants o bé en determinades plantes que s’han adaptat a atraure exclusivament dípters: plantes amb corol·la poc profunda i amb els òrgans sexuals assequibles (plantes miòfiles).
Pel que fa als lepidòpters, la majoria de les papallones diürnes i algunes de les nocturnes (especialment els esfíngids), es comporten com a florícoles i s’alimenten exclusivament del nèctar que aspiren amb l’espiritrompa. Els ropalòcers (diürns), tot i que són força mòbils, no visiten gaires flors, perquè tenen pocs requeriments energètics, però en canvi són importants en la pol·linització de determinades flors, posseïdores de corol·les tubulars i profundes, o que amaguen el nèctar en punts on no arriben altres insectes (plantes psicòfiles). Els esfíngids, per contra, tenen requeriments energètics elevats i alhora són bons voladors, de manera que visiten un nombre elevat de flors; són pol·linitzadors especialment eficaços i gairebé exclusius de les anomenades plantes falenòfiles, que obren les seves corol·les tubulars únicament de nit.
Els himenòpters constitueixen, sens dubte, el principal grup d’insectes pol·linitzadors. Convé separar dos grups d’himenòpters, segons la seva manera d’actuar en relació amb la pol·linització.
El primer grup reuneix els símfits (el subordre sencer), la sèrie dels terebrants i la major part de la sèrie dels aculeats (apòcrits). Aquests tenen en comú les peces bucals, de tipus mastegador, modificades per a llepar, però curtes, i també el fet que les seves larves no s’alimenten de pol·len ni de nèctar (tret d’algunes rares excepcions) i, per tant, els adults únicament recullen aliment per a ells mateixos. Els esfècids, eumènids i vèspids són, segurament, els que hom veu més freqüentment damunt les flors, però no presenten adaptacions especials a la vida florícola i la seva importància en la pol·linització pot ésser comparada a la dels dípters.
El segon grup reuneix tots els apoïdeus (és a dir, la resta dels aculeats no inclosos en el primer grup), que són els insectes més adaptats a explotar el pol·len i el nèctar, de manera que són també els millors pol·linitzadors. Hom calcula que pertanyen a aquest grup entre el 70 i el 95 % dels insectes pol·linitzadors de les regions temperades. Contràriament a allò que passa en la resta dels himenòpters, les larves dels apoïdeus s’alimenten, com els adults, de pol·len i nèctar, i les femelles construeixen nius, que abasteixen amb l’anomenat pa d’abella, que no és més que pol·len aixafat i amassat, barrejat amb nèctar. Això fa que els adults hagin de visitar moltes més flors, ja que han d’aportar aliment als nius. Les espècies cleptoparàsites i els mascles de les espècies no paràsites, com que no s’ocupen de l’alimentació de les larves, resulten menys eficaços com a pol·linitzadors. D’altra banda, els apoïdeus tenen les peces bucals de tipus mastegador llepador i, de vegades, molt llargues, per tal de poder accedir als nectaris profunds. Alhora presenten el cos densament recobert de pèls (de vegades, pèls especials), que faciliten l’adhesió dels grans de pol·len, i fins i tot les femelles de moltes espècies tenen uns òrgans especials per a transportar el pol·len recollit, que s’anomenen escopes i que se situen a les potes del davant (en les abelles podilègides, com Andrena, Halictus, Colletes) o a les esternites abdominals (en les abelles gastrilègides, com Megachile, Osmia, Anthidium). Altres, com l’abella de la mel (Apis mellifica) i els abellots (Bombus) transporten el pol·len ja amassat en les corbícules, que fan com una mena de cistellets, situats a les tibies posteriors.
Els apoïdeus tenen els òrgans dels sentits molt desenvolupats i una capacitat d’aprenentatge molt elevada respecte de la resta dels insectes antòfils, i això els ha permès manipular amb destresa les flors amb mecanismes de pol·linització complexos i de distingir bé entre les diferents espècies vegetals (són especialment constants). Per això, les plantes adaptades a la pol·linització per abelles (plantes mel·litòfiles) tenen les flors amb els nectaris amagats i la seva manipulació requereix una certa força i precisió per part de l’insecte. Alhora, la capacitat pol·linitzadora dels apoïdeus s’accentua en les espècies socials. En el cas de l’abella de la mel, les dances de reclutament permeten concentrar un gran nombre d’individus en les plantes més productives, de manera que aconsegueixen així explotar-les amb més eficàcia que les altres espècies d’abelles.
L’adequació a l’ambient
Els insectes constitueixen el grup animal més diversificat ecològicament. Tot i que són fonamentalment terrestres, n’hi ha grups sencers que són aquàtics, o que en són, almenys, en les fases larvals o juvenils, especialment a les aigües continentals. Els insectes viuen a tot el planeta i s’han adaptat a viure als ambients més variats: des de les regions àrtiques fins a les equatorials, des dels boscos tropicals fins als deserts més àrids, des de la vora de la mar fins a l’alta muntanya, des de les aigües continentals fredes fins a les aigües termals, i, fins i tot, algunes espècies, han colonitzat el domini marí. La gran importància numèrica dels insectes es deu, en una gran part, a la seva plasticitat evolutiva, que es tradueix en tota una sèrie d’adaptacions morfològiques i fisiològiques, i a la seva gran capacitat de dispersió.
Els factors físics més importants que determinen la presència d’un organisme en un ambient determinat són: la temperatura, la humitat i el fotoperíode.
Els insectes, com tota la resta dels organismes, poden ésser euriterms, és a dir, capaços de suportar canvis importants respecte de la temperatura òptima de desenvolupament, o bé estenoterms, és a dir, incapaços de tolerar oscil·lacions de temperatura que no siguin molt petites. Tot i que no regulen la seva temperatura interna, que depèn de la temperatura ambiental, els insectes poden modificarla de maneres diferents. En uns casos, en els dies freds augmenten la seva temperatura corporal per mitjà de moviments musculars, com és el cas de molts insectes voladors, que mouen les ales abans d’iniciar el vol per tal d’atènyer una determinada temperatura; en altres casos, simplement es posen al sol i retenen l’escalfor per mitjà de pèls especials o esquames. En el sentit contrari, els insectes que viuen a zones molt caloroses solen enterrar-se o amagar-se en les hores de més calor per tal d’evitar la deshidratació, i són actius quan el sol es pon. La temperatura òptima de desenvolupament varia en les diferents espècies, cosa que s’ha pogut comprovar en insectes que viuen en un mateix biòtop: cada espècie inicia la seva activitat quan la temperatura adquireix un determinat valor; com a casos extrems, podem esmentar el d’alguns dípters (alguns culícids), que són actius en les regions àrtiques a zero graus, i el d’uns altres dípters (alguns quironòmids), que viuen a les aigües termals i poden desenvolupar-se entre els 49 i els 51 graus. En general, quan la temperatura baixa respecte de la temperatura òptima, també baixa l’activitat dels organismes, mentre que quan puja s’incrementa aquesta activitat.
Pel que fa a la humitat, els insectes poden ser eurihigrobis, quan resisteixen variacions importants d’humitat, o bé estenohigrobis, quan només en toleren petits canvis. En uns casos estan adaptats a viure en ambients molt secs, mentre que en d’altres només poden viure en ambients saturats d’humitat. D’altra banda, el grau d’humitat té relació amb la temperatura ambiental. Lligada a la temperatura i la humitat, la resistència dels organismes a la deshidratació és variable; en general, els insectes que viuen en ambients molt secs tenen la cutícula molt endurida i totalment impermeable; en altres casos, la cutícula és tova però impermeable a causa del gran desenvolupament de l’epicutícula. D’altra banda, quan la cutícula no és impermeable, els insectes es veuen obligats a viure en ambients molt humits, cosa que evita la seva deshidratació; per aquesta raó, moltes larves i adults cerquen refugi en el medi edàfic, i fins i tot a les cavitats subterrànies, ja que fora d’aquests ambients moren deshidratats.
La importància biològica de la llum és variable. Hi ha espècies, acostumades a un fotoperíode diürn, que poden desenvolupar-se en la foscor, mentre que, per contra, d’altres necessiten l’existència del fotoperíode per a desenvolupar-se. Respecte d’aquest factor, els insectes poden ser eurífots o bé estenòfots, segons quina sigui la seva capacitat d’adaptació a les variacions quantitatives de llum. Hom entén per fotoperíode el període diürn de les hores de llum respecte de les hores de foscor. Hi ha organismes que requereixen per a desenvolupar-se un fotoperíode llarg, el corresponent als dies de primavera o d’estiu, mentre que d’altres en requereixen un de curt, com per exemple el corresponent als dies de final de la tardor, i encara, n’hi ha que són indiferents al fotoperíode. El fotoperíode és d’una importància vital per al determinisme de la diapausa i també influeix sobre el polimorfisme.
Els insectes poden reaccionar de maneres diferents als estímuls lluminosos; alguns en resulten atrets (tenen fototropisme positiu), mentre que d’altres són repel·lits (tenen fototropisme negatiu). Alguns, especialment els lepidòpters nocturns, són atrets per la llum artificial, sobretot quan és de longitud d’ona curta (llum ultraviolada). El grau de fototropisme varia a cada espècie i es relaciona amb la llargada d’ona de la llum i amb les condicions ambientals (temperatura, humitat, presència de la lluna, nebulositat, etc.); per tant, el fototropisme positiu és més gran en les nits càlides i humides, de baixa pressió, i sense vent ni lluna.
Els recursos defensius
Un aspecte important de la supervivència dels organismes són les estratègies defensives respecte dels seus possibles depredadors. Moltes vegades, la resposta dels insectes a la presència d’un depredador és la fugida, reacció que és desencadenada per estímuls visuals, auditius o químics (feromones d’alarma); únicament produeixen feromones d’alarma els insectes socials i gregaris. En altres casos la reacció davant del perill els fa reunir-se en grups nombrosos (gregarisme). Alguns tenen mitjans ofensius o defensius per a superar el perill, com per exemple secrecions tòxiques, tegument resistent, actituds d’intimidació, immobilització reflexa, etc. Finalment, la cripsi i el mimetisme constitueixen els mecanismes de defensa més sofisticats.
Molts insectes dels anomenats picadors es defensen inoculant substàncies tòxiques als seus possibles agressors per mitjà de les peces bucals (el cas dels dípters i els heteròpters, entre altres) o bé per mitjà d’un ovipositor modificat, que es connecta amb les glàndules del verí (cas dels himenòpters) o d’uns pèls urticants (les erugues de molts lepidòpters). Les secrecions tòxiques d’algunes glàndules tegumentàries (repugnatòries o repel·lents) sovint molt pudents, també col·laboren a la defensa; els uns les projecten a distància (alguns coleòpters, els escopeters), d’altres, davant d’una situació de perill expulsen "sang", que és només hemolimfa amb substàncies tòxiques, per mitjà d’un fenomen anomenat autohemorrea, que té lloc en certes membranes articulars i en les vesícules exèrtils i que és conegut especialment en coleòpters (coccinèl·lids, crisomèlids, melòids, etc.) i en alguns ortòpters; així mateix, alguns insectes, especialment ortòpters, quan estan en perill, deixen anar una saliva que és tòxica i irritant.
Hi ha insectes (lepidòpters, ortòpters i alguns coleòpters) que són capaços d’acumular en el cos productes tòxics, en general procedents de la seva dieta alimentària; la presència d’aquestes substàncies fa que siguin de gust desagradable o, fins i tot, tòxics per als seus possibles depredadors. Aquests insectes solen ésser de colors vius, advertidors de la seva condició de tòxics, i per això es considera un cas de mimetisme.
Les actituds de defensa poden ser actives o passives. En el primer cas, els insectes, especialment màntids (els pregadéus) i ortòpters (els llagosts) intenten augmentar la seva envergadura tot estenent les ales, mentre adopten una postura amenaçant per a espantar al seu depredador; d’altres emeten sons amb la mateixa funció d’espantar i desconcertar l’enemic. Entre les actituds de defensa passiva, hi ha, per exemple, la de fer-se el mort, també anomenada immobilització reflexa, que és molt corrent en els insectes i que pot considerar-se com un fenomen de cripsi, ja que facilita que l’insecte passi desapercebut; altres insectes, especialment dictiòpters, insectes bastó i ortòpters, quan són atacats per un depredador, s’amputen un o més apèndixs (autotomia reflexa), que després són capaços de regenerar. La duresa del tegument i la possibilitat d’encaixar tots els apèndixs en la carcassa del cos fent com una mena de bola és un altre element que es pot considerar defensiu; de vegades, el cos i els apèndixs tenen espines i altres formacions punxants, amb la mateixa funció.

L’homotípia és un tipus de cripsi pel qual els insectes imiten la forma i el color del substrat sobre el qual viuen. La fotografia ens mostra diferents espècies de papallones fulla, exòtiques (gènere Kallima), el color i els dibuixos de les quals varien d’una posta a l’altra, i fins i tot en individus nascuts d’una mateixa posta, presumiblement com a sistema per a evitar un model concret i augmentar, així, la seva eficàcia críptica.
Jordi Vidal/Col·lecció Vilarrúbia.
Els insectes solen cercar refugis temporals o permanents, per tal de protegir-se dels seus depredadors: s’amaguen sota les pedres i les escorces, a les molses i al sòl, entre la fullaraca i la vegetació etc. De vegades segreguen substàncies protectores (laques i ceres) amb les quals s’empastifen tot el cos, o es col·loquen excrements al damunt, restes de les preses, pedretes, trossos de plantes, etc., tot per tal de passar desapercebuts. De tota manera, molts insectes construeixen activament els seus propis refugis. Hi ha una gran diversitat d’especialitzacions en la construcció de refugis, sobretot en les fases larvals, per a la qual fan servir tota mena de materials cimentats amb saliva, excrements o amb fils de seda (per exemple, en tricòpters, i alguns lepidòpters i coleòpters); d’altra banda, la nimfosi dels holometàbols sempre té lloc dins de capolls o en llocs protegits. En alguns casos els insectes construeixen nius col·lectius, per exemple les bosses de la processionària del pi, dins les quals viuen un gran nombre d’erugues.
La cripsi i el mimetisme tenen com a finalitat alterar el comportament dels seus depredadors. La cripsi permet a l’insecte de no ésser reconegut pel depredador com una possible presa, mentre que en el mimetisme, encara que el depredador reconeix l’insecte, el desprecia perquè l’associa a organismes que tenen caràcters ofensius o defensius i que li són tòxics.
Els organismes críptics tenen aspectes, colors, formes de moviment i olors que els fan poc aparents, de manera que es confonen amb el medi. L’homocromia i imitació dels colors del medi és el sistema críptic més estès; sol acompanyar-se de la immobilització reflexa en situacions de perill i, sovint, el motiu del dibuix consisteix en taques que trenquen el contorn visual de l’animal, de manera que aquest es confon amb el substrat; aquest tipus de coloració s’anomena disruptiva i el trobem a diferents grups d’insectes. D’altra banda, alguns insectes poden canviar de color en funció dels tons del substrat i del fotoperíode; i el recobriment de moltes larves d’insecte amb excrements o restes de mudes també es pot interpretar com un cas de cripsi. Un altre tipus de cripsi és l’homotípia o imitació de formes d’objectes naturals, a més de la imitació del color; així, és conegut de tots el cas dels insectes que imiten branquetes (els insectes bastó, els nostres cavalls de faves), fulles (els insectes fulla, lepidòpters, ortòpters, etc.); de vegades, la imitació és tan perfecta que fins i tot mostren trossets trencats i taques que imiten l’atac de fongs.
Els insectes proveïts de defenses pròpies, sigui a partir d’armes o d’òrgans defensius o bé pel fet de tenir un gust dolent, que sol ésser tòxic, evolucionen en el sentit de fer-se ben visibles en el medi on viuen, atès que no tenen cap motiu per a témer els depredadors; generalment tenen colors molt vistents (advertidors o aposemàtics) i en general es mouen lentament, són diürns i solen viure en agregats o colònies. La coloració advertidora pressuposa un aprenentatge i per això es dirigeix a depredadors evolucionats, en general ocells i mamífers, bé que també poden presentar aquesta capacitat alguns amfibis i invertebrats; els depredadors aprenen a deixar en pau els organismes vistents, pel perill que els suposen. La coloració aposemàtica combina colors de contrast màxim, que alhora destaquen sobre el color del medi, com, per exemple, combinaciones de negre amb groc o taronja, que són ben uniformes en els insectes gràcies al petit nombre de possibilitats que permet la fórmula òptima; aquests colors són corrents en molts himenòpters (abelles, vespes, abellots, etc.), en alguns heteròpters, alguns lepidòpters (helicònids, danàids, etc.) i coleòpters (lícids), tots proveïts de recursos de defensa. Les característiques aposemàtiques també poden afectar les olors i l’emissió de sons. És força corrent que diferents espècies proveïdes d’elements protectors adoptin el mateix aspecte i color, encara que pertanyin a grups molt diversos. L’evolució de les espècies aposemàtiques amb el mateix aspecte extern no és totalment independent i hom diu que formen part d’un cercle sinaposemàtic i també que constitueixen un tipus de mimetisme anomenat müllerià.
Hi ha altres insectes que, tot i ser inofensius, adopten, externament, el mateix aspecte que els insectes aposemàtics, la qual cosa els dona una certa protecció; també poden imitar-ne els sons i les olors. En aquest cas, hom parla de mimetisme batesià, fenomen que per a ésser efectiu, requereix unes condicions determinades: l’espècie model (la perillosa) i la còpia (la inofensiva) han de coincidir geogràficament i ecològicament. Són nombrosos els casos de mimetisme batesià entre els insectes; els més coneguts són de papallones (lepidòpters) de les famílies dels pièrids i papiliònids, inofensives, que imiten les de les famílies dels helicònids i danàids (aposemàtics); molts dípters, heteròpters, coleòpters i ortòpters inofensius copien la forma i els colors de les vespes i les abelles (himenòpters), proveïdes de fibló.
L’organització social
El grau de psiquisme que tenen certs insectes és elevat. Alguns són capaços d’aprendre i memoritzar fets determinats, la qual cosa els permet modificar o adaptar la seva conducta a les noves condicions. S’ha comprovat que els insectes són capaços d’orientar-se, i ho fan de diferents maneres; les abelles, quan surten del rusc, resten una estona voleiant-lo pel damunt per tal de memoritzar el color i la forma de l’entorn i, quan no hi ha punts de referència, es guien per la posició del sol. D’altra banda, les mateixes abelles són capaces de transmetre’s informacions, com per exemple, el lloc on hi ha menjar, la distància a la qual es troba, etc., a través d’un llenguatge fet de moviments. Hi ha insectes que poden arribar a tenir una noció exacta del temps, capacitat que ha rebut el nom de «rellotge biològic».
Les abelles surten del rusc just a l’alba, però si el rusc és transportat a una altra regió, elles continuen regint-se pel cicle del país d’origen i triguen uns quants dies a modificar el seu horari. D’altra banda, l’emergència o naixement dels adults a partir de la darrera fase juvenil en molts grups d’insectes es limita a unes hores determinades del dia. El grau de complexitat que pot atènyer el comportament dels insectes és ben manifest en els insectes socials.
En general, els insectes únicament es reuneixen amb altres individus de la seva mateixa espècie durant l’època de la reproducció, i, de vegades, un temps més, mentre tenen cura de les postes i les cries. En molts casos, els insectes són insociables, i mostren un territorialisme exagerat que els porta a atacar els intrusos, siguin o no de la seva mateixa espècie.
En els insectes gregaris, els individus d’una mateixa espècie es reuneixen sota un motiu principal i determinant, que és l’existència de condicions adequades per a viure, siguin de microclima, quantitat d’aliment, etc. Aquestes reunions es mantenen mentre duren les condicions, però es dispersen tan bon punt aquestes canvien. Hi ha molts casos de gregarisme en fase larval o juvenil i alguns es dissolen en atènyer l’estadi d’adult, com passa en la processionària del pi, i d’altres es mantenen, com és el cas dels pugons. També hi ha casos en què el gregarisme es limita a unes hores determinades del dia, com per exemple, el cas de les danses crepusculars dels mosquits i les danses dels efemeròpters i alguns odonats. Un tipus particular de gregarisme és el dels grups formats per individus en repòs, que es poden limitar al període nocturn o bé, el cas més general, perdurar durant tota l’estació desfavorable, generalment tot l’hivern; aquest tipus de gregarisme s’observa en alguns heteròpters (Pyrrhocoris), alguns himenòpters (Ammophila) i la majoria dels coccinèl·lids (coleòpters). El gregarisme té, encara, característiques especials en alguns ortòpters, en els quals hi ha una alternança de generacions solitàries i de generacions gregàries; les gregàries volen en grans grups i poden provocar flagells als conreus i als arbres; és ben conegut el cas dels llagosts (Locusta migratoria), que en fase gregària fan migracions i constitueixen greus flagells, que afecten, sobretot, el continent africà.
Els insectes socials no poden viure aïllats dels seus congèneres a causa dels estrets llaços intraspecífics que s’estableixen entre els individus de la colònia. En els casos més especialitzats formen societats homogènies, constituïdes pels descendents d’una sola parella (societat monogènica) o de més d’una (societat poligènica). Les relacions entre els individus que formen la colònia poden ésser molt complexes i comporten la diferenciació de castes que fan funcions diferents; únicament formen societats complexes els tèrmits i alguns himenòpters. En alguns casos, les societats són temporals, generalment anuals (cas de les vespes i abellots), i es formen a l’època favorable, fundades per una femella que ha estat fecundada prèviament. Altres són permanents o plurianuals, com el cas de les formigues, els tèrmits i les abelles, i és en aquest cas que es diferencien les castes; en principi, n’hi ha dues: una de fèrtil i una altra d’estèril, però aquesta darrera pot subdividir-se en altres castes, que fan funcions diferents. La casta fèrtil és formada per pocs individus que s’encarreguen de la reproducció i tenen característiques morfològiques i fisiològiques especials: les femelles tenen l’abdomen hipertrofiat, una gran longevitat i fecunditat, etc. La casta estèril reuneix la majoria dels individus de la colònia, que en els tèrmits són mascles i femelles, i en canvi en formigues i abelles són només femelles; són individus normals però amb les gònades no funcionals, per un fenomen de regressió controlat per diversos factors (dieta alimentària durant el desenvolupament, acció de feromones, etc.). La casta estèril, en les societats més evolucionades, pot subdividir-se en altres dues castes: la de les obreres i la dels soldats, i aquestes també es poden diferenciar en altres tipus segons les funcions que fan. El conjunt de les castes estèrils és molt especialitzat i tenen cura dels reproductors (la reina o els reis), transporten els ous a les cel·les de cria, cerquen aliment, tenen cura de les larves i de la neteja i la defensa de la colònia. La casta fèrtil només s’ocupa de la reproducció, segons la seva composició hom distingeix dos tipus de societats: les matriarcals (les dels himenòpters), a les quals els mascles només viuen un temps molt curt, i les conjugals (les dels tèrmits), a les quals els mascles són presents d’una manera permanent a la colònia.
Per a aconseguir la perfecta organització de la societat cal la coordinació de tots els individus que la integren, i això fa indispensable la capacitat de comunicació. Per això, els insectes socials tenen els òrgans sensorials molt desenvolupats i un psiquisme elevat. Pel que sembla, les feromones juguen un paper important en el perfecte funcionament d’aquestes societats.