Els crustacis són, després dels insectes, el grup d’artròpodes que ha tingut més èxit evolutiu, si jutgem pel nombre d’espècies existents (entorn de 30 000) i per la diversitat d’ambients on es troben. Es tracta d’un grup quasi exclusivament aquàtic, adaptat perfectament, tant a les aigües marines com a les continentals. En aquest sentit, hom considera que, per l’espai que ocupen i per la funció que hi fan, són els equivalents als insectes, fonamentalment terrestres, en el medi aquàtic. De fet, també hi ha espècies de vida terrestre, però en molts casos la seva permanència al medi terrestre és molt limitada i necessiten retornar a l’aigua periòdicament, com passa en algunes espècies de decàpodes del litoral marí. Únicament dins dels isòpodes, hi ha espècies que s’han adaptat plenament a la vida terrestre, en condicions precàries, però, comparat amb d’altres grups d’artròpodes de vida terrestre. D’altres grups, com els amfípodes, els ostràcodes i els cladòcers, tenen una presència més aviat anecdòtica al medi terrestre. El parasitisme és una forma de vida relativament estesa entre els crustacis, i ha donat lloc a una gran diversificació en alguns grups. Com passa en general en tots els grups paràsits, aquest tipus de vida ha induït a nombroses modificacions anatòmiques, de diferent nivell segons el grau d’adaptació. En els casos extrems, en espècies endoparàsites, fins i tot es fa difícil de reconèixer la condició d’artròpodes en la seva morfologia, i la seva identificació només és possible en les fases larvàries, que corresponen a la morfologia típica del grup.
Característiques del grup
Fins i tot si hom prescindeix de les espècies paràsites, la variabilitat morfològica dels crustacis es mostra molt elevada, motiu pel qual es fa difícil d’establir una diagnosi precisa del grup, si no és per un nombre reduït de caràcters, que en general resulten poc conspicus a l’observació directa. Malgrat aquesta dificultat, hom considera els caràcters següents com a definidors del grup dels crustacis.
Pel que fa a la morfologia, el cèfalon de les formes adultes és format per l’àcron i 6 metàmers, separats en dues regions: una preoral i una altra postoral; la regió oral consta dels metàmers preantenular, antenular i antenal, i la regió postoral dels metàmers mandibular, maxil·lular i maxil·lar. Les mandíbules són de tipus gnatobàsic, és a dir, que la seva part més desenvolupada és la basal i correspon a les endites, mentre que la resta de l’apèndix pot ésser modificat, amb tendència a la reducció o fins i tot a la desaparició. En relació al desenvolupament embrionari, és característic l’origen d’algunes zones del cos a partir de determinades cèl·lules de l’ou; en els grups on el desenvolupament embrionari és breu, de l’ou en surt una larva anomenada naupli. A la larva naupli, les antenes són postorals, cosa que ha donat lloc a una certa controvèrsia al voltant de la natura exacta i la disposició dels segments que formen el cèfalon.
A més de les característiques esmentades, n’hi ha d’altres, potser no tan generals, que es troben, però, molt esteses entre els crustacis i que, per tant, es poden considerar representatives: la colonització d’ambients aquàtics i la respiració branquial; la divisió del cos en un nombre variable de metàmers que s’agrupen en tres tagmes o regions, cèfalon, perèion i plèon, que en alguns grups poden mostrar algunes modificacions, com és la reunió del cèfalon amb un o més metàmers del perèion per a formar el cefalotòrax; la situació dels orificis genitals fora del darrer segment del cos, el qual conté, en canvi, l’anus; i la presència d’òrgans excretors segmentaris i d’origen mesodèrmic.
Morfologia
El tegument i la closca

La morfologia dels crustacis varia molt d’uns grups als altres. La imatge típica és associar la idea de crustaci a la idea de decàpode, i imaginar que tenen «aspecte de gamba», com la gambeta il·lustrada (Palaemonetes zariquieyi), procedent del delta de l’Ebre. Hi són constatables, certament, els elements morfològics externs de la classe (la possessió de dos parells d’antenes, la closca impregnada de sals, els apèndixs birramis, etc.), però presentats sota una aparença ben distinta a la que adopten en el cas dels ostràcodes o dels copèpodes, per exemple.
Enric Curto.
La cutícula dels crustacis és feta d’una sèrie de capes de quitina, segregades per l’epidermis, i es diferencia de la cutícula de la resta dels artròpodes pel fet de mostrar una elevada calcificació. En realitat aquesta és una característica no tan estesa com el nom del grup sembla indicar, ja que en molts dels seus representants el tegument és poc calcificat. D’entre les diferents capes que formen la cutícula és especialment important l’epicutícula doble, a la qual manquen les ceres impermeables, presents a la resta de grups d’artròpodes i que contribueix a regular el balanç hídric. Possiblement, aquesta és una de les causes principals que expliquen el poc èxit evolutiu dels crustacis en el medi aeri, on és indispensable una bona impermeabilització. Les capes més internes de la cutícula formen la procutícula, que al seu torn és feta de diverses capes de calcificació creixent de dins cap enfora.

Els colors de la closca dels crustacis no sempre són invariables per a cada espècie. Al contrari, moltes vegades, diferents individus d’una mateixa espècie (o, encara, un mateix individu al llarg de la seva vida) poden mostrar tonalitats i dibuixos distints, els quals, per tant, no tenen valor taxonòmic. Així, en l’isòpode Sphaeroma serratum, que reprodueix la il·lustració, hom observa una gran variabilitat en la distribució de les taques de la cara dorsal del cos, que és conseqüència d’un polimorfisme genètic.
Ramon Torres.
Com ja hem indicat, la pretesa robustesa de l’esquelet dels crustacis no és tal, ja que al costat de formes amb cutícula engruixida de fins uns quants mil·límetres (cas dels decàpodes), n’hi ha d’altres, generalment planctòniques i petites, a les quals l’esquelet és una fina cutícula totalment transparent i mancada de calcificació. En les formes més robustes, hom aprecia a la cara interna el desenvolupament d’apodemes i trabècules que serveixen de punt d’inserció als potents músculs, a més de formar un endosquelet relativament desenvolupat.
Els colors que adopten les closques dels crustacis són deguts generalment a l’existència de pigments, que poden allotjar-se en unes cèl·lules molt vacuolitzades, que es distribueixen per la cutícula i que són regulades per activitat hormonal; el pigment més comú és la zoeritrina. En les formes planctòniques, especialment en els copèpodes, és freqüent l’existència de gotes d’oli (en els d’aigua dolça) o de ceres (en els marins) que els dona un color vermell intens o blavós, i que és degut al carotenoide astaxantina o a d’altres de composició semblant. A més d’actuar com a substàncies de reserva i contribuir a la flotabilitat de l’animal, en els llacs d’alta muntanya i en les aigües molt somes, aquests pigments tenen un valor protector contra l’alta intensitat de les radiacions lumíniques i les radiacions ultraviolades. En altres casos, per contra, la manca de pigmentació pot servir de protecció contra la depredadó, especialment si la detecció és visual.
La closca dels crustacis és una de les seves estructures morfològiques característiques. De fet, la closca és una expansió del tegument de la regió posterior del cèfalon, que s’estén en totes les direccions, però especialment cap enrere, fins a cobrir bona part del cos. La closca pròpiament dita implica la fusió de, com a mínim, un metàmer del perèion al cèfalon, és a dir, la formació del cefalotòrax. No obstant això, en alguns grups evolucionats, la closca es perd secundàriament, com passa en els amfípodes i els isòpodes. En aquests casos, hi ha una estructura semblant, però més petita, que només cobreix el cap, anomenada escut cefàlic. L’escut cefàlic és format per la unió de les tergites dels metàmers cefàlics i dona al cèfalon un aspecte unitari semblant al del cap dels insectes o d’altres grups d’artròpodes.
Les dimensions i la forma de la closca són molt variables. En els casos més simples es tracta d’una petita placa corbada cap al cos que s’eixampla en direcció al perèion, i també lateralment i cap endavant. Quan aquesta closca és més desenvolupada, com passa en els eucàrides, forma una cuirassa protectora que s’estén cap enrere fins a cobrir tot el perèion, cap endavant per a formar el rostre, i pels costats, fent una cambra branquial per la qual circula l’aigua. En d’altres crustacis, la closca és bivalva, és a dir, són dues plaques articulades per una xarnera a la part dorsal, que permet que s’obrin i es tanquin per mitjà de músculs situats en el segment maxil·lar. Aquest tipus de closca es pot desenvolupar molt més extensament i pot arribar a cobrir tot el cos de l’animal, de manera que aquest adopta un aspecte semblant al dels mol·luscs bivalves; és el cas dels crustacis concostracis i ostràcodes. En altres grups, la situació no és tan espectacular, tot i que també cobreix una part important del cos, com passa en els leptostracis. Finalment, en els cirrípedes, la closca és considerablement modificada, i forma l’anomenat mantell, que, a més de cobrir tot el cos, segrega unes plaques calcàries externes que protegeixen completament l’animal. Mentre que en les espècies de closca simple la cavitat que queda entre la paret externa del cos i la interna de la closca serveix principalment per a la funció respiratòria, en el cas dels crustacis bivalves es relaciona més directament amb la reproducció, ja que sovint serveix de cambra incubadora o fins i tot de punt d’allotjament de les gònades, els cecs intestinals o d’altres parts del cos.
L’existència de closca en molts dels grups de crustacis i alhora en grups ancestrals d’artròpodes, com per exemple els trilobits, ha estat motiu perquè alguns especialistes la considerin com un caràcter primitiu en els artròpodes, perdut secundàriament en determinats grups, com els artròpodes terrestres. Contra aquesta teoria hom argumenta que, precisament, els crustacis més primitius, els cefalocàrides, no tenen closca.
Les regions dels cos i la metamerització
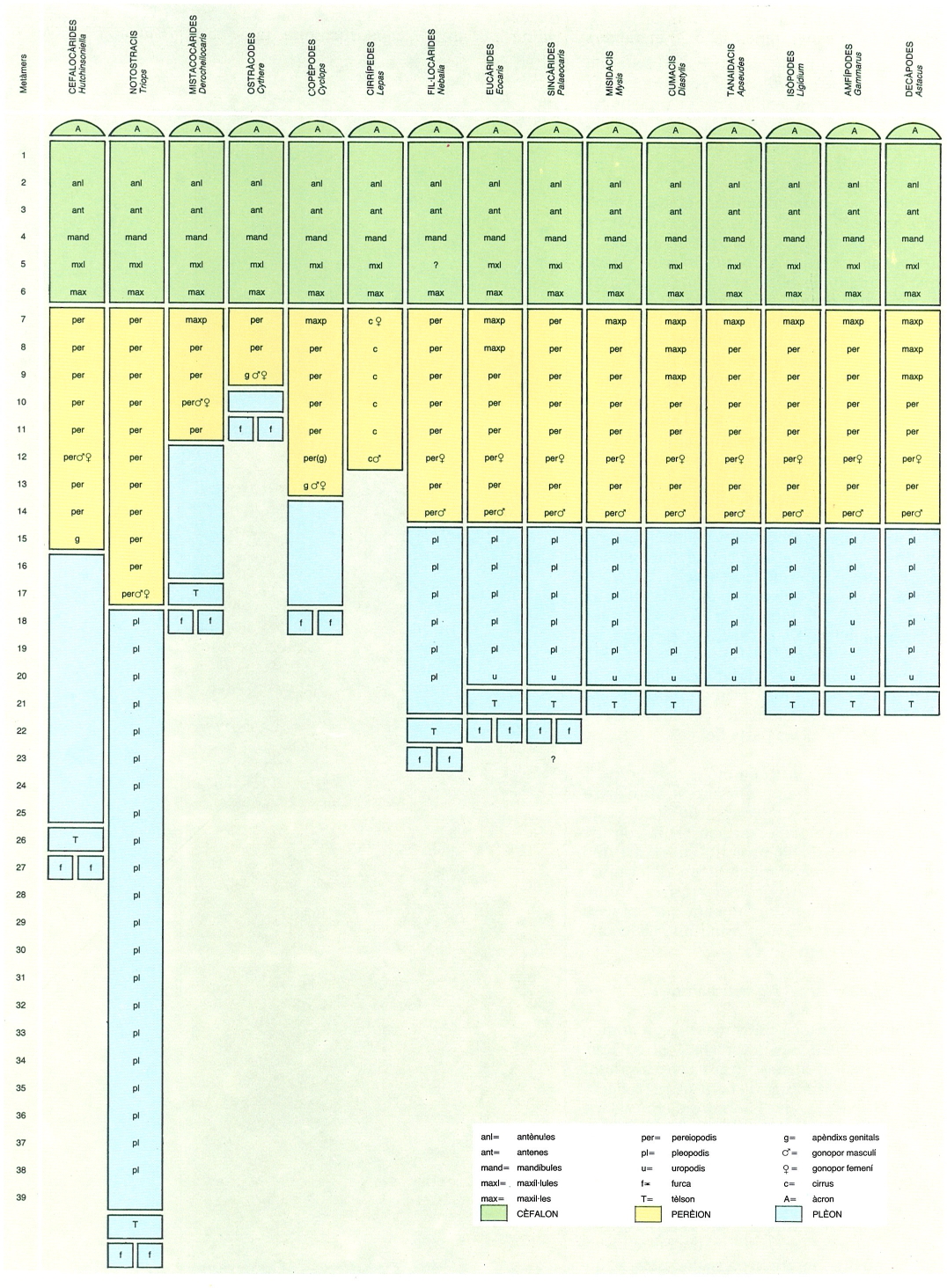
Nombre i distribució de segments en diversos gèneres representatius dels principals grups de crustacis, amb indicació dels apèndixs de què cadascun és portador. Hom consigna, també, l’emplaçament dels porus genitals i els apèndixs modificats per a fer funcions reproductores. Els diferents colors indiquen els segments corresponents al cèfalon, al perèion i al plèon. Dins de cada grup poden trobar-se combinacions diferents de les indicades ja que la distribució dels apèndixs sol ésser un caràcter molt variable, cosa que s’accentua especialment en el cas dels cumacis i el dels tanaidacis.
Maber, modificat a partir de Moore 1969.
El cos dels crustacis és format per un nombre variable de metàmers intercalats entre l’àcron i el tèlson. En alguns grups, els considerats com a més primitius (cefalocàrides, concostracis i notostracis), el nombre d’aquests metàmers pot arribar a ser superior a 50, mentre que en d’altres, com els cladòcers i els ostràcodes, no n’hi ha més de deu. La pèrdua de metàmers es considera una tendència cap a formes més evolucionades, cosa que no sempre s’adapta al grup dels crustacis; efectivament, en una posició intermèdia entre els extrems ara dits, se situen els malacostracis, que són considerats com els crustacis més evolucionats i, en canvi, tenen 19 o 20 metàmers. La identitat dels metàmers no és sempre evident, com passa també a la resta d’artròpodes, perquè la divisió general del cos queda subordinada a l’especialització regional o tagmatització. D’aquesta manera, el cos dels crustacis queda dividit en tres tagmes: cèfalon, perèion i plèon. El cèfalon és format per l’àcron i sis metàmers, el primer dels quals, anomenat preantenular, no porta apèndixs. La divisió d’aquesta regió en dues, la preoral i la postoral, suposa també una divisió funcional dels apèndixs situats a cada zona: a l’anterior hi ha els apèndixs sensorials (antènules i antenes), a la posterior els bucals (mandíbules, maxíl·lules i maxil·les). El perèion i el plèon porten apèndixs anomenats, respectivament, pereiopodis i pleopodis, els quals tenen una funció molt més diversificada i més relacionada amb la forma de vida de cada espècie.
La divisió del cos en els tres tagmes esmentats moltes vegades resta emmascarada a causa de l’existència del cefalotòrax. El cefalotòrax és el resultat de la cefalització d’un o més metàmers del perèion i del desenvolupament de la closca a partir del cèfalon vers la part posterior del cos. Actualment hom accepta que és resultat de la fusió del cèfalon amb un o més segments del cos, però també és corrent considerar que només quan el cèfalon s’uneix amb la totalitat del perèion (cosa que passa únicament en els eucàrides) es pot parlar de cefalotòrax. En aquesta obra utilitzarem la primera accepció.
La fusió de metàmers també és corrent en el plèon. Generalment es produeix entre el darrer segment i el tèlson, i es forma el pleotèlson que, a més, pot tenir una furca al final, que pot ser articulada (en els isòpodes) o no serne (en els copèpodes). Alguns grups (per exemple els ostràcodes) tenen el plèon molt reduït, i només s’aprecia un sac no segmentat; en altres casos, com en els cirrípedes, fins i tot pot arribar a desaparèixer.
Els apèndixs i les formacions apendiculars

Els apèndixs dels crustacis són típicament birramis. Únicament el primer parell d’antenes és format per una sola branca, i el segon parell, també visible a la fotografia del cap d’un escamarià (Nephrops norvegicus), mostra l’estructura bífida característica. Les mandíbules, maxil·les i maxil·lípedes, de petites dimensions, no són visibles externament. El primer parell de potes del perèion, acabat en pinça com tots els altres, apareix com a unirrami perquè té una de les dues branques formant part del sistema branquial. L’estructura dels apèndixs dels decàpodes, com l’escamarià, correspon al tipus estenopodial.
Amadeu Tardiu.
Juntament amb la tagmatització del cos, es produeix la modificació dels apèndixs de cada metàmer en relació a diferents funcions. Seguint el paral·lelisme amb l’evolució dels metàmers, hom pot dir que en les formes més primitives, els apèndixs tendeixen a ser iguals o molt semblants, mentre que en les espècies més avançades es van transformant i s’adeqüen a funcions específiques. És lògic pensar que, a mesura que s’ha anat produint una evolució dels hàbits de cada espècie, els membres respectius s’han anat ajustant a funcions cada vegada més concretes, segons la seva localització en el cos. Entre aquestes funcions hi ha la sensorial, la masticatòria, la de captura i manipulació de l’aliment, la locomotora (en diferents variants), la respiratòria, la reproductora i d’emmagatzematge dels ous, i també una combinació d’algunes d’aquestes funcions. Cada adaptació ha anat presentant modificacions, cada vegada més allunyades d’allò que hom considera l’estructura bàsica de l’apèndix dels crustacis. Així, en alguns casos, únicament un estudi detallat a partir del desenvolupament larvari permet seguir l’evolució d’un apèndix i la identificació de cadascuna de les seves parts.

Els anomenats cirrus dels cirrípedes, dels quals veiem a la fotografia el percebe (Pollicipes cornucopia) constitueixen una modificació especial dels apèndixs de tipus fil·lopodial, que s’oposa als de tipus estenopodial o apèndixs cilíndrics i articulats típics.
Josep M. Barres.
Tret de les antènules, la resta dels apèndixs dels crustacis són birramis, almenys en les primeres fases del desenvolupament. L’estructura bàsica de l’apèndix birrami consta d’una part unirràmia, anomenada protopodi, per mitjà de la qual l’apèndix s’insereix al metàmer corresponent, i és feta de tres artells (de vegades, reduïda a dos o a un). Al damunt del protopodi s’assenten dues branques, una interior o endopodi, i una exterior o exopodi. A partir d’aquesta estructura es poden produir modificacions que fan que arribin a desaparèixer algunes de les parts i se’n desenvolupin d’altres de manera extraordinària. Per exemple, és freqüent que en el protopodi es desenvolupin uns sortints, coneguts amb el nom d’endites quan es troben al costat intern i exites quan són a l’extern. En alguns casos, aquestes protuberàncies creixen considerablement i arriben a tenir el paper més important de tot l’apèndix en relació amb la seva adaptació funcional. Així, en les mandíbules, les endites basals, anomenades gnatobases, s’encarreguen de mastegar l’aliment. En altres casos, l’exita distal adquireix una forma foliàcia o ramificada i fa una funció fonamentalment respiratòria, ja que actua com una mena de brànquia; en aquest cas, s’anomena epipodi. Pel que fa a la desaparició d’alguna d’aquestes parts, és important destacar la de l’exopodi, perquè fa que l’apèndix es mostri unirrami, com és el cas dels crustacis decàpodes.
Malgrat la variabilitat ja esmentada, en general, els apèndixs dels crustacis responen a dos tipus estructurals: l’estenopodial i el fil·lopodial. Els apèndixs estenopodis es caracteritzen per la seva estructura, més o menys robusta, en la qual les diferents parts estan ben articulades i són mogudes per una musculatura potent. Els apèndixs fil·lopodis tenen una estructura molt més laxa i les articulacions menys marcades, de manera que mantenen la seva forma gràcies a la pressió que l’hemolimfa exerceix al damunt de les parets internes. Des del punt de vista de la forma, hom pot dir que els estenopodis generalment són cilíndrics i tenen el tegument dur, mentre que els fil·lopodis són plans i estrets, i tenen el tegument prim i poc calcificat.
Des d’un altre punt de vista, la diferenciació i la diversificació dels apèndixs al llarg del cos es relacionen directament amb una organització avançada. En un primer esquema de diferenciació, cal distingir entre els apèndixs cefàlies i els de la resta del cos.
Els apèndixs cefàlics poden situar-se en una posició anterior respecte de la boca (apèndixs preorals) o posterior (postorals). Els apèndixs preorals són bàsicament sensorials: són les antenes, que es troben en nombre de dos parells en els crustacis (cas únic, en el món dels artròpodes). D’aquests dos parells, el primer constitueix les antènules, que són apèndixs unirramis i multiarticulats. Excepcionalment, en alguns malacostracis, aquest apèndix pot tenir dos i, fins i tot, tres flagels multiarticulats. En els copèpodes les antènules adquireixen una gran importància i diversificació; en efecte, a més d’ésser un òrgan sensorial, les espècies planctòniques les utilitzen per a nedar, i els mascles d’algunes espècies les tenen modificades per a subjectar les femelles durant la fecundació. Les antenes constitueixen el primer parell d’apèndixs birramis i, per tant, són homologables als apèndixs de la resta del cos. Són de funció gairebé exclusivament sensorial, però en alguns grups, com els concostracis, els cladòcers i els ostràcodes, són els principals apèndixs locomotors. En les larves nauplianes, les antenes s’encarreguen també de la natació, cosa que també passa en els estudis larvaris dels anostracis i dels eufausiacis.
Els apèndixs postorals o bucals solen relacionar-se amb l’alimentació. Són de tipus gnatobàsic, és a dir, que realitzen la seva funció per mitjà de les endites, que són extraordinàriament desenvolupades. En canvi, les dues branques de l’apèndix fan un paper secundari i no és rar que es trobin totalment o parcialment atrofiades. En general, hom pot dir que aquests apèndixs són molt plàstics pel que fa a la seva morfologia, com a resposta al tipus d’adaptació alimentària. Les mandíbules se situen immediatament a continuació de la boca i, en principi, són de funció masticatòria tot i que en les larves nauplianes formen part de l’aparell locomotor. En la majoria dels casos, les mandíbules són formades per un protopodi, que porta les endites molt desenvolupades i esclerotitzades. La part distal del protopodi es continua en un epipodi curt, que dona lloc al palp mandibular, el qual de vegades pot mancar. Les maxíl·lules i les maxil·les són d’estructura molt semblant a les mandíbules, és a dir, són formades per un protopodi gros, que de vegades és articulat per a facilitar el moviment. Les branques acostumen a ser poc desenvolupades, especialment l’exopodi. La funció d’aquests apèndixs depèn del tipus d’alimentació. En les espècies filtradores hi ha grups de sedes que fan funció de filtre, i en les carnívores tenen forma de garres amb les quals capturen i manipulen l’aliment. Altres vegades apareixen sedes o branques especialitzades a netejar la resta dels apèndixs, o que produeixen corrents d’aigua vers la boca o les brànquies.
Els apèndixs no cefàlics també presenten diferents estratègies d’adaptació, algunes ben esteses entre els diferents grups de crustacis. D’una banda, els primers pereiopodis tenen tendència a transformar-se en apèndixs bucals auxiliars, anomenats maxil·lípedes el primer parell i gnatopodis la resta, que serveixen per a la manipulació de l’aliment. D’altra banda, la resta dels apèndixs del perèion no implicats en l’alimentació solen relacionar-se amb la locomoció, sigui per a caminar o per a nedar. En alguns grups hi ha adaptacions especials, com als paracàrides, en les femelles dels quals formen, juntament amb les pereionites, la cambra incubadora o marsupi; en general, el grau de diferenciació d’aquests apèndixs depèn del seu nombre, ja que, quan n’hi ha pocs, acostumen a ser diferents, com passa en els cladòcers i els ostràcodes. A més, els apèndixs del plèon o pleopodis solen mancar en la majoria dels crustacis no malacostracis, tret del darrer parell, que sol ésser ben desenvolupat i constitueix el dels uropodis; quan la resta dels pleopodis són desenvolupats tendeixen a transformar-se en apèndixs copuladors en els mascles i ovígers en les femelles.
Organització interna
Els sistemes de relació i control
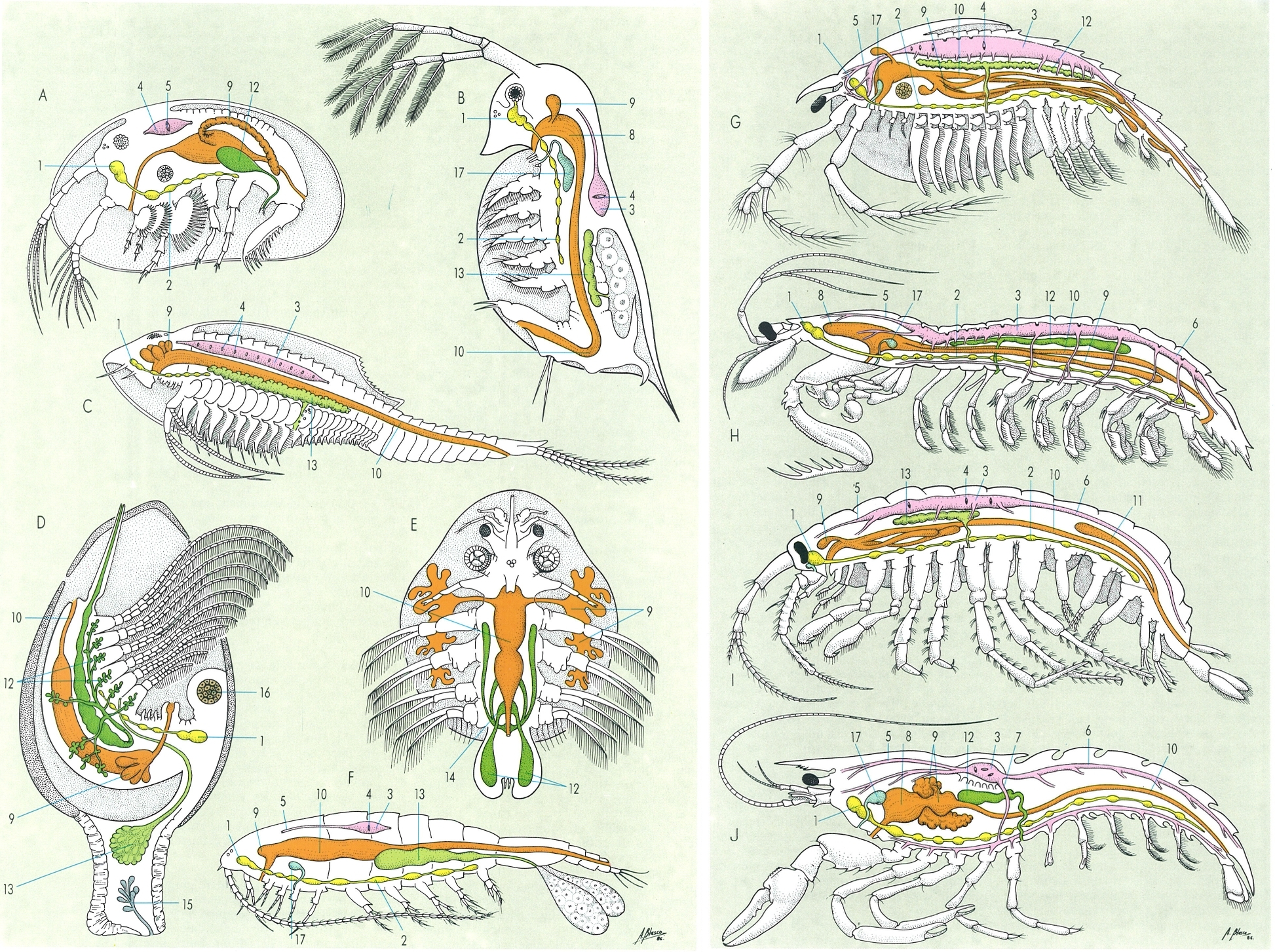
Anatomia de diferents grups de crustacis: A ostràcode, B cladòcer, C notostraci, D cirrípede, E branquiür, F copèpode, G fil·locàride, H hoplocàride, I amfípode, J decàpode. Hom hi distingeix: 1 cervell, 2 cadena nerviosa ventral, 3 cor, 4 osffol, 5 aorta anterior, 6 aorta posterior, 7 aorta descendent, 8 estómac, 9 glàndula de l’intestí mitjà, 10 intestí, 11 cec posterior dorsal, 12 testicle, 13 ovari, 14 vesícula seminal, 15 glàndula de ciment, 16 múscul adductor, 17 glàndula maxil·lar.
Amadeu Blasco, a partir de R. Siewing 1963.
Els sistemes de relació i control en els crustacis tenen una estructura general que correspon a la típica dels artròpodes. El sistema nerviós mostra una estructura bàsica formada per un gangli supraesofàgic, un collar periesofàgic i una cadena ventral doble de ganglis metamèrics, units per comissures. A partir d’aquesta estructura s’estableixen diverses modificacions, les quals mostren una tendència cap a la centralització, tant per fusió de ganglis com per supressió. En els crustacis més primitius (com en els cefalocàrides i els branquiòpodes), els glanglis presenten la màxima diferenciació, ja que en ells el tritocervell encara es troba separat de la massa ganglionar anterior que comprèn el protocervell i el deutocervell. A partir del collar periesofàgic, els ganglis corresponents als metàmers bucals i toràcics encara estan separats, mentre que els abdominals tendeixen a fusionar-se en una cadena única. La centralització del sistema nerviós s’inicia amb la formació d’una massa ganglionar única, que ja incorpora les tres parts del cervell, segueix amb la formació del cefalotòrax i amb la desaparició d’alguns metàmers toràcics i abdominals. Precisament, el grup més evolucionat de crustacis (els decàpodes) mostra dues estructures ben diferents: tots els ganglis ventrals centralitzats en una sola massa (en els braquiürs) i ganglis ventrals molt individualitzats (a la resta del grup).
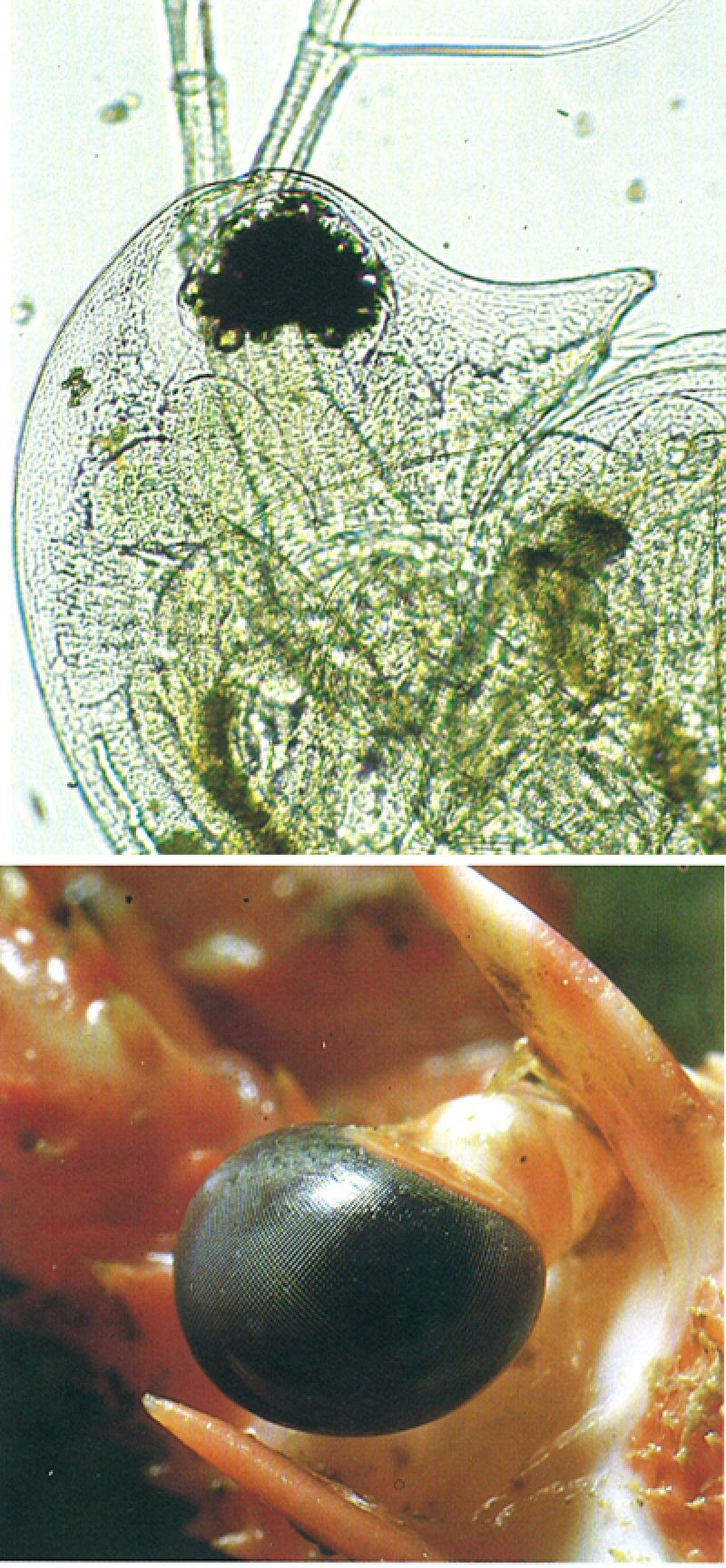
Els ulls dels crustacis presenten diferents configuracions segons els grups, com s’evidencia en els dos exemples il·lustrats. L’ull dels cladòcers, com el de les dàfnies (Daphnia, a dalt), és peculiar ja que és l’únic cas en què els dos ulls laterals han estat fusionats en un de sol i formen una mena de bola, que permet visió lateral i frontal; d’altra banda, hom pot veure clarament a la fotografia la distribució de les facetes al voltant de la concentració de pigment. En els decàpodes, els ulls compostos pedunculats, com els de la llagosta comuna (Palinurus elephas, a baix) són l’exemple del cas més representatiu dels crustacis superiors.
Miquel Blasi i Enric Curto.
L’encèfal dels crustacis mostra tres regions ben desenvolupades, com passa en els insectes i els miriàpodes: el protocervell, el deutocervell i el tritocervell. Cadascun d’aquests centres nerviosos s’encarrega d’innervar una regió definida del cap i els seus corresponents apèndixs. El protocervell innerva els glomèruls dels cossos pedunculats; d’ell surten els nervis òptics i les connexions al sistema glandular del peduncle ocular (òrgan x, glàndula del si) i, tot i que la seva funció no és del tot coneguda, hom li atribueix el control de l’activitat reflexa. El deutocervell és molt desenvolupat gràcies a l’existència dels lòbuls olfactoris, que innerven les terminacions quimioceptores de les primeres antenes, alhora que s’encarrega del moviment d’aquests apèndixs. Finalment, el tritocervell fa una funció semblant però en relació amb el segon parell d’antenes.
El sistema simpàtic té la missió d’innervar l’intestí, el cor, les gònades i la musculatura. La seva estructura és variable i molt poc coneguda fins aquest moment. A grans trets, és format per un gangli que es connecta al collar periesofàgic, i un altre petit gangli labial, que se situa per sota dels lòbuls encefàlics. D’aquests ganglis en surten innervacions que van cap al tub digestiu anterior i mitjà, i cap al cor i les gònades. En els malacostracis, aquest sistema és complementat per una xarxa nerviosa semblant que parteix del darrer gangli abdominal i controla el darrer tram de l’intestí.
A més de rebre estímuls i d’emetre impulsos, el sistema nerviós s’encarrega de regular moltes activitats fisiològiques per mitjà de la neurosecreció. Tota una sèrie de glàndules situades en diferents zones del cos fan, entre altres activitats encara no ben conegudes, el control de les mudes, la regulació de la pigmentació del cos i dels omatidis, de l’activitat del cor i de l’aparició de caràcters sexuals secundaris. En l’actualitat, del sistema endocrí dels crustacis, únicament en coneixem l’estructura i, en part, la funció, en el cas d’algunes espècies de malacostracis de dimensions grans, especialment idònies per a l’experimentació; a tota la resta del grup, i també en les fases del desenvolupament larval i juvenil, en desconeixem les característiques.
Les glàndules o òrgans del sistema endocrí que hom coneix en els decàpodes són les que descriurem a continuació. En primer lloc, la regió medul·lar de l’òrgan X, situada al peduncle ocular, és formada per un grup de cèl·lules nervioses situades a la zona medul·lar, que es projecta des del cervell fins a la base nerviosa de l’ull. En segon lloc, la glàndula del si és una petita cavitat, situada també al peduncle ocular, que rep àxons i conductes secretors de diferents zones de l’encèfal i altres glàndules pròximes; funciona com un lloc on s’emmagatzemen les hormones que segrega la zona de l’òrgan X i les dels ganglis supraesofàgic i ventral, i es troba molt vascularitzada per tal de facilitar l’abocament d’hormones al corrent sanguini. Les papil·les sensorials de l’òrgan X constitueixen un tercer tipus; algunes cèl·lules sensorials de la paret del peduncle ocular projecten terminacions nervioses cap a la zona medul·lar de l’òrgan X, on tenen una activitat neurosecretora; tot i que és una zona histològicament molt complexa (gràcies a la diversitat de cèl·lules secretores), se li atribueix la producció de l’hormona encarregada d’inhibir la producció d’altres hormones a l’òrgan Y, reguladores de les mudes, a més de produir d’altres hormones, reguladores del desenvolupament i la maduració de l’aparell reproductor. Els òrgans postcomissurals se situen a la comissura postesofàgica, on arriben àxons de cèl·lules neurosecretores del tritocervell i, possiblement, de l’estomodeu, i d’on, alhora, parteixen terminacions nervioses vers els sins sanguinis; les hormones que produeix aquesta regió serveixen per a controlar la distribució de pigment en els cromatòfors i els omatidis. L’òrgan pericardial és format per un plexe nerviós situat a la cavitat pericardial, on les cèl·lules secretores es troben en contacte amb la sang; produeix hormones que estimulen l’activitat del cor. Un altre tipus de glàndules són les ventrals que formen l’òrgan Y, i que són dues petites glàndules situades a prop dels músculs de les mandíbules, originades a partir de l’ectoderma de la paret del cefalotòrax; a aquesta zona arriben àxons del gangli subesofàgic que segreguen les hormones encarregades de desencadenar les mudes i regenerar apèndixs. Diferentment als òrgans equivalents d’altres grups d’artròpodes, en els crustacis no desapareixen en l’estat adult, ja que els crustacis muden al llarg de tota la vida. Les glàndules androgèniques són formades per una filera de cèl·lules situades a banda i banda de les terminacions dels conductes deferents, en els mascles; hom els atribueix un paper de reguladores de la diferenciació sexual en el mascle, a partir de la larva. En alguns crustacis, pot haver-hi encara un altre tipus de secreció hormonal: els ovaris segreguen hormones que controlen l’aparició de caràcters secundaris en les femelles.
En els crustacis, els òrgans dels sentits són d’estructura i funció similars, si no idèntiques, als de la majoria d’artròpodes en conjunt, i s’encarreguen de proporcionar informació de l’entorn i de la seva pròpia activitat. Hi ha, però, algunes estructures particulars, com els estatocists, que només es troben en els malacostracis. Els òrgans receptors més comuns són mecanoceptors, que es troben àmpliament distribuïts per tot el cos, i que poden ser de morfologia diferent, amb especialitzacions concretes a diferents estímuls. A l’interior del cos hi ha òrgans equivalents a aquests, però que s’estimulen segons l’activitat muscular; són els anomenats receptors de tensió que, juntament amb els propioceptors o sensors de les articulacions, fan que l’animal tingui informació constant sobre els reflexos posturals i el control de la mobilitat del cos. A més de sensors aïllats, hi ha també òrgans complexos formats per la integració d’unitats senzilles del tipus mecanoceptor. En els crustacis són dels tipus següents: els estatocists, que són una invaginació de la paret del cos que forma com una petita cavitat oberta a l’exterior i entapissada de sedes sensorials, dins de la qual hi ha granets de sorra o alguns cossos durs segregats pel mateix animal que, segons la seva posició, informen de la posició del cos i la direcció del moviment, alhora que actuen com a òrgan d’equilibri; aquests òrgans només es troben a la base de les antenes o a l’abdomen dels malacostracis; els aparells estridulants, presents en alguns decàpodes, els permeten de percebre el soroll que fan picant al terra per mitjà d’estructures complexes formades per grups de sedes sensorials especialitzades. Un altre grup interessant d’òrgans sensorials de la superfície del cos són els quimioceptors, que generalment se situen a les antenes, les antènules i els apèndixs bucals; es tracta d’un sistema sensorial que permet discriminar entre diferents compostos químics. La seva estructura és més complexa que la dels mecanoceptors, però es desconeix la manera com es fa el procés d’identificació dels compostos.
Els ulls són els òrgans sensorials més especialitzats. En els crustacis, com en la resta d’artròpodes, poden ser de dos tipus: ocel·les i ulls compostos. Els ocel·les són les estructures més simples, i generalment s’agrupen formant els ulls centrals del cap, de nombre i estructura diferents segons cada grup. El tipus d’ull central més estès és l’ull nauplià, format per la unió de tres ocel·les, que és el que es troba en totes les larves naupli. Generalment és un ull únic, situat a la part frontal mitjana del cap. En alguns grups de crustacis (els ostràcodes, els copèpodes i els braquiürs) aquest ull és l’únic que persisteix en l’estat adult.
Els ulls compostos són estructures simètriques situades a banda i banda del cap, bé que, secundàriament, es poden desplaçar cap al centre del cos, sense perdre mai la simetria (en cumacis i notostracis) i, ocasionalment, arriben a fusionar-se i fan un sol ull esfèric (en els cladòcers o puces d’aigua). En el desenvolupament larvari, els ulls compostos apareixen més tard que els ocel·les, però acaben desplaçant-los en importància, ja que aquests arriben fins i tot a desaparèixer, tret en el cas que no hi hagi ulls compostos. En alguns crustacis (anostracis i molts malacostracis), els ulls compostos es troben situats a l’extrem d’unes protuberàncies que poden ser mòbils, els peduncles oculars, la finalitat dels quals és doble: d’una banda, donen mobilitat a l’ull i, per tant, augmenten el camp visual, i, de l’altra, són la zona d’ubicació de diferents glàndules neurosecretores. D’altra banda, els ulls compostos també funcionen com a òrgans de posició i d’orientació en relació amb la direcció i la intensitat de llum que reben; aquesta adaptació és especialment important en els crustacis nedadors que, a més, fan migracions verticals diàries, com passa en moltes espècies planctòniques de cladòcers, copèpodes, eufausiacis, misidacis i cumacis, principalment.
En els casos més perfeccionats (en els decàpodes), el pigment ocular és capaç de desplaçar-se segons la llum que percep per mitjà d’un complex sistema de causació hormonal, en el qual intervenen secrecions neuronals. Alhora, tot aquest sistema es troba connectat amb les cèl·lules pigmentàries de la cutícula, de manera que l’animal pot regular la pigmentadó del cos segons els estímuls que percep.
Cada ull compost és format per un conjunt d’unitats independents anomenades ommatidis. El nombre d’ommatidis és molt variable, ja que pot oscil·lar entre 4 (en alguns isòpodes) i més de 10 000 (en decàpodes). Aquesta variabilitat té relació amb l’agudesa visual de cada espècie, però en general les formes nedadores i les carnívores tenen més ommatidis als ulls compostos que no les bentòniques, les filtradores o les herbívores.
L’aparell digestiu
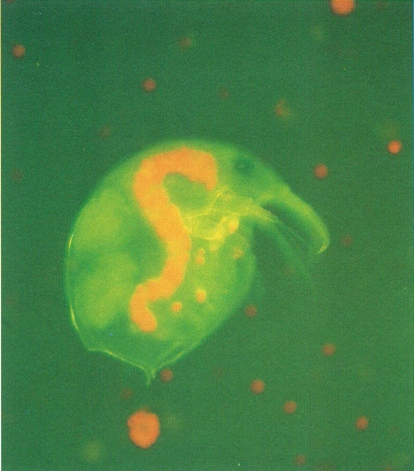
La forma del tub digestiu, en el cas dels cladòcers (a la fotografia, Bosmina longirostris), pot veure’s a través del tegument ja que apareix sovint tenyit de verd a causa del material vegetal acumulat, encara no digerit (principalment, algues clorofícies).
Miquel Blasi.
L’aparell digestiu dels crustacis s’inicia amb dues estructures externes que limiten la boca, pel davant (el labre) i pel darrere (els paragnats), originades respectivament a partir del lòbul cefàlic i de la regió esternal. Com passa en la resta d’artròpodes, hom pot diferenciar en el tub digestiu tres regions diferents: una anterior o estomodeu, originada a partir de l’ectoderma, una de mitjana, gairebé sempre derivada de l’endoderma, i una de posterior o proctodeu, derivada de l’ectoderma. Les dues zones extremes, derivades de l’ectoderma, van cobertes de cutícula.
La part anterior del tub digestiu és la que mostra més variabilitat estructural. En els entomostracis micròfags (com els copèpodes), es redueix a un tub curt voltat d’una forta musculatura que ajuda a deglutir l’aliment. No obstant això, en la majoria de les espècies hi ha sedes filtradores o dents quitinoses que classifiquen l’aliment per grandàries, i la trituradó s’inicia abans d’entrar aquest a l’intestí mitjà. Per tal de facilitar aquesta masticado a l’esòfag hi ha unes dilatacions, semblants a cambres amb diverticles i canals envoltades d’una forta musculatura, que formen el molinet gàstric. Aquestes cambres no són constants en totes les espècies, sinó que poden canviar molt fins i tot entre espècies semblants, si tenen un règim alimentari diferent. En els malacostracis aquest molinet gàstric ateny la màxima diversificació, i arriba a tenir dues cambres ben diferenciades, una d’anterior, o estómac cardíac, i una altra de posterior, o estómac pilòric, que solen especialitzar-se, respectivament, en la trituració i la filtració. Cada cambra porta unes plaques esclerotitzades, dents trituradores, fileres de sedes filtradores, vàlvules (esfínters) i tota una sèrie d’estructures que es mouen per músculs independents, cosa que dona lloc a estructures molt complexes i especialitzades.
L’intestí mitjà s’encarrega de la secreció d’enzims digestius i d’absorbir l’aliment. Per això, apareixen en aquesta zona divertides i cecs que augmenten considerablement la superfície d’absorció. La llargada i el diàmetre de l’intestí mitjà, com el nombre de cecs, és molt variable, i depèn de la qualitat i l’estructura de l’aliment ingerit. En alguns casos és molt curt, de manera que es redueix a l’espai suficient per a poder-s’hi desenvolupar un parell de cecs intestinals. La digestió de l’aliment és externa, per la qual cosa també solen tenir glàndules secretores que es diferencien dels cecs perquè són molt més ramificades. La diversificació d’aquesta zona és molt elevada: en els cladòcers hi ha dos cecs dirigits vers la zona cefàlica, i a la resta de crustacis hi ha un tub que forma un llaç (en les espècies detritívores); en els anostracis hi ha nombrosos cecs lobulats, i en els notostracis i els concostracis els cecs són ramificats.
El tub digestiu posterior presenta una estructura més uniforme: es redueix a un tub estret, mancat de divertides i revestit de cutícula. La seva funció és la de formar els excrements i impulsar-los cap a l’exterior per mitjà de moviments musculars. En aquesta zona es fa la secreció d’un moc que reuneix els excrements en paquets (els anomenats "fecal pellets"), que, en ésser expulsats a l’exterior del cos, cauen cap al fons ràpidament.
L’aparell respiratori
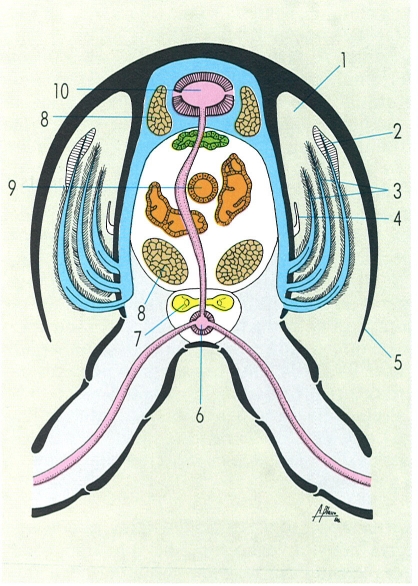
Secció transversal d’un cranc de riu feta a l’alçada del segon parell de pereiopodis que mostra les diferents posicions que poden tenir les brànquies dins la cambra branquial i la seva relació amb el sistema circulatori. Hom hi distingeix: 1 cambra branquial, 2 podobrànquia, 3 artrobrànquia, 4 pleurobrànquia, 5 branquiostegita, 6 artèria subneural, 7 cadena nerviosa, 8 múscul extensor, 9 intestí, 10 cor.
Amadeu Blasco, a partir d’originals diversos.
La respiració, en les espècies de petites dimensions i tegument poc calcificat (copèpodes i malacostracis) es fa a través de tota la paret del cos, cosa que també s’observa en les espècies paràsites. No obstant això, és més aviat excepcional, ja que en general la cutícula dels crustacis és gruixuda i hi apareixen regions primes especialitzades en l’intercanvi de gasos. No hi ha un òrgan respiratori únic. Fins i tot les estructures més comunes, les brànquies, poden tenir un origen molt divers. No hi ha dubte que la diversificació més gran pel que fa a l’origen i l’estructura de les brànquies es fa en els decàpodes, cosa lògica si pensem que són els més grossos i, per tant, els que tenen unes necessitats absolutes d’oxigen més grans.
Sigui quin sigui l’origen de les brànquies, sempre es tracta d’expansions del tegument, més o menys ramificades, en les quals la cutícula és molt fina, de manera que la hipodermis es troba a prop de la superfície i, per tant, hi ha una gran vascularització. Hi ha, també, altres zones del cos que tenen aquestes característiques i la mateixa funció, però no adopten forma de brànquia, com és el cas de la paret interna de la closca, que és un sistema de respiració especialment important en els concostracis, els cladòcers, els ostracodes i els cirrípedes, és a dir, en aquells grups que mostren el cos tancat dins d’unes valves o plaques. Una tendència que s’observa en diferents grups de crustacis és la de situar les superfícies respiratòries en posició cada vegada més interna, protegides, a més, per alguna cobertora; això és el que ha originat les cambres branquials, que només són cavitats formades per l’expansió del tegument per a cobrir les brànquies. Aquesta adaptació ha suposat, a més, l’aparició d’apèndixs que impulsen l’aigua cap a la cambra branquial i en renoven la de l’interior; en els cumacis, els misidacis, els eufausiacis i els decàpodes, aquesta funció la fan els endopodis del segon parell de maxil·les. La respiració per mitjà dels pleopodis és més aviat excepcional en els crustacis: únicament els isòpodes i els estomatòpodes són capaços d’efectuar-la. D’altra banda, els ostràcodes són els únics que fan servir l’abdomen, a més d’altres zones del cos, com a superfície respiratòria.
Les brànquies dels decàpodes es mostren diversificades. Segons la seva localització al cos se’n distingeixen tres tipus: les pleurobrànquies, que són expansions de la paret lateral del cos; les artrobrànquies, que són membranes articulars entre la paret del cos i l’apèndix; i les podobrànquies, que són zones situades a l’apèndix.
La respiració aèria no ha suposat gaires canvis en l’estructura de l’aparell respiratori, ja que tant els isòpodes com els decàpodes han aprofitat les cambres branquials, que han passat a una situació més interna al cos. Alhora, han desenvolupat noves estructures, diferents a l’un i l’altre grup. En els isòpodes hi ha pseudotràquees, molt ramificades, situades als exopodis dels pleopodis. Els decàpodes terrestres han resolt el problema d’una manera més imperfecta, a base de recobrir la cambra branquial amb papil·les molt vascularitzades, que intercanvien gasos directament de l’aire a un ambient saturat d’humitat.
L’aparell circulatori
El sistema circulatori dels crustacis és format, bàsicament, per un tub dorsal que corre per sobre del tub digestiu, proveït d’un eixamplament, que és el cor. En la seva forma més simple, el cor, inclòs en el si pericardial, presenta una o més obertures anomenades ostíols, i ocupa una bona part de la llargada del cos. Del cor pot sortir una aorta anterior, que es dirigeix cap a la regió encefàlica, i de vegades també una aorta posterior, que arriba fins a l’extrem del plèon. La resta del sistema circulatori és obert i la sang es mou a través de sins i de llacunes hemocèliques, que banyen les vísceres, abans de retornar al cor a través del si pericàrdic i dels ostíols. A partir d’aquest esquema, general en els grups més primitius (com els branquiòpodes anostracis), hi ha nombroses simplificacions secundàries en relació amb les dimensions del cos, que afecten principalment els no malacostracis. Una primera tendència d’aquest procés de reducció és la del cor, el qual originàriament és allargat i envoltat d’una musculatura espiral que produeix les contraccions; en els grups més primitius el nombre d’ostíols és elevat (18 parells en els anostracis i 14 parells en els notostracis), però es va reduint en altres grups fins arribar a un sol parell (en els cladòcers, els ostràcodes i els branquiürs). Excepcionalment, aquesta tendència arriba a casos límit, com en alguns grups de copèpodes, als quals el cor desapareix i la circulació es fa per mitjà de moviments de l’intestí. Encara més excepcionalment, en els cirrípedes és tot el sistema circulatori el que ha desaparegut, cosa que no és tan rara en les espècies paràsites.
El sistema arterial també presenta una certa variabilitat. En general és poc desenvolupat o manca totalment en els grups que tenen el cor situat a prop del cèfalon. No obstant això, quan aquest es desplaça cap a la part posterior del cos (per exemple, en calanoides i en branquiürs), hi ha una aorta anterior que condueix l’hemolimfa cap a les proximitats de l’encèfal.
L’hemolimfa és un fluid pàl·lid, en el qual el pigment que transporta els gasos és l’hemocianina. Ocasionalment, hi ha una certa concentració d’hemoglobina, que pot emmascarar l’hemocianina. Això és especialment important en les espècies que viuen en ambients amb concentració baixa d’oxigen, com passa en algunes del gènere Daphnia, pròpies de la zona profunda d’alguns llacs. Per l’hemolimfa circulen diferents tipus de leucòcits, els quals es distingeixen per la seva morfologia.
Els malacostracis mostren un sistema circulatori peculiar, molt desenvolupat com a conseqüència, principalment, de la necessitat de transportar l’oxigen ràpidament i a una certa distància de les brànquies. En primer lloc, cal destacar que la grandària del cor i el nombre d’ostíols són diferents segons el grau d’evolució del grup: en els més primitius, el cor és gran i porta nombrosos parells d’ostíols (13 en els leptostracis), mentre que en els evolucionats és petit i amb pocs ostíols (fins a un mínim de tres parells). En segon lloc, han desenvolupat considerablement el sistema arterial i, a més de les artèries anterior i posterior, n’hi ha unes quantes de laterals, que surten a banda i banda del cor o bé de les mateixes artèries anterior i posterior; el nombre d’aquestes artèries és variable, des d’un màxim de 13 en els leptostracis fins a un minin d’una en els eucàrides. Les artèries laterals es divideixen ràpidament en dues branques, que es dirigeixen cap als apèndixs del perèion i el plèon (la branca apendicular) i cap a l’intestí, les gònades i, de vegades, al sistema nerviós (la branca visceral). L’artèria o aorta anterior sol ser llarga, cosa que obliga en la majoria dels grups (tret dels cumacis i els tanaidacis) a l’existència d’un segon òrgan pulsador a l’altura de l’estómac, l’anomenat cor anterior, el qual restableix la pressió arterial perduda per la fricció de l’hemolimfa al llarg de tot el seu recorregut; aquesta artèria desemboca, com en la majoria dels crustacis, a les proximitats de l’encèfal, al qual irriga. A la cara ventral del cos i immediatament per sota del cordó nerviós corre l’artèria neural, que és la que irriga els ganglis nerviosos; neix de la branca visceral d’una de les artèries laterals. Quan només hi ha una artèria lateral, cosa que passa en els decàpodes i, per tant, manquen les corresponents branques apendiculars, els apèndixs són irrigats per branques de l’artèria subneural. Tot i que en els malacostracis el sistema circulatori és més complex, també és obert, com a la resta dels crustacis, ja que les artèries van a parar als sins i llacunes pels quals l’hemolimfa flueix als òrgans. Per exemple, en els decàpodes, l’hemolimfa flueix de les branques de l’artèria neural cap als sins esternals i laterals, on s’irriguen les brànquies, i després passa al si pericàrdic i torna a entrar al cor a través dels ostíols per tal d’iniciar un nou cicle.
Els òrgans excretors
Els òrgans excretors més importants dels crustacis són les glàndules antenals i les maxil·lars, bé que altres zones del cos poden tenir superfícies per les quals hom ha detectat activitat excretora. Malgrat la semblança d’origen, estructura i funcionament d’ambdós tipus de glàndules, és molt rar que es trobin juntes en estat funcional. La glàndula antenal és l’òrgan excretor predominant en les larves nauplianes i també es troba present en els adults dels eucàrides i alguns peracàrides. La glàndula maxil·lar, per contra, es troba en els adults de la majoria dels crustacis no malacostracis, i en els cumacis, tanaidacis i isòpodes. Finalment, alguns altres grups (com els ostràcodes i els leptostracis) manquen d’un i de l’altre tipus.
Les glàndules antenals i maxil·lars són, en realitat, nefròcits d’origen celòmic i d’estructura senzilla. A partir del celoma es forma un sac terminal, en el qual se situen les cèl·lules excretores, que desemboca en un nefroducte —també celòmic— al seu costat distal, que condueix a una bufeta urinària proveïda d’un por excretor, el qual s’obre a la base de l’apèndix corresponent. La variabilitat d’aquesta estructura depèn de la grandària del sac terminal i de la llargada i la diferenciació de zones en el nefroducte. En els malacostracis, el sac terminal té molts plecs que augmenten considerablement la superfície excretora, i el nefroducte presenta en el seu tram més intern una gran dilatació, anomenada glàndula verda, que té una gran activitat excretora. En canvi, la llargada del nefroducte es relaciona molt més amb la salinitat de l’aigua del medi on viu cada espècie; en les espècies d’aigua dolça és molt més llarg que en les d’aigua marina.
Altres zones del cos poden tenir també una certa activitat excretora, bé que d’una manera secundària respecte a alguna altra funció; per exemple, en els círrípedes hom creu que hi ha excreció a través de la paret de l’intestí mitjà. També pot ser important, segons que sembla, l’excreció a través del tegument, especialment en aquelles zones del cos on la cutícula és poc calcificada, com passa en les brànquies i en tots els epitelis respiratoris.
Els crustacis excreten compostos nitrogenats en forma d’amoníac. Es tracta d’un sistema poc evolucionat, que possiblement ha limitat l’adaptació dels crustacis al medi aeri. Una part molt important de l’activitat excretora es relaciona amb l’osmoregulació, cosa que fa que cada espècie hagi desenvolupat les seves pròpies adaptacions, les quals depenen més de l’ambient on viuen que no del grup al qual pertanyen. Com a exemples il·lustratius, podríem citar les adaptacions dels decàpodes a l’aigua dolça en la figura del cranc de riu, o les d’altres espècies que viuen en aigües hiperhalines, com l’artèmia (Artemia salina).
L’aparell reproductor
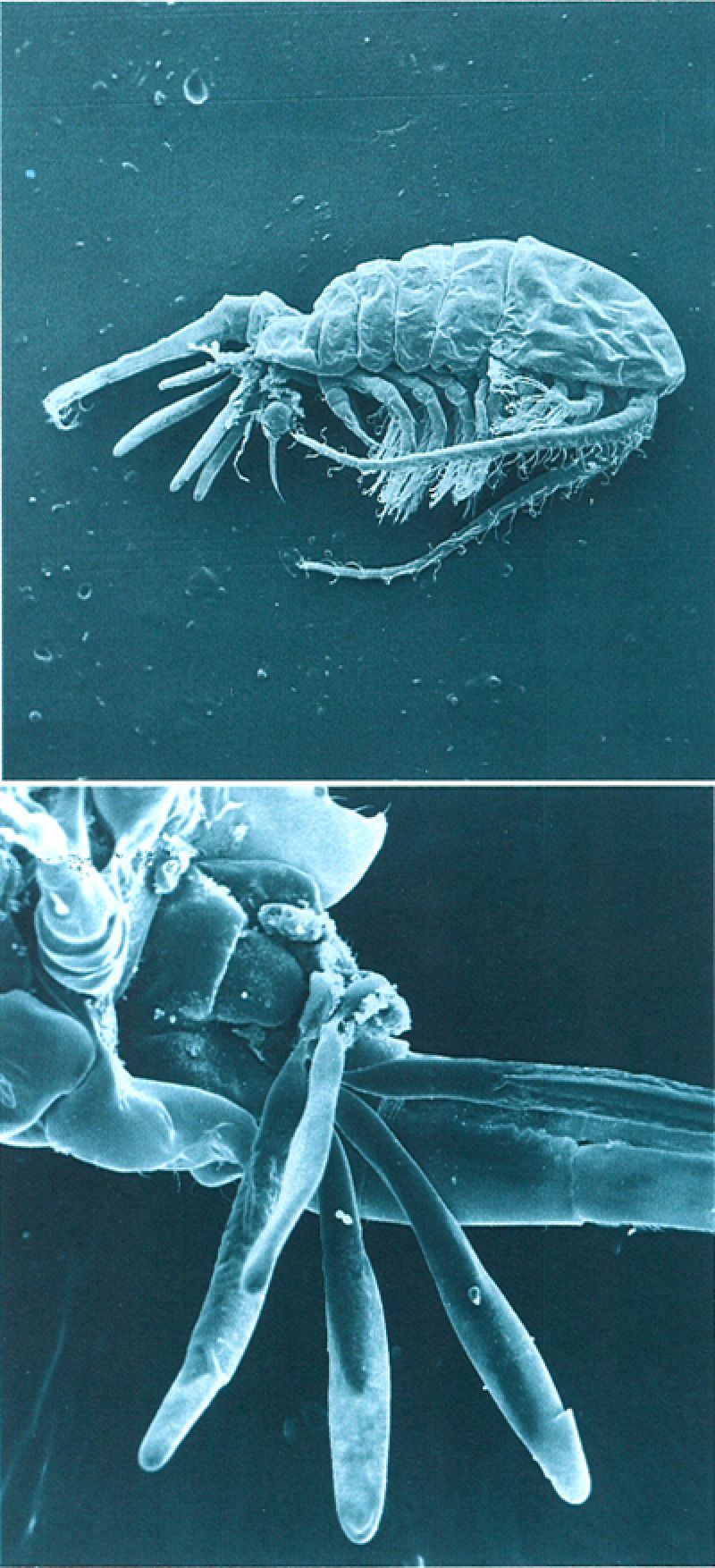
La formació d’espermatòfors és característica d’alguns grups, com el dels decàpodes i el dels copèpodes. Així, no és rar de veure, en una mostra de plàncton, femelles de copèpode amb uns quants espermatòfors (a dalt) provinents de diferents mascles, i d’unes dimensions considerables, adherits a la seva part posterior (a baix, en detall). Hom desconeix encara la manera com els espermatozoides, desproveïts de moviment, deixen l’espermatòfor per a arribar fins a l’òvul.
Mercè Durfort.
L’aparell reproductor s’organitza, bàsicament, d’una manera semblant en tots els crustacis: un parell de gònades tubulars, de les quals neixen dos gonoductes, que s’obren a l’exterior per mitjà d’uns gonopors situats a l’altura del perèion. Les gònades i una part important dels gonoductes són originats a partir del celoma i se situen lateralment al tub digestiu a la regió del cefalotòrax o del perèion. A partir d’aquest esquema es poden produir algunes modificacions, com la tendència dels dos testicles a fusionar-se en un de sol, i també dels dos gonoductes, que es poden fusionar totalment o parcialment al llarg del seu recorregut. Altres modificacions són degudes a canvis introduïts en el pla d’organització del cos. Per exemple, en les espècies de vida paràsita, les gònades es desenvolupen extraordinàriament i poden arribar a ocupar una bona part del cos; en els ostràcodes, a causa de la reducció i simplificació de la segmentació del cos, una gran part de les vísceres, incloses les gònades, se situen entre les parets de la closca; finalment, en els cirrípedes toràcics, l’hermafroditisme ha portat al desplaçament dels ovaris cap a la zona del peduncle o del disc basal, mentre que els testicles queden situats en posició normal, al costat del tub digestiu.
Els gonoductes acompleixen una funció més complexa que no sols la de transportar els òvuls i els espermatozoides a l’exterior del cos. Una part de les cèl·lules epitelials que recobreixen el seu interior són de tipus glandular i produeixen secrecions, importants per a preservar les cèl·lules germinals. De vegades, aquestes cèl·lules es poden diferenciar del propi gonoducte i actuar com a glàndules, més o menys independents físicament, però en una relació estreta a través de les secrecions que produeixen. Als mascles d’alguns grups (com els copèpodes), els espermiductes produeixen espermatòfors, que són càpsules protectores de les masses d’espermatozoides. A les femelles, els gonoductes juguen un paper semblant en relació amb la protecció de l’òvul, ja que, a més de segregar el vitel·lus, també produeixen una coberta quitinosa que protegeix els ous destinats a resistir períodes de dessecació, o els ovisacs, dins dels quals algunes femelles transporten els ous fins al moment de l’eclosió.
La situació dels gonopors en el cos dels crustacis és variable, i és un caràcter que separa diferents grups taxonòmics, com per exemple les subclasses. Tret d’algunes excepcions, l’obertura genital dels mascles sol situar-se al darrer metàmer del perèion, i el de les femelles, quan no coincideix en el mateix metàmer, se situa en posició una mica anterior.
Biologia
Reproducció
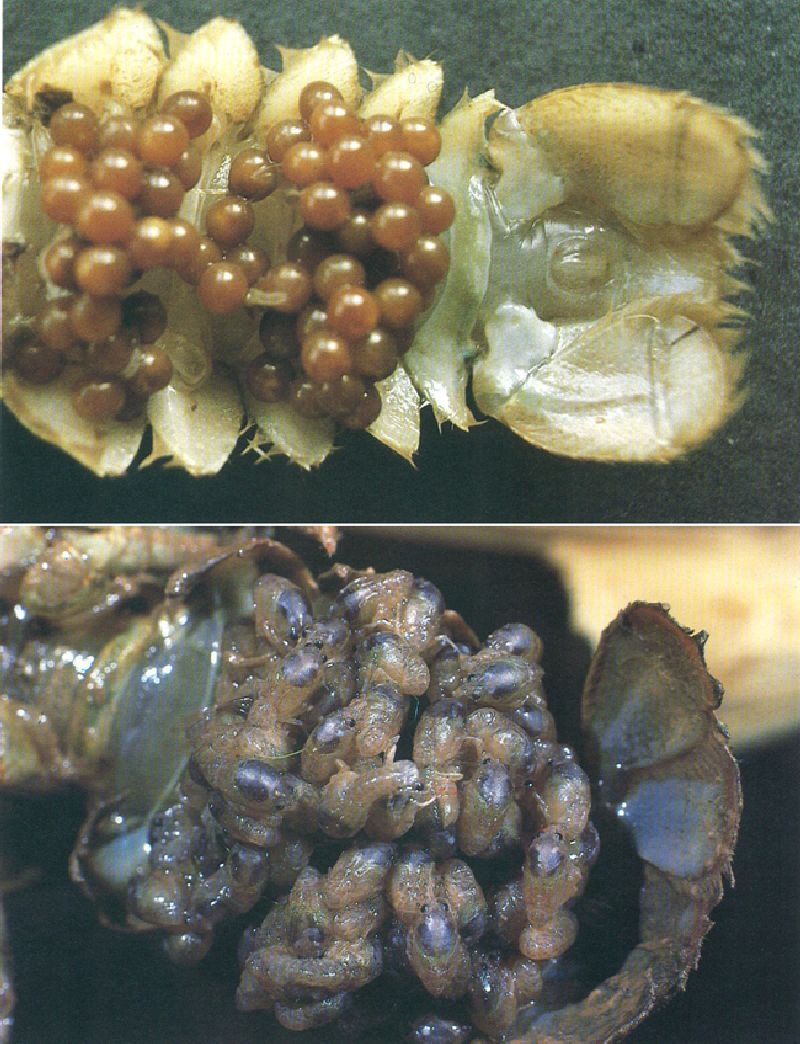
La fixació dels ous fecundats a la part ventral del perèion és gairebé general dins dels crustacis decàpodes, com mostra la fotografia (a dalt) d’un cranc de riu (Austropotamobius pallipes lusitanicus). Tanmateix, els crancs de riu són els únics crustacis que presenten desenvolupament postembrionari directe, és a dir, que dels ous neixen petits individus morfològicament molt semblants als adults (a baix), en lloc d’aparèixer les larves nedadores que caracteritzen la resta dels crustacis.
Josep M. Barres i Oriol Alamany.
En general, en els crustacis els sexes són separats, és a dir, que hi ha individus mascles i individus femelles; només en alguns casos hi ha hermafroditisme, com en els cirrípedes, cosa que es relaciona amb el tipus de vida sèssil o paràsita, de la majoria de les seves espècies. En altres grups (branquiòpodes, ostràcodes, tanaidacis, isòpodes i alguns decàpodes) hi ha espècies que tenen una alternança entre la reproducció sexual i la partenogenètica, cosa que s’ha interpretat com una adaptació a la vida en condicions molt fluctuants; per exemple, a les aigües continentals és on aquest fenomen és més manifest, especialment en les espècies que viuen a les aigües temporals, de tal manera que la partenogènesi s’ha associat a la producció d’ous de desenvolupament ràpid i la reproducció sexual a la producció d’ous capaços de resistir la dessecació (ous durables).
Les femelles solen tenir alguna estructura especial per a transportar els ous, de morfologia diversa, com els marsupis, les cambres incubadores, estructures especials situades entre els pereiopodis o els pleopodis, ovisacs, la cavitat del mantell, etc. Sovint aquest transport continua després de l’eclosió dels ous, de manera que, moltes vegades, quan els joves abandonen la mare, ja han passat bona part del seu desenvolupament postembrionari.
Els branquiürs i els anaspidacis, en la majoria dels eufausiacis i ostràcodes, i en alguns copèpodes i decàpodes, solen deixar els ous lliures a l’aigua o a la superfície de pedres o de vegetació submergida.
Desenvolupament embrionari
La manera com es fa la segmentació de l’ou en els crustacis és un tema d’una gran controvèrsia, ja que hi ha una gran variabilitat. En l’actualitat hom accepta que, diferentment d’allò que passa en els altres artròpodes, en els crustacis la segmentació és total, desigual i espiral a les primeres fases. Tot i que el grup que presenta aquest tipus de desenvolupament embrionari d’una manera més característica és el dels cirrípedes, s’observa una organització similar a la majoria dels branquiòpodes, els ostràcodes, copèpodes, els anaspidacis, els eufausiacis i alguns, molt pocs, decàpodes. En aquests grups, l’ou és de tipus telolecític, és a dir, que té el vitel·lus acumulat al pol vegetatiu i, després de la segmentació, apareix una blàstula buida per dins, que no sembla tenir relació amb la formació d’una primera larva nauplíana. En els grups de crustacis més evolucionats, els ous són centrolecítics i la segmentació és superficial, tot i que també s’han observat formes de transició, amb una segmentació mixta total-superficial. En qualsevol cas, cal insistir en el fet que la segmentació és molt variable, cosa que, paradoxalment, contrasta amb la gran uniformitat de desenvolupament en les fases posteriors a la de blàstula. En efecte, aquesta uniformitat és tan marcada que els mapes de destinació presumible que indiquen la descendència de cada blastòmer ja és definit en la fase de blàstula, i constitueix una característica definitòria dels crustacis.
Desenvolupament postembrionari
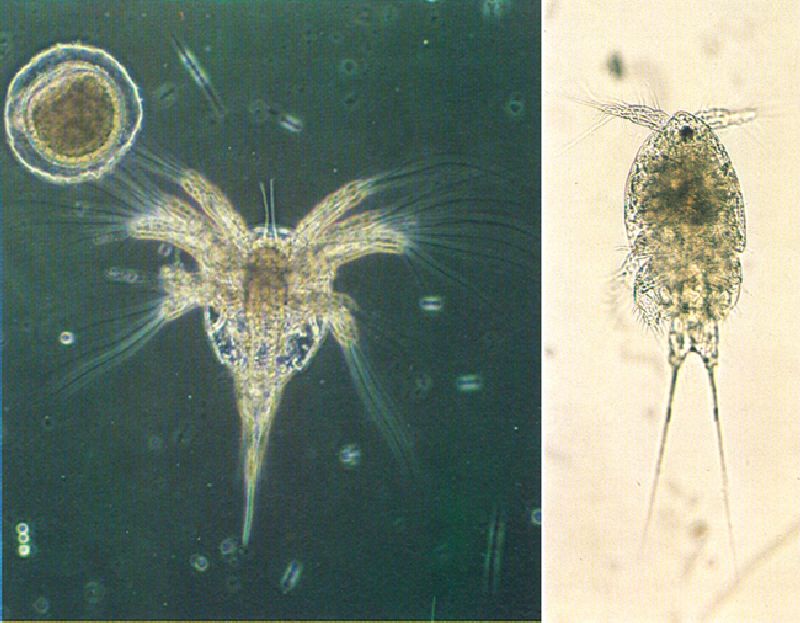
La forma larval que segueix a la fase d’embrió en la majoria dels crustacis és la de naupli, que pot prendre la forma de vida lliure (a l’esquerra), o bé pot transcórrer a l’interior de l’ou. A partir d’aquesta fase se succeeixen altres formes larvats, algunes peculiars d’uns grups determinats: és el cas de la larva magalopa, exclusiva dels decàpodes, dels ostracodits dels ostràcodes, o dels copepodits (a la dreta) exclusius dels copèpodes (noteu en aquesta fotografia l’ull únic, en posició central i pigmentat de vermell, que caracteritza les larves).
Xavier Palaus i Mercè Durfort/Xavier Vidal
El grau de desenvolupament que arriben a atènyer els embrions abans de sortir de l’ou és molt variable; depèn, en gran mesura, de la forma de vida de cada espècie.
En alguns casos, de l’ou surten individus de morfologia molt semblant als adults i amb un mateix nombre de metàmers; en aquest cas hom parla de desenvolupament directe i epimòrfic. Aquest tipus de desenvolupament no és molt estès entre els crustacis i, pel que sembla, respon a un tipus de vida força particular, en el qual la rapidesa amb què es produeixen les noves generacions és l’estratègia fonamental per al seu èxit. Els cladòcers en són el millor exemple, bé que també presenten un desenvolupament més o menys directe d’altres grups, com els leptostracis, els anaspidacis, força espècies de peracàrides i alguns decàpodes.
En la majoria dels crustacis, el desenvolupament és anamòrfic, ja que de l’ou surten individus de morfologia diferent a la dels adults i amb un nombre de metàmers inferior al d’aquests, que han de passar diferents estadis larvaris abans no completin el seu desenvolupament.
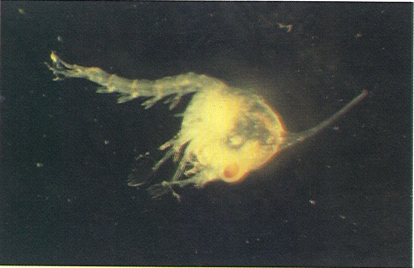
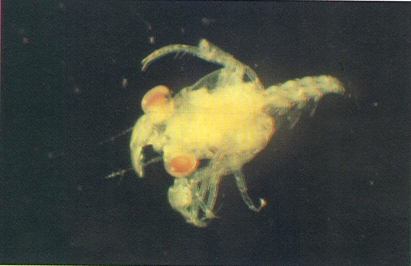
Les fases larvals dels decàpodes representen estadis diferents de desenvolupament, amb formes prou allunyades de les que caracteritzaran el crustaci adult. La il·lustració recull una larva en la fase de zoe (a dalt), i una altra en la fase de megalopa (a baix), aquesta darrera de braquiür.
Josep M. Gili
El nombre de formes larvàries pot variar molt segons cada grup i, naturalment, depèn del moment en què s’interromp el desenvolupament embrionari dins de l’ou. Tot sovint la primer fase larvària correspon a una larva anomenada naupli, que té una estructura molt simple, amb només tres parells d’apèndixs funcionals (antènules, antenes i mandíbules) i els seus tres corresponents metàmers. Aquesta larva, que té una forma ovalada o arrodonida, és la més típica i la més estesa dins dels crustacis, ja que forma part del desenvolupament dels ostràcodes, els copèpodes, els branquiürs, els cirrípedes, els eufausiacis i alguns decàpodes molt primitius. La fase següent a la de naupli és la de metanaupli, que té el mateix nombre d’apèndixs funcionals, però ja té nous esbossos meta mèrics i també d’altres apèndixs en formació incipient; els cefalocàrides, els mistacocàrides i els anostracis, notostracis, concostracis i el cladòcer Leptodora kindti (dins dels branquiòpodes) surten de l’ou en aquest estadi larval. La fase del metanaupli, però, no la passen tots els grups de crustacis, sinó que després del naupli poden aparèixer també altres formes larvàries, característiques de cada grup.
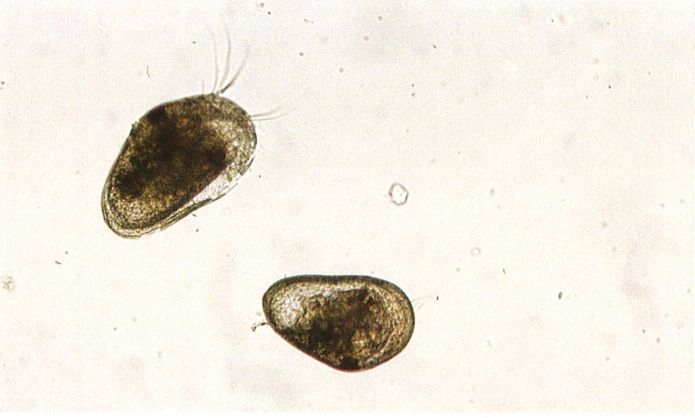
Els ostracodits, pertanyents a l’espècie Cypridopsis newtoni en el cas de la fotografia, són la fase de desenvolupament intermèdia entre la larva naupli i la d’adult. Externament els caracteritza la forma de la part posterior del cos, més estreta que l’anterior.
Mercè Durfort.
En el cas dels copèpodes, després de la fase del metanaupli apareix una fase anomenada de copepodit, peculiar del grup, que es caracteritza pel fet de tenir la forma de l’adult, però sense tenir els metàmers del plèon totalment desenvolupats ni tampoc els apèndixs del perèion. D’altra banda, després de la larva naupli, els cirrípedes i els ostràcodes tenen un estadi larvari exclusiu, la larva cipris, que té la closca bivalva molt desenvolupada, tant que cobreix totalment el cos de l’animal. En els malacostracis, després de la fase de naupli (quan n’hi ha, que no és sempre) apareix una fase de zoe, a la qual la larva ja té pereiopodis funcionals, el plèon en metamerització molt avançada i els ulls compostos laterals; aquesta fase té una gran diversitat d’estadis, amb lleugeres diferències morfològiques segons el nombre de mudes sofert, que canvien també segons cada grup taxonòmic. Com a conseqüència d’aquesta gran diversitat, sovint es designen amb noms específics: s’utilitzen els termes protó-, meta-, eu-, meso-, epi- i antizoe, o també, pro-, proto-; meso- i metamisi per a designar algunes de les larves que es diferencien en aquesta fase. En els decàpodes branquiürs encara hi ha, a més, una darrera fase larvària, que correspon al període en què tots els apèndixs ja són funcionals (pleopodis inclosos), però encara no s’ha assolit l’estadi d’adult; aquesta fase és anomenada per alguns zoòlegs fase de postlarva, però en realitat el nom més estès és el de megalopa.
Ecologia
Els crustacis són majoritàriament aquàtics, cosa que contrasta amb la resta de grans grups o classes d’artròpodes. De les gairebé 30 000 espècies de crustacis conegudes, menys de l’1, 1 % han passat a la vida terrestre i, encara, en condicions molt precàries. Els isòpodes i els decàpodes són els grups de crustacis que contenen més espècies terrestres (prop d’un miler cadascun), mentre que entre els amfípodes, ostràcodes i els cladòcers la representació de formes aèries és, més que res, simbòlica. A l’aigua és on els crustacis han demostrat una capacitat d’adaptació més gran, especialment a la mar. En efecte, dels grans grups de crustacis, únicament els branquiòpodes són majoritàriament d’aigua dolça; els cefalocàrides, els remipedis, els mistacocàrides i els cirrípedes només tenen representants marins, mentre que la resta dels grans grups (ostràcodes, copèpodes, branquiürs i malacostracis) es troben distribuïts tant pel mar com per les aigües dolces.
La vida a les aigües dolces significa una adaptació a condicions fluctuants, una gran capacitat per a resistir la dessecació estival de l’aigua i, alhora, una gran facilitat per a dispersar-se i colonitzar nous ecosistemes. A la mar, en canvi, la variabilitat de condicions ambientals és reduïda, i gairebé tot l’esforç evolutiu s’ha dedicat a l’especialització en determinades funcions tròfiques o en formes de vida específiques; el resultat d’aquesta especialització a la mar es pot resumir indicant que pràcticament no hi ha cap ambient marí que no sigui ocupat per crustacis. Les espècies pelàgiques, especialment els copèpodes i els eufausiacis, es troben en els primers graons de totes les cadenes tròfiques oceàniques, mentre que les bentòniques, especialment els peracàrides, amb una alimentació sovint de tipus detritívor, juguen un paper molt important en el reciclatge de la matèria. Aquesta plasticitat adaptativa també queda reflectida en la gran diversitat d’espècies paràsites: prop de 4000 espècies de crustacis són paràsites, que exploten tota mena d’animals marins i, fins i tot, altres espècies de crustacis; els copèpodes, amb 3000 espècies paràsites, són el grup més diversificat, bé que els cirrípedes també tenen representants paràsits i els branquiürs constitueixen l’únic grup exclusivament de vida paràsita.
En el bentos, ultra els cirrípedes, que fan vida sèssil, és a dir, que viuen fixats damunt del substrat del fons, hi ha molts altres grups de crustacis que es mouen sempre molt a prop del fons. Tampoc no és infreqüent el cas de les formes que s’amaguen en galeries, que s’alimenten o simplement viuen en aquest ambient, i constitueixen una part important d’allò que hom anomena la fauna intersticial. Precisament, la comunicació que hi ha entre les aigües intersticials marines i les continentals ha facilitat la colonització de les aigües subterrànies per algunes espècies d’origen marí. De fet, la intercolonització entre l’aigua de mar i les aigües subterrànies continentals és un procés que continua, però que poques vegades ha tingut èxit. Per exemple, els cladòcers, tot i que tenen diverses espècies marines, mai no han assolit a la mar la importància que tenen en les aigües continentals. També per a les espècies marines, la colonització de les aigües continentals representa un procés difícil, que ha estat intentat repetidament per alguns grups com, per exemple, el dels copèpodes. En la majoria dels casos, hom considera que el principal factor limitant és la manca d’adaptació a les condicions ambientals, però en realitat pot ser-ho l’existència d’espècies competidores que ja es troben adaptades i que frenen l’evolució de les espècies invasores i les restringeixen als llocs d’origen. Els millors exemples d’invasió s’han produït després de grans canvis climàtics (les glaciacions), que suposaren la desaparició de moltes de les espècies potencialment competidores.
Pel que fa a la distribució dels crustacis com a grup, hom pot parlar d’una tendència al cosmopolitisme en el medi aquàtic, cosa que es pot considerar com una prova de l’èxit que han obtingut en aquest ambient. No obstant això, alguns grans grups de crustacis, malgrat el seu interès sistemàtic i filogenètic, mostren una escassa diversificació i una distribució limitada; per exemple, els cefalocàrides i els remipedis, els grups de sincàrides i pancàrides i els espeleogrifacis mostren una distribució molt restringida a determinades zones geogràfiques. En relació amb aquest darrer punt, cal indicar que el nostre coneixement dels crustacis és encara limitat i que l’exploració de nous ambients o l’observació minuciosa d’altres ambients ja coneguts fan que es descobreixin noves espècies, de característiques fins ara desconegudes, que obliguen els zoòlegs a crear nous tàxons per a situar-los dins de l’esquema sistemàtic dels crustacis. Potser la subclasse dels remipedis constitueix l’exemple més recent d’aquest fenomen, i alhora el que ens és relativament més pròxim, ja que la seva primera espècie fou descoberta en una cova submarina de les Bahames l’any 1981, i la segona, i darrera fins ara, es trobà també en una cova submarina (la cova de Los Jameos del Agua), aquest cop de Lanzarote, l’any 1984.
Sistemàtica
La classificació dels crustacis constitueix un tema d’un cert interès pel fet que, en l’actualitat, es troba en una fase de revisió. D’una manera sintètica, hom pot dir que la sistemàtica dels crustacis és sotmesa a dues pressions: l’una externa, que es produeix com a resultat de l’elecció entre diverses teories filogenètiques que hi ha al voltant de l’origen dels artròpodes, i l’altra interna, que és resultat de l’agrupament dels crustacis en tàxons menors segons diferents criteris morfològics.
Ja hem discutit les diferents teories que actualment s’accepten per a explicar l’origen dels artròpodes, i les conseqüències que se’n deriven per a la seva classificació. L’elecció de la teoria monofilètica porta implícita la consideració dels crustacis com una classe i també l’existència de la categoria de subclasse immediatament inferior. Malgrat això, alguns autors han elevat els crustacis a la categoria taxonòmica de subfílum, fílum o superclasse i, en aquest cas, s’utilitza la de classe per als nivells immediatament inferiors de la seva classificació.
La problemàtica que planteja la classificació interna dels crustacis es deu, en gran mesura, a la seva gran variabilitat. La mateixa diagnosi dels crustacis, com a grup, mostra fins a quin punt és difícil trobar característiques comunes a tots; aquesta manca de criteris sistemàtics clars pot explicar en gran part les vicissituds per les quals ha anat passant la classificació dels crustacis al llarg del temps.
Les unitats principals
La divisió dels crustacis en entomostracis i malacostracis com a sinònim de crustacis "inferiors" i "superiors" respectivament, va persistir fins a començament de segle. En l’actualitat, malgrat que s’accepta el caràcter artificial dels entomostracis com un grup zoològic, encara és corrent que s’utilitzi, com a denominació vulgar per a fer referència als crustacis no malacostracis. La base de classificació moderna del grup fou establerta per Calman l’any 1909 i divideix els entomostracis en cinc subclasses: branquiòpodes (Branchiopoda), ostràcodes (Ostracoda), copèpodes (Copepoda), branquiürs (Branchiura), i cirrípedes (Cirripedia), als quals se sumava ja, com a subclasse, la dels malacostracis (Malacostraca). Posteriorment, i d’acord amb els nous descobriments, s’han anat afegint tres noves subclasses, compostes de poques espècies, de descripció recent: els cefalocàrides (Cephalocarida), els remipedis (Remipedia) i els mistacocàrides (Mistacocarida).
En contrast amb l’artificiós grup dels entomostracis, els malacostracis constitueixen un grup homogeni, amb relacions morfològiques ben definides i amb el grau d’evolució més alt dins dels crustacis. Aquest grup comprèn al voltant de 22 000 espècies i, amb la categoria taxonòmica de subclasse, comprèn sis grans grups d’una gran versatilitat d’adaptació a tots els ambients aquàtics. És entre els malacostracis que hi ha les úniques espècies de crustacis que han colonitzat el medi terrestre d’una manera temporal (els crancs del litoral marí) o permanent (els porquets de sant Antoni). Aquesta gran versatilitat adaptativa al medi ha portat a una gran diversificació anatòmica, malgrat la qual el grup manté la seva unitat morfològica. En efecte, d’una manera característica, els malacostracis tenen el cos dividit en un nombre gairebé constant de segments (20 o 21), repartits en regions o tagmes de la manera següent: 6 segments al cèfalon, 8 segments al perèion i set segments al plèon en els fil·lòpodes i els misidacis lofogàstrides, i sis a la resta dels grups. També és constant i característica la distribució i la quantitat dels apèndixs dels diferents tagmes: al cèfalon hi ha un parell d’ulls compostos, un parell d’antènules i un parell d’antenes, aquestes birràmies i multiarticulades (l’exopodi de les antenes es pot transformar en una esquama o placa antenal dita escafocerita, absent en els fil·locàrides, pancàrides i cumacis); al voltant de la boca hi ha les mandíbules, les maxí·lules i les maxil·les. Al perèion hi ha vuit parells d’apèndixs birramis, un per segment; quan els primers segments són units al cèfalon i formen el cefalotòrax, els apèndixs corresponents es transformen segons la seva funció en maxil·lípedes o gnatopodis, o bé resten com a apèndixs locomotors. El plèon pot portar apèndixs o no, però quan els porta, són de funció respiratòria. El darrer parell d’apèndixs, els uropodis, juntament amb el tèlson formen el ventall de la cua. Els sexes són separats, bé que hi ha alguns casos d’hermafroditisme, per exemple entre els isòpodes i els tanaidacis. Els gonopors se situen sempre al sisè metàmer toràcic en les femelles i al vuitè en els mascles.
Hom considera la subclasse dels malacostracis dividida en sis superordres: els fil·locàrides (Phyllocarida), hoplocàrides (Hoplocarida), els sincàrides (Syncarida), els pancàrides (Pancarida), els peracàrides (Peracarida) i els eucàrides (Eucarida). Al grup dels eucàrides pertanyen els ordres del decàpodes (Decapoda), a més dels eufausiacis (Euphausiacea), i els amfionidacis (Amphionidacea, aquests exòtics i considerats, fins a ben poc, dins dels decàpodes). Els eucàrides són considerats els crustacis de més èxit evolutiu a causa de la gran diversificació assolida pels decàpodes. Com a resultat de la seva gran capacitat d’adaptació a tota mena d’ambients aquàtics i, excepcionalment, a la vida terrestre, són de morfologia molt variada. Entre les característiques principals del grup destaca el gran desenvolupament de la closca, que es projecta en sentit posterior fins a cobrir totalment el perèion, amb el qual se solda dorsalment; es tracta, doncs, del cefalotòrax més desenvolupat de tots els crustacis. A la part anterior de la closca se situen els ulls compostos, que són molt desenvolupats, pedunculats i mòbils, i també les antenes, amb el protopodi format per dos artells. Entre els apèndixs bucals destaca la manca de lacinia mòbil de les mandíbules. Els eucàrides no tenen cambres incubadores o marsupis per a transportar els ous, sinó que els porten entre els pleopodis. El desenvolupament larvari, d’altra banda, es considera el més complex de tots els crustacis. Dels ous neixen larves postnauplianes que, en etapes successives, poden passar per 11 estadis larvaris diferents abans d’arribar a adults, en els casos més complexos.
Tendències actuals
El principal inconvenient de la classificació de Calman és la manca de criteris sobre els quals establir les relacions filogenètiques dins del grup. Altra vegada sorgeix, doncs, el problema de la gran variabilitat dels crustacis i la manca d’unitat en els criteris sobre els quals es podrien basar les relacions entre els diferents grups. A tall d’exemple, i com una revisió històrica, descriurem a continuació alguns dels grups que hom ha proposat per a relacionar les diferents subclasses considerades dins dels crustacis.
D’una banda, els toracòpodes (Toracopoda) reunirien els cefalocàrides, els branquiòpodes i els malacostracis segons la base que tots tenen com a característica comuna la presència de pereiopodis fil·lopodials. Aquests apèndixs es considerarien, doncs, segons aquest criteri, com els apèndixs més primitius i, per tant, relacionats estretament amb els que suposadament tenien els crustacis ancestrals.
El nom de gnatostracis (Gnatostraca) s’aplica a un grup semblant a l’anterior, que reuneix els cefalocàrides i els branquiòpodes, però el qual, a diferència d’aquell, exclou els malacostracis, segons el criteri de considerar el grau de reducció de les maxíl·lules i les maxil·les.
Els pal·liats (Palliata) correspondrien a una suposada línia evolutiva de la closca per a donar lloc als diferents tipus que hi ha, actualment, entre els crustacis. Segons aquest criteri en aquest grup hom reuneix els branquiòpodes, els ostràcodes, els cirrípedes i els malacostracis.
El terme de copèpodes (Copepoda) emprat en sentit ample s’aplicaria per a reunir els mistacocàrides i els branquiòpodes amb els copèpodes, grups que mostren alguns caràcters molt semblants, fins al punt que aquells dos, quan varen ésser descrits per primera vegada es van considerar inclosos dins del grup dels copèpodes. Aquesta inclusió de tres subclasses en un sol grup no alteraria, però, el seu "status" actual.
La denominació de branquiòpodes (Branchiopoda) emprada en sentit ample per a reunir branquiòpodes, ostràcodes i leptostracis és molt inversemblant, ja que és basa exclusivament en el fet que totes tres subclasses tenen closca bivalva.
Els maxil·lòpodes (Maxillopoda) constitueixen una nova agrupació que és, de totes les esmentades fins ara, la més acceptada actualment. Ja hem dit que els copèpodes, els mistacocàrides i els branquiürs presenten una sèrie de característiques comunes que han justificat la creació d’un grup nou. En realitat, els cirrípedes també s’hi haurien d’encloure seguint criteris semblants, fonamentats en l’estructura del cos i dels apèndixs cefàlics.
La classificació dels crustacis és força complexa a causa de la gran quantitat de formes d’organització biològica que presenten les altrament molt nombroses espècies existents. La llista recull i jerarquitza els diversos grups, fins a la categoria taxonòmica d’infraordre, segons un dels molts criteris que han estat utilitzats pels diferents autors, concretament l’observat en aquesta obra. Les abreviatures corresponen a les categories de classe (CI), subclasse (SCI), superordre (SpO) ordre (O), subordre (SO), infraordre (IO) i superfamília (SpF).
| Cl. crustacis (Crustacea) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Scl. branquiòpodes (Branchiopoda) | ||||||||
| O.anostracis (Anostraca) | ||||||||
| O.anostracis (Anostraca) | ||||||||
| O.notostracis (Notostraca) | ||||||||
| O.cladòcers (Cladocera) | ||||||||
| Scl. ostràcodes (Ostracoda) | ||||||||
| O.miodocopis (Miodocopa) | ||||||||
| O.cladocopis (Cladocopa) | ||||||||
| O.platicopis (Platícopa) (* ) | ||||||||
| O.podocopis (Podocopa) | ||||||||
| Scl. mistacocàrides (Mistacocarída) | ||||||||
| Scl. copèpodes (Copepoda) | ||||||||
| O.calanoides (Calanoida) | ||||||||
| O.harpacticoides (Harpactícoida) | ||||||||
| O.ciclopoides (Cyclopoida) | ||||||||
| O.pecilostomatoides (Poecylostomatoida) | ||||||||
| O.sífonostomatoides (Siphonostomatoida) | ||||||||
| O.monstril·loides (Monstrilloida) | ||||||||
| O.mormonil·loides (Mormonilloida) | ||||||||
| O.misofrioides (Mysofrioida) (* ) | ||||||||
| Scl. branquiürs (Branchiura) | ||||||||
| Scl. cirrípedes (Cirrípedia) | ||||||||
|
O.ascotoràcics (Ascothoracica)
|
||||||||
| O.acrotoràcics (Acrothoracica) |
SO.pigòfors (Pigophora)
|
|||||||
| SO.apigòfors (Apigophora) | ||||||||
| O.toràcics (Thoracica) | SO.lepadomorfs (Lepadomorpha) | |||||||
| SO.verrucomorfs (Verrucomorpha) | ||||||||
| SO.balanomorfs (Balanomorpha) | ||||||||
| SO.braquilepadomorfs (Brachylepadomorpha) (+ ) | ||||||||
| O.rizocèfals (Rhizocephaia) | SO.centrogònides (Kentrogonida) | |||||||
| SO.acentrogònides (Akentrogonida) | ||||||||
| Scl. malacostracis (Malacostraca) | ||||||||
| SpO.fil·locàrides (Phyllocarida) | ||||||||
| O.leptostracis (Leptostraca) | ||||||||
| O.himenostracis (Himenostraca) (+ ) | ||||||||
| O.arqueostracis (Archaeostraca) (+ ) | ||||||||
| SpO.hoplocàrides (Hoplocarida) | ||||||||
| O.estomatòpodes (Stomatopoda) | ||||||||
| SpO.sincàrides (Syncarida) | ||||||||
| O.anaspidacis (Anaspidacea) (* ) | ||||||||
| O.estigocaridacis (Stigocaridacea) (* ) | ||||||||
| O.batinel·lacis (Batinellacea) | ||||||||
| SpO.pancàrides (Pancarida) | ||||||||
| O.termosbenacis (Thermosbaenacea) | ||||||||
| SpO.peracàrides (Peracarida) | ||||||||
| O.espeleogrifacis (Spelaeogryphacea) (* ) | ||||||||
| O.misidacis (Mysidacea) | SO.lofogàstrides (Lophogastrida) | |||||||
| SO.misides (Mysida) | ||||||||
| O.cumacis (Cumacea) | ||||||||
| O.tanaidacis (Tanaidacea) | SO.monoconòfors (Monokonophora) | |||||||
| SO.diconòfors (Dikonophora) | ||||||||
| O.isòpodes | SO.gnatidèids (Gnathiidea) | |||||||
| SO.anturoidèids (Anthuroidea) | ||||||||
| SO.microcerberidèids (Microcerberidea) | ||||||||
| SO.flabel·lífers (Flabellifera) | ||||||||
| SO.oniscoidèids (Oniscoidea) | ||||||||
| SO.valvífers (Valvifera) | ||||||||
| SO.freatoicidèids (Phreatoicidea) (* ) | ||||||||
| SO.asel·lotes (Asellota) | ||||||||
| SO.epicaridèids (Epicarídea) |
SpF. bopirins (Bopyrinae)
SpF. criptoniscins (Cryptoniscinae)
|
|||||||
| O.amfípodes (Amphípoda) | SO.gammaridèids (Gammaridea) |
SpF. lisianassoideus (Lysianassoidea)
SpF. gammaroideus (Gammaroidea)
SpF. leucotoideus (Leucothoidea)
SpF. talitroideus (Talitroidea)
|
||||||
| SO.hiperidèids (Hyperiidea) | IO.fisostomats (Physostomata) |
SpF. escinoideus (Scinoidea)
SpF. lanceoloideus (Lanceoloidea)
|
||||||
| IO.fisocefalats (Physocephalata) |
SpF. vibilioideus (Vibilioidea)
SpF. fronimoideus (Phronimoidea)
SpF. liceopsoideus (Lycaeopsoidea)
SpF. platisceloideus (Platysceloidea)
|
|||||||
| SO.caprellidèids (Caprellidea) | ||||||||
| SO.ingolfiel·lidèids (Ingolfiellidea) (* ) | ||||||||
| SpO.eucàrides (Eucahda) | ||||||||
| O.eufausiacis (Euphausiacea) | ||||||||
| O.amfionidacis (Amphionidacea) | ||||||||
| O.decàpodes (Decapoda) | SO.dendrobranquiats (Dendrobranchyata) |
SpF. peneoideus (Penaeoidea)
SpF. sergestideus (Sergestidae)
|
||||||
| SO.pleociemats (Pleocyemata) | IO.estenopodideus (Stenopodidea) | |||||||
| IO.carideus (Caridea) | ||||||||
| IO.astacideus (Astacidea) | ||||||||
| IO.talassinideus (Thalassinidea) | ||||||||
| IO.palinurs (Palinura) | ||||||||
| IO.anomurs (Anomura ) | ||||||||
| IO.braquiürs (Brachiura) | ||||||||
Els grups de crustacis presents als Països Catalans
Els crustacis presents als Països Catalans pertanyen als grups que citem a continuació i que comentarem en els capítols posteriors. Dins dels crustacis no malacostracis tenim els branquiòpodes, majoritàriament d’aigües continentals, de basses i llacunes, entre els quals hi ha les curioses tortuguetes i les puces d’aigua; els ostràcodes, petits crustacis bivalves que viuen tant a les aigües dolces com a la mar; els mistacocàrides, petits organismes que viuen a l’aigua que mulla la sorra de la platja, a la vora de les onades; els copèpodes, que constitueixen una gran part de la biomassa del zooplàncton marí; els branquiürs, paràsits, poc representats a les nostres latituds, i els cirrípedes marins, de vida paràsita o sèssils, com els bàlans i els peus de cabrit. Dins dels crustacis malacostracis, trobem els leptostracis (fil·locàrides), un grup de crustacis marins poc important numèricament però molt interessant des del punt de vista zoològic, ja que es consideren els malacostracis més primitius; els estomatòpodes (del grup dels hoplocàrides), organismes marins entre els quals és ben coneguda la galera; els batinel·lacis, grup de sincàrides que són importants com a fòssils i dels quals, en canvi, coneixem actualment pocs representants, d’un gran interès biogeogràfic; els termosbènids (pancàrides), organismes molt poc abundants, que viuen a les aigües termals i a les coves marines; els misidacis (peracàrides), crustacis d’una gran importància quantitativa als ambients marins; els cumacis (peracàrides), marins i principalment bentònics; els tanaidacis (peracàrides), igualment habitants dels fons marins, poc representats a les nostres mars; els isòpodes (peracàrides), grup molt extens, i d’una gran diversificació, amb representants marins, d’aigua dolça i també terrestres; els amfípodes (peracàrides), principalment marins, molt importants numèricament. Dins dels malacostracis eucàrides, coneixem els abundants eufausiacis, que neden fent grans moles, entre els quals hi ha els que constitueixen l’aliment dels cetacis i de molts peixos ("krill"), i els decàpodes, els més coneguts de tots els crustacis: crancs, gambes, llagostes marines, etc.