
El gran nombre de teranyines que trobem al bosc a determinades èpoques, especialment a final d’estiu, ens fa adonar de la importància numèrica d’un grup d’organismes que la resta de l’any semblen absents i els quals, a causa de llurs hàbitats i costums, passen per ésser considerats artròpodes poc abundants, en contra de la presència ben ostensible i gairebé constant dels insectes al medi aeri i dels crustacis al medi aquàtic.
Xavier Moreno.
D’entre el conjunt de formes artropodianes, actuals i fòssils, es pot destacar un ampli grup d’espècies, els quelicerats, que mostren una sèrie de trets estructurals comuns, el més significatiu dels quals és, sens dubte, la possessió d’un parell d’apèndixs, anatòmicament preorals, en forma de pinça o ganxo: els quelícers; altres trets acompanyants són l’absència d’antenes, la divisió del cos en tagmes o regions, la posició, constant, de l’orifici genital, etcètera.
Aquestes i altres adaptacions revelen un caràcter eminentment depredador, que els aràcnids comparteixen amb els merostomes; la disjunció entre ambdós grups es fonamenta en criteris adaptatius, funcionals, que deriven del seu hàbitat —terrestre o marí, respectivament— i de l’escassetat de dades paleontològiques, que emmascara els orígens i les connexions evolutives entre ambdós blocs.
Seguint criteris tradicionals, més assentats en l’ús que no pas en les tendències fílètiques actuals, hom divideix el bloc dels artròpodes quelicerats en dues fraccions, amb categoria de classe: els merostomats i els aràcnids. En l’actualitat, la morfologia, l’embriologia i la paleontologia disposen de raonaments que poden posar en dubte aquesta divisió simplista. Tanmateix, no havent-hi tampoc evidències absolutes entorn d’altres esquemes alternatius, ens sembla suficient assenyalar, ací, aquestes vacil·lacions i, alhora, adoptar aquesta divisió senzilla, que s’adapta millor a la funció d’aquesta obra.
Característiques del grup

Els grans trets morfològics que caracteritzen els aràcnids poden veure’s sobre un model, en aquest cas, una aranya del gènere Araneus: són visibles a la fotografia la divisió del cos en dues parts, la concentració dels apèndixs a la part anterior, la presència de quelícers i pedipalps, i el nombre de segments de les potes, generals a tots els aràcnids, i d’altres caràcters, com l’absència de segments visibles a tot el cos i la formació de fils de seda, que són propis d’alguns dels grups.
Rafael Campillo.
Les aranyes, els escorpins, els opilions, els àcars i altres grups menors solen agrupar-se en els esquemes sistemàtics en una sola unitat taxonòmica, amb categoria de classe, la classe dels aràcnids. Tots tenen un conjunt de trets o caràcters comuns, que destacarem a continuació com una descripció definitòria del grup. Són quelicerats adaptats a la vida terrestre; tenen el cos constituït per dos tagmes, un d’anterior, anomenat prosoma, i un altre de posterior anomenat opistosoma, per bé que no sempre sigui nítida la separació anatòmica d’ambdues parts. El prosoma porta sis parells d’apèndixs, dels quals, els quatre darrers estan adaptats a la locomoció (potes caminadores); el primer parell d’apèndixs prosòmics són els quelícers, que van inserits per davant de la boca, mentre que el segon parell, generalment anomenat pedipalps, és de morfologia molt variada. L’opistosoma porta l’orifici anal en posició terminal o subterminal i l’orifici genital, i encara, de vegades, vestigis apendiculars molt modificats; l’orifici genital conserva una posició morfològica constant: sempre a la cara ventral del segon segment opistosòmic. La respiració es fa per mitjà de tràquees o filotràquees, i només excepcionalment és cutània, en algunes formes de petites dimensions; l’excreció es fa per mitjà de tubs de Malpighi i de glàndules coxals, principalment l’intestí mitjà (mesodeu) diferencia una glàndula hepatopancreàtica, constituïda per diverticles molt nombrosos i ramificats. Són gonocòrics i de desenvolupament directe; són, majoritàriament, depredadors i el seu sistema d’alimentació consta d’una fase prèvia de digestió externa.
Morfologia
Les regions del cos i la metamerització
Els aràcnids mostren una gran diversitat de formes, però mantenen sempre una mateixa organització general. La divisió del cos en dues regions o tagmes (prosoma i opistosoma) sol trobar-se emmascarada per modificacions secundàries, que tendeixen, d’una banda, a la pèrdua de la identitat dels metàmers que els constitueixen, de l’altra, a la fusió d’ambdues regions, amb pèrdua de llur demarcació anatòmica, i, ocasionalment, al desdoblament de l’opistosoma en un mesosoma, ample, i un metasoma, estret.
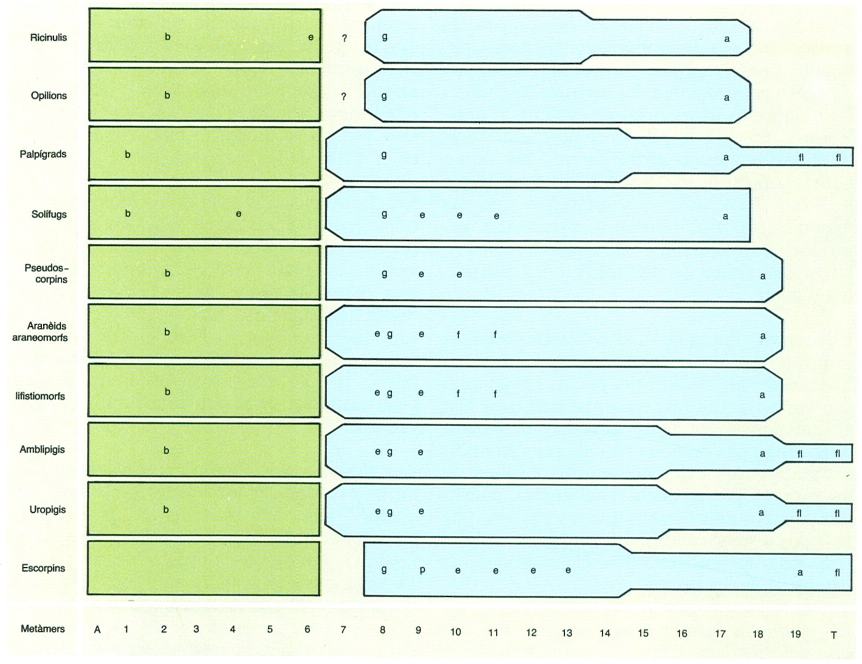
Nombre de segments del cos en els diferents grups d’aràcnids i la seva distribució en tagmes o regions, amb indicació de la posició de la boca (b), l’anus (a), els estigmes respiratoris (e), els gonopors (g) i altres estructures més específiques dels diferents grups, com les fileres (f), el flagel (fl), el fibló (fi) i les pintes (p). La reducció dels segments compresos entre l’àcron (A) i el tèlson (T) s’indica amb la disminució de l’amplada de la taca de color fins a la seva desaparició. Els colors indiquen els segments corresponents al prosoma (verd) i a l’opistosoma (blau).
Maber, a partir d’originals diversos.
La pèrdua d’identitat dels metàmers per la modificació dels seus elements externs i interns és una de les dificultats de la seva interpretació morfològica. Per això, aquesta interpretació s’ha de fer per mitjà d’hipòtesis, que suposen diferenciacions, supressions, desplaçaments, reutilitzacions, desdiferenciacions, etc. Únicament algunes vegades aquestes hipòtesis es converteixen en evidències, en reconstruir-se sèries fílètiques a les quals és possible una homologació perfecta, o bé quan l’estudi de l’ontogènia descobreix posicions o parentius que són críptics en els adults. Cal no perdre de vista que en els aràcnids, com a la resta dels artròpodes, el principi bàsic estructural, és a dir, la metamerització, subordina la seva plasticitat i capacitat adaptativa a un principi funcional: la tagmosi.
L’entitat funcional de cadascun dels dos tagmes diferenciables és clara. Mentre que el prosoma s’organitza estructuralment per servir a funcions sensorials i de coordinació, d’ingestió de l’aliment i de locomoció, a l’opistosoma es localitzen funcions vegetatives: circulació, respiració, digestió interna, reproducció, etc.
Rarament són nítids els límits entre el prosoma i l’opistosoma, tret de les aranyes i els amblipigis, en els quals hi ha un pedicel·le o estrangulament que fa de pont. Externament, en el prosoma s’aprecia una tendència acusada cap a la coalescència de les plaques tergals i esternals, i una reducció de les zones membranoses pleurals. D’aquesta manera, tret d’alguns grups (solífugs, esquizòmids i palpígrads), les tergites prosòmiques formen un sol escut, l’anomenat escut prosòmic. El prosoma, així, es converteix en una estructura rígida i compacta. Per contra, l’opistosoma té una clara tendència a reduir les esclerites que, en molts casos, es conserven com a testimonis de la metameria en detriment d’un tegument membranós i plegat, que permet les enormes distensions que imposen els fenòmens reproductius o alimentaris.
La participació dels apèndixs en les tasques de captura de l’aliment i en la seva elaboració i ingestió, afegeix un factor clar de modificació de l’estructura metamèrica primària: d’una banda, el desplaçament dels quelícers, apèndixs primordials en aquesta funció, per davant de l’orifici bucal; de l’altra, l’apropament de les coxes dels pedipalps, i de vegades, també, dels dos primers parells de potes caminadores cap a la boca, i la seva diferenciació com a estructures gnatocoxals. Aquest apropament comporta simultàniament un allargament de les estructures coxals, les quals convergeixen cap a la boca, de manera que la seva zona esternal es redueix i empeny cap endavant elements esternals posteriors. Així, podem trobar, a la cara ventral, una placa esternal única, dues plaques o la seva absència total, o també la presència d’elements esternals opistosòmics en una posició anatòmicament prosòmica.
És pràcticament impossible deduir amb seguretat, a través dels elements externs, quants i quins són els metàmers que integren el tagma anterior del cos. Els apèndixs són testimonis innegables dels seus metàmers respectius, però hi ha certs dubtes i una manca d’arguments per a considerar si la regió preoral és constituïda o no a partir de diversos metàmers. Per tant, hi ha distintes hipòtesis, que es fonamenten, sobretot, en l’anatomia comparada del sistema nerviós, en l’embriologia i en la interpretació de les diferents estructures apendiculars.
La hipòtesi més generalitzada, tot i que probablement és massa simplista, és la que emeteren Holmgren i Hamstron l’any 1928, i que ha estat suportada per autors clàssics com Millot (1949), Snodgrass (1952) i Kaestner (1956). Es fonamenta en la interpretació del sistema nerviós, i sosté que el prosoma dels aràcnids és constituït per sis metàmers, precedits per una zona acronal i distribuïts de la manera següent: l’àcron constitueix l’extrem anterior del prosoma, porta els lòbuls protocerebrals (el deutocervell és pràcticament inexistent) i es construeix a expenses de fragments mesodèrmics despresos del primer metàmer; el segment quelicèric, que porta els quelícers íntimament units a la zona acronal, que seria el primer segment del cos i els segments neuromèrics del qual, prèviament postorals, es desplacen cap endavant i deixen una comissura infraesofàgica; així constitueixen els lòbuls tritocerebrals. El segon segment porta els pedipalps, els quals s’innerven a la massa nerviosa infraesofàgica; el tercer, el quart, el cinquè i el sisè segments porten, respectivament, el primer, el segon, el tercer i el quart parells de potes caminadores i s’innerven, igualment, a la massa infraesofàgica, que representa la coalescència d’un nombre variable de parelles de neuròmers.
A l’opistosoma, la deducció metamèrica no és tan complicada perquè disposem d’arguments anatòmics i embriològics més bons. Les dificultats més grans es troben en la reducció i la simplificació dels darrers metàmers del tagma i, també, del primer, que l’uneix al prosoma. Embriològicament, hom arriba a diferenciar 12 o 13 somites, encara que en alguns casos no tants. A vegades, el tèlson morfològic es conserva en forma de fibló (en els escorpins) o de filament multiarticulat en d’altres (palpígrads). L’anus ocupa una posició terminal, lleugerament desplaçada envers la cara ventral, després del darrer somita. L’orifici genital s’obre en el segon segment opistosòmic, en posició ventral.
El segment anomenat pregenital (el primer de l’opistosoma) generalment experimenta una reducció pel fet de fer de punt d’unió amb el prosoma, bé estrenyent-se en forma de pedicel·le (en aranyes i amblipigis) o bé perdent la placa tergal, mentre que la seva part esternal es veu empesa cap al prosoma (en escorpins i opilions).
En els segments postgenitals sol haver-hi un desenvolupament preponderant dels somites intermedis, mentre que els metàmers terminals es redueixen, es telescopitzen o desapareixen; aquesta tendència és màxima en els àcars.
Els apèndixs i les formacions apendiculars
Els apèndixs són formacions primàries de caràcter metamèric i, per tant, són molt valuosos en els jocs d’interpretació morfològica.
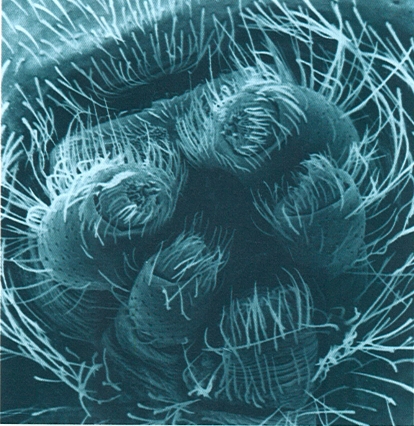
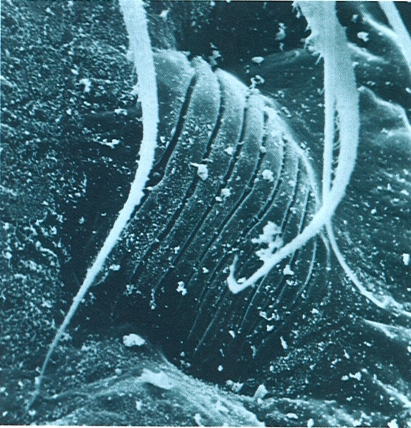
Detall de les fileres d’una aranya (Amaurobius), vistes al microscopi electrònic d’escandallatge (× 100) Noteu l’estructura cònica d’aquests apèndixs modificats i la presència al seu interior de les nombroses fúsules per les quals emergeix la seda.
Carles Ribera.
El prosoma mostra, invariablement a tots els aràcnids, sis parells d’apèndixs: quelícers, pedipalps i parells primer, segon, tercer i quart de potes caminadores. El caràcter apendicular d’aquestes estructures és evident. L’opistosoma, en canvi, pot presentar una sèrie d’estructures que, aparentment, no tenen aquest aspecte i, malgrat tot, cal considerarles com a apèndixs morfològics o derivats apendiculars. Ens referim a estructures respiratòries anomenades fil·lotràquees, a les fileres, les pintes (o rastrells) i les plaques operculars.
Es reserva el nom de quelícers exclusivament al primer parell d’apèndixs del prosoma, situats al davant de l’orifici bucal. En general són curts i petits (excepcionalment, són ben desenvolupats en alguns grups, com els solífugs, en alguns opilions i en algunes aranyes) i orientats cap a la cara ventral. Acaben en un ganxo o pinça (queia o subquela), constituït pels dos únics artells que els formen o bé pels dos distals, quan són formats per tres artells. La seva funció bàsica és la subjecció i la trituració de les preses.

Visió general d’una aranya (Zoropsis spinimanus) que mostra les característiques dels dos primers parells d’apèndixs dels aràcnids, els quelícers, curts, robusts i prensors, i els pedipalps, d’aspecte més semblant a les potes, de vegades acabats en pinça, però generalment de funció sensorial.
Ramon Torres.
Els pedipalps formen el segon parell d’apèndixs del prosoma. Les seves coxes se situen a banda i banda de la boca i generalment tenen diferenciades unes làmines maxil·lars anomenades gnatocoxes. Són, sens dubte, els apèndixs més heterogenis, en consonància amb l’heterogeneïtat de funcions que arriben a desenvolupar. En els escorpins i els pseudoscorpins atenyen un desenvolupament considerable, sovint amb la diferenciació d’una queia terminal, però generalment són més petits que les potes caminadores i assumeixen funcions sensorials. Generalment consten de cinc artells. L’artell terminal o tars mostra diferenciacions notables, com els òrgans en ventosa (en els solífugs), el bulb copulador (en els mascles de les aranyes), etc.
Els quatre parells d’apèndixs restants són més homogenis i assumeixen funcions locomotores. Excepcionalment, es pot produir una inversió funcional entre els pedipalps i el primer parell de potes, amb els canvis estructurals consegüents. Les potes locomotores són apèndixs llargs, d’artells cilíndrics, en nombre de set, anomenats coxa, trocànter, fèmur, patel·la, tíbia, metatars i tars. Les ungles terminals es poden interpretar com un pretars.
Les fil·lotràquees, també anomenades pulmó en llibre, tenen valor morfològic d’apèndixs opistosòmics. Són presents en els escorpins (segments 4t, 5è, 6è i 7è), aranyes (segments 2n i 3r), amblipigis, uropigis i esquizòmids. La seva estructura es tractarà més endavant, en parlar dels elements respiratoris.
En els escorpins, el segon segment opistosòmic destina els esbossos metamèrics apendiculars a la diferenciació de les valves que formen l’opercle genital, mentre que els esbossos apendiculars del tercer diferencien unes estructures molt peculiars, de funció sensorial, que pel seu aspecte extern reben el nom de pintes. També són derivats apendiculars les estructures còniques i generalment biarticulades anomenades fileres, a través de les quals les aranyes emeten la seda. Totes aquestes estructures es tractaran en detall en l’estudi dels diferents grups.
Organització interna
El tegument i la musculatura
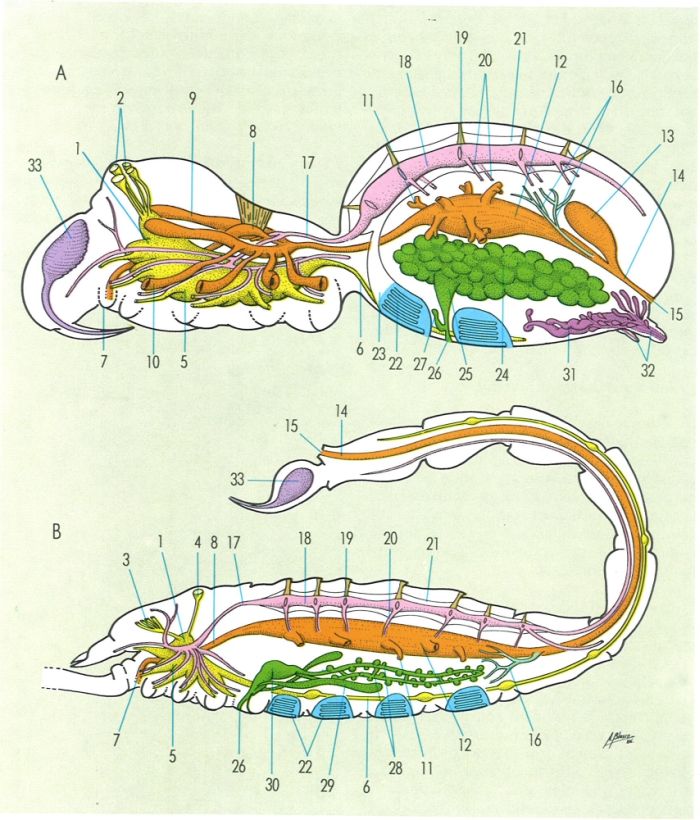
Anatomia dels aràcnids, vista sobre dos models: una aranya (A) i un escorpí (B), i indicació dels principals elements interns: 1 cervell, 2 ulls, 3 ulls laterals, 4 ulls dorsals, 5 massa ganglionar infraesofàgica, 6 cadena nerviosa ventral, 7 boca, 8 faringe. 9 cecs laterals, 10 cecs apendiculars, 11 cecs hepatopancreàtics, 12 intestí, 13 ampolla rectal. 14 recte, 15 anus, 16 tubs de Malpighi, 17 aorta anterior, 18 cor o vas dorsal, 19 ostíol. 20 arteriola metamètica, 21 pericardi. 22 fil·lotràquees, 23 si pulmonar, 24 ovaris, 25 oviducte, 26 orifici genital, 27 espermateca, 28 testicles, 29 glàndules accessòries, 30 atri genital. 31 glàndules sericígènes. 32 fileres. 33 glàndula de verí.
Amadeu Blasco, original de l’autor.
El tegument dels aràcnids, com el de la resta dels artròpodes, comprèn un epiteli simple —la hipodermis— que descansa damunt d’una membrana basal, i que és recobert per una cutícula quitinosa, de gruix variable, segregada per la capa ectodèrmica.
La cutícula serveix de punt d’inserció a la musculatura, i a diferents punts del cos es diferencien estructures apodemals, les principals de les quals són les següents: els apodemes coxals, situats a la cara anterior de les coxes dels apèndixs; els apodemes del clipi, al marge anterior de l’escut prosòmic; i els apodemes tergals, que generalment consisteixen en un engruiximent central, sagital (fòvea), o bé en engruiximents laterals. Aquestes estructures apodemals, juntament amb l’anomenada endosternita constitueixen l’esquelet intern dels aràcnids. L’endosternita és una formació secundària, d’origen conjuntiu (mesodèrmic), que es diferencia a la zona ventral de confluència de la musculatura prosòmica, per sota del tub digestiu. És present únicament en alguns grups.
La musculatura té un caràcter bàsicament metamèric, però l’efecte de la tagmosi, particularment en el prosoma, comporta modificacions importants (desplaçaments, hipertròfies, reduccions, etc.) que emmascaren el seu caràcter primari. Això s’acompleix en el cas dels músculs anomenats suspensors de l’endosternita, els verticals, els horitzontals, que, en definitiva, corresponen a la musculatura dorsiventral, i els longitudinals, de caràcter metamèric. Altres músculs importants a la zona del prosoma són els músculs extrínsecs dels diferents apèndixs i també els intrínsecs, i els músculs de la faringe, molt desenvolupats, ja que fan possible el bombeig de líquids o purés alimentaris.
A l’opistosoma es manté amb més nitidesa l’ordenació muscular metamèrica (músculs dorsiventrals i músculs longitudinals), tot i que també aquests són afectats per la reducció de la zona terminal.
Els sistemes de relació i control
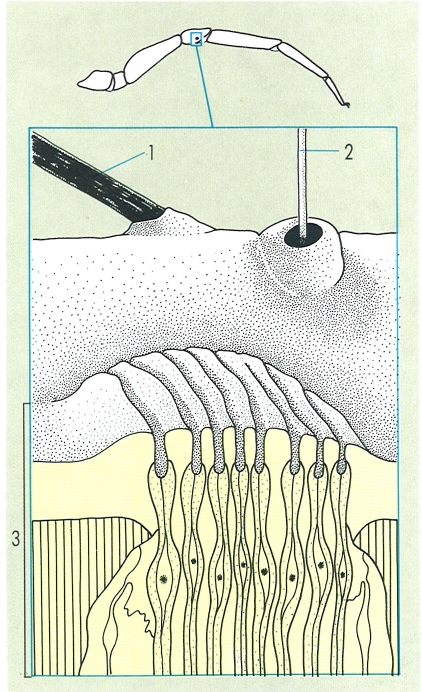
Diferents formacions tegumentàries pròpies dels aràcnids i la seva localització en un apèndix qualsevol del cos: 1 pèl o seda sensorial, 2 tricobòtria, 3 òrgan liriforme.
Gustavo Hormiga.
El sistema nerviós central consta, com a la resta dels artròpodes, d’una massa ganglionar encefàlica unida a la cadena ventral per un collar que envolta l’esòfag. Com a norma general, en els aràcnids hi ha un escurçament de la cadena ventral que comporta una gran concentració de neuròmers, els quals es fusionen i fan una massa nerviosa infraesofàgica voluminosa i complexa.
La constitució del cervell és semblant en els diferents grups, però hi ha certes divergències pel que fa a la seva interpretació. El protocervell és format per dos lòbuls ben desenvolupats, als quals es distingeixen, almenys, els centres visuals, el cos central i els cossos pedunculats. Bàsicament són centres d’associació de la visió. El deutocervell és molt reduït, pràcticament englobat pel protocervell i el tritocervell. El tritocervell, també anomenat centre quelicèric pel fet que innerva fonamentalment els quelícers, és molt desenvolupat; la seva comissura transversal és infraesofàgica, cosa que és un clar indici de la seva condició morfològica postoral.
El sistema estomatogàstric constitueix la porció nerviosa que innerva la part estomodeal del tub digestiu. Té un funcionament autònom i regula d’una manera reflexa la túnica muscular que l’envolta. És en relació amb el sistema central a través del gangli innominat (centre de caràcter vestigial), que connecta amb el gangli frontal a través del nervi recurrent. El gangli frontal presenta connectius cap al deutocervell i el tritocervell.
Hi ha òrgans endocrins situats a la part anterior del prosoma, per sobre de la massa encefàlica (els òrgans de Schneider 1 i 2), i a l’opistosoma. Els òrgans de Schneider 1 són els més desenvolupats i mantenen connexió amb les diferents parts del cervell i la massa infraesofàgica; alhora es relacionen amb els òrgans de Schneider 2, que també són connectats al deutocervell o centre rostral. La informació que hom posseeix sobre aquests sistemes simpàtic i neuroendocrí és molt limitada.
Els aràcnids mostren tres tipus d’òrgans sensorials generalitzats: els ulls, els òrgans liriformes i les setes o pèls sensorials. A més, hi ha, a determinats grups, altres òrgans sensorials particulars i característics, com les pintes dels escorpins, els òrgans en raqueta i els òrgans en campana dels solífugs, l’òrgan tarsal de les aranyes, les vesícules de Haller i els òrgans estigmàtics dels àcars, entre d’altres.
La presència de setes en el cos dels aràcnids és, en general, abundant, i moltes vegades tenen funció sensorial mecanoreceptora i quimioreceptora. Mereixen una menció especial les tricobòtries, que són uns pèls fins i molt llargs, l’aurèola membranosa basal dels quals els caracteritza.
Òrgan liriforme d’aràcnid (Dysdera) vist al microscopi electrònic d’escandallatge (× 800). Dins de cadascuna de les petites fissures de què es compòn aquesta petita fosseta olfactòría, hi ha una cèl·lula nerviosa capaç de percebre els estímuls químics procedents de l’exterior.
Carles Ribera.
Els òrgans liriformes es troben preferentment als apèndixs, però també al cos. Es tracta de fenedures aïllades o agrupades en disposició paral·lela, que porten una petita dilatació mitjana on s’allotja la terminació d’una cèl·lula sensorial protegida per una cutícula fina. La distribució d’aquestes estructures és constant en els diferents grups. Únicament els palpígrads i els ricinulis no en tenen.
Els aràcnids no gaudeixen d’un equipament fotoreceptor gaire diferenciat, però tenen algunes particularitats en aquest sentit. Tret d’algunes rares excepcions (adaptacions a la vida cavernícola, per exemple), presenten un o dos tipus d’ulls: laterals i mitjans. Els ulls laterals són estructuralment molt simples, ja que es diferencien gràcies a un engruiximent ectodèrmic lleugerament enfonsat, al qual es distingeix una capa retinal, protegida per una lent o còrnia. Els ulls mitjans sorgeixen d’una invaginació ectodèrmica, prèviament engruixida, que es recobreix a ella mateixa, de manera que s’hi diferencien tres capes cel·lulars, i de l’extrem de la invaginació en sorgeix un gangli òptic en relació amb el protocervell. La capa externa origina la còrnia; la capa intermèdia, que adopta una posició invertida, origina la retina; la capa interna, post-retinal, és en general una capa pigmentària, reflectant, que condiciona una visió indirecta. Els ulls són, sempre, de tipus rabdomèric, i es localitzen a la part anterior del prosoma.
L’aparell digestiu
La majoria dels aràcnids són carnívors i depredadors; subjecten i trituren les seves preses amb els quelícers i els pedipalps. Contra allò que podria suposar-se, ingereixen únicament aliment líquid, per la qual cosa procedeixen abans a triturar les preses i a fer-ne una digestió externa, gràcies als líquids digestius que regurgiten al damunt dels materials triturats. Les glàndules salivals són molt desenvolupades.
Aquesta pasta predigerida passa a la cavitat prebucal, delimitada pels quelícers, els elements coxals i esternals dels pedipalps i els primers parells de potes. En aquesta cambra sovint hi ha raspalls de pèls que efectuen processos de filtració. L’estomodeu pròpiament dit comprèn una faringe aspiradora (musculatura de bombeig) i un conducte esofàgic que arriba fins a la zona endodèrmica.
El mesodeu pot presentar una sèrie de diverticles, amplament ramificats, que penetren els apèndixs, i hi fan com una mena de reservori. La seva porció més desenvolupada i constant és l’anomenada glàndula hepatopancreàtica, una estructura esponjosa que omple la cavitat general de l’opistosoma. És constituïda per una multitud de divertides molt ramificats, de parets glandulars que segreguen sucs digestius i alhora fan funcions d’absorció.
L’intestí posterior o proctodeu és curt i s’obre a l’exterior a través de l’anus, en posició terminal o subterminal, amb una ampolla rectal que controla la defecació.
L’aparell respiratori
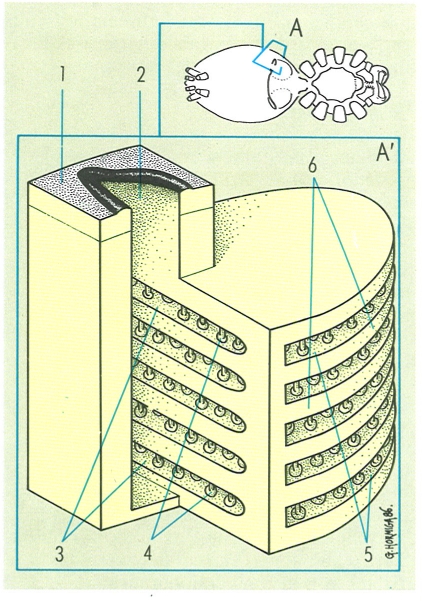
Les fil·lotràquees o pulmons en llibre són estructures típiques dels aràcnids. Morfològicament considerades com a derivats dels apèndixs de l’opistosoma. es presenten per parelles a la cara ventral del cos (A). En l’esquema d’una fil·lotràquea (A’) hom distingueix: 1 superfície cuticular externa, 2 estigma fil·lotraqueal, 3 cambra aèria. 4 columnetes. 5 cambra hemàtica. 6 laminetes cuticulars.
Gustavo Hormiga, a partir d’originals diversos.
Els aràcnids respiren per mitjà de pulmons laminare o fil·lotràquees, que poden anar acompanyats o ésser substituïts per un sistema traqueal. Els escorpins, els amblipigis, els esquizòmids, els uropigis, entre altres, porten exclusivament fil·lotràquees, mentre que els àcars, els opilions, els pseudoscorpins, els solífugs i els ricinulis tenen exclusivament tràquees. Les aranyes, al llarg de la seva gran variació sistemàtica, mostren totes tres possibilitats. Els palpígrads no tenen òrgans respiratoris diferenciats.
Anatòmicament, les fil·lotràquees sorgeixen d’una invaginació del tegument que diferencia un vestíbul o atri. En aquest atri, la paret anterior forma una sèrie d’invaginacions secundàries de parets molt fines, paral·leles i equidistants, superposades com els fulls d’un llibre, que es mantenen separades per mitjà de petites columnetes cel·lulars. La part interna de les laminetes és banyada per l’hemolimfa, en relació amb un si sanguini ventral. La circulació de l’hemolimfa i la ventilació pulmonar es fan principalment per l’alternació rítmica de pressió i depressió que es deriva de la contracció del vas dorsal. El nombre de laminetes és variable.
Les fil·lotràquees es localitzen, per parelles, a la cara ventral de l’opistosoma, i es troben clarament metameritzades. És clar el valor mofològic d’aquestes estructures que representen apèndixs opistosòmics molt diferenciats i involucionats, homologables a les potes branquials que tenen els grups merostomats. L’estructura anatòmica no recorda gens l’estructura apendicular, però la seqüència ontogènica permet de catalogar-los com a apèndixs veritables, involucionats a les primeres etapes i incorporats a les zones laterals de l’esternita corresponent i que diferencien, només, l’estructura respiratòria de les laminetes posteriors.
El sistema traqueal dels aràcnids és comparable, d’una manera general, al dels insectes, però deriva, en part, del sistema fil·lotraqueal, al qual substitueix en un grau variable. Consisteix en un sistema de túbuls profusament ramificats a diferents nivells, que es mantenen obert pels tenidis espiralats o circulars. Estructuralment, és un sistema molt més senzill que el dels insectes, però la seva base funcional és semblant.
L’aparell circulatori
L’aparell circulatori dels aràcnids és obert. Comprèn un cor o vas dorsal, envoltat d’un si pericàrdic, i d’una sèrie de vasos principals, no capil·laritzats, que s’obren a una xarxa de sins hemolimfàtics.
El cor és un tub de parets contràctils, situat en posició sagital, a la meitat anterior dorsal de l’opistosoma. Primàriament, el cor és una estructura segmentada, constituïda per subunitats o petits ventricles, cadascun dels quals porta un parell d’ostíols, i un parell d’arterioles metamèriques. La tendència general, ja esmentada, de reducció de la metameria opistosòmica, també afecta el vas dorsal, fins al punt que arriba a desaparèixer en alguns àcars.
L’hemolimfa que envolta el vas dorsal, inclosa a la cambra pericàrdica, penetra al cor pels ostíols i és impulsada cap endavant per una aorta posterior, i cap als costats per arterioles metamèriques que irriguen les vísceres. L’hemolimfa queda lliure a la cavitat general del cos i s’acumula en un gran si ventral, en relació amb els sins pulmonars. L’acció del cor bombeja l’hemolimfa des dels sins pulmonars fins a la cambra pericàrdica, a través de les artèries pulmonars.
Els òrgans excretors
En els aràcnids, els òrgans excretors, poden ser de dos tipus: glàndules coxals i tubs de Malpighi.
Les glàndules coxals són derivats celomàtics i, per tant, es poden qualificar d’urocels. Es disposen fent parells metamèrics en el prosoma, en nombre variable a cada grup. Cada glàndula coxal consta d’un sac inicial mesodèrmic, unit per un canal curt a un tub cargolat i apilotat (el laberint), la part terminal del qual s’obre a l’exterior després d’una dilatació o veixiga. Els orificis excretors es localitzen a la base de les coxes.
Els tubs de Malpighi són estructures tubulars, cegues, flexuoses, que en nombre d’un parell o dos s’obren a la part terminal del mesodeu. Es localitzen a l’opistosoma, entre els diverticles de la massa hepatopancreàtica. La llum del tub és ciliada. Eliminen els productes residuals nitrogenats en forma de cristalls de guanina.
L’aparell reproductor
En els aràcnids, els sexes són separats i és freqüent el dimorfisme sexual, que en alguns casos es tradueix en desproporcions en les dimensions del cos, en la forma o en les dimensions dels quelícers, i també en la pigmentadó i ornamentació del cos.
Les gònades són parelles i ocupen les zones laterodorsals de l’opistosoma. Cada gònada es perllonga cap endavant per un gonoducte que ateny la zona anteroventral (el segon segment) de l’opistosoma, es fusionen i entren en relació amb un atri, previ a l’orifici genital. En aquest atri, de complexitat molt variable, aboquen tota una sèrie de glàndules accessòries, responsables de la formació dels espermatòfors, quan n’hi ha.
La reproducció

El desenvolupament dels aràcnids és ràpid i no acostuma a comprendre fases amb morfologies diferents, si bé hi ha estadis postembríonaris separats per mudes. Durant el primer i el segon d’aquests estadis, als quals correspon la fotografia, les petites aranyes es mostren gregàries, atès que acaben de sortir dels ous. A partir de la segona o tercera muda. però, es dispersen i inicien la seva vida independent.
Ramon Torres.
Els aràcnids mostren un sistema reproductor típicament biparental, que és conseqüència del seu gonocorisme estricte, i que comprèn una fase de fecundació (amb una sèrie de conductes molt estereotipades i interessants), la posta (i la cura dels petits, generalment perllongada, a càrrec de la femella) i els aspectes ontogènics o de desenvolupament pròpiament dit, que condueixen a la formació de nous individus.
La fecundació és interna, però generalment és indirecta, bé per mitjà d’espermatòfors, bé per estructures transmissores de l’esperma, no relacionades amb les vies genitals (per exemple, el bulb copulador de les aranyes). Aquest sistema no exclou, sinó que reforça, una conducta d’aparellament ritualitzada, de la qual parlarem més endavant.
La majoria dels aràcnids són ovípars, condició que es considera primària, tot i que hi ha alguns casos d’ovoviviparisme i de viviparisme (alguns escorpins i àcars). En general, les femelles disposen d’espermateques, cosa que les capacita per a tenir una o més postes. Els ous, en general, són protegits per una coberta de seda o d’alguna altra substància, i són vigilats per la femella.
L’etologia reproductora dels aràcnids sol ésser bastant complexa. Moltes vegades, el mascle ha de desenvolupar un sistema de senyals, de tipus vibratori (transmissió per l’aire, l’aigua o la tela), sonor (òrgans estridulants diversos) o de tipus visual (moviments estereotipats dels pedipalps i les primeres potes, lliurees pigmentaries característiques) als quals cal que la femella respongui favorablement per tal que l’aparellament s’efectuï amb èxit. La gamma de possibilitats en aquest sentit és immensa: des d’una desproporció de grandàries entre ambdós sexes i el comensalisme del mascle, que viu al damunt de la femella, passant per mecanismes més o menys complexos d’ofrena, fins als atacs fulminants i per sorpresa a les femelles, que provoquen en aquestes reaccions de submissió. Tot i que en alguns casos hi ha una introducció directa de l’esperma a les vies genitals femenines, gràcies a l’existència d’òrgans copuladors (en els opilions), en els aràcnids són molt freqüents els mecanismes de transmissió indirecta de l’esperma, bé per estructures diferenciades a altres parts del cos (el bulb copulador de les aranyes), bé per la diferenciació d’espermatòfors (cas dels pseudoscorpins, per exemple).
Les femelles fan una o més postes, que generalment protegeixen o recobreixen de diverses cobertes de seda o d’altres substàncies, i que rarament abandonen. Al contrari, la femella sol vigilar-les de prop, i fins i tot les transporta damunt seu, o bé resta, inactiva, al seu costat, fins que s’esdevé el seu exhauriment alimentari i la seva mort. En produirse l’eclosió, el desenvolupament de la cria és molt ràpid. Les formes juvenils es mantenen agrupades durant un temps breu (fins a la segona o tercera muda) i, després, es produeix la dispersió, moment que és crític per a la majoria dels individus, ja que únicament un petit nombre assoleix l’estadi adult.
Desenvolupament embrionari i postembrionari
La segmentació i la gastrulació
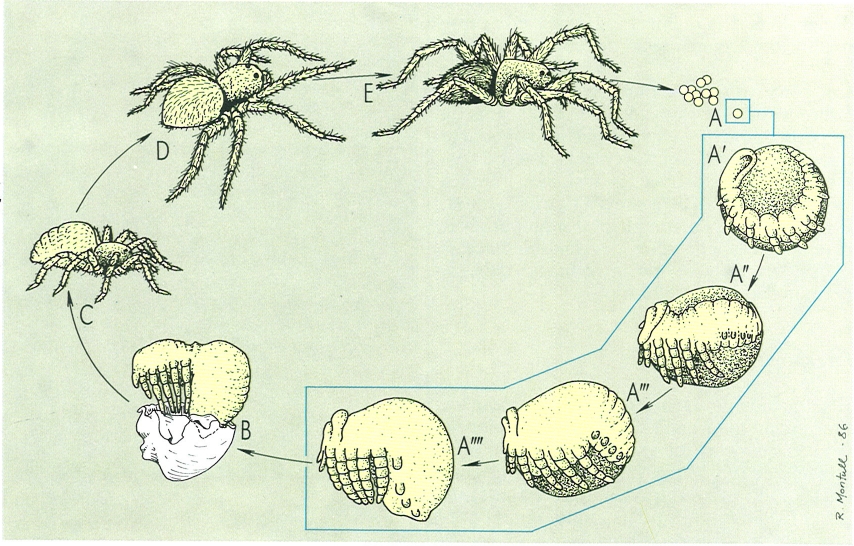
Desenvolupament embrionari i postembrionari d’una aranya. Les aranyes ponen un gran nombre d’ous (A), que recobreixen de fils de seda. Durant el desenvolupament embrionari s’esdevé la inversió de l’embrió (A’, A", A’", A""), després de la qual la massa de vitel·lus queda completament tancada en el mesodeu i es comença a diferenciar l’organització interna del cos i també l’externa (apareix el prosoma, l’opistosoma i els esbossos dels apèndixs). L’eclosió de l’ou dóna lloc a l’emergència d’una fase immòbil, la prelarva (B), a la qual segueix una primera fase juvenil, mòbil (C). Els joves experimenten un creixement a base de mudes que els porta a l’etapa de subadult (D) i finalment a la d’adult (E), sexualment madura.
Servei de Fotografía/Román Montull, a partir de fonts diverses
Els ous, generalment, són centrolecítics o, almenys, rics en vitel·lus. Hom ha detectat l’existència de quatre tipus de segmentació en els diversos grups d’aràcnids: segmentació total (rara i només en ous pobres en vitel·lus); mixta (típica d’aranyes, pseudoscorpins i alguns palpígrads), per la qual, després de les primeres divisions, els blastòmers interns fusionen el seu deutoplasma en una massa única comuna; superficial (en opilions, àcars, solífugs i algunes aranyes); i discoïdal (rara en els invertebrats, es dóna en certs escorpins, que presenten ous telolecítics).
La segmentació condueix a la formació d’una capa de blastòmers perifèrics que envolten la massa vitel·lina. A la zona ventral de l’embrió es diferencia, per efecte de la proliferació molt activa dels blastòmers, un disc germinatiu primordial separat en dues meitats simètriques per un lleuger solc sagital. És en aquest disc on la proliferació cel·lular dóna lloc a una delaminació, a la qual la capa externa es constitueix en ectoblast, mentre que internament es diferencia l’endomesoblast, que posteriorment es delimita amb més nitidesa.
És probable que la condició genèrica comuna als diferents grups sigui una segmentació total, amb la diferenciació d’un blastoderma perifèric i un endoderma vitel·lí interioritzat. La proliferació del disc germinatiu delimitaria les dues capes restants, l’ectoderma i el mesoderma, per delaminació. Però el que és cert és que es fa difícil establir un esquema comú i diferenciar quins són els caràcters cenogenètics i quins els palingenètics en aquestes fases decisives del desenvolupament.
La metamerització de l’embrió
El disc germinatiu, situat a la part ventral de l’embrió, s’allarga alhora que es produeix una fragmentació progressiva de les dues bandes mesodèrmiques i una diferenciació progressiva dels elements metamèrics de davant cap endarrere. Els acúmuls mesodèrmics metameritzats s’excaven i es diferencien les bosses celomàtiques que, per mitjà d’una prolongació digitiforme, indueixen a la formació dels primers esbossos apendiculars. Els esbossos dels quelícers i els pedipalps són els més avançats en aquest procés de diferenciació, però hi ha acúmuls mesodèrmics, previs als quelícers, de valor morfològic discutit.
Prèviament a la metameria real, les bandes mesodèrmiques es fragmenten, en molts casos, en diversos macrosomites, que posteriorment se subdivideixen i donen lloc als metàmers reals. Els somites prosòmics es diferencien abans que els opistosòmics, i els esbossos apendiculars es formen més aviat com més gran hagi de ser la seva dimensió posterior. També es formen, en la seva posició metamèrica corresponent, els esbossos de les estructures apendiculars opistosòmiques que, més endavant, originaran estructures ben diferents.
Els annexos embrionaris
El desenvolupament en un medi hostil com el terrestre fa que en aquests grups trobem algunes estructures que serveixen de protecció o que faciliten la viabilitat de l’embrió; per exemple, les estructures que diferencien alguns escorpins (serosa i amni) les cobertes fol·liculars, o el pseudoamni (en els pseudoscorpins) i altres estructures (en els àcars), totes protegides per la coberta del còrion. Són interessants determinades estructures accessòries de caràcter cenogenètic que diferencien alguns embrions com, per exemple, els òrgans laterals dels solífugs, l’aparell suctor dels pseudoscorpins, els òrgans d’eclosió de determinats grups (opilions, aranyes), i els òrgans absorbents i l’aparell dorsal dels escorpins.
Els fenòmens de blastocinesi
L’allargament de la banda germinativa i, posteriorment, la metamerització de l’embrió, fan que aquest se situï entorn de la massa vitel·lina en una posició corbada característica. La porció terminal de l’opistosoma en alguns grups (particularment, aquells que tenen un metasoma diferenciat) se separa de la massa vitel·lina per manca d’espai i es corba o flexiona en disposició ventral. No obstant això, el cas més curiós és el que protagonitzen les aranyes amb el fenomen de blastocinesi, conegut com a inversió. Aquest fenomen consisteix en una separació ventral de les dues meitats de la banda germinativa després d’haver-se produït la metamerització, de manera que aquelles es desplacen a banda i banda de l’esfera vitel·lina fins a reunir-se a la part dorsal, però mantenint-se sempre units els dos extrems anterior i posterior de l’embrió. En acabar-se aquest procés, l’embrió mostra una curvatura dorsal que fa que els seus extrems s’uneixin en el pla sagital, i tendeix a cobrir progressivament la part ventral, de manera que la massa vitel·lina queda englobada pels elements endoblàstics i, per tant, inclosa a la cavitat de l’arquènteron.
L’organogènesi
L’ectoderma, a més de donar origen al tegument, també en dóna al sistema nerviós, als òrgans dels sentits, als derivats apendiculars de l’opistosoma (com les estructures filotraqueals, les glandulars) i a l’estomodeu i el proctodeu del digestiu, principalment. En el sistema nerviós, la cadena ventral i els centres cerebroides es formen d’una manera independent i, posteriorment, una vegada interioritzats, entren en relació, bé com a invaginacions semicirculars (protocervell), bé com a engruiximents longitudinals metameritzats (neuròmer de la cadena ventral). En les formacions oculars, pel que fa a la seva ontogènia, hom pot parlar de dos tipus d’ulls: mitjans i laterals, com ja hem comentat anteriorment. En el cas de les estructures filotraqueals, l’esbós apendicular primari s’inclina sobre el tegument i, a la seva part posterior, es diferencien una sèrie de plecs. D’aquesta manera queda interioritzat l’esbós d’allò que seran els sacs pulmonars.
L’estomodeu es forma a partir d’una invaginació ventral en el lòbul anterior, alhora que es diferencien els apèndixs prosòmics. Aquest esbós, interioritzat, s’allarga i s’envolta de mesoderma, que originarà una musculatura potent, alhora que se subdivideix, almenys, en una porció esofàgica i una part faríngia. El proctodeu es forma, molt tardanament, a partir d’una lleugera i breu invaginació de l’extrem posterior.
L’endoderma, que genera la porció mitjana del tub digestiu (el mesodeu), pel que sembla, es genera a través d’elements cel·lulars difosos a la massa vitel·lina que es concreten en un epiteli circumdant, o bé per un procés morfogènic a partir de dues zones polars, anterior i posterior que, per proliferació, arriben a embolcallar la massa vitel·lina i entren en relació amb l’estomodeu i el proctodeu respectivament; l’esbós posterior dóna lloc, igualment, als tubs de Malpighi. De tota manera, aquest procés no és un fenomen general a tots els grups.
Com a derivats mesodèrmics, a més de les bosses celomàtiques, sorgeixen el sistema circulatori, les glàndules coxals, l’aparell genital i la musculatura. Cada antímer, situat inicialment a la cara ventral, a la banda germinativa, es fragmenta en tres parts: la part dorsal creix cap al dors i origina el vas dorsal; la part ventral dóna origen a les glàndules coxals, en el prosoma, i a les gònades de l’opistosoma; les parts mitjanes indueixen la formació dels apèndixs i donen lloc a la seva musculatura. Els derivats celomàtics sofreixen una dissociació primerenca, de manera que el celoma es confon amb la cavitat general del cos.
Les fases juvenils

Durant el creíxement, els aràcnids, com tota la resta d’artrópodes, muden la cutícula vella per una de nova per a iniciar un nou estadi de desenvolupament. La fotografia mostra una exúvia d’aranya, de la qual s’ha desprès la part posterior, corresponent a l’opistosoma. Noteu la persistència de la forma del cos i els apèndixs en la cutícula i la dels elements cuticulars externs.
Ramon Torres.
S’acostuma a dir que els aràcnids presenten un desenvolupament directe, ja que l’individu que surt de l’ou després de l’eclosió, té una gran semblança amb l’adult, i el seu creixement, a través d’una sèrie de mudes, simplement completa la fàcies general i estructural d’aquest darrer. No obstant això, aquestes formes que sorgeixen de l’ou, en general, són força desvalgudes i imperfectes, atès que la seva organogènesi encara no és acabada (manquen els ulls, les potes no són ben perfilades, etc.). D’altra banda, en els pseudoscorpins i els àcars hi ha una sèrie de transformacions en aquesta fase del desenvolupament, que permeten parlar d’una metamorfosi i, per tant, de desenvolupament indirecte; la conseqüència és un desacord considerable en la terminologia que designa aquestes fases postembrionaris (prelarves, larves, nimfes, etc.).
Ecologia i comportament
L’interès ecològic dels aràcnids radica en el seu paper de consumidors secundaris, ja que constitueixen un graó important de les cadenes tròfiques dels ecosistemes terrestres. Ja hem indicat que es tracta de petits depredadors, que porten a terme una missió insubstituïble en el control natural de les poblacions d’insectes.
La seva actuació, d’una manera paradoxal, és poc aparent, però una anàlisi detallada de qualsevol ecosistema terrestre en posa en evidència la importància. Es tracta d’un grup poc perceptible als nostres ulls en el medi natural, però els sistemes de mostratge especials i les diferents metodologies que permeten la seva quantificació ens parlen invariablement de la seva presència, la seva diversitat i àdhuc la seva densitat poblacional (especialment en el cas dels opilions i les aranyes). En aquest sentit, s’han difós alguns càlculs teòrics, com el de Turnbull (1973), segons el qual —en condicions òptimes— i per a un medi herbaci, s’aprecien densitats de 130 individus per metre quadrat. Si hom considera que una sola aranya pot consumir 0,1 g al dia, la conclusió és que les aranyes que viuen en una hectàrea consumirien 47 500 kg (1,3 × 106 × 0,1 × 365 = 4,75 × 107).

Els hàbits depredadors, generals a tots els aràcnids, s’acompanyen de característiques morfològiques peculiars, tant pel que fa a la qualitat prensora de determinats apèndixs, com per la coloració críptica que els facilita l’operació de caça. Ambdós fenòmens queden ben il·lustrats a la fotografia de l’escorpí groc (Buthus occitanus), presa al massís de Garraf (a dalt), i en la d’un tomísid (Synaema globosum, a baix), capaç de variar el color de fons del cos, blanquinós o groguenc, segons els colors del substrat que el sosté.
Oriol Alamany i Josep M. Moraleja.

El grau de cripsi que poden aconseguir els aràcnids és especialment gran en les aranyes atès que presenten tota una gamma de colors molt variada, en relació als de la resta dels grups. En el cas dels tomísids que viuen a les flors, dels quals veiem dos exemplars a les fotografies (Thomisius albus), aquesta capacitat d’adequació del color esdevé singularment remarcable.
Josep M. Barres i Josep M. Moraleja.
Com passa en altres grups, en els aràcnids cada espècie es troba adaptada a un medi concret, de manera que són relativament poques les que tenen un nínxol ampli. Pràcticament les 50 000 espècies que pertanyen a aquest grup mostren un nínxol ecològic ben definit, del qual no es poden sostreure fàcilment. En contraposició a aquest fenomen adaptatiu, la diversitat és molt gran, particularment entre les aranyes, de manera que cada ecosistema sol posseir una fauna pròpia característica i estable.
Els aràcnids, en línies generals, solen adoptar estratègies reproductores que impliquen, entre altres aspectes, una descèndencia molt abundant, seguida d’una elevada mortalitat de les fases juvenils. Els seus cicles biològics s’adapten d’una manera clara a les fluctuacions generals del medi, especialment a les regions temperades, on el desenvolupament s’accelera o es retarda en funció de les variacions fenològiques en les condicions fisiogràfiques. D’aquesta manera, ens trobem, independentment de la longevitat individual (en general escassa, de menys d’un cicle anual) amb un o més períodes reproductors al llarg de l’any que, segons els casos, se situen a la primavera, l’estiu o la tardor, i a"amplitud molt variada (formes euricrones, estenocrones, diplocrones, etc.).
Ignorem, encara, molts aspectes de l’activitat dels aràcnids. Hom va descobrint, cada vegada més, fenòmens d’inactivitat de caràcter diapàusic. Però també hi ha casos d’una aparent activitat durant tot el cicle, vinculada a fenòmens de migració vertical, especialment en formes diminutes que poden utilitzar sense dificultats els medis hipogeu i endogeu, a més del medi epigeu superficial; en el cas de l’alta muntanya, també hi ha casos d’activitat sota la neu.

Les formes de vida aquàtiques són excepcionals dins dels aràcnids. Únicament entre els àcars hi ha un grup de vida veritablement aquàtica, el dels hidràcars, al qual correspon l’exemplar de la fotografia de dalt, pertanyent al gènere Arrenurus, que és molt comú a les basses i les llacunes d’aigües dolces. D’altra banda, entre les aranyes, a part del gènere Argironetta, absent del nostre país i l’únic que viu veritablement a l’aigua, n’hi ha algunes que menen una vida relacionada amb els ambients aquàtics, que hom pot veure de vegades caminant per damunt de l’aigua, a les zones marginals, com el licòsid de la fotografia de baix.
María José Gil i Marcel·la Chinchilla.
Els aràcnids tenen una activitat críptica o poc aparent, cosa que, no hi ha dubte, es relaciona amb la seva condició depredadora. Per aconseguir l’aliment utilitzen tècniques molt variades, fins al punt que aquestes i les seqüències depredadores poden, per elles mateixes, caracteritzar molts grups sistemàtics. La depredació, en els aràcnids, no és exhaustiva sinó oportunista, tant si utilitzen trampes com si no. Aquests animals reuneixen dues característiques: una golafreria exagerada quan disposen d’aliment abundant (cas en què el seu límit, pel que sembla, ve marcat per processos interns de tipus neuroendocrí i pel grau de distensió opistosòmica) i una capacitat de dejuni admirable que, en casos extrems pot portar a l’exhauriment de les reserves i a un "stress" alimentari que els pot suposar la mort. La tècnica de caça predominant sol ésser la de l’aguait i l’atac fulminant més o menys espectacular de les preses. Un exemple manifest és el de les aranyes de les flors (fonamentalment del grup dels tomísids), que adopten tons mimètics i una immobilitat absoluta, cosa que els permet de situar-se a la corol·la de les flors esperant l’arribada de les preses. També és coneguda la potencialitat de les aranyes de fabricar teles o teranyines de seda. Aquestes teles, d’estructura molt variada, serveixen alhora d’habitacle i de trampa de caça, i la seva eficàcia radica en bona mesura en la seva situació i orientació. La cacera a l’aguait també és present en altres grups d’aràcnids i és en consonància amb l’extraordinari desenvolupament del sistema sensorial, particularment el mecanoceptor, capaç de detectar les mínimes vibracions de l’entorn.
El fet de dominar les preses en un temps molt curt juga un paper fonamental en l’èxit de la depredació. Es per això que alguns aràcnids han desenvolupat fortes pinces en els pedipalps, o quelícers robusts, o bé glàndules que segreguen un verí capaç d’immobilitzar les preses; en altres casos, les emboliquen i les subjecten amb la seda que emeten les fileres. La pròpia condició de depredadors fa que els aràcnids siguin animals solitaris, capaços d’aniquilar els seus propis congèneres en cas d’interpretar-los com una presa. Per això, en aquest grup, els fenòmens socials o el gregarisme són excepcionals, bé que es donen en algunes aranyes i opilions. Això exigeix, igualment, tot un ritual complex per a l’aparellament; en molts casos, els mascles adults alteren la seva conducta i cercen activament les femelles per copular, procés en el qual juguen un paper decisiu l’emissió de feromones que es dipositen a la seda o al substrat, o que es volatitzen. El mascle no té cap altra missió biològica que la fecundació de la femella, fins al punt que en molts casos no s’alimenta i es mor poc temps després.
Són pocs els coneixements que tenim pel que fa als depredadors naturals dels aràcnids. Deixant de banda alguns exemples clàssics i vistents com el dels himenòpters pompílids o el dels dípters oncòdids, cal no despreciar la parasitosi per part de nematodes, que en molts casos es troben envaint l’opistosoma fins a matar el seu hoste. És probable, però, que siguin els ocells els consumidors més eficaços d’aràcnids, gràcies a la consistència tova dels teguments i al fet que els seus verins són inactius per via digestiva.
Distribució geogràfica

El curiós mecanisme de l’aerodispersió, que permet a les aranyes joves allunyar-se de l’àrea geogràfica materna, col·labora d’una manera activa a perfilar l’àrea de distribució de les espècies. A la fotografia podem veure una aranya, en un dels seus primers estadis de desenvolupament, en la posició d’esperar ésser dispersada pel vent.
Ramon Torres.
Poca cosa es pot afirmar, de caràcter general, al voltant de la distribució geogràfica dels aràcnids. L’àrea de distribució de les nombroses espècies que els constitueixen sol ésser limitada (fins i tot en espècies de ràpids i eficaços mecanismes de dispersió). No obstant això, l’elevat grau de diversificació assolit en alguns casos (fonamentalment, aranyes i àcars) han fet possibles adaptacions molt heterogènies, de manera que poden considerarse casos de cosmopolitisme, representats a pràcticament tots els ambients terrestres. Per contra, els grups que no gaudeixen d’un grau de diversificació tan gran es troben relegats a les zones tropicals i intertropicals més càlides. L’àrea de distribució és condicionada, en gran mesura, per dos factors: els mecanismes habituals de dispersió que puguin posseir, i les limitacions fisiològiques de cada espècie, especialment les que condicionen la seva capacitat de reproducció. En el primer d’aquests aspectes, els aràcnids es manifesten molt heterogèniament, ja que hi ha des de formes estàtiques, que només utilitzen un desplaçament lent sobre el substrat fins a d’altres que es deixen portar pel vent i poden desplaçar-se fins a centenars de quilòmetres del seu lloc de naixement en molt poc temps. En el segon aspecte, pel que sembla, es troben molt més limitats. En línies generals, es pot dir que els aràcnids són formes preferentment tropicals o subtropicals, ja que és en aquestes latituds on es troba la seva diversitat més gran, i únicament alguns grups (aranyes, opilions i àcars) han diferenciat formes capaces d’adequar-se a les àrees temperades o fredes del medi terrestre i fins i tot diversificar-s’hi.
Els aràcnids presents als Països Catalans
Set dels onze ordres d’aràcnids que comprenen formes vivents tenen representació a la fauna dels Països Catalans. A més de les aranyes típiques (aranèids), aquests grups inclouen els escorpins, ben coneguts de tothom; els pseudoscorpins, que, per la seva forma semblen petits escorpins sense cua; els solífugs, organismes veritablement rars al nostre país; els palpígrads, petits aràcnids habitants de les coves; els opilions, sovint confosos amb les aranyes per la seva forma, comuns i de les mateixes dimensions que aquestes; i els àcars, grup molt divers al qual pertanyen, entre altres, les paparres dels gossos. Entre els àcars hi ha l’únic grup d’aràcnids adaptat perfectament a la vida aquàtica; tota la resta són animals terrestres i, majoritàriament, predadors. De tots aquests grups, ens n’ocuparem en les pàgines successives, després de fer un petit recordatori dels camins que ha seguit el seu estudi al nostre país.
Sistemàtica i filogènia
Hom reuneix, dins de la classe dels aràcnids, els ordres següents: àcars (Acari), amblipigis (Amblypygi), aranyes (Araneae), opilions (Opiliones), palpígrads (Palpigradi), pseudoscorpins (Pseudoscorpiones), ricinulis (Ricinulei), esquizòmids (Schizomida), escorpins (Scorpiones), solífugs (Solifugae) i uropigis (Uropygi). Els ordres restants corresponen exclusivament a formes fòssils: els custaràcnids, el haptòpodes, els antracomarts, els arquitarbs, i els trigonotarbs.
La diversitat sistemàtica de la classe dels aràcnids ofereix poques dificultats a l’hora d’acceptar la individualitat de cadascun dels seus ordres i, en tot cas, únicament la complexitat i la originalitat dels àcars ha donat origen a un desdoblament sistemàtic en alguns esquemes recents. No obstant això, l’establiment de connexions evolutives que pugui explicar aquesta diversitat constitueix una de les incògnites de l’aracnologia. Són diverses i desiguals les hipòtesis emeses en aquest sentit, i únicament les afinitats entre alguns parells d’ordres semblen constituir fets ben establerts.
Savory (1971) distingeix cinc línies bàsiques, sense especificar, però, cap connexió entre elles. La línia que sembla ésser bàsica o central és encapçalada pels aranèids (com a grup més important), ordre que es relaciona estretament amb els amblipigis, si es prescindeix de la sèrie de caràcters apomorfs que els individualitzen. Un segon binomi s’integra en aquesta línia d’evolució: uropigis i esquizòmids, grups clarament emparentats, tot i que actualment es consideren com a ordres independents. A aquest complex de formes s’uneixen els palpígrads que, malgrat les seves adaptacions al medi edàfic, han conservat alguns trets d’organització que els apropen a les suposades formes primitives d’aquesta línia bàsica d’aràcnids. Savory situa en aquesta línia, també, l’ordre fòssil dels custaràcnids. Una segona línia d’evolució, que mostra caràcters propis i una gran cohesió, és constituïda, en essència, pel binomi opilions-àcars. Ambdós ordres es relacionen clarament a través dels subordres cifoftalms-notostigmates, respectivament. A aquest bloc de formes s’assimilen dos altres grups: l’ordre dels ricinulis i l’ordre fòssil dels arquitarbs. Savory defensa al seu esquema el parentiu entre els escorpins i els pseudoscorpins. Ambdós grups es relacionen amb els solífugs que, en ostentar una sèrie de caràcters morfològics considerats més primitius, representen un pas previ en el procés evolutiu. Aquest conjunt constitueix la tercera línia evolutiva. Les dues línies restants són integrades únicament per ordres fòssils: d’una banda, l’ordre dels trigonotarbs, de característiques peculiars i, de l’altra, els heptàpodes i els antracomarts, les connexions morfològiques dels quals permeten reunir-los en una cinquena línia d’evolució dins dels aràcnids.
Hi ha, però, altres punts de vista, que valoren d’una manera diferent el conjunt de caràcters dels aràcnids. Per exemple, les classificacions evolutives de Petrúnkevitx, Dubin, Grasshoff, Van der Hammer, etc., donen lloc a dendrogrames diferents. En definitiva, a tots aquests esquemes hi ha un problema de base: la connexió filètica entre tota aquesta sèrie. Tampoc manquen arguments que discuteixen la unitat filètica de la classe dels aràcnids, tal i com taxonòmicament és considerada en aquesta obra. La dificultat més coneguda en aquest sentit recau en les afinitats amb altres grups de quelicerats merostomats (els euriptèrids) que s’observen entre els escorpins o les peculiaritats de la línia opilions-àcars contra l’estoc bàsic format pels palpígrads, aranèids i amblipigis.