
Còpies en guix dels «Würzburger lügensteine» (105-110 mm d’altura), els falsos fòssils que Johann Beringer figurà en la seva extensa obra «Lithographiae Wirceburgensis» (1726) ignorant que es tractava d’un intent deliberat dels seus col·legues universitaris de desacreditarlo. Quan se n’adonà, Berlinger va comprar tots els exemplars del seu llibre que va poder i els va destruir, per la qual cosa l’obra esdevingué una de les curiositats més singulars de la literatura paleontològica.
Jordi Vidal / MGB.
La paleontologia és una ciència que estudia els fòssils com a elements per a entendre la vida en el passat geològic i com a eina per a explicar la diversitat de la vida actual. Al llarg de la història de la paleontologia hom ha passat de considerar els fòssils com a «ludus naturae» (jocs o capricis de la natura) a veure-hi els representants d’antigues catàstrofes que havien assolat el nostre planeta (teories catastròfiques) o bé la prova de l’existència de l’evolució. Gairebé tots els qui generaren teories d’una tendència o altra en el camp de la paleontologia o de la geologia, utilitzaren i utilitzen els fòssils per a reforçar les seves tesis. Fins i tot, darrerament, les de l’Archaeopteryx (l’ocell més antic que es coneix, del Juràssic superior), han estat considerades falses, com si es tractés d’un producte artesanal preparat pels científics per a donar suport a les seves teories. Res no és més lluny de la realitat ja que, si bé és cert que, històricament, hi ha hagut certs fraus en aquest camp (d’altra banda ben coneguts, com els falsos fòssils de Beringer, al segle XVIII; o el crani de l’home de Piltdown, a Anglaterra, al segle passat), aquests no han fet sinó refermar totes les precaucions dels paleontòlegs des del moment de recol·lectar un fòssil, en la seva minuciosa documentació i en la seva identificació i adscripció, sia a una cenozona o a una biozona paleontòlogica, sia a un nivell, pis o període geològic.
Els fòssils i la fossilització
Concepte de fòssil
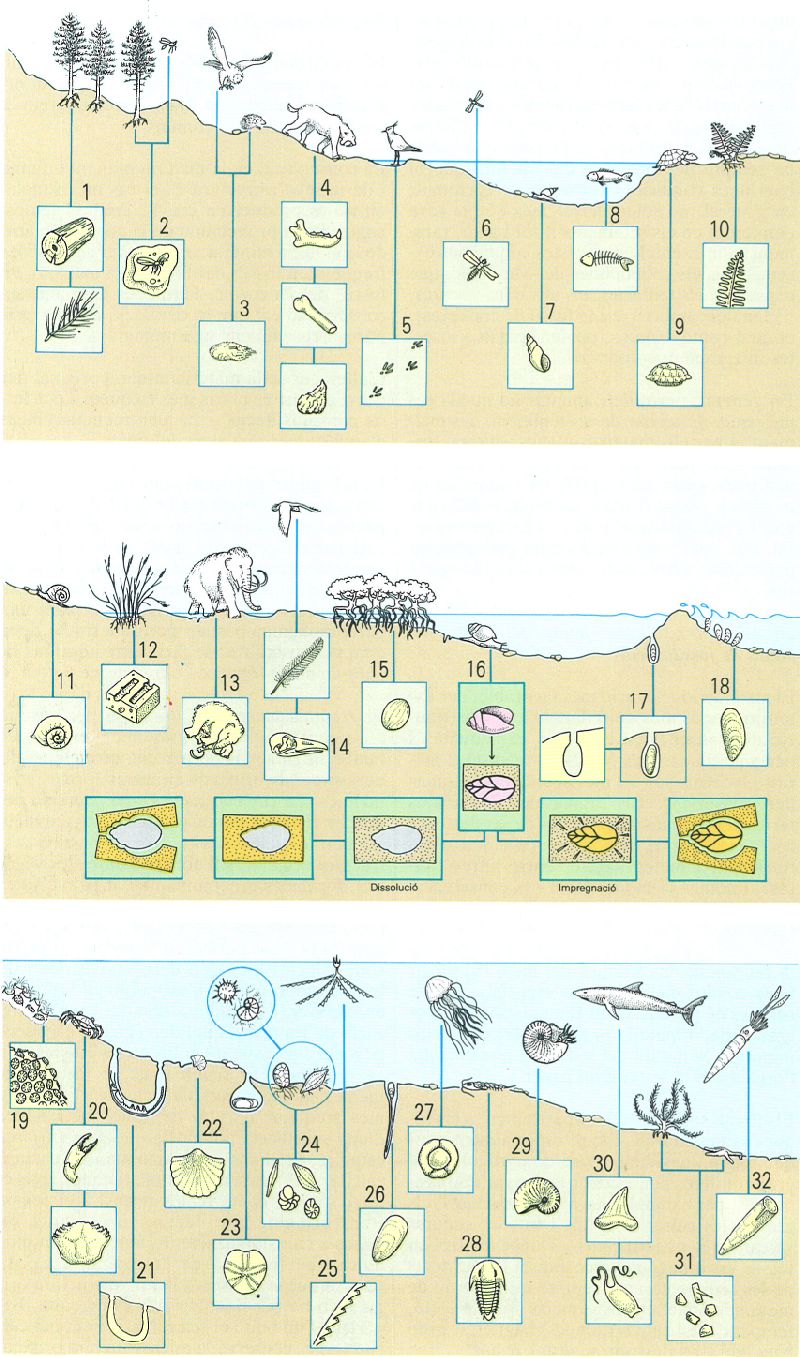
Síntesi de reconstrucció paleotafonòmica. En ambients continentals i litorals (a dalt i al mig), els vegetals són representats per troncs i fulles (1), frondes carbonitzades (10), herbes impregnades fent travertins (12), fruits (15), espores i pol·len; els invertebrats, per gasteròpodes, terrestres (11) o de zones fluvials o límniques (7), i insectes en ambre (2) o en sediments de gra molt fi (6); i entre els vertebrats, plomes i ossos (14), icnites (5) i egagròpiles d’ocell (3), dents de mamífers (4) o grans exemplars sencers, conservats en gel, pous d’asfalt o antigues torberes (13); ossos, i excepcionalment ous, d’amfibis i rèptils (9), esquelets sencers de peixos (8), de vegades amb escates. En els ambients marins, els foraminífers són abundants formant lumaquel·les (24); els cnidaris deixen simples impressions (27) o grans esculls coral·lins (19); també s’hi troben braquiòpodes (26) i lamel·libranquis fixats en substrats durs (18) o al seu interior (17), on deixen marques d’ocupació, o en substrats tous (22); gasteròpodes (16), cefalópodes nautiloïdeus i ammonoideus (29), i fragmocons de coleoïdeus (32); pinces i closques de decàpodes (20), excavacions d’altres crustacis (21), exosquelets o exúvies de trilobits (28), esquelets calcaris d’equinoderms, disgregats (31) o sencers (23), i estipes o rabdosomes carbonitzats de graptòlits (25); i dents i postes de selacis (30). Al centre de la figura (fons verd) es mostra el procés general de petrificació: un cop mort l’organisme i enterrat en el sediment pot produir-se la impregnació (sals minerals de l’aigua del sediment rebleixen els porus de les parts dures, o bé les sals que formen l’esquelet són reemplaçades per altres del sediment i en resulta un fòssil i un motlle extern) o la dissolució (el buit deixat per l’organisme és ocupat per substàncies minerals, i se n’obté un motlle intern i un d’extern).
Amadeu Blasco, original de l’autor.
El mot fòssil (del llatí fossilis, excavat) ha estat utilitzat de moltes maneres per diferents autors. La majoria accepten que un fòssil ha de ser una evidència de l’existència d’un organisme que va viure en el passat. Alguns autors especifiquem més: també ha de subministrar alguna idea de la natura (forma, dimensions, etc.) d’una part de l’organisme o de tot sencer. A més, un fòssil ha de tenir una certa edat; aquest requeriment és tan intangible i indeterminable que fixar el límit entre el que és no fòssil, subfòssil i fòssil en una etapa determinada de la història de la conservació d’un organisme pot resultar completament arbitrari. També hi ha qui insisteix que el fòssil ha d’haver estat preservat en materials de l’escorça terrestre per agents i processos naturals. Malgrat això, sol aplicar-se el terme de fòssil només quan la matèria orgànica desapareguda de l’organisme ha estat reemplaçada per compostos minerals, és a dir, quan s’ha petrificat o transformat en pedra, tot i que també sol emprar-se en casos d’organismes momificats en asfalts o conservats en gel d’una antiguitat considerable.
El procés de fossilització o tafonòmic d’un organisme requereix tot un seguit de circumstàncies. En primer lloc, cal que l’organisme mort quedi ràpidament enterrat en un sediment (de dimensions de gra tan fines com sigui possible) que, en recobrir-lo, el protegeixi dels elements destructors, com l’erosió, etc. Efectivament, quan un organisme mor, les seves parts toves són atacades i assimilades ràpidament per l’acció de bacteris i d’altres organismes necròfags, de manera que gairebé sempre desapareixen del tot fora que hi hagi unes condicions realment excepcionals; en general, no solen deixar més que la seva empremta en els sediments i, tot i això, rarament. Són exemple d’aquestes condicions excepcionals els cefalòpodes del Mesozoic, que conserven els tentacles, els ulls o les vísceres, o el mateix ambre (resina fòssil de coníferes), el qual conté espores, pol·len, pètals i insectes íntegrament conservats.
En general, però, dels animals no queda res més que els teixits de sosteniment, normalment ja per ells mateixos càlcics, silicis o fosfàtics, i les parts dures: closques de microorganismes, esquelets de polípers, conquilles de mol·luscs, ossos, dents i escates de vertebrats, etc. La substància original, de caràcter mineral, s’ha conservat o bé ha estat reemplaçada per calcita, sílice, guix, pirita o altres minerals.
Medis de fossilització
El medi de fossilització més favorable, per extensió, condicions de sedimentació i distribució d’organismes en el seu si, és l’aquàtic, i per aquesta raó, els fòssils d’organismes marins i lacunars són els més abundants i els més ben conservats. Els organismes anomenats terrestres són més susceptibles de ser destruïts (per putrefacció, necrofàgia, dificultat d’enterrament i meteorització, entre altres causes), i només la momificació i la conservació en gel els ofereixen grans possiblitats de preservació.
A les condicions físiques del medi cal afegir unes condicions químiques especials, ja que cal que es produeixi un reemplaçament progressiu de la matèria orgànica per compostos minerals; la natura del sediment que engloba l’organisme hi té un gran paper.
Els fòssils solen aparèixer bàsicament a les roques sedimentàries, i molt especialment a les que tenen unes dimensions de partícula o de gra prou fines per a poder preservar els organismes: per exemple, les argiles, els llims, els gresos, les calcàries i, fins i tot, les cendres volcàniques estratificades. També es troben fòssils en roques metamòrfiques (procedents de les sedimentàries), sempre que el grau de metamorfisme (transformació per pressió, temperatura i canvis químics) hagi estat prou baix per a no destruir aquestes restes.
Mecanismes de fossilització



Els resultats finals de la tafonomia poden ser variats, com ens il·lustren aquestes imatges. La primera (a dalt) és una vista dorsal d’una tortuga (Trionyx marini) procedent de l’Oligocè d’Almatret (Segrià) que mesura uns 20 cm de llargària, i és un bon exemple de carbonització, és a dir, de reemplaçament amb pèrdua d’elements originals i enriquiment en altres elements. La segona (al mig) mostra un exemplar de Pseudohaploceras (5 cm), ammonoïdeu de l’Aptià inferior de la Vall de Cabó (Alt Urgell), al qual s’evidencia clarament el procés de piritització (substitució per pirita, en aquest cas ja alterada a limonita) del motlle intern de les primeres voltes del cefalòpode; la closca original, d’aragonita, ha desaparegut, bé per recristal·lització a calcita i posterior dissolució d’aquesta, bé per dissolució directa de l’aragonita. La tercera imatge (a baix) correspon a un motlle intern de Scutella faurai del Tortonià de Montjuïc (Barcelona), d’uns 11 cm de llargària; la mateixa aigua del sediment que provocà la dissolució de la closca d’aquest equinoïdeu ha dissolt la sílice dels sediments (gresos silicis) i l’ha dipositada en forma d’òpal, impregnant els espais buits del motlle intern.
Jordi Vidal / MGB-UAB
Hi ha diferents mecanismes relacionats amb diversos aspectes químics que fan que un organisme esdevingui fòssil. Enumerarem a continuació els més comuns.
La conservació de la matèria mineral original i de la seva estructura és el cas més simple, on no es produeixen canvis. Amb el temps, augmenta la probabilitat de la no-conservació de la matèria mineral original; per això és més freqüent en fòssils del Pleistocè i més rara en fòssils del Paleozoic. Perquè es doni aquesta conservació cal que la circulació d’aigua pels porus del sediment sigui mínima.
L’alteració de la matèria mineral original per canvi químic és el cas més freqüent i pot ferse per substitució o bé per reemplaçament d’aquesta per una altra del medi.
La substitució pot donar com a resultat la conservació de l’estructura original o no; en el primer cas, es produeix una substitució molècula per molècula; en el cas de boscos o vegetals silicificats, la degradació de les parets de les cèl·lules vegetals tendeix a provocar la pèrdua de la cel·lulosa original, deixant una paret de lignina bastant porosa a través de la qual poden infiltrar-se fàcilment solucions riques en sílice (òpal).
El reemplaçament pot donar-se per pèrdua d’elements originals i enriquiment en d’altres també originals, o bé per una reorganització dels minerals originals en altres formes cristal·lines. La carbonització és un exemple del primer tipus, que sol produir-se a pressió i temperatura elevades i fa que es conservi l’estructura original; sol donar-se en restes vegetals, formades predominantment per cèl·lules i lignina, i en alguns invertebrats rics en quitina, com ara els artròpodes terrestres i els graptòlits. La pèrdua d’elements químics, com oxigen i hidrogen dels polisacàrids i de les escleroproteïnes, a més d’un enriquiment en carboni, dóna com a resultat la carbonització de les restes que, de vegades, s’evidencien per una capa de grafit en les roques que els contenen. El segon tipus de reemplaçament s’anomena recristal·lització i l’exemple més freqüent n’és el de fases metastables d’una substància polimorfa que passa a fases estables; és el cas de l’aragonita que passa a calcita. La naturalesa del medi aquós amb el qual són en contacte les restes, i el temps transcorregut, són factors importants en aquesta recristal·lització. El fenomen pot produir-se per inversió de l’aragonita a calcita (cas en què es conserva la microstructura original) o bé per dissolució de l’aragonita, l’aparició d’un buit i el seu reompliment per calcita (no es conserva la microstructura original i apareix l’estructura espàtica de la calcita). En organismes silicis, la recristal·lització afecta el pas d’òpal a la calcedònia.
Aquestes substitucions o reemplaçaments poden fer-se també en altres materials, com la sílice, la pirita i la marcassita, el fosfat càlcic o el guix. Parlem, per exemple, de silicificarió o de piritització per a denominar el conjunt de canvis químics produïts en el fòssil.
La impregnació pot aparèixer en qualsevol dels casos anteriors quan la resta esquelètica queda reforçada per la precipitació de minerals addicionals (que es troben en l’aigua del medi) en els espais porosos deixats per la matèria orgànica en els esquelets. Entre els minerals d’impregnació més freqüents hi ha la calcita i la sílice.
La incrustació fa que el material esquelètic quedi envoltat de crostes, especialment de calcita, produïdes o bé per activitat algal, o bé per precipitació química d’aigües sobresaturades (ooides i travertins).
La conservació indirecta resulta d’una diagènesi desmineralitzadora que provoca una dissolució de les restes i l’aparició dels anomenats motlle intern i motlle extern. El motlle intern és format per sediments o per un farciment cristal·lí precipitat en closques molt tancades, que eren a l’interior de la resta abans de la dissolució. El motlle extern és la marca deixada per la superfície externa de la resta en els sediments que la contenien; aquest motlle pot fornir molta informació sobre el fòssil que l’originà i permet de treure’n un positiu per mitjà de l’ús de guix, plastilina, silicones o altres matèries plàstiques. En el cas dels vertebrats, poden produir-se motlles endocranials, és a dir, motlles de la cavitat interna del crani, que informen sobre les circumvolucions cerebrals de l’animal i, per tant, del grau d’evolució de les espècies fossilitzades. Les dissolucions que originen els motlles poden donar-se després que l’esquelet hagi romàs en estat fòssil un cert temps a causa de l’aigua que circula pels sediments.
La conservació excepcional és referida a la que ha preservat les parts toves dels animals. En són exemples els insectes inclosos en ambre, la inclusió en nòduls de sílice de flagel·lats de la creta, d’algues cianofícies i verdes i de bacteris del Precambrià i d’alguns graptòlits, els mamuts i rinoceronts llanuts congelats, la conservació en basses de quitrà de vertebrats i invertebrats, la momificació per mort en ambient desèrtic i deshidratació dels teixits d’alguns dinosaures, i la impressió de les parts toves (jaciments d’Alcover i Mont-ral, de Rúbies, de Libros, de Solnhofen o de Holzmaden).
Els icnofòssils

La varietat d’icnofòssils que hom pot trobar al camp és elevada. Les icnites evidencien el pas de vertebrats damunt de sediments poc consolidats, normalment en medis continentals o de transició; a dalt veiem les d’un vertebrat proveït de mans pentadàctiles del Buntsandstein des Recó de S’Algar (Mallorca), que hom atribueix genèricament a Cheirotherium; al mig a l’esquerra, icnites procedents de l’Eocè superior de Ledia (Navarra), que corresponen típicament a ocells i que són d’adscripció taxonòmica més difícil. Però els icnofòssils no apareixen només com a marques en el substrat sinó també damunt d’altres organismes: la closca del gasteròpode de la fotografia (× 2) del mig (a la dreta) ha sofert l’acció de diferents organismes, entre ells els litòfags; aquesta és una de les maneres en què els organismes contribueixen als processos sedimentaris tot reciclant matèria d’origen orgànic en bioclasts. Pel que fa a les pistes d’arrossegament (a baix a l’esquerra), Paleodictyon té forma de xarxa hexagonal i encara no ha pogut ser adscrita a cap organisme; l’exemplar procedeix de les turbidites eocenes de la carretera de Campdevànol a Gombreny, i mesura uns 40 mm. Helminthoides (a baix a la dreta) ha estat considerat com una marca deixada sobre el fons per algun gasteròpode en el seu procés alimentari; l’exemplar ve de les turbidites santonianes del barranc de Montsor (Pallars Jussà) i fa uns 175 mm d’altura.
Feliu Calafat, Bartomeu Saez, Oriol Riba, Jordi Vidal / Col·lecció Jordi Martinell i UAB.
La paleoicnologia estudia la naturalesa i el comportament dels animals del passat per mitjà de les traces, les pistes, les perforacions, les marques i altres restes que són producte de la seva activitat. Són el que hom anomena icnofòssils («trace fossils»), i representen marques d’allotjament i/o desplaçament en medis tant aquàtics com terrestres, i són especialment predominants en els primers. També formen part dels icnofòssils restes com els copròlits, les egagròpiles i els ous, que apareixen com a resultat de l’activitat alimentària i reproductora dels vertebrats.
S’han conservat generalment en les interstratifícacions de dos tipus de sediments diferents (marques exògenes) o dins d’un estrat (marques endògenes), i s’han generat quan aquest es trobava en un estat plàstic o susceptible de ser retreballat, cosa que, en molts casos, ha provocat la biotorbació (destrucció de les làmines) total o parcial d’un sediment; fins i tot, en certs casos, després de la diagènesi d’un substrat determinat (cas de les perforacions de litòfags). Moltes vegades, la situació espacial dels motlles o contramotlles d’aquestes marques s’utilitza com un criteri de polaritat dels sediments que les contenen.
Encara que els inicis de la paleoicnologia es remunten al primer quart de segle XIX, l’estudi d’aquestes restes s’ha revaloritzat els darrers vint-i-cinc anys, fins a considerar-se com una eina important per a una més bona comprensió de certs ambients sedimentaris del passat, i com un dels elements més importants per a entendre el comportament dels animals que hi visqueren. I és així perquè aquestes marques, perforacions, pistes i traces solen ser tan característiques que, malgrat desconèixer amb exactitud l’animal que les originà, poden ser utilitzades per a definir biofàcies i, de vegades, com a fòssils guia per a determinar l’edat geològica dels nivells que les contenen. Moltes vegades, la clau per a la interpretació d’aquestes marques s’ha trobat en l’estudi i la investigació de les que deixen els organismes actuals.
Per a la nomenclatura dels icnofòssils hom segueix les mateixes normes de nomenclatura binomial, tot i que es tracta de paratàxons. Entre els innombrables sistemes de classificació que poden utilitzar-se, el que sembla ajustarse més bé a la finalitat de la paleoicnologia és el de la classificació genètica i ecològica, sense oblidar-ne d’altres, com la morfològica i preservacional (que, malgrat la seva utilitat, es fonamenta en caràcters descriptius força arbitraris) i la filogenètica (que disposa encara de poques dades i que és bastant incerta a l’hora d’atribuir les marques a espècies concretes).
La classificació genètica i ecològica proposada per Seilacher (1964) i modificada posteriorment pretén agrupar els icnofòssils d’invertebrats en sis grups que, malgrat que s’encavalquen lleugerament, són d’utilitat. Un primer grup és el de les marques d’arrossegament (Repichnia), i inclou les pistes d’elements que es desplacen per una superfície, per exemple, Cruziana o Scolicia. Un altre grup és el de les marques de repòs (Cubichnia), algunes de les quals apareixen com a continuació de les marques d’arrossegament; per exemple, Rusophycus, Cardioichnus o Asteriacites. Un tercer grup són les marques de detritívors epibentònics (Parcichnia); algunes traces, com Helminthoides, s’assemblen molt a les que deixen els gasteròpodes actuals en els fons marins en la seva recerca d’aliment. Les marques d’alimentació (Fodichnia) abasten les excavacions endobentòniques efectuades per detritívors generalment de vida epibentònica però que excaven per a cercar l’aliment; entre altres inclouen tubs en forma de lletra U, com Rhizocorallium, i estructures radials proveïdes d’una gran quantitat de ramificacions, com Chondrites. Un altre grup són les marques d’ocupació (Domichnia), que són excavacions («burrows») i perforacions permanents realitzades per animals suspensívors que viuen dins dels sediments però que retenen per a la seva alimentació les partícules en suspensió que hi ha a l’aigua; els tubs poden ser des de verticals subcilíndrics, com a Skolithos, fins a tubs en forma de lletra U. Les marques d’escapament (Fugichnia) són tubs verticals que suggereixen moviments en vertical, ascendents o descendents, que feien alguns organismes a mesura que el sediment s’acumulava o s’erosionava per tal de no ser completament sepultats o quedar al descobert (Diplocraterion, per exemple). Aquesta classificació no és exhaustiva, ja que avui encara hi ha moltes marques que no poden ser incloses en aquests grups, com és el cas de Paleodictyon, que té forma de xarxa hexagonal.
En fàcies continentals són més corrents les icnites o petjades de vertebrats, i són relativament comunes les corresponents a rèptils, amfibis, ocells i, més rarament, mamífers. Produïdes sobre terrenys plàstics per animals que es desplaçaven mitjançant les extremitats, cada petjada correspon a la impressió d’una pota, cosa que demostra la seva presència en certs ambients i això aporta dades sobre l’anatomia de l’aparell locomotor (nombre i disposició dels dits), la forma i la mecànica de locomoció, i relacions d’associació entre elements faunístics diversos. Són característiques les petjades d’ocells amb la impressió de tres dits dirigits endavant i un endarrere.
Tal com sol passar en el cas dels invertebrats, les petjades només es troben excepcionalment associades a restes fossilíferes dels animals que les produïren, si bé és cert que s’han pogut atribuir amb més certesa un nombre més gran d’icnites que d’altres marques als animals que les generaren.
Altres icnofòssils són els copròlits, les egagròpiles i les restes d’ous. Els copròlits són els excrements fòssils de vertebrats, comunament de peixos, rèptils i, fins i tot, mamífers, que ofereixen la possibilitat de conèixer la dieta alimentària dels animals que els originaren pel fet de contenir elements o restes dels organismes que en formaven part, susceptibles de fossilitzar-se. També és el cas de les egagròpiles, producte característic dels ocells rapinyaires que, no podent digerir les plomes, els pèls i els ossos de les seves preses (altres ocells i petits mamífers), els expulsen en forma de boles que conserven l’esquelet, moltes vegades sencer. Entre els icnofòssils que evidencien activitat reproductora destaquen les postes, generalment de peixos, i els ous. Dels ous, se’n conserva la closca, bé que la majoria de vegades esclafada pel pes dels sediments, i generalment corresponen a rèptils, com el cas dels dinosaures de la conca de Tremp i, més rarament, a ocells.
Els principis i les lleis de la paleontologia
La investigació paleontològica es regeix clàssicament per un seguit de principis i de lleis, propis els uns i compartits amb la biologia els altres; la majoria de les lleis són empíriques i han sorgit del mateix estudi dels fòssils. Basades en diferents disciplines físico-químiques, biològiques i geològiques i a l’empar d’aquests principis i lleis, han sorgit diferents disciplines d’estudi dins del camp de la paleontologia, com la paleoecologia, la paleoetologia, la paleofisiologia (paleopatologia, paleoteratologia, paleoneurologia, paleopsicologia i paleobioquímica), la paleobiogeografia, la biostratigrafia i la paleontologia evolutiva.
El principi de l’actualisme biològic
L’actualisme paleontològic estableix que els organismes amb restes fossilitzades es regien per les mateixes lleis biològiques que els éssers vius actuals. Les necessitats fisiològiques eren similars i la seva organització anàloga i equivalent a l’actual. Així, doncs, associem les valves fòssils d’un lamel·libranqui a les petxines actuals, de la mateixa manera que unes dents fòssils les fem correspondre a un vertebrat. De la presència d’eriçons fòssils en uns sediments deduïm el seu caràcter marí, atesa l’exclusivitat dels representants actuals d’aquest grup d’organismes per al medi marí, i podem estendre aquesta consideració sobre les característiques del medi a la resta dels organismes fòssils que apareixen en el jaciment. Seguint el mateix principi, podem atribuir les closques d’ous fòssils a animals que presentaven un tipus de reproducció ovípara.
El principi de la no-generació espontània
L’actualisme implica la continuïtat del procés vital al llarg del temps geològic, sense interrupció, de tots els grups d’organismes des de la seva aparició fins a la seva extinció. Qualsevol ésser viu procedeix d’uns antecessors, i això ha estat així d’un manera continuada, i per això els organismes actuals són el resultat de generacions successivament anteriors, i cada vegada més antigues.
El principi de l’anatomia comparada
L’anatomia comparada entre animals i vegetals actuals permet de constatar les analogies al mateix temps que les característiques diferenciadores dins de cada grup o entre els elements dels quals es componen. Així, per exemple, les valves dels lamel·libranquis són suficients per a diferenciar aquests organismes entre ells i permeten, per tant, d’atribuirlos el nom de l’espècie a la qual pertanyen. Fins i tot en els vertebrats, restes fragmentàries d’elements esquelètics (per exemple, les dents) poden ser prou característiques per a diferenciar les espècies que es troben en un jaciment.
Més problemàtica pot semblar l’adscripció a un grup o una espècie fòssil quan els seus representants ho són d’un grup extingit, com és el cas dels arqueociats o dels trilobits. En aquests casos, cal recórrer a la comparació amb grups que ofereixen similituds o analogies, fins a arribar (quan és possible) a una interpretació correcta. Encara hi ha dubtes en grups com els graptòlits, els conodonts o els carpoïdeus; concretament, aquests darrers han estat considerats equinoderms, però encara resta oberta la possibilitat que es tracti de cordats primitius.
El principi de la correlació orgànica
Enunciat per Cuvier, estableix que gràcies a una part o a una estructura d’un fòssil, no tan sols podem arribar a conèixer-lo sinó que, a més, podem deduir quins altres elements el conformen o, d’entre altres elements, quins en formen part. Per exemple, diem que a un animal fòssil que posseeix urpes li correspon de tenir una dentició típica de carnívor, mentre que si té peülles li correspon una dentició de tipus herbívor.
El principi de la cronologia relativa
Estableix que l’ordre de superposició dels estrats correspon a l’ordre cronològic en què es dipositaren, és a dir, els més antics se situen sota els més moderns. En conseqüència, si no hi ha hagut cap inversió de les capes, els fòssils que apareixen en un estrat determinat són posteriors o més moderns que els que trobem en els estrats subjacents i anteriors o més antics que els dels estrats suprajacents. Aquest principi procedeix del camp de l’estratigrafia i fou enunciat pel geòleg danès Steno, al segle XVIII. És important per a poder establir seqüències d’organismes al llarg del temps, les quals aporten dades filogenètiques i d’ordre evolutiu, ja que per a valorar-les cal estudiar la seva ordenació cronològica.
Les lleis paleontològiques
Una primera llei estableix que la durada de les espècies en les èpoques geològiques ha estat limitada. Tant les espècies com els diferents grups taxonòmics de categoria més àmplia compleixen un cicle que inclou les fases d’aparició, desenvolupament, dispersió, predomini, declinament i extinció. Alguns organismes són anomenats pancrònics o fòssils vivents perquè han perdurat a través del temps amb variacions mínimes; en són exemples el celacant, el nàutil, el límul, els escorpins, etc. Però, si bé és cert que en aquests i en d’altres casos les variacions morfològiques han estat mínimes, no ho és menys que les espècies s’han anat modificant al llarg de la història de la vida a la Terra i que, per tant, han estat limitades com a tals a diferents àmbits de temps geològic. Aquesta llei és d’aplicació fonamental per a establir els fòssils característics de determinades zones o períodes, i també com a base per a establir les variacions de les espècies en el temps, és a dir, la seva evolució.
La segona llei estableix que les diferències entre les faunes fòssils i les actuals són més acusades com més antigues són les faunes fòssils. Així s’expressa una circumstància que s’acompleix en el conjunt d’una fauna o d’una flora fòssil; només en els organismes pancrònics es fa més difícil d’avaluar aquestes diferències.
Els animals de les faunes més recents tenen formes més variades que no les de les faunes més antigues. Aquesta tercera llei va lligada a l’anterior, i no es constata tan sols que les formes són més variades sinó que, a més, ha anat augmentant progressivament el nombre de tàxons gràcies a l’aparició de nous grups biològics que superen en nombre els extingits.
Una quarta llei estableix que els animals més perfectes (d’organització més complexa) són relativament més recents. El desenvolupament dels diferents grups biològics ha seguit un ordre creixent de complexitat anatòmica o d’efectivitat amb relació al seu mode de vida especial. La regressió que s’observa en certs òrgans i estructures i que implica una simplificació anatòmica no suposa cap excepció a aquesta llei, ja que els òrgans simplificats solen ser més eficients per a l’animal que els posseeix, o bé es tracta d’estructures anatòmiques que han perdut la seva funció primitiva i ja no són útils a l’animal; per exemple, la reducció del nombre de dits i el desenvolupament i la complicació de les molars en els èquids, la reducció parcial o total de la dentició en els ocells i els cetacis, etc.
La cinquena llei diu que des del moment de l’aparició d’un grup biològic fins a la seva extinció, no hi ha hagut interrupció en la seva existència. És una conseqüència del principi de l’actualisme, que proclama la continuïtat dels processos vitals. Si hi ha mancances en el registre fòssil de certs grups biològics, es deu al caràcter incomplet de la informació paleontològica existent, i a mesura que les investigacions es desenvolupen sorgeixen nous descobriments que omplen aquestes llacunes.
Per la sisena llei, cada espècie ha arribat a l’existència coincidint en l’espai i en el temps amb una altra de preexistent. Les espècies fòssils no es troben aïllades, sinó que les més properes es troben en les mateixes localitats o en localitats veïnes. La seqüència natural d’espècies per ordre d’afinitat ho és també temporalment i geogràficament, de manera que cada espècie ha tingut una antecessora o una altra de pròxima, de la qual s’originà. Aquesta llei neix de l’observació de les condicions d’aparició dels fòssils i de les seves associacions recíproques.
Segons la setena llei, l’ordre d’aparició dels representants d’un cert tipus d’animals es reprodueix en les fases per les quals passa el desenvolupament individual dels més perfectes i més recents del mateix grup. Aquesta llei, també anomenada llei de la recapitulació, permet deduir que l’ontogènia és una reproducció abreujada de la filogènia (llei biogenètica fonamental de Haeckel).
La distribució geogràfica dels grups biològics ha sofert canvis en el decurs del temps. En èpoques geològiques successives, els fòssils d’un grup biològic determinat es troben en àrees geogràficament diferents, la qual cosa suposa que ha variat la seva àrea inicial de dispersió per migració en determinades direccions. En tots els grups fòssils es pot comprovar aquest fet, fins i tot en molts organismes de tipus sedentari (bentònics, sèssils) que, gràcies al caràcter nectònic (nedador) o planctònic de les seves larves, han pogut colonitzar nous espais. Evidentment, aquestes migracions són més espectaculars en el cas dels vertebrats, per la mateixa facilitat de locomoció que els caracteritza.
La classificació paleontològica
Per a la classificació paleontològica, com per a la biològica, s’utilitzen la sistemàtica i la taxonomia. La sistemàtica pretén assignar a cada individu el nom (l’espècie) que li correspon i situar-lo dins d’un grup, mentre que la taxonomia ordena i agrupa les espècies.
En paleontologia s’utilitza una classificació morfològica, que d’uns quants anys ençà és ajudada per l’aplicació de criteris biomètrics (paleontologia quantitativa). Quan un paleontòleg descobreix un seguit d’elements faunístics o florístics fòssils i intenta descobrir les espècies que representen, ha d’agrupar els diferents individus segons les seves similituds morfològiques i formar uns grups que es diferencien els uns dels altres per un determinat conjunt de característiques peculiars. Les diferències entre aquests grups són molt evidents en alguns casos, mentre que en d’altres són menors i això comporta un risc de subjectivitat. Aquests grups es consideren inicialment com a espècies diferents, que després cal identificar per mitjà del reconeixement de les seves característiques en publicacions i monografies paleontològiques que tracten del grup o els grups representats de la mateixa edat que el descobert. Aquest procés d’identificació fa que alguns grups es mostrin idèntics a espècies ja descrites o bé mostrin només variacions menors. D’altres, en els quals no són visibles totes les característiques necessàries per a una determinació segura, o en la identificació de les quals hi ha un cert grau d’incertesa, es poden adscriure a una espècie afegint una abreviatura entre el nom genèric i l’específic o bé després del genèric: aff. (affinis, afí a), cf. (confer, semblant a), sp. inc. (espècie incerta), o sp. ind. (espècie indeterminada). També sol ser comuna l’anomenada nomenclatura oberta, per la qual s’escriu l’abreviatura sp. a continuació del nom genèric; el seu sentit és, doncs, que els exemplars responen a les característiques d’aquell gènere, però que hom no n’ha determinat l’espècie.
D’altres grups d’espècimens poden resultar nous, és a dir, desconeguts fins aquell moment. En aquest cas cal erigir-ne les espècies noves: cal figurar l’holotipus i els paratipus tot donant una descripció tan àmplia com sigui possible de la morfologia del fòssil; indicar la localitat tipus i l’edat de la sèrie tipus o jaciment i donar nom a l’espècie tot indicant la col·lecció o el museu que en conserva l’holotipus i la sèrie, amb els corresponents números de registre. També cal donar l’etimologia del nom i les sinonímies (noms que havia rebut anteriorment) i indicar les diferències respecte de les espècies més semblants o pròximes. Cada espècie nova s’ha d’atribuir a un gènere, i si aquest també s’ha de definir com a gènere nou, cal crear-lo amb les mateixes consideracions que si es tractés d’una espècie. Per a erigir una nova espècie a partir dels exemplars trobats poden seguir-se diferents camins: escollir una sèrie tipus o hipodigma (conjunt d’exemplars sobre els quals hom descriu l’espècie); o escollir un holotipus (exemplar de la sèrie que representa la mitjana dels individus que la formen). Quan s’escull un holotipus, la resta dels individus de la sèrie tipus s’anomenen paratipus i donen el ventall de característiques i variació que l’holotipus tot sol no dóna. Si no s’escull un holotipus, tots els individus de la sèrie s’anomenen sintipus. Si posteriorment es tria d’entre aquests un element de la mateixa significació que l’holotipus, rep el nom de lectotipus. Un neotipus és l’exemplar seleccionat dins la sèrie tipus (o no) quan l’holotipus o fins i tot la sèrie s’ha perdut o s’ha destruït. Un tipoide és un exemplar que s’afegeix a la sèrie tipus perquè presenta una determinada característica que es troba més ben desenvolupada. Finalment, els topotipus són tots els representants de l’espècie que provenen de la mateixa localitat que la sèrie tipus.
Sovint, el paleontòleg s’enfronta al fet de treballar amb restes fòssils que o bé no poden ser relacionades amb l’ésser viu al qual pertanyien, o bé, tot i saber a quin organisme corresponen, no són atribuïbles a una determinada espècie. La primera possibilitat es dóna, per exemple, en la majoria dels icnofòssils, i molt especialment en el cas de les pistes i de les marques ja que, si bé hom les pot classificar morfològicament, generalment no poden ésser adscrites amb certesa a un organisme concret. Dins la segona opció es troben, per exemple, les espícules d’esponges i d’alcionaris, les esclerites dels holoturoïdeus, molts apticus, opercles de gasteròpodes marins, otòlits, peus d’equinoïdeus, dents de selacis, megàspores, micròspores i grans de pol·len. També en aquest cas, hom fa servir la nomenclatura binomial, encara que les espècies i els gèneres no tenen el mateix sentit que en la nomenclatura biològica, sinó que representen un nom convencional, d’utilitat pràctica. D’aquesta manera, en lloc d’usar tàxons s’empren els anomenats paratàxons, que segueixen el criteri de classificació morfològica. Per a molts d’aquests paratàxons s’utilitzen terminacions en -ites i -ichnites; així, per exemple, les petjades fòssils reben el nom genèric d’icnites.