Els organismes del fitoplàncton
El fitoplàncton és format per organismes unicel·lulars autòtrofs i d’altres que, encara que siguin palesament heteròtrofs en tot o en part, deriven amb tota evidència de manera directa d’organismes autòtrofs. Són petits (generalment entre 2 i 200μm, rarament fins a més d’1 mm) i, en moltes espècies, les cèl·lules, tot i conservar la seva independència, van unides en cadenes o filaments, de vegades coberts de mucilag, que poden arribar a ser visibles a ull nu.
Tot i que la sistemàtica adoptada en aquesta obra és la que parteix de la divisió dels éssers vius en cinc regnes, proposada per Robert H. Whittaker el 1959, d’acord amb la qual la majoria dels organismes del fitoplàncton no són pas plantes —com permetia de suposar el seu nom—, sinó protoctists, la denominació tradicional de fitoplàncton per a designar els productors primaris planctònics manté encara tota la seva vigència, malgrat la incorrecció taxonòmica que implica el seu sentit etimològic de “plàncton vegetal”. El mateix seria aplicable al zooplàncton o “plàncton animal”, també amb una important participació de protoctists.
Els cianobacteris i els protocloròfits
Els elements més petits del plàncton fotosintetitzador són els cianòfits, cianofícies o cianobacteris, representats principalment per organismes procariotes de formes arrodonides (Synechocystis) o allargades (Synechococcus); fan aproximadament 1,5 μm de diàmetre o encara menys. Aquests organismes són esclaus de la viscositat de l’aigua i els és difícil la renovació de la capa hídrica de pocs micròmetres que els envolta amb tenacitat. Si es busquen, es troben gairebé arreu en concentracions que poden arribar a atènyer fins a centenars de milers de cèl·lules per mil·lilitre, encara que normalment les concentracions són molt més baixes. Quan hom va començar a fixar-s’hi, se’ls atribuí una fracció exagerada de la producció primària; posada la qüestió en els seus justos termes, hi poden contribuir en una proporció d’entre el 20 i el 40%, ben sovint menys, segons sigui localment la seva abundància. Per la seva naturalesa i persistència, es troben, més o menys inactius, en un gran gruix de l’aigua, de manera que gairebé a tot arreu n’hi ha, sempre en condicions d’assimilar així que l’aigua arriba a nivells il·luminats. Aquests organismes poden ser ingerits, juntament amb altres bacteris, per petits flagel·lats heteròtrofs, relativament abundants i molt diversificats al plàncton.
Els cianobacteris tenen els pigments en tilacoides estesos en la perifèria de les cèl·lules, no en cromatòfors discontinus, cosa que dóna a les cèl·lules una coloració difusa. Contenen clorofil·la a i diferents ficobilines i carotinoides. Les ficobilines els donen l’exclusiva, entre els components del fitoplàncton, en la utilització de la radiació en la faixa intermèdia de la llum verdosa. A més dels cianobacteris, en el plàncton fotosintetitzador hi ha altres organismes procariotes d’aspecte semblant, perquè també són extremament menuts (entre 0,6 i 0,8 µm), però dotats de clorofil·la b. Són els protocloròfits, que també estan força estesos als oceans. La seva organització ha estat comparada a la d’un plastidi envoltat per una paret cel·lular.
A les aigües dolces, els cianobacteris de mida cel·lular més gran —i per tant amb una organització que s’allunya encara més de la dels altres procariotes— són comuns i diversificats, però no passa així a la mar, tret del cas de les parts més dessalades de la mar Bàltica. Una de les poques excepcions és el gènere Trichodesmium, que fa llargs filaments units en feixos i que és especialment freqüent en aigües tropicals estratificades, sovint en la part externa o lateral de les zones d’aflorament. S’ha dit que aquest organisme pot fixar nitrogen atmosfèric, com fa gairebé la generalitat dels cianobacteris, però no hi ha acord pel que fa al cas. Òbviament, l’esquifida presència d’aquests cianobacteris a les aigües marines lliures deu constituir una prova que el nitrogen no és pas un element limitant major en els oceans. Ben al contrari, en les aigües dolces sotmeses a processos de desnitrificació i enriquides amb fòsfor —canvis avui deguts majorment a l’acció humana— els cianòfits es multipliquen amb ufana.
Cal notar que els feixos de Trichodesmium, visibles a ull nu, suporten altres organismes, com ara hidraris i fins i tot fongs, cosa que els hauria de donar el caràcter d’una mena de líquens pelàgics. Els seus flocs es desfan ràpidament quan es remena l’aigua i, en destriar-se uns filaments dels altres, el seu conjunt escapa a la visibilitat. Richellia forma filaments curts, que generalment porten en un extrem un heterocist (cèl·lula diferenciada, assimiladora de nitrogen) i acostumen a trobar-se com a simbionts interns dins de les cèl·lules de diatomees d’un cert volum.
Els dinòfits
Els dinòfits, pirròfits, dinofícies, dinoflagel·lats o dinomastigotes (el “Handbook of Protoctista”, editat el 1989 per L. Margulis, J. O. Corlyss, M. Melkonian i D. J. Chapman, reconeix els criptòfits com un phylum o tipus independent de protoctists) són eucariotes, però amb un nucli peculiar, que es pot interpretar com a particularment primitiu, amb cromosomes sense histones, que s’acostumen a veure molt bé durant la interfase de les divisions i donen llavors al nucli un aspecte que recorda un cabdell de fil, caràcter que pot resultar útil per a confirmar la naturalesa de cèl·lules dubtoses. En diferents espècies es reconeixen graus diversos de poliploïdia que es poden manifestar en un polimorfisme de mida, amb una freqüència aparentment més gran de les cèl·lules de més grandària durant l’estació freda.
L’organització dels dinòfits és molt variada: són un grup amb una ingent capacitat evolutiva, probablement els dominadors més antics del plàncton marí. En canvi no han arribat a donar una organització pròpiament pluricel·lular. Serien, com si diguéssim, una potencial línia filètica avortada, probablement perquè altres formes d’organització —els eucariotes genuïns— li passaren al davant. Les poques formes que s’han aproximat a una organització pluricel·lular són paràsites d’animals planctònics, com copèpodes i apendiculàries.
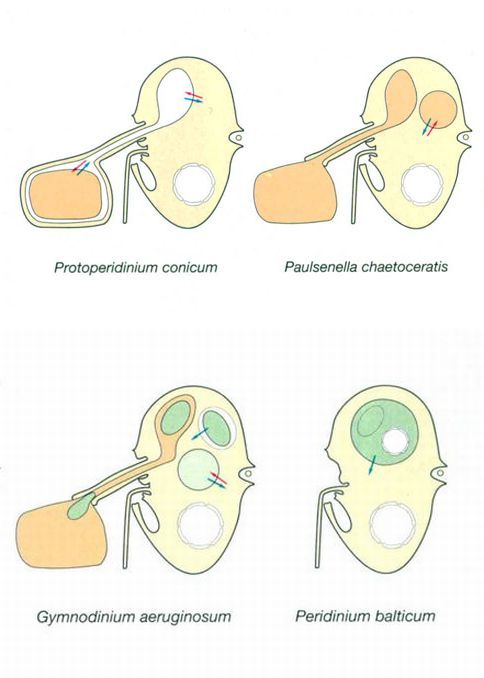
Jordi Corbera, a partir de fonts diverses
No tots els dinòfits són organismes fotosintetitzadors involucrats en la producció primària de les aigües on viuen: molts, tant marins com d’aigües dolces, tenen diferents formes de vida heterotròfica, més variades i complexes en el medi marí. Alguns, com Protoperidinium conicum, poden estendre i retreure una mena de pseudopodi en forma de vel (pallium) amb el qual envolten altres cèl·lules, principalment diatomees, que digereixen extracel·lularment. Altres, com Paulsenella chaetoceratis, emeten pseudopodis xucladors que penetren al citoplasma de les seves preses i el xuclen per digerir-lo intracel·lularment (mizocitosi). El mateix fa, encara que sigui d’aigua dolça, Gymnodinium aeruginosum, però en aquest cas utilitza el pseudopodi per adquirir cloroplasts de la seva víctima, que conserva vius i funcionals mentre digereix la resta del contingut del citoplasma. Un cas ben diferent és el de Peridinium balticum, que allotja un petit simbiont intracel·lular fotosintetitzador del qual obté l’aliment que altrament no podria sintetitzar. També hi ha dinòfits que es limiten a fagocitar altres organismes i alguns que funcionen com a paràsits intracel·lulars que s’alimenten del citoplasma aliè que els envolta.
Actualment mostren, en conjunt, un espectre funcional molt ampli, més diversificat a les aigües marines que no pas a les dolces: des d’organismes autònoms fotosintetitzadors, normalment flagellats, fins a cèl·lules força grosses (0,5 mm) que es comporten essencialment com a animals. Les del gènere Noctiluca, efectivament, ingereixen fins animals “superiors” com apendiculàries o ous de seitó. Els dinòfits de característiques més primitives —i generalitzades— tenen cromatòfors amb clorofil·les a i c i carotenoides (peridina i altres d’especials). El seu color és generalment verd groguenc o brunenc, però en proliferacions massives, algunes espècies es veuen rogenques (“marees roges”).
Les cèl·lules són sempre de forma asimètrica. La conformació més generalitzada en les famílies més nombroses inclou un solc equatorial d’extrems desviats (helicoide destre o sinistre) que conflueix o s’interseca amb un altre solc longitudinal. A l’àrea on els dos solcs s’encreuen, s’implanten dos flagels més o menys diferenciats. Es tracta d’una regió peculiar, en la qual es poden formar pseudòpodes de citoplasma clar, linears o bé en forma de vels extensos que permeten una digestió extracellular. També hi desemboquen cavitats intracel·lulars (púsules). S’han descrit així mateix, de diferents dinoflagel·lats, altres diferenciacions molt avançades com, per exemple, ocels capaços de percebre la llum i tentacles de diversa maniobrabilitat. També és freqüent la producció de llum, sovint en orgànuls diferenciats (escintil·lons).
Hi ha diversos dinòfits, dels gèneres Prorocentrum, Dinophysis, Gonyaulax, Alexandrium, Pyrodinium, tots ells planctònics, i encara d’altres, com els Gambierdiscus i algunes formes de Prorocentrum que viuen sedentaris, fixats sobre Sargassum, sobre altres algues macroscòpiques marines i fins i tot sobre granets de sorra, que produeixen tòxics específics molt potents, de composició química força diversa, amb efectes negatius sobre el funcionalisme d’alguns animals. Són compostos d’una gran varietat (saxitoxines i altres), molècules amb anelles que contenen nitrogen, de pesos moleculars que rarament baixen dels 1 000 daltons. Aquestes substàncies poden arribar fins a l’organisme humà a través de cadenes tròfiques de les quals formen part, generalment, lamel·libranquis (intoxicacions com a conseqüència de la ingestió de musclos o de cloïsses) o peixos (“ciguatera” en alguns països tropicals). Altres encara (Cochlodinium) generen a l’interior de les seves cèl·lules uns bastonets (tricocists) de diverses menes que poden projectar-se a l’exterior i produeixen filaments enganxosos, dels quals se suposa que també poden ser tòxics, que s’enganxen als filaments branquials dels peixos i en perjudiquen el funcionament.
La superfície de les cèl·lules dels dinòfits té una coberta de conformació variable, que va des d’una làmina simple fins a una estructura complexa formada per diverses plaques distintes i unides en sutures sovint amb moltes complicacions. De vegades la coberta inclou una regió preformada per la qual la cèl·lula, després de prendre una forma esferoïdal i envoltada d’un nou embolcall relativament prim, pot abandonar la vella coberta, talment un pilot d’un caça que es projecta fora del seu avió. La cèl·lula pot passar a una forma quiescent i aleshores el revestiment es fa de més durada, acumulant materials que s’assemblen a l’esporopol·lenina de les parets dels grans de pol·len i de les de moltes espores vegetals, si més no per la seva resistència, la qual permet que es conservi molt de temps. Altres dinòfits es calcifiquen a la seva perifèria.
Hi ha un bon registre fòssil de dinòfits, que és testimoni de l’antiguitat del grup; restes de dinòfits com els actuals es coneixen del Silurià, i formes pròximes, com alguns acritarcs, són encara molt més antigues, del Cambrià, si no d’abans. La seva edat també és testimoniada per la seva antiquíssima adopció, com a simbionts, pels coralls i altres animals marins des dels seus orígens evolutius. Simbionts, d’altra banda, que són sorprenentment uniformes i que generalment reben el nom de Symbiodinium pseudoadriaticum. L’estudi acurat de simbionts procedents de diferents hostes mostra que les diferències entre unes i altres soques són mínimes.
En el plàncton marí actual, els dinòfits o dinoflagel·lats es retroben arreu, representats per organismes que generalment fan entre 10 i 200 μm, i de formes de nutrició molt variades. Hi ha més formes heteròtrofes del que es podria creure, amb alimentació interna (ingestió) o externa (el vel de molts Protoperidinium). Poden contenir o transportar simbionts interns o semiexterns (com els cianobacteris d’Histioneis). L’assimilació força avançada d’alguns simbionts per part de dinòfits ha produït veritables quimeres o organismes mosaic. Els dinòfits de nutrició heterotròfica són particularment diversificats i abundants als nivells anomenats crepusculars, per sota del màxim de clorofil·la, en aigües estratificades.
El flagel transvers faria rodar la cèl·lula al voltant del seu eix àntero-posterior; etimològicament, el prefix dino-, de dinòfit, deriva del verb grec “dino” (δινῶ), que significa girar com una baldufa, i no pas de “deinos” (δεινόϛ), que significa terrible, com en dinosaure. L’esmentat flagel transvers és normalment el més robust, i a la seva acció s’oposa sovint la forma aplanada o irregular de la cèl·lula, que acostuma a ser d’una gran riquesa de detall (Ceratium, Dinophysis, Ornithocercus, etc.), sempre dins d’una asimetria peculiar. La conseqüència d’això és una intensificació de la fluència de l’aigua sobre la cèl·lula, cosa que augmenta l’absorció; al mateix temps, el moviment del flagel longitudinal permet mantenir el nivell apropiat d’il·luminació o de concentració de nodriment dins de l’aigua i fins i tot regular moderades migracions verticals.
Els criptòfits
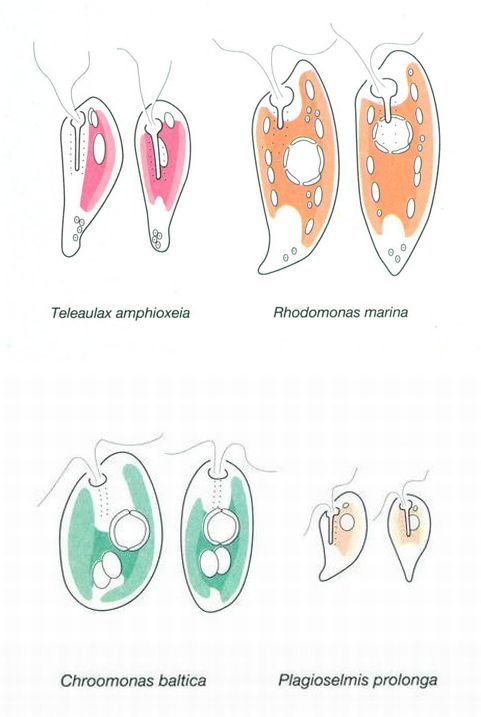
Representacions esquemàtiques d’alguns criptòfits. Tots són petits, sobretot Plagioselmis prolonga, i tots ells de colors diferents, com correspon a la diversitat de pigments que poden posseir. Cada una de les figures aparellades són la mateixa cèl·lula vista des d’un angle diferent.
Jordi Corbera, a partir de dades de Hill
Els criptòfits són organismes que, en una seqüència de dimensions i complicació cel·lular creixent, mostren, com a característiques comunes, la mida petita (ordinàriament entre 5 i 20 μm, menors a la mar que a les aigües dolces), i les cèl·lules asimètriques més o menys allargades, sovint afuades per l’extrem posterior. Tenen dos flagels que surten d’una mena de “faringe”, amb la qual van associats bastonets que es poden disparar o projectar a l’exterior com els tricocists dels dinòfits. Posseeixen també cromatòfors amb pigments complexos i, per tant, de colors variats: verdosos, rovellats, i fins i tot blavissos i rosats. Hi ha formes heterotròfiques, algunes de parentiu dubtós com Leucocryptos marina, un organisme incolor força freqüent; altres s’alimenten de bacteris. S’ha provat que molts criptòfits, potser tots, són veritables quimeres o organismes complexos, amb assimilació organísmica d’antics simbionts. El “Handbook of Protoctista”, editat el 1989 per L. Margulis, J. O. Corlyss, M. Melkonian i D. J. Chapman, reconeix els criptòfits com un phylum o tipus independent de protoctists.
Abans s’esmentaven molt poc en el plàncton marí però, en realitat, hi són freqüents. Acostumen a presentar-se formant núvols discontinus en aigües superficials, sovint després d’una barreja local intensa. Mereixen ser destacats els gèneres Cryptomonas, Rhodomonas i Hemiselmis.
Els crisòfits o cromòfits
El “Handbook of Protoctista”, editat el 1989 per L. Margulis, J. O. Corlyss, M. Melkonian i D. J. Chapman, fa dels haptòfits o primmesiòfits i de les diatomees o bacil·lariòfits dos tipus independents de protoctists, mentre que inclou en els crisòfits (que tracta igualment com a phylum independent) els silicoflagel·lats com a classe, amb el nom de dictiofícies. Aquí, més que aplicar-lo a un grup taxonòmic determinat, emprem aquí aquest terme col·lectiu com a inclusiu, entre altres de menys importants, dels grups dels haptòfits —que inclouen els coccolitoforals—, les diatomees i els silicoflagel·lats, els quals hauran de reestructurar-se taxonòmicament quan es coneguin millor i s’hagi arribat a un consens sobre les afinitats entre petits flagel·lats molt diversos, cada vegada més ben estudiats mercès a la microscòpia electrònica*. En general es tracta d’organismes amb cromatòfors groguencs (clorofil·les a i c i fucoxantina com a pigments bàsics) que mai no acumulen midó com a reserva, amb flagels diferenciats (quan existeixen), de plasma clar i amb la cèl·lula coberta de petites escates riques en detalls estructurals, de vegades mineralitzades, que es poden estendre en grau divers sobre els flagels. El seu estudi exigeix l’ús de la microscòpia electrònica, i per motius tècnics la informació segura sobre les seves distribucions per força ha de ser relativament recent.
En les diatomees, que s’inclouen en aquest col·lectiu o gran grup i són de mida relativament grossa, la silicificació de la coberta és total. Aquesta coberta consisteix en dues teques (estructures que es coneixen des de fa més de dos segles) que acostumen a encaixar l’una en l’altra com una capsa amb la seva tapa. La mida més gran de les cèl·lules, el caràcter de les valves, l’absència de formes flagel·lades, tret del cas d’algunes espècies heterogamètiques, i el caràcter diploide de les cèl·lules vegetatives han fet que les diatomees s’hagin pogut considerar com a equivalents als zigots d’altres crisòfits. També són relativament més recents; la seva expansió més gran s’escaigué al començament del Terciari, i es podria associar amb el desenvolupament de regions marines amb afloraments intensos. Les diatomees són, doncs, en una paraula, postdinosaurianes.
Els haptòfits o primmesiòfits

Chrysochromulina és un gènere de primmesiòfits amb reputació d’“alga assassina” encara que no es coneix de quina manera ocasiona les mortaldats de peixos que se li atribueixen. Als mesos de maig i juny del 1988 una sobtada proliferació de C. polylepis afectà una extensa àrea dels estrets danesos i øresund i de les costes del S de Noruega i causà una gran alarma i una certa mortaldat de peixos, deguda més a l’obstrucció de les brànquies pel mucilag o a exhauriment local de l’oxigen en alguna badia per la descomposició de la massa d’algues i mucilag encallats, que a cap toxina identificable. El dibuix (a l’esquerra) mostra una altra espècie de Chrysochromulina, una mica més gran que C. polylepis, la qual, de tant en tant, genera taques semblants a la Mediterrània occidental.
Ramon Margalef i Editrònica
Els haptòfits o primmesiòfits són un grup de crisòfits de cèl·lules més aviat petites (10-25 μm) dotades normalment de dos flagels funcionals com a tals i d’un tercer filament que sovint es veu entortolligat en espiral i que no funciona com a flagel ni en té l’estructura, ja que no té els dos túbuls centrals que es troben en els flagels normals a més dels nou parells perifèrics. El nom d’haptonema que se li donà suggereix que podria servir per a fixar la cèl·lula a un substrat, però el cert és que en alguns primmesiòfits (Chrysochromulina) serveix per a atreure, aconduir adequadament els bacteris dels quals s’alimenta, i facilitar-ne la ingestió.
Phaeocystis pot donar cèl·lules flagel·líferes, però la forma més comuna és la de cèl·lules esfèriques, molt petites (2-4 μm) i sense flagels, immerses en una matriu mucilaginosa que arriba a formar masses importants. Obstrueixen les malles de les xarxes de plàncton i fins i tot les brànquies dels peixos. Des de fa molt de temps els pescadors sabien que, a la mar del Nord, els peixos defugen les aigües en què abunda Phaeocystis. Encara que es troba també regularment a la Mediterrània, les proliferacions massives d’aquestes algues es donen amb més freqüència o són més importants a les mars fredes, incloent-hi les antàrtiques, on la seva abundància s’havia relacionat amb l’escassetat de bacteris trobats en el tub digestiu d’animals als quals arriba material sintetitzat per l’alga. Tant Phaeocystis com algunes espècies de Chrysochromulina s’han considerat com a tòxiques i es produí una considerable alarma a l’estiu del 1988 —i els successius— a la mar del Nord, pel fet que alguna espècie de Chrysochromulina es presentà en concentracions que localment arribaren a les 80 000 cèl·lules per mil·lilitre. Hi ha moltes espècies d’aquest gènere, des de les que fan pocs µm de llargada i tenen l’organització reduïda al mínim, fins a les de 13 μm o més de diàmetre.
El grup més important d’haptòfits planctònics i autòtrofs, que són bons productors primaris, és sens dubte el de les coccolitoforals, de cèl·lules cobertes de plaquetes de carbonat càlcic o coccòlits. En la major part de les seves cèl·lules no es veuen flagels funcionals. Hi ha casos de polimorfisme, o d’alternança entre formes d’organització diferent o, si més no, amb plaquetes diferents. Les coccolitoforals de plaques més extraordinàriament desenvolupades (Scyphosphaera, que té unes peces calcàries que semblen talment vasos de cristall tallat, Rhabdosphaera, Discosphaera, etc.) semblen aparèixer cap a la fi de les successions ecològiques, quan les cèl·lules es divideixen més a poc a poc i els vaga d’anar dipositant carbonat càlcic i potser algun altre element químic associat. Per contra, les espècies que es multipliquen a velocitat fantàstica, com ara Emiliania huxleyi, que és segurament l’organisme més nombrós en el plàncton marí, i potser arreu del món, tenen les plaquetes més primes (particularment primes i més fàcilment disgregables quan la velocitat de multiplicació cel·lular és màxima) i solen fer uns pocs micròmetres, de manera que rarament es poden estudiar i identificar correctament sota el microscopi ordinari; entre els coccòlits que s’acumulen en el sediment de la Mediterrània occidental, més del 80% pertanyen a E. huxleyi. Les coccolitoforals es multipliquen amb profusió en condicions apropiades, sovint properes a fronts. S’ha dit que certa terbolesa de l’aigua observable des d’avions, per exemple en els fronts de marea a l’W del canal de la Mànega, es deu a proliferacions de coccolitoforals. La presència de plaquetes de carbonat càlcic deu augmentar la densitat de les cèl·lules, i en pot afavorir la sedimentació en aigua ascendent. D’altra banda, contenen una fracció de carboni que es retira de la circulació quan les plaquetes o coccòlits se sedimenten i alleugereixen així una mica l’efecte hivernacle. Les espècies de coccolitoforals són molt antigues i de gran dispersió, i han anat canviant regularment al llarg dels temps geològics, de manera que són fòssils molt adients per a l’estudi de la distribució i la diversificació dels tipus de plàncton al llarg del temps.
Els bacil·lariòfits o diatomees
L’altre grup significatiu de crisòfits és el dels bacil·lariòfits o diatomees. Aquests organismes, d’ençà que es coneixen, sempre han cridat l’atenció per la seva coberta silicificada, que sovint es conserva en sediments antics. La seva estructura, sorprenentment rica i delicada, era apreciable amb els instruments òptics d’ús comú des del segle XIX, i convertí l’estudi de les diatomees en tasca d’afeccionats, que començaven per bullir les diatomees en àcid per deixar netes les cobertes. Per això la taxonomia del grup ha estat treballada meticulosament i, en alguns casos, fins a un punt excessiu en el seu detall, amb moltes sinonímies, com esdevé amb tots els organismes que tenen la fortuna o la dissort de tenir parts dures, com per exemple els mol·luscs. La microscòpia electrònica ha ajudat a resoldre moltes estructures petites, algunes d’elles força importants per a entendre particularitats de la vida de les diatomees. Perquè la seva coberta no és inert, i el conjunt de porus i cambres d’aquesta té certament una funció significativa en la nutrició; sovint canalitza la producció de cordons de mucilag que uneixen unes cèl·lules amb unes altres o que s’inflen i formen grans masses mucilaginoses (Chaetoceros socialis, Thalassiosira). La matriu orgànica de la coberta cel·lular diposita àcid silícic hidratat (SiO2 + nH2O), o òpal, polimeritzat, en forma d’una xarxa molecular contínua. Material orgànic, si més no en forma d’una pel·lícula externa, ocupa les valències lliures i n’evita la dissolució en aigües alcalines insaturades de silici. Segons les espècies i les condicions de vida, la part silícia és més o menys gruixuda. Cada població hi fa de més o de menys i el silici rarament és un factor limitant seriós. En el plàncton marí, especialment si les diatomees creixen ràpidament o si l’aigua conté una concentració baixa de silici (com s’escau, per exemple, a la Mediterrània, on l’alcalinitat és lleugerament superior a la mitjana oceànica i la concentració de calci també és més alta), sovint les cèl·lules són poc silicificades i es veuen flexibles. De fet, a la Mediterrània, només arriben a conservar-se en el sediment cobertes d’aquelles formes particularment gruixudes, que moltes vegades vénen del litoral, com la generalment difosa Paralia sulcata o les formes de resistència (hipnòcits) de Chaetoceros i d’alguns altres gèneres comuns. També arriben sovint a la mar valves de diatomees d’aigua dolça.
Les diatomees són organismes generalment grossos, de grandària que pot arribar a ser mil·limètrica. Es classifiquen en dos grups principals: les centrals, amb valves de simetria radiada, o circulars, de reproducció oògama, i les pennades, de valves allargades, en la reproducció de les quals no apareixen formes flagel·líferes, sinó que se solen formar gametangis que copulen sencers. Hi ha, a més, Phaeodactylum, un curiós organisme univalvar, comú en bassiols de les costes rocoses i que es fa bé en aquaris, però que només accidentalment s’ha reconegut en aigües lliures. La gran majoria de les diatomees del plàncton marí són centrals, principalment dels gèneres Thalassiosira, Coscinodiscus, Chaetoceros o Rhizosolenia (gènere avui subdividit). Les pennals acostumen a ser més costaneres, com ara Asterionella, Thalassionema o Thalassiothrix.
Les diatomees s’havien vist sempre com l’equivalent marí de les gramínies, productors inofensius i generosos, amb l’únic vici de dipositar silici. Recentment s’ha trobat que algunes espècies de Nitzschia produeixen tòxics potents, com és l’àcid domoic, un aminoàcid que, entre altres efectes, té el de ser causa d’amnèsia en els humans. Les cèl·lules d’algunes diatomees voluminoses, principalment del gènere Rhizosolenia, que són vacuolades i que suren durant l’estiu en aigua pobra de nutrients, allotgen cianobacteris fixadors de nitrogen (Richellia intracellularis). Altres diatomees s’associen amb ciliats que ajuden a mantenir-les en suspensió. Sovint es troben cadenes de cèl·lules buides de diatomees del gènere Dactyliosolen que porten tot a l’entorn cèl·lules flagel·lades; aquestes han estat descrites amb el nom de Solenicola, però se sospita que podrien pertànyer al cicle de la mateixa diatomea.
Els dictiocòfits o silicoflagel·lats
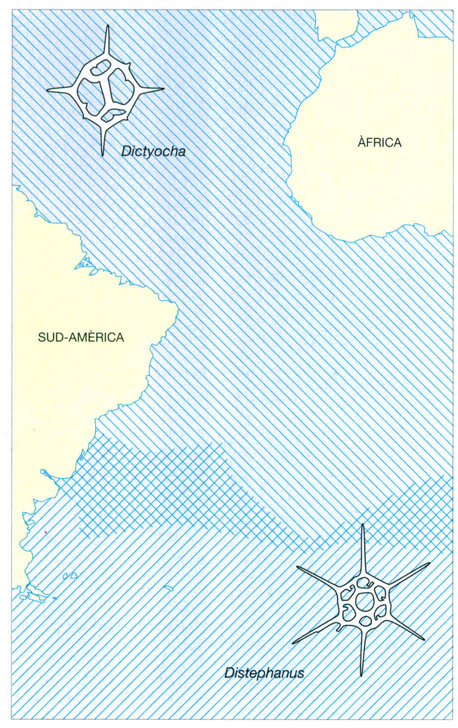
Alguns silicoflagel·lats només poden viure en uns intervals de temperatura molt definits. Hi ha una gran correspondència entre la distribució dels dos grans grups de silicoflagel·lats (els de quatre cares, atribuïts al gènere Dictyocha, i els de sis cares, atribuïts al gènere Distephanus) i la temperatura de l’aigua, fins al punt que es pot dir que els del tipus Distephanus es troben generalment en aigües de temperatura inferior a 15°C, mentre que els del tipus Dictyocha no poden viure per sota els 10°C. La transició entre ambdues formes pot ser sobtada, com passa a la mar del Japó, o poden coexistir en zones d’aigües de temperatures compreses entre els seus respectius límits, com passa a l’Atlàntic sud-occidental.
Jordi Corbera, a partir de dades de Lipps
Els dictiocòfits són protoctists flagel·lats de mida mitjana (20-70 μm), amb membrana feble, cromatòfors nombrosos i un esquelet intern reticulat molt característic, generalment amb simetria quadrangular o hexagonal, fet d’estructures tubulars de sílice (òpal), que són més compactes i es conserven millor que les cobertes de les diatomees. Abans de la divisió, es forma dins de la cèl·lula un segon esquelet amb simetria especular respecte al ja existent, llevat del cas d’algunes poblacions en moments de ràpid creixement que perden l’esquelet. Se’n coneixen restes fòssils des del final del Mesozoic ençà, i la seva expansió ha estat paral·lela a la de les diatomees, però, a diferència d’aquelles, es manifesten com un grup morfològicament molt conservatiu i pobre d’espècies. D’entre les més comunes, l’espècie de forma quadrangular Dictyocha fibula sempre s’ha comportat com una mica més termòfila que l’hexagonal Distephanus (= Dictyocha) speculum.
Els prasinòfits
Les algues verdes pròpiament dites (cloròfits), amb predomini del color verd donat per les clorofil·les i amb presència de midó, són d’aigua dolça. En el plàncton marí es troben els prasinòfits, un grup germà, ecològicament paral·lel però diferent dels cloròfits per característiques citològiques*. Són freqüents organismes amb quatre flagels iguals (Pyramimonas, Tetraselmis) i sovint amb escatetes superficials, esteses moltes vegades fins i tot damunt dels flagels. Les espècies del gènere Halosphaera són cèl·lules esfèriques bastant grans (40-100 μm de diàmetre) que donen petites cèl·lules propagatives multiflagel·lades.
El “Handbook of Protoctista”, editat el 1989 per L. Margulis, J. O. Corlyss, M. Melkonian i D. J. Chapman, inclou en el tipus dels cloròfits la classe de les prasinofícies (=prasinòfits) i la considera com l’origen filogenètic de la resta de classes dels cloròfits.
Els euglenòfits i els rafidòfits
Els euglenòfits són flagel·lats d’organització complexa, amb una coberta feta de làmines elàstiques ajustades lateralment que permet contínues deformacions de la cèl·lula. Tenen generalment dos flagels o, en tot cas, almenys un de funcional quan el segon ha estat reduït secundàriament. Tenen pigments verds i glúcids de reserva (paramil). Són freqüents en el plàncton marí d’aigües eutròfiques, com el que es troba en els ports. Una espècie característica és Eutreptiella gymnastica. El “Handbook of Protoctista”, editat el 1989 per L. Margulis, J. O. Corlyss, M. Melkonian i D. J. Chapman, reconeix els euglènids (=euglenòfits) com un phylum o tipus independent de protoctists.
Un altre phylum de protoctists que viu en condicions semblants a les dels euglenòfits és el petit grup dels rafidòfits*** (l’obra citada més amunt reconeix els rafidòfits com un phylum o tipus independent de protoctists, si bé alguns autors prefereixen tractar-los com una classe dins dels cloròfits), representat principalment per les espècies del gènere Chattonella, al qual pertanyen probablement també algunes espècies descrites amb els noms genèrics d’Olisthodiscus i Hornellia.
Els simbionts d’organització simplificada
Cèl·lules senceres que es poden reconèixer com a pertanyents a diferents grups dels descrits (criptòfits, per exemple) i d’altres amb diferents graus de simplificació (reduïdes sovint a simples cromatòfors) es troben en qualitat de simbionts en ciliats (Mesodinium) i en altres organismes.
Cèl·lules no tan disminuïdes o modificades, ben identificables encara pel que fa a les seves afinitats, viuen a l’interior de foraminífers, acantaris i radiolaris; la seva contribució a la productivitat primària del plàncton pot ser important, i molts dels organismes que les hostatgen han arribat a obtenir així una independència tròfica comparable a la dels coralls. Diversos organismes dels grups dels criptòfits i dels dinòfits són veritables quimeres, integrades per components associats, però d’origen genètic molt diferent.
La distribució i la diversitat del fitoplàncton
La densitat de les poblacions de fitoplàncton (microplàncton i nanoplàncton) en la zona fòtica sol estar compresa entre 10 i 1 000 cèl·lules per mil·lilitre. Una fracció considerable d’aquestes cèl·lules correspon a organismes que són parcialment o totalment heterotròfics. El nombre de cèl·lules del picoplàncton (cianòfits i protocloròfits) pot ser entre 10 i 1 000 vegades més gran (104-106 per mil·lilitre), però es tracta d’organismes molt petits, cada un dels quals fa entorn d’una mil·lèsima, només, del volum mitjà de les cèl·lules del nanoplàncton. Les poblacions en aigües profundes són més esparses, però mai no es pot dir fins on arriben. Sempre es troben en fondària, fora de la llum, cèl·lules d’espècies variades.
Les menudes cèl·lules del picoplàncton o ultraplàncton rarament reben nom específic, com a molt un nom genèric, encara que se sap que n’hi ha formes diverses, ja que no totes les poblacions es comporten, en cultiu, de manera igual. Els organismes més grossos fan de més bon descriure, i el seu estudi començà i progressà en una època en la qual la taxonomia es valorava relativament més que no pas actualment. Hi ha obres clàssiques, publicades entre el final del segle passat i mitjan segle actual, amb belles il·lustracions i monografies força acurades. Moltes espècies no han estat mai descrites, com resulta evident a tot recercador que tracti d’aprofundir en el coneixement del fitoplàncton d’una localitat qualsevol. Molts grups, com els criptòfits i diversos grups de petits flagel·lats, incloent-hi alguns que tenen parts dures, com els coccolitoforals, són particularment mal coneguts, com també moltes formes heterotròfiques de dinòfits, nombroses i freqüents, especialment en els nivells de penombra.
El plàncton amb la diversitat més alta del món
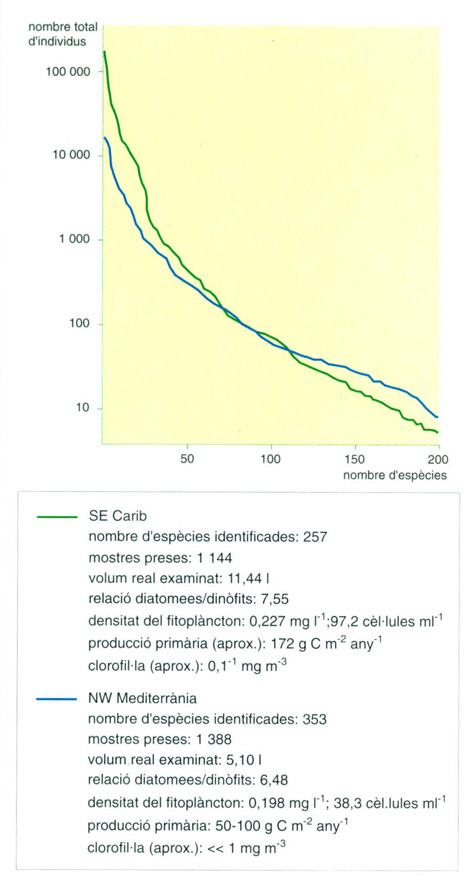
Els mateixos conjunts de mostres, on s'han ordenat les espècies del fitoplàncton per ordre decreixent de numerositat total.
Editrònica, a partir de dades de l’autor
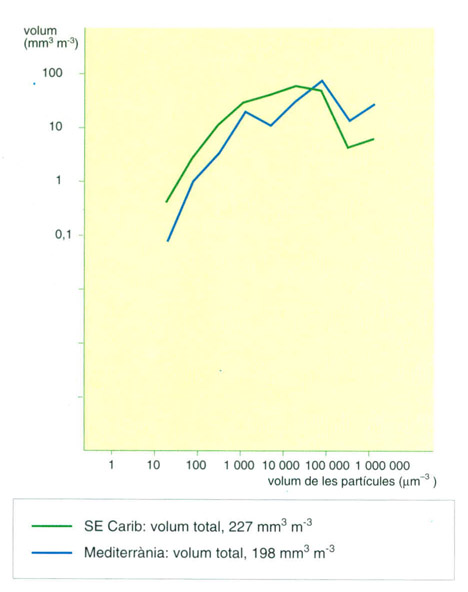
Comparació de la distribució dels dos conjunts de mostres, de la Mediterrània i del mar Carib, en grups caracteritzats pel volum de les cèl·lules. Al Carib, les cèl·lules petites predominen i les més grosses tenen una representació relativament menor.
Editrònica, a partir de dades de l’autor
Mediterrània i Carib: són les mars on la diversitat del plàncton és de les més altes del món, probablement només superada per alguns indrets de l’oceà Pacífic.
Els gràfics adjunts mostren dues comparacions entre mostres de plàncton de dues àrees, una a la Mediterrània nord-occidental i l’altra al SE del mar Carib (1963 i 1965). Solament es prenen en consideració les 200 espècies més abundants en cada col·lectiu. L’abundància és presa en escala logarítmica (esquerra). Els valors trobats són molt semblants en tots dos casos, amb lleuger avantatge per a la Mediterrània.
Baixa diversitat, àmplia distribució
Alain Sournia, amb alguns col·laboradors, publicà el 1991 un cens de les espècies conegudes en el fitoplàncton marí, que donà una xifra molt baixa, inferior a les 5 000 espècies, i molt inferior al nombre d’espècies conegudes en el fitoplàncton d’aigua dolça. Sorprèn aquesta pobresa relativa, si es pensa en la riquesa d’espècies que es troba en qualsevol família d’insectes. El baix grau de diferenciació específica en els oceans podria ser una conseqüència de la barreja intensa de l’aigua, que s’oposa a l’isolament genètic. Es pot entendre que els llacs hagin estat més efectius a l’hora de confinar les poblacions i d’estimular l’especiació. No obstant això, la “flora” planctònica marina ha anat canviant, més aviat ràpidament, en el decurs dels temps geològics i no és pas que tingui com a característica peculiar un baix grau congènit d’especiació.
En conclusió, la diversitat biològica pot ser inferior a la que seria esperable, encara que no totes les espècies es troben arreu. Segurament la continuïtat de totes les aigües oceàniques i els seus moviments rebaixen la importància dels factors de segregació i d’especiació que, en ambients continentals o d’aigua dolça, reflecteixen més la història i la geografia del passat. És clar que el coneixement encara és imperfecte, per falta de descripcions prou acurades i pel poc ús que es fa de criteris taxonòmics basats en diferències bioquímiques i genètiques. No es pot negar que hi ha moltes espècies de diatomees australs i que força dinòfits, principalment dels gèneres Ornithocercus, Amphisolenia i Histioneis, es coneixen solament en aigües fortament estratificades de les mars tropicals. Alguns organismes que estan en el mateix cas, com Dinophysis miles, fins i tot mostren clines o gradients de forma sobre extensions considerables.
Els ecosistemes planctònics
El cert és que en el plàncton no hi ha ecosistemes tan diversos i tan ben ancorats en àrees geogràfiques definibles com, posem per cas, el bosc tropical humit o la tundra àrtica. Però és cert que cada extensió oceànica té un estil propi de condicions i de dinàmica, i això defineix una participació més abundant d’unes espècies i menor d’altres.
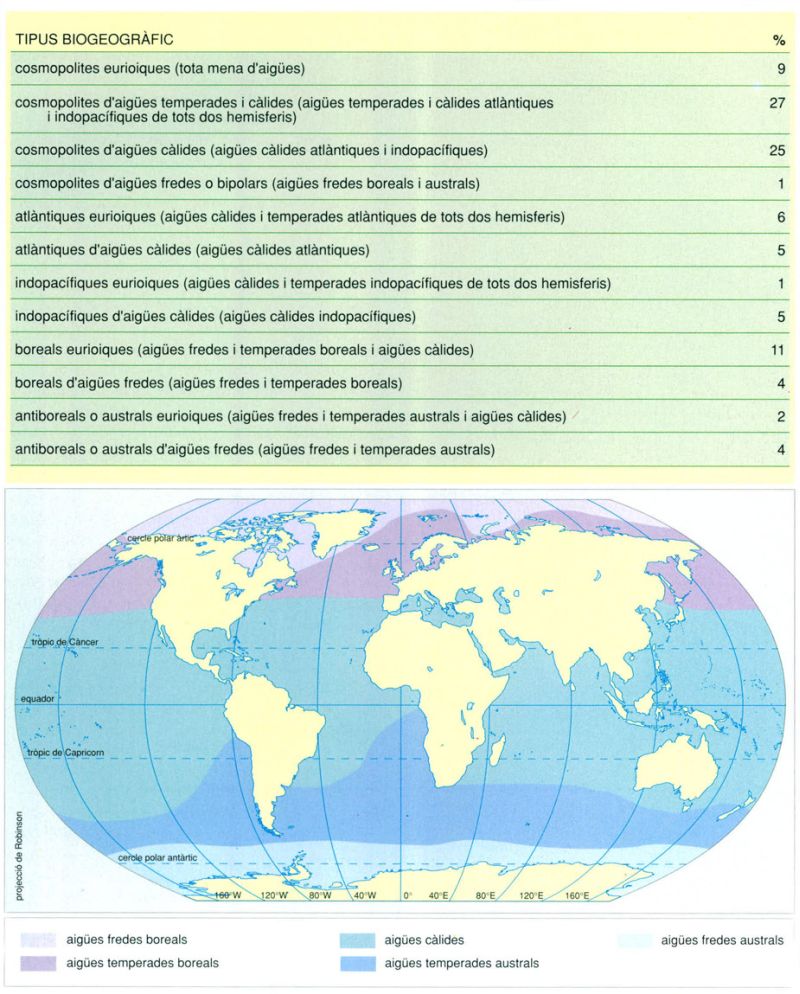
Tipus biogeogràfics i àrees de distribució del fitoplàncton. La distribució geogràfica de les espècies del fitoplàncton marí encara no es coneix prou per a poder fer generalitzacions que vagin més enllà d’una simplificació provisional que permeti l’ordenació còmoda de les dades existents. La que es dóna aquí es basa en la divisió dels oceans en cinc grans dominis: aigües fredes boreals, aigües temperades boreals, aigües càlides, aigües temperades australs i aigües fredes australs. Aquests dominis permeten de caracteritzar dotze tipus d’àrees biogeogràfiques en les quals es poden incloure totes les espècies conegudes de fitoplàncton marí. S’indica el tant per cent aproximat d’espècies de cada tipus.
Editrònica, a partir de dades compilades per l’autor
Sembla que no hi ha major dificultat en mantenir un “banc genètic” prou ric per a poder colonitzar efectivament l’aigua en qualsevol nova situació que es presenti. Amb relació a les aigües roges, s’ha especulat sobre el significat que pogués tenir la disponibilitat d’un nombre suficient de cèl·lules de resistència en el sediment dels fons propers als llocs on el fenomen es presenta. Se’n troben efectivament, però potser no són indispensables, perquè en estudis prou minuciosos de llocs molt diversos sempre es troben cèl·lules vives de la majoria de les espècies habituals en la flora local, si no de totes, les quals proliferaran de manera alternativa al llarg de les successions anuals o de temporada. Les cèl·lules quiescents són relativament nombroses en aigües profundes, on és evident que no estan fotosintetitzant, malgrat que es mantinguin vives.
Si el nombre d’espècies és relativament restringit i les diferències geogràfiques petites, és clar que uns tipus semblants de comunitats es poden reconèixer en llocs molt diversos. Si es volen o es poden distingir diferents comunitats, aquestes representen sovint diversos estadis de successions ecològiques, típiques del plàncton, que van de poblacions de baixa diversitat, sovint riques en diatomees (o en coccolitoforals), fins a situacions en què les poblacions són localment heterogènies i de més alta diversitat, especialment enriquides en dinòfits.
La taxonomia no està de moda, i l’estudi dels organismes del fitoplàncton sovint presenta dificultats, ja que són petits i fràgils, i les tècniques que demanen la fixació, el seccionament i l’estudi al microscopi electrònic volen, si més no, paciència, habilitat i temps. Però altres preocupacions de la biologia actual incideixen més positivament sobre l’anàlisi de les poblacions de fitoplàncton com, per exemple, en l’estudi de la possible existència de clons o races diferents dintre de cada una de les espècies “clàssiques”, que es manifestaria principalment en propietats fisiològiques i bioquímiques. La majoria de les “espècies” en les quals s’han cercat aquests clons o races, que són les que es cultiven amb més freqüència en els laboratoris, com ara les dels gèneres Phaeodactylum, Emiliania, Gonyaulax, Skeletonema, Thalassiosira i Prorocentrum, demostren diferències fisiològiques entre soques, pel que fa a la dependència respecte a temperatura i salinitat, a la capacitat heterotròfica i als requeriments de vitamines, la toxicitat i d’altres propietats que, certament, defineixen les probabilitats de supervivència alternativa de les respectives poblacions en un ambient natural mai uniforme.
Són prou conegudes algunes diferències comparables entre parells d’espècies que, encara que sovint es troben juntes, en realitat divergeixen per les seves preferències, com és el cas de Ceratium furca i Ceratium fusus, la primera més termòfila, menys halòfila, amb requeriments més elevats de fòsfor i amb més activitat de multiplicació en la superfície, o el de Nitzschia delicatissima i les espècies del grup seriata d’aquest mateix gènere, les segones pròpies d’aigües més fredes i profundes, amb més gran contingut salí i amb més fòsfor. Observacions locals repetides en el temps suggereixen moltes altres divergències ecològiques del mateix tipus. En organismes de morfologia molt senzilla, com ara els Synechocystis i Synechococcus del fitoplàncton, caldrà recórrer sempre a tècniques bioquímiques, fisiològiques i genètiques per a provar-ho.
Censos procedents d’una mateixa regió es poden combinar en un gràfic que arrenglera en ordre decreixent els logaritmes de la representació numèrica de cada una de les espècies, ordenades de més a menys freqüents. Això dóna una bona expressió de la diversitat. De vegades surten relatives sorpreses. Per exemple, la Mediterrània resulta una de les mars de diversitat més alta, superior fins i tot a la de qualsevol mar comparable i fins i tot més “tropical”, com la de les Antilles, si més no pel que fa a l’extensió meridional d’aquesta. Aquesta diferència està d’acord amb el fet que la densitat de plàncton i la seva productivitat és una mica superior al Carib que no a la Mediterrània.
La diversitat i la producció
En general, hi ha una relació inversa entre la diversitat i la producció biològica, i encara més que amb la producció, amb les seves fluctuacions, la freqüència i la intensitat de les quals, en els oceans, estan fortament correlacionades amb la mateixa producció. Tots els moments de màxima producció es caracteritzen per poblacions en les quals hi ha una o poques espècies extremament abundants (francament dominants), seguides per un llarg reguitzell d’espècies de reserva, però de moment poc representades.
Les esmentades relacions es visualitzen de manera adient representant gràficament les relacions entre nombre d’individus i nombre d’espècies. Mostren la irreversibilitat que caracteritza tots els canvis reals, i especialment dels que es refereixen a sistemes vius. L’augment de la biomassa —i de la densitat total del plàncton— es fa amb un nombre d’espècies restringit; assolit el màxim de biomassa, va augmentant el nombre d’espècies que hi contribueixen i, a la fi, baixa la biomassa present, però queden esparsos suficients individus d’un bon seguit d’espècies que hauran de facilitar el fonament de proliferacions futures.
Aquesta població barrejada, rica d’espècies, però majorment en estat quiescent, que podríem comparar amb un “banc de llavors”, se situa en aigües on arriba poca llum i és naturalment en aquest nivell on es manté sovint una diversitat global màxima, que és el sostre asimptòtic dels espectres de diversitat locals. Aquest mecanisme de persistència és comparable amb el de la vida en els rius, a les vores dels quals i a diversos nivells, sovint en un gruix insignificant de sòl i fins i tot a considerable distància de l’aigua corrent, es mantenen a disposició una àmplia reserva de gèrmens de tota mena d’espècies, que expliquen la recolonització continuada dels rierols de muntanya i dels rius de temporada.
La reserva d’alta diversitat és alimentada de manera alternativa i discontínua amb els resultats dels assaigs d’evolució que contínuament s’emprenen en les porcions més actives de l’ecosistema, que és on es fan els experiments ecològics, genètics i evolutius que, en el seu cas, enriquiran genèticament la diversitat de la mateixa reserva que es manté en el temps. Aquesta és una de les funcions de les ràpides i abundants proliferacions de fitoplàncton que, ací i allà, es donen contínuament a l’oceà.
La producció primària del fitoplàncton
Els productors primaris del plàncton són petits: la majoria fan entre 2 i 200 μm, i com que no tenen fibres, ni vasos, ni fusta, permeten enfocar i entendre el tema de la producció millor que la vegetació terrestre. Estan suspesos a l’aigua, on troben nodriment i llum.
Els pigments fotosintetitzadors
La clorofil·la i altres pigments actius en la fotosíntesi són fàcils d’extreure amb dissolvents adients i també és fàcil avaluar-los, tant colorimètricament com per la seva fluorescència. Al volum 1 d’aquesta mateixa obra (pàgs. 180 a 190) hi ha informació sobre la distribució de la llum al si de l’aigua i sobre l’estructura i la funció de l’aparell de la fotosíntesi, aplicable a la biologia del fitoplàncton. En el fitoplàncton marí, la relació entre el pes total de la clorofil·la (que per altra banda conté, ella mateixa, un 44% de carboni) i el del carboni orgànic se situa generalment entre 1:30 i 1:300, una proporció molt més elevada que la que es pot trobar en la vegetació terrestre pel fet que aquesta conté una fracció gran d’estructures de suport i transport desproporcionadament riques de carboni.
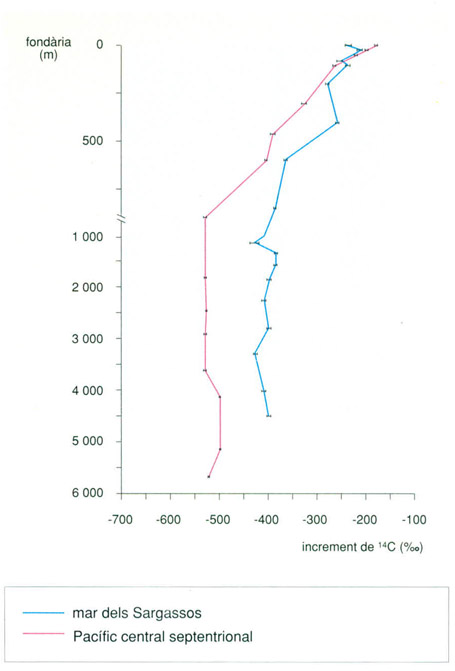
Contingut de 14C en el carboni orgànic dissolt, oxidable per la radiació ultraviolada en mostres del nord del Pacífic central (31°N, 159°W) i de la mar dels Sargassos, a l’Atlàntic (31°50’N, 63°30’W). Els valors de 14C són indicatius de l’edat de la matèria orgànica dissolta. Com que el tant per mil d’aquest isòtop minva a mesura que creix la fondària, la matèria orgànica és més vella com més avall es prenen les mostres, tal com posa de manifest el gràfic. De les dades també es pot concloure que a les aigües profundes del Pacífic hi ha més matèria orgànica dissolta que no pas a l’Atlàntic, fet que confirma la relativa joventut d’aquest oceà. Als 1 000 m hi ha un canvi en l’escala vertical.
Editrònica, a partir de fonts diverses
El biòleg nord-americà Alfred C. Redfield proposà, per al fitoplàncton, una relació mitjana entre les concentracions relatives de C:N:P, en àtoms, de 106:16:1, que és generalment acceptada i se’n fa molt ús. Multiplicant pels respectius pesos atòmics, es tenen les relacions en pes. En la vegetació terrestre, la quantitat relativa de carboni és almenys de dues a cinc vegades més gran, cosa que està d’acord amb el que s’acaba de dir. No és, doncs, correcte, comparar el fitoplàncton amb l’herba d’un prat polvoritzada i posada en suspensió; perquè la comparança fos correcta, el material esmicolat s’hauria de limitar als teixits fotosintetitzadors de les fulles. La vegetació epicontinental conté sovint entre 1 i 1,5 g de clorofil·la per metre quadrat. En els oceans, el fitoplàncton actiu s’estén en un gruix d’uns 100 m, que coincideix aproximadament amb l’alçada dels arbres més elevats; però la quantitat total de clorofil·la present és més petita i generalment no passa de 0,1 g per metre quadrat.
Els pigments presents en els sistemes fotosintetitzadors fan que siguin efectius els fotons que corresponen a longituds d’ona entre 400 nm (blau) i 800 nm (roig); aquesta és la radiació activa en la fotosíntesi. Hi ha un buit relatiu en l’absorció cap a la banda de major transparència de l’aigua (blau-verdós), centrada vers 520 nm; en tal segment de l’espectre solar poden absorbir radiació les ficobilines pròpies dels cianobacteris. I sorprèn que aquesta finestra s’aprofiti poc, ja que els cianòfits tenen una representació relativament migrada en les aigües marines, a diferència del que passa en les epicontinentals, on són força abundants.
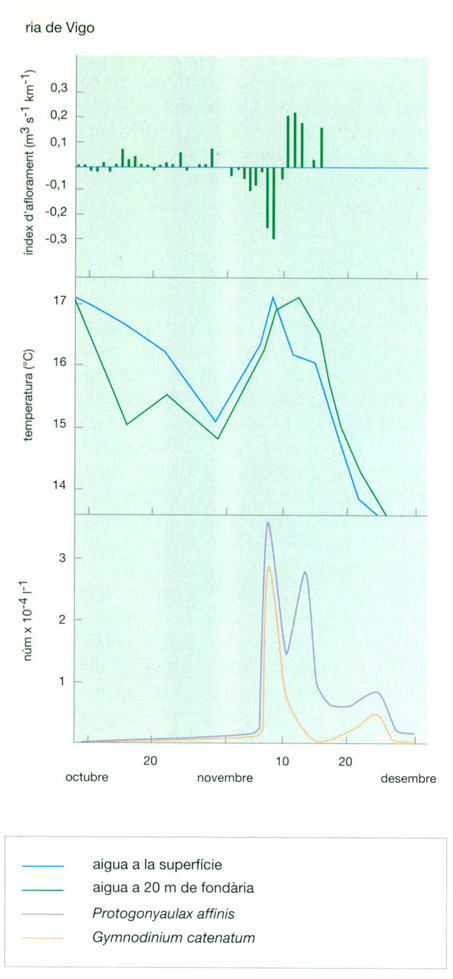
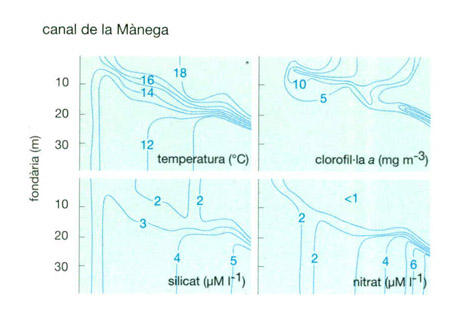
Vectors condicionadors de la distribució del fitoplàncton. A dalt, relació entre la fluctuació de les condicions ambientals i la presència de dinoflagel·lats tòxics (Gymnodinium catenatum) i (Protogonyaulax affinis) a la ria de Vigo a la tardor del 1985. L’aigua en relatiu repòs (índex d’aflorament baix) coincideix amb una temperatura superficial alta i el desenvolupament de dinoflagel·lats tòxics. L’aparició de l’aflorament costaner introdueix aigua més freda que acaba fent desaparèixer la “purga de mar”. A baix, altres exemples entre clorofil·la, nutrients i llum. Les observacions fetes al canal de la Mànega pel juliol del 1976 permeten de veure com la tendència és arribar més o menys ràpidament a una situació tal que, on hi ha molta llum, ja no queden elements nutritius. A baix, perfils més detallats de la mar Bàltica, on els pics de clorofil·la s’acumulen entorn del gradient màxim de temperatura.
Jordi Corbera, a partir de S. Fraga, Derenbach i Pingree
La radiació s’expressa correctament com a energia o es compta com a fotons (μE), cosa que es justifica perquè, dins dels límits de la radiació activa en la fotosíntesi, sembla que tots el fotons tenen una efectivitat semblant, independentment de llur energia respectiva. La distribució vertical de la llum disponible dins de l’aigua és funció de la seva transparència; a una fondària entre dues i dues vegades i mitja la màxima de visió del disc de Secchi es reconeix una zona crítica, on sovint té lloc una part considerable de la producció, en coincidència amb el màxim profund de clorofil·la. El carboni assimilat per gram de clorofil·la rarament excedeix 4 g per hora de llum, i davalla de pressa (logarítmicament) per sota d’uns 60 μE per metre quadrat i per segon (uns 8 000 “lux”).
La producció s’estudia a través del bescanvi d’oxigen, o de carboni marcat (14C) en petits volums confinats d’aigua amb el seu plàncton. La fotosíntesi inclou una reacció fotoquímica, la qual, com a tal, depèn menys de la temperatura que les fases ulteriors del metabolisme. La rapidesa amb què es consumeixen els nutrients continguts a l’aigua de mar permet completar la imatge. Hom ha intentat, a partir de la concentració dels pigments o, més recentment, de la seva fluorescència, fer estimacions de la producció. Es comprèn que hi hagi un desig de trobar algun indicador de la producció real que es pugui percebre des de l’espai, però, de moment, les relacions entre la productivitat potencial i la producció real són indirectes i es veuen dubtoses.
El paper dels nutrients
Si es posen en paral·lel dues llistes per comparar les concentracions dels diferents elements químics en els organismes i en l’aigua de mar i s’anoten els respectius quocients, es veu que gairebé sempre el fòsfor resulta l’element relativament més escàs i, per tant, el que deu limitar o controlar la producció. Com en una cadena de producció industrial, la velocitat de l’operació depèn d’aquell component que es proporciona amb més gran parsimònia. El carboni no sembla ser mai limitant, si no és per alguna raó de fisiologia específica que només excepcionalment es dóna en determinats organismes, ni tampoc ho són els elements que donen la salinitat a l’aigua, com el calci, el magnesi, l’estronci, el sodi, el potassi, el clor, el fluor, etc.
No s’acaba d’entendre com i per quines raons, a partir de cert moment, molts planctòlegs marins (Harvey, el 1955, posava correctament el fòsfor davant del nitrogen) s’han expressat com si creguessin que el nitrogen fos l’element que més sovint limita la producció primària marina. És cert que la quantitat de nitrogen dissolt en forma combinada (nitrat, nitrit, amoni) pot ser relativament petita, sovint com a conseqüència de desnitrificació deguda a l’acció de bacteris que fan passar el nitrogen de combinat a molecular, el qual, com a gas, es dissol en l’aigua. Però, pel que fa a aquesta forma molecular, com a gas dinitrogen (N2), l’aigua està en equilibri amb l’atmosfera, i això vol dir que l’aigua de mar normalment conté uns 13 mg (=10,5 cm3) de nitrogen per litre. El nitrogen combinat dissolt en l’aigua marina acostuma a ser només entre 1/100 i 1/1 000 de la concentració de nitrogen gasós. Costa creure que, si el nitrogen fos l’element limitant, no hi hagués hagut una invasió més gran de cianobacteris en el plàncton marí, semblant a la que s’observa en les aigües continentals i avui dia prou evident en les aigües enriquides en fòsfor. En elles no basta el nitrogen combinat, i els cianòfits fixen l’atmosfèric, amb el resultat que els llacs eutròfics se n’omplen. Hi ha cianobacteris en nombre considerable una mica pertot, però a la mar, les proliferacions significants i visibles de cianòfits filiformes (Trichodesmium o Oscillatoria; Nodularia) són bén escadusseres i sovint es detecten en llocs on precisament no hi falta nitrogen combinat, que tal vegada hi és per l’activitat passada de les poblacions dels mateixos cianobacteris.
Concentració de fòsfor i fertilitat
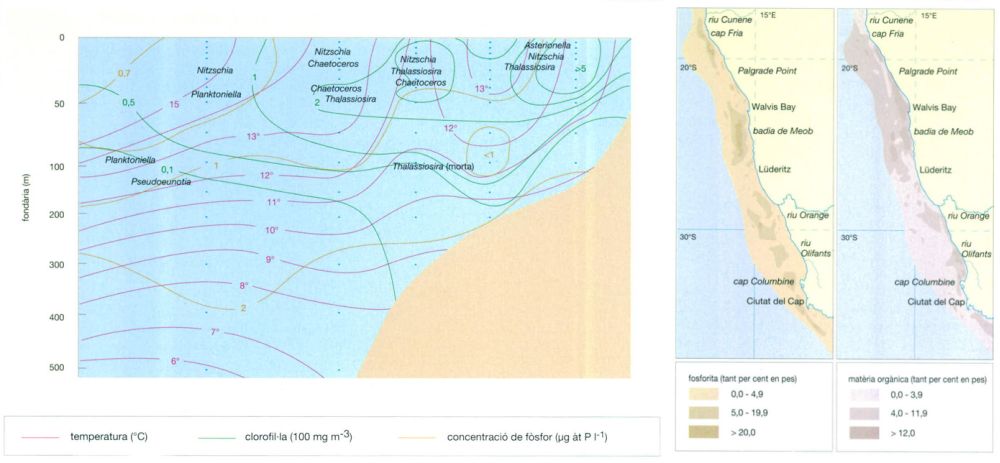
Jordi Corbera, a partir de dades de l’autor
La concentració del fòsfor a les aigües marines és decisiva per a la seva fertilitat. Però, tot i que a escala global el contingut de fòsfor de les aigües dels oceans és considerable (una mitjana de 2,3-2,5 micrograms-àtom de fòsfor per litre), les concentracions de fòsfor a les aigües superficials és negligible, ja que gairebé tot el que hi arriba és usat immediatament pels organismes. Només a les àrees d’aflorament el fòsfor procedent de les aigües profundes i dels sediments rics de nutrients en duu una mica més a la superfície. Però fins i tot en aquestes àrees, el fòsfor es concentra sobretot a les aigües profundes, tal com posa de relleu la superposició de les concentracions de fòsfor a disposició del fitoplàncton i de clorofil·la en un recorregut al llarg del paral·lel 26°S, prop de la ciutat de Lüderitz, a Namíbia, a l’àrea d’aflorament del SW d’Àfrica, durant la primavera austral. En aquesta àrea, com en altres àrees d’aflorament, la importància de la producció no dispersada ni consumida determina l’acumulació als sediments del fons de matèria orgànica i de fosforites, tal com mostren els mapes.
En el gràfic, corresponent a una presa de mostres feta del 19 al 21 de setembre de 1985, s’indiquen també els llocs on dominen determinats gèneres o espècies de diatomees de comportament diferent en relació amb la riquesa de nutrients. Thalassiosira es multiplica ràpidament (màxims de clorofil·la) i serveix d’aliment als copèpodes; a l’extrem contrari, Planktoniella, tot i tenir una organització semblant, es multiplica més lentament i té una mena de “faldilles” que en dificulten la ingestió, i és més freqüent mar enfora, on decau la intensitat de l’aflorament.
Les concentracions de fòsfor són baixes i resulten decisives per a la fertilitat de l’aigua. Si bé els nivells on no arriba la llum, en els grans oceans, en contenen una considerable reserva, amb concentracions de 30-60 mg per metre cúbic (a la Mediterrània, en canvi, només és d’uns 5 mg per metre cúbic), en els estrats il·luminats l’acció dels organismes manté sovint la concentració de fòsfor a nivells inapreciables. Part del fòsfor del qual poden disposar els organismes es pot trobar en forma orgànica, o poc reactiva, de la qual cosa en resulten dificultats suplementàries per a la determinació. Ordinàriament, les concentracions de fòsfor en les aigües superficials són negligibles; quan n’hi arriba una mica, s’usa ràpidament.
Els respectius comportaments del nitrogen i del fòsfor al mar presenten acusats contrastos. El primer dóna compostos solubles i circula entre aigua i atmosfera. El fòsfor, en canvi, es troba en els organismes en forma de compostos de l’àcid fosfòric, sense reduir-se mai a nivell elemental, i a l’aigua de mar s’hi troba combinat amb bases orgàniques o com a ortofosfat (HPO4=). Com que dóna compostos poc solubles, el seu cicle transcorre fonamentalment entre l’aigua i el sediment, i contínuament se’n perd o queda immobilitzat per aquesta via. El fòsfor actua en la dinàmica de la biosfera com el gran regulador o esmorteïdor.
Tots els sediments contenen fòsfor, i molt especialment els que es formen sota les aigües molt productives, en les zones d’aflorament, que són rics en fosforites. En aquestes mateixes regions, els ocells marins treuen fòsfor dels oceans i l’immobilitzen temporalment en el guano, contribuint així a rebaixar la productivitat dels sistemes pelàgics als quals deuen la seva vida. Aquest comportament és equivalent al dels petits crustacis planctònics migradors que, amb els seus excrements compactats, acceleren l’exportació del fòsfor cap a les profunditats més grans, retardant igualment la dinàmica global. És com si el cicle del fòsfor estigués sempre subjecte a pagar un tribut.
Aquesta mena de regularitats no sempre són benvingudes. Quan s’ha intentat fertilitzar artificialment àrees marines confinades per a cultivar organismes marins, s’ha trobat que una fracció considerable del fòsfor afegit es perd ja en el primer cicle, precipitant com a fosfat insoluble, de manera que no s’aconsegueix estabilitzar la productivitat a un nivell desitjablement alt. La continuïtat de la vida demana, per tant, la redissolució i la mobilització de més fosfat, i això, en bona part, s’esdevé sobre els continents.
Les correlacions significants no s’estableixen entre concentracions, sinó que cal basar-les en processos ajustats, per exemple, entre el consum de fosfat i el creixement del plàncton. La relació, com és natural, és la d’un procés assimptòtic, que s’alenteix a mesura que augmenta la massa dels organismes i disminueix la concentració de l’element que es va assimilant. A més, en cada moment, pot presentar-se alguna altra limitació, cosa que no exclou que hi hagi molts elements o compostos en excés.
És costum prendre com un indicador útil aquella concentració d’un element a la qual correspon una velocitat d’absorció que és la meitat de la màxima possible. Una cèl·lula mitjana o típica de fitoplàncton conté entre 0,3 i 2 pg (picograms, 1 pg = 10-12g) de fòsfor i la concentració de semisaturació pot ser entre 0,02 i 0,5 μg at P per litre o μM P (o senzillament, entre 0,6 i 15 μg de fòsfor per litre). Per al nitrogen, la constant de semisaturació varia segons el tipus d’ió o de molècula assimilada, però és unes deu vegades més gran que per al fòsfor, cosa que no ha d’estranyar si es recorda la relació de Redfield.
Repetidament s’ha pensat que la cadena de producció es podia trencar per l’escassetat d’algun element necessari que no fos ni el fòsfor ni el nitrogen, tal vegada per un component que es troba en petita quantitat, com el cobalt, el seleni i encara d’altres que semblen necessaris per a la vida de les plantes i els animals marins. L’aigua de mar és una mena d’extret de la part sòlida de la Terra, i sembla que els conté a tots en quantitat suficient, encara que localment hi podria haver alguna mancança relativa. Els anys vuitanta s’havia assenyalat el ferro com a possible limitant del creixement del fitoplàncton, i es va proposar enriquir els oceans australs amb compostos solubles d’aquest metall, per incrementar la producció de fitoplàncton. Així es contribuiria a fixar una part de carboni i tal vegada ens alliberaria del malson de l’efecte hivernacle. Però no hi ha raons convincents per afirmar que el ferro sigui limitant enlloc.
Cal afegir que el joc de les vitamines opera també en el plàncton marí i que hi ha molts organismes la vida dels quals requereix que trobin a l’entorn i ja sintetitzades determinades molècules orgàniques: cobalamines, per exemple. Però aquesta mena de relacions rarament deuen canviar les condicions de producció més comunes, ja que sempre hi ha prou varietat d’organismes perquè el seu conjunt pugui produir sota les condicions més generalitzades, tal com s’han presentat.
La producció nova i la producció reciclada
La producció primària del fitoplàncton és la base de la vida marina. S’hi afegeix el subsidi, relativament petit, que els oceans reben dels continents en forma de matèria dissolta, que arriba amb els rius. Toca entre 1 i 2 g de carboni orgànic per metre quadrat de mar i any. Una gran part d’aquest material és biològicament inaprofitable de manera directa, i no fa més que descompodre’s lentament a l’aigua.
No és fàcil organitzar críticament tota la informació disponible sobre producció. S’ha suggerit una mitjana mundial entorn d’uns 100 g de carboni fixat per columna de metre quadrat de secció i per any. Xifres d’aquesta mena demanen integrar, d’una manera fiable, un seguit de mesures de producció fetes a diverses fondàries, entre la superfície de la mar i el límit inferior de la fotosíntesi. Però la major part de les dades publicades sobre producció dels oceans es refereixen a producció neta del fitoplàncton, descomptant la respiració de les mateixes algues i, en realitat, tenen un elevat grau d’incertesa.
Els experiments fets per mesurar la producció posen de manifest les complicacions del reciclatge i les dificultats que comporta la seva quantificació. Regularment, per a aquesta mena de mesures, el plàncton, amb l’aigua, es confina en una ampolla. Es recomana separar prèviament els animals grossos, colant l’aigua per una malla que deixi passar el fitoplàncton, però que retingui els animals. Tot plegat resulta una tasca contradictòria i definitivament impossible, perquè hi ha algues, com són diverses diatomees amb llargs apèndixs, que fan més embalum que molts animals. A vegades aquella precaució no es pren i, en l’espai confinat del flascó, es verifica el metabolisme de tot un ecosistema complet sense que se’n puguin discernir les diferents partides. Si traiem els animals grossos, en queden de petits i en qualsevol cas resulta impossible deslliurar-se dels bacteris, que no es poden segregar fisiològicament de manera efectiva i pràctica de les algues planctòniques. Tots els experiments per mesurar la producció, sigui per bescanvi d’oxigen, sigui amb 14C, que tal vegada és un procediment encara més difícil d’interpretar, expressen el resultat d’una activitat biològica complexa, en la qual mai no ha cessat el reciclatge intern de part dels elements. La mateixa dificultat es fa palesa en grau extrem en sistemes simbiòtics molt integrats, com ara els coralls o diferents protoctists planctònics no fotosintètics dotats de simbionts, que manifesten a l’exterior una producció neta negligible, perquè interiorment recirculen els elements químics mercès a l’impuls de l’energia de la llum.
S’ha fet notar una internalització semblant dels cicles de la selva amazònica que, pràcticament, ni dóna oxigen ni pren CO2. Al capdavall, la Terra entera recicla i arribem a la conclusió que la distinció entre producció “bruta” i producció “neta” és funció de la delimitació intel·lectual que fem de l’entitat estudiada i de la interpretació que en fem. El funcionament real dels ecosistemes fa inoperants molts dels nostres esquemes conceptuals.
De manera més pragmàtica, l’espai pelàgic es pot dividir en estrats superposats. El nivell que se situa just per sota del límit fins al qual la llum és suficient per a la fotosíntesi resulta especialment important i ens separa una regió superior fòtica i una d’inferior afòtica, aquesta darrera sense llum o amb molt poca. El funcionament normal de l’ecosistema pelàgic té com a resultat net transportar els nutrients cap avall i acumular-los en el compartiment inferior.
D’unes dècades ençà, és usual deixar exposats, per dies o per setmanes, a fondàries normalitzades, col·lectors del material que se sedimenta contínuament: plàncton mort o les seves parts dures, excrements, material que arriba transportat per l’aire (el pol·len de pi, per exemple, es troba gairebé a tot arreu), etc. Els resultats són sovint incerts per inclinació dels recipients i per interferència amb corrents horitzontals però, en general, s’observa que tant la quantitat de material com el seu valor nutritiu disminueixen a mesura que augmenta la profunditat.
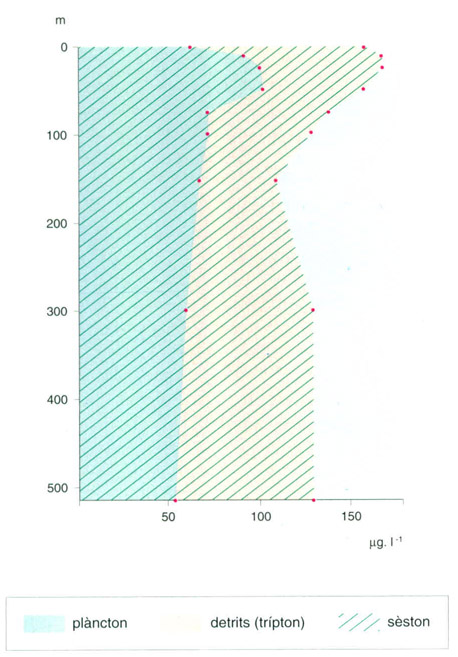
Distribució vertical del trípton (sèston que no és plàncton), en pes sec, i de mitjana, en aigües oceàniques de l’W i NW d’Austràlia, segons observacions d’A. Hagmeier (estiu del 1961), en una escala logarítmica. El material en suspensió, a més de plàncton viu, consta de partícules no vives, la informació sobre les quals és encara molt minsa.
Editrònica, a partir de fonts diverses
Una considerable fracció dels elements (nitrogen, fòsfor, etc.) s’assimila a la zona fòtica: el fitoplàncton és menjat per animalets, i aquests poden excretar voluntàriament o involuntàriament (en ser menjats) en el mateix nivell o més avall. Petits productors primaris, com cianobacteris —i també altres bacteris fotosintetitzadors— són ingerits per minúsculs flagel·lats i ciliats, en un cicle qualificat de “microbià”, perquè són organismes petits els que reciclen una part de la producció primària. Aquests organismes es poden trobar dispersos, però més sovint van associats amb material mort aglevat en flocs, a vegades grossos i visibles en forma de “neu marina”. La quantitat de material orgànic mort, detrític, en suspensió en l’aigua —el trípton— és com deu vegades el volum del plàncton, cosa de la qual no podem fer-nos idea si limitem el coneixement del plàncton, per una part al retingut per la tela d’una xarxa, i per una altra a cianobacteris i altres bacteris fotosintètics minúsculs. Al seu torn, aquest material detrític absorbeix compostos químics diversos i pot concentrar secrecions d’organismes. Tot plegat, un món crepuscular i discret, però important.
A les partides anteriors convé afegir la fracció de la producció primària total que serveix per a alimentar, principalment durant les hores de fosca, molts petits crustacis que migren alternativament amunt i avall, cosa que comporta a l’estrat superior una pèrdua més definitiva d’elements nutritius essencials. Més que per ells mateixos, aquests crustacis són importants pels seus excrements compactats, que se sedimenten amb considerable rapidesa fins que es desfan, en arribar a fondàries considerables, per l’acció de bacteris que sempre es troben adherits a la pel·lícula que els envolta i els manté compactes. Això sí, sempre que els excrements no siguin capturats per altres copèpodes més pobres que viuen a una fondària més gran i que se n’aprofiten.
A partir de determinada fondària, la vida es manté perquè rep una contribució o un subsidi de més amunt (l’única excepció coneguda són els “oasis” dels grans fons, mantinguts per quimiosíntesi entorn de surgències submarines, que es descriuen al capítol 3.5, àmbit 1, d’aquest mateix volum). Però aquell mannà deixaria de caure tan bon punt s’interrompés el procés de producció primària, per falta de nodriment. La reintroducció de nodriment mineral en superfície és necessària perquè no s’interrompi el cicle de producció i requereix l’energia que es manifesta en la circulació marina, energia que qualifiquem d’externa o exosomàtica, per contrast amb l’energia interna o endosomàtica associada amb la llum i el metabolisme dels organismes.
En cada localitat i per a cada temporada es pot determinar, si més no en teoria, un nivell que correspon al “centre de gravetat” entorn del qual es distribueix la funció de producció primària, com també un altre baricentre o centre de gravetat entorn del qual es distribueix l’activitat respiratòria del sistema. La distància entre els dos centres de gravetat esmentats es relaciona amb el camí que recorre, per terme mitjà, un àtom de carboni, des del lloc on s’assimila fins a aquell on es respira. Aquesta distància té relació amb el grau de superposició en la distribució dels factors de producció. Els dos centres de gravetat esmentats mai no podran coincidir, però la distància entre ells serà petita en un sistema fortament agitat i en canvi serà més gran, fins i tot molt més gran, en un sistema més estratificat. Si distingim compartiments, qualsevol d’ells té un reciclatge intern i una baga externa formada per vies d’exportació i d’importació, equivalents si la situació es manté més o menys estacionària.
És natural que el límit inferior de la zona on és possible la fotosíntesi resulti un lloc particularment adient per a situar una frontera imaginària o conceptual. Sempre hi haurà una pèrdua més o menys gran de material orgànic cap avall, vers les profunditats tenebroses. Si es manté un estat estacionari al llarg de períodes més o menys prolongats, cal que en el compartiment superficial entri un nodriment supletori equivalent (certes quantitats de P i N), sigui per transport horitzontal, per afloraments locals, o bé per barreja vertical de temporada. Aquesta aportació permet una producció qualificada de nova, per contrast amb la part de la producció mesurada a les capes superficials, que s’associa amb el reciclatge biològic que té lloc dins mateix dels estrats superficials.
La relació entre la producció nova i la total s’acostuma a designar amb la lletra f, i és una característica local important dels ecosistemes marins. El valor de f és alt en sistemes molt barrejats, i més aviat baix (per exemple entorn de 0,1-0,3) en ecosistemes que passen molt de temps estratificats i que tenen un metabolisme de reciclatge relativament intens a dalt, tal vegada perquè s’hi han seleccionat organismes (nedadors) que tenen menys tendència a perdre’s per sedimentació. Diversos oceanògrafs, i especialment R.C. Dugdale, R.W. Eppley i Goering, han remarcat que els fluxos de les diverses formes de nitrogen podrien servir per a estimar el valor de f si s’accepta que el reciclatge del nitrogen afecta principalment el nitrogen amoniacal —com és l’excretat per animals i altres heteròtrofs— i que la producció nova es basa en la recepció de nitrogen en forma de nitrat, procedent d’algun altre lloc, normalment de les capes d’aigua inferiors.
En les mars estratificades o en les temporades en què l’aigua està estratificada, hom reconeix un nivell molt definit, sovint esquinçat sobre el pla horitzontal, just dessota del màxim de clorofil·la, i considerablement enriquit de nitrit (6 a 20 mg de N per metre cúbic en la Mediterrània). Allí on es travessa el fort gradient de disminució de la llum, i aquesta va esdevenint insuficient per a la fotosíntesi, amb l’augment sobtat de la concentració de nutrients encara no utilitzats (la nutriclina), les algues del plàncton que es van sedimentant passen a una zona en la qual la llum resulta insuficient per a la producció fotosintètica, però en canvi poden absorbir nitrat i procedir al primer pas de la seva reducció química, que és a nitrit, tot i que no tenen prou llum per assimilar-lo i probablement el deixen anar, cosa que explicaria la seva acumulació en un nivell molt precís. És clar que secundàriament s’hi pot afegir una activitat bacteriana d’oxidació de nitrogen amoniacal. Cal esperar que l’estrat de màxim de nitrit serà mès contrastat quan el valor de f sigui baix.
La dinàmica de la producción fitoplanctònica
El mateix Victor Hensen (1835-1924) que, el 1887, introduí la denominació de plàncton, va emprendre estudis força minuciosos del de la badia de Kiel, iniciant així una aproximació quantitativa a l’estudi del plàncton i tractant d’entendre les causes ambientals i les interaccions entre diferents organismes implicats en la dinàmica de la vida pelàgica. Hensen també va intervenir en la Planktonexpedition del National (1889), que fou la primera expedició oceanogràfica monogràfica sobre el plàncton marí i que inicia una aproximació quantitativa global (a tot l’Atlàntic) a l’estudi d’aquest. Contemporàniament, i encara més en entrar el segle XX, l’interès per conèixer les causes de les variacions en les poblacions de peixos féu que en moltes localitats costaneres —Ostende, a Bèlgica, el 1843; KonkKerme (Concarneau), a Bretanya, el 1859; Rosgo (Roscoff), també a Bretanya, el 1871; Woods Hole, a Massachusetts, el 1873; Nàpols, a Itàlia, el 1874 —s’instal·lessin laboratoris, i una de les seves funcions fou sempre estudiar les fluctuacions locals de les poblacions de plàncton, en les quals, prou correctament, es veia el fonament de la fertilitat de les aigües. El que començà fent un calendari, acabà en un estudi de la causació.
Les variacions de la producción en el temps i en l’espai
A les regions temperades, si més no prop de les costes, era evident una gran abundància de plàncton —primer diatomees i després copèpodes— a la primavera, quan el dia s’allargava i la temperatura de l’aigua en superfície anava augmentant. Ja des del 1906 W. Natanson tenia ben clar que l’augment de fertilitat era degut principalment a l’aproximació a la superfície il·luminada d’aigües de procedència profunda i, per tant, riques encara en nodriment mineral. Els comentaris sobre la producció nova i la producció reciclada, de la darrera secció, són de plena aplicació a les situacions més estables que vénen després; però les grans proliferacions de vida marina sempre ocorren a batzegades, i el seu origen queda sovint incert.
Les observacions de H.H. Gran i T. Braarud (1935), fetes primer a la costa de Noruega i després esteses al golf de Maine, posaren de manifest la falta d’uniformitat del màxim planctònic de primavera, cosa que els portà a reconèixer els factors que en controlen la producció. En aigua molt turbulenta i amb el seu contingut de nodriment uniformitzat en tota la fondària, el fitoplàncton augmenta primer en les àrees menys profundes, on el fons posa un límit inferior a la possible dispersió del plàncton de la columna d’aigua cap a nivells poc il·luminats. Si la fondària excedia de 100 m i l’aigua era molt agitada, cada cèl·lula, de mitjana, passava massa poc temps sota una llum suficient per a poder-se multiplicar indefinidament. En aquestes situacions es retardava l’arrencada d’una producció neta, amb excedent. Però la gran producció arrencava decididament tan bon punt avançava la primavera i la llum penetrava més, també perquè començava l’estratificació tèrmica de l’aigua que limitava l’extensió de la mescla vertical.
Perquè es dispari la producció, cal que la fondària de barreja, mesurada a partir de la superfície, no excedeixi de certa relació amb la fondària de compensació, que és aquella on la producció és igual a la respiració del sistema, de manera que no deixa producció neta. Segons el noruec Harald Sverdrup (1953), aquella relació límit se situaria entre 1,5 i 5,5, segons quines siguin altres circumstàncies locals. A mesura que la relació va baixant, per augment de la llum i per decreixement del gruix de l’estrat fortament agitat, la producció massiva de fitoplàncton s’estén gradualment, de les regions més properes a la línia de costa i menys profundes a les més profundes situades mar endins.
Aquesta visió, des de terra, es millora contemplant el que passa des d’alta mar. Els centres dels oceans, de fa temps, havien estat reconeguts com a particularment erms: mai no s’hi desenvolupaven denses poblacions de diatomees ni grans moles de peixos. La mar dels Sargassos ha estat un exemple clàssic d’aquesta situació, al qual sovint es fa referència. Tanmateix, en data relativament recent, s’hi ha reconegut una activitat gens menyspreable, en la qual tenen un paper important els minúsculs organismes del picoplàncton.
La producció i les àrees d’aflorament
Una situació força diferent s’observa en les àrees d’aflorament o d’“upwelling”. En elles l’acció dels vents, interactuant amb la direcció de corrents costaners i en conjunció amb forces associades a la rotació de la Terra, allunya de la costa les aigües superficials, que van essent substituïdes per aigües profundes més riques en nutrients. D’aquesta manera s’alimenta una producció primària elevada durant llargs períodes ininterromputs. Aquestes regions sempre han estat centres d’activitat pesquera.
Es pot calcular la velocitat d’ascensió de l’aigua profunda i la quantitat de nodriment que injecta en la zona fòtica, com també l’energia cinètica procedent del sistema atmosfera-hidrosfera que realitza localment el treball mecànic que és suport d’una producció primària elevada i seguida. Aquella energia exosomàtica és, pel cap baix, d’entre 25 i 50 vegades l’energia de la llum que intervé en la producció biològica. Una relació semblant es retroba en la vegetació terrestre, on l’energia exosomàtica es comptabilitza principalment com a evapotranspiració. En el cas de l’agricultura s’hi afegeix a més l’energia auxiliar invertida en regar, fertilitzar i treballar la terra.
L’aplicació al plàncton dels models d’interacció ecològica formulats per Alfred J. Lotka (1925) i Vito Volterra (1926) no es féu immediatament, malgrat que Volterra s’inspirà específicament en un problema de producció pesquera. La dificultat bàsica és que aquells models s’adreçaven a descriure només la interacció entre espècies i conjunts d’espècies, com a entitats extretes del seu marc, i no tenien en compte ni l’organització de l’espai ni l’energia externa, la importància de la qual és òbvia en les poblacions marines.
El model de la columna d’aigua
Gordon A. Riley introdueix l’espai de manera subreptícia, a través de la sedimentació i de la difusió, aquesta darrera associada amb l’energia de la turbulència. Les conclusions d’una aproximació a l’ecologia del plàncton atlàntic, per Riley, Stommel i Bumpus (1949), proposaven un model segons el qual: dF/dt = (r–gZ)F - V dF/dz + Azd2F/dz2, on, si convé, farem: r–gZ = r’. En aquesta expressió, F representa el fitoplàncton, Z el zooplàncton, V és la velocitat de sedimentació (= 0,001-0,02 cm per segon) i A és el coeficient de difusió vertical, que és una mesura de la turbulència; z indica que es refereix a la dimensió vertical. A, inicial d’Austausch, o canvi, és un coeficient de difusió, que expressa la transmissió de calor, salinitat o moviment en un líquid, i les seves dimensions són g/cm i segon (com la viscositat dinàmica, dividit per la densitat, que és aproximadament 1, resulta la viscositat cinemàtica, de dimensions cm2 s-1). El plàncton és escàs prop de la superfície, no per excés de llum, sinó per la discontinuïtat que representa la superfície del mar, per sota de la qual es va sedimentant el plàncton, però on no n’hi entra si no és per divisió de les cèl·lules. A és pot calcular de manera aproximada mesurant la transmissió de calor cap avall, a través d’un determinat gradient tèrmic. Referit a la vertical (Az) i a la zona fòtica de la Mediterrània, s’obtenen valors inferiors a 2 entre maig i juliol, de 2 a 5 en abril-maig i en juliol-agost, i més alts que 5 entre setembre i abril, quan les aigües són més agitades i també més productives. En la direcció horitzontal, Ax, ateny valors molt més alts.
En la forma abans proposada, el model descriu tot just una columna vertical d’aigua i no es pot aplicar directament a un món real, en el qual importen molt les diferències horitzontals o entre columnes adjacents. L’estructura física de la mar és molt complicada, amb discontinuïtats gairebé tan mòbils i tan imprevisibles com aquelles a què ens té acostumats l’atmosfera. De manera que predir l’evolució de les poblacions planctòniques resulta tan difícil com predir el temps que farà.
Les heterogeneïtats i les asimetries
D’altra banda, el món pelàgic conté un conjunt de reaccions químiques que tenen lloc en un espai que es va estructurant progressivament, com passa en alguns sistemes de tipus col·loïdal. Realment, la successió ecològica en els sistemes pelàgics es fa en un medi que es va estructurant o, si més no, que es va estratificant tèrmicament d’una manera progressiva.
Les consideracions de Smoluchowski (1918) pel que fa al cas ja suggerien expressar la producció P com dF/dt = P = A·C, expressió en la qual A continua representant l’energia que es dissipa en un moviment turbulent de barreja i C és la covariància o grau de superposició en la distribució dels factors de producció, bàsicament clorofil·la, llum i nutrients. Si hi ha reciclatge extern, s’ha de considerar una baga que passa per fora de l’espai de referència i que uneix les importacions (I) i les exportacions (E); aleshores P és la producció que es reté localment: P + E = I + AC i P = A·C. En la seva forma simplificada, sense la baga de reciclatge, aquesta expressió es pot derivar respecte al temps, i aleshores dóna:
d2F/dt2 = dP/dt = C dA/dt + A dC/dt,
que expressa, potser amb alguna elegància, la successió, permetent interpretar com se sol combinar el decrement de l’energia disponible en la difusió o turbulència, amb una disminució de la covariància en les distribucions dels factors de producció. Aquesta disminució de la covariància equival a un augment de la segregació entre els elements, interacció que contribueix a la producció com resulta evident: sempre es tendeix que, allí on hi ha llum, no hi quedin nutrients, i que aquests només s’acumulin allí on no arriba la llum.
Model de la formació de remolins i de reticles d’aigües descendents envoltant petits remolins d’aflorament d’aigües profundes a l’hemisferi nord. La força del vent genera moviments horitzontals que, en compondre’s amb el moviment de rotació de la Terra, origina alternativament remolins de sentit de gir oposat. Els anticiclònics (els que giren en sentit horari a l’hemisferi nord i antihorari a l’hemisferi sud) tendeixen a estratificar aigua més lleugera al seu centre i a eixamplar-se fins a fusionar-se lateralment els uns amb els altres. Els ciclònics (que giren en els sentits oposats als assenyalats a cada un dels respectius hemisferis) actuen en el seu centre com un xuclador d’aigua profunda, amb la qual cosa porten a nivells il·luminats aigua més rica de nutrients i mostren una fertilitat més gran, però tendeixen a estrènyer-se i a mantenir-se separats al si de l’estructura reticular derivada dels remolins anticiclònics.
Jordi Corbera, a partir de dades de l’autor
Totes les heterogeneïtats que es produeixen mostren asimetries. Per exemple, si es formen remolins de diferents sentits de gir, per interferència amb la rotació de la Terra es trenca la indiferència primitiva en qualsevol mosaic irregular, en el sentit que els centres de més gran producció associats amb els remolins ciclònics (sentit oposat a les busques del rellotge en l’hemisferi nord) es manifesten en forma de taques isolades en un fons més indiferent, i no pas de la manera inversa.
Actualment, estructures d’aquest tipus es poden reconèixer en imatges obtingudes des de l’espai. Ara i abans, en la rutina oceanogràfica, s’han fet i es fan anàlisis en continu o transsectes (temperatura, salinitat, clorofil·la mesurada generalment per fluorescència) a partir dels quals es pot postular una estructura general bidimensional raonable en la qual s’espera que tinguin cabuda i trobin interpretació les heterogeneïtats revelades pels segments lineals analitzats. L’única estructura compatible resulta ser la formada per taques discontínues més productives, esparses sobre un fons de productivitat o de biomassa més baixa. Les esmentades taques de fertilització coincideixen, òbviament, amb remolins ciclònics, encara que no sempre.
La transformació logarítmica de les concentracions de plàncton X [x =log (X+1)] al llarg d’un transsecte és la que fa més simètrics els gràfics i fa pensar en la manera com moltes distribucions de plàncton en l’espai recorden els perfils fractals de les cadenes de muntanyes. Si s’estenen a dues dimensions mostren cims que apareixen isolats o discontinus. Cims que, en el nostre cas, corresponen a centres d’alta producció, des dels quals es dispersen les poblacions de fitoplàncton. Aquest és un estil molt generalitzat a la naturalesa i segurament el més comú en la distribució de la producció marina, en la qual sempre s’ha fet notar la manifestació de densificacions locals o taques.
Avui dia, els ordinadors ofereixen moltes possibilitats encoratjadores que permeten simular com el plàncton es podria multiplicar i evolucionar d’acord amb suposicions definides. És natural que els models de fluxos i salinitats, on hi ha continuïtat, resultin més acceptables que els models biològics, en els quals l’esdevenir històric hi intervé amb totes les seves incerteses. L’escassetat de dades reals porta sovint a excedir-se en les interpolacions, no sempre correctes, si es té present la freqüència de discontinuïtats i pertorbacions locals. A qualsevol escala les perspectives de predicció correcta són encara poc falagueres.
Els processos de successió i els tipus biològics
Els ecosistemes mai no conserven per molt temps la mateixa organització. Si es mantenen actius en un entorn raonablement estable, sempre poden seguir ocupant l’espai amb estructures lleugerament diferents (encara que equivalents) que impliquin un menor cost de manteniment, o bé enriquir la pròpia complexitat sense augmentar el cost energètic. Normalment aquests canvis van associats amb el gradual consum i disminució de la quantitat de nodriment que hi ha a disposició. Tot això constitueix l’essència mateixa de la successió ecològica que, en el plàncton, ofereix la particularitat, fonamentada en l’escala temporal pròpia de l’ecosistema pelàgic, que tots els esdeveniments i els canvis van molt més ràpids que en ecosistemes bentònics o continentals. La rapidesa més gran facilita la manifestació de diferències locals.
La successió en el fitoplàncton
La successió en el plàncton mai no té característiques constants ni uniformes entre punts separats, encara que siguin veïns. Les diferències locals generalment s’han generat en el temps, sigui perquè els canvis han anat més de pressa en uns llocs que en uns altres, sigui perquè seqüències equivalents s’han iniciat en moments diversos, tot en dependència de diferències locals en l’agitació de l’aigua, la concentració de nutrients i les distribucions de la llum. Els canvis són sempre asimètrics: els canvis graduals, autoorganitzatius i relativament lents són truncats o interromputs per pertorbacions aleatòries i sobtades, que solen acompanyar qualsevol intensificació de la mescla turbulenta de l’aigua o desplaçaments de masses d’aigua d’un nivell a un altre.
Temps i espai es combinen en la dinàmica de les poblacions planctòniques. Més d’una vegada s’han comparat les proliferacions de fitoplàncton a núvols, perquè les cèl·lules es multipliquen més allí on hi ha moviments d’aigua de direccions i intensitats apropiades i que són particularment efectius quan apropen nodriment vers la llum. Però tots els moviments, siguin unidireccionals, com els corrents marins i els afloraments, siguin alternatius i confusos, com ara la turbulència, depenen d’energia externa que, al cap i a la fi, ve del Sol.
L’aigua és de densitat una mica més baixa que la dels mateixos organismes, amb un lleuger gradient d’augment cap avall, i rep energia lluminosa, tèrmica i mecànica, principalment a nivell de la superfície. El creixement és intens mentre hi hagi coincidència, en l’espai, de cèl·lules (o de clorofil·la), llum i nutrients. L’aigua tendeix a escalfar-se en superfície i a estratificar-se per densitats i això s’oposa a la barreja vertical local. La mescla és feble i pot originar-se de manera indirecta, associada a moviments de transport horitzontal.
Una barreja vertical massa intensa, en aigües profundes, fa que cada cèl·lula del fitoplàncton, per terme mitjà, gaudeixi de la llum durant una fracció massa breu del seu temps, amb el resultat d’un creixement negligible o nul i la dispersió excessiva de les poblacions. Si el gruix de l’estrat de mescla excedeix en cert factor (1,5 a 5 vegades, segons altres circumstàncies) la fondària de compensació —aquella a la qual la producció biològica i la respiració s’anivellen— hi ha massa pèrdues i el plàncton no augmenta. Però encara que es donin les condicions òptimes, la coincidència o l’alta covariància en la distribució dels factors o agents de producció davalla aviat, sigui perquè la població se sedimenta i passa a nivells en els quals la llum és insuficient, sigui per esgotament dels nutrients o, més generalment, per tot plegat.
Producció i òxids de silici
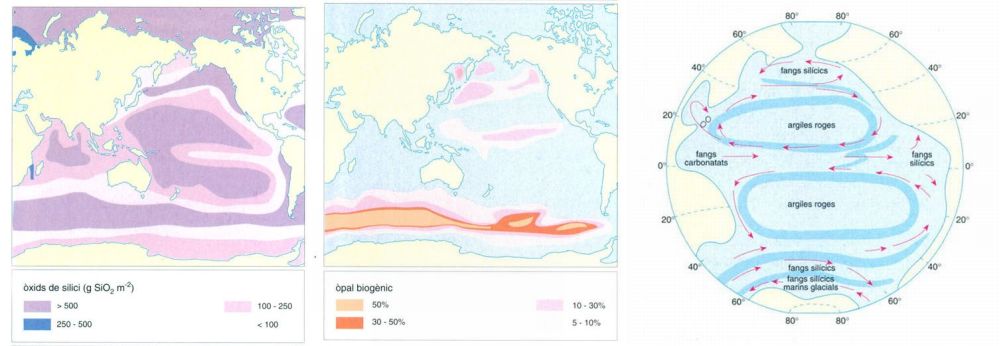
Editrònica, a partir de dades de Lisitzin
Comparació entre la producció i la presència d’òxids de silici a la superfície dels sediments a l’oceà Pacífic i a l’Índic, i model generalitzat de l’acumulació de sediments als fons marins.
La producció d’òxids de silici pel fitoplàncton (principalment per les diatomees i més secundàriament pels silicoflagel·lats i els radiolaris, més nombrosos a les aigües equatorials) s’expressa en grams d’òxid de silici per metre quadrat i any, mentre que la de sílice amorfa (òpal) s’expressa en tant per cent del sediment sec lliure de carbonat. Els valors màxims de contingut de sílice amorfa al sediment se situen entre la convergència i la divergència antàrtiques, però una mica desviats en relació amb les zones de màxima producció. Es troben màxims relatius de producció que es poden relacionar amb les àrees d’aflorament i de divergència d’aquests oceans i que es reflecteixen també en els sediments de la franja equatorial del Pacífic oriental. En un model generalitzat de sedimentació oceànica, els fangs silicis tendirien a acumular-se en les àrees d’aflorament de les ribes orientals i sota les divergències equatorials, mentre que als centres dels hemioceans de cada continent, reflectint la seva escassa productivitat, s’acumularien argiles roges, pobres de matèria orgànica.
Un principi fonamental de la vida planctònica és que tot àtom d’un element que passa del medi al cos d’un organisme probablement retornarà del mateix organisme o d’un altre (un planctòfag o un depredador, per exemple) al medi, a un nivell més baix, és a dir, més a prop del centre de la Terra que el nivell en el qual fou assimilat. El resultat net és que hi ha un transport generalitzat cap avall dels elements continguts a l’aigua i que són necessaris per a la vida. La situació relativament estacionària —amb els seus alts i baixos fugissers o locals— a què s’arriba és el resultat de la interacció o la interferència entre processos biològics i moviments de l’aigua. Ordinàriament el grau de major activitat biològica s’estableix a nivells gairebé crepusculars des del punt de vista de la disponibilitat de llum: entre 40 i 100 m de fondària. Aquest estrat, de pocs metres de gruix, continua mantenint una producció biològica modesta, estimulada pel cicle diari d’activitat i per possibles ones internes, entre la zona superior plenament il·luminada i eixorca, i la zona inferior rica en nutrients, però fosca. Com que el creixement és controlat pel subministrament de nodriment, no cal estranyar-se que el plàncton vegetal, en multiplicar-se sota una llum crepuscular, faci aparentment mal ús de la llum, que en realitat sobra.
La màxima quantitat i activitat del fitoplàncton, doncs, se situa a certa fondària, no perquè la llum en superfície sigui nociva per massa intensa, sinó perquè la superfície de la mar, a més de tenir el nodriment exhaurit, és una discontinuïtat a partir de la qual els organismes se sedimenten, però a la qual no n’arriben més si no és venint de més avall i encara, generalment, només per mescla turbulenta de l’aigua. De fet, la major part de la llum que arriba a l’oceà és simplement absorbida per l’aigua i no resulta de cap profit directament per a la vida. Es comprèn que la quantitat de clorofil·la que es troba per metre quadrat als oceans sigui, en general, més baixa (i sovint moltíssim més) que la que conté la vegetació epicontinental. Normalment, per unitat de superfície, és entre 1/10 i 1/20 de la que es troba en la vegetació terrestre.
Perquè el fitoplàncton produeixi cal que coincideixin organismes, llum i nutrients. Els organismes són, en general, més densos que l’aigua i, en conseqüència, hi ha un flux imparable de dalt a baix, encara que sovint retardat, mercès als variats artificis que faciliten la natació o la flotació passiva, tan fatal com la mort de cadascú de nosaltres.
El retorn dels elements químics necessaris a les aigües superficials il·luminades i la continuació del cicle de la vida pelàgica depenen de l’energia exosomàtica o externa. La productivitat primària o producció del fitoplàncton esdevé proporcional al subministrament dels elements necessaris, en primer lloc el fòsfor, després el nitrogen i encara d’altres. Aquest sembla l’ordre corrent, encara que no s’exclou que localment es pugui canviar i passar a ser limitant algun element diferent del fòsfor. El carboni no sembla que ho sigui mai, o sols localment i per a organismes que hagin evolucionat donant per pressuposat un gran excés de carboni en forma de CO2, i que es trobarien en inferioritat en aigües massa alcalines en les quals una fracció més gran del carboni inorgànic és present en forma de bicarbonat (HCO3-).
Cap situació no es manté estacionària, persistent i uniforme per ella mateixa. La naturalesa, en general, i molt visiblement la mar, es mou a batzegades. Des de les maltempsades a les ones, superficials o internes, introdueixen les seves discontinuïtats, a diverses escales. Aquestes intermitències afecten més els nivells superficials que reben llum i que són els únics productius. Certament existeix una vida abissal prou rica i variada, alimentada per material que cau dels nivells superiors, i aquest mannà que ve del cel ho acaba fent de manera força uniforme, perquè integra molts petits episodis productius i dispersos. Aquests episodis activen, aquí i allà, la zona fòtica, dintre un marc de temps i d’espai suficientment ampli per a permetre integrar i fer uniforme, mentre fa via vers el fons, el flux d’una part de la producció neta generada en la superfície.
Les estratègies natatòries
Cada un dels majors grups taxonòmics representats en el fitoplàncton mostra certa “vocació”, com a conseqüència de característiques fonamentals (proveïts d’un llast d’òpal o de calcita), o com a resultat d’una evolució secundària adaptativa (mida o presència d’apèndixs diversos). Generalment, els representants de cada grup precís viuen i es multipliquen millor —i per tant se seleccionen— sota una constel·lació definida de condicions ambientals, o bé en determinats moments de les seqüències successionals associades amb els respectius episodis productius, la duració dels quals és variable, generalment entre setmanes i mesos.
Si l’aigua és nutritiva en superfície, la producció va ràpida. Més encara si l’aigua és ascendent i es renoven contínuament els nutrients allí on la llum és intensa; en tal situació és profitós deixar-se sedimentar a contracorrent en el si de l’aigua que puja, mantenint un nivell de lluminositat acceptable. En aigua més reposada o més pobra, les cèl·lules no aconsegueixen dividir-se tan de pressa i val la pena no perdre nivell i augmentar, en la mesura que sigui possible, la capacitat d’absorció del poc aliment que queda. És acceptable destinar una part de l’energia a mantenir la cèl·lula en moviment, mitjançant flagels, i encara millor si la natació determina remolins que augmentin l’absorció; això es pot aconseguir per mitjà de conformacions de la cèl·lula que ens podrien semblar capricioses, però que, sens dubte, expressen un gran respecte per les lleis de la hidrodinàmica. Les mateixes formes, potser amb petits afegitons, poden tenir ensems una funció de defensa o de dissuasió enfront d’enemics potencials.
Els tipus biològics principals que s’assenyalen a continuació són termes d’una seqüència, més que no pas classes discontínues, encara que l’evolució divergent que les distancia hagi pogut incloure la resolució necessària d’alguna disjuntiva. Diatomees i coccolitoforals són organismes poc o gens mòbils i amb llast, que es poden multiplicar ràpidament i que se sedimenten també amb facilitat; els silicoflagel·lats també potser es podrien associar al mateix grup ecològic. Tots són organismes com els que, en dinàmica de poblacions, s’acostuma a dir que practiquen l’estratègia de la r: multiplicar-se de pressa, aprofitant el nodriment del moment, i que passi el que hagi de passar. D’altra banda, hi ha els organismes típicament nedadors, representats fonamentalment pels dinòfits, molt antics, els vells senyors del plàncton, als quals es poden unir altres grups menors; en general, la seva supervivència s’associa amb l’estratègia de la K, on una multiplicació relativament parsimoniosa es combina amb una natació constant o alguna altra manera de sobreviure en un entorn relativament pobre en recursos. El fitoplàncton marí és generalment molt barrejat, però és notable la relativa alternança, en el temps i en l’espai, del domini dels dos grups assenyalats. Els del segon grup, especialment els dinòfits que són més diversificats, poden persistir sempre en poblacions més o menys riques i fluctuants. Tan bon punt les condicions externes faciliten accelerar la producció, a la vegetació preexistent s’afegeix el resultat de ràpides proliferacions, sovint molt locals, de diatomees, o també de coccolitoforals o de Phaeocystis.
Les regularitats i les variacions
Segons les localitats varia l’estil del canvi en les condicions de vida. Cada any es poden repetir aproximadament i en seqüències semblants les mateixes situacions pròpies de les diferents estacions: a l’hivern hi pot haver barreja intensa de l’aigua, marcada per poblacions de diatomees, i la resta de l’any l’aigua s’estratifica i gairebé es buida de vida en els estrats més superficials. Entre situacions extremes s’observen diferències de producció com d’1 a 10 i encara més grans.
Sovint s’observen proliferacions de fitoplàncton irregularment distribuïdes i que varien ràpidament: hi contribueixen els remolins menors (entre 10 i 20 km de diàmetre) que es generen sota l’acció de vents locals. Si hi ha remolins de diferent sentit de gir, els ciclònics fan pujar aigua en el seu centre, són més productius i solen contenir més diatomees; els anticiclònics són molt més passius. Una altra modalitat de circulació es genera també sota l’impuls del vent i té la forma de bandes paral·leles estretes (d’uns centenars de metres) i allargades, que constitueixen les anomenades cèl·lules de Langmuir, visibles sovint des d’avions, especialment prop de les costes, on tenen l’aspecte de faixes d’amplada de prop de centenars de metres, alternativament lluents (superfície neta, aigües ascendents, amb més diatomees) i més opaques (aigües descendents i amb una proporció més gran d’organismes nedadors).
Els transsectes d’estudi que creuen una regió amb moltes estructures heterogènies donen seqüències quantitatives que tenen la forma del perfil de cadenes de muntanyes. Són compatibles amb una distribució que consisteixi en taques d’alta producció, aïllades i discontínues, com són també els típics remolins ciclònics, i disperses sobre un fons de productivitat menys excitada.
Els fronts marins es descriuen en els límits de corrents o bé allí on es toquen corrents o masses d’aigua de moviments diferents, convergents, divergents o amb lliscament lateral, els quals, en llur aiguabarreig, fan un treball i poden aportar nodriment. Aquest nodriment, sovint complementari, fa que siguin llocs de producció biològica més elevada; poden contenir proliferacions de diatomees o de coccolitoforals, talment com les regions més genuïnes d’aflorament amb forta presència i fins predomini de diatomees. Els fronts de marea, amb aigua barrejada, allotgen sovint denses poblacions de coccolitoforals.
En situacions d’estratificació hi pot haver una aportació seguida de nodriment, introduïda sovint horitzontalment, tal com passa en badies o altres sistemes costaners de característiques adients, sovint en règim d’estratificació persistent, generada per l’advecció o transport superficial d’aigua menys salada. Allí es mantenen (sovint es multipliquen copiosament) organismes nedadors que arriben a produir “aigües” o “marees roges”, en les quals són freqüents dinòfits tòxics. Ja es pot imaginar que hi ha regions marines especialment afectades per aquestes condicions i això es pot haver agreujat en incrementar-se els fluxos de nutrients que vénen de terra. Una intensa barreja vertical de l’aigua acaba generalment amb aquesta mena d’episodis.
Sovint es fa la distinció entre el plàncton nerític o costaner i l’oceànic o d’alta mar. Els marges dels continents interfereixen amb la dinàmica marina, especialment en les costes amb marees, i l’energia de la interacció produeix un treball de barreja d’aigua, amb la fertilització local que comporta. Aquesta condueix, moltes vegades, a proliferacions irregulars de diatomees que coincideixen amb els moments en què el vent o la mar mouen i mesclen intensament les aigues menys profundes. L’estudi de situacions d’aquesta mena ha ajudat a aclarir els mecanismes bàsics de la producció primària marina.
Sabem que les característiques del medi marí són molt variables, tant en cada volum d’aigua com en l’estil dels canvis. Existeix un gran nombre d’organismes diferents i molts fugen del comportament més generalitzat en el grup taxonòmic al qual pertanyen, cosa que fa difícil trobar regularitats de validesa general. Les diatomees es multipliquen ràpidament, però algunes espècies persisteixen en cultius contínuament il·luminats mentre que altres no prosperen si no estan subjectes a un ritme diari o circadiari en el qual s’alternin llum i foscor. També se sedimenten ràpidament i, en el seu model del plàncton atlàntic, Riley, Stommel i Bumpus usaven una velocitat de sedimentació per a les diatomees de 10-15 cm per hora a 0°C, i el doble d’aquesta velocitat a 25°C. Altres observacions diverses queden entre 1 i 50 cm per hora. Però espècies definides tenen recursos propis i s’escapen del que podria semblar una llei general. Diatomees oceàniques de gran volum contenen vacúols de fluid més lleuger que l’aigua marina, el manteniment del qual exigeix un treball metabòlic suplementari (per exemple, substituir cations metàl·lics per amoníac). Durant l’estiu, en aigües superficials, sobreviuen també diatomees que s’associen amb ciliats tintínnids. Aquests funcionen com a “motors fora borda” que les mantenen en suspensió (Chaetoceros, més rarament Planktoniella). Altres diatomees (Thalassiosira) excreten cordons de mucilag i fan masses que potser també romanen més bé en suspensió o, si més no, canvien la forma de vida habitual d’aquests organismes. Phaeocystis és un altre organisme les cèl·lules del qual estan sovint cobertes de mucilag.
Així doncs, la condició fonamental per a la producció del plàncton vegetal és la presència de nutrients en els nivells il·luminats, és a dir, una elevada coincidència (expressable per una covariància, C) en les distribucions de llum, nutrients i cèl·lules. El manteniment de la necessària energia exosomàtica per a seguir barrejant l’aigua és donat per una mesura de la difusivitat turbulenta (A). Combinant de les maneres possibles dues categories de cada un d’aquests descriptors es pot construir un gràfic amb quatre compartiments que ajuda a resumir diversos aspectes de la biologia del fitoplàncton.
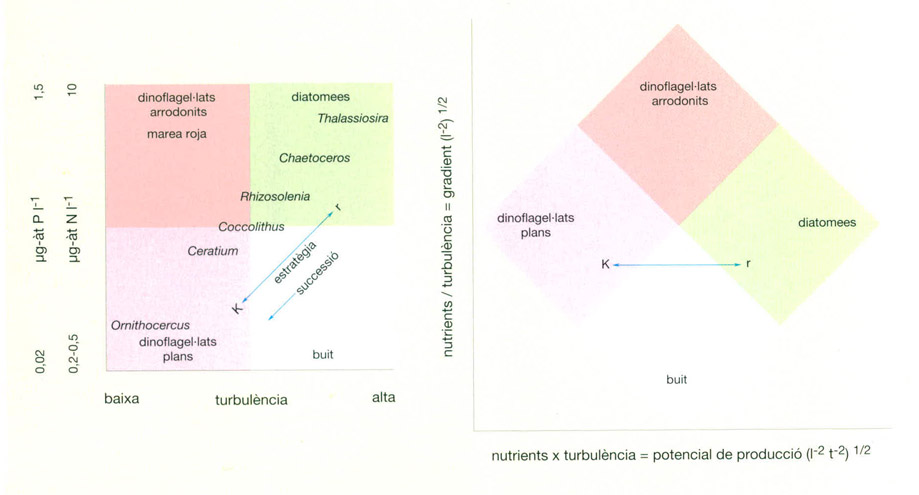
Representació segons diferents descriptors de les principals característiques del fitoplàncton (“mandales” del plàncton). En la seva primera versió, els descriptors són la presència de nutrients i la turbulència. Dues de les caselles, la de dalt a la dreta i la de baix a l’esquerra, corresponen respectivament a les etapes inicial i terminal de la successió normal, la primera caracteritzada per una turbulència important i una important persistència del nodriment a les capes il·luminades (com també per un predomini d’espècies oportunistes, amb una elevada taxa de reproducció), i la segona per baixa turbulència i exhauriment de nutrients (com també pel predomini d’espècies més especialitzades i persistents, amb baixa taxa de reproducció). El mateix encasellat permet encabir dues condicions més, que podrien semblar anormals, però que no són pas rares. La de baix a la dreta correspondria a la situació tipificada per les aigües molt agitades de primavera d’una mar nòrdica o antàrtica, pràcticament mancades de nutrients i d’organismes (efecte Gran-Braarud). La de dalt a l’esquerra es dóna en badies o altres aigües estratificades properes a les costes, on les aigües superficials reben nodriment suplementari, sovint d’origen continental; en aquestes situacions proliferen i s’acumulen els dinòfits nedadors i sovint tòxics de les marees roges. El mateix diagrama es pot girar 45° en el sentit de les agulles d’un rellotge, canviant les coordenades, que passen a ser respectivament el quocient nutrients/turbulència i el producte nutrients/urbulència, que aleshores corresponen més aviat a l’estratificació i a la productivitat, respectivament. L’eix de la successió (i del canvi del predomini d’estratègies de reproducció) discorreria en aquest cas de dreta a esquerra.
Jordi Corbera, a partir de dades de l’autor
El mateix diagrama es pot girar 45°, canviant les coordenades, que passen a ser respectivament el quocient C/A i el producte A·C, que aleshores corresponen millor a l’estratificació i a la productivitat. En diagrames independents es poden marcar les distàncies estadístiques entre diferents espècies, basades en la probabilitat de coincidència de les mateixes espècies en mostres d’aigua reals. Si una representació simplificada d’aquests tipus es deixa projectar de manera raonable sobre un diagrama C-A com l’esmentat, es reforça el significat d’aquesta mena de representacions, que algú proposà anomenar mandales del plàncton.