Les selves oceàniques
Els esculls coral·lins són un cas especial de bioherms o estructures de carbonat càlcic d’origen biològic, resistents a l’onatge. Formacions marines com aquestes s’originen dels esquelets cimentats d’organismes vius coneguts com a hermatípics, o formadors d’esculls, i de les seves restes mortals. Hi ha esculls construïts per diverses menes d’organismes hermatípics com ara les ostres, els gasteròpodes vermètids, els poliquets serpúlids, les algues del coral·ligen, particularment rodòfits, els hidrozous i els coralls. Aquests últims són, de molt, els més estesos, i formen els que probablement són els més diversos i acolorits dels ecosistemes marins, l’equivalent marí de la selva equatorial plujosa.
Els esculls coral·lins segurament representen el cim de la vida en el domini marí. A més, tots els altres esculls d’origen orgànic són força limitats en la seva distribució, mentre que els coral·lins dominen la biota bentònica virtualment a totes les regions costaneres d’aigües tropicals. En els esculls corallins, els coralls hermatípics formen tant el marc físic com la base tròfica de l’ecosistema sencer. Moltes vegades s’hi associen els anomenats coralls de foc (Millepora, Stylaster), que no són pròpiament coralls, sinó hidrozous.
Els tipus d’escull coral·lí
Els esculls coral·lins han estat agrupats a grans trets en tres tipus principals: esculls de franja, esculls de barrera i atols, si bé, per a certs estudis, aquests tres tipus encara són subdividits en diferents subtipus atenent a la seva morfologia i gènesi.
Els esculls de franja es troben molt a prop de terra, de vegades tan a prop que deixen amb prou feines una petita albufera de pocs metres d’amplada entre els esculls i la riba. Són força comuns, i apareixen en qualsevol lloc on comencen a desenvolupar-se esculls coral·lins.
Els esculls de barrera es troben molt més allunyats de terra, com en el cas de la Gran Barrera australiana, que és no tan sols l’escull coral·lí més gran del món, sinó potser l’estructura biològica més gran que existeix. La Gran Barrera australiana s’estén més de mil quilòmetres, vorejant la costa est d’Austràlia a una distància que excedeix en molts casos els cent quilòmetres, sense una veritable albufera o “lagoon” entre la costa continental i l’escull coral·lí.
L’últim grup, els atols o atol·lons, són els més singulars de tots els esculls. La seva forma és aproximadament circular i s’hi enclou una llacuna poc fonda (“lagoon”). Els diàmetres dels atols varien entre uns metres i setanta quilòmetres. Hi ha un centenar d’atols, setanta d’ells, aproximadament, a l’Indo-Pacífic i la resta a l’oceà Pacífic i el Carib. La forma regular i poc usual dels atols va cridar l’atenció de Darwin durant el seu viatge en el “Beagle” i el va portar a desenvolupar una teoria que n’explicava la forma, basada en la seva formació. Va suggerir que tots els atols s’haurien originat com illes volcàniques que haurien emergit del fons de l’oceà, privats de tota mena de vida a causa del seu origen igni. En arribar a la superfície, o molt a prop, esdevenien un substrat adequat per a ser colonitzats, majoritàriament, però no exclusivament, per organismes bentònics de propàguls planctònics. Els coralls tenen efectivament aquest tipus de larves planctòniques que viuen lliures —les seves plànules—, a més de disposar d’altres formes de multiplicació, que es detallaran més endavant. Aquests coralls i la fauna i flora associades es desenvoluparien fins a esdevenir esculls de franja, que encerclarien l’illa cònica volcànica. Els canvis subsegüents en l’altura de l’illa i el nivell de la mar, resultat bé d’increments del nivell de la mar, bé de subsidència de l’illa, menen a la desaparició de l’illa mateix, mentre que el creixement cap amunt del corall segueix el creixent nivell de la mar. Si l’índex del creixement cap amunt de l’escull coral·lí no s’adiu amb l’enfonsament de l’illa, l’escull està condemnat, i morirà. En els atols existents, no va ser aquest el cas, evidentment i, una vegada enfonsada l’illa, l’escull ha encerclat la llacuna on abans hi havia el volcà. La teoria de Darwin, proposada el 1842, finalment va ser verificada cent anys després, quan una prospecció de perforació a la llacuna de l’atol Eniwetok, feta específicament per provar aquesta teoria, va arribar a roca basàltica d’origen volcànic a una fondària de 1 400 m, sota una capa d’escull coral·lí fòssil.
La distribució geogràfica
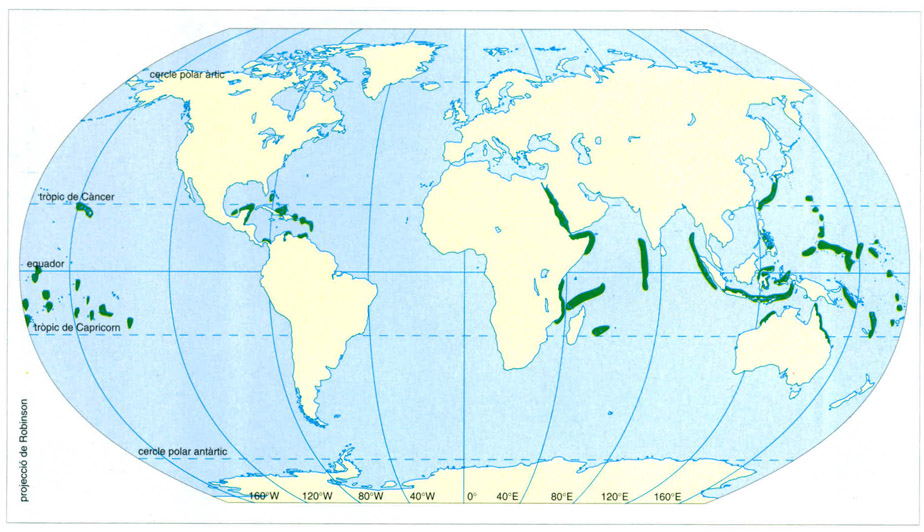
Distribució mundial dels esculls madrepòrics. La influència dels corrents freds a les façanes occidentals dels continents es fa ben palesa en la distribució dels madreporaris hermatípics, força més abundants a les façanes orientals.
Editrònica, a partir de fonts diverses
D’esculls coral·lins fòssils, se’n troben a molts llocs; molts d’ells actualment força allunyats de qualsevol mar. Els esculls més primerencs que apareixen al registre fòssil són de l’Ordovicià, fa quaranta milions d’anys. Es tracta d’esculls construïts per hermatips diferents als que coneixem en els esculls actuals, però com que en els fòssils que trobem gairebé mai no s’han conservat els teixits blans, no podem estar segurs de les afinitats taxonòmiques d’aquests constructors d’esculls primerencs. Tot i això, a partir únicament del que permeten inferir les restes dels seus esquelets, es considera avui que pertanyen a grups de coralls ja extingits: els rugosos i els tabulats. A partir del Juràssic podem trobar esculls coral·lins en els quals grups de coralls encara existents avui, i fins i tot gèneres i espècies idèntics als actuals, fan un paper d’importància creixent. El grup principal de coralls hermatípics dels esculls actuals són els escleractiniaris, també coneguts com a madreporaris, que apareixen més abundantment des del Cenozoic. De les 7 500 espècies de corall conegudes, unes 5 000 s’han extingit i 2 500 han sobreviscut i poden ser trobades als esculls actuals.
Els esculls de corall viu es troben principalment entre les latituds 30°N i 30°S, o sigui entre els tròpics de Càncer i de Capricorn. Tot i això, es troben esculls coral·lins molt lluny d’aquests límits actuals. Ja que la posició dels pols i també la de l’equador han canviat durant els temps geològics, quan ja existien els coralls i els esculls de corall, probablement aquests grans canvis de distribució dels esculls coral·lins són un resultat dels canvis concomitants en la distribució dels models de clima mundial durant el mateix període. Actualment, encara que algunes espècies de corall es troben fora dels límits descrits, no són hermatípics, ja que sobreviuen i creixen com a colònies aïllades que no es desenvolupen mai com a estructures d’esculls contínues i estables.
Les qüestions més importants sobre la distribució global dels esculls coral·lins són dues: quins són els factors ambientals que fixen els límits del desenvolupament i la supervivència dels esculls coral·lins i com es poden explicar els models actuals de diversitat de coralls.
Les condicions ambientals i la disponibilitat de nutrients
L’absència evident d’esculls coral·lins al bentos de les aigües costaneres immediatament al nord i al sud dels tròpics indica certament una relació amb la temperatura, però l’evidència experimental directa per provar aquest agent causal no és, malauradament, concloent. S’ha suggerit que les temperatures inferiors a 15°C impedeixen als coralls la captura eficient de preses i la reproducció. Tot i això, un cert nombre de coralls ha sobreviscut a exposicions d’unes hores a temperatures menors que aquestes. Sembla que, basant-nos en coneixements actuals, podem dir que el límit tèrmic crític per a la distribució d’esculls coral·lins es pot fixar en la isoterma mínima hivernal de 15°C. Fins ara no s’han trobat aigües marines que resultin massa càlides per a la formació d’esculls, però, tanmateix, sí que s’han acumulat evidències de perjudicis extensos i de la subsegüent mortalitat en esculls després de canvis de la circulació oceànica que comporten temperatures més elevades a la regió danyada. Aquests esdeveniments, durant els quals les temperatures oceàniques havien excedit entre 1 i 3°C, els valors mitjans d’aquella àrea i d’aquella estació van durar des d’unes setmanes fins a uns mesos. Basant-se en aquesta correlació i en alguns estudis de laboratori, s’ha suggerit que els esculls corallins poden ser registres a llarg termini de l’escalfament de les aigües oceàniques.
Tanmateix, hi pot haver efectes indirectes de temperatura sobre els coralls. En aigües tropicals tan sols hi ha una diferència mínima, si és que n’hi ha, entre les temperatures d’estiu i les d’hivern. La intensa insolació manté la tebior de la part superior de la columna d’aigua i, a causa de l’expansió termal, també la manté més lleugera que l’aigua profunda freda subjacent. Aquesta situació és anomenada estratificació tèrmica i és un factor permanent de les mars tropicals. L’estratificació impedeix la barreja vertical entre les aigües tèbies superiors i les que hi són subjacents. Els organismes del fitoplàncton, les algues microscòpiques que són els principals productors dels oceans, han d’extreure de l’aigua que els envolta els compostos i els nutrients necessaris per al seu creixement i la seva multiplicació. Ja que el fitoplàncton necessita, per a la fotosíntesi, llum i CO2, que és abundant en gairebé totes les situacions marines, són les aigües superiors i il·luminades les que s’han desproveït d’aquests nutrients.
Aquest tipus d’aigües pobres de nutrients, oligotròfiques, no poden sustentar el creixement de fitoplàncton abundant, cosa evident gairebé a simple vista, ja que aquest tipus d’aigües són extraordinàriament transparents i característicament blaves, color de les aigües marines netes, en comptes de tenir el color verdós i tèrbol típic de l’aigua “fèrtil”, rica en fitoplàncton. Aquestes aigües tropicals oligotròfiques s’anomenen “deserts blaus” i, atesa la migradesa de vida que contenen, es mereixen ben bé aquest nom. Com qualsevol ecosistema, els oceans també depenen de la fotosíntesi de les plantes com a única font d’energia i, per tant, si manca el fitoplàncton, les mars tropicals no poden sustentar vida abundant de cap altre grup que en depengui, directament o no, en particular el zooplàncton, els animals de vida lliure que pasturen el fitoplàncton, i tots els animals més grossos que en mengen, com ara els peixos petits, i els seus depredadors, i tots els carnívors marins més grossos, inclosos, encara que siguin animals terrestres, els humans.
Com veurem, és precisament aquesta manca de nutrients i de zooplàncton el que va menar a l’evolució de la simbiosi entre zooxantel·les i coralls, i al consegüent domini dels coralls en les aigües tropicals poc fondes. Tan sols en aquestes condicions especials d’aigües tèbies, oligotròfiques, transparents i pobres de fitoplàncton i zooplàncton, els coralls i els esculls coral·lins es troben en avantatge competitiu enfront d’altres comunitats bentòniques, especialment enfront de les algues. En medis marins, dins del “cinturó de corall” on hi ha afloraments, no hi ha esculls coral·lins. En aquestes regions, els corrents oceànics porten aigües riques de nutrients des de les profunditats cap a la superfície il·luminada. En aquestes zones l’aigua és tèrbola a causa dels enormes creixements de fitoplàncton, i al bentos, tan sols pot arribar-hi una mica de llum. A més, en aquestes aigües les algues es troben en clar avantatge competitiu sobre els coralls (els quals creixen molt lentament), ja que en una regió d’aflorament, amb abundants nutrients, les algues poden créixer i ofegar l’escull. La temperatura exerceix un efecte addicional, si bé indirecte, sobre la distribució dels coralls, en controlar la barreja vertical, o la manca d’aquesta, i la distribució de nutrients a la columna d’aigua.
Potser és en part a causa dels afloraments que, en general, no hi ha esculls coral·lins a les costes occidentals dels continents, mentre que abunden a les costes orientals. Aquest és el cas d’Austràlia, Àfrica i gran part d’Amèrica. Els grans sistemes fluvials també causen buits o interrupcions en els sistemes d’esculls, altrament ben continus. La reducció de la salinitat, la sedimentació i l’increment dels índexs de càrrega de nutrients pel corrent d’aigua terrestre poden ser, junts o per separat, les causes fonamentals d’aquestes interrupcions.
Fins i tot dins de la regió on la temperatura de l’aigua i els baixos nivells de nutrients afavoreixen la dominància dels esculls coral·lins del bentos de poca fondària, encara interactuen factors addicionals que són determinants per a decidir el resultat definitiu de la lluita pel substrat. L’abundància relativa d’herbívors pot inclinar la balança entre els coralls i les diferents comunitats d’algues.
La diversitat específica i el capteniment dels coralls
Una darrera gran qüestió en la zoogeografia dels coralls és la distribució poc homogènia de la diversitat d’espècies de coralls o, en termes menys rigorosos, la distribució d’espècies i gèneres de coralls. Hi ha diferències marcades entre el domini atlàntic i l’indo-pacífic. Això va dur a una controvèrsia sobre la qüestió de si la regió d’alta diversitat Indo-Pacífica és un centre d’evolució des de la qual les espècies de corall es van estendre i moltes simplement van desaparèixer pel camí, o si aquesta regió d’alta diversitat és un “refugi” on les espècies que van evolucionar en altres llocs s’han acumulat a causa de les condicions favorables, qüestió encara no resolta.
Hi ha una altra qüestió relacionada amb aquesta: la colonització per coralls de substrats adequats de creació recent, com ara illes volcàniques emergents, lluny d’esculls existents. Fa temps que se sap que les plànules de corall s’uneixen al plàncton i, com a tal, són transportades pels corrents marins. Però encara no se sap gaire de la capacitat d’aquestes larves per sobreviure a aquests viatges. Els estudis de laboratori han provat que les plànules d’algunes espècies de coralls poden sobreviure durant períodes de prop de tres mesos abans de fixar-se al substrat. Si durant aquest temps arriben a substrats adequats i inhabitats, poden esdevenir l’inici d’un escull naixent. El transport de plànules a distàncies de més de 1 000 km es poc probable, a causa de les velocitats dels corrents oceànics. Tanmateix, estudis recents han posat de manifest que fragments de pedra tosca flotants podrien rebre la implantació d’alguna plànula que hi comencés el desenvolupament d’una colònia coral·lina. Aquesta colònia, o potser colònies d’espècies diferents, podrien ser transportades durant mesos o anys, al llarg de milers de quilòmetres. Una vegada arribada a terra o enfonsada a causa del pes creixent podria esdevenir el començament d’un escull, o simplement afegir espècies a un de ja existent. Com que la pedra tosca procedeix de les erupcions de volcans marins o terrestres, i cada volcà i cada erupció tenen les seves empremtes característiques o composició química, és possible datar-ne cada tros. Basant-se en aquestes evidències, i recol·lectant coralls “raiers” a les platges d’illes del Pacífic, ha estat proposat fins i tot un model de colonització d’illes per coralls i esculls.
La disposició en fondària
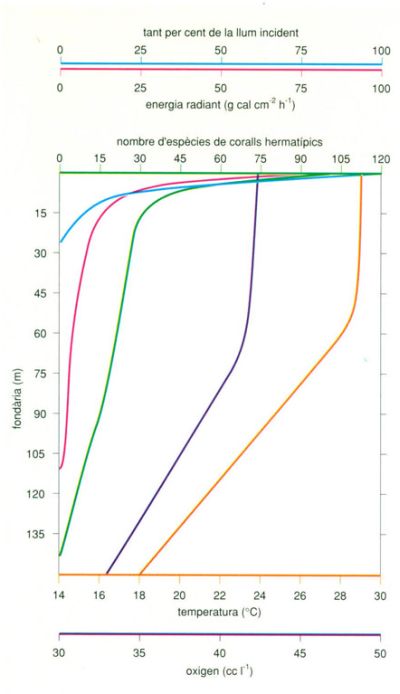
Distribució del nombre d’espècies de coralls hermatípics en relació amb la fondària i comparació amb la d’altres paràmetres ambientals. Els únics paràmetres que mostren models de distribució comparables són la llum i l’energia radiant, cosa que posa de relleu la dependència dels madreporaris (o més aviat de les zooxantel·les que hi viuen en simbiosi) respecte de la llum.
Editrònica, a partir de dades de Stoddart
A escala local, el factor que determina la distribució dels coralls i de cada escull és la fondària. Els esculls comencen pràcticament a la superfície de la mar, arriben al seu màxim desenvolupament en els primers metres superiors, i a poc a poc van decreixent a mesura que augmenta la fondària, tant en vigoria com en abundància d’espècies. Encara que algunes espècies coral·lines poden viure a profunditats de més de cent metres, com el corall Leptoseris fragilis, que viu en aigües profundes a la mar Roja, no són espècies hermatípiques, ja que no formen esculls. I espècies que normalment són hermatípiques, com per exemple, per no moure’ns de la mar Roja, Stylophora pistillata, que en aigües poc fondes és un element important en l’estructura de l’escull, cap al límit inferior de la seva distribució no en forma.
L’únic paràmetre ambiental que segueix un model batimètric similar és la llum. La llum s’atenua ràpidament en tots els medis aquàtics, amb un índex que depèn de la transparència de l’aigua. Altres paràmetres que se sap que afecten el creixement i la distribució dels esculls, o que s’ha suggerit que ho fan, com ara la temperatura, els nutrients i el zooplàncton, no mostren una relació profunda que podria explicar el que es pot observar als esculls en qualsevol lloc. La base d’aquesta dependència dels esculls en la llum s’explicarà detalladament més endavant, en altres seccions. De tota manera, el límit de profunditat per al desenvolupament dels esculls coral·lins depèn de la transparència de l’aigua, que és el paràmetre que determina l’extensió de la penetració de la llum a qualsevol profunditat. Aquesta correlació estreta entre la llum subaquàtica i el desenvolupament dels esculls és determinada, com veurem més endavant, per la capacitat fotosintètica dels simbionts algals dels coralls: les zooxantel·les.
L’ecosistema coral·lí
L’ecosistema dels esculls coral·lins té tots els paràmetres que defineixen un ecosistema madur en els termes de les ja clàssiques definicions de Margalef i d’Odum. Té una biomassa alta i constant i la respiració és gairebé igual a la productivitat, és a dir que els esculls de corall estan en un estat gairebé estable, en el qual la respiració consumeix tota la biomassa produïda i utilitza tota l’energia incorporada per fotosíntesi. La biodiversitat dels esculls de corall és alta: de fet, és de les més altes que podem trobar enlloc del nostre planeta, un tret considerat corol·lari de maduresa als ecosistemes.
La complexitat i les interrelacions
Quan sorgeix una massa de terra a causa d’una erupció volcànica en latituds adequades, comença un procés sense aturador de colonització de plànules de corall i propàguls de diversos organismes bentònics. Aquests organismes sèssils són seguits d’altres, també sèssils, i d’alguns de mòtils, que depenen dels primers per a aixoplugar-se i per a alimentar-se. Amb el temps, s’hi fixen més espècies, i la competència pel substrat i per l’energia disponibles redueix els recursos a l’abast de cada espècie. Espècies que practiquen l’estratègia de la r (descendència nombrosa, fàcilment dispersable, creixement ràpid, vida curta, etc.) o espècies oportunistes, són substituïdes per altres, altament especialitzades, que practiquen l’estratègia de la K. L’especialització acota el nínxol ecològic de cada espècie i redueix l’encavalcament entre uns i altres, la qual cosa resulta en un creixement del nombre d’espècies amb poblacions petites. Això minimitza la competència interspecífica i redueix les poblacions de cada espècie particular.
Com que són escassos els nutrients en les aigües circumdants i la majoria estan retinguts com a part de la biomassa viva, la producció de biomassa nova és limitada i regulada per la taxa d’alliberament de nutrients de l’escull. L’alliberament i la mineralització de nutrients, especialment el nitrogen i, en menor grau, el fòsfor, tenen lloc mercès a la mort dels diferents organismes de l’escull i l’acció de molts animals bioerosionadors, que perforen i dissolen l’esquelet calcari dels coralls de l’escull. Aquests organismes són fonamentalment les algues i les esponges perforadores, i també els percebes i les cloïsses que viuen entre els coralls.
A la majoria d’ecosistemes d’esculls de corall desenvolupats, la base energètica de l’ecosistema de l’escull prové de les zooxantel·les, les algues endosimbiòtiques que viuen dins de les cèl·lules del corall. A més d’aquests productors primaris, els esculls de corall donen aixopluc a moltes algues: algues cespitoses, humils i inconspícues; algues endolítiques, amagades dins l’esquelet del corall; rodofícies hermatípiques i calcàries del coral·ligen; clorofícies que es calcifiquen i moltes altres algues fixades a roques nues o a corall mort de l’escull. Els fons sorrencs propers de poca fondària estan coberts de fanerògames marines.
Tots aquests organismes fotosintetitzadors, en un medi amb un zooplàncton relativament escàs, contribueixen a l’alta productivitat primària global de l’escull. Aquests fotosintetitzadors, o productors, suporten a la llarga tots els múltiples consumidors de l’escull, incloent-hi els herbívors que mengen fanerògames i que mengen algues, els carnívors, els detritívors, i els descomponedors. Els herbívors, que pasturen i brostegen les algues i les fanerògames marines, inclouen els que s’alimenten per filtració, com els musclos i les cloïsses, el zooplàncton, i els eriçons de mar, gasteròpodes, peixos, tortugues i dugongs brostejadors.
Tots els depredadors depenen d’aquests herbívors o d’altres carnívors d’ordre més baix, per a l’alimentació. Entre els depredadors hem de comptar els coralls, les clavellines de mar, els poliquets i les rajades manta, que mengen zooplàncton, i els peixos planctívors. Als esculls habiten molts peixos depredadors com amfosos, morenes, peixos lleó, peixos escorpí, peixos pedra, peixos lloro, taurons, rajades i escurçanes, tots ells carnívors (segon nivell tròfic de l’escull, després dels productors primaris i dels herbívors). Als esmentats cal afegir molts invertebrats carnívors, com ara estrelles de mar, cargols i llimacs de mar, i molts crancs i gambes. El detritus o matèria particulada orgànica morta satisfà les necessitats d’encara més habitants dels esculls. Són detritívors prominents els cogombres de mar i també alguns eriçons de mar excavadors, que viuen al fons.
Les catàstrofes naturals
Com que els coralls i els ecosistemes dels esculls de corall han evolucionat durant llargs períodes de condicions estables, estan mal adaptats per a acarar pertorbacions catastròfiques. Com que són organismes marins que viuen en un medi de salinitat molt constant, els coralls són molt sensibles a qualsevol exposició a l’aigua dolça. Una forta tempesta sobre l’illa hawaiana d’Oahu va fer que es formés una capa d’aigua dolça d’1 m de gruix sobre els esculls de corall de la badia Kaneohe. Aquesta capa s’hi va quedar uns dies i va provocar la mort de gairebé tots els coralls d’aigües somes, cosa que va afectar especialment les parts més altes. També s’han descrit casos de mort de coralls a causa de les pluges durant la marea baixa en la Gran Barrera australiana.
A la mar Roja, els coralls gairebé mai no estan exposats, ni tan sols durant la marea baixa. Tot i això, la combinació d’una marea baixa amb vents del nord va comportar el 1970 una exposició d’una setmana a l’aire, incloses les hores de més calor de migdia. El resultat va ser la mort massiva de tots els coralls de la part superior de l’escull del golf d’Aqaba fins al punt que la pudor de la descomposició del corall va impregnar l’aire de tota la regió.
Els huracans també causen destrosses als esculls. Molts esculls com el de la badia Discovery, a Jamaica, i els esculls del Carib mexicà a Yucatán van ser colpejats molt durament el 1988 per l’huracà Gilbert, i encara ara no s’han recuperat. Les espècies ramificades, com la banya de cérvol (Acropora cervicornis), que abans de l’huracà dominaven els paisatges de l’escull, van ser les que van patir més. La causa va ser, no tan sols el fet que eren les més abundants, sinó també per la morfologia, que els exposa a energies molt altes a causa de la disposició de les branques que, sota condicions d’altes energies d’onada i velocitats del corrent, tendeixen a fer alçaprem sobre la seva base, més estreta i fràgil.
Val la pena de mencionar que l’efecte d’aquestes dues catàstrofes sobre els respectius sistemes d’escull i el consegüent curs de la recuperació són totalment diferents. El sistema de la mar Roja tenia, abans de la marea baixa, una diversitat d’espècies molt alta, al voltant d’unes cent espècies de coralls hermatípics, i la catàstrofe les va afectar totes en la mateixa mesura. Tot i això, una vegada despullada l’àrea, va començar la colonització per una espècie de corall oportunista, Stylophora pistillata, que ha esdevingut dominant, si bé atès el temps que ja ha passat des de la marea baixa del 1970, ha començat a decréixer, i la diversitat d’espècies augmenta gradualment. Al Carib, l’escull era un peu quasi monospecífic de banya de cérvol (Acropora cervicornis). Quan va ser eliminat, les espècies més petites i compactes que havien estat ofegades per l’ombra de les seves branques van començar a créixer. El resultat de l’huracà va ser, doncs, un augment de la diversitat d’espècies, en comptes d’una reducció com en el cas de la mar Roja. Amb el temps, les poblacions d’Acropora cervicornis es recuperen, i probablement tornaran a dominar.
La gran mortalitat per emblanquiment (pèrdua de les zooxantel·les dels coralls) ocorreguda recentment a nombrosos esculls també s’ha considerat una conseqüència de fenòmens naturals. Tanmateix hi ha evidències convincents que aquest descoloriment va ser el resultat d’increments regionals de la temperatura de l’aigua. Aquest escalfament oceànic, de prop de 2 a 3°C, podria haver estat lligat, al seu torn, amb un episodi d’oscil·lació d’El Niño del sud. Aquest és un canvi dels models de corrents de l’oceà Pacífic meridional, que alguns investigadors suggereixen que podria ser un resultat —o una prova— de l’escalfament global de la Terra.
Un cas interessant de danys naturals a un escull de corall s’ha vist l’hivern de 1992-93, a la mar Roja. Aquell any l’hivern va ser més llarg i més fred a la regió que cap que hagi estat registrat, i com a resultat, la barreja d’aigües va ser més fonda i va durar més temps del normal. Això va donar com a resultat la injecció d’una quantitat molt més gran de nutrients de les aigües fondes situades per sota de la termoclina cap a les aigües superfícials il·luminades. Aquest episodi d’“eutrofització natural” va tenir dues conseqüències: en les aigües normalment pobres de nutrients i cristal·lines del nord de la mar Roja es va desenvolupar un “bloom” de fitoplàncton que va limitar la llum que arribava als coralls. Aquest problema va ser agreujat encara més pel desenvolupament d’una coberta contínua d’algues, principalment Enteromorpha i Ulva, que van créixer fins al punt d’ofegar els esculls de corall. Les més perjudicades van ser les petites colònies d’espècies ramificades, com Stylophora pistillata, que van patir una pèrdua de fins al 40%.
En general, sembla que en la majoria de casos els sistemes d’esculls tenen la capacitat de recuperar-se després d’una pertorbació natural, encara que sigui majúscula. És l’exposició crònica a agents d’estrès subletals antropogènics allò que redueix la capacitat dels esculls per sobreposar-se als desastres naturals. Aquest va ser clarament el cas de la mar Roja, on després de la mortalitat de la marea baixa del 1970, els coralls d’aigües molt contaminades i visitades mai no van recuperar l’esplendor original. En canvi, els esculls remots en llocs poc o gens freqüentats són, avui dia, vint anys després de l’episodi, tan diversificats i robustos com abans.
Els organismos dels esculls coral·lins
Als esculls coral·lins no hi ha únicament coralls, és clar. Ben al contrari, com ja ha estat apuntat, la diversitat de les formes biològiques hi és enorme. Els coralls, tanmateix, en són els animals més emblemàtics. I potser també els vistosíssims peixos coral·lins. En tot cas, els invertebrats hi són molt abundants i diversos.
L’ecosistema dels esculls coral·lins, a més de ser construït per coralls que, és clar, també són invertebrats, és habitat per miríades d’altres animals de gairebé tots els grups que es troben en el reialme marí. Poden ser classificats segons el seu grau d’associació amb l’escull, el nivell tròfic que hi ocupen, segons si són constructors (hermatípics) o no, o segons si contribueixen al seu bon funcionament o si, al contrari, són bioerosionadors que ataquen l’estructura de l’escull. Els habitants invertebrats de l’escull podrien ser tractats fins i tot, és clar, d’acord amb les seves afinitats taxonòmiques. Qualsevol d’aquestes classificacions, a causa de l’immens nombre d’organismes implicats, va molt més enllà de l’abast d’aquesta obra; a més, a causa de l’emplaçament remot de molts dels esculls del món, un gran nombre de les espècies que els habita encara no han estat descobertes ni descrites. És clar que en molts casos, fins i tot en aquells organismes (relativament pocs) que han estat descrits, se sap poc de la seva biologia, cicle vital, nivell tròfic i col·locació dins de l’ecosistema dels esculls corallins. Ens limitarem, doncs, a alguns dels invertebrats més visibles o més importants dels esculls, i il·lustrarem amb ells alguns grups clau o les funcions que acompleixen en l’ecosistema.
Tanmateix, un cas especial a remarcar és el dels bacteris, els quals, tot i que apareixen en concentracions relativament baixes a les aigües oligotròfiques a tocar dels esculls, hi són presents, i sens dubte ajuden l’ecosistema de l’escull a extreure de l’aigua alguns nutrients, la baixa concentració dels quals els fa gairebé inaccessibles al fitoplàncton, als coralls, a les algues i fanerògames marines. Hi ha, sens dubte, poblacions bacterianes associades a la copiosa secreció mucosa dels coralls i a les funcions regeneratives de nutrients que ocorren als espais interns dels esculls i les colònies de coralls. El paper important que fa aquest grup amb prou feines s’ha començat a estudiar.
Els coral·laris o antozous
Els coralls, malgrat ser sèssils durant gairebé tota la seva vida, són animals. No tan sols, amb molt poques excepcions, no tenen capacitat de locomoció, sinó que les seves formes i els seus colors recorden els de les plantes i les flors. Aquesta és, probablement, la raó del nom donat al grup sencer quan van ser descoberts: antozous, que en grec vol dir “animals en forma de flor”. Els coralls són celenterats, és a dir, animals multicel·lulars de pla corporal simple, basat en una simetria radial. El seu cos es desenvolupa a partir de dues capes de cèl·lules: una externa, l’ectoderm, i una interna, l’endoderm. Aquestes dues capes invaginades formen com un tub de doble paret, obert per un extrem. Aquesta obertura, la boca, voltada de tentacles que serveixen per a capturar l’aliment, té les funcions tant d’ingestió de l’aliment com d’excreció dels residus. En la majoria dels celenterats (i entre ells els coralls) els tentacles són armats de nombroses cèl·lules urticants destinades a la captura i la immobilització de la presa.
Els tres grups principals de cnidaris es componen d’organismes aquàtics, predominantment marins. Són els escifozous, o meduses; els hidrozous, inclosos els coralls de foc i les hidres; i els antozous, dels quals els més coneguts són les anemones de mar i els coralls. Els celenterats tenen una alternança de generacions entre una fase sexual, com a meduses amb motilitat, i una fase sèssil, asexual, com a pòlips. La distinció entre els tres grups es basa en la importància relativa de les fases de medusa i de pòlip: en els escifozous predomina la fase medusa, tant en termes de mida com en durada del cicle vital; en els hidrozous predomina la fase pòlip, ja que la fase medusa és minúscula i de vida curta; i en els antozous la fase medusa no existeix.
Gairebé tots els coralls són colonials, i es desenvolupen a partir d’un òvul fertilitzat, fins a arribar a una mena de larva anomenada plànula, que és capaç de nedar lliurement. Una vegada trobat el substrat adequat, la plànula perd motilitat, es fixa, i evoluciona fins a la fase pòlip, que aleshores creix vegetativament fins a arribar a ser una colònia amb la forma típica de cada espècie, però que també es pot modificar per les condicions ambientals.
Dins dels coralls es distingeixen encara dos grups diferents: octocoral·laris i hexacoral·laris. Els membres del primer grup tenen vuit tentacles semblants a plomes, mentre que els del segon grup, els hexacoral·laris, tenen sis tentacles simples o un nombre múltiple de sis.
Els octocoral·laris o coralls tous
Els octocoral·laris, a l’inrevés dels hexacoral·laris, no tenen esquelets de carbonat càlcic i generalment no són (amb poques excepcions) hermatípics. Els seus elements esquelètics poden ser espícules individuals, característiques i flexibles, o esquelets durs i cornis, en algun cas usats en joieria, especialment el corall vermell (Corallium rubrum) i els coralls negres o antipataris (Antipathes). Tots aquests coralls, coneguts també com a coralls tous, són part important de cada escull, encara que no contribueixen a la seva arquitectura.
Tots ells s’alimenten probablement de zooplàncton, i molts tenen zooxantel·les simbiòtiques, com els hexacoral·laris. Alguns tenen formes i colors vistosos, i les espècies que no tenen zooxantel·les poden trobar-se a grans fondàries on creixen en forma de fines xarxes perpendicularment als corrents predominants, i s’alimenten filtrant el zooplàncton de l’aigua. Alguns membres d’aquest grup produeixen substàncies actives contra els depredadors que actualment són estudiades per diverses companyies farmacèutiques com a possibles fonts d’antibiòtics. Els diversos coralls tous competeixen amb els hexacoral·laris i altres organismes hermatípics pel substrat adequat on fixar-se. La majoria dels membres d’aquest grup es reprodueixen sexualment de diverses maneres. Els sexes poden estar separats o no i, en el cas més simple, els productes sexuals es deixen anar a l’aigua, on es realitza la fertilització. En moltes espècies, però, els òvuls es retenen en cavitats especials, que és on té lloc la fertilització. L’òvul fertilitzat evoluciona fins a ser una plànula que, quan és madura, es deixa anar a l’aigua.
Els hexacoral·laris o madrèpores
Els hexacoral·laris (i, en particular, els escleractiniaris) són, de molt, els constructors d’esculls més importants. Aquest grup disposa actualment d’unes 2 500 espècies, que varien de forma, color, longevitat i biologia. La unitat bàsica de construcció d’aquests coralls és la mateixa que es troba en els coralls tous: el pòlip. Aquest pòlip s’origina d’una plànula produïda sexualment que, una vegada fixada, comença a construir un esquelet adherit al substrat. Els substrats preferits són les roques netejades per abrasió pels peixos o pels eriçons de mar i els esquelets de coralls morts. Aquests esquelets, encara que finalment s’envolten de teixit animal viu, estan cimentats a les roques, i són, doncs, topològicament externs, a diferència del que passa amb els esquelets dels vertebrats, que són interns. A mesura que creix l’esquelet del corall, s’hi forma una depressió en forma de copa en la qual viu el pòlip. Aquesta depressió, anomenada calze, té particions disposades radialment, els septes, que corresponen a estructures de teixit animal, els mesenteris. Aquests, i els filaments mesentèrics que s’hi adhereixen, creen una cavitat del cos que es prolonga fins a la boca, la funció principal de la qual és digestiva. El zooplàncton que toca els tentacles causa una descàrrega de les cèl·lules urticants; aquestes injecten el seu verí que immobilitza o mata la presa, la qual és portada a la boca amb els tentacles, i digerida per l’acció de l’endoderm. Les restes de la presa sense digerir s’evacuen per la boca. En algunes espècies, els filaments mesentèrics es poden estendre per la boca a una distància considerable i s’usen contra colònies coral·lines competidores. En aquesta mena d’encontres, l’espècie menys “agressiva” acaba per ser digerida per aquests filaments mesentèrics, o danyada per l’acció de “tentacles escombra” especials. Interaccions com aquesta, i també la superposició, determinen la jerarquia de dominància entre espècies de corall en competència en les comunitats d’esculls.
Els pòlips individuals es divideixen quan arriben a una mida determinada, que és característica per a cada espècie. Aquests pòlips normalment mantenen contacte entre ells al si de la colònia en creixement mitjançant una continuïtat dels respectius teixits. La colònia es desenvolupa prenent una forma i assolint una grandària que són característiques per a cada espècie, encara que poden ser modificades fins a un cert punt com a resposta a factors ambientals, especialment a l’energia de les onades i a la intensitat i la direccionalitat de la llum subaquàtica. La forma, la mida i el model dels septes del calze, i també les distàncies i les disposicions dels calzes en la colònia són els trets més importants en què es basen la taxonomia i la identificació de les diferents espècies de corall.
En algunes espècies no hi ha pòlips individuals discrets, fet que duu a la coneguda morfologia de la colònia del cervell de mar (Platygyra meandrina). Alguns coralls són un sol pòlip, que pot arribar a mides de fins a 50 cm, com alguns del gènere Fungia. Les espècies de corall no sols difereixen en la morfologia de la colònia, ans també en la respectiva taxa de creixement que, mentre que en algunes de les espècies massives que creixen molt lentament no ultrapassa uns pocs mil·límetres per any, en algunes espècies ramificades de creixement ràpid pot superar els 10 cm per any.
Els coralls hexacoral·laris es reprodueixen sexualment de maneres similars a les descrites en els octocoral·laris. Sempre que la fertilització és externa, la coordinació de l’emissió d’òvuls i esperma és crítica. A la Gran Barrera australiana, totes les espècies fan la fresa durant tan sols unes nits, presumiblement sincronitzades per una fase de la lluna. En altres localitats no es coneixen freses en massa semblants, i la reproducció, fins i tot en espècies que en la Gran Barrera participen en les “orgies” de fresa en massa, es distribueix al llarg de molts mesos. És clar que els òvuls i l’esperma, rics de proteïnes, són una delícia per a tots els planctívors, que s’asseguren d’acudir als esculls coral·lins a l’època de la reproducció.
La plànula que resulta de la fertilització és amb prou feines visible a ull nu i és exposada als mateixos perills que la resta del plàncton marí. Pot nedar per moviment ciliar, encara que és transportada a grans distàncies pels corrents marins. Té boca i normalment és capaç d’alimentar-se durant la fase planctònica. Una vegada fixada, fa la metamorfosi a la fase pòlip, que comença a dipositar l’esquelet calcari en el substrat i, si sobreviu, es desenvoluparà fins a ser una colònia.
L’esquelet de carbonat càlcic dels coralls hermatípics, especialment els ramificats, es trenca fàcilment, com a resultat de tempestes, i també, cada cop més, d’accions humanes. Aquestes branques trencades, si cauen en un substrat i a una profunditat adequats, poden formar colònies noves. Aquest tipus de fragmentació és, en certes espècies i localitats, una manera important de reproducció. Totes les colònies filles d’una situació que s’origina per fragmentació d’una colònia única són, és clar, genèticament idèntiques. Aquest tipus de reproducció contribueix a la formació d’extenses situacions monospecífiques, mentre que la reproducció sexual dóna oportunitats d’incrementar el “pool” genètic en les poblacions, i també permet el transport a llarga distància d’espècies i el seu eventual establiment en regions noves.
Les zooxantel·les
En el cas únic de l’escull de corall, les algues, que són els principals productors primaris i que suporten amb la seva fotosíntesi la vida de l’ecosistema sencer, viuen dins de cèl·lules d’animals, concretament de cèl·lules dels coralls mateixos. Aquestes algues són organismes unicel·lulars que pertanyen al grup dels dinòfits o dinoflagel·lats, al qual pertanyen alguns dels components més importants del fitoplàncton. Però, mentre que els dinoflagel·lats que viuen lliures tenen cadascun dos flagels que els donen mobilitat, els que viuen com a simbionts endozoics dels coralls, les zooxantel·les, normalment són sèssils.
Aquestes algues microscòpiques tenen un diàmetre d’unes 10 µm i poden arribar a densitats de més d’un milió de cèl·lules per centímetre quadrat de corall. Les zooxantel·les viuen en una associació simbiòtica molt estreta amb els coralls, de la qual tots dos socis es beneficien de tal manera que permet a tots dos o a la seva associació, la colònia de corall zooxantèl·lica, de colonitzar i dominar hàbitats en els quals cada soci per ell mateix no podria reeixir. En la seva associació, són les zooxantel·les les que posen la seva capacitat fotosintetitzadora al servei de l’associació, proveint l’animal hoste de molècules orgàniques riques d’energia, produïdes per la fotosíntesi. En lloc de ser retinguts per l’alga, aquests compostos s’escapen de les cèl·lules algals i són posats a l’abast de l’hoste animal.
Es pensa que aquest procés, anomenat translocació, resulta d’una combinació de dos factors. Les zooxantel·les, a causa del baix nivell de nitrogen i fòsfor al seu abast, no poden usar tot el carboni que obtenen per fotosíntesi, i excreten al seu medi (el citoplasma de les cèl·lules del corall) compostos rics d’energia i pobres de nitrogen. Al seu torn, el corall, mitjançant un anomenat “factor hoste”, indueix un augment de l’excreció en les zooxantel·les. Els compostos translocats, especialment en aigües somes i ben il·luminades, són suficients per a proveir totes les necessitats tant del simbiont com de l’hoste, com també el creixement respectiu i la reproducció d’un i altre. A canvi, les zooxantel·les tenen accés als productes residuals metabòlics del corall, rics de nutrients.
Aquests productes, perjudicials per al corall, són aprofitats eficientment per les zooxantel·les. Així, en comptes de ser excretats i perdre’s per dilució en les aigües oceàniques circumdants, pobres de nutrients, poden ser reciclats i utilitzats totalment al si de la colònia coral·lina i la comunitat de l’escull. Així i tot, per més que la simbiosi entre coralls i les zooxantel·les assegura als primers una font d’energia en aigües pobres en zooplàncton, i minimitza les pèrdues de nutrients del sistema, aquesta relació no pot proveir els nutrients, altres que el carboni, que són necessaris per al creixement. Aquests encara s’han d’adquirir, bé dels nutrients dissolts a l’aigua, gairebé tots inorgànics, per part de les zooxantel·les, bé de la digestió de preses capturades pels pòlips. Atès que tant nutrients dissolts com preses a l’abast normalment escassegen als tròpics, els que hi ha, s’aprofiten àvidament i els nutrients d’una i altra procedència s’emmagatzemen per a un ús prolongat.
La densitat de població de les zooxantel·les normalment és estable per a cada espècie de corall i en condicions ambientals donades, i fluctua entre centenars de milers i uns quants milions de cèl·lules per centímetre quadrat de corall. Aquesta densitat de població es manté com un balanç entre la taxa de creixement de les zooxantel·les (que tendeix a incrementar la seva densitat), la del corall (que tendeix a reduir-la), la pèrdua de zooxantel·les que resulten expulsades de la colònia i possiblement també, encara que no s’ha pogut demostrar per ara, la digestió d’algunes zooxantel·les per part de l’hoste. Els principals factors que afecten el creixement de la població de zooxantel·les són la disponibilitat de nutrients i la intensitat lumínica. En experiments en què s’han incrementat deliberadament els nivells de nutrients, especialment nitrogen i fòsfor, el creixement de la població de zooxantel·les es descontrola i les algues, en comptes de translocar els productes de la fotosíntesi, els retenen i els usen per a créixer aprofitant els nutrients que ara abunden.
Situacions d’aquest tipus es produeixen quan l’activitat o la desídia humanes donen com a resultat augments en els nivells de nutrients a l’aigua, és a dir eutrofització. Increments de fins a cinc vegades el nombre de zooxantel·les en els teixits dels coralls es poden percebre visualment com un palès enfosquiment de les colònies. Aquest procés és perjudicial per als coralls, ja que la població creixent de zooxantel·les reté els productes de la fotosíntesi en comptes de translocar-los a l’hoste. Això sense comptar que la població més densa de zooxantel·les requereix, per a la fotosíntesi, una proporció de carboni més alta que la disponible. Això comporta un decreixement de l’activitat fotosintetitzadora de les zooxantel·les, del qual resulta una reducció encara més gran de la seva contribució al benestar del seu hoste.
Una altra expressió del col·lapse de la regulació de la població de zooxantel·les és el fenomen conegut com emblanquiment, terme que vol descriure la pèrdua de zooxantel·les dels coralls a l’aigua circumdant, la qual esdevé tèrbola i marronosa a causa de la presència d’un gran nombre d’algues, ara mòtils. Si la colònia afectada no és recolonitzada per zooxantel·les residuals que queden als teixits, la colònia morirà.
Els altres celenterats
A més dels mateixos hexacoral·laris, que són els constructors principals dels esculls, d’altres celenterats zooxantel·lats també fan una contribució menor a la formació d’aquests. D’aquests “falsos coralls”, l’orgue de mar (Tubipora musica), un octocoral·lari, i el corall de foc comú, l’hidrozou Millepora dichotoma, són els més prominents. Aquests últims formen una zona de Millepora en molts esculls de la mar Roja. Els coralls de foc tenen moltes cèl·lules urticants potents que usen per immobilitzar preses (i possiblement per defensar-se), que poden inflingir cremades força doloroses i de llarga durada a nedadors incauts.
Els esculls són habitats per moltes espècies d’anemones de mar com la gran Condylactis gigantea, que viu en una simbiosi mútua amb el peix pallasso Amphiprion bicinctus. Aquesta anemone també acull nombroses zooxantel·les a les seves cèl·lules. En fons tous al costat d’esculls coral·lins, en llacunes d’atols i al curiós “llac de les meduses” de Palau hi ha moltes meduses; algunes s’alimenten de zooplàncton i peixos petits, d’altres, com Cassiopea andromeda, també acull zooxantel·les i s’alimenten primordialment dels productes de la seva fotosíntesi. Aquesta medusa pren el sol en aigües poc fondes i assolellades, com cap altra medusa, en posició “panxa enlaire”, exposant al sol els seus tentacles plens de zooxantel·les.
Els poliquets
Un dels grups d’invertebrats més plens de color dels esculls coral·lins són els plomalls de mar, un grup de cucs que s’alimenten filtrant l’aigua i que, després d’una existència larval de vida lliure, fan la metamorfosi a adults sèssils. Aquests cucs adults viuen en un tub calcari encastat en l’esquelet del corall, que esdevé part de l’estructura de l’escull. Un plomall de brànquies disposat en espiral i en constant moviment acompleix alhora les funcions respiratòries i de captura d’aliment.
Algunes espècies, com Spirobranchus giganteus, mostren un sorprenent devessall de colors vius, sense cap relació amb el color de la colònia coral·lina hoste. S’ha suggerit que aquests colors realment protegeixen la població en conjunt, ja que els depredadors que són capaços d’associar un color determinat a la seva presa favorita no faran cas dels individus de color diferent. Aquests poliquets abunden força als esculls i poden assolir densitats de moltes desenes en la mateixa colònia coral·lina. La seva distribució indica una forta preferència pels coralls vius sobre qualsevol altre substrat. Potser un moviment dels cilis dels seus tentacles també crea petits corrents que porten preses a l’abast dels tentacles dels coralls. A més a més, tot aliment que capturen i paeixen aquests (i qualssevol altres) organismes filtradors, enriqueix de nutrients les aigües en la proximitat immediata dels esculls. Aquests nutrients deriven de preses que arriben d’aigües relativament allunyades, de manera que se sumen als disponibles per al metabolisme de les zooxantel·les, i per a la posterior aportació d’aquestes als coralls simbionts.
Els mol·luscs
Els mol·luscs inclouen animals tan coneguts com cloïsses, ostres, cargols, llimacs, calamars i pops. Molts d’ells tenen capacitat de secretar una closca calcària protectora.
Aquest fílum tan divers dóna representants a tots els tipus d’assentaments funcionals de l’escull i del seu sistema. N’hi ha que són organismes hermatípics integrats a l’escull, d’altres que són agents bioerosius que perforen els esquelets del corall. Hi ha espècies que acullen zooxantel·les, n’hi ha d’herbívors i de carnívors, de sèssils (ostres, cloïsses, alguns gasteròpodes) i també àgils nedadors (nudibranquis), de cos nu o dotats de closca consistent. Poden dissimular la seva presència amb colors esvaïts, o manifestar-la amb colors vius iridiscents.
Un dels bivalves més impressionants dels esculls coral·lins és la tridacna o cloïssa gegant (Tridacna magna). És dels mol·luscs més grossos, i arriba a tenir mides de més d’un metre i un pes de més de quatre-cents quilograms. Aquestes mides es troben als oceans Índic i Pacífic, però, a causa del temps molt llarg que cal per assolir-les i de la pesca intensiva, escassegen els exemplars d’aquestes dimensions. Des que les descobriren els mariners europeus, van esdevenir trofeus preuats per oferir a catedrals, esglésies i santuaris on feien el paper de fonts baptismals o de piques d’aigua beneita, i també un motiu ornamental força valorat de l’art barroc i el rococó. Pescades des d’antic per les poblacions locals per la seva carn i les seves petxines, actualment hom fa esforços seriosos per conrear-les a diferents centres de Palau i altres països de l’Indo-Pacífic. Es conreen tant per al consum humà com per a l’aprofitament de les petxines. Les destinades a l’alimentació humana es cullen a l’edat de 2 a 3 anys, mentre que les destinades a l’explotació de les closques resulten més valuoses com més velles i grosses són. Aquests bivalves també tenen zooxantel·les, encara que, a l’inrevés dels coralls, no són endocel·lulars, sinó que viuen en els fluids corporals. Les algues els proporcionen la major part de l’alimentació, i la resta l’obtenen de plàncton filtrat de l’aigua.
Altres mol·luscs sèssils comuns en els esculls són alguns gasteròpodes vermètids, com Dendropoma maxima que, després d’una fase larval planctònica en què neda lliurement, es fixa a l’escull i forma una closca irregular que s’estén fent com una xarxa sobre la superfície del corall. Obté l’aliment estenent a la superfície circumdant una mena d’ampla xarxa del mucus que secreta pel peu i que periòdicament ingereix, juntament amb el plàncton que hi hagi quedat atrapat. Les closques d’aquests vermètids, que arriben a fer diàmetres de 3 cm i llargàries de 50 cm, igual que les petxines de les tridacnes, esdevenen a la llarga part de l’escull.
Al contrari de les cloïsses gegants i dels vermètids, que esdevenen part de l’estructura calcària de l’escull, entre els mol·luscs hi ha espècies que realment afebleixen l’esquelet coral·lí, perforant-lo activament mitjançant la dissolució del carbonat càlcic de què és construït. Un exemple de bioerosionador com aquest és el diminut (de la mida d’un gra d’arròs) dàtil de mar Lithophaga lessepsiana, que viu als esquelets calcaris de coralls vius de diferents espècies, com per exemple Stylophora pistillata, el seu hoste favorit. Aquest dàtil de mar, com tots els seus parents, s’alimenta per la filtració de plàncton de l’aigua que circula pels seus sifons i per les seves brànquies. Altres bivalves perforadors prefereixen hostes diferents. Una colònia gran de Stylophora pistillata pot contenir centenars de Litophaga lessepsiana que, a la llarga, conduiran a la fragmentació i a la mort de la colònia. Tot i això, sembla que en colònies coral·lines sanes els petits corrents que porten aliment i oxigen als dàtils de mar són benèfiques per al corall, les zooxantel·les del qual també aprofiten àvidament tot l’amoni que excreta la cloïssa.
Un altre grup de mol·luscs perjudicials —molt més que els diminuts dàtils de mar de què hem parlat— és el d’aquells que, com els gasteròpodes del gènere Drupella, principalment D. cornus, s’alimenten activament del teixit viu del corall. Infestacions de poblacions coral·lívores com aquestes, que creixen explosivament, poden tenir efectes devastadors en comunitats senceres d’esculls. Molts altres cargols de vistós acoloriment brostegen les algues d’alguns esculls coral·lins o s’alimenten de diversos animals que viuen a l’escull mateix o al seu voltant. Entre aquests hi ha els cònids, principalment els del gènere Conus, que són verinosos. La seva ràdula s’ha desenvolupat fins a esdevenir una probòscide, altament especialitzada, comparable a una agulla hipodèrmica, que fan servir per injectar un verí mortal tant a preses com a atacants. Aquests cargols, d’una gran bellesa, han estat preuats pels col·leccionistes, la qual cosa n’ha determinat l’extinció en moltes localitats. El seu verí potent pot ser mortal fins i tot per als humans. Altres cargols d’escull més o menys vistosos, com les porcellanes (Cypraea), també són recol·lectats localment, principalment per vendre’ls en grans quantitats als turistes, sovint ben lluny de les localitats d’origen.
Altres mol·luscs molt visibles que viuen als esculls són els nudibranquis. Aquests gasteròpodes no tenen la protecció d’una closca dura i, en molts casos, es refien de ser verinosos o repugnants. Fan exhibició dels seus colors vius, que poden funcionar com a advertiment que recorda als depredadors potencials el gust que fan. Altres espècies combinen l’excreció de sabors desagradables amb un camuflatge perfecte, la qual cosa els permet d’harmonitzar sense falla amb els esculls circumdants.
Els equinoderms
Els equinoderms són un fílum exclusivament marí, i moltes de les seves espècies tenen representació a les comunitats d’esculls coral·lins. Gairebé tots els membres d’aquest fílum comencen la seva vida com a larves planctòniques després d’una fertilització externa. D’adults mostren un pla corporal extern basat en una simetria radial característica. La seva locomoció és per mitjà de “peus” contràctils especials, característics dels equinoderms, anomenats peus ambulacrals. Aquests peus consisteixen en una combinació d’un sistema combinat hidràulico-muscular, cadascun d’ells mogut per un bulb que força l’entrada d’aigua al peu mentre els músculs d’aquest en determinen la direcció de l’extensió. Els peus acaben en una punta amb un disc de succió que permet a l’animal d’agafar-se a les superfícies i moure’s, o aixecar i moure objectes petits i aliments. Aquest sistema únic aquo-vascular és el mitjà de transport d’estrelles de mar, cogombres de mar i garotes. Els equinoderms principals que viuen als esculls pertanyen a totes les classes principals del fílum: crinoïdeus o clavellines de mar, cogombres de mar, estrelles de mar, ofiures i garotes o eriçons de mar, cap dels quals no és hermatípic. En realitat, tant la corona d’espines (Acanthaster planci) com alguns eriçons de mar del gènere Diadema, alguns dels pitjors enemics de l’escull, són membres d’aquest fílum.
Les clavellines i les holotúries
Les clavellines de mar o crinoïdeus articulats (Antedon, Leptometra), acolorides i delicades, figuren entre les més belles visions nocturnes de l’escull. Tenen cinc braços, cadascun subdividit en dos prop de la base. Aquests animals, que s’alimenten de plàncton, remenen les aigües de l’escull amb els deu braços com plomes, filtrant-ne criatures minúscules que fan anar cap a la boca, dirigida cap amunt. S’agafen al substrat de l’escull amb cirrus com pinces, que també els permeten de desplaçar-se des de les clivelles on s’amaguen durant el dia cap als seus emplaçaments nocturns de cara a mar oberta. Aquest comportament sembla adreçat a evitar els peixos depredadors diürns, i alhora a capitalitzar l’ascensió vertical nocturna del zooplàncton cap a l’escull, relativament menys profund.
En els fons sorrencs del voltant dels esculls, derivats en part de l’erosió de l’escull mateix, es poden veure molts cogombres de mar (Stichopus, Thelenota, Holothuria), alguns dels quals poden arribar a fer 1,5 m de llargària, arrossegant-se pel fons tou, ingerint la sorra o el llot i digerint-ne qualsevol matèria orgànica que hi hagi. Com que s’arrosseguen en la direcció de la seva boca tentaculada, el seu pla corporal s’ha elongat, i algunes espècies fins i tot han desenvolupat una simetria bilateral secundària. Algunes es protegeixen amb verins potents, mentre que altres responen als possibles atacs de maneres força inusuals. N’hi ha que expulsen les vísceres, que deixen a mans de l’hipotètic predador, ja que poden regenerar-les subseqüentment. D’altres enreden els enemics en xarxes de filaments llefiscosos que es formen quan uns òrgans específics, els anomenats tubs de Cuvier, són expel·lits per l’anus i, en trencar-se, deixen anar un líquid proteic que es coagula en contacte amb l’aigua. Algunes espècies de cogombre de mar són un veritable requisit gastronòmic a l’Àsia oriental i a les illes del Pacífic. En la seva preparació cal prendre precaucions especials per a evitar-ne els efectes tòxics.
Les estrelles de mar
Les estrelles de mar (Acanthaster, Protoreaster, Culcita, Linckia) són habitants comuns dels esculls coral·lins. Aquests equinoderms tenen una simetria radial característica, i normalment cinc braços, encara que algunes espècies en tenen múltiples, fins a quaranta, o bé no en tenen cap i adopten formes sensiblement pentagonals amb els costats lleugerament apuntats o convexos. Estan envoltades per plaques protectores, i es mouen, com els eriçons i els cogombres de mar, per mitjà de peus ambulacrals. Les estrelles de mar són carnívors actius que devoren animals sèssils o de moviment lent, els quals són ingerits o digerits per l’estómac evertit de l’estrella, que els envolta parcialment. Moltes estrelles de mar mengen cloïsses i musclos que obren lentament, fent força amb els seus peus ambulacrals, fins que la cloïssa es cansa i s’obre.
Atesa la devastació d’enormes extensions d’esculls, especialment a la Gran Barrera australiana, ocasionada per la corona d’espines (Acanthaster planci) i la preocupació mundial per la supervivència mateixa dels esculls, en parlarem amb una mica més de detall. La corona d’espines és una estrella de mar grossa que pot arribar a fer diàmetres de fins a 75 cm en trenta anys. L’adult té 16 o 17 braços, dotats de fortes espines verinoses, i es regenera fins i tot de fragments de braços, la qual cosa la fa pràcticament immortal. Una femella adulta pot deixar anar milions d’òvuls en el decurs d’unes setmanes cada estiu. Els òvuls fecundats es transformen al cap de 2 a 4 setmanes en larves planctòniques fins que arriben als esculls, s’hi fixen, i comencen la metamorfosi. Els joves s’alimenten uns mesos de plàncton i després canvien de dieta i comencen a consumir el seu aliment bàsic: els coralls. La corona d’espines és un coral·lívor consumat, amb una impressionant sèrie d’enzims específics per a digerir les ceres dels coralls. Les seves preferències es decanten pels coralls ramificats, encara que en condicions epidèmiques ataca qualsevol corall. En condicions normals la densitat de població de l’estrella és molt baixa, i a les aigües indo-pacífiques occidentals, on no s’ha observat cap destrucció, es va estimar que tan sols hi havia 6 animals per quilòmetre quadrat. Quan hi ha una epidèmia poblacional, els nombres augmenten explosivament de 3 a 4 ordres de magnitud, i arriben fins a 15 000 individus per quilòmetre quadrat. Si els adults despullen prop d’1 m2 d’escull al dia, un escull de la Gran Barrera australiana de 10 km2 resultaria destruït en uns dos anys.
Les primeres epidèmies de corona d’espines acuradament documentades van ser el 1957 a les illes Ryukyu, al sud del Japó. Els anys següents, a partir del 1960, es van documentar nombroses epidèmies a la Gran Barrera australiana, a les illes Marianes, Carolines i Marshall, a Nova Bretanya, a Malàisia, a les Fiji, a les Filipines, a Samoa occidental, a Tahití, a Hawaii i a Sri Lanka. A la Gran Barrera australiana, l’impacte destructor va ser tan greu que el govern australià va nomenar una comissió especial per estudiar i, si fos possible, posar remei al problema. Segons la versió més acceptada, els episodis recents d’epidèmies de corona d’espines resulten de l’acció humana. Les causes suggerides serien, bé la contaminació marina tan estesa, que reduiria la resistència dels coralls i del sistema d’esculls, bé la reducció de les poblacions d’enemics naturals d’Acanthaster planci. Un exemple d’aquest tipus d’enemics és el cargol gegant Charonia tritonis, que s’alimenta àvidament de corona d’espines, i que actualment és a frec de l’extinció a causa de la intensificació de les captures, tant per al consum humà com per al tràfic de records turístics. En el cas d’altres enemics especialitzats de la corona d’espines, com els peixos Arothron hispidus, un tetraodòntid, i Cheilinus undulatus, un làbrid, també se suposa una implicació humana en la reducció de la densitat de les seves poblacions.
Les garotes
Un altre grup d’equinoderms que es troba als esculls coral·lins és el de les garotes o eriçons de mar. Aquests tenen una gran closca calcària formada per un seguit de plaques que tanca dins seu la totalitat dels òrgans interns, coberta d’espines mòbils característiques, diferents per a cada espècie. Aquestes espines van des de les d’Heterocentrotus mammillatus, massives i de més de 10 cm de llargària i 1 cm de gruix, que s’usen per fer mòbils, sonalls de porta i elements de joieria, passant per les agulles primes i afuades de Diadema setosum i altres congèneres, de 20 cm de llargària, les espines de molts eriçons d’escull, amb 1 o 2 cm de llarg per 2 o 3 mm de gruix (Echinometra), fins a algunes que han estat reduïdes a la coberta vellutada i pilosa que es troba en alguns pobladors dels fons arenosos dels esculls com ara els anomenats “sand dollars” o “lochas de playa” (Echinarachnius, Mellita) i altres espècies que s’encauen.
Molts eriçons tenen, a més de les espines i els apèndixs ambulacrals, pedicel·laris, uns apèndixs especials que també són part del sistema ambulacral, coronats, a la seva punta, amb unes valves triples que formen una pinça i que, en algunes espècies, s’usen per netejar les espines, mentre que en d’altres, com per exemple Asthenosoma varium, estan equipades amb potents glàndules verinoses. Les garotes tenen una boca amb cinc dents, situada a la seva cara inferior, que utilitzen per rascar la superfície viva de l’escull, que és formada majoritàriament d’algues, però que també pot incloure diversos animals bentònics, com ara coralls joves. Com tots els equinoderms, els eriçons de mar tenen fertilització externa i larves planctòniques.
Pel seu paper erosionador i empobridor dels esculls coral·lins, són especialment importants els eriçons del gènere Diadema. Si es trepitgen accidentalment o es toquen sense cura, aquests eriçons de mar de color morat negrós infligeixen, amb les espines afuades del gruix d’un cabell, ferides doloroses que triguen molt a guarir. Aquestes garotes brostegen en grans ramats per les bases dels esculls i grups de coralls, i erosionen tant coralls vius com morts. El resultat visible de la seva activitat ha estat anomenat “aurèola de Randall” i a la llarga condueix a la fragmentació i la mort de coralls sencers. La majoria dels altres eriçons de mar també són bioerosionadors de corall, ja que s’alimenten brostejant el corall viu. També debiliten l’estructura dels esculls perquè fan forats i clivelles on s’amaguen durant el dia.
Aquests equinoderms serveixen d’aliment a nombrosos peixos que han trobat maneres enginyoses de superar les seves defenses. Els peixos ballesta (Balistes) projecten per la boca un fort corrent d’aigua que capgira els eriçons de mar, en deixa exposades les àrees del voltant de la boca que no tenen espines, per les quals és més fàcil que el peix obri la closca. Fet això, molts altres peixos s’ajunten al festí sobre la víctima indefensa.
Les ofiures
Els últims representants, i els menys conspicus, de la classe dels equinoderms comuns als esculls són els ofiuroïdeus. Són, en canvi, els més àgils de tots ells, gràcies als cinc braços llargs i flexibles que els impulsen. S’alimenten principalment de detritus, encara que es pensa que un grup, els frinofiúrids, que tenen els cinc braços primaris profusament ramificats, poden filtrar plàncton. Aquests darrers són habitants força conspicus dels esculls a la nit. Durant el dia s’amaguen en clivelles fondes de les roques, però, del vespre fins a l’alba, fan exhibició de la seva espectacular randa de braços entortolligats en espiral de 70 cm de diàmetre.
Els crustacis
Hi ha nombrosos representants als esculls de corall d’aquest important grup d’artròpodes. Alguns simplement afegeixen notes de color a l’exhibició enlluernadora dels esculls, mentre que d’altres fan papers importants en el sistema. Molts crustacis petits, com també les fases larvals de moltes espècies grosses, formen part del zooplàncton que és, en graus diversos, l’aliment de l’escull. Les nombroses espècies de coralls, lliris de mar, peixos, bivalves i molts altres planctívors del sistema d’esculls, tant si es tracta de filtradors com si són caçadors actius de zooplàncton, fan de l’escull un mur de boques a l’aguait del zooplàncton, constituït majoritàriament de crustacis.
Tal com passa amb els crustacis planctònics, els cirrípedes, el grup dels percebes i els glans de mar, no estan associats únicament amb els esculls coral·lins. Són habitants comuns a l’intermareal de tota mena de costes rocoses en gairebé totes les mars, i també habiten molts altres substrats, vius i inanimats, des de vaixells i molls fins a tortugues i balenes. A més dels glans de mar que cobreixen costes rocoses pelades al costat d’esculls, d’altres prosperen sobre corall mort o en soques de manglars propers. Són d’especial interès els que es fixen en l’esquelet del corall i a la llarga s’hi encasten. Aquest grup és format primordialment per espècies cadascuna de les quals viu en una sola espècie de corall, o en espècies molt pròximes.
No s’entén encara com és que les larves planctòniques poden arribar a trencar les barreres formidables de les cèl·lules urticants de coralls i hidrocoral·laris, i a penetrar al teixit de l’hoste i fixar-se al seu esquelet. Tot i que els glans de mar, en alguns casos, deformen la colònia, cosa especialment palesa en el cas de Savignum milleporae sobre el corall de foc (Millepora dichotoma), pot ser que tant el gla de mar com el corall es beneficiïn de l’associació. Com en tantes simbiosis mútues trobades als esculls de corall, el corall dóna el substrat i l’aixopluc, mentre que el gla de mar, que crea microcorrents amb els cirrus, captura i digereix el zooplàncton. Els subproductes metabòlics d’aquesta digestió enriqueixen el micromedi de la colònia de corall. Els nutrients excretats pel gla de mar són assimilats per les zooxantel·les, i posteriorment es faciliten al corall hoste en el corrent de translocació abans esmentat. Tots els percebes són hermafrodites i, com que la fertilització és interna, s’han de fixar molt acostats entre ells perquè els gàmetes es puguin trobar. Quan dels ous fecundats en neixen les larves, aquestes s’incorporen al plàncton i més tard es fixen al substrat, normalment al costat d’individus de la seva mateixa espècie, els quals són capaços de detectar pels compostos específics que excreten en baixíssimes concentracions.
Una altra associació peculiar entre crustacis i corall és la del “gall crab” o cranc de gal·la (Hapalocarcinus). La femella d’aquest crustaci es fixa a l’enforcament d’una ramificació d’un corall, el qual, a causa de la irritació que experimenta, es modifica fins a incloure el cranc en una mena de gal·la. El zooplàncton del qual s’alimenta i l’oxigen li arriben per les obertures que queden a la seva “presó”. La fertilització i l’alliberament de les larves també es fan per aquests porus.
Una estranya associació simbiòtica mútua que dóna protecció als coralls contra la corona d’espines (Acanthaster plancii) és la d’alguns crancs, la majoria del gènere Trapezia, amb coralls ramificats pocil·lopòrids (del gènere Styllophora, per exemple). Aquests crancs diminuts (2 a 3 cm) viuen encauats entre les branques dels coralls, on troben aixopluc i aliment, i els indueixen a secretar una mucosa, que arrepleguen amb les seves potes modificades. Quan el corall és atacat per una corona d’espines, els crancs surten dels caus i s’enfilen branques amunt i estenen les pinces per ferir i allunyar l’atacant i tallar-li les espines. S’ha demostrat que, si s’eliminen experimentalment els crancs, la colònia de corall queda indefensa i és probable que sucumbeixi a un atac de corona d’espines.
Altres exemples impressionants d’aquest tipus d’associació són les delicades gambetes netejadores del gènere Stenopus, que romanen amagades a les clivelles de l’escull i sota els coralls, sense mostrar més que les fines i llargues antenes que anuncien als peixos els “serveis” que els poden oferir: bo i alimentant-se d’ectoparàsits i trossets de pell desenganxada dels peixos que atreuen, ajuden a mantenir la salut dels seus “clients”. Com veurem més endavant, hi ha altres peixos que es guanyen la vida d’una manera semblant.
Hi ha espècies de gambetes que només es poden trobar sobre hostes o associats específics o al costat d’ells. Per exemple, la vellutada anemone de mar Cryptodendrum adhesivum allotja dues espècies de gambes delicades, gairebé transparents, que viuen a la seva superfície, dels gèneres Periclimenes i Thor. La naturalesa de l’associació no és gens clara, però podria ser que les gambes guanyessin protecció i s’alimentessin d’una mucosa que excreta l’anemone. En oferir alimentació experimentalment a l’anemone, un cranc fort i robust, de la família dels xàntids, repetidament va forçar la boca de l’anemone per robar-li la delícia a penes ingerida. Aquesta relació es coneix com cleptoparasitisme, parasitisme de robatori.
En fons plans al costat d’esculls, es troben caus on conviuen parelles de gambes i gòbit. Les gambes tenen pinces fortes, com pales, que usen per cavar el cau comú, però són cegues, i es refien de la visió dels seus peixos associats per prevenir-se de perills i buscar aliment, que després es reparteixen peix i gamba. Les gambes tenen antenes llargues i primes amb les quals perceben els moviments del peix. Aquestes relacions involucren diverses espècies de gòbits i gambes en diferents regions i zones d’esculls.
Encara hi ha un altre grup de crustacis, els crancs ermitans, presents a gairebé totes les mars del món. Aquests crancs tenen un abdomen tou, que per a molts dels depredadors famolencs dels esculls és una veritable temptació. Aquesta part vulnerable de la seva anatomia és la que els ermitans amaguen en conques buides de cargols morts. A mesura que va creixent el cranc, es muda a una “casa nova” més gran. Un ermità molt gros dels esculls, Dardanus, no es refia d’aquesta protecció i col·lecciona fins a una dotzena d’anemones molt urticants, que trasplanta a la seva closca, un sistema de defensa realment formidable.
A molts esculls es poden trobar diverses espècies de llagostes (Panulirus japonicus, P. longipes, P. ornatus, P. versicolor, etc.), que normalment s’amaguen en escletxes fosques i fondes durant el dia, i surten a l’exterior de l’escull a la nit, per alimentar-se. Aquests crustacis són molt valorats arreu com una menja deliciosa. L’expansió del comerç mundial i els turistes gastrònoms que arriben als esculls més remots ha tingut com a resultat la sobreexplotació mundial de la pesca de totes les espècies de llagosta.
Els peixos
Els peixos són un dels grups més visibles, abundants i rics en espècies, i particularment importants per a l’ecologia de l’escull, i també per a l’economia de les societats humanes que viuen prop dels esculls. Els milers d’espècies de peixos dels esculls coral·lins van des de les que depenen totalment de l’escull fins a les que hi tenen una interacció més aviat escassa o ocasional.
El comportament reproductor dels peixos d’escull abasta tots els models coneguts en aquests animals. Tot i això, es poden fer algunes generalitzacions. Molts peixos d’escull inverteixen un gran esforç en la cura dels ous, posant-los en llocs protegits, com els peixos pallasso (Amphiprion), que els fixen a superfícies rere l’anemone de mar dins de la qual viuen. D’altres, com els peixos ballesta (Balistes, Canthidermis, Melichthys), fan nius on la femella pon els ous, els quals es fertilitzen en un ritual complex; després, el niu es protegeix de depredadors i els pares el mantenen oxigenat dirigint amb la boca raigs d’aigua als ous fins a l’eclosió. L’ampla difusió d’aquest model reproductor en els esculls sembla concordar amb la teoria ecològica segons la qual en els ecosistemes madurs, com els esculls coral·lins i les selves plujoses tropicals, moltes espècies mostrarien estratègies de la K (vida llarga, pocs descendents, important inversió paternal en la cura de la descendència, etc.).
Moltes espècies de peixos d’escull canvien de sexe en el decurs de la seva vida. Per exemple, el peix daurat de la mar Roja (Anthias squamipinnis) comença la vida com a femella en una mola que viu als voltants d’un massís de corall. Un banc com aquest pot ser format per desenes o fins i tot centenars de femelles. Planen damunt seu uns quants mascles que tenen una coloració distintiva molt diferent, adornats amb apèndixs de remolc: les aletes dorsal i caudal. Com que els mascles estan més exposats que les femelles pel fet de viure més amunt, fora de la mola i de la seguretat relativa que aquesta dóna, és molt més probable que siguin devorats per depredadors de l’escull (neros, per exemple). Quan desapareix un mascle, la femella més grossa i dominant, canvia de sexe i es torna mascle. El potencial de canvi de sexe es manté mercès a la presència de gònades sense desenvolupar del sexe oposat. El predomini de les gònades d’un o altre sexe és induït pels canvis socials o ambientals per via hormonal.
Altres peixos d’escull, incloent-hi moltes espècies de neros (Epinephelus, Mycteroperca) i també algunes de peixos papallona i peixos pallasso (Chaetodon, Amphiprion), viuen en parella, que pot durar, bé un cicle reproductor estacional, bé, com en el cas dels neros, tota la vida.
Els menjadors de corall: peixos lloro i peixos papallona
Els dos grups més importants de peixos menjadors de corall són els escàrids o peixos lloro i els quetodòntids o peixos papallona.
Els abundants peixos, típicament robustos, de la família dels escàrids, anomenats genèricament peixos lloro (Scarus, Scarops, Bolbometopon, Sparisoma, Cryptotomus, etc.) per la característica forma de la boca i el seu vistós acoloriment, són de dimensions que van des dels 10 cm fins a 1 m; hi ha des d’espècies que viuen en parella fins a espècies gregàries que formen grans grups, amb colors que van des del negre i el marró apagat fins a un arc de Sant Martí de blaus brillants, morats, verds i grocs. Tots els membres de la família tenen les dents de les mandíbules superior i inferior fusionades en un bec de lloro fort i poderós amb què roseguen el corall, del qual es mengen el teixit i l’esquelet i hi deixen uns senyals característics. També mengen algues, que arrenquen del substrat. A la nit dormen dins de clivelles de l’escull, embolcallats en un capoll de mucosa que secreta la seva pell.
Els quetodòntids, encertadament qualificats de peixos papallona (Chaetodon, Chelmon, Heniochus, Forcipiger, etc.), són un grup de peixos molt vistosos i acolorits, subel·líptics, comprimits lateralment, els adults dels quals fan de 10 a 20 cm. La seva anatomia és molt adequada per a maniobrar amb precisió entre els coralls i els passadissos estrets dels esculls, i les seves dents abrasives (el nom de quetodòntids significa dents de cerra) els serveixen per a rosegar el teixit del corall i d’altres organismes bentònics.
Els menjadors d’algues: peixos barber
Entre els peixos que viuen en moles que neden per les cares dels esculls que miren a mar oberta, són freqüents les espècies de peixos barber (Acanthurus, Zebrasoma, Prionurus, Axinurus, etc.). El nom prové de les espines, afilades com navalles, que hi ha en el peduncle que connecta el cos i la cua. S’alimenten d’algues, que paeixen amb l’ajuda d’un grup de protists recentment descoberts, que viuen al seu sistema digestiu. Hom pot veure els peixos barber, cap a la posta de sol, nedant a corrua feta des d’arreu de l’escull fins a convergir en un punt de trobada. Quan hi arriben, la mola de milers d’individus comença lentament a girar formant una massa cònica. Com més s’incrementa la densitat, més de pressa neden, el remolí esdevé un deliri i els peixos de més amunt surten de la superfície de l’aigua repetidament. Uns quants mascles envolten una femella a punt de fresa i simultàniament es deixen anar òvuls i esperma. L’aigua es torna tèrbola de gàmetes, cosa que fa feliços els molts planctonívors que esperen el seu torn. La pau i la calma tornen aviat a l’escena, però l’orgia es repeteix cada dia durant uns tres mesos.
Els grans depredadors: taurons i rajades
Cap discussió sobre peixos d’escull de corall no seria completa sense una referència als taurons i els seus parents, les rajades. Els taurons i les rajades no són, de cap manera, propis únicament dels esculls. Tot i això, algunes espècies de taurons sí que en són exclusives, i circulen per les aigües del costat de mar oberta, i atemoreixen peixos, nedadors i cabussadors; altres, simplement les visiten (Carcharinus leucas, Negaprion brevirostris, Gynglimostoma cirratum, Triaenodon obesus, Sphyrna zygaena, etc.). La majoria de taurons no s’aventuren més enllà de la cresta de l’escull si no és amb marea alta, i fins i tot això, només ho fan espècies petites, com el tauró de punta negra (Carcharhinus limbatus) i individus joves d’altres espècies. Els d’espècies més grans i més ràpides recorren els costats de mar oberta d’esculls i espadats. Llocs com l’anomenat Blue Corner a Palau (Carolines) i Ras Mohammed a la península de Sinaí (Egipte), es consideren miradors privilegiats per a observar taurons.
Les rajades i les escurçanes són habitants comuns dels fons arenosos, tant als “lagoons” com als costats de mar oberta dels esculls. Aquests peixos cartilaginosos posseeixen una versió extremament aplanada del pla corporal dels taurons. Les més grans del grup, les mantes (Manta, Mobula), semblen una majestuosa catifa voladora, i tenen una envergadura de fins a 4 m. Contràriament als seus trets impressionants, són els habitants més innocus dels esculls; s’alimenten de plàncton que filtren de les aigües litorals. Les escurçanes (Dasyatis, Urolophus) també són dorsalment aplanades i habitants dels fons, on s’alimenten bàsicament de mol·luscs, les closques dels quals trituren amb les dents hemisfèriques. Les cues, com fuets, tenen uns aguts fiblons, que fan servir quan se senten amenaçades, i que poden infligir ferides greus. En alguns membres d’aquest grup, aquests fiblons fins i tot tenen una glàndula de verí a la base per afegir efectivitat a aquesta arma poderosa.
Les moles de nedadors: damisel·les, anfosos i peixos pallasso
Moltes espècies de peixos neden per les cares exteriors dels esculls en moles d’un nombre variable d’individus, des d’uns quants fins a molts milers. Aquest comportament de banc pot ser limitat a l’etapa juvenil, a l’etapa de fresa o ser permanent per tota la vida. Es pensa que les moles donen molts avantatges, des de reduir la resistència hidrodinàmica, la qual cosa estalvia energia metabòlica, fins a atemorir depredadors, que les veuen com una gran criatura amenaçadora. La gran quantitat de peixos que s’associen en forma de moles també pot distreure l’atenció del depredador o, com en els ramats d’herbívors terrestres, facilitar el sacrifici dels lents i malalts i la conservació dels més ben adaptats.
Hi ha espècies que usen l’escull com a refugi dels depredadors i que formen moles que es retiren a amagar-se entre les ramificacions dels coralls al més mínim senyal de perill, com pot ser una ombra que els cau al damunt. Aquests peixos, normalment molt petits, viuen permanentment dins d’un territori, una colònia de corall determinada. En la majoria de casos, la mola viu a la mateixa colònia de corall al llarg de tota la vida d’aquesta. Normalment, una mola d’una espècie determinada de peixos prefereix una espècie determinada de corall. Exemples típics en són les moles de diferents pomacèntrids, les menudes i acolorides damisel·les, com ara Chromis caerulea que, de vegades, fa moles de centenars d’individus que viuen en una colònia d’Acropora, o les moles petites de la damisel·la ratllada (Dascyllus aruanus), que també viuen normalment entre les branques d’Acropora. Aquests peixos només s’aventuren a deixar l’escull per capturar aliment i defensen el seu territori d’intrusos aliens, normalment contra peixos de la mateixa espècie. L’àrea defensada es troba dins una àrea més gran en què s’obté l’aliment. Una mica menys intensa és la l’associació entre peixos en espècies com el serrànid Anthias squamipinnis, el peix daurat de la mar Roja, que té un territori format per tot un massís de corall, o una part d’un escull. Un massís o part d’escull d’aquest tipus és una comunitat de corall amb múltiples espècies, en la qual la mola es refugia. Sota aquests massissos, o a tocar, es poden trobar, a més de les espècies esmentades anteriorment, tots els planctívors, les parelles territorials o els simples depredadors. D’aquests cal destacar els acolorits anfosos o neros d’escull (Epinephelus, Mycteroperca), que s’aparellen per tota la vida.
Un exemple especial i altament conspicu d’un grup de peixos territorials d’escull amb “adreça fixa”, són els anomenats peixos pallasso (Amphiprion, Premnas), que viuen en una associació simbiòtica amb les anemones de mar. Aquest grup inclou nombroses espècies comunes a tots els esculls de corall, que viuen en parella o en petites moles en simbiosi específica amb anemones de mar. La naturalesa d’aquestes simbiosis no és completament clara, però en la majoria dels casos inclou la protecció mútua contra depredadors, cosa aparentment més obligada per part dels peixos, ja que cap peix pallasso adult es troba mai sense una anemone hoste, mentre que són força comuns els grups d’anemones sense peixos pallasso. Si més no en alguns casos, els peixos s’han de “condicionar” a les seves anemones per immunitzar-se contra les cèl·lules urticants d’aquestes. D’altra banda, sembla que també hi ha relacions nutricionals entre els dos socis. En molts casos s’ha observat que els peixos, una vegada tips, porten l’aliment que els sobra a la boca de les anemones, que se l’empassen àvidament. Els peixos pallasso no tan sols s’amaguen dins de l’anemone quan són amenaçats, sinó que també fixen els ous a la paret darrere l’anemone, on estan protegits fins a l’eclosió. Altres peixos vivament acolorits, com el peix àngel emperador (Pomacanthodes imperator), es poden veure pasturant durant molts anys en un mateix tros d’escull.
S’ha suggerit que la coloració tan viva de molts peixos actua de senyal de perill per als intrusos de la mateixa espècie, en el sentit que els adverteixen que el territori està ocupat. L’etòleg austríac Konrad Lorenz va inventar el terme “colors de cartell” (“Plakatfarbe”), per a aquest tipus de coloració. En alguns casos, incloent-hi el del peix àngel emperador, els joves de l’espècie tenen un altre color completament diferent al dels adults, que possiblement els protegeix dels adults més grossos. Els diferents colors fan que semblin membres d’una espècie diferent i, per tant, que no siguin competidors potencials. Més recentment, Paul Ehrlich ha argumentat que alguns dels peixos més agressivament territorials tenen una coloració més grisa, mentre que els més acolorits semblen no ser territorials, o fins i tot viuen en moles.
L’art del camuflatge: escorpènids i cavallets de mar
En contrast evident amb les miríades d’espècies de peixos de colors vius que campen pels esculls, hi ha molts mestres de l’engany i el camuflatge. En particular els peixos pedra (Synanceia verrucosa i altres espècies de sinancèids), els peixos escorpí (diverses espècies dels gèneres Dendrochirus i Scorpaena i altres escorpènids) i els peixos granota (família dels antennàrids). Tots ells es refien de la seva capacitat de mimetisme gairebé perfecte amb el rerefons de l’escull per amagar-se dels enemics i les preses. Esperen, pacientment emboscats, fins que una presa incauta hi passa a prop; el depredador, normalment dropo, obre l’enorme boca i de fet la hi xucla. Molts d’aquests peixos tenen glàndules verinoses en la base de les seves espines dorsals i són un perill real per a qualsevol humà que els pot trepitjar accidentalment. La fiblada pot causar un dolor agudíssim i greus trastorns, fins i tot paràlisi i mort. El camuflament gairebé perfecte d’aquests peixos de vegades es fa encara més elaborat amb l’ús de coloracions de “flash”. Quan un peix escorpí o un peix pedra abandona la immobilitat que el fa invisible contra el fons, revela els colors llampants de les aletes desplegades, i aquest “flash” sobtat desapareix també sobtadament quan el peix es para, deixant l’observador perplex, sense saber què se n’ha fet, de tot aquell color.
Els dissenys de camuflatge poden variar des de l’aspecte d’un roc cobert d’algues, com en el cas del peix pedra, fins al d’esponges de colors vius, com en el cas del peix granota, o grapats d’algues flotants, imitades amb tots els detalls, com en el cas de Phyllopterix foliatus, un cavallet de mar d’aigües australianes. Altres espècies semblen fulles flotants i tota la seva anatomia es reorganitza per servir a la disfressa: els cossos s’aplanen i s’elonguen, el color es fa verdós, amb taques realistes i, detall aquest el més impressionant de tots, neden en una direcció perpendicular a l’eix del cos, com passa amb alguns centríscids; el cap apunta cap avall i l’aleta caudal que els impulsa, molt reduïda, es doblega en un angle recte respecte al cos.
Altres dissenys de color són pensats per dissimular els contorns generals del cos del peix amb diversos dibuixos geomètrics de ratlles i taques de color. En realitat alguns dels autors que rebutgen la teoria de Konrad Lorenz sobre la “coloració de cartell”, asseguren que els colors vius de tants peixos d’escull actuen més aviat com els dissenys que es pinten als avions, vehicles i edificis militars, per dissimular els seus contorns. Un altre truc és el d’amagar l’ull. Els ulls, a causa de les seves propietats òptiques i el seu color, destaquen de l’entorn, i criden l’atenció de l’observador. A més, alguns depredadors, incloent-hi el martinet d’escull (Egretta gularis), intenten llancejar els peixos pels ulls. És usual, doncs, amagar l’ull amb una franja fosca, o tenir-ne un de fals en un lloc distant del cos.
Les espines i els verins: diodòntids, tetraodòntids i peix escorpí
L’escull és tan summament ric d’espècies que les estratègies per defensar-se dels depredadors potencials més aferrissats són molt nombroses. L’espectacular peix escorpí (Pterois volitans), que sembla de randes, té glàndules verinoses a la base dels radis de les aletes. És clar que pot nedar majestuosament serè, fent evident el seu verí per la coloració, i engolint peixets amb tota la calma. Molts nedadors i escafandristes han estat atrets per la seva bellesa i la seva pausada navegació per les aigües, i han intentat atrapar-lo o tocar-lo amb les mans nues. La fulminant fiblada inesperada de les seves espines dorsals és una lliçó dolorosa i potencialment mortal per als incauts. Una estratègia completament diferent és la del llenguado de Moisès (Pardachirus marmoratus) de la mar Roja. Aquest peix té com a depredadors els taurons, i els combat amb una secreció dèrmica verinosa capaç de paralitzar-los la mandíbula un verí neurotòxic que s’estudia actualment com un possible repel·lent de taurons per a ús humà.
Els peixos globus (Arothron i altres gèneres de la família dels tetraodòntids) i els peixos eriçó (Diodon hystrix i altres diodòntids) usen una combinació única d’armes per aturar els enemics. Tenen la capacitat d’inflar-se d’aigua, i incrementar així el volum i canviar la forma elongada pròpiament pisciforme per una de globus el·líptic en el cas dels peixos globus, o una esfera gairebé perfecta en el cas dels peixos eriçó. En aquests últims, el fet d’inflar-se fa també que les espines, fortes i gruixudes, s’aixequin d’una posició paral·lela a l’eix del cos a una de perpendicular. Els noms de les famílies diodòntids i tetraodòntids al·ludeixen al fet que aquests peixos tenen totes les dents fusionades en dues (una a cada mandíbula) en el cas dels diodòntids, o quatre (una a cada costat de cada mandíbula) en el dels tetraodòntids. Aquests darrers, a més dels seus mecanismes defensius anatòmics i etològics, també produeixen un dels verins més potents coneguts, la tetradoxina, emprada en medicina i en la recerca neurofisiològica.
Les tortugues i els dugongs
Altres organismes més grossos de l’entorn dels esculls són les tortugues i els dugongs, a frec de l’extinció a causa de la sobreexplotació de la pesca.
Les tortugues sempre han estat preuades per les seves closques, que en alguns llocs del Pacífic eren d’enorme importància en la manufactura de joies, ornamentació i objectes de culte, i que també es comercialitzaven com a material per a la manufactura de pintes i muntures d’ulleres, i per la seva carn, tinguda per exquisita arreu del món. Aquests rèptils longeus, que pesen centenars de quilos, arriben a fer, per exemple en el cas de la tortuga verda (Chelonia mydas), una longitud de closca de més d’1 m i poden arribar a viure fins a cent anys. Tanmateix, a més de ser objecte d’una sobreexplotació pesquera temerària, les urbanitzacions a primera línia de mar han destruït la majoria de platges arenoses que necessiten per a pondre els ous.
Els dugongs (Dugong dugon), considerats per molts com a inspiradors d’antigues llegendes de sirenes, són grans mamífers marins que arriben a fer 2,5 m de llargària i de 250 a 400 kg de pes, que pasturen les praderies de fanerògames marines. Com a comentari interessant, els israelites, quan vagarejaven per les costes de la mar Roja al Sinaí, van usar pells de dugong per a la tenda que empraven per al culte en el desert. Aquest santuari portàtil va ser substituït segles més tard, quan els israelites ja estaven establerts a la Terra Promesa d’Israel, pel temple de Salomó.