La vida al fons de la mar
El fons de la mar és un dels reductes de la biosfera menys assequibles directament als humans; la seva extensió és enorme: ocupa aproximadament un 70% de l’escorça terrestre. Els humans, tot i la impossibilitat de colonitzar aquests grans espais, n’exploten una gran part, ja sigui en forma de pesca, ja en la d’extracció de recursos minerals. Aquesta explotació no es correspon, però, amb un nivell de coneixements comparable al que hom té dels ecosistemes terrestres. Hi ha encara grans buits, en efecte, sobre el funcionament dels ecosistemes submergits i, fins i tot, dels organismes que els habiten.
La diversitat dels fons marins és extraordinària, ja que abasta des de llocs ben il·luminats fins a zones en la foscor més absoluta, temperatures des de més de 30°C fins a inferiors als 0°C, pressions que poden ultrapassar les 1 000 atmosferes, aportacions de nutrients i moviments de l’aigua radicalment diferents, fons tous i fons durs, etc. Aquest ventall tan ampli de condicions ecològiques es reflecteix en una diversitat paisatgística igual o superior a la dels ecosistemes terrestres a escala mundial, i en una diversitat de classes o fílums d’organismes sense parió. Tots els organismes que viuen associats al fons de la mar i que, per tant, ocupen la interfase aigua-sediment o aigua-roca s’anomenen organismes bentònics. Tots ells, juntament amb el medi físic que els envolta, constitueixen el sistema bentònic o bentos.
Els productors primaris: bacteris, algues i algunes herbes
Els productors primaris del món bentònic són principalment organismes fotosintetitzadors, el quals tenen una distribució limitada a la zona més superficial, il·luminada. Bacteris quimioautòtrofs, com els que viuen associats a les fumaroles de les dorsals oceàniques, constitueixen els únics productors primaris bentònics no fotosintetitzadors. La seva producció, però, tot i ser del més gran interès com a font d’aliment d’un dels ecosistemes més peculiars i interessants de la biosfera, és irrellevant en el conjunt del medi marí. Altres bacteris quimioautòtrofs que viuen en els fons sedimentaris obtenen el poder reductor de molècules inorgàniques reduïdes, presents en el sediment com a resultat de la descomposició de la matèria orgànica causada per altres processos bacterians i, per tant, no es tracta d’una producció estrictament “nova”. Fanerògames, algues, cianobacteris i determinats bacteris fotosintetitzadors del sofre es consideren els productors primaris bentònics per excel·lència.
Les fanerògames marines són plantes d’origen terrestre, posteriorment adaptades a la vida aquàtica, que han reeixit especialment en els fons sedimentaris superficials, on la competència per l’espai amb les algues els ha estat favorable mercès a les arrels, que els permeten fixar-se al fons i extreure’n nutrients.
Les algues, molt diversificades, són els principals productors primaris en la majoria d’ecosistemes bentònics. Les algues verdes (cloròfits), brunes (feòfits) i vermelles (rodòfits) poblen amb abundància les roques submergides d’arreu del món, sempre que hi hagi un mínim de llum per a realitzar la fotosíntesi. Les diatomees (crisòfits) són molt importants en fons sedimentaris, mentre que les zooxantel·les (dinòfits) són de gran importància en els esculls coral·lins, on viuen com a endosimbionts dels coralls.
Els cianobacteris (abans anomenats cianòfits, cianofícies o algues verd blaves) són presents una mica pertot, però són especialment abundants en condicions ecològiques extremes com les que es donen en les zones d’estuari, els “lagoons” tropicals, les àrees hipersalines o els indrets rics de nutrients.
Finalment, els bacteris fotosintetitzadors del sofre apareixen només en fons sedimentaris, tot concentrant-se en aquell nivell del sediment on les condicions són pràcticament anòxiques, ja que utilitzen l’àcid sulfhídric com a donant d’electrons i el transformen en sofre o en sulfat.
Els consumidors i els descomponedors: animals, fongs i bacteris
La major part dels fons oceànics es troben a fondàries considerables més enllà d’on hi ha llum suficient per a sustentar la vida dels organismes fotosintetitzadors. En efecte, la profunditat mitjana dels oceans és d’uns 4 000 m, mentre que la capa fòtica, aquella que permet la penetració de la llum, no ultrapassa els 100 m de mitjana. La resta és el regne de la foscor total, només interrompuda pels esclats de bioluminiscència d’alguns organismes abissals.
Això provoca que la vida sota la zona il·luminada depengui de fonts d’energia externa al sistema. L’entrada d’aliment és sempre condicionada per la capacitat productiva de la zona fòtica i arriba sempre transportada per l’aigua. Aquesta matèria en suspensió pot ser directament consumida per uns organismes especialment adaptats a capturar-la —els suspensívors— o sedimenta en el fons marí i allí és utilitzada pels organismes sedimentívors o directament degradada pels descomponedors.
La dels suspensívors és una estratègia tròfica importantíssima en el medi aquàtic, encara que sigui pràcticament inexistent en el medi terrestre, on l’única (bé que llunyana) similitud es troba en els aràcnids teixidors de xarxes per a capturar insectes voladors. La categoria d’organismes suspensívors és conceptualment molt àmplia, ja que, de fet, engloba tant animals filtradors com depredadors. Els filtradors típics separen del medi per filtració partícules sempre de mida petita (bacteris, matèria orgànica particulada, fitoplàncton); en són exemples característics les esponges, els ascidis i molts mol·luscs bivalves. Altres concentren les partícules en suspensió prop de la boca servint-se de les brànquies (poliquets), dels cirrus (cirrípedes) o dels braços (crinoïdeus); i encara uns darrers, els pogonòfors, tenen uns tentacles on queden enganxats els organismes planctònics, els quals són digerits externament. Els suspensívors depredadors, en canvi, són organismes colonials amb pòlips o zooides que capturen les seves preses (principalment zooplàncton) mitjançant tentacles o estructures tentaculars; diversos grups de cnidaris (hidrozous, alcionaris, antipataris, coral·limorfaris, zoantaris, madreporaris) i els briozous es poden incloure en aquesta categoria. Hom també pot dividir els organismes suspensívors en dues categories, segons que l’energia necessària per al moviment de l’aigua és proveïda pels mateixos animals (cas dels filtradors actius) o pels corrents marins (cas dels filtradors passius).
Els organismes detritívors, que s’alimenten de restes orgàniques, tenen una especial rellevància en el bentos, ja que s’alimenten de les restes de la producció bentònica com, per exemple, els cadàvers dels organismes pelàgics que s’hagin dipositat al fons de la mar. Molts dels organismes detritívors del bentos són sedimentívors, ja que ingereixen el sediment, del qual assimilen la matèria orgànica i n’expulsen la part mineral. D’aquesta manera aprofiten tant les restes orgàniques com els petits animals i microorganismes que viuen en el sediment. Determinats grups de poliquets, mol·luscs bivalves, holoturioïdeus, equinoïdeus, tanaidacis i isòpodes constitueixen el nombre més gran d’organismes sedimentívors.
Les comunitats bentòniques també acullen herbívors i carnívors. Els herbívors són nombrosos a les zones rocoses més superficials, a la zona fòtica, on viuen les algues. Les garotes, determinades espècies de peixos, alguns mol·luscs gasteròpodes i poliplacòfors i certs crustacis s’alimenten d’algues i de fanerògames marines. Els carnívors, també abundants, depreden tant sobre la fauna sèssil com sobre la vàgil; els actiniaris o anemones de mar, alguns poliquets, molts crustacis, mol·luscs gasteròpodes i cefalòpodes, els asteroïdeus i ofiuroïdeus, i un elevat nombre de peixos, en són exemples.
Finalment, els organismes descomponedors estan associats a la matèria orgànica morta i s’encarreguen de degradar-la fins a convertir-la de nou en compostos inorgànics (mineralització). Com en els ecosistemes terrestres, els organismes descomponedors dels fons marins són constituïts bàsicament per bacteris i alguns fongs.
Els factors ambientals
Les característiques de les comunitats bentòniques que s’instal·len sobre un fons marí determinat depenen, en gran mesura, de les seves característiques ambientals. El grau d’estructura, la complexitat i el funcionament d’una comunitat és imposat per una sèrie de factors abiòtics; i això, tant pel que fa als seus valors mitjans com a la seva variació espacial i temporal. El resultat final, és a dir, la comunitat que en resulta, depèn, però, de les interaccions biòtiques que s’estableixen entre els diferents organismes que estan, a priori, adaptats a les condicions ambientals imposades pels factors abiòtics, amb la modulació imposada per les corresponents restriccions biogeogràfiques.
És important de remarcar que, en determinats casos, són més importants les variacions en els paràmetres ambientals que els seus valors mitjans. Molt sovint les variacions i els valors màxims i mínims de temperatura, llum, hidrodinamisme, salinitat, o qualsevol altre factor ambiental, limiten el creixement dels organismes, o determinen la dinàmica de les comunitats bentòniques. Així, per exemple, les condicions ambientals mitjanes de temperatura, llum i nutrients que hom troba a les aigües de l’illa de Cabrera, a la Mediterrània occidental, i a moltes zones de la Mediterrània central i oriental, possibilitarien el desenvolupament d’esculls coral·lins. Això no obstant, les temperatures mínimes hivernals de 14 o 15°C que es mantenen durant almenys tres mesos impedeixen la vida dels coralls hermatípics en aquesta zona. L’absència de poblaments de laminàries gegants al Pacífic de la península de Califòrnia cap al S, sembla estar determinada principalment per la durada del període de l’any amb aigües relativament calentes i pobres de nutrients.
El substrat físic
La naturalesa física del substrat separa els fons tous o sedimentaris dels fons durs o rocosos. En termes generals, l’estabilitat dels primers és molt inferior a la dels segons, i això condiciona enormement els tipus d’organismes que hi viuen, l’estructura tròfica i l’ocupació de l’espai. En els fons durs hi dominen els organismes sèssils que creixen sobre el substrat, recobrint-lo o fent-lo servir de suport. L’ocupació de l’espai és, doncs, del substrat cap a l’aigua. En els fons tous, en canvi, els organismes, generalment mòbils, es concentren a l’interior o a la part superficial del sediment; per això les característiques físiques (naturalesa, constitució granulomètrica) i les químiques (oxigenació, potencial redox, contingut de matèria orgànica) del sediment tenen una incidència fonamental sobre el poblament d’organismes que s’hi estableixen.
En els fons rocosos superficials, les macroalgues són importantíssimes com a compartimentadores de l’espai i com a font d’aliment, i possibiliten l’existència d’una sèrie d’herbívors i detritívors que incorporaran aquest nodriment als altres nivells tròfics. En els fons sorrencs no poblats per fanerògames marines o per determinades algues clorofícies capaces d’establir-s’hi, les algues unicel·lulars que s’hi desenvolupen no hi tenen cap funció estructural i són consumides pels sedimentívors. Les roques de les aigües profundes, on no arriba prou llum o no gens, en canvi, són dominades per suspensívors que incorporen al bentos la matèria orgànica en suspensió a l’aigua.
D’altra banda, en els fons tous de sediments fins i relativament rics en matèria orgànica, els sedimentívors són els organismes més abundants, mentre que en determinats indrets de sediment gruixut i relativament pobres en matèria orgànica, l’estratègia tròfica dominant pot ser la dels suspensívors. La conversió d’un fons tou en un fons dur o, si més no amb les característiques biòtiques pròpies d’un fons dur, és possible i va lligada a l’activitat edificadora dels organismes. En són exemples la fixació del sediment per part dels herbeis de fanerògames marines, l’edificació d’estructures organògenes per acumulació d’esquelets d’algues carbonatades que viuen lliures en els fons sedimentaris, o —si més no parcialment— la creació de substrat dur per l’activitat dels coralls hermatípics i les algues carbonatades en els esculls coral·lins. La conversió inversa és també possible i, encara que pot anar també lligada a l’activitat dels organismes (per exemple, l’erosió provocada per certs peixos herbívors en els esculls coral·lins), sovint és causada per processos físics d’erosió i de rebliment.
La llum
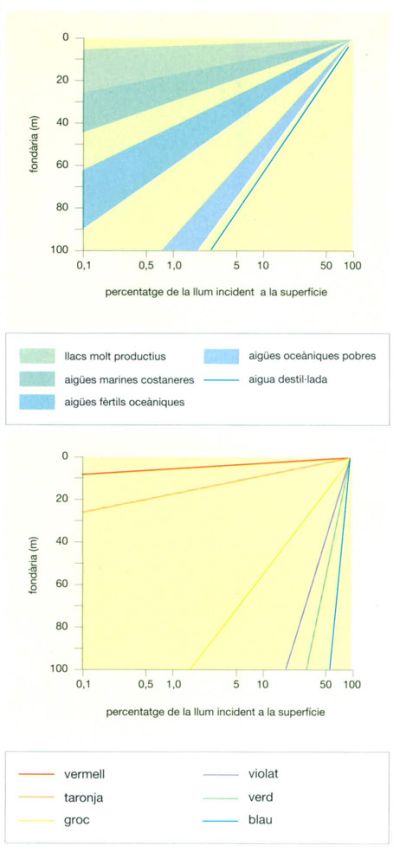
Perfils d’extinció de la llum i de decrement de la intensitat dels colors a mesura que augmenta la fondària. L’extinció és molt més severa a les aigües productives, i per això mateix més tèrboles, que no pas a les aigües pures i poc fèrtils.
Jordi Corbera, a partir de fonts diverses
La llum té una importància fonamental en el bentos marí. La difusió de la llum a l’aigua depèn de la transparència d’aquesta, la qual és determinada per les partícules en suspensió. En qualsevol cas, sigui l’aigua clara o tèrbola, l’extinció de la llum en fondària segueix una funció exponencial i els vegetals tenen limitada la seva distribució als fons marins il·luminats. Segons la seva distribució i la seva resposta a la llum, hom parla d’organismes fotòfils (amants de la llum) o esciòfils (amants de la foscor).
Recentment hom ha trobat algunes espècies d’algues adaptades a viure a intensitats de llum molt baixes. Això no obstant, la possibilitat de vida autòtrofa fotosintetitzadora està generalment limitada a aigües superficials, i no acostuma a sobrepassar els 200 m en mars tropicals i subtropicals, els 120 m en mars temperades, i els 60 m en mars fredes. La dominància de les algues enfront d’altres organismes bentònics té una amplitud encara més reduïda. Per tant, si tenim en compte l’extensió de la mar i l’elevada fondària mitjana dels oceans, veiem que la part del fons marí on la producció primària té una importància significativa és molt minsa.
Això té diverses implicacions de la màxima importància en l’estructura tròfica dels ecosistemes bentònics, ja que la principal font d’energia passa de ser la llum en els ecosistemes superficials a ser les partícules capturades pels organismes suspensívors o dipositades al fons i consumides pels sedimentívors en els ecosistemes profunds. Evidentment aquest canvi és gradual, de tal manera que la importància de la producció primària autòctona és màxima en els primers metres i disminueix progressivament cap al fons.
Ultra aquesta distinció entre “fons il·luminats” i “fons foscos”, dins la zona fòtica, la diferent intensitat de llum d’uns llocs a uns altres també té una importància extraordinària, ja que és la principal responsable dels canvis que s’observen en la composició específica de les comunitats (sobretot en les algues i en els animals amb cèl·lules vegetals simbionts, però també en d’altres organismes), en la seva estructura espacial, la seva capacitat productiva, la seva dinàmica i les relacions biòtiques que s’estableixen entre els organismes. En serien exemples les diferentes bandes o horitzons que s’estableixen en les comunitats que es desenvolupen sobre zones rocoses a mesura que augmenta la fondària o bé el gradient de poblaments bentònics que hom troba a les parets de les coves, principalment causat per la diferent disponibilitat de llum.
L’hidrodinamisme
L’hidrodinamisme també és un factor important en el bentos marí. L’agitació renova la capa d’aigua immediatament adjacent als organismes i afavoreix tant la captació d’anhídrid carbònic i nutrients per part de les plantes, com les partícules alimentàries en el cas dels organismes suspensívors. Ara bé, un excés d’agitació és contraproduent, ja que els individus han de tenir una morfologia especialment adaptada a la força del corrent per a evitar de ser arrencats. A més, si es produeix un intens hidrodinamisme en un fons tou o en un de rocós envoltat de sorra, el moviment i la resuspensió del sediment dificulta la vida en aquest ambient. De fet, una de les zones més desfavorables per a la vida és la dels fons sorrencs situats a flor d’aigua en costes batudes, ja que la gran mobilitat del sediment i la manca d’aliment són determinants de la seva baixa diversitat i biomassa.
La zona superficial acostuma a ser la més agitada, però aquesta agitació és turbulenta i disminueix exponencialment en fondària. Això es reflecteix en la distribució de les garotes o altres organismes vàgils que no suporten una agitació intensa. Molt importants són els corrents de marea, direccionals, els quals també són especialment intensos a les zones més superficials, principalment en zones estuàriques o d’escassa fondària. Més al fons, però només en zones concretes, poden originar-se forts corrents causats per fenòmens locals o per causes de tipus geostròfic; aquestes zones profundes afectades per corrents tenen també uns poblaments ben diferenciats, constituïts per espècies anomenades reòfiles (amants dels corrents). Així, per exemple, els briozous, esponges incrustants i madreporaris generalment dominants a les parets situades entre 40 i 90 m de fondària a la Mediterrània central, són substituïts per gorgònies i esponges globuloses en les zones afectades per corrents forts.
Els nutrients i la matèria orgànica
Les comunitats d’organismes bentònics també estan fortament influïdes per la disponibilitat de nutrients i matèria orgànica, ja que molt sovint en determinen la seva capacitat productiva. Les algues laminarials, per exemple, són exclusives de mars temperades o fredes, riques de nutrients, mentre que són raríssimes o totalment absents en zones tropicals i subtropicals, on la pobresa de nutrients de l’aigua impedeix el seu desenvolupament o el limita a les aigües més riques del fons.
El creixement de les algues i les fanerògames marines sovint és determinat per la seva capacitat d’assimilar nutrients i el seu cicle estacional reflecteix, en molts casos, les variacions en el contingut de nutrients de l’aigua. De fet, el màxim creixement de moltes algues fotòfiles de mars temperades que creixen a poca fondària es produeix en coincidència amb el màxim de nutrients dissolts a l’aigua (generalment nitrats, però també fosfats) i no del màxim d’irradiància. La presència o l’absència de coralls productors d’esculls s’ha associat generalment a la temperatura de l’aigua, però la riquesa de nutrients també és un factor determinant per al creixement d’aquests organismes, com ho reflecteix la seva absència o abundància molt menor en les zones tropicals riques de nutrients. El grau d’eutròfia o oligotròfia d’un indret determina també el poblament algal que s’hi desenvolupa, ja que les algues oportunistes, de creixement ràpid, eliminen els poblaments més madurs, d’algues més grosses i de creixement generalment més lent, en situacions eutròfiques.
La quantitat de matèria orgànica particulada en suspensió a l’aigua és fonamental com a determinant de la densitat i el tipus d’organismes suspensívors. Així, per exemple, el canvi en espècies, grups d’organismes i recobriment que hom observa en endinsar-se en una cova submarina és determinat principalment per una menor disponibilitat d’aliment per a la fauna cavernícola. Ja hem comentat també que en els fons tous, les zones amb un sediment ric en matèria orgànica són favorables al desenvolupament dels organismes sedimentívors, mentre que els sediments amb poca matèria orgànica afavoreixen els suspensívors.
La temperatura
La temperatura és un altre factor important en el bentos marí, ja que determina la distribució de moltes espècies, tant geogràfica com vertical.
Pel que fa a la distribució geogràfica, l’exemple més clàssic és la coincidència entre la temperatura mínima de 20°C i la distribució dels esculls corallins arreu del món. Hi ha, a més, moltíssimes espècies la distribució de les quals sembla limitada per les temperatures màximes o mínimes anuals. Això no obstant, i principalment pel que fa a les algues, els límits geogràfics sovint atribuïts a la temperatura també poden estar causats per la riquesa de nutrients a l’aigua, ja que les aigües fredes en són usualment riques i les aigües càlides n’acostumen a ser pobres. Tot i això, és evident l’existència d’una limitació en la distribució de les espècies a causa de la temperatura. Nombrosos gèneres d’algues estan restringits a zones càlides (per exemple, Halimeda, Caulerpa, Udotea, Dictyosphaeria, Turbinaria) i altres a zones fredes (Desmarestia, Iridaea, laminarials). Semblantment passa amb qualsevol grup faunístic; és especialment conegut el cas dels peixos. Fins i tot l’abundància i el límit nord de determinades espècies de peixos d’afinitats tropicals oscil·la anualment a la Mediterrània occidental, com és el cas del fadrí (Thalassoma pavo) o la vaca serrana (Serranus scriba).
La formació d’una termoclina durant els mesos estivals a totes les mars temperades condiciona l’amplitud batimètrica d’algunes espècies, en impedir el seu creixement per sobre o per sota d’aquest nivell. N’és un clar exemple la distribució de diversos endemismes d’algues mediterrànies, entre ells Laminaria rodriguezii, que mai no apareixen per sobre del nivell estival de la termoclina. Altres organismes presents a la Mediterrània, com determinats braquiòpodes (Megerlia truncata, Gryphus vitreus), viuen només en aigües profundes, però mai en coves submarines, cosa que suggereix que la temperatura és el factor limitant de la seva distribució.
Les pertorbacions
La incidència de pertorbacions o de factors catastròfics d’origen natural o induïts per l’activitat humana és fonamental per a molts poblaments. Les comunitats situades sobre la plataforma continental són les més afectades per aquests esdeveniments. En canvi, les condicions ambientals per sota dels 200 m de fondària són molt més regulars, i la probabilitat d’aquests fenòmens és molt més baixa. Temporals, minves, períodes inusuals de gran terbolesa o claredat de l’aigua o, fins i tot, esdeveniments amb un període pluriennal, tenen una incidència considerable en els poblaments bentònics.
A la Mediterrània, la dinàmica anual de la comunitat dominada per l’alga coneguda com pèl sauper (Cystoseira mediterranea) depèn de la incidència de les minves. Com que és un poblament que no va més enllà del metre de fondària, períodes llargs de mar encalmada i un nivell de la mar excepcionalment baix (causat per la presència d’un centre d’altes pressions) durant la primavera poden provocar la dessecació i la mort dels branquillons d’aquesta espècie i influir així negativament en la producció global i en la resta d’espècies de la comunitat.
La incidència de forts temporals també pot modificar extraordinàriament l’estructura d’algunes comunitats, tal com va ocórrer, per exemple, al gener del 1988 a les costes de Califòrnia amb un temporal qualificat com el pitjor en 200 anys. L’efecte sobre els “boscos” de laminàries gegants (Macrocystis pyrifera) fou qualificat de dramàtic, ja que va causar una mortalitat sense precedents (propera al 100%) en aquestes algues pluriennals. A continuació l’alga Desmarestia ligulata va recobrir tot el substrat disponible, però poc a poc el “bosc” de Macrocystis es va anar reconstituint fins a assolir recobriments comparables als d’abans de la tempesta al cap d’un any i mig.
Una altra causa de pertorbació important en les comunitats bentòniques poden ser episodis climàtics de periodicitat irregular però pluriennal, com El Niño. Tot i que el principal efecte s’observa en les comunitats planctòniques i incideix molt negativament en determinats peixos pelàgics d’interès comercial, El Niño afecta també les comunitats del bentos, ja que provoca una minva molt important de la matèria orgànica disponible per al sistema bentònic provinent del món pelàgic i afecta també negativament el creixement dels macròfits litorals (Macrocystis i altres laminarials). Les repercussions d’aquest fenomen van ser molt serioses per als poblaments de laminàries gegants de les costes californianes a l’hivern de 1982-83, que van quedar molt malmeses i van trigar dos anys a presentar indicis de recuperació.
La salinitat i la sedimentació
Hi ha encara altres paràmetres que afecten les comunitats bentòniques, com són la salinitat i la taxa de sedimentació, amb importància, sobretot, en ambients costaners.
Les variacions de salinitat afecten els organismes que viuen en ambients estuàrics o en zones tancades com les petites albuferes o els “lagoons” tropicals. La incidència sobre la resta d’ambients bentònics és desconeguda i, en tot cas, molt baixa.
La taxa de sedimentació pot ser localment intensa i afectar qualsevol organisme sèssil, sigui perquè quedi enterrat, sigui, en el cas dels organismes suspensívors, perquè resultin obturats els mecanismes de filtració. Alguns organismes, com ara moltes algues dels gèneres Caulerpa, Dasycladus, Halimeda i Penicillus, suporten o estan especialment adaptats a ambients amb taxes de sedimentació elevades. En canvi, en altres ambients, la manca de sedimentació pot comportar l’erosió del sediment, amb importants repercussions tant per als animals que puguin viure dins seu com per als organismes que viuen a la seva superfície.
La relació entre els organismes
Els paràmetres ambientals condicionen en gran mesura l’estructura i la dinàmica de les comunitats, però són, en darrer cas, les interaccions biòtiques les que determinen moltes de les característiques dels poblaments que s’estableixen sobre el fons marí. Hem de tenir en compte, però, la diferència en la importància dels factors abiòtics i els biòtics. Per exemple, un fons sedimentari situat a 4 000 m de fondària, amb una temperatura de 2°C i totalment fosc tindrà un poblament bentònic radicalment diferent al d’un escull de corall o un “bosc” de laminàries, i la seva estructura i dinàmica reflectirà les diferents característiques ambientals dels dos tipus de fons. A més, aquestes comunitats no són intercanviables, és a dir cap d’elles no pot succeir-ne una altra en un temps a escala ecològica o humana. En canvi, un “prat” d’algues erectes pot esdevenir una zona rocosa amb un recobriment algal constituït només per algues coral·linàcies incrustants a causa d’una lleu modificació en el comportament, en la intensitat de depredació o en el reclutament de les garotes, els principals herbívors de molts sistemes costaners. En aquest cas, el poblament d’algues erectes i la roca amb coral·linàcies i garotes són dues comunitats intercanviables, ja que és possible el pas d’una a l’altra.
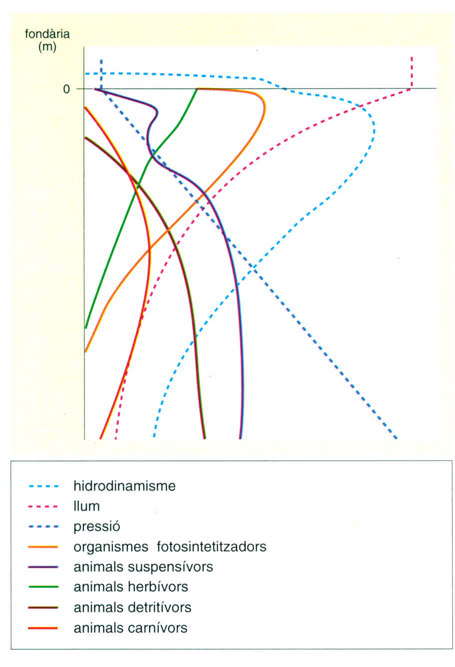
Variació de les estratègies tròfiques i dels factors ambientals relatius al bentos marí en funció de la fondària (tanmateix, les variacions estacionals de la temperatura, de la disponibilitat de nutrients i de l’existència de matèria orgànica particulada són tan o més accentuades que aquesta variació en fondària).
Editrònica, a partir de fonts diverses
Actualment s’admet, doncs, que els factors abiòtics determinen el possible “pool” d’espècies que es poden desenvolupar en un indret deteminat, com també els tipus de comunitats que es poden establir. Això no obstant, els factors biòtics poden ser molt importants com a estructuradors de les comunitats bentòniques. De fet, la importància relativa d’uns i altres sembla dependre del sistema, de tal manera que en les comunitats més estructurades són les relacions biòtiques les que determinen la constitució final dels poblaments, mentre que en les comunitats menys madures els factors físics tenen una importància més gran.
Els mecanismes biològics que determinen la complexitat dels ecosistemes marins són infinitament més diversos que els paràmetres ambientals esmentats abans. De fet, abasten les característiques de morfologia, creixement, reproducció, capacitat de dispersió, alimentació, excreció i mortalitat de totes i cadascuna de les espècies presents. Mitjançant aquestes característiques, les espècies interaccionen entre elles de maneres diferents i, si bé algunes relacions tenen poca o cap importància, d’altres són vitals per a entendre la constitució final de l’ecosistema.
La depredació
L’entrada d’energia i la seva transferència són dos punts fonamentals en un ecosistema. Del que hem dit en parlar de les estratègies tròfiques es pot deduir que els organismes fotosintetitzadors, els suspensívors i els detritívors són les possibles vies d’entrada energètica. Suspensívors i detritívors s’alimenten, almenys parcialment, de matèria produïda a la zona pelàgica i la integren així al sistema bentònic, mentre que algues i altres fotosintetitzadors produeixen matèria orgànica a partir de diòxid de carboni, aigua, llum i nutrients. La depredació dins del sistema bentònic se centra, doncs, en la matèria proporcionada per aquestes tres estratègies tròfiques i es transmet pels herbívors i els carnívors.
Model de Witman
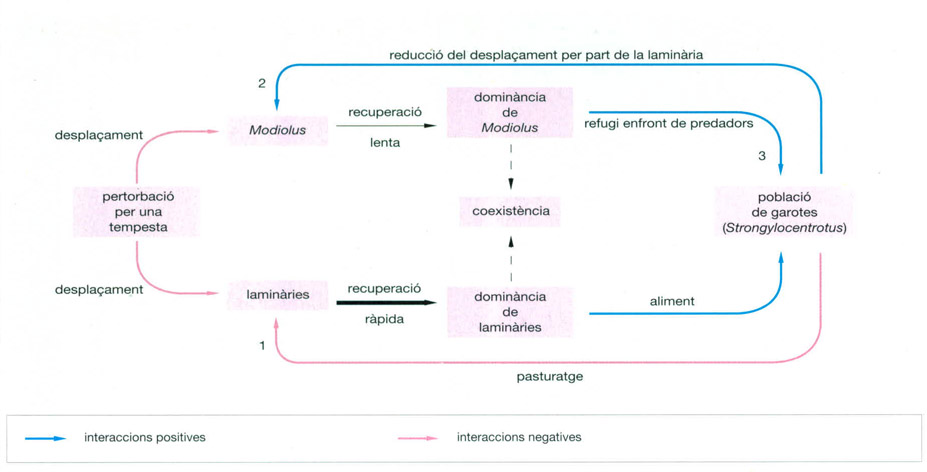
Jordi Corbera
Model conceptual d’interaccions entre laminàries, garotes (Strongylocentrotus) i musclos (Modiolus), elaborat per Witman, on les relacions es classifiquen com a positives o negatives. Com s’ha demostrat en experiments basats en l’extracció de garotes del seu hàbitat, el pasturatge d’aquests animals té un efecte negatiu sobre les laminàries (1), perquè minva el llindar inferior de distribució de l’alga. Pel que fa a la relació entre Modiolus i Strongylocentrotus, hi ha un fenomen de retroalimentació positiva (2 i 3), ja que el musclo proporciona un refugi a l’equinoderm davant dels seus depredadors, especialment en el cas de les garotes petites i, en el cas de les de mida més gran, sempre minva la possibilitat d’ésser menjades o arrencades per les tempestes; la relació Modiolus-Strongylocentrotus és, per tant, de mutualisme, ja que ambdues espècies en treuen profit. Quan s’ha tractat de recolonitzar les clarianes produïdes pel desplaçament o arrencament ocasionat per una tempesta, s’ha vist que les laminàries es veuen afavorides per la seva ràpida capacitat de recuperació i colonitzen l’espai. La dificultat del musclo per a reomplir les clarianes produïdes per l’arrencament fa que, segons la pressió del pasturatge, Modiolus perdi espai davant de la laminària recolonitzadora.
Hi ha moltíssimes referències de la importància d’aquestes relacions en l’estructura del bentos marí, però potser l’exemple més alliçonador és el dels poblaments de laminàries (gèneres Macrocystis, Nereocystis, Laminaria), les garotes (gènere Strongylocentrotus) i les llúdries marines (Enhydra lutris) de la costa oest d’Amèrica del Nord. Les garotes són els principals consumidors d’algues, encara que en situacions normals s’alimenten principalment de trossos d’algues a la deriva. Si aquesta font d’aliment és insuficient, les garotes es mengen les algues vives i poden devastar totalment un “bosc” de laminàries. Les llúdries marines i determinades estrelles de mar (Pycnopodia helianthoides) s’alimenten principalment de garotes, i en limiten el creixement de les poblacions. La caça indiscriminada de les llúdries durant el segle passat i a l’inici del present va comportar canvis importants en la distribució i l’estructura dels “boscos” de laminàries, mentre que la protecció d’aquests animals ha contribuït a regenerar aquests poblaments d’algues.
La importància de les garotes i dels seus depredadors com a espècies-clau en les comunitats bentòniques infralitorals de substrat rocós és general i ha estat remarcada tant en les comunitats de laminarials de l’Atlàntic i del Pacífic, com als esculls coral·lins —on els peixos herbívors comparteixen amb les garotes el paper de controladors de l’abundància i la distribució de les algues— o a les comunitats algals mediterrànies. A molts indrets de les illes Canàries, a l’Atlàntic, per exemple, l’extraordinària abundància de la garota Diadema antillarum entre els 2 i els 40 m de fondària impedeix la proliferació de comunitats algals ben estructurades, excepte en aquelles zones que tenen un fort hidrodinamisme; la causa d’aquesta densitat extraordinària de garotes a les Canàries es desconeix, però és possible que vagi lligada a la sobrepesca dels seus principals depredadors.
Nombroses estratègies permeten als organismes bentònics escapar-se de la depredació: la cripsi, el mimetisme, el fet d’amagar-se en anfractuositats i esquerdes de les roques, l’elaboració de closques, esquelets durs, espines o espícules, l’activitat nocturna, l’agregació, l’enterrament, els moviments ràpids, la defensa química i moltes d’altres. La diversitat d’organismes que conviuen en un sistema bentònic va associat a una diversitat d’estratègies defensives, de tal manera que la màxima complexitat s’observa en els ecosistemes més estables i madurs, i la mínima en comunitats dominades per unes poques espècies que interactuen fortament entre elles i amb el medi físic.
La competència
Els organismes bentònics competeixen gairebé sempre pels recursos limitants com la llum, els nutrients, l’aliment o l’espai. Les diferents morfologies de les algues responen sempre a un compromís entre l’adaptació als factors ambientals i la necessitat de competir amb més èxit amb les altres algues. El creixement en alçada, per exemple, com en els arbres de la selva tropical, és una resposta a la competència per la captació de llum. Excel·lents exemples de com pot actuar la selecció natural en aquest sentit es poden trobar en les algues laminarials. La majoria de gèneres d’aquest ordre de feòfits com ara Nereocystis, Macrocystis o Laminaria i moltes fucals (Sargassum, Cystoseira) formen “boscos” densos que redueixen molt la llum que arriba al substrat, la qual cosa afecta de manera decisiva el reclutament, el creixement, la reproducció i la supervivència de les algues més petites. L’eliminació natural o experimental de la coberta d’algues grans permet la fixació i el creixement d’un gran nombre d’espècies.
La competència per l’aliment és molt usual entre els animals bentònics, els quals sovint exploten els mateixos recursos alimentaris. Algunes vegades aquesta competència fa que espècies que utilitzen el mateix recurs tinguin distribucions diferents, però en d’altres, les espècies coexisteixen. Un dels tòpics encara no solucionats en biologia marina és com explicar l’alta diversitat d’espècies que aparentment utilitzen el mateix recurs alimentari en una mateixa comunitat. Probablement la resposta es troba en el fet que el recurs alimentari és lleugerament diferent i el que en realitat manca és un coneixement més detallat de la biologia de cada espècie. Així, per exemple, l’alta diversitat de detritívors que apareixen en les comunitats fangoses de molta fondària, sembla que té per causa una diferenciació específica en la ingestió de partícules del sediment d’una mida o d’una altra. Altres vegades, la coexistència és possible gràcies a la diferent resposta de les espècies competidores a lleugers canvis en els paràmetres ambientals o a d’altres factors biòtics.
Les comunitats bentòniques són sovint un espai saturat, on les espècies competeixen fortament per l’espai. Probablement un dels millors exemples d’aquesta saturació de l’espai el proporcionen els poblaments de suspensívors de les zones poc il·luminades de parets verticals i d’entrades de coves submarines d’arreu del món. Esponges, cnidaris, poliquets, briozous i tunicats formen un recobriment policrom continu en aquests ambients. La seva bellesa no reflecteix, però, la situació real de lluita contínua per recobrir o evitar ser recobert, ni la duresa de la competència, que es produeix ja des dels estats larvaris, per ocupar els buits que es produeixen ocasionalment. Les armes d’aquests organismes sovint són subtils, però evolutivament molt “treballades” i mortíferes: determinats metabòlits secundaris que actuen inhibint el creixement d’altres espècies. Moltes vegades els compostos responsables d’aquesta mena de defensa química serveixen també per a evitar la depredació, la qual cosa afavoreix encara més la persistència i la competitivitat dels organismes que les tenen. Altres comunitats on la competència per l’espai és palesa són les zones rocoses amb algues, els herbeis de fanerògames marines, els esculls de corall i, en general, qualsevol ambient on la disponibilitat d’aliment no sigui limitant.
El parasitisme, la simbiosi, el comensalisme i el mutualisme
Com en el medi terrestre, el parasitisme és un tipus d’interacció biòtica molt àmplia que afecta potencialment tots els organismes d’un ecosistema. Aquest pot estar provocat per microorganismes, però també per diversos tipus d’invertebrats. L’actuació de determinats paràsits pot tenir una importància definitiva en l’estructuració de les comunitats. Tornant a l’exemple de les garotes i les comunitats d’algues, sembla que la recuperació dels boscos de laminàries de la costa W d’Amèrica del Nord després d’un estadi de dominància de garotes és possible mercès a una epizoòtia d’aquestes, resultant d’un període de temperatures anormalment elevades.
Un altre exemple de la importància de les malalties en l’estructuració de les comunitats han estat els canvis ocasionats en els esculls coral·lins del Carib després de la mort de les poblacions de la garota Diadema antillarum durant l’any 1983: el recobriment de les algues ha disminuït significativament, mentre que el dels coralls ha augmentat.
La simbiosi pot interpretar-se també com una relació biòtica important en determinar les característiques estructurals d’un ecosistema marí. Els casos més rellevants els tenim en les simbiosis entre algues unicel·lulars i determinats organismes heteròtrofs. Els coralls amb zooxantel·les simbionts en són l’exemple més paradigmàtic i important, ja que això els ha permès de colonitzar uns biòtops en principi reservats a les algues. Però no únicament en els esculls coral·lins hi ha exemples de simbiosi algues-animals, sinó que en les mars temperades també hi ha nombroses esponges (Ircinia, Cliona, Chondrilla) i zoantaris (Anemonia, Anthopleura) que s’aprofiten d’algues microscòpiques endosimbionts per a augmentar la seva producció en zones il·luminades i resultar competitives enfront de les macroalgues. Més curiós encara és la supervivència i l’activitat productiva de cloroplasts d’algues clorofícies (Acetabularia, Udotea, Halimeda) consumides per mol·luscs opistobranquis (Elysia, Bosellia) en les cèl·lules de la glàndula digestiva d’aquests animals.
Les relacions de comensalisme i mutualisme són molt abundants en el bentos. Els organismes filtradors com les esponges, els tunicats i determinats mol·luscs bivalves tenen sempre una gran varietat d’organismes comensals en el seu interior que engloben grups tan diversos com els protozous, amfípodes, copèpodes, lamel·libranquis, crancs, cucs, i fins i tot peixos. En els fons tous, les galeries construïdes pels filtradors són ocupades per un elevat nombre de comensals. Tenim exemples de mutualisme en els organismes netejadors, tan abundants en els peixos, o en la clàssica associació entre els xufancs (Dardanus arrosor) i determinades anemones (Calliactis parasitica).
Altres interaccions
Molt sovint, com passa també al medi terrestre, l’existència d’unes espècies, generalment de mida gran i dominants, permet l’existència d’altres espècies, o fins i tot comunitats senceres, a causa dels microhàbitats que creen. El bentos marí és ple d’exemples d’aquesta mena.
Els “boscos” d’algues erectes creen unes condicions vora el substrat que impedeixen el creixement d’altres algues que necessiten molta llum per a créixer (fotòfiles), però que afavoreixen, en canvi, les algues anomenades esciòfiles —amants de baixes irradiàncies— i molts organismes suspensívors que, sense aquestes grans algues, no podrien competir en condicions favorables amb altres petites algues fotòfiles que els recobririen.
L’anomenat coral·ligen de la mar Mediterrània és una estructura biogènica constituïda bàsicament pels esquelets d’algues carbonatades, pròpia de fondàries on arriba entre el 2 i el 0,05% de la llum superficial. La complexitat estructural d’aquest ambient, totalment foradat, permet la coexistència d’un elevadíssim nombre d’espècies, sovint més típiques de zones més profundes.
Als marges continentals de la costa atlàntica d’Amèrica del Nord, el peix Lopholatilus chamaeleonticeps excava forats de 4 m de diàmetre i 3 m de fondària en el sediment, que atreuen una gran varietat de crancs, peixos, equinoderms i altres organismes bentònics que generalment estan distribuïts de manera esparsa pel fons marí.
Com a darrer exemple de control o modificació del fons marí pels organismes que hi viuen, podem comentar la ja clàssica explicació de la regulació dels poblaments bentònics de fons tou per part dels suspensívors o dels sedimentívors. En general, els sediments fins de la plataforma continental tenen un alt contingut de matèria orgànica, les poblacions bacterianes hi són més nombroses i els organismes sedimentívors es veuen afavorits. L’activitat dels sedimentívors remou el sediment i això obtura els mecanismes de filtració dels suspensívors i dificulta la fixació de les seves larves. En canvi, els suspensívors abunden en els sediments més gruixuts, on hi ha menys matèria orgànica i els sedimentívors tenen menys menjar. Com que el substrat és estable, els suspensívors no tenen dificultats en establir-s’hi i impedeixen la introducció de larves de sedimentívors amb el continu filtratge de l’aigua. Veiem, doncs, que l’activitat de determinades estratègies facilita la seva pròpia continuïtat i dificulta possibles canvis.
El paisatge bentònic
La presència d’una espècie en un indret determinat no depèn només dels factors ambientals i de les interaccions biòtiques, sinó que és importantíssima la seva distribució geogràfica. L’especiació té lloc en un indret geogràfic determinat i sota unes condicions ambientals també més o menys concretes, però això no vol dir que les espècies només puguin viure en aquests ambients.
La història evolutiva
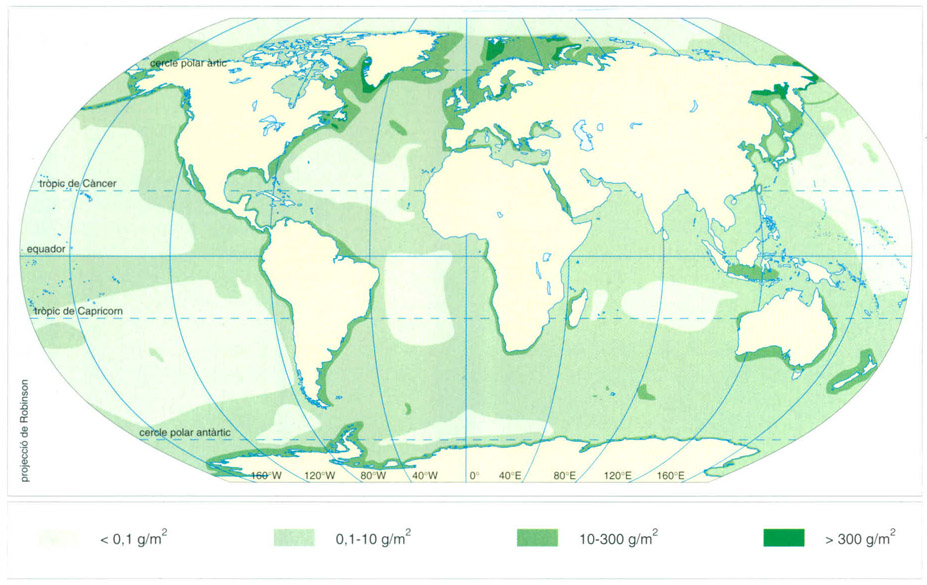
Distribució de la biomassa de la macrofauna als diversos oceans, expressada en grams de pes fresc per m2.
Editrònica, a partir d’un original de Zenkevitch, 1971
De fet, hi ha moltes espècies vicariants que ho són per raons històriques de l’aïllament de les seves poblacions, però que es desenvolupen en condicions ambientals molt similars. Aquest és el cas de moltes espècies de l’Atlàntic i del Pacífic, tant tropical com temperat, tant de nivells superficials com profunds: pertanyents a un mateix gènere, que ha sofert una radiació específica diferent en un i altre oceà, les espècies ocupen actualment hàbitats semblants i tenen una actuació semblant dins dels respectius ecosistemes.
Altres vegades, al contrari, les espècies tenen una distribució geogràfica àmplia, i tot i que sovint tenen una ecologia relativament coincident, de vegades l’hàbitat que ocupen és diferent a causa de les interaccions biòtiques que s’estableixen en els diferents llocs.
Finalment, en determinades ocasions, espècies que han evolucionat en una zona geogràfica determinada poden estar ben adaptades a les condicions d’altres zones i, en el cas de ser introduïdes, poden esdevenir les espècies dominants lluny de la seva àrea natural. Aquest és el cas de nombroses introduccions intencionades o accidentals provocades pels humans, com les de l’alga bruna Sargassum muticum, el creixement de la qual ha desplaçat alguns poblaments d’algues a les costes atlàntiques, o l’expansió de la cloïssa japonesa (Ruditapes philippinarum) a costa de les cloïsses autòctones a les costes sorrenques de l’Adriàtica septentrional.
Així, doncs, la presència o l’absència de determinades espècies en ambients determinats es pot deure a raons biogeogràfiques, i les comunitats són com són a causa, en part, de la seva història evolutiva. Això no obstant, i tot i que les espècies difereixen d’una mar a una altra, d’un oceà a l’altre, l’estructura de les comunitats i els processos que hi tenen lloc són semblants, sempre que les comunitats estiguin afectades per factors ambientals similars. És a dir, un poblament de laminàries, una comunitat de suspensívors sobre substrat rocós, un fons de sorra litoral o un fons fangós d’aigües profundes, tindran una composició específica diferent a les illes Britàniques, Sud-àfrica, les costes de Califòrnia o el Japó, però les estratègies tròfiques, els tipus d’organismes, la compartimentació de l’espai i els processos que la regulen, el flux energètic i la dinàmica de les poblacions, tenen una coherència prou gran dins de cada sistema per a permetre’n una explicació global. A l’inrevés, l’estudi d’un cas concret permet extrapolar alguns dels resultats obtinguts a sistemes semblants d’altres àrees geogràfiques.
La zonació batimètrica
La màxima variació en les condicions ambientals en la mar es produeix seguint l’eix vertical. La fondària, encara que pròpiament no és un paràmetre ambiental, porta associat el canvi de la majoria de factors que afecten decisivament els organismes, com la llum, l’hidrodinamisme, la temperatura, l’esdeveniment de fenòmens catastròfics i, en menor grau, els nutrients i la matèria orgànica. La variació en els valors d’aquests paràmetres porta associat un canvi en les relacions biòtiques dels organismes, i això provoca un canvi en les poblacions d’animals i plantes que va associat a la fondària. Als nostres ulls, aquest canvi es reflecteix principalment en la dominància de les espècies principals. Les zones de distinta dominància es distribueixen en franges o horitzons situats perpendicularment a l’eix batimètric. A aquest patró de distribució tan característic, més palès encara a la zona intermareal, i propi de qualsevol indret de la biosfera sotmès a un gradient ecològic fort, se’l coneix amb el nom de zonació.
Evidentment, els patrons de zonació varien geogràficament i d’acord amb els paràmetres biòtics i abiòtics. Així, és diferent la zonació que s’observa en una costa mediterrània de la que hi ha a les illes Britàniques, i ambdues són diferents de les que es donen a les costes pacífiques d’Amèrica del Nord o al S d’Austràlia. També varia aquesta zonació si el fons és dur o tou, si està molt o poc exposat a les onades, si el lloc és més o menys ric en nutrients, o si l’acció de predació per part de les garotes (per posar un exemple de factor biòtic) és important o no.
Contràriament al que passa a la zona intermareal, de la qual s’han fet nombrosos estudis, de la zona infralitoral n’hi ha pocs que posin de manifest les causes reals d’un model de zonació determinat. Al golf de Maine, a l’Atlàntic nord-occidental, els fons rocosos estan dominats per dues laminàries (Laminaria digitata i L. saccharina) a les fondàries compreses entre 4 i 8 m, i per una determinada espècie de musclo (Modiolus modiolus) a les compreses entre 11 i 18 m. Els musclos que creixen ocasionalment en aigües superficials són recoberts per les laminàries i són arrencats, quan hi ha temporal, a causa de la força que exerceixen les onades directament sobre les laminàries i indirectament sobre els musclos. Els espais buits que deixen els musclos arrencats són ràpidament recolonitzats per les laminàries, però no pels musclos, que tenen un creixement més lent. Així, doncs, els musclos no poden ser dominants a dalt perquè són arrencats pels temporals i perquè les laminàries són competitivament superiors en l’ocupació de l’espai. Però, d’altra banda, la llum disminueix en fondària i amb ella el creixement de les dues Laminaria; per sota dels 11 m, aquest queda tan disminuït que no pot compensar el consum que fan les garotes (Strongylocentrotus droebachiensis) d’aquests feòfits. Les garotes són més abundants a la comunitat de musclos que a la de laminàries per tal com hi troben més bon recer per a resistir el fort hidrodinamisme i per a escapar-se de la depredació. L’eliminació experimental de les garotes a la comunitat de musclos provoca un elevat reclutament de laminàries, la qual cosa fa augmentar extraordinàriament la mortalitat de musclos per arrencament. Així, doncs, en fondària, la interacció musclos-garotes facilita la coexistència d’ambdues espècies i impedeix la colonització per part de Laminaria.
En aquest cas es veu com llum, hidrodinamisme, pertorbacions temporals, herbivorisme i protecció de la depredació intervenen per produir un model de zonació, en un excel·lent exemple de la interacció de diferents factors. La relativa complexitat del model, tot i tractar-se d’unes comunitats molt senzilles, i la complicació d’experimentar en el món subaquàtic, explica la raresa d’estudis semblants, però també ens fa veure la importància de la integració dels factors ambientals, de les interaccions biòtiques, dels cicles de vida i de creixement, per a comprendre els motius de la distribució de les espècies en el bentos.
Els sistemes i els estatges
Atenent a les característiques dels ambients i als organismes que hi viuen, hom distingeix tot un seguit d’estatges o zones, distribuïdes d’acord amb la fondària, que són vàlides per a qualsevol indret. En una primera gran divisió se separen el sistema litoral del sistema profund. El sistema litoral inclou els fons de la plataforma continental, la qual abasta aproximadament des de la cota 0 de la línia de costa fins als 200 m de fondària. A les mars d’aigües transparents, les formes de vida fotoautòtrofes són viables en tota l’extensió de la plataforma continental, però en mars d’aigües tèrboles, productives, la distribució dels vegetals té una amplitud inferior. El sistema profund s’inicia en el talús continental i en ell no és possible la fotosíntesi. Llevat dels poblaments de les fumaroles de les dorsals oceàniques, tota l’energia prové de la matèria produïda a la zona fòtica. El bentos profund és, doncs, un sistema totalment heterotròfic, la dinàmica del qual és regida per processos que passen molt més amunt.
El sistema litoral comença al límit inferior de les marees i acaba sovint amb la total desaparició de les algues. Per això rep també el nom de sistema fital. Clàssicament es divideix en dues zones o estatges: l’infralitoral i el circalitoral. L’amplitud de tots dos estatges és condicionada per la transparència de l’aigua i es relaciona amb la quantitat de llum que hi arriba. La seva delimitació sobre fons rocosos és regida per la dominància d’algues fotòfiles (zona infralitoral) o algues esciòfiles i organismes suspensívors (zona circalitoral). El canvi entre infralitoral i circalitoral és imprecís, però se situa aproximadament allà on arriba ja únicament entre l’1 i el 5% de la llum superficial.
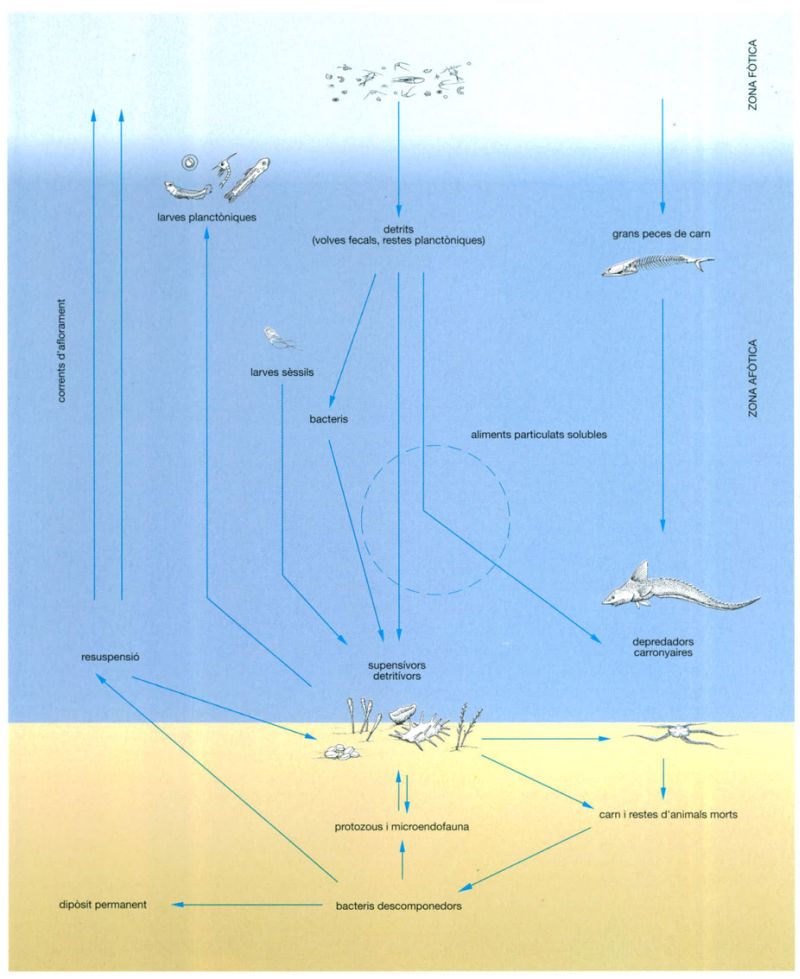
Relacions tròfiques entre els organismes que habiten els ambients bentònics abissals, és a dir els fons de les grans fondàries oceàniques.
Jordi Corbera, a partir de fonts diverses
El sistema profund es divideix alhora en tres grans zones o estatges: batial, abissal i hadal. L’estatge batial s’inicia en acabar-se l’estatge circalitoral i comprèn, bàsicament, les comunitats del talús continental i de les seves immediacions, fins a aproximadament els 3 000 m. La zona abissal ocupa la zona central dels oceans, fondàries compreses entre els 3 000 i els 6 000 m, mentre que la zona hadal comprèn només les profundes fosses oceàniques. Les diferències entre aquestes zones són principalment de tipus tròfic, ja que hi ha una disminució d’aliment en fondària, la qual cosa repercuteix en una menor biomassa i densitat d’individus.
Alguns casos exemplars
Sobre fons rocosos litorals, i exceptuant la zonació que hom observa en els esculls coral·lins on els coralls tenen una gran importància en la zona ben il·luminada, el patró de zonació que s’estableix arreu és fins a cert punt semblant. Com a exemples de zonació podem comentar el que es troba al sistema litoral sobre substrat rocós en tres indrets ben diferents però igualment ben estudiats: la Mediterrània nord-occidental, les costes de la Bretanya francesa, a l’Atlàntic nord-oriental, i les de la regió californiana de Monterey, al Pacífic nord-oriental.
A la zona superficial, als llocs moderadament batuts, immediatament per sota de les franges d’algues intermareals, s’instal·la una comunitat de distribució batimètrica reduïda, dominada per una o poques espècies de feòfits, exclusiva d’aquest nivell (Cystoseira mediterranea o C. stricta a la Mediterrània, Laminaria digitata i Alaria esculenta a les costes de Bretanya, Egregia menziesii a les de Monterey). Una mica més avall, la comunitat es diversifica, amb el desplaçament de les espècies superficials i l’entrada de moltes noves espècies d’algues i d’animals. A la Mediterrània, per exemple, hi ha una zona dominada per altres espècies de Cystoseira, per dictiotals (un altre ordre de feòfits) i per Halopteris scoparia (un feòfit de l’ordre dels esfacelarials). A la Bretanya el relleu l’agafen Saccorhiza polyschides i Laminaria hyperborea, mentre que a Monterey s’hi desenvolupen espectaculars “boscos” de Macrocystis pyrifera i altres laminarials (Laminaria setchellii, L. farlowii, Dictyoneurum californicum, Eisenia arborea, i altres).
La part inferior de l’estatge infralitoral té una composició bastant diferent. A la Mediterrània apareixen un nombre molt elevat d’espècies, algunes de les quals constitueixen fàcies determinades (Codium vermilara, Dictyopteris membranacea, Cystoseira). A la Bretanya, Laminaria hyperborea fa poblaments poc densos on tenen cabuda altres algues de mida més petita (Dictyota dichotoma, Dictyopteris membranacea, Cryptopleura ramosa). A Monterey continua havent-hi Macrocystis, però Nereocystis luetkeana esdevé sovint l’espècie dominant; en l’estrat inferior abunden les laminarials Pterygophora californica i Costaria costata.
A la zona circalitoral el gradient de llum està esmorteït i no hi ha una zonació tan clara com a l’infralitoral; tot i que es manté un patró de zonació, aquest està desdibuixat per la dominància a taques de determinades espècies. L’estatge circalitoral és marcat per un progressiu desplaçament de les algues pels animals. A la Mediterrània nord-occidental, el gran desenvolupament en gruix de les algues coral·linàcies incrustants dóna lloc a una comunitat molt particular, el coral·ligen, el qual té un recobriment d’algues esciòfiles (Lithophyllum, Mesophyllum, Halimeda, Peyssonnelia) a l’estatge circalitoral superior o és dominat per poblaments de suspensívors (gorgònies com Paramuricea i Eunicella, corall vermell, esponges) a l’estatge circalitoral inferior. La diversitat hi és molt elevada. A les costes de Bretanya i a les de Monterey, el circalitoral superior és constituït per poblaments mixtos d’algues coral·linàcies (Mesophyllum, Lithothamnium, Lithophyllum), rodofícies laminars (Rhodymenia, Fryeella, Phycodrys) i animals suspensívors (esponges com Axinella, alcionacis com Alcyonium o Paralcyonium), però sense constituir una comunitat tan complexa com a la Mediterrània; al circalitoral inferior, en canvi, la dominància animal (coralls blancs com Dendrophyllia, gorgònies, esponges, antipataris —és a dir coralls espinosos—, etc.) és pràcticament total.
En els fons tous, l’extinció de la llum cap al fons té poca importància, ja que les algues no hi tenen cap paper estructural i la zonació batimètrica esdevé confusa o desapareix. Paràmetres que varien associats a la fondària, com l’hidrodinamisme o la quantitat de matèria orgànica, poden fer-ho, tant o més, en sentit horitzontal, i els corrents marins o la història geològica poden canviar les característiques del sediment més intensament que el que pugui fer-ho la fondària. Tot i això, si suposem una homogeneïtat a nivell horitzontal, la fondària porta associada un decreixement en la mida mitjana dels grans de sediment de tal manera que al llarg de la plataforma continental s’observa un gradient des de sorra gruixuda a llim en augmentar la fondària, sovint acompanyat d’un enriquiment de matèria orgànica ocasionat pel decreixent hidrodinamisme i la consegüent manca de rentatge.
La diversitat de tipus de fons marins que hom pot trobar és immensa, amb la particularitat que, essent la màxima variabilitat de condicions ambientals al llarg de l’eix vertical, en una mateixa localitat geogràfica es poden trobar moltes comunitats o zones diferents. Les causes d’aquesta zonació han estat atribuïdes a la disminució de la matèria orgànica en fondària, al canvi en el tipus de sediments i als corrents i a l’efecte de la pressió sobre els sistemes enzimàtics. Aquesta diversitat paisatgística és superior a la del medi terrestre, on la variació en els paràmetres ambientals es produeix a una escala diferent.