El cicle de la matèria
Si l’anatomia de la selva plujosa és marcada per l’elevada diversitat que presenta, la seva fisiologia es caracteritza pel seu enorme dinamisme. La selva plujosa és una màquina metabòlica trepidant que fa ostentació de la seva puixança: tot o quasi tot és a la vista, res o ben poca cosa passa sota terra. Així com els sòls dels boscos temperats són un contenidor de nutrients i un reactor que governa la fisiologia forestal, manifestada a penes en una part aèria barroca però nutritivament modesta, els sòls de la selva plujosa gairebé només són la peanya de l’ecosistema. La selva plujosa viu del terra cap amunt, i és per això que sol tenir tantes dificultats, si és destruïda, a autoregenerar-se.
El capteniment de la biomassa
La competència per la llum comporta en els sistemes terrestres un estirament de la vegetació per atansar les fulles a la font lumínica. Aquest increment d’alçada implica l’allargament del sistema de transport vertical d’aigua i nutrients i, al capdavall, representa un retard en el cicle material del sistema, perquè les estructures que s’edifiquen han de ser resistents a la destrucció. De tot plegat resulta un augment de la biomassa de fusta que és paral·lel a l’increment de l’àrea de fulles per unitat de superfície. D’entre els sistemes terrestres, és a la selva plujosa, pràcticament lliure de limitacions de llum, d’aigua o de temperatura, on s’aconsegueixen les alçades màximes i la biomassa acumulada és més gran.
D’altra banda, a ningú no passa per alt que els vegetals de la selva, de fullatge esponerós i aparent perennitat, al cap d’un temps han d’haver produït noves fulles i generat un gruix de fulles velles, inútils per a la fotosíntesi. El verd constant, divers però immutable, que copsa el visitant ofereix claps: arbres amb el fullam marcint-se i arbres nus apareixen voltats de vegetació exultant, rutilant en la seva vigoria. I encara més: un mateix peu pot presentar, fet sorprenent en altres biomes, branques amb fulles marcides al costat de branques esfullades i altres encara de ben actives. La monotonia del clima n’és la causa. L’absència d’un període marcat d’eixut o d’una estació freda que actuïn de referents per a les plantes fa que cada individu floreixi i grani obeint un ritme propi, en algun cas ajustat a les lleus variacions que el clima equatorial pot oferir. Fins i tot alguns arbres efectivament caducifolis o semicaducifolis que es fan a les selves plujoses poden perdre totalment la fulla, però ho fan sense seguir pautes estacionals.
La selva, efectivament, entapissa, per bé que no amb l’espectacularitat dels boscos caducifolis, implacablement el sòl amb una pluja de fulles. Flors, fruits i branquillons (troncs i branques d’una manera més esparsa) s’afegeixen a aquest al·luvió incessant. Tot aquest material, inapte (llevat dels fruits) per a les funcions que acomplia, conté moltes molècules orgàniques, que conserven encara força energia en els seus enllaços. Són l’estoig, així mateix, de molècules inorgàniques i de nutrients indispensables per al conjunt dels organismes del bosc. Però el sòl de la selva no acull només residus vegetals: ossos, pèl, escates i, sobretot, excrecions constitueixen la col·laboració de la fauna a la configuració edàfica. Tot i així, la virosta, d’origen vegetal, n’és amb escreix l’agent més important.
Els processos mineralitzadors
D’un any per l’altre el volum de restes que fan cap al sòl és força semblant (la màxima diferència observada entre un mínim i un màxim anuals no ha arribat a un 12%). Tota aquesta matèria morta que el bosc fabrica podria romandre al sòl inalterada fins a constituir veritables muntanyes de residus, però una munió d’organismes (col·lèmbols, tèrmits i altres insectes, milpeus, àcars, fongs, bacteris, etc.) s’encarrega de descompondre-la fins a convertir-la a diòxid de carboni i sals minerals.
L’acció descomponedora difon diòxid de carboni a l’atmosfera. Per estimar l’activitat descomponedora global del sòl un mètode força senzill és avaluar la quantitat d’aquest gas que el sòl exhala. Si tenim en compte que la temperatura i la humitat són força constants, la respiració del sòl no deu fluctuar gaire en el transcurs de l’any. Convé fer les mesures sovint. En una selva baixa de Java hom ha mesurat una taxa de respiració de 224 mg de CO2 per m 2 i hora, que correspon a 10,7 t per hectàrea i any. Si tenim en compte que el ritme d’acumulació de virosta en aquests boscos tropicals és entorn de 10,9 t per hectàrea i any, és clar que el balanç anual en aquesta selva és un increment molt lleu de matèria orgànica al sòl. Però la taxa de descomposició no és sempre tan alta. En una selva de l’Amazones, situada aigües avall de l’aiguabarreig dels rius Branco i Negro, s’ha mesurat una taxa de 5 t per hectàrea i any. Altres informacions mostren que la taxa anual de descomposició de la virosta a les selves s’acosta al 80%. Aquest fet, afegit al ritme lent de polimerització i d’humificació, explicaria la minsa quantitat d’humus que s’hi observa.
Els agents de la descomposició són invertebrats i microorganismes sapròfags. En alimentar-se, a més d’obtenir energia, trenquen progressivament les molècules orgàniques fins a aconseguir-ne la mineralització. El procés generalment l’inicien animals invertebrats, sobretot àcars i col·lèmbols, els quals deixen la matèria a punt per a l’actuació de fongs i bacteris. Les possibilitats químiques de degradació que aquests darrers posseeixen són molt grans. Són capaços d’escindir la cel·lulosa, la molècula que constitueix la paret de les cèl·lules vegetals i, per tant, la fibra més abundant a les fulles. Algunes espècies de fongs, encara, disposen d’enzims capaços de destruir la lignina i d’utilitzar-ne l’energia. Aquests fongs són essencials en els boscos ja que la lignina, una molècula especialment invulnerable, és la fibra de la fusta.
Encara dins dels organismes descomponedors convé fer esment dels tèrmits que, en alguns casos, arriben a constituir el 70% del pes dels invertebrats de la fullaraca. A les selves plujoses del Zaire i de la Costa d’Ivori hom ha comptat unes 870 colònies de tèrmits per hectàrea (una biomassa de 16 t per hectàrea), capaces de consumir un terç de les restes de matèria orgànica. N’hi ha que disposen de protozous o de bacteris simbiòtics en el sistema digestiu que els permeten fer aquesta digestió; d’altres fan cultius de fongs (Termitomyces, Xylaria) sobre amuntegaments de boletes de fusta mastegada, empassada i defecada i més tard s’alimenten d’aquest material un cop transformat pels fongs que hi creixen; les nimfes són alimentades amb els conidiòfors d’aquests fongs, uns filaments portadors d’espores asexuades que intervenen en el cicle reproductor del fong, que cal suposar que són particularment nutritius.
Els cucs de terra, que excaven galeries al sòl, intervenen també en els processos de transformació de les restes, bé sigui directament, en la mineralització del nitrogen, bé indirectament, afavorint l’oxigenació del sòl. Efectivament, la transformació de la matèria orgànica fins a convertir-la en inorgànica (utilitzable pels vegetals) exigeix el consum d’oxigen; els canals que els cucs produeixen permeten l’airejament del sòl i, per tant, la difusió de l’oxigen.
De tota manera, convé no oblidar que, al capdavall, la part més important de la descomposició depèn dels microorganismes (fongs i bacteris, en conjunt anomenats microflora) i no dels invertebrats (microfauna). Si hom suprimeix la microfauna d’un sòl la taxa de descomposició (mesurada en pèrdua de pes) continua essent significativa. A la selva intertropical la importància dels fongs, que actuen sobretot en les primeres fases de descomposició, és més gran que no pas al bosc temperat.
La fusta i la virosta, factors de control
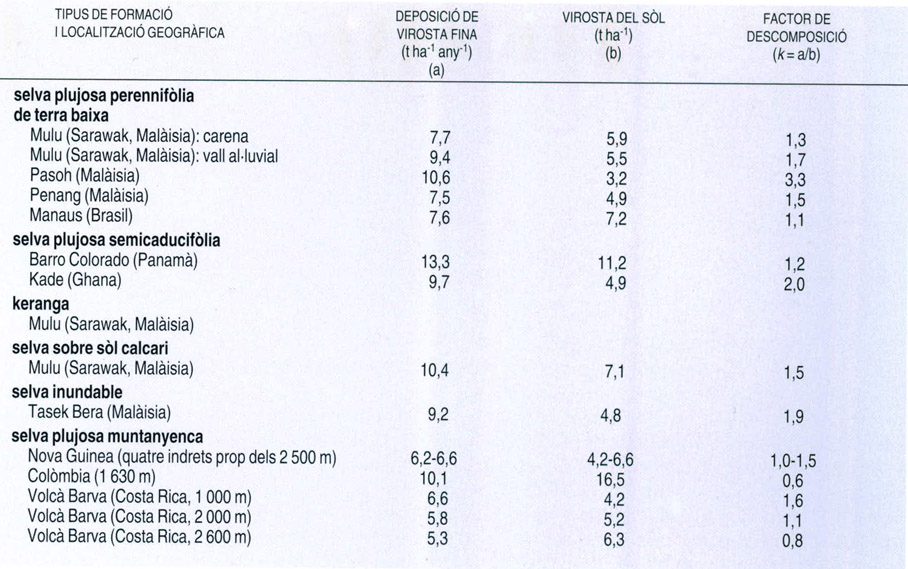
Dades de caiguda, quantitat mitjana al sòl i taxes de descomposició de la virosta fina a la selva plujosa de diferents estacions de mostratge. Entre les diverses àrees recollides hi ha tota mena de selva: des de la selva plujosa de terra baixa a la selva semicaducifòlia, bosc inundable indonèsic de les riberes fluvials i bosc nebulós.
Dades elaborades per T.C. Whitmore a “Ecosystems of the World”
El volum de virosta consumida pels descomponedors determina les reserves d’elements i molècules inorgàniques que romandran atrapats entre les fulles seques, fora de l’abast de les arrels. A la selva equatorial, on la humitat i la temperatura són altes, la fracció de nutrients retinguts a la virosta és menor que en els altres components. Hi ha casos excepcionals, però: a Surinam, en una pluviïsilva que ha acumulat un gruix d’un metre de virosta, la fracció més important ben segur que no és a la vegetació! Excepcions a banda, hom considera que al bosc tropical humit els nutrients poques vegades romanen més d’un any a la virosta (les mesures que s’han fet per al fòsfor, el calci, el magnesi i el potassi van dels dos mesos i mig als catorze mesos). Tal com veurem més endavant, és aquesta rapidesa en els cicles dels elements allò que fa possible la fertilitat manifesta de la selva plujosa.
És un fet obvi que la velocitat de descomposició de les fulles és molt més gran que la de la fusta. Si en una selva primària de Guatemala calen de 150 a 300 dies perquè es descomponguin completament les fulles, calen, en el mateix bosc, uns quants anys per a eliminar del tot les restes de fusta. La velocitat dependrà de la mida dels fragments: els més petits, és clar, seran els primers a desaparèixer. A la Costa d’Ivori, hi ha branques d’1 a 2 cm de diàmetre que perden en mig any un terç del seu pes sec (el pes que resulta després de descomptar l’aigua). A Nigèria, en una selva una mica més humida, blocs de fusta de 25 cm de diàmetre poden trigar 7 mesos a perdre un quart del seu pes sec. Això no obstant, a la jungla del Panamà un arbre sencer es descompon en menys de deu anys.
L’aportació de la fusta a la matèria orgànica del sòl, si bé és menor que la de les fulles, és, tot i així, molt important. De les prop 20 t per hectàrea i any de matèria orgànica que van a parar al sòl en una selva intertropical, gairebé un terç corresponen a fusta morta. La descomposició de la fusta s’esdevé principalment a la superfície del sòl. A El Verde (Puerto Rico) al cap de l’any es generen uns 600 g per metre quadrat de restes llenyoses, dels quals 55 g corresponen a troncs i branques, 145 g a les rabasses i 400 g a les branques fines. A la selva de Pasoh (Malàisia) van al sòl cada any 330 g per metre quadrat de branques de diàmetre superior a 10 cm.
D’altra banda, el ritme de descomposició depèn d’una colla de factors: en un mateix indret, en anys diferents, la descomposició completa de les fulles pot trigar de 2,5 fins a 6 mesos. No cal dir que també varia segons la regió i segons l’espècie de què es tracti: les fulles de palmeres, per exemple, són més resistents (1/5 de temps més, aproximadament, que les d’altres espècies). A El Verde hom ha observat que la velocitat de descomposició de les fulles d’una espècie pot arribar a ser 2,5 vegades superior a la d’una altra. Altres factors que influeixen en aquesta velocitat són l’edat de les fulles, que hi influeix negativament, i la humitat que, en el cas que hi hagi fluctuació anual, l’accelera. La relació entre precipitacions i velocitat de descomposició podria explicar les variacions interanuals. La pluja intervé també indirectament, a través de la fauna i dels microorganismes.
L’administració dels nutrients
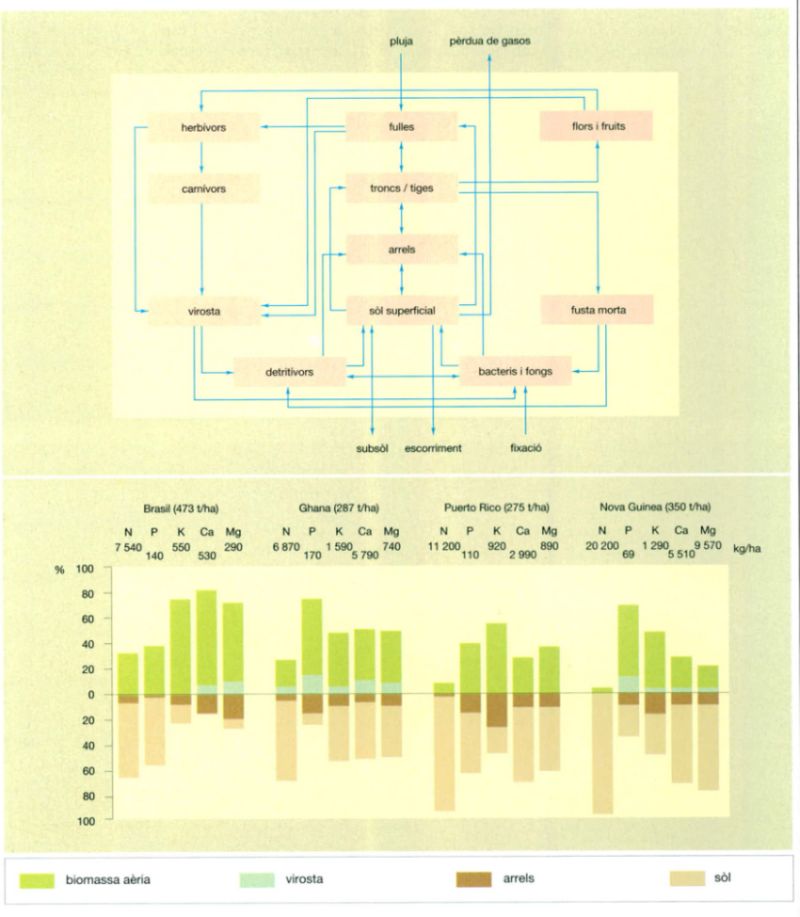
El cicle dels nutrients a la selva es pot esquematitzar d’una manera general segons el gràfic superior. L’inferior presenta el valor, en kg per ha, dels principals nutrients (nitrogen, fòsfor, potassi, calci i magnesi) corresponents a quatre estacions de mostratge de selves de diferents parts del món, per a mostrar la distribució dels nutrients inorgànics per sobre i per sota de la capa del sòl. S’hi indiquen també els valors de biomassa (entre parèntesis). És bo de fer notar que la idea tan estesa que la major part dels nutrients de la selva es troba en la biomassa no sempre és certa. Cal fer notar també que el sòl és una capa d’entre 0 i 0,3 m en tots els casos excepte el de Puerto Rico, que és de 0 a 0,25 m.
Jordi Corbera, a partir de fonts diverses]
El resultat final dels processos de descomposició de la matèria orgànica és el retorn al sòl dels nutrients (fosfats, nitrats, potassi, etc.) en condicions de ser novament absorbits per les arrels. Les condicions dels sòls tropicals, on, dins la fracció argilosa, predomina la caolinita (amb poca capacitat per a adsorbir cations), i on la freqüència i la quantitat de precipitacions són tan elevades, fan indispensable la ràpida recuperació dels nutrients per part dels organismes abans que s’esdevingui la lixiviació.
L’absorció radicular
Això explicaria l’extraordinària abundància d’arrels somes (en un bosc de la Costa d’Ivori entre el 20% i el 50% de les arrels es troben en els 10 cm primers de fondària) que s’estenen i s’imbriquen en oberta competència, a l’encalç dels escassos nutrients lliures. Molt sovint, a les selves edificades en sòls especialment pobres, el terra s’ofereix torturat per centenars d’arrels que serpentegen retortes i s’agleven, fixades per la lignificació, en una febrosa recerca per una terra gairebé exhausta.
L’absorció que fan les arrels tanca un cicle, el dels nutrients, que pot presentar diverses variants. S’esdevé un cicle curt quan la descomposició afecta restes d’origen vegetal: en ser consumides, els nutrients són alliberats al sòl i ràpidament reincorporats a la biomassa forestal a través de les cabelleres radicals. Si els vegetals són consumits per herbívors (o les seves restes per sapròfags) i aquests, al seu torn, per altres animals, s’inicia una cadena en la transferència dels nutrients que, alhora que en pot retardar la reincorporació, determina un cicle complex. La complexitat encara augmenta si hi intervenen paràsits.
Als arbres dels boscos tropicals és molt freqüent la presència de micorizes. Tal com el nom indica es tracta d’una relació de simbiosi entre un fong i les arrels d’un vegetal: mentre la xarxa d’hifes facilita la captació de les sals al vegetal, aquest forneix al fong compostos orgànics. Hom no descarta que en alguns casos es pugui establir un flux directe de nutrients, sense mineralització intermèdia, entre la virosta i les arrels, per via de l’agent descomponedor: les micorizes del fong. Aquesta canalització directa dels nutrients adquireix una transcendència cabdal en aquests ambients on, com acabem de veure, les reserves del sòl són ben migrades.
La circulació dels elements nutritius
A les selves intertropicals són els mateixos organismes els qui acabalen els nutrients. Lluny de constituir reserves estàtiques s’hi esdevé una cinètica molt important. D’una manera global podríem dir que els nutrients tenen un temps mitjà de permanència d’uns quinze anys a la vegetació i un període inferior a un any a la virosta. Convé, però, fer-hi precisions. En un estudi fet en un bosc del Panamà per tal d’estimar el temps de renovació de fòsfor, calci, magnesi i potassi, els resultats eren temps de permanència que anaven dels tres mesos per als fruits, uns dotze mesos per a les fulles, set mesos de mitjana per a la virosta i de dotze a quinze anys per a la fusta. Les dades obtingudes a Puerto Rico i a Ghana són força coincidents amb les del Panamà. Els troncs i les branques, doncs, constitueixen les principals reserves. Els segueixen en importància les arrels i, a més distància, les fulles, les flors i els fruits. Dins l’estructura del bosc la màxima acumulació correspon a les zones inferiors.
D’entre els diversos nutrients, el nitrogen té un capteniment i unes característiques particulars. A més de ser fonamental per als vegetals, hi ha organismes capaços de fixar-lo a partir de l’atmosfera i posar-lo a l’abast de les arrels. Així, hi ha lleguminoses tropicals (algunes de les quals, no gaires, són arbres) que fan simbiosi amb Rhizobium o amb altres fixadors. Altres plantes tropicals no lleguminoses dels gèneres Dioscorea, Cycas i algunes rubiàcies disposen de nòduls fixadors de nitrogen a les arrels, les tiges o les fulles. Hi ha encara bacteris de vida lliure que també són fixadors i que representen una font important de nitrogen per a l’ecosistema (entre 4 i 24 g de N per metre quadrat i any). Pel que fa a la mineralització, el nitrogen es perd molt aviat en el procés de descomposició de les fulles i retorna al sòl.
Les pèrdues de fertilitat
Com a resultat dels circuits dels elements en la selva plujosa, les aigües de les fonts, allà on la selva és encara ben constituïda, solen ser molt pobres de sals. Les lleugeres pèrdues de nutrients que hom detecta podrien ser restituïdes pels ions de l’aigua de la pluja. A la xarxa fluvial de Mondo, a Java, s’ha observat que, en el residu soluble, només es perdia àcid silícic, i encara de manera força variable segons el cabal del riu: unes 0,4 tones per hectàrea i any.
En oposició a aquesta situació, la tala i la crema del bosc comporten grans pèrdues, sigui per transport dels nutrients amb la fusta, sigui per rentatge de les cendres a les àrees incendiades. Una zona talada i destinada reiteradament al cultiu, esdevé, després d’uns quants cicles, extremament pobra, capaç només de suportar un poblament herbaci (Imperata o Gleichenia a Malàisia, Pteridium al Brasil). Tot plegat fa que només les zones amb aportacions alienes de nutrients (dipòsits al·luvials, adob amb cendres volcàniques noves) o amb característiques particulars (sòls volcànics recents) permetin la utilització continuada de les terres després d’haver-ne foragitat el bosc.
Metabolisme exaltat, productivitat modesta
És molt estesa la concepció que la selva és un bosc que anualment experimenta un gran increment de volum. Aquesta convicció, que és només una veritat a mitges, pot molt ben ser que provingui de l’observació de la vegetació des dels cursos d’aigua, des de les pistes que travessen la selva o des de les clarianes; és a dir, des de les vores de la selva plujosa estant, allà on la llum entra generosa i on les espècies pioneres, heliòfiles molt sovint, prosperen amb abundància i s’estenen pertot amb avidesa. Certament, però, els arbres grans (de quaranta anys o més) que conformen el bosc no destaquen pas pel seu creixement excepcional, ja que amb prou feines sobrepassen els tres mil·límetres anuals d’increment del diàmetre de la soca. Més que no la producció, allò que fa del bosc tropical una comunitat tan notable és la gran biomassa que acumula.
Posats a l’altre extrem, l’alt grau de complexitat i estructutura que mostra no ens l’han de fer veure com un bosc estàtic, aturat en el seu envitricollament, ja que cada 40 o 100 anys tota la massa mor i es renova. Els canvis que s’hi produeixen s’esdevenen a diversos nivells, d’espècie, de comunitat o del bosc sencer, i, en una escala més breu de temps, caldria afegir-hi els canvis fenològics (producció de fulles, flors i fruits) que són característics de cada individu.
La dinàmica de la selva i la productivitat
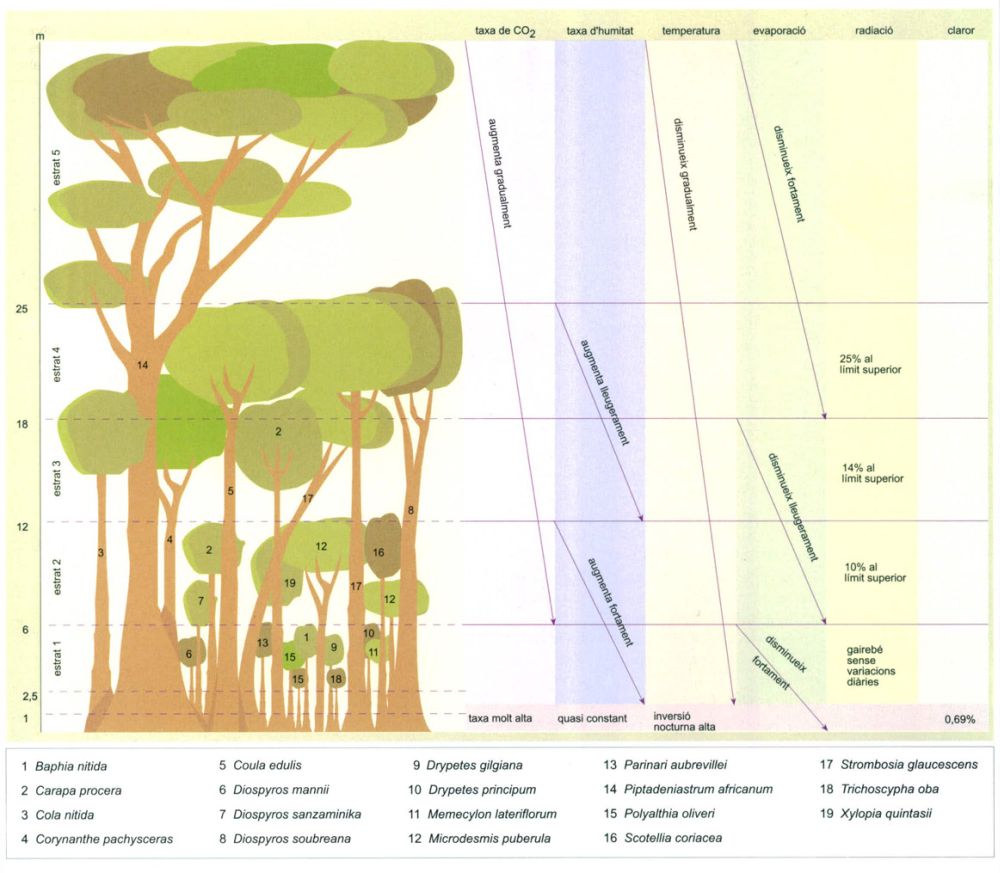
Variació de diversos paràmetres ecològics en els diferents estrats de vegetació a la selva plujosa tropical. L’esquema correspon a una selva plujosa perennifòlia de la reserva del bosc Tai, a la Costa d’Ivori, i s’hi indiquen les espècies principals que formen l’estructura dominant en cada estrat. S’ha omès la vegetació que hi ha per sota dels 2,5 m.
Editrònica, a partir d’un original de Bourgeron i Guillaumet, 1981
Aquest model de bosc dinàmic, més mudable del que a primer cop d’ull pot semblar, tampoc no és homogeni en l’espai i pot oferir alhora diverses fesomies que corresponen a estadis diversos en la consecució d’una hipotètica fase clímax, culminació de la seva maduresa. D’una manera esquemàtica hom distingeix tres etapes: la fase oberta, posterior a la caiguda d’arbres o a una intervenció humana discreta, on els arbres encara no han atès els tres metres d’alçària, la fase de regeneració, durant la qual els arbres creixen molt en gruix i en longitud, i la fase madura, la darrera: l’increment d’alçada disminueix fins a aturar-se i l’augment del diàmetre del tronc es manté, per bé que molt atenuat, ratllant l’asímptota. Aquesta fase última, si tenim en compte la seva longevitat, és la que ocupa una àrea més extensa de la superfície forestal. La fase oberta, per contra, afectaria només una desena part del bosc.
La producció i la productivitat
La producció de matèria orgànica de la selva plujosa és molt diferent segons que ens trobem en una o altra de les fases. Aquí, inicialment, ens fixarem en la producció del bosc madur. La captació d’energia lumínica que fan els vegetals (Vegeu també La fotosíntesi biosfèrica) s’utilitza en la síntesi de molècules orgàniques destinades bé a subvenir els requeriments del propi manteniment, bé a augmentar la biomassa (de fusta o de reserves). Hom anomena producció primària bruta el conjunt de la matèria orgànica sintetitzada i producció primària neta la que resulta de restar d’aquella la fracció esmerçada en la respiració i que és la que es tradueix en un augment de biomassa.
A les regions intertropicals, la radiació és màxima: reben el doble de l’energia que arriba a la latitud 60°N o S. S’esperaria que la producció de biomassa per unitat de superfície hi fos màxima. Doncs bé, en una selva de la Costa d’Ivori la producció neta anual és de 9 t /ha, una xifra no gaire més alta que la d’una fageda de Dinamarca. Algunes mesures fetes en altres tipus de bosc tropical donen resultats poc superiors a aquest. L’explicació es pot trobar en l’elevadíssima taxa de respiració de la vegetació tropical; s’estima que en els boscos madurs la respiració pot arribar a ser del 70% de la producció primària bruta! El residu destinat a la síntesi de cel·lulosa i proteïnes és, comparativament, ben migrat.
Les escasses estimacions de la producció primària bruta de què hom disposa per als boscos tropicals donen un valor mitjà de 86 t per hectàrea i any. Pel que fa a estimacions de la producció primària neta, hom no disposa de gaires més dades, ja que molts estudis tenen en compte només les espècies susceptibles d’explotació comercial. Tot i així, s’ha proposat un valor mitjà de 20 t per hectàrea i any, amb valors que oscil·larien entre les 9 i les 32 t per hectàrea i any (entre les 25 i les 30 t per hectàrea i any segons altres fonts). Els valors més alts corresponen a les àrees de pluviositat més elevada i més constant, i també caldria tenir en compte la fertilitat del sòl. En els boscos explotats, on predominen els arbres de menys de 20 anys, aquests valors poden ser encara més elevats.
Per oferir una informació més completa de la dinàmica productiva, resulta molt interessant relacionar la producció neta del bosc amb la seva biomassa. S’obté així un índex de l’eficiència del sistema: la productivitat. És semblant a l’establiment d’una taxa d’interès, resultat de comparar els rendiments amb el capital que els origina. En una població d’algues unicel·lulars que es divideixen cada 24 hores, la productivitat diària és del 100%. La productivitat de la vegetació terrestre, que fa una gran inversió en estructures de sosteniment i distribució, pot variar dins d’un interval molt ampli, del 2 al 100% anual, sempre molt per sota dels sistemes aquàtics. Això significa una taxa de renovació que variaria dels 50 anys a 1 any. El bosc tropical, doncs, se situaria, malgrat que posseeix una taxa de síntesi molt elevada, dintre dels sistemes amb una productivitat menor.
D’altra banda, el creixement en gruix i en alçària dels arbres de la selva primària és menor que el d’alguns arbres (pollancres, eucaliptus, alguns pins) de zones temperades. Per contra, la producció total de matèria orgànica (producció primària bruta) d’un arbre tropical duplica, i fins triplica, la d’un arbre de les regions temperades. La biomassa dels boscos tropicals, expressada en pes sec, oscilla entre 300 i 400 t per hectàrea (entre 350 i 550 t per hectàrea segons alguns autors), el 90% de la qual correspon a branques i troncs, les estructures més perennes. La fracció de fulles, el material més apte per a insectes i altres herbívors, representa ben poc del total: només el 2%.
La distribució de la productivitat
Per a fer un estudi complet de la producció caldria mesurar l’augment anual de pes de les fulles, de fusta i d’arrels a més de tenir en compte el consum dels animals i les pèrdues d’estructures mortes. Només en el cas de la fusta, i per raons òbvies d’explotació forestal, hom disposa de tècniques per a avaluar-ne l’increment; cal fer mesures periòdiques del diàmetre (i en ocasions també del perímetre) de la soca a l’alçada del pit de l’observador, és a dir, aproximadament a 1,3 m del sòl, com també de l’alçada sempre que és possible; després, s’han de fer els càlculs de biomassa utilitzant índexs diferents segons les espècies. Aquestes estimacions, ultra servir únicament per a la fusta, són força imprecises. Molts arbres tropicals, a més d’això, tenen contraforts a la base que es poden estendre fins a tres i quatre metres de l’eix i que fan difícils les mesures. El creixement individual d’un arbre, tant a curt com a llarg termini, pot ser molt irregular i la longevitat és prou gran perquè les variacions climàtiques l’hagin pogut influir.
El temps de renovació de les estructures renovables (fulles, fruits, etc.) és molt més breu que el de les estructures permanents (troncs, branques, arrels). La producció també serà diversa per als components diferents. Fins i tot en els boscos madurs la producció de massa llenyosa aèria suposa la part més gran de la producció neta. La producció de fulles en constituiria el 35%, mentre que la producció d’arrels, de la qual hom disposa de molt poques dades, podria representar només un 7%. En els arbres corpulents, una desena part de la fusta es descompon abans de caure i, de la resta, van a terra al cap de l’any més de 3 t de fusta morta de diàmetre superior a 10 cm.
Resulta més senzill d’avaluar el que hom anomena coberta morta, que és tot allò que acull el sòl de la selva. Per bé que només inclou fulles, flors, fruits i branquillons (exclou la fusta aèria i subterrània), resulta una bona estimació de la producció del component més dinàmic. Les variacions que s’observen d’un any a l’altre (al voltant del 10%) les podrien provocar, entre altres factors, les espècies de defoliació massiva.
Els condicionadors de la producció primària
Dins del bosc, la producció és condicionada per una colla de factors. La disponibilitat de llum, o el grau d’ombra, influeix el creixement; així, els arbres d’ombra creixen, en proporció mitjana, només un terç del que ho fan els que són exposats al sol: 0,7 mm anuals, els de diàmetre inferior a 5 cm situats en una ombra densa, i poc més d’1 mm anual per als arbres més grans també a l’ombra. En un bosc obert, en canvi, el creixement pot ser molt més gran. Algunes espècies de Flindersia, un gènere de rutàcies difós per les selves plujoses de les Moluques, Nova Guinea, Austràlia i Nova Caledònia, poden arribar a tenir un increment del diàmetre de 2,5 mm l’any, si creix en paratges oberts i assolellats, mentre que, a l’ombra, el creixement és gairebé nul. Per a la producció de fusta se seleccionen les espècies heliòfiles i de clarianes: a les Filipines, per exemple, hom recorre sovint a la mimosàcia Albizia falcataria, que ateny 20 m en dos anys i que cada vuit és aterrada. Hom selecciona així mateix, per a l’explotació, espècies pioneres de la successió secundària: la limba (Terminalia superba) a l’Àfrica, o el tec (Tectona grandis) a l’Extrem Orient.
L’estructura de la vegetació
L’estructura de la selva, amb arbres (en particular bombacàcies, dipterocarpàcies i lleguminoses) que emergeixen de la coberta principal, afavoreix l’heterogeneïtat. Les fulles superiors d’aquests arbres, exposades a la radiació directa, semblen mostrar poca sensibilitat estomàtica; la transpiració intensa que s’hi esdevé els converteix en metxes gegantines. Moltes de les seves fulles són esclerofil·les, amb cutícules gruixudes, i mostren una elevada reflectància. D’aquesta manera, la coberta, tot i constituir un filtre, permet l’entrada de força llum vers la vegetació inferior. Els arbres alts, exposats a una irradiació abundant, contribueixen més a la fotosíntesi que no els del sotabosc ombriu: a Cambodja representen més de la meitat de la producció del bosc, tot i disposar de només una cinquena part de la superfície foliar. Sembla que les espècies de capçada estreta, amb un quocient entre la superfície d’assimilació i el volum respiratori relativament alt, tenen una productivitat més elevada que no les espècies de capçada ampla, amb un volum respiratori més gran.
La disponibilitat d’aigua
Si la disponibilitat hídrica no és limitant (sense arribar als extrems de l’entollament), la densitat d’arbres emergents augmenta, com també la densitat de lianes i d’epífits; augmenta, en definitiva, l’índex de l’àrea foliar de la coberta i, per tant, l’ombra. La biomassa del bosc haurà augmentat mentre que la productivitat no haurà variat gaire. Dins dels vegetals, sobretot epífits, que no disposen d’aigua abundant, és corrent la utilització de l’anomenat metabolisme àcid de les crassulàcies (CAM, en sigles angleses) una modalitat de metabolisme encara més eficient que la C4 que constitueix un mecanisme d’adaptació a l’eixut: bromeliàcies, cactàcies i algunes falgueres, igual que les crassulàcies (els crespinells, per exemple), obren els estomes de nit per deixar entrar el diòxid de carboni (que guardaran en forma d’àcid màlic) i els tanquen de dia per evitar les pèrdues d’aigua; l’àcid màlic acumulat als vacúols permet continuar la fotosíntesi en aquestes circumstàncies.
La disponibilitat de llum
La capacitat màxima de fotosíntesi varia molt d’unes espècies a les altres i mostra una clara relació amb els ambients que ocupen: mentre que és molt elevada en les espècies pioneres (26 mg per decímetre quadrat i hora), és menor en conjunt per als arbres de la selva (6-24 mg per decímetre quadrat i hora) i força reduïda en les plantes dels estrats inferiors (2-4 mg per decímetre quadrat i hora), que amb poca llum aconsegueixen el seu màxim. Els estadis joves de totes les plantes de la selva poden funcionar amb poca llum: l’ambient ombriu i la reduïda biomassa comporten una taxa de respiració força atenuada que els permet un cert balanç positiu en la producció neta.
L’efecte de la temperatura
La temperatura és un altre factor que cal tenir en compte. L’absència d’un període fred evita qualsevol interrupció en el ritme anual de producció. Aquesta és potser la principal diferència dels boscos tropicals en relació amb els boscos temperats i, no cal dir-ho, en relació amb els boscos de coníferes de la taigà. Tant és així que, si no hi ha interrupcions estacionals ocasionades per una certa eixutesa, la fusta d’aquests arbres de la selva no sol presentar anells de creixement: les espècies perennifòlies, en concret, rarament fan anells observables i fins n’hi ha de caducifòlies que tampoc no en fan. D’altra banda, tot i que encara no ha estat del tot verificat el fet que la temperatura elevada comporti un increment de la respiració, la major part d’autors ho accepta com a realitat. Així, les nits fresques podrien reduir significativament les pèrdues per respiració.
Dins del bosc, el gradient tèrmic és molt marcat: en un migdia assolellat, a les capçades superiors hi pot haver una temperatura de 32°C i una humitat del 60%, mentre que al sotabosc, sense circulació d’aire i amb una ombra densa, la temperatura pot ser de 26°C i la humitat del 90%, l’atmosfera xafogosa tan divulgada de la selva.
La producció secundària
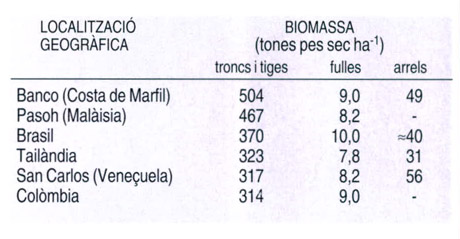
Valors de biomassa per a diferents estructures vegetals, en tones de pes sec per hectàrea, corresponents a diferents punts de selva plujosa tropical. La interpretació de les dades de biomassa de què hom disposa actualment és difícil, ja que sovint es barregen dades obtingudes amb mètodes diferents de mostratge. No obstant això, sí que hi ha algunes tendències generals, com ara valors superiors en tiges i troncs que en arrels, i valors superiors per a arrels que per a fulles. Paral·lelament, el contingut de nutrients als troncs i les tiges tendeix a ser superior al dels altres components dels vegetals de la selva.
Dades elaborades per Frank B. Golley
Si bé les dades de què hom disposa sobre la producció primària són escasses i incompletes, en el cas de la producció secundària encara n’hi ha menys. Per bé que la informació de la sistemàtica, de la distribució i fins de l’ecologia de molts animals és abundant, no existeix a hores d’ara cap estudi complet de les diverses espècies d’heteròtrofs d’un sol bosc tropical. Només hi ha estimacions de la producció, tant dels individus com de les poblacions, fetes a partir de la despesa metabòlica anual. Però dins d’un mateix nivell metabòlic el ritme de producció varia segons que es tracti d’organismes homeoterms, de despesa més elevada, o d’organismes poiquiloterms. Entre els consumidors, encara, l’assimilació del material ingerit és molt variable; així, mentre que alguns vertebrats assimilen entre el 80 i el 90% de l’aliment, en organismes detritívors (isòpodes, miriàpodes) l’assimilació sol ser només d’entre el 10 i el 30%.
Les dificultats no apareixen només a l’hora d’avaluar l’increment global de la biomassa animal sinó que ja comencen en l’intent de conèixer la mateixa biomassa (coneixement d’altra banda necessari, juntament amb el de la densitat, per a establir la producció de la fauna). Alguns autors, tot i això, donen estimacions, que reconeixen incompletes i preliminars, de la zoomassa. Així, per exemple, s’han donat valors de 210 kg per hectàrea per a una selva plujosa de plana a la “terra firme” del centre de l’Amazònia, i de 118 kg per hectàrea per a una selva plujosa de peu de muntanya a Puerto Rico. Aquests valors de biomassa animal total són inferiors a la biomassa dels vertebrats de les sabanes. De fet, però, els grans vertebrats de la sabana tindrien ben poques possibilitats d’alimentar-se a la selva, on els herbívors es nodreixen a 30 m enfilats dalt dels arbres i on la fusta i les fulles ofereixen més resistència a ser consumides que no l’herbam tofut de la sabana o la praderia.
Els insectes sobretot, però també els petits mamífers, són els herbívors principals del bosc. A l’illa de Barro Colorado (Panamà) s’ha calculat una zoomassa no voladora de 53 kg per hectàrea, de la qual el 60% correspon als peresosos (Bradypus) i el 12% a les mones udoladores (Alouatta), tots ells consumidors de fulles i ben adaptats a la vida arborícola. Hi ha ben poca informació de la biomassa foliar consumida pels herbívors. A Costa Rica i al Panamà s’han fet estimacions del 6 i el 10% respectivament, xifres que semblen ben minses si es té present que constitueixen l’entrada principal de matèria i energia vers els altres nivells tròfics. També cal considerar els mecanismes defensius, làtexs i metzines, que els vegetals fabriquen per evitar de ser consumits. Malgrat que amb una biomassa reduïda (infravalorada segons molts autors), aquest nivell alimentari exerceix un cert control dels processos productius. Els insectes fitòfags, per exemple, podrien actuar com a reguladors de la producció. Hi ha vegetals que han evolucionat conjuntament amb un seguit de fitòfags que, en determinar la mort de l’arbre, milloren la disponibilitat de llum i d’aigua dels supervivents i alhora forneixen una clariana adobada per a les llavors.
La major part de la biomassa d’heteròtrofs, però, és al sòl, i és quatre o cinc vegades més gran que la d’herbívors i carnívors plegats: 165 kg per hectàrea a la selva amazònica, on la biomassa d’herbívors és de 30 kg per hectàrea i la de carnívors de 15 kg per hectàrea, o de 80 kg per hectàrea en una selva baixa on la biomassa d’herbívors és de 25 kg per hectàrea i la de carnívors de 10 kg per hectàrea. D’entre la fauna del sòl destaquen els tèrmits, que poden ser molt importants en alguns boscos, els oligoquets i els microartròpodes; aquests darrers, tot i tenir una funció poc coneguda, són molt abundants al sòl i a la coberta morta on s’han fet recomptes de desenes de milers d’individus.
Les estratègies i els nínxols ecològics
La selva plujosa intertropical és la màxima expressió de la biodiversitat. El fet és reconegut, però les seves causes no són tan òbvies com es tendeix sovint a pensar: més energia disponible, producció de biomassa més elevada, clima estable durant un període prou llarg de temps... Amb tot això no n’hi ha prou per a explicar-ho. Ben al contrari, suscitaria la pregunta de per què una o unes poques espècies particularment ben adaptades a les condicions climàtiques de la selva plujosa no esdevenen dominants unes com a productors, unes altres com a consumidors primaris i unes altres com a consumidors secundaris i com a descomponedors, i s’estenen per àmplies àrees climàticament i edàficament prou homogènies en comptes de diversificar-se els éssers vius fins al punt que ho fan en aquest bioma de les selves plujoses.
La diversitat i la història
En aquest bioma i en tots els altres. Perquè, fet i fet, al món hi ha molt poques espècies àmpliament distribuïdes i representades per molts individus mentre que per contra són més nombroses les espècies d’àrea més restringida i representades per efectius menys nombrosos. L’evolució és un procés que s’autoaccelera i, en un context de clima estable amb estacionalitat poc acusada, les possibilitats d’especialitzacions que ajudin a defugir la competència de les altres espècies, encara que sigui a condició de circumscriure fins a l’extrem el propi nínxol ecològic, és molt elevada. D’això en resulta un gran nombre de nínxols, molts d’ells extremament limitats i especialitzats, alguns duts al límit de l’imaginable per l’extravagància o per la bellesa.
Hi ha autors que han volgut veure en aquest nombre tan elevat de nínxols i d’espècies el reflex d’una hipotètica estabilitat d’aquest sistema d’ençà del començament del Cenozoic, uns 65 milions d’anys enrere, durant els quals hauria romàs com a conjunt inalterat, no pertorbat per les variacions climàtiques d’altres latituds. Tanmateix avui sabem que no ha estat així. Les selves plujoses tropicals haurien tingut un màxim d’extensió al començament de l’Eocè, en què sembla que arribaren fins als 30° de latitud, si bé les latituds pròpiament equatorials, llavors de clima més eixut, eren ocupades per formacions més obertes. A partir de l’Eocè mitjà (fa uns 50 milions d’anys), coincidint amb una tendència planetària del clima a fer-se més calent però també més eixut, l’àrea de les selves plujoses tropicals es reduí. A partir del començament de l’Oligocè, fa uns 34 milions d’anys, nous canvis climàtics es traduïren en una disposició de les selves tropicals restringida a les latituds més equatorials, sense ultrapassar els 15° de latitud, una extensió més reduïda i tot que la present, que arriba fins als 20-25° de latitud. De llavors ençà, els canvis climàtics i les modificacions de l’àrea dels boscos plujosos s’han succeït.
En particular, als pics de les glaciacions, sembla que les selves plujoses quedaren reduïdes en més d’una ocasió a àrees molt limitades, els anomenats refugis plistocens, envoltades de boscos tropicals estacionals. La hipòtesi dels refugis plistocens es va proposar inicialment per a l’Amazònia, on s’observà que algunes àrees presentaven un elevat grau d’endemisme i una gran diversitat d’espècies de diferents grups animals (ocells, rèptils, papallones, algunes famílies de plantes, etc.), mentre que els territoris que les envoltaven presentaven un grau d’endemisme i una diversitat força més baixos. A l’Amèrica Central i meridional s’han identificat una bona colla d’aquests centres de diversitat que podrien correspondre a àrees de refugi de la selva plujosa durant els períodes més desfavorables del Plistocè, des dels de Yucatán fins als de la “mata” atlàntica brasilera i des dels del Chocó, a la costa pacífica de Colòmbia, fins als de les Guaianes, passant pels de l’orient equatorià, el Napo peruà o el sud-oest de Pará, al Brasil. A l’Àfrica s’haurien reconegut tres d’aquests centres, un a l’alta Guinea, l’altre als confins entre el Camerun i el Gabon i el tercer als marges orientals de la conca del Zaire. Així doncs, si bé la història explica les variacions que ha sofert la riquesa d’espècies a les selves plujoses, l’argument de l’estabilitat només seria aplicable a aquests comptats refugis plistocens i només per un període molt més breu que els suposats 65 milions d’anys d’ençà del començament del Cenozoic.
Tampoc l’explica completament, encara que certament hi té un pes indubtable, el fet que siguin les regions equatorials les que rebin més radiació solar i les que disposin de pluges més abundoses. En definitiva, les que, en igualtat de condicions pel que fa a nutrients del sòl, poden ser més productives i poden desenvolupar-se més en alçada. D’altra banda, així com la producció depèn de la superfície, tant pel que fa a la disponibilitat d’energia com d’aigua de pluja, l’acumulació de biomassa (i també de necromassa) no queda limitada de la mateixa manera sinó que es pot desplegar també en la tercera dimensió de l’espai. Així ho fan els arbres, en efecte, per als quals la competència per la llum és una pressió selectiva decisiva, punt central de l’organització dels ecosistemes forestals, dels quals les selves plujoses són l’exemple més acabat.
Els grans arbres dominants de les selves plujoses optimitzen al màxim la formació d’estructures verticals que permetin a les seves fulles captar la llum necessària i alhora que en privin a les dels arbres veïns, i això fins al límit que ho permetin les disponibilitats d’aigua i nutrients, la capacitat de suport dels materials i de l’estructura dels troncs (la fusta, en el cas dels boscos) i la capacitat de transport dels vasos que pugen aigua i nutrients des de les arrels fins a les fulles.
El predomini de les plantes llenyoses
Així, els arbres, en la seva lluita per assolir la llum que els convé i amb la seva capacitat d’organitzar les tres dimensions de l’espai de l’ecosistema fins a alçades que atenyen sovint els 35 m a l’alçada del cobricel i els 50 m a les capçades dels emergents, són els primers factors determinants d’una certa heterogeneïtat d’aquest espai i, en conseqüència, d’una diferenciació primària d’ambients, de nínxols ecològics, font d’una diversitat creixent.
La varietat de les formes arbòries
La diversitat mateixa dels arbres de les selves plujoses resulta ja de bon començament aclaparadora. Mentre que als boscos temperats, sovint dominats per una sola espècie o per ben poques, la xifra de 10 espècies diferents d’arbres en una hectàrea ja és molt elevada, als boscos tropicals se sobrepassen amb facilitat les 100 espècies de més de 10 cm de diàmetre per hectàrea. Això significa també una àmplia gamma de materials químicament diversos, variació en els substrats per als epífits i una estratificació que, en constituir microambients, estimula l’especiació en els organismes que els habiten. Les condicions de llum, temperatura i humitat poden variar espectacularment de les capçades al sòl, a les diferents alçades dels troncs i de les capçades, fins i tot en cada enforcadura d’una branca o en cada forat de cada tronc. D’altra banda, si la pressió selectiva primària és per a les plantes llenyoses la competència per la llum, un cop les grans espècies arbòries han tancat un cobricel amb les seves capçades, són unes altres les pressions selectives dominants per al conjunt de les plantes vasculars (adaptació a l’aprofitament de la llum escassa del sotabosc, a l’accés al capdamunt del cobricel, a la vida epifítica, al parasitisme).
L’abundància d’epífits i lianes
Un element especialment característic de les selves és, així, el dels epífits, les plantes que viuen damunt dels arbres, els quals els ofereixen un suport en la volta il·luminada del bosc, sense causar-los directament cap perjudici. Fora dels tròpics només es troben microepífits (líquens i molses sobretot, i algunes falgueres). Els macroepífits es troben en gran abundància a les selves plujoses, ja que se’n compten unes 28 000 espècies repartides en 65 famílies. La de les orquidàcies, la més rica, inclou més espècies epífites que no pas terrestres, i a les selves americanes, hi són nombroses les bromeliàcies.
Entre els epífits es pot fer una distinció per estrats: els de la coberta, molt ben adaptats per a absorbir i guardar l’aigua, presenten característiques semblants a les plantes d’indrets eixuts (fulles dures i lluents, o bé extremament primes) obligats com són a resistir la irradiació intensa, el vent i les sequeres ocasionals; els del sotabosc, en canvi, que han de suportar molta humitat i subsistir amb poca llum, tenen fulles delicades, fan gutació per eliminar aigua, sovint les fulles els acaben en punta per assecar-se aviat i mostren la superfície arrugada o vellutada per augmentar l’àrea d’intercepció de llum. Les branques antigues, fortes i robustes, de més fàcil colonització, suporten una gran massa d’epífits. Les escorces clivellades són avantatjoses per a les arrels rampants i els circells.
Per poder subsistir en unes condicions no habituals en els vegetals, els epífits han desenvolupat diversos mecanismes. Un pot ser tenir tiges o fulles adaptades per recollir aigua i així poder esmorteir els canvis de disponibilitat d’aigua i de nutrients. Un altre pot ser presentar dos tipus d’arrels, l’un per aferrar-se a l’hoste i l’altre que penetra l’humus o bé s’estén lliurement en l’aire per capturar, talment una esponja, la humitat de l’aire. La simbiosi amb altres organismes també és molt freqüent: les micorizes d’un fong els poden fornir nutrients, especialment en els primers estadis, o les formigues hi poden transportar engrunes de sòl i restes de virosta per tal d’establir-hi el niu. Això sense comptar tot un estol d’altra fauna associada (altres insectes i fins petites granotes) que troba, a més d’aixopluc, la humitat i l’aliment que necessita en aquesta mena de bosquetó suspès. Moltes espècies animals hi poden completar tot el seu cicle vital sense haver-se’n de moure.
Les lianes, especialment abundants als tròpics, contribueixen a l’aspecte dens i misteriós d’aquest bosc. Com els epífits i els hemiepífits, defugen la foscor dels estrats baixos. És per això que inverteixen en estructures de creixement ràpid i en mecanismes que els permeten agafar-se als arbres alts que, tot i ser els que dominen el bosc, els serveixen de suport per a enfilar-s’hi tortuoses. A les selves baixes, al voltant d’un 8% de les espècies segueixen aquesta estratègia i la proporció encara és més gran al bosc secundari.
El capteniment de la fauna
Amb la multiplicitat d’hàbitats i de recursos que ofereix el món vegetal, no és pas sorprenent que també la diversitat de la fauna sigui elevada i ocupi nínxols d’especialització inimaginable en altres biomes, a la qual contribueixen també les condicions de llum i humitat pròpies del bioma. La riquesa comença per la fauna edàfica, composta per una gran diversitat d’àcars i col·lèmbols que viuen sobre fongs lignícoles. Els verms, per contra, hi són poc representats, mentre que formigues i tèrmits, tal com ja s’ha dit, sorprenen per l’abundància (arriben a constituir tres quarts de la zoomassa edàfica) i són l’aliment d’óssos formiguers i pangolins, com també de llangardaixos arborícoles. Els tèrmits són essencials en la descomposició de la fusta mentre que les formigues, especialment les armades, són carnívores d’una ferocitat temible.
Els avantatges de la humitat ambiental elevada
L’elevada humitat ambiental permet que els animals de pell nua i prima no corrin perill d’assecament i els obre les portes d’hàbitats insospitats en altres latituds: invertebrats propis de les aigües dolces (planàries, sangoneres) poden viure entre les branques uns quants metres per damunt de la superfície del sòl, als estrats baixos del bosc; verms propis de la fauna edàfica troben dalt de les capçades, a la virosta retinguda pels epífits, un hàbitat perfectament apropiat.
Pel que fa als vertebrats s’hi fa, per exemple, una gran varietat de granotes, algunes de les quals fan un cicle completament “terrestre” en realitzar tot el procés de metamorfosi dins l’ou. Els fitotelmes, cavitats destinades a recollir aigua de fulles, branques o inflorescències de moltes plantes epífites, com la majoria de les bromeliàcies, constitueixen una font d’aigua i de nutrients per a larves de mosquits i per a diminutes granotes, com també un refugi en condicions de sequera per a molts animals de petites dimensions.
L’èxit de la petitesa
La reducció de la mida és un altre dels recursos dels animals per a poder explotar encara més nínxols: esquerdes insignificants, petites concavitats, pilositats de fulles o d’animals, cavitats corporals d’animals, plomes, etc. Tot serveix com a hàbitat, com a aliment, o de totes dues maneres, per a servar una forma o altra de vida. Com també ajuda a la supervivència de molts animals de les selves el recurs de la discreció i l’ambigüitat: hàbits nocturns, formes i coloracions críptiques i mimètiques. O a l’inrevés: l’exhibició de coloracions advertidores de la pròpia perillositat o mimètiques d’altres organismes efectivament perillosos.
El triomf de l’arboricolisme
En un medi forestal tan complex i intricat no és estrany que la proporció de fauna arborícola sigui elevada en tots els grups animals, dels més petits als més grans. Ja s’ha esmentat la presència, molts metres per sobre del sòl, de fauna edàfica o pròpia de les aigües superficials, però molts insectes, fins i tot alguns habitualment tan terrícoles com formigues i tèrmits, també fan nius a molts metres d’alçada, dalt dels arbres. En un estudi fet a la Guyana, mentre que es trobaren 31 espècies de mamífers arborícoles, només se’n trobaren 23 que visquessin a peu pla.
Tots els animals arborícoles presenten adaptacions anatòmiques (mans, urpes i cues prènsils, ungles poderoses, expansions cutànies per planar, etc.) que els ajuden a moure’s a través de les branques dels arbres amb gran eficàcia. Els ocells, per exemple, són molt abundants als estrats superiors, mentre que ben poques espècies viuen o ponen els ous a terra. L’avifauna tropical, tot i ser una limitada fracció de la biomassa, mostra una gran diversitat en relació amb la de les regions temperades. En molts casos han estat les relacions recíproques entre dues o més espècies les que han conduït a una especiació que té molt de coadaptació: insectes, ocells i rats-penats pol·linitzadors, tota casta d’animals disseminadors de fruits i llavors, pugons criats com a bestiar per formigues, etc.
Les diferents ofertes alimentàries
I això que només una petita part de la biomassa vegetal és veritablement accessible als herbívors, i encara aquesta només per als veritablement especialitzats. La composició química de les fulles les fa sovint inadequades, si no tòxiques, excepte per a molt comptats consumidors especialistes, la majoria insectes. Els animals més grans, incapaços de mastegar-les prou finament per a trencar les parets de cel·lulosa i alliberar-ne els nutrients, necessiten l’ajut de bacteris que viuen al seu sistema digestiu i amb els quals estableixen una simbiosi.
Pel que fa als depredadors, tot i que són més variats i més nombrosos que en altres boscos, a les selves tenen una diversitat menor que la dels herbívors, si més no en nombre d’espècies. L’àmplia riquesa d’insectes afavoreix el predomini dels insectívors, la majoria dels quals són així mateix invertebrats. Les característiques de l’aliment determinen les dimensions reduïdes de la majoria de depredadors, amb excepcions tan notables com els grans fèlids, que són caçadors de rosegadors grans, remugants, tapirs i pècaris. Entre els carnívors del bosc tropical, destaquen també nombrosos ocells rapinyaires que s’alimenten de mamífers i d’ocells arborícoles petits. Les temperatures altes i constants dels boscos tropicals fan possible, d’altra banda, en contraposició amb la miniaturització, força més generalitzada a la majoria dels grups animals, l’existència de formes gegantines d’insectes i d’altres invertebrats que han intercanviat els papers amb els vertebrats, i són capaces d’alimentar-se d’ocells petits, sargantanes, rosegadors menuts i fins de petites serps.
El mite de l’estabilitat
A les selves plujoses és on el quocient entre producció i biomassa (P / B), és a dir, la taxa de renovació, assoleix els valors més baixos. Ja s’ha explicat que això implica un control més gran dels cicles dels elements químics, fins al punt que es pot dir que es tracta d’un ecosistema tancat, en el qual tot es recicla. Potser per això hom havia associat de manera massa simplista la complexitat i la diversitat del bioma de les selves plujoses amb una suposada gran estabilitat. La realitat és que un sistema amb un espectre més ampli d’espècies i una xarxa més complexa d’interaccions té una vulnerabilitat més elevada que els sistemes menys diversos i més simples enfront d’aquelles pertorbacions que són d’una freqüència insuficient per haver estat interioritzades pel sistema o bé d’una intensitat desacostumada. La selva plujosa tropical resulta en realitat, doncs, més fràgil que no pas els sistemes forestals temperats, els quals, malgrat la seva complexitat menor, tenen una capacitat més gran de regeneració enfront de catàstrofes incidentals com un incendi o una tala.
En circumstàncies normals, la feble oscil·lació que s’esdevé d’una manera natural en els paràmetres ambientals dels ambients tropicals humits permet que la selva plujosa, estable només dintre d’uns límits relativament restringits, pugui prosperar i persistir. Efectivament, les pertorbacions hi són localitzades i de poca amplitud. Els llamps, el vent, la modificació del curs d’alguns rius, per exemple, poden determinar l’aparició de clarianes allà on els arbres han estat abatuts, però aquests claps pertorbats no ocupen normalment més enllà de l’1 o el 2% de la superfície de la selva. Malgrat aquesta relativa exigüitat, tals processos de destrucció localitzada seguida d’un procés de restauració gradual del bosc original, eviten que les selves plujoses en el seu conjunt romanguin massa temps sense renovar-se. De fet, encara que la superfície efectivament pertorbada sigui escassa en cada moment, l’extensió dels claps en reconstrucció és força superior, del 5 al 20% de la superfície total, atesa la lentitud del procés de successió consecutiu a una pertorbació.
En definitiva, doncs, els cataclismes naturals, que es produeixen amb una certa regularitat, menen el bosc a una diversificació d’estadis que coexisteixen i el complexifiquen: estadis inicials, amb poca diversitat i molt mudables, s’ofereixen voltats d’estadis madurs, més persistents i diversos, i encara d’estadis de degradació manifestament empobrits. En el cas dels grans rius de plana, per exemple, que descriuen meandres i van modificant gradualment el seu curs, la disposició topogràfica de la vegetació reflecteix la cronologia de la successió secundària: a mesura que els aiguadeixos fan enretirar el curs del riu, la nova ribera és colonitzada en faixes paral·leles a la riba, de manera que les etapes més inicials de la successió queden a tocar d’ella i les més avançades en queden més allunyades. Recíprocament, allí on el riu erosiona la riba, sovint talla perpendicularment les faixes successives que representen etapes més o menys avançades de successions secundàries precedents. Però una estabilitat dinàmica així exigeix unes condicions ambientals quasi constants (requisit que el clima del bioma de les selves plujoses sol oferir) i alhora que les pertorbacions no ultrapassin un cert llindar; altrament, els canvis que sobrevenen són grans i perdurables.