Cadascú al seu lloc, però més insectes que altra cosa
L’inventari de les espècies animals vivents, sobretot la de les que viuen a les selves equatorials i tropicals, és ben lluny de ser complet. Les dramàtiques destruccions sofertes pel patrimoni forestal del nostre planeta han portat a l’extinció d’un nombre considerable de formes animals —i vegetals—, abans que la seva existència fos documentada i reconeguda, però això no obstant sabem que les selves plujoses equatorials representen, absolutament, el més ric i prestigiós dipòsit de biodiversitat animal de tot el planeta.
És difícil valorar exactament el nombre i, en certs aspectes, la naturalesa de les espècies animals encara per descriure. Només hem de pensar que les selves equatorials africanes no ens han revelat fins al segle XX l’existència d’animals tan grans i vistosos com l’ocapi (Okapia johnstoni), descrit per primera vegada l’any 1901, l’hiloquer o senglar gegant (Hylochoerus meinertzhageni), que ho fou el 1904, o el paó del Zaire (Afropavo congensis), incorporat als annals de la ciència el 1936.
L’abundància marejadora dels insectes
En termes numèrics, a la incomparable varietat de formes animals de les selves plujoses contribueixen sobretot els insectes, amb un nombre d’espècies difícilment determinable, però estimat avui dia entre un mínim de 2 milions (si més no, tres vegades el nombre total de les espècies actualment conegudes, que s’acosten a les 800 000), fins a un màxim de 80 milions, que correspondria a quasi 100 vegades el nombre de les espècies fins ara inventariades. Els insectes, d’altra banda, representen gairebé a tot arreu la fracció més important de la fauna dels ecosistemes terrestres, cosa deguda a factors diversos, entre els quals les seves dimensions i la seva especialització alimentària.
Els insectes, en efecte, tenen habitualment dimensions que oscil·len entre pocs mil·límetres i pocs centímetres. Això els permet constituir poblacions locals no gaire petites, capaces de sobreviure fins i tot en àrees reduïdes, amb una disponibilitat de recursos alimentaris que seria massa modesta per mantenir en vida un poblament d’animals més grans. La seva especialització alimentària, d’altra banda, és sovint extrema, tant pel que fa a les espècies vegetarianes com pel que fa a les que s’alimenten d’altres animals. En aquelles, l’especialització es refereix tant a la selecció de les plantes alimentàries com a la manera d’alimentar-se’n. A les selves tropicals, entorn d’una única espècie arbòria, es pot desenvolupar l’existència sencera d’un miler d’espècies diferents d’insectes. Per començar, aquelles que s’alimenten de fulles xuclant-ne els sucs, rosegant-ne el limbe a partir del marge o des d’una de les cares (preferentment des de la inferior), minant-hi una galeria en el gruix subtil de la mateixa fulla o, encara, induint la fulla mitjançant secrecions fetes a propòsit, a transformar-se en una gal·la. A continuació, aquelles que ataquen les arrels, o la tija, o bé les flors o els fruits. I, finalment encara, aquelles que esperen que la fusta d’una planta moribunda o morta sigui atacada pels fongs.
Certament, una part considerable d’aquests insectes pot alimentar-se a costa de plantes d’espècies diferents (generalment, però, pertanyents al mateix gènere o, almenys, a la mateixa família); el percentatge d’insectes vegetarians monòfags, és a dir, vinculats per a la seva alimentació a una única espècie vegetal, s’estima, per a les selves plujoses, entre l’1 i el 5%. En tot cas, a més de la incertesa d’aquestes estimacions, que es basen en coneixements encara molt fragmentaris, s’ha de recordar l’enorme diversitat de la flora d’aquests ambients, on només les espècies arbòries, que són les que hostatgen una fauna associada més rica, es compten ja per milers (40 000 espècies pel cap baix). És precisament de consideracions d’aquest tipus que en resulten algunes de les més raonables estimacions globals sobre biodiversitat proposades aquests últims anys.
El poblament harmònic i el poblament disharmònic
Davant de l’enorme diversitat d’espècies animals que pobla les selves plujoses, s’imposa, però, introduir un factor diferencial. D’una banda tenim el poblament de les selves africanes i, sobretot, de les de l’Àsia sud-oriental, en les quals la diversitat del poblament zoològic no és donada sols pel grandíssim nombre d’espècies presents, sinó també per la diversitat de grups que hi contribueixen. D’altra banda, ben al contrari, tenim el poblament de les selves sud-americanes i, encara més, de les de Madagascar, de Nova Guinea i d’Austràlia, en les quals el nombre d’espècies és elevat, però hi contribueix un nombre relativament modest de tàxons de nivell superior al de l’espècie. Els biogeògrafs expressen aquesta diferència definint com a harmònic un poblament del tipus present en les selves africanes i asiàtiques i disharmònic un poblament del tipus present en les altres àrees.
Aquesta diferència té, naturalment, raons històriques. El poblament d’Austràlia, de Nova Guinea i de Madagascar es ressent de la condició insular, més o menys antiga, d’aquestes àrees, on la colonització no ha estat possible per part de grups que, en canvi, sí que han contribuït al poblament de les selves asiàtiques i africanes. També l’Amèrica del Sud, tot i haver conegut moltes vegades, ni que fos que de manera interminent, enllaços amb l’Amèrica del Nord que han permès intercanvis faunístics importants entre les dues àrees, ha vist desenvolupar les seves faunes, sobretot la de la selva plujosa, en condicions d’aïllament.
Tanmateix, val la pena recordar que la presència, a l’Amèrica del Sud, d’un animal com el jaguar (Panthera onca) s’ha de relacionar amb una onada de poblament relativament recent. A l’inrevés, el fet que Madagascar (juntament amb alguna de les illes Comores) sigui avui dia la pàtria exclusiva dels lèmurs no és pas el resultat d’una història evolutiva acomplerta exclusivament en aquella illa, sinó el fruit de la progressiva extinció dels lemúrids en altres parts del Vell Món, de manera que els últims representants només sobreviuen avui a Madagascar i a Maore (Mayotte).
Els uniformes i les disfresses
En un ecosistema extraordinàriament ric d’espècies, com és el de les selves plujoses, el risc de ser víctima d’un depredador és, òbviament, molt alt. Això explica la riquesa i la complexitat de les adaptacions defensives, o preventives, que han desenvolupat els més diversos grups zoològics. La situació s’acaba de complicar pel fet que moltes adaptacions importants per a la defensa d’una possible presa són al mateix temps adaptacions ofensives que faciliten, al mateix animal, l’actuació com a predador en relació amb altres preses.
Només cal fixar-se, per exemple, en el cas dels camaleons, un grup de saures arborícoles particularment difós entre les selves plujoses de l’Àfrica equatorial i, sobretot, de Madagascar (dos terços de les espècies del món). Els camaleons es mouen molt poc i amb una increïble lentitud, comportament sorprenent tractant-se de saures. No és gens fàcil observar-los entre les branques i el fullam on passen el temps, sobretot perquè la seva coloració es confon en gran manera amb la del fons. És ben coneguda la capacitat dels camaleons per a canviar de color d’acord amb el de l’ambient que els envolta. En aquest notable comportament hi ha implicats els ulls, que indiquen a l’animal cada variació cromàtica del medi, i els cromatòfors, és a dir les cèl·lules pigmentades de la pell, l’estat de contracció o de distensió dels quals és el responsable dels canvis cromàtics de l’animal. En qualsevol cas, aquest se n’aprofita tant en el seu paper de predador com en el de presa. De fet, d’una banda, és important per al camaleó no fer-se notar pels insectes dels quals, immòbil, està a l’aguait. Tan bon punt els té a una distància convenient, els captura amb un rapidíssim moviment de la llarga llengua viscosa, que projecta endavant en menys d’un vintè de segon sobre la seva víctima i retira no menys ràpidament. D’altra banda, la capacitat del camaleó de romandre immòbil i ajustar la seva coloració a la del medi redueix el risc de ser víctima d’algun dels nombrosos depredadors (mamífers, ocells, serps) a mercè dels quals es pot trobar.
Camuflatges d’aquesta mena estan molt difosos per la selva plujosa, encara que en general no es dóna la capacitat, pròpia dels camaleons, de canviar de color segons les necessitats. Així i tot, resulta difícil dubtar de l’eficàcia de les coloracions protectores de nombrosíssims insectes, la forma de les ales dels quals (o, fins i tot, la forma del cos) contribueix a fer més complet el camuflatge. Són exemples força notables i dels més coneguts els insectes bastó i els insectes fulla de l’ordre dels fasmòpters. Rígids i angulosos com petites branquetes sense fulles, els primers inclouen entre altres, amb la seva trentena de centímetres, el més llarg dels insectes (Pharnacia serratipes); dels segons són remarcables les espècies del gènere Phyllium, les curtes ales de les quals simulen a la perfecció la nervadura de les fulles. També s’observen casos semblants en molts ortòpters, sobretot de l’Àsia sud-oriental, o també en algun lepidòpter, com les papallones índies del gènere Kallima, la cara superior de les ales de les quals traeix el parentiu amb les vistoses vanesses, mentre que la cara inferior, amb les ales tancades, fa l’insecte virtualment idèntic a una fulla morta, inclòs el pecíol.
Un resultat anàleg l’aconsegueixen també molts mamífers, mitjançant el desenvolupament de ratlles o de sèries de taques, que difuminen els contorns de les seves siluetes, tal com poden aparèixer a distància, entre els clarobscurs de la selva. Aquest, per exemple, sembla que és el significat de les vistoses ratlles verticals, alternativament clares i fosques, que trobem, no únicament a la pell del tigre (Panthera tigris) sinó també en alguns antílops africans com el bongo (Tragelaphus euryceros) o el duiker zebra (Cephalophus zebra), que precisament viuen en la part més frondosa de la selva, o les dels quarts posteriors de l’ocapi (Okapia johnstoni), un dels mamífers més grans de la selva plujosa africana.
Els colors d’advertiment
Tanmateix, no sempre els colors o els dibuixos s’utilitzen per a fer menys visible l’animal als ulls d’un possible depredador o d’una possible presa. A vegades, l’estratègia adoptada és exactament la contrària. En altres paraules, per a molts animals esdevé molt important ser vistos, i no sols per membres de la pròpia espècie.A les selves equatorials, en efecte, hi ha un gran nombre d’espècies amb coloracions aposemàtiques o d’advertiment, com les que en les selves americanes identifiquen, fins i tot a distància, la presència de les serps corall (Micrurus), de mossegada molt verinosa, o la d’una petita granota arborícola del gènere Dendrobates o d’algun altre de la mateixa família, amb una pell molt rica de secrecions tòxiques que pot resultar un material preciós, per a algunes poblacions humanes locals, per a fabricar fletxes enverinades. Aquests animals estan proveïts de temibles defenses naturals, però també per a ells, i no únicament per als seus adversaris, és sens dubte preferible no haver-les de fer servir. És raonable, doncs, interpretar la vistosa coloració roja i negra de molts dendrobàtids, o el característic dibuix d’anells rojos, blancs i negres de les serps corall, com a mitjans per a avisar de lluny el potencial agressor que no és aconsellable jugar-se-la amb un animal capaç d’inflingir una mossegada enverinada o de causar vòmits (o greus intoxicacions) a qui se’n vulgui alimentar, abans d’haver d’arribar a un enfrontament directe.
Tanmateix, sovint la presència de coloracions vistoses comporta utilitzacions molt més subtils. Aquesta circumstància fou descoberta, precisament en el cor de la selva plujosa amazònica, per un dels seus primers grans exploradors, el naturalista anglès Henry W. Bates (1825-92), a propòsit d’alguns gèneres molt comuns de papallones diürnes d’ales negres, vellutades, amb vistoses taques roges i blanques que havia descobert durant els seus viatges per l’Amèrica del Sud del 1848 al 1859. A la selva amazònica, entre les papallones amb aquesta coloració més difoses i abundants hi ha les del gènere Heliconius, les erugues de les quals es desenvolupen nodrint-se de fulles de Passiflora. Són papallones del tot immenjables per les seves carns tòxiques. Tanmateix, Bates descobrí que el mateix patró de coloració aposemàtica es manifesta en altres papallones, com ara les de les famílies dels pièrids i els papiliònids, sense cap parentiu especial amb els helicònids i, a diferència dels Heliconius, perfectament comestibles. La semblança entre les espècies inofensives i Heliconius no és de cap manera casual, com ho mostra el fet que cada espècie “copia” precisament la coloració i el dibuix de l’espècie d’Heliconius amb la qual conviu. Als ulls d’un depredador es confonen, doncs, el missatge aposemàtic “correcte” d’una papallona immenjable i l’“enganyós” d’una papallona comestible, que redueix el risc de ser efectivament menjada mercès a la seva extraordinària semblança amb l’espècie tòxica. Per tal que el mimetisme batesià funcioni, l’espècie copiada ha de ser força més abundosa que la inofensiva ja que, si no, els depredadors detectarien l’engany.
Qui es menja qui
Totes les xarxes tròfiques són més o menys complexes. Sempre hi ha animals carnívors i herbívors, éssers especialitzats a devorar només una sola mena de preses i d’altres capaços d’ingerir pràcticament qualsevol cosa. A la selva plujosa també és així, però potser encara més que en altres biomes. Sense moure’ns del món dels insectes, és fàcil trobar especialitzacions alimentàries concretes que es troben a la base de l’enorme riquesa d’espècies present a les selves plujoses.
Els insectes paràsits i predadors
Sense insistir en els vegetarians, dels quals ja s’ha parlat extensament amb relació als seus aspectes coadaptatius amb les plantes, no es poden deixar de banda els depredadors i els paràsits.
D’aquests últims, la fracció més important és representada pels himenòpters, sovint de minúscules dimensions, el grau d’especialització dels quals és generalment encara més gran que el que vincula moltes de les seves víctimes a les respectives plantes d’on obtenen l’aliment. De fet, el cicle alimentari d’aquestes minúscules vespes ha de sincronitzar-se oportunament amb el de les seves víctimes, la localització de les quals, per altra banda, resulta tan dificultosa com pugui resultar-ho la recerca de fulles o de flors d’una determinada planta. En particular, els més petits d’aquests himenòpters (fins a mig miler d’espècies diferents entre les frondoses branques de la capçada d’un únic arbre, a les selves de Borneo!) es desenvolupen dins dels ous d’altres insectes: un únic ou de papallona, o de xinxa, pot contenir prou aliment per a una d’aquestes vespes, que acompleixen dins d’ell tot el seu desenvolupament larval i en surten, ja adultes, tot just per a acoblar-se i per anar a cercar, les femelles, altres ous de la mateixa xinxa o papallona, en els quals dipositar-hi els seus propis ous.
Aquesta mena d’adaptacions i la complexa xarxa de relacions tròfiques que en resulta, encara que no són exclusives del bioma de les selves plujoses, hi eviten el creixement explosiu de les poblacions. Molt més característiques són, tanmateix, altres relacions, que expliquen per elles mateixes la diversificació d’algunes famílies d’insectes en milers i milers d’espècies.
Una d’aquestes relacions és la que vincula les plantes del gènere Ficus i els insectes agaònids. El gènere Ficus té aproximadament unes 800 espècies, quasi totes pròpies de les selves plujoses, sobretot de l’Àsia sud-oriental. N’hi ha que són arbres gegantins i d’altres que són petites mates, moltes són lianes, fins i tot epífites, però totes produeixen les característiques inflorescències en siconi formades per un gran nombre de floretes obertes cap a l’interior d’una cavitat que comunica amb l’exterior mitjançant una obertura apical molt estreta. Per a garantir una fecundació creuada, cal que el pol·len produït per les flors masculines dins d’una d’aquestes inflorescències arribi a les flors femenines d’una altra. Es tracta d’una empresa molt difícil que només pot ser realitzada exclusivament per pol·linitzadors força especialitzats, com són els agaònids, minúscules vespes (un mil·límetre de llarg o poc més) que acompleixen el seu cicle vital sencer a les figues. Per a dipositar els ous, les femelles d’aquests insectes han de sortir de la figa on s’han desenvolupat i penetrar en una altra, de la qual asseguren així la pol·linització. Cada espècie de ficus té la seva vespa pol·linitzadora o fins i tot més d’una. L’estretíssima relació entre tots dos grups d’organismes explica així, alhora, la riquesa d’espècies de Ficus i la d’agaònids.
Cicle biològic d’una vespa pol·linitzadora de Ficus
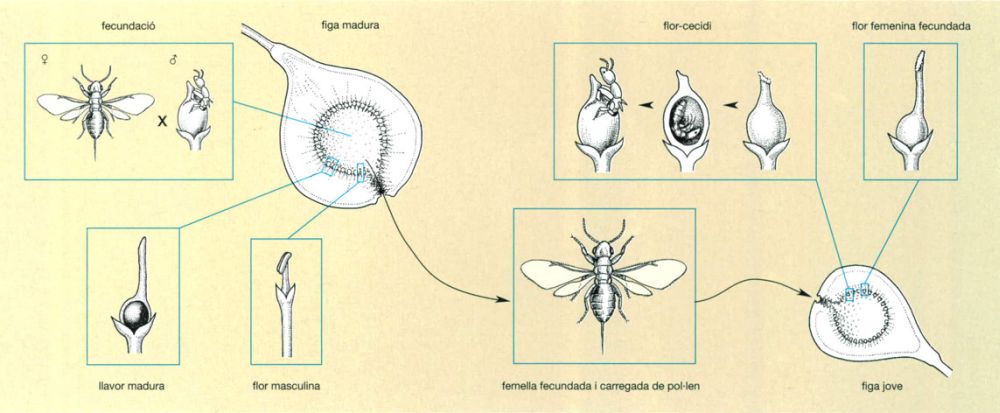
Jordi Corbera, a partir de fonts diverses
Les serps constrictores i les verinoses
A les selves plujoses, les serps són dels vertebrats que assoleixen un paper més singular com a depredadors. Al si d’aquest bioma, i no tan sols entre les formes més fines i lleugeres, han evolucionat formes d’ofidis de singular agilitat per a moure’s entre les branques. Entre les serps de la selva també s’han generalitzat les coloracions críptiques, que en aquest cas tenen essencialment valor ofensiu: són una útil precaució per a no ser vistes abans d’hora per les víctimes escollides. Entre les més significatives cal esmentar el pitó reticulat (Python reticulatus), el pitó verd arborícola (Chondropython viridis), la boa maragdina (Corallus caninus), les serps liana (Ahaetulla prasina, Dryophis) i la serp fuet verda (Oxybelis fulgidus).
Les tres primeres espècies pertanyen a la primitiva família dels bòids. Són serps massisses, que maten la presa mecànicament, estrenyent-la entre els seus anells. El pitó reticulat (Python reticulatus), força difós a l’Àsia sud-oriental i a l’arxipèlag indonesi, és la serp més gran després de l’anaconda comuna (Eunectes murinus), un altre bòid, en aquest cas americà; s’alimenta de preses proporcionades al seu cos, que engoleix senceres, encara que pesin 50 kg!
Notablement més petits són els altres dos bòids esmentats. La boa maragdina (Corallus caninus) viu a les selves de l’Amèrica del Sud, mentre que la pàtria dels pitons verds arborícoles (Chondropython viridis) són les selves d’Austràlia septentrional i de Nova Guinea. La lliurea adulta d’aquestes dues serps és sorprenentment semblant, d’un verd brillant amb dibuixos blancs sobre el dors, però les cries l’assoleixen mitjançant una fase juvenil en què la respectiva coloració és completament diferent: vermella en el cas de la boa maragdina, groga (més rarament d’un roig rajola) en el del pitó verd arborícola.
La serp liana (Ahaetulla prasina) està dotada de dents verinoses. És una serp de fins a 2 m de llarg, amb una àrea de difusió semblant, a grans trets, a la del pitó reticulat. És una espècie força àgil i veloç, amb un cap de forma poc freqüent en una serp, que s’estreny, cap endavant, en forma cònica, fins al punt de permetre als ulls una visió binocular (i per tant estereoscòpica), circumstància força oportuna per a un depredador de sargantanes i de granotes arborícoles. També és verinosa la mossegada de la serp fuet verda americana (Oxybelis fulgidus), en la qual trobem, possiblement, la més completa adaptació a la vida arborícola que hagi estat desenvolupada per una serp. El seu cos, extraordinàriament afinat, pot assolir els 2 m de llargada. La coloració és d’un verd herba brillant al dors, amb un fi marge groc clar que la separa del groc verdós del ventre: una òptima coloració críptica per a un animal que viu al bell mig del fullam dels arbres. És un animal actiu tant de dia com de nit, implacable predador de sargantanes, granotes i nius d’ocells.Encara que són més freqüents en altres biomes, alguns crotàlids viuen també a les selves plujoses del Nou Món i hi representen una de les més temudes presències animals. La més gran entre aquestes serps és la surucucú (Lachesis muta) que, amb els seus 3 m o més de llarg, només és superada, entre les serps verinoses, per la cobra reial (Ophiophagus hannah) de l’Àsia meridional, que pot assolir una llargada de 4 m o més. El verí de la surucucú, d’altra banda, és força menys perillós que el d’una altra serp de les selves equatorials americanes, la barba groga o punta de llança (Bothrops atrox) que, tot i ser la meitat de llarga que la primera, és la responsable de la majoria de morts per mossegada d’ofidi de tot el continent americà. La surucucú és ovípara, mentre que la barba groga és vivípara, tal com és habitual entre els crotàlids.
Els mamífers i els ocells d’alimentació especialitzada
També es poden observar notables especialitzacions en l’alimentació dels mamífers de la selva plujosa. Aquest és el cas, per exemple, dels peresosos (Bradypus, els peresosos de tres dits, amb tres espècies, i Choloepus, els peresosos de dos dits, amb dues), típics habitants de les selves humides americanes. En particular, els peresosos de tres dits (Bradypus) tenen una clara predilecció per les fulles de les cecròpies (Cecropia). Aquesta especialització alimentària es troba probablement en relació amb el nivell d’activitat singularment baix d’aquests animals, de ben segur els més sedentaris de tots els mamífers, que només comptades vegades fan desplaçaments diaris superiors a una trentena de metres. Els peresosos passen la major part del seu temps penjats de les branques, gràcies a les seves ungles fortíssimes. Sobre els seus llargs pèls es desenvolupa habitualment una pàtina d’algues, semblant a la que es fa, al si de l’ambient tan humit de la selva, sobre l’escorça dels arbres o sobre qualsevol altre substrat inert. El pèl sedós del peresós esdevé així, més que res, tot un ecosistema en miniatura, del qual participa fins i tot una eruga que s’alimenta de les descamacions cutànies del mateix peresós.
Una altra especialització alimentària és la dels mamífers i els ocells que s’alimenten de formigues i de tèrmits. Aquests darrers, per exemple, representen l’aliment quasi exclusiu de l’ós formiguer (Myrmecophaga tridactyla) i el predominant de les altres dues espècies de la mateixa família dels mirmecofàgids: el tamàndua (Tamandua tetradactyla) i l’ós formiguer nan (Cyclopes didactylus), anomenat a Colòmbia “serafín de platanal” pel seu aspecte angelical de boleta de pelfa. El paper dels óssos formiguers a les selves sud-americanes l’assumeixen, al Vell Món, els pangolins (Manis), menjadors de formigues amb el dors recobert d’escates, alguns dels quals, encara que no tots, són típics habitants de les selves plujoses.
Ja s’ha esmentat la peculiar activitat depredadora, a les selves africanes i americanes, de les temibles fileres de formigues viatgeres, com Eciton burchelli i Labidus praedator. L’onada de petits organismes com ara aranyes, coleòpters i altres invertebrats i petits vertebrats terrícoles que fugen desesperadament de les formigues sense parar esment en cap altre possible predador ofereix una bona font d’aliment a alguns d’aquests predadors especialitzats precisament a seguir els desplaçaments de les colònies de formigues viatgeres. Aquesta és l’especialització d’algunes espècies d’ocells de la família dels formicàrids, en particular l’ocell formiguer de corona negra (Pittasoma michleri) i diverses espècies de griva formiguera (Formicarius).
Les arrels alimentàries dels primats
La relació primària dels primats, tant prosimis com simis, amb l’ambient de la selva plujosa és ben establerta. Tots els simis sud-americans estan vinculats al bioma de la selva plujosa; només al Vell Món (tant a l’Àfrica com a l’Àsia), algunes espècies de primats han colonitzat secundàriament ambients més oberts o zones forestals més temperades. Semblant a la situació sud-americana seria, en canvi, la de Madagascar, on les selves plujoses, desgraciadament tan malmenades, allotgen la major part de les espècies de prosimis vivents.
Un grup zoològic que pot ajudar-nos a reconstruir algunes etapes importants de les vissicituds evolutives que des d’un típic mamífer terrestre, com podria ser una musaranya, han portat a l’organització dels primats pròpiament dits, amb les seves sofisticades adaptacions a la vida arborícola, és el de les tupaies o musaranyes arborícoles (Tupaia, Dendrogale, Urogale). I la veritat és que, entre els nombrosos grups de mamífers que viuen avui a les selves equatorials, les tupaies representen certament un dels menys vistosos. La quinzena d’espècies conegudes tenen unes dimensions corporals d’entre 10 i 20 cm de llarg, més de 10 a 25 cm de cua, i el seu aspecte no s’allunya gaire del d’una musaranya. Juntament amb talps, eriçons i musaranyes, en efecte, les tupaies havien estat sovint enquadrades, en el passat, en l’ordre dels insectívors. Però en les classificacions modernes se’ls reconeix una col·locació diferent, formant un ordre independent, el dels escandents, i sembla que se’ls accepta un parentiu particularment estret amb els primats.
Les tupaies són molt àgils enfilant-se per les branques, però les seves extremitats no són prènsils. Tot i una certa mobilitat recíproca dels dits, per enfilar-se s’han de valer sobretot de les ungles llargues i esmolades. Tampoc la cua, llarga i força mòbil, no s’ha transformat en òrgan prènsil, i durant els desplaçaments pot ser, com a molt, utilitzada com a contrapès variable per equilibrar la resta del cos. Aquestes limitacions mecàniques no són gaire greus ateses les modestes dimensions dels animals. Les tupaies són animals diürns, molt actius, amb un ull que sembla que és capaç de distingir els colors, capacitat no gaire freqüent entre els mamífers. Llurs hàbits alimentaris, que corresponen als dels animals dels quals versemblantment deriven, són sobretot insectívors. Alimentar-se d’insectes és, en tot cas, una de les poques bones solucions per a un animal de sang calenta i de dimensions molt petites, amb un metabolisme força elevat que requereix una contínua aportació d’aliments molt energètics. Els insectes, juntament amb altres menjars rics de calories, com el nèctar o els líquids ensucrats produïts pels insectes que xuclen la saba de les plantes i en concentren els sucres, constitueixen també una fracció rellevant de l’alimentació dels primats més petits, com els lèmurs ratolins (Microcebus) de Madagascar, d’unes dimensions semblants pràcticament a les d’una tupaia. En canvi, un simi de grans dimensions pot acontentar-se fins i tot amb un aliment menys nutritiu però més fàcil d’obtenir, com són les fulles i els fruits dels arbres.
Tant entre els lemúrids de Madagascar com entre els simis platirrins sud-americans, s’observa una notable radiació d’adaptació que afecta, en particular, les especialitzacions alimentàries. Sens dubte el comportament més extrem és el de l’ei-ei (Daubentonia madagascariensis). L’aspecte d’aquest lèmur és tan insòlit que fou pres inicialment per un rosegador, en comptes d’un primat. Aquesta atribució errònia fou suggerida probablement per la dentició, en particular per les quatre incisives esmolades; a més, li manquen els ullals i en total només té 18 dents. Però encara més singular que la dentició és l’estructura de les mans i els peus. Tots els dits són bastant llargs i prims i acaben en urpes recorbades, a excepció del polze del peu, que posseeix, per contra, una ungla plana. Particularment prim és el tercer dit de la mà, que l’ei-ei utilitza com una eina especialitzada. Els components de la seva alimentació són dos: fruita i larves d’insectes que viuen sota les escorces. Per alimentar-se de fruita, l’animal trenca la pell amb les dents i després hi introdueix el tercer dit de la mà, amb el qual rasca la polpa tova, que es porta després a la boca.També fa servir el mateix dit per arribar als estrets amagatalls que hi ha entre escorça i fusta, refugi de larves xilòfagues de què també s’alimenta.
Un univers d’adaptacions admirables
A la selva plujosa, quasi tota la vida animal es desenvolupa al cobricel i a les capçades dels estrats més elevats del sotabosc, entre els alts troncs columnars i l’intrincat brancam de les capçades. La manca de vida a peu pla i en el subsòl té, naturalment, les seves causes i les seves no menys paleses i importants conseqüències. En terra, manca aquell espès estrat de fulles mortes i d’altres detrits que, generalment, en els ambients forestals temperats s’acumula als peus dels arbres i que lentament és transformat per una legió de microorganismes, de fongs i de petits invertebrats, bo i permetent el desenvolupament d’una comunitat rica i variada. A la selva plujosa, una part del detrit orgànic és interceptat, immobilitzat i transformat abans d’arribar a terra i roman enredat entre les cavitats dels troncs o a les axil·les de les grans branques, entre els coixinets de molsa o entre les fulles i les arrels de les innombrables plantes epífites. El que arriba a terra es descompon molt ràpidament.
En qualsevol cas, a dalt, entre les branques frondoses, hi ha força més per menjar del que hi pugui haver a terra, i un ambient més complex i més estructurat. Un ambient on construir-se un niu, on cercar amagatall o, al contrari, aguaitar una presa. Per això, la major part dels animals de la selva presenten adaptacions específiques per a una vida entre les branques. Aquestes adaptacions són particularment singulars i notables en el cas dels quadrúpedes, que semblen, en principi, més aptes per a una vida a peu pla.
Moure’s entre les branques
Les adaptacions més vistoses són les relacionades amb la locomoció. Cues prènsils, dits oposables i ungles molt fortes i llargues són les solucions més esteses i eficaces. Cadascuna ha aparegut més d’una vegada al llarg de la història evolutiva en llinatges ben diferents, fins i tot s’ha repetit en diferents grups d’una sola classe com és el cas dels mamífers. Les estratègies adoptades pels animals de les selves plujoses del Vell Món amb relació a la locomoció presenten un ventall de solucions força més ampli que el que es pot trobar a l’Amèrica del Sud. Certament, en un i altre espai abunden els grimpadors, però algunes solucions, com la cua prènsil, no tenen la mateixa importància al Vell que al Nou Món.
La cua com a extremitat
Les cues prènsils, per exemple, semblen una especialitat dels mamífers de les selves equatorials americanes, però les trobem en grups molt diversos: en les àgils mones aranya (Ateles), en un carnívor nocturn de mida mitjana com el “kinkajú” o ós meler (Potos flavus), en alguns marsupials com els opòssums o sarigues (Didelphis), o fins i tot en un rosegador com el coendú o porc espí arborícola de cua prènsil (Coendou prehensilis). Aquest darrer, un animal de talla relativament gran (de 30 a 60 cm de llargada del cos fins a la base de la cua i fins a 5 kg de pes), presenta també unes ungles fortes i robustes, com les que es troben en alguns simis sud-americans com els tamarins (Saguinus) i, sobretot, en els dos óssos formiguers arborícoles (Tamandua tetradactyla i Cyclopes didactylus) i en els peresosos (Bradypus i Choloepus).
La braquiació dels simis
Una altra important adaptació per a la vida entre les branques és el fenomen anomenat de braquiació, és a dir, la capacitat de penjar-se de les branques mitjançant les extremitats i de desplaçar-se d’aquesta manera de capçada a capçada. La braquiació, pròpia dels simis, a més d’un bon desenvolupament de cames i braços, exigeix una mà de dits oposables, prènsil en definitiva. Una mà de polze oposable als altres quatre dits —quasi com la humana—, però també un peu de dits oposables: molts mamífers arborícoles són en realitat quadrumans (aquesta característica és general en quasi tots els primats, amb la sola excepció del bipedisme humà). Una modificació semblant es dóna també a les quatre extremitats dels camaleons, que són de bon tros les més ben adaptades a la vida arborícola de les de tots els saures. Entre els camaleons, però, la disposició dels dits és diferent de la dels mamífers quadrumans: a les extremitats anteriors d’aquests rèptils hi ha oposats els tres primers dits contra els altres dos, mentre que a les extremitats posteriors els dos primers dits s’oposen als tres restants.
Entre els primats, les espècies arborícoles més difoses de l’Àsia sud-oriental es mouen a força de braços, tant els gibons (Hylobates), que assoleixen una obertura de braços d’1,5 m i un pes de 13 kg, com els orangutans (Pongo pygmaeus), el mascle dels quals pot assolir la considerable alçada de 150 cm i un pes de 90 kg pel cap baix. Les nou espècies conegudes de gibons habiten a les selves de l’Àsia sud-oriental i, sota molts punts de vista, s’han de considerar els més àgils d’entre tots els mamífers. Es mouen entre els arbres utilitzant les extremitats superiors, i es llancen sovint al buit, des d’una branca a una altra fins a distàncies d’uns 3 m; amb el salt, al qual recorren menys sovint, poden assolir els 9 m. És ben difícil veure per terra un gibó o un orangutan, a diferència del que s’observa entre els simis antropomorfs africans, és a dir, entre les dues espècies de ximpanzés —el ximpanzé comú (Pan troglodytes) i el ximpanzé nan o bonobo (P. paniscus)— i el goril·la (Gorilla gorilla), del qual es distingeixen diferents subspècies, només algunes de les quals viuen la selva plujosa.
Comparació entre els peus i les mans dels simis antropomorfs i els humans
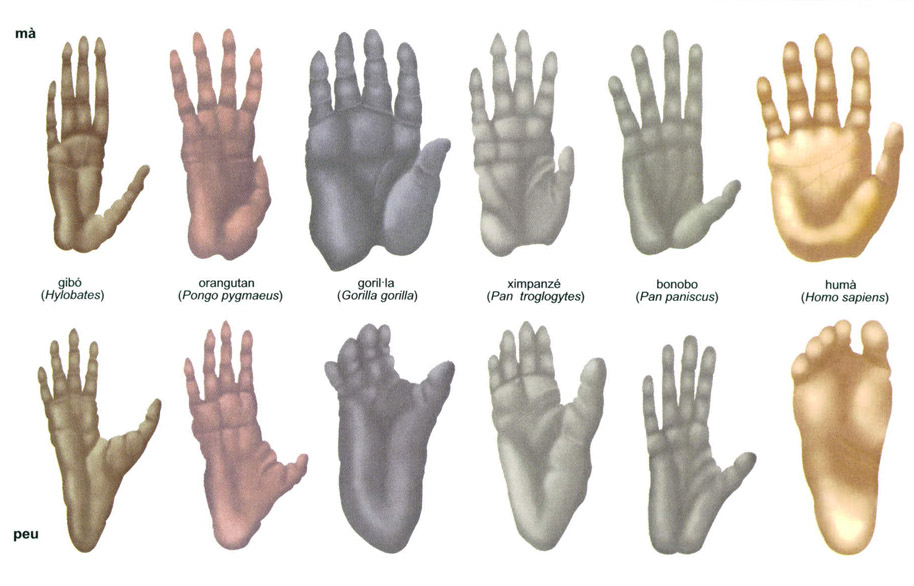
Editrònica, a partir de fonts diverses
Els vols planats
És singular el nombre d’animals de les selves plujoses asiàtiques que han desenvolupat dispositius per a poder realitzar vols planats. Aquest comportament afecta, dins de l’àrea en qüestió, dos grups diferents de mamífers i dos gèneres de saures, a més d’un gènere de granotes i encara, per sorprenent que resulti, algunes espècies de serps. És una estratègia comuna de tots aquests animals l’eixamplament o l’aplanament del cos a fi de crear una mena de paracaiguda, capaç de sostenir l’animal en l’aire, mentre es llança des d’una branca més alta cap a una de més baixa o cap a terra. Les modalitats amb què s’assoleix aquest objectiu varien segons els casos.
Els mamífers capaços de planar
Una de les transformacions anatòmiques que fan possible el vol planat entre els mamífers consisteix en el desenvolupament d’un patagi lateral, és a dir, d’un plec cutani que s’estén, a banda i banda del cos, entre els membres anteriors i els posteriors. Aquest és el cas, per exemple, dels esquirols voladors. A les selves de l’Àsia sud-oriental, aquest grup d’esquirols és representat pels gèneres Aeromys, Petaurista, Hylopetes, Petinomys, Pteromysus, Petaurillus i Iomys. Tanmateix, els esquirols voladors no són pas exclusius de les selves plujoses tropicals: n’hi ha gèneres diferents que són presents a les selves temperades de l’hemisferi septentrional, tant a Euràsia com a l’Amèrica del Nord.
Per contra, és exclusiu de les selves de l’Àsia sud-oriental el minúscul ordre dels dermòpters, que comprèn només dues espècies, conegudes amb el nom de lèmurs voladors: Cynocephalus volans, que viu a la part meridional de les illes Filipines, i C. variegatus, present a la part meridional de Tailàndia i d’Indo-xina, a la península de Malaca, a Sumatra, Java, Borneo i a diverses illes menors. L’estratègia planadora dels lèmurs voladors es basa en la possessió d’unes membranes cutànies que uneixen el cap amb els membres anteriors, aquests amb els posteriors, i els posteriors amb la cua. Són animals de posició sistemàtica incerta, probablement propers als insectívors, encara que la seva alimentació és fonamentalment vegetariana, a base de brots tendres, flors i fulles. Nocturns, de 34 a 42 cm de llargada, més una cua de 17 a 27 cm, i amb un pes d’entre 1 i 1,7 kg, els lèmurs voladors passen les hores del dia dins de cavitats d’algun arbre, o penjats del fullatge d’una palmera ben alta. De nit fan llargs vols planats amb els quals poden superar distàncies de 100 m o més, sense perdre pràcticament alçada.
El vol planat ha evolucionat també en un tercer grup de mamífers, en aquest cas marsupials, amb una estratègia força paral·lela a la dels esquirols voladors. Es tracta dels opòssums voladors (Petaurus, Petauroides, Acrobates), amb una àrea de difusió que s’estén pel nord d’Austràlia, Nova Guinea i les veïnes illes Bismarck.
Els rèptils i els batracis que volen
Entre els rèptils, es troben formes capaces de planar en gèneres de dues famílies de saures (agàmids i gecònids) i d’una família de serps (colúbrids).
A la família dels agàmids pertanyen els dragons voladors de l’Àsia sud-oriental (Draco), sens dubte els animals que posseeixen el patagi més sofisticat. Quan s’estan immòbils sobre una branca, aquests petits rèptils (22 cm de llargada, dels quals 14 cm corresponen a la cua) tenen si fa no fa l’aspecte d’una sargantana, i encara poc perceptible a causa de la seva coloració críptica. Però el seu aspecte es transforma completament quan l’animal, estirant les costelles fins a posar-les en posició perpendicular a l’eix del cos, estén el seu patagi vistosament acolorit i es llença a l’aire tot d’una, prenent l’aspecte d’una gran papallona de colors brillants. L’espècie més coneguda, Draco volans, té el patagi de color blau amb taques negres; en altres espècies del mateix gènere la coloració pot ser groguenca o fins i tot roja. La vistosa coloració de l’animal volant desapareix així que aterra, tan ràpidament com s’havia manifestat. Naturalment, tot això, a més de la dificultat de seguir un animal capaç de llançar-se al buit d’improvís, contribueix a desorientar qualsevol possible depredador. Els vols planats dels dragons voladors tenen generalment una amplitud de 10 o 20 m, el que necessita per arribar d’un arbre a un altre, però poden assolir-ne fins a 100 m, naturalment amb una notable pèrdua d’alçada. Tanmateix, el petit rèptil no té cap dificultat per guanyar una posició més alta, grimpant amunt veloçment per les branques. El dragó volador no utilitza el patagi només per a fugir, sinó també com a defensa enfront dels enemics i fins i tot en les parades nupcials: mascle i femella despleguen alhora les respectives membranes vistosament acolorides i s’observen recíprocament; després la femella, si està disposada a l’acoblament, roman ferma sobre la branca, amb les ales replegades, i el mascle la sobrevola i se li enfila al dors, sempre amb el patagi desplegat, i finalment procedeix a la inseminació. La femella pon generalment quatre ous cada vegada, que s’obren després al cap d’un o dos mesos. El dragó volador és insectívor i s’alimenta quasi exclusivament de formigues, que captura amb la llengua.
És més simple el patagi dels gecònids planadors del gènere Ptychozoon, de la Malàisia peninsular, Sumatra, Java, Borneo i illes dels estrets de Malaca. En aquests rèptils planadors existeixen reduïdes expansions cutànies al costat del coll, al llarg dels flancs i de la cua, com també al llarg de les extremitats, que acaben en cinc dits palmats. Els vols planats d’aquests gecònids són molt més modestos que els dels dragons voladors; arriben com a màxim a uns quants metres.
En el cas de les serps voladores (Chrysopelea), l’aplanament del cos necessari per a sostenir-se a l’aire s’obté per un aixecament de les costelles, per efecte del qual la part anterior del cos s’aplana i el ventre esdevé lleugerament còncau. Les serps voladores tenen una mossegada verinosa, però el seu verí no és gaire potent. Un cop capturada la presa, que pot ser un ocell petit, una sargantana, un ratolí o un rat-penat, la serp la manté ferma entre les mandíbules i l’estreny entre els seus anells, fins a matar-la.
Una membrana estesa entre els llargs dits (una solució força evident per a un amfibi) és finalment el que permet “volar” a les petites granotes voladores (Rhacophorus), les més famoses de les quals són probablement la granota voladora de Reinwardt (R. reinwardti), que viu a Java i a Sumatra i que fa uns 8 cm de llargada, i la granota voladora de Wallace (R. nigropalmatus), que viu a Borneo.
La vida nocturna
Una part força important de la vida animal de la selva té com a escenari les hores de la nit. Un primer senyal del desvetllament de moltes criatures, en correspondència a la posta, l’assenyala un canvi sobtat dels sons de la selva.
Les granotes raucadores i els insectes lluminosos
La major part de les granotes, per començar, fa sentir els seus raucs sobretot a les hores de la nit. Es tracta generalment de concerts amb totes les de la llei, no únicament per la sonoritat dels intèrprets, sinó també per la seva diversitat: una zona ben reduïda de la selva pot allotjar fins a una cinquatena d’espècies diferents de granotes raucadores. Cada espècie, naturalment, té el seu precís repertori de vocalitzacions, cadascun d’ells d’una importància vital per a l’espècie que l’emet, ja que és el mitjà per al recíproc reconeixement i per a la formació de les parelles. Una altra adaptació clàssica a la vida nocturna, certament presentada també per aquestes granotes, és el desenvolupament d’ulls de grans dimensions: entre les coses que poden ser importants de valorar per a una granota arborícola, hi ha la distància del blanc cap al qual vol saltar. Es poden trobar adaptacions encara més òbvies en la forma dels dits, generalment eixamplats a la punta i dotats d’un coixinet que secreta un moc adequat per afavorir la fixació de la granota arborícola a la superfície de les fulles.
Una altra presència molt sonora, a l’espessor de la selva, al costat de la de les granotes arborícoles, és la de cigales i grills. Però certament moltíssims altres animals estan actius, durant la nit, encara que la seva presència no causi altres rumors que febles brogits o ocasionals sorolls provocats per una caiguda. El silenci afavoreix tant els depredadors que van de caça, com les seves possibles preses. Si els mascles de les granotes, de les cigales i dels grills fan sentir els seus cants, és només perquè no tenen cap altre mitjà eficaç per trobar parella. Objectiu, aquest, que altres animals de la nit realitzen mitjançant raigs de llum, tal com fan les lluernes.
Entre els insectes productors de llum de les selves equatorials, els més vistosos són els “cucujos”, “cucubanos” o “bichitos de luz” americans (Pyrophorus), coleòpters de la família dels elatèrids que s’han fet un lloc dins del folklore local antillà. Hom diu que els negres de les Antilles capturaven aquests coleòpters atraient-los amb carbons incandescents fixats a l’extrem d’un bastó i després se’n servien per il·luminar les seves cabanes: també, embolicats en tul i fixats als cabells o als vestits de les dones, han estat utilitzats com a joies vivents en festes i balls tradicionals.
Els depredadors de la nit
La nit, tanmateix, és sobretot l’escenari de les incursions silencioses dels depredadors, des dels grans felins com el tigre o el jaguar, fins a les serps, els dragons o les escolopendres. La presència i l’activitat nocturna d’aquests animals són fàcilment perceptibles a l’entorn de qualsevol implantació humana, allà on un llum encès esdevé el centre d’atracció d’una multitud d’arnes, borinots i altres insectes, la presència dels quals atreu al seu torn la dels seus petits depredadors, com dragons i escolopendres, eventualment seguits per altres visitants més amenaçadors. D’aquests, els més temibles són en general els felins. No tots els representants d’aquesta família estan lligats al bioma de les selves plujoses, ni tots tenen hàbits exclusivament nocturns, però no hi ha dubte que entre els felins trobem òptims exemples d’adaptació a una activitat depredadora nocturna en un ambient selvàtic.
Per a aquests animals són molt importants les prestacions de llurs òrgans sensorials i sobretot dels ulls. Aquests estan col·locats en tal posició que els permeten una visió binocular, condició indispensable per a calcular correctament la distància a què es troba la presa a encalçar. També és molt important la capacitat d’adaptar l’ull a condicions de diversa intensitat lluminosa. Si aquesta és molt baixa, la nineta pot dilatar-se molt i així pot utilitzar de la millor manera possible la poca radiació disponible. Un estrat de cèl·lules reflectores, col·locades darrere de la retina, reflecteix els raigs que arriben a aquesta, redoblant-ne a la pràctica la intensitat. També són força útils els llargs bigotis que els permeten una delicada sensibilitat tàctil, suggerint al felí quins són els espais pels quals pot avançar, fins i tot en condicions de gran obscuritat. Altres adaptacions, com les ungles retràctils, són massa conegudes per haver-hi d’insistir.
Els felins, en definitiva, tenen una important presència en quasi totes les selves asiàtiques, africanes o americanes. A l’Àsia, tret del tigre (Panthera tigris), del lleopard (P. pardus), l’àrea del qual s’estén fins a l’Àfrica, i del lleopard nebulós (Neofelis nebulosa), són representats per espècies de dimensions modestes, com el gat de jungla (Felis chaus), el gat marbrat (F. marmorata) i el gat daurat asiàtic (F. temmincki). A les selves africanes trobem també, a més del lleopard, el gat daurat africà (F. aurata), mentre que els majors felins americans són, a part el jaguar (Panthera onca), l’ocelot (F. pardalis), el margai (F. wiedii) i el jaguarundí (F. yagoaroundi).
Els termoscopis de les serps
Moltíssimes serps estan actives de nit. No obstant això, entre aquests rèptils no s’aprecien mai ulls grans i sortints com en moltes granotes, o en algun mamífer nocturn com els lemúrids. De fet, les serps disposen d’altres solucions per a poder localitzar amb precisió les preses durant les seves caceres nocturnes.
En primer lloc, moltes espècies poden dilatar sensiblement, en condicions d’escassa lluminositat, la seva nineta, de manera semblant a com ho fan els felins. Les serps, però, recorren més aviat a altres sentits més que no pas a la vista. Per exemple, a l’olfacte, gràcies a un òrgan particular (òrgan de Jacobson) format per dues cavitats sensorials presents al paladar, al sensible epiteli de les quals arriben les partícules oloroses recollides a l’aire o al medi mitjançant l’incessant moviment de la llarga llengua bífida.
Però algunes serps disposen, a més, d’uns òrgans sensorials veritablement únics al món animal: es tracta de termoscopis, és a dir, d’òrgans que identifiquen les radiacions tèrmiques emeses per un cos calent. Mitjançant aquests òrgans la serp pot localitzar una víctima (un mamífer o bé un ocell) que tingui una temperatura més alta que la del medi que l’envolta. Hi ha termoscopis més aviat rudimentaris que es troben presents en diferents espècies de pitons, sota la forma d’unes plaques labials peculiars, però la forma més especialitzada d’aquests òrgans és pròpia de la família dels crotàlids, a la qual pertanyen entre d’altres les serps cascavell. Un termoscopi és una petita fossa sensorial encaixada en la corresponent depressió de l’os maxil·lar i subdividida en dues cambres, una d’externa i una d’interna, mitjançant una membrana molt fina i amb moltes terminacions nervioses, prop d’un miler per mil·límetre quadrat. Aquestes terminacions nervioses estan connectades al nervi trigemin i són sensibles a les diferències de temperatura, fins i tot molt petites (0,003°C!), entre la cambra externa i la interna.
Els primats noctàmbuls
Entre els primats han evolucionat costums nocturns en diferents grups. Un primer exemple el representarien els lèmurs, si més no els de petites dimensions, com els lèmurs ratolí (Microcebus, Phaner), els lèmurs nans (Mirza, Cheirogaleus, Allocebus) o els lèmurs mostela (Lepilemur). En els lèmurs, tanmateix, sembla haver-se verificat (cas força singular) un retorn als costums diürns, o almenys crepusculars, entre les espècies de dimensions més grans (Lemur, Eulemur, Hapalemur, Varecia, Petterus), versemblantment menys subjectes als riscos de depredació que les espècies més petites.
També són nocturnes les quatre espècies conegudes de tarsis (Tarsius), característics habitants de les selves de les illes Filipines, de les Cèlebes, de Borneo i de Sumatra, uns petits primats arborícoles que fan tot just de 8,5 a 17 cm de llarg, més 18 a 28 cm de cua. Tenen les extremitats molt llargues, sobretot les posteriors, i dits llargs i fins, amb les puntes dilatades; els ulls són molt grans, com sovint succeeix entre els mamífers actius durant les hores nocturnes, o entre els rapinyaires nocturns, amb els quals tenen en comú la capacitat de girar el cap quasi 180° i el sigil dels desplaçaments. Es mouen sobretot amb àgils salts de prop d’un metre, i poden cobrir, en una sola nit, distàncies de fins a prop d’un quilòmetre. Igualment són de costums nocturns els gàlags (Galago, Euoticus, Otolemur, Galagoides), uns altres prosimis, aquests africans, emparentats amb els lèmurs, la majoria dels quals viuen en típics ambients de selva equatorial. Tenen, igual que els tarsis, ulls grans i botits.
En canvi, entre els simis pròpiament dits, els hàbits nocturns són una excepció, representada fonamentalment pels duruculís (Aotus), uns petits simis americans de grans ulls.
Les adequacions de l’avifauna
Per als animals típicament terrestres com els rèptils o els mamífers, l’adaptació a la vida de la selva es fa notar sobretot amb relació a les peculiars solucions locomotrius que els permeten una vida entre les branques dels arbres. Naturalment, són ben diferents les pressions de selecció que han modelat els diferents grups d’ocells que s’han adaptat a viure a la selva plujosa. En qualsevol cas, la presència dels ocells a les selves plujoses és massiva i extremament diversificada. Una modesta parcel·la de selva amazònica, per exemple, pot allotjar una avifauna tan rica d’espècies (400 o 500) com la de tot Europa.
L’evidència dels becs
Entre les adaptacions més conspícues a la vida a la selva en el cas dels ocells, hi ha les referents al règim alimentari i, com a reflex d’aquest règim, la forma del bec. Per a moltes espècies frugívores, sobretot pel que fa als grans gèneres de coloms i tórtores de l’Àsia sud-oriental i del Pacífic tropical (Ducula, Treron, Ptilinopus), sembla que tot es limiti a un problema de dimensions, que permetin evitar una coincidència massa estreta amb altres espècies del mateix grup: cada parcel·la de selva pot allotjar-ne fins a tres o més espècies, cadascuna de mida diferent i amb uns becs semblants, però de llargada proporcionada a la grandària de l’ocell.
En altres casos, la mateixa naturalesa dels aliments preferents ha imposat una dràstica reestructuració del bec, com és el cas dels papagais, als quals els és possible l’accés a la polpa sucosa d’una llavor encara que estigui protegida per una closca molt dura, o dels colibrís, ben adaptats a succionar els nèctars de les flors tubuloses. Menys fàcil d’explicar és l’origen del bec, enorme però molt lleuger, dels tucans i dels calaus, en qualsevol cas escandalosament vistents.
Dos casos singulars: l’hoatzín i el casuar
A les mateixes selves amazòniques que representen el cor de l’àrea geogràfica, totalment americana, dels colibrís, hi ha un altre ocell prou insòlit, l’hoatzín (Opisthocomus hoazin), la posició sistemàtica del qual s’ha estat debatent constantment des del moment de la seva descripció, feta el 1776. A causa d’una superficial semblança amb els cràcids, una família sud-americana de l’ordre dels gal·liformes (al qual pertanyen galls, faisans, paons i perdius), l’hoatzín havia estat atribuït sovint, sense gaire convicció, a aquest ordre, però investigacions basades en mètodes moleculars d’hibridació del DNA suggereixen més aviat una afinitat amb els cucúlids, la família dels cucuts. El tret més singular de l’hoatzín és que, de jove, presenta, al primer i el segon dit de l’ala, unes ungles molt desenvolupades i dotades d’una musculatura especial que en permet el moviment. Aquestes ungles fan que el jove hoatzín pugui —cas únic entre els ocells— penjar-se a les branques dels arbres. Al cap de tres semanes d’haver sortit de l’ou, aquestes ungles cauen i l’ala pren un aspecte més normal. L’hoatzín, que assoleix una llargada d’uns 60 cm, freqüenta els indrets de la selva d’aigües abundoses, al llarg dels rius, i de bona gana s’hi cabussa i hi neda amb una certa desimboltura. La presència d’ungles a les ales d’aquest ocell sovint ha estat considerada una característica de gran primitivisme, que recordaria una condició semblant a la de l’ocell més antic conegut, l’Archaeopteryx del Mesozoic, però aquesta comparació només es pot fer en sentit analògic, sense cap implicació filogenètica homològica directa. Però és que l’hoatzín també és remarcable per la seva fisiologia digestiva: és un dels rars ocells que s’alimenta exclusivament de fulles, raó per la qual disposa d’un esòfag i un estómac eixamplats en els quals una població bacteriana simbiòtica fermenta la matèria vegetal ingerida, tal com fan els remugants, permetent-li així l’assimilació dels aliments. Tant és així, que l’hoatzín fa olor de fems de vaca i, a causa de la llarga durada de les seves digestions (dos dies), la seva capacitat de vol queda molt limitada mentre paeix.
Decididament, però, encara és més primitiva la posició dels casuaris (Casuarius), grans ocells sense ales, vagament semblants a l’estruç, que tenen una àrea de difusió que abasta Nova Guinea, l’Austràlia septentrional i algunes illes i arxipèlags veïns. De les tres espècies conegudes, almenys una, C. unappendiculatus, de Nova Guinea, pot ser considerada com un habitant de les selves plujoses. El casuari és un fort ocell corredor, capaç de córrer a gran velocitat fins i tot per entre la intricada vegetació de la selva. Durant molt de temps s’ha pensat que precisament en relació amb això hagués desenvolupat el vistós elm ossi que li corona el cap. Segons una interpretació corrent, de fet, aquesta robusta cresta òssia li protegiria el cap dels violents xocs contra les plantes amb què el casuari ha de topar inevitablement durant els seus veloços desplaçaments pel frondós sotabosc. Tanmateix, unes observacions recents indueixen a pensar que la funció principal d’aquest “casc” és en realitat la d’una mena de pala, amb la qual el casuari remou la sorra i els detrits de terra, en cerca de fruits caiguts.
Les estructures de nidificació
Finalment, cal fer referència als comportaments nidificadors de molts ocells de les selves plujoses. Moltíssimes espècies, naturalment, utilitzen les cavitats naturals que troben als troncs o bé n’excaven de noves. Aquest comportament, però, no afecta únicament els grups que solen recórrer a aquest tipus de nidificació, com ara els picots o els raspinells neotropicals de la família dels dendrocolàptids, sinó també els representants d’algunes famílies que típicament construeixen nius de tota una altra mena. És el cas, per exemple, dels anomenats ocells forners sud-americans (família dels furnàrids), que generalment fan uns nius de fang massís que, assecats al sol, assoleixen una consistència molt notable, mentre que les espècies que viuen a la selva nien dins de cavitats dels troncs.
Més singulars encara resulten els nius dels oriols americans (Psarocolius) de la família dels ictèrids. Aquests ocells nien en colònies sobre grans arbres, sovint isolats, a les branques dels quals pengen en grups els seus grans nius, d’una llargada que pot arribar a assolir el metre i mig. Són nius en forma de sac, suspesos d’un peduncle força prim, que per la seva factura resulten pràcticament inaccessibles a la depredació per part de les serps i els mamífers que cacen entre les branques dels arbres. En tot cas, la singularitat de molts nius d’ocell és un espectacle més dels molts que ofereix la selva plujosa equatorial.
El ritme de la vida animal
La selva plujosa, bé que mancada d’una estacionalitat clara, està sotmesa a ritmes, sovint de natura interna dels organismes, tal com ja s’ha posat de relleu a propòsit de la vegetació. En el cas de la fauna, la situació es repeteix. Alguns casos són tan espectaculars que mereixen un tractament específic.
El cas de les formigues viatgeres africanes
Un dels flagells més notables de la selva plujosa africana són les formigues viatgeres del gènere Anomma. Aquestes formigues són el resultat d’un llarg procés evolutiu completament aliè a l’experimentat pel llinatge dels ocells o dels mamífers. Es disposen en eixams de lluitadores que són capaces de caçar preses molt més grans que elles atacant-les concertadament en grup, al contrari de moltes altres formigues que cacen individualment i que poden obtenir preses una mica més grans que elles. Les colònies d’Anomma lluitadores poden ser formades per fins a 20 milions de germanes, o el que és el mateix, per 20 milions de boques, de mandíbules i d’agullons.
La massa de formigues escombra tot el terra de la selva, empassant-se la fullaraca i les plantes petites a un ritme de més de 20 m per hora, en un front d’entre 10 i 20 m. En una activitat frenètica, les formigues obreres avancen i reculen sense parar, funcionant d’acord amb la voluntat col·lectiva de la colònia, trabada per les hormones gasoses secretades per les seves companyes. A mesura que s’acosta l’onada de formigues, els animals amb possibilitats de retirar-se ho fan, marcant el progrés de la colònia amb frènetics salts i voleiades. La majoria de les víctimes de les formigues són immòbils (larves d’insecte poc amagades, pollets abandonats al niu pels seus pares) o també les criatures lentes i dissortades que no poden volar o que, confoses per la por, han corregut en la direcció equivocada. Són pocs els vertebrats adults que són atrapats per l’eixam, excepte en aquells casos que es trobin debilitats o amb deficiències o, en el cas de les serps, que tinguin els reflexos alentits mentre estan digerint un bon àpat. Quan les formigues aconsegueixen una presa animal en cobreixen tot el cos, la paralitzen i l’esmicolen. Els fragments del cos de la víctima són transportats per l’escamot atacant cap a l’eixam, i d’allí, transportats per una espessa columna de formigues cap al quarter general o “campament” temporal de les formigues. Un cop arriba allí, les part nutritives són processades per a alimentar la innombrable progènie.
Una colònia d’Anomma té una existència rítmica. Cada una de les colònies té una reina, un individu monstruosament fecund que fa més de 5 cm i que és més gran que cap altra formiga al món. Aquestes reines poden pondre de 3 a 4 milions d’ous cada mes, la majoria dels quals són produïts en períodes de 5 o 6 dies escalonats aproximadament cada 3 setmanes. Segons la disponibiiltat d’aliment, les formigues romanen al mateix “campament” entre una setmana i tres mesos, període que correspon a l’anomenada “fase sedentària”. Durant aquest temps les formigues excaven i ocupen un elaborat sistema de túnels subterranis que poden penetrar fins a 4 m sota terra. Aquests segurs campaments subterranis contrasten amb els que construeixen les seves parentes sud-americanes, les formigues del gènere Eciton, els campaments de les quals s’eleven mitjançant amples estructures verticals en forma de columna i que són aixecades només amb els cossos de les mateixes formigues enllaçats per a formar torres.
Durant la fase sedentària, els eixams d’Anomma surten del campament per a tornar-hi a entrar més tard diàriament. Mentrestant, els ous comencen a obrir-se i les larves són alimentades tan profusament com ho permet la població d’animals susceptibles de ser capturats com a presa de la rodalia. Abans d’arribar a l’edat adulta, però, les larves d’obreres han de convertir-se en pupes, i és l’acumulació d’una població de pupes prou gran el que fa que les formigues hagin d’emigrar del campament. Carregant la reina i la seva progènie, la colònia sencera reprèn el seu camí per la selva fins a trobar una nova residència uns centenars de metres més enllà. En aquesta migració participen tantes formigues que la columna que formen en desplaçar-se de campament a campament sovint dura uns quants dies.
Les migracions d’ocells
A una escala molt diferent, però amb semblances pel que fa a la manera d’alimentar-se el fenomen de la migració dels ocells també contribueix a instaurar ritmes en l’ecologia de la selva plujosa. Molts dels ocells que hi habiten són migradors, és a dir, o bé neixen a la selva plujosa i volen cap a latituds temperades durant l’estiu per aprofitar-hi l’abundància estacional d’insectes, o bé neixen a l’estiu temperat i es protegeixen de l’hivern a la selva plujosa.
Uns estudis d’anellament han demostrat que sovint, després d’haver volat milers de quilòmetres, els ocells retornen exactament al mateix indret de la selva d’on sortiren, exactament als mateixos arbres o a la mateixa zona del sotabosc. Aquesta fidelitat demostra fins a quin punt resulta conservadora la natura: si un hàbitat és predicible, per què s’hauria de córrer el risc de visitar altres arbres que no siguin els que ja van mantenir viu l’ocell l’any precedent? És millor deixar les provatures per a aquells que no tenen cap elecció, per als individus que han estat desplaçats dels seus arbres per un rival territorial o per una calamitat ecològica important.
La migració té sentit per als ocells de la selva plujosa per dues raons. En primer lloc, els estius temperats ofereixen una gran abundància d’aliment potencial, atès que l’activitat vegetativa de les plantes, i per tant la producció es concentra en uns pocs mesos, durant els quals les poblacions d’insectes també pugen a xifres astronòmiques; l’augment d’aliment a l’estiu sempre tendeix a ultrapassar la capacitat de les poblacions locals per a explotar-lo de manera eficient. En contrapartida, la variació estacional de les fonts d’aliment a la selva plujosa tropical és trivial comparada amb la de les àrees temperades; si un ocell sap què ha de menjar i com trobar-ho, i què ha d’evitar i com reconèixer-ho, mai no li faltarà aliment ni s’exposarà a cap risc. Així, la selva plujosa és un bon indret per sobreviure mentre s’espera l’estiu, o fins i tot, mentre l’ocell sigui prou adaptat a les peculiaritats de la selva, per a criar-hi. Tanmateix, les mateixes peculiaritats de la selva plujosa, com ara les defenses químiques sofisticades, fan que els recursos alimentaris per a cada espècie particular a la selva plujosa es trobin en quantitats molt limitades, cosa que restringeix dràsticament la taxa de creixement de la població de qualsevol ocell i també la seva densitat de conjunt.
Els ocells migradors de les selves plujoses equatorials aprofiten un ecosistema bipolar molt ample: en un pol hi ha la predicible superabundància estacional de l’estiu temperat, a l’altre la no menys predicible i en aquest cas permanent —no estacional— abundància, dins d’una línia d’austeritat, de la selva plujosa. Mitjançant els viatges d’un pol a l’altre, els ocells poden maximitzar les seves aptituds reproductives individuals i, incidentalment, la taxa de creixement de les seves poblacions.
Però la migració també té el seu preu, que inclou els contratemps d’un viatge llarg, la competència amb els rivals més ràpids, les exigències de l’orientació i la bona memòria, i els requeriments d’un comportament social i d’alimentació flexible. Invertint en aquestes adaptacions, els migradors necessàriament han de convertir-se en generalistes, i la selva plujosa quasi no té lloc per als generalistes. Els ocells que no són migradors i que viuen i creixen a la selva plujosa durant tot l’any tendeixen a ser especialistes; la pressió de selecció els encamina a menjar i a fer cada cop menys coses, però amb més eficiència.
Que hi hagi especialistes a centenars, distribuïts ajustadament en l’hàbitat de la selva plujosa, deixa poc espai per a competidors menys especialitzats. Això, al seu torn, fa que la selva plujosa sigui encara menys hospitalària per als migradors del que podria ser i fa encara més atractiva l’opció de marxar cap a l’estiu temperat. Aquesta via és oberta, tanmateix, a poques espècies, ja que la majoria dels ocells de la selva plujosa (com la majoria dels altres organismes d’aquest bioma) més aviat evolucionen en el sentit d’un refinament en l’especialització al llarg de les generacions.