L’economia i el desenvolupament dels pobles que viuen a les selves
La selva plujosa tropical no és un hàbitat gaire favorable per als vertebrats terrestres. Com que la major part dels aliments d’origen vegetal disponibles es troben a les capçades dels arbres, són sobretot ocells, rèptils (especialment serps), rates-pinyades i mamífers arborícoles (majorment simis) els vertebrats que exploten millor aquests recursos. Els humans no hi troben, doncs, un ambient gaire fàcil de poblar. Això no vol dir que no hi visqui gent extraient-ne els recursos.
Els nòmades de la selva
El fet que a les selves puguin viure poblacions humanes, generalment marginals i greument amenaçades i amb una tecnologia molt reduïda (en alguns casos no coneixen l’ús de la metal·lúrgia, si bé solen posseir alguns estris de ferro importats), tot i que sobrevisquin en nombre molt reduït, posa de manifest l’efectivitat de les adaptacions culturals humanes pre-agrícoles i pre-industrials. Els caçadors i recol·lectors poden sobreviure mercès a la baixa densitat de població i a les seves habilitats a l’hora d’extreure recursos del medi. En l’actualitat estan amenaçats, no per l’ambient natural, sinó pels seus veïns agricultors i ramaders, per governs mal dirigits i per una sèrie d’influxos culturals occidentals que per a ells són destructius.
La majoria dels caçadors-recol·lectors i molts dels agricultors itinerants no tenen assentaments estables, ja que s’instal·len temporalment en petits campaments fets amb materials poc duradors que proporciona la mateixa selva; aquest és el cas, per exemple, dels pigmeus de les selves africanes. Les densitats d’aquestes poblacions són molt baixes, de prop d’un habitant per cada 4 o 5 km2. Per tant, les àrees de selva per on es desplacen poden considerar-se virtualment desertes de poblament humà. I encara més si pensem que, en l’actualitat, si més no pel que fa als pigmeus africans i algunes altres ètnies de caçadors-recol·lectors de les selves plujoses, com els aeta de Luzon o els penan de Borneo, els seus campaments mai no estan gaire allunyats dels assentaments de pobles agricultors més sedentaris amb els quals tenen establertes relacions permanents d’intercanvi, i dels quals obtenen també productes agrícoles.
Els caçadors sedentaris i els agricultors itinerants
Com hem apuntat abans, a la pràctica, són molt pocs els pobles de les selves plujoses dedicats exclusivament a la cacera i la recol·lecció itinerant. La majoria de caçadors i recol·lectors practiquen també alguna forma d’agricultura, encara que sigui itinerant. Es tracta, aleshores, de pobles d’hàbits més o menys migradors, però no exactament de nòmades.
A moltes àrees de l’àmbit de les selves plujoses es repeteix força el model d’una densitat de població molt condicionada per la fertilitat relativa dels sòls que són posats en conreus i també parcialment per les tècniques agrícoles aplicades. Així, a Malaca, mentre que els semang caçadors-recol·lectors tenen unes densitats de població no gaire allunyades de les que hem indicat per als humans d’economia de cacera i recol·lecció (1 habitant per 4 o 5 km2), tant els semang agricultors itinerants com els jakun, horticultors, tenen densitats de població fins a 10 o 20 vegades superiors.
Les ciutats colonials
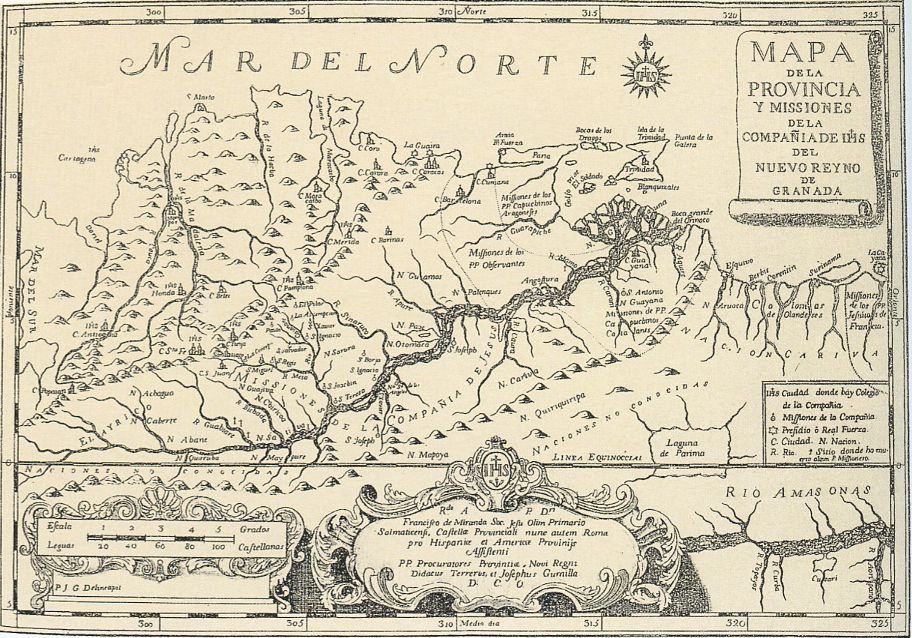
La implantació de missions en plena selva representà sovint una punta de llança del sistema d’ocupació humana occidental en l’àmbit de la pluviïsilva. Aquest mapa de mitjan segle XVIII, pertanyent a l’obra de Josep Gumilla “El Orinoco ilustrado y defendido” (Madrid, 1741) mostra les diferents missions, sobretot jesuítiques, que existien aleshores a l’àrea orinòquica.
Biblioteca de Catalunya
En la mesura que l’àmbit de les selves plujoses és percebut com a poc acollidor per als humans, es tendeix a identificar-lo com un “desert verd”. Si bé la imatge pot ser vàlida per a algunes àrees de la “terra firme” amazònica, dels interfluvis de la conca del Zaire o per a algunes zones de l’interior de Malaca, d’Indoxina o de les grans illes insulíndiques, difícilment es pot conciliar amb altres àrees que figuren entre les més densament poblades del planeta, com poden ser Singapur, algunes àrees de Vietnam, de Java o de les altres illes d’Indonèsia, les quals forneixen grans contingents de transmigrants, o algunes àrees costaneres de l’àmbit de les selves plujoses d’Àfrica i del Brasil. En tot cas, les poblacions autòctones, més o menys densament, ocupen un espai al si de les selves plujoses d’arreu del món.
Des de temps passats, en algunes àrees de l’àmbit de les selves plujoses s’han desenvolupat civilitzacions urbanes que han concentrat en determinades àrees poblacions considerables. Aquest seria el cas de la civilització maia, de les ciutats iuoruba, a l’Àfrica, i de l’imperi khmer, al SE asiàtic. Però, a partir de l’expansió colonial europea dels darrers 500 anys, un nou tipus d’implantació urbana començà a aparèixer a l’àmbit de les selves humides. Unes implantacions que inicialment tenien establerts més lligams amb una llunyana metròpoli que amb el rerepaís que les envoltava, amb una població majoritàriament vinguda d’aquella metròpoli o portada d’alguna altra banda, amb unes activitats econòmiques totalment diferents de les de les poblacions tradicionalment establertes en el seu entorn més proper.
Les primeres grans ciutats colonials de la selva nasqueren a Amèrica. Ja en el seu primer viatge, el 1492, Colom deixà establerta una part dels seus tripulants en un punt de la costa septentrional de la Hispaniola que anomenà Navidad (prop de l’actual localitat de Cap-Haïtien, a Haití), però aquest primer establiment europeu a Amèrica fou destruït i els seus habitants morts abans que hi tornés el descobridor. Aquest, en el segon viatge, fundà La Isabela, i més tard, el seu germà Bartomeu, Santo Domingo. Aquests establiments foren les primeres bases de la conquesta i la colonització castellana del Nou Món abans que, a partir dels primers anys del segle XVI, s’establissin els primers nuclis colonials a terra ferma del continent americà, els més importants dels quals, però, evitaren l’àmbit de les selves plujoses que els era força hostil i al qual no eren adaptats. Els primers establiments portuguesos a les costes atlàntiques del Brasil, en canvi, es feren majoritàriament dins l’àmbit de la “mata” atlàntica, com és el cas de São Vicente i São Paulo (1532), Santos (1545), Bahia (1549), Recife (1563) o Rio de Janeiro (1565).
Tot i que els portuguesos, en la seva exploració de la costa occidental africana ja havien anat establint algunes factories i forts (São Jorge da Mina o Elmina, a la costa de l’actual Ghana, el 1482; São Tomé, el 1493; Mpinda, a la desembocadura del Zaire, el 1501), fou la mateixa colonització americana la que estimulà la creació de les primeres implantacions colonials a la costa africana del golf de Guinea destinades a assegurar el subministrament d’esclaus per a mà d’obra a les colònies americanes. Al llarg dels segles XVII i XVIII totes les potències europees n’establiren i fou des d’aquests establiments que s’inicià, al segle XIX, l’exploració i la penetració colonial de l’interior del continent africà. També al S i SE d’Àsia s’establiren, des del segle XVI, factories colonials d’aquest tipus, malgrat l’existència prèvia d’una certa xarxa urbana autòctona; seria el cas de Colombo, fundada pels portuguesos a Sri Lanka el 1505, o de Batàvia, l’actual Djakarta, fundada pels holandesos a Java el 1619.
La xarxa urbana moderna
Les àrees urbanes implantades a l’àmbit de les selves plujoses equatorials són nombroses i amb històries molt diverses. La majoria comparteixen un tret comú amb les altres ciutats del Tercer Món: l’explosiu creixement demogràfic dels darrers decennis i l’aclaparador èxode rural que han d’acollir. Són, en efecte, centres relativament més desenvolupats i més ben connectats amb els centres de decisió de les antigues o de les noves metròpolis que el territori rural fortament subdesenvolupat que les envolta.
Al començament, les ciutats colonials eren poc més que un port, una guarnició, uns magatzems i una administració, però moltes de les americanes, amb el desenvolupament de les colònies de les quals eren porta d’entrada i de sortida de tot el comerç, esdevingueren aviat veritables ciutats. Durant el segle XIX, amb la difusió de la navegació a vapor i el necessari establiment d’una xarxa d’escales d’avituallament, algunes, principalment les sotmeses a la colonització britànica (Singapur, Hong Kong), es consolidaren com a centres comercials i la majoria no han deixat de créixer a partir de la descolonització. Djakarta, la Batàvia colonial, també ha tingut un creixement més acusat a partir de la descolonització (1949).
De la vintena d’àrees urbanes d’aquest àmbit que superen avui el milió d’habitants, ben poques tenen una història que es remunti a més de 500 anys, i la majoria eren nuclis insignificants al començament del segle XIX, i ciutats ben secundàries al començament, i fins i tot a mitjan segle XX. Ha estat el procés postcolonial de la segona meitat del segle XX el que ha conduït a un model d’urbanització dependent, amb elevades taxes de creixement, forts desequilibris socials i àmplies capes de població marginada. São Paulo, avui la tercera àrea urbana del món, ja començà a expandir-se al segle XVII, quan succeí Bahia com a capital del Brasil, i també rebé un impuls important de l’expansió del conreu del cafè entre el 1870 i el 1929, però quan ha accelerat enormement el seu creixement ha estat durant la segona meitat del segle XX. Rio de Janeiro, també capital del Brasil del 1763 fins al 1960, conegué un primer creixement durant la segona meitat del segle XVIII mercès al conreu de la canya de sucre i del cotó i al descobriment de les mines de Minas Gerais, visqué hores d’esplendor com a residència de la cort portuguesa i com a capital de l’imperi brasiler del 1808 al 1889 i no alentí el seu creixement (no tan accelerat, tanmateix, com el de São Paulo) ni tan sols amb la pèrdua de la capitalitat del Brasil el 1960.
Els darrers nouvinguts i les transmigracions
Les colonitzacions dels darrers 500 anys han portat a l’àmbit de les selves plujoses poblacions més o menys nombroses de procedència exterior. Aquestes migracions han estat casos aïllats, i els més notoris han estat el de blancs europeus (sobretot mediterranis) i negres africans (principalment de l’Àfrica occidental) a les Antilles, a Amèrica Central i a Amèrica del Sud, principalment a la façana atlàntica; i el de xinesos septentrionals a l’Àsia meridional, sobretot a Malàisia i a Singapur, on constitueixen minories importants i influents.
Més recentment encara, sovint per iniciativa dels governs, han començat a produir-se fenòmens de transmigració des d’algunes àrees superpoblades de l’àmbit a altres més aviat poc poblades. Aquest procés és particularment rellevant a Indonèsia, on aquests programes ja foren iniciats en època colonial (1905) pels holandesos i han significat el trasllat i el reassentament, amb el suport governamental, de més de 2 milions i mig de persones, procedents principalment de Java però també de Madura, Bali i Lombok, en àrees prèviament poc o gens poblades de Sumatra, Kalimantan (Borneo), Cèlebes i Irian Jaya (la meitat occidental de l’illa de Nova Guinea). Això sense comptar els emigrants espontanis que, sense acollir-se a cap programa governamental, han seguit el mateix camí. El nombre d’emigrants creixé espectacularment durant el període 1979-86, durant el qual la transmigració afectà unes 60 000 famílies per any, amb una inversió pública de prop de 10 000 dòlars nord-americans per família. El ritme s’alentí, a partir del 1986, a causa de la caiguda d’ingressos per la venda de productes petroliers que experimentà el govern indonesi i s’estabilitzà en una xifra inferior a les 10 000 famílies per any, una reducció de població que no arriba a compensar, tanmateix, el vertiginós creixement vegetatiu de la població de Java.
La colonització de les pluviïsilves
Gairebé tots els pobles que viuen a les selves d’Àfrica, el SE d’Àsia, Oceania i Amèrica Central i del Sud han deixat de banda la cacera i la recol·lecció i han adoptat alguna mena d’horticultura, si bé poden també dedicar-se a la cacera; en l’actualitat hi ha pocs casos de pobles de la selva que sobrevisquin només de la cacera i la recol·lecció d’aliments silvestres. Els que ho fan sovint comercien amb pobles agricultors establerts en petits nuclis, però són capaços de trobar el seu propi aliment dins la selva, cosa que ens mostren el seu règim alimentari i la seva tecnologia. Aquests colons, des que arribaren a la selva, han après a integrar-se en l’entorn, a adaptar-s’hi i a gestionar-lo de manera que quedi protegit i alhora que se’n puguin extreure uns beneficis per a sobreviure i comerciar a petita escala. Avui s’estima que dins del bioma de les selves plujoses viuen fins a 50 milions de persones; són pobles que han desenvolupat estils de vida en harmonia amb el seu ambient i que no sotmeten els recursos a pressions excessives.
Un segon tipus de colonització és el dut a terme des del segle XV, que es tracta d’una colonització expansiva, de frontera. Tots els nous colonitzadors que avui tallen i cremen selva al Brasil per a establir-hi camps de conreu i pastures insostenibles pertanyen a aquest segon tipus però han tingut il·lustres precedents en els darrers cinc segles. De tota manera, tant al segle XVI com al segle XX (i probablement al XXI) la necessitat de terres haurà empès un nombre sempre creixent de pobladors cap a l’interior de les pluviïsilves. Els nouvinguts normalment no posseeixen ni els coneixements ni les tècniques adients per a utilitzar els recursos i els serveis que els proporciona el bosc tropical humit, i contribueixen a la seva ràpida destrucció.
Una observació a fer referent als estudis arqueològics de les selves plujoses és que les característiques ecològiques no permeten de mantenir restes orgàniques molt de temps sense que es descomponguin. Com diuen els ecòlegs, la vida a la selva capitalitza tota la matèria orgànica, i la incorpora al cicle biològic degradant-la de pressa per efecte dels descomponedors, fongs i bacteris.
Els humans a les selves africanes
La migració humana cap a l’interior de les selves plujoses va començar en temps prehistòrics. Els primers testimonis d’humans moderns a l’àmbit actual de les selves plujoses equatorials, però, no són restes òssies, sinó testimonis culturals. Durant la darrera glaciació, fa entre 40 000 i 30 000 anys, aparegué a la part oriental de la conca del Zaire (llavors ocupada probablement per una vegetació sabanoide) una població humana amb una cultura de la qual ens han arribat estris de pedra i d’os, alguns en aparença de caràcter no estrictament utilitari, sinó ornamental (cultura sangoana). Alguns autors han atribuït als humans d’aquesta cultura els primers assaigs deliberats de domesticació d’una espècie vegetal, concretament la carbassera vinetera (Lagenaria siceraria), després difosa a les altres àrees tropicals, i present tant a Nova Guinea com a l’Amèrica tropical, des de dates molt remotes.
Els tipus humans de les selves africanes
Encara que no es pot assegurar que la regió de l’E del Zaire on s’han trobat restes de la cultura sangoana estigués ocupada per selves fa 40 000 anys, ni que els humans que l’han produïda fossin els ancessors dels pigmeus ni de cap altre poble de caçadors-recol·lectors o d’agricultors itinerants actual, el fet és que les selves plujoses africanes són relativament més pobres d’espècies del que s’esperaria i una de les causes que han estat invocades per explicar-ho no és altra que la intensa i prolongada influència dels humans. Fins i tot hi ha qui pensa que a l’Àfrica no subsisteix cap resta de selva intacta o primària, sinó que totes les selves plujoses són selves secundàries més o menys velles i, per això mateix, en un permanent estat de transició pel que fa a la composició florística, com a conseqüència d’una explotació mil·lenària.
En les selves africanes s’han desenvolupat civilitzacions que han concentrat densitats de població importants en nuclis urbans, com seria el cas de les ciutats i estats ioruba, i del regne edo de Benín, que cresqueren entorn d’Ifé, el seu comú centre religiós, entre el segle XII i el XVIII. El país ioruba i el reialme de Benín s’estenien (i s’estenen) per tot el SW de l’actual Nigèria a occident del riu Níger i arribaven per l’W fins al riu Mono, desbordant les fronteres occidentals nigerianes actuals i ocupant part dels actuals estats de Benín (només una part del territori del qual estigué sota la sobirania de l’antic reialme d’aquest nom) i Togo. Com a mínim des dels segle XII o XIII, a partir de la fundació d’Ifé, començaren a crear-se en aquest territori nombroses ciutats estat que han anat coneixent al llarg de la història successius períodes d’esplendor i de decadència. Força abans de l’arribada dels primers europeus, al final del segle XV, aquesta era la regió més urbanitzada de l’Àfrica al S del Sàhara, amb ciutats molt extenses i amb una doble línia de muralles. Malgrat una decadència generalitzada a partir del final del segle XVII, la majoria d’aquestes ciutats arribaren fins al període colonial i moltes han conegut un nou esplendor sota aquell règim o després de la descolonització, fins al punt que avui dues d’aquestes ciutats (Ibadan i Lagos) ultrapassen el milió d’habitants, una tercera (Ogbomosho) ultrapassa el mig milió, i una vintena més ultrapassen els 100 000 habitants. El territori en conjunt té una densitat de població que s’acosta als 200 habitants per km2, una xifra comparable a la d’Itàlia.
Els pigmeus centrafricans
Les poblacions més importants de pigmeus són les de les selves africanes: mbuti (o bambuti) de l’Ituri, al Zaire, i binga (o babinga) de l’extrem septentrional de la república del Congo i les regions adjacents del Zaire, de la república Centrafricana, del Camerun i del Gabon, on alguns petits nuclis arriben al centre i el SE del país. Tot i que molts d’ells, sobretot els binga, comencen a sedentaritzar-se, les seves formes de vida tradicionals són totalment nòmades i viuen agrupats en bandes que apleguen una vintena de famílies i que constitueix una unitat de subsistència, tant pel que fa a l’obtenció com al consum d’aliments. Pel que fa a la morfologia, sembla que hi ha una correlació entre l’estatura dels pigmeus africans i el grau de barreja amb altres poblacions locals.
Els pigmeus, en efecte, viuen en petites bandes d’una vintena de famílies, cada una de les quals pot explotar un extens territori, fins i tot de més d’un miler de quilòmetres quadrats. Normalment s’estableixen per un temps, per períodes d’un o dos mesos, en campaments formats per una vintena de cabanes de forma hemisfèrica construïdes simplement amb branques corbades i entreteixides, clavades a terra i recobertes amb fulles de tilipi o mongongo (Megaphrynium macrostachyum i altres marantàcies afins), amb les obertures orientades cap al centre de la clariana on el campament ha estat instal·lat, generalment prop d’un curs d’aigua. Els homes utilitzen per caçar l’arc (els efe) o la xarxa (els mbuti) i exploten la mel dels ruscos d’abelles, mentre que les dones pesquen i recullen tota mena d’aliments d’origen vegetal i animal (fulles, arrels i tubercles comestibles, bolets, cargols, insectes) sense allunyar-se gaire del campament.
Els humans a les selves asiàtiques
El vast territori que comprèn l’àmbit de les selves plujoses al SE d’Àsia té un poblament molt divers. La majoria de les poblacions humanes són integrades per xantoderms, o mongoloides, però també resten poblacions d’humans diminuts: els negritos de les Filipines i altres grups emparentats del SE asiàtic. Els habitants de les selves asiàtiques viuen, en certs casos, de la cacera i la recol·lecció. Els semang de Malaca s’apleguen en bandes de 6 a 12 nuclis familiars amb vincles de parentiu entre elles. Cada una explota en exclusiva un territori d’uns 50 km2 dins del qual construeixen campaments temporals de cabanes fetes de branques i fulles. L’estil constructiu és diferent del dels pigmeus africans; no es tracta de cabanes hemisfèriques, amb l’estructura feta de branques corbades i clavades a terra pels dos extrems, sinó d’uns paravents oberts cap a l’espai comú suportats per una estructura que consisteix en tres o quatre estaques gruixudes clavades a terra amb una lleugera inclinació cap a la clariana que centra el campament, l’extrem lliure de les quals reposa sobre l’enforcadura d’uns puntals inclinats en direcció oposada. A les estaques que suporten el paravent es lliguen branques travesseres i finalment es fa un entreteixit de fulles i lianes.
Fora d’Àfrica, les poblacions més semblants morfològicament a les dels pigmeus africans són els escassos supervivents (jarawa, andaman i onge) dels antics habitants de les illes Andaman (golf de Bengala), si bé se’n diferencien per les proporcions més gràcils del cos i en particular del cap, no tan gros en relació amb el cos, i del nas, no tan ample i aixafat com el dels pigmeus africans. Altres poblacions d’humans diminuts de les selves equatorials de l’Àsia meridional són els semang, abans esmentats, que viuen a les selves del centre de la península de Malaca frontereres entre Tailàndia i Malàisia, i els negritos o aeta de les Filipines, que viuen principalment a les muntanyes de l’E de Luzon, però també en altres àrees de la mateixa illa i en altres illes menors. En algunes illes de la Sonda (Timor, Sumba, Badjawa, Celebes, Flores), a Sumatra i a Nova Guinea han estat assenyalades poblacions de petita talla, com també a les muntanyes Nilgiri, al S de l’Índia, i en algunes àrees del sud de Cambodja. Totes aquestes poblacions diminutes sembla que són els descendents de la primera onada d’homes moderns que arribà al sud i el sud-est asiàtic, que quedaren desplaçats en zones menys acollidores per les poblacions neolítiques posteriors de parentiu genètic amb les poblacions mongoloides de més al nord.
Els humans a les selves oceàniques
Els primers humans arribaren a les costes de Nova Guinea fa, pel cap baix, 50 000 anys, però no hi ha testimonis de la seva penetració en les terres altes d’aquesta illa fins fa uns 26 000, tot i que ja havien arribat fa uns 40 000 anys a Austràlia i en fa més de 30 000 a Nova Irlanda. Tot plegat, com també les formes de vida dels escassos pobles caçadors-recol·lectors que subsisteixen avui a les selves plujoses equatorials, fa pensar que els humans han arribat tardanament a ocupar i explotar aquests ambients i que ho han començat a fer només marginalment o forçats per circumstàncies adverses (conflictes amb altres grups humans, exhauriment o limitació dels recursos de les àrees ocupades prèviament a la sabana adjacent, etc.).
Els melanesis són probablement els humans més plenament lligats a les selves plujoses tropicals oceàniques. Tots ells viuen en aquest àmbit o bé en el de les selves nebuloses de les terres altes equatorials, llevat d’uns quants que viuen en àrees tropicals d’estació seca més accentuada, que corresponen a l’àmbit dels boscos monsònics o de les sabanes. Al NE d’Austràlia, precisament en àrees de selva plujosa de l’altiplà d’Atherton (Atherton Tableland), entorn del llac Barrine, i també més al N, fins a la capçalera del riu Daintree, hi ha algunes poblacions, principalment els ngatjan, els tjapukai i els wultura, de característiques semblants a les dels semang i els negritos. Aquestes poblacions, anomenades barrineanes, s’estima que apleguen uns 500 individus sense barreja amb les poblacions veïnes; hom pensa que podrien ser, doncs, les romanalles de les primeres onades d’humans moderns del remot continent austral, els avantpassats dels quals podrien haver-se establert a l’altiplà d’Atherton fa prop de 40 000 anys.
Els humans a les selves americanes
Sembla que els testimonis humans més antics de l’Amèrica tropical són els de l’abric de Pedra Furada, a l’estat de Piaui, al Brasil, als quals alguns autors atribueixen una antiguitat d’entre 32 000 i 23 000 anys (còdols treballats, restes de focs i fragments de roca pintada), si bé hi ha molta polèmica sobre aquestes datacions. Aquests jaciments són en una àrea de “caatinga” que versemblantment no ha estat mai ocupada per selva, i que avui es troba allunyada 700 o 800 km de les selves més properes.
La majoria de caçadors i recol·lectors de les selves americanes practiquen actualment alguna forma d’agricultura, encara que sigui itinerant. Es tracta, aleshores, de pobles d’hàbits itinerants, però no exactament de nòmades. Aquest seria el cas de bona part dels pobles de la “terra firme” amazònica. Quant a l’hàbitat i l’ocupació de l’espai, no hi ha grans diferències entre ells i els nòmades purs, bé que els seus poblats són normalment més estables i, de vegades, són reaprofitats parcialment després d’un temps d’abandó. A la “terra firme” amazònica les densitats de població no són gaire diferents de les de les selves de l’Ituri o de Malaca, i els poblats consisteixen en una o més cabanes de planta oval, rectangular o amb un extrem arrodonit, cada una ocupada per diverses famílies i amb espais diferenciats segons els usos o les persones a qui estan destinats.
A la “várzea”, la plana inundable de l’Amazònia, les densitats de població ja són més elevades (uns 15 habitants per quilòmetre quadrat), i al llarg de les riberes els assentaments humans són més nombrosos i més permanents que a la “terra firme”, com correspon a una situació en la qual la terra té una fertilitat més gran, renovada per les periòdiques revingudes del riu, de manera que les clarianes obertes al bosc poden ser conreades indefinidament sense que calgui obrir-ne de noves al cap d’un temps i abandonar les antigues. Els recursos animals i vegetals espontanis de la selva també són més abundants, variats i presents al llarg de tot l’any.
Un cas peculiar, que ha estat àmpliament discutit per molts antropòlegs, seria el dels warao del delta de l’Orinoco que, tot i ser plenament caçadors-recol·lectors, són sedentaris i, a diferència d’altres pobles caçadors-recol·lectors, viuen en societats igualitàries, amb escassa o nul·la diferenciació social. Entre els warao hi ha caps, consellers i sacerdots (“wisidatu”) que no participen a les tasques que exigeixen esforç físic però controlen els processos d’obtenció i conservació del recurs bàsic, que per a ells és el moll d’una palmera d’aigual, el “moriche” (Mauritia flexuosa).
Les poblacions d’amerindis de les selves plujoses no estan tan diferenciades de les de la resta del continent americà com ho estan les melanoafricanes respecte dels habitants dels deserts o les regions mediterrànies de l’Àfrica septentrional. Com que la major part de les poblacions ameríndies procedeixen de grups de pobladors relativament poc nombrosos i d’arribada relativament recent al continent americà procedents d’Àsia, les divergències genètiques entre unes poblacions i unes altres no són tan grans com les que es troben entre els pobladors d’altres continents. Malgrat tot, els habitants d’Amèrica Central són més semblants genèticament als pobladors del subcontinent nord que als de l’Amèrica del Sud. Per contra, la seva fragmentació i les migracions gairebé ininterrompudes dels darrers mil·lennis han estat la causa d’una gran varietat lingüística.
L’ocupació humana de les pluviïsilves de l’Amèrica del Sud és més recent; els primers immigrants que van arribar a l’Amazònia ho van fer a través d’Amèrica Central, fa entre 12 000 i 15 000 anys. L’agricultura de l’Amazònia deu datar de fa 5 000 anys. Després que la primera onada de gent arribés a la regió, l’Amazònia devia resultar relativament poc afectada per les immigracions subsegüents, fins a l’època de la colonització europea.
Quan els primers europeus van arribar a l’Amèrica del Sud, l’Amazònia era la llar de diverses cultures natives diferents i complexes, amb produccions artístiques sofisticades i extenses xarxes comercials. S’estima que aleshores a l’Amazones vivien 8 milions de persones, però aquest nombre va disminuir ràpidament amb la persecució, l’esclavitud, les guerres i les malalties introduïdes pels europeus. Avui hi queda aproximadament el 4% del nombre de pobladors indígenes que hi havia a l’arribada dels europeus a Amèrica. Uns quants grups petits d’indis continuen sense tenir cap contacte, o molt poc, amb el món exterior, però la seva existència tradicional és precària a causa de l’estesa desforestació dels boscos de la regió.
El “boom” del cautxú, a primers d’aquest segle, va atreure molta gent cap al Brasil, i avui els descendents d’aquests i d’altres immigrants són els habitants del bosc. Els “caboclos”, mestissos d’europeus i indis, viuen de l’agricultura, la caça i la recol·lecció de productes del bosc. Tenen amplis coneixements sobre els recursos forestals dels quals depenen i entenen els processos ecològics de la pluviïsilva.
Durant els últims vint anys la colonització de l’Amazònia s’ha incrementat espectacularment. Pagesos immigrants, treballadors rurals sense terres i miners han estat animats a traslladar-se cap a zones cada vegada més remotes de les pluviïsilves. La política del govern brasiler ha afavorit aquests nous emplaçaments perquè signifiquen un mitjà d’alleugerir les pressions resultants d’una distribució injusta de les terres. Al Brasil, per exemple, menys de l’1% dels terratinents controla quasi la meitat de la terra cultivable del país. Els pagesos pobres sense terres han estat estimulats a desplaçar-se des del sud i el nord-est del país cap a l’interior dels estats amazònics. A Colòmbia el 80% de la terra cultivable està en mans del 10% dels propietaris, i els habitants es veuen obligats a desplaçar-se. Malauradament, els nous habitants de l’Amazònia no tenen els mateixos coneixements sobre l’ambient de la pluviïsilva que els indis, els “caboclos” i altres grups que tradicionalment han habitat el bosc.
Les adaptacions dels humans dins l’àmbit de les selves
Malgrat les diferències físiques i culturals que poden trobar-se entre les diferents poblacions d’humans moderns, el cert és que sembla que hi ha acord que el parentiu de la població del globus no remunta més enllà dels últims 100 000 o 150 000 anys. És des d’aqueixa data que, creuant deserts, travessant oceans, instal·lant-se en jungles o en els extrems de latitud més elevada, han adquirit aspectes i han desenvolupat maneres de viure ben diferents, condicionades per les exigències de l’entorn.
En la seva expansió pel planeta, els humans distribuïren els gens polimòrfics (que ens conten les diverses relacions de parentiu entre les poblacions) i veieren modelada la seva fesomia per efecte de la selecció natural, quan s’instal·laren en un determinat bioma suficient temps perquè pogués actuar. És a dir, tant les característiques genètiques de l’espècie (els polimorfismes: molècules amb variació genètica neutra, però de funció coincident), com les fenotípiques, són les que marquen les diferències entre els humans; però, malgrat el que es podria intuir, són les característiques genètiques les que marquen les diferències causades pels 100 000 (o més) anys d’història de l’espècie, les que permeten de reconstruir els esdeveniments històrics; mentre que els caràcters visibles (que varien, a més, de manera contínua) indiquen on han viscut els darrers ancessors. En conseqüència, poblacions que visquin en el mateix tipus d’entorn però separades des de llarg temps, poden molt bé coincidir en l’aspecte físic i divergir força en el genètic. Aquest és el cas dels africans i dels habitants de Nova Guinea: aparentment s’assemblen força, però genèticament, els papuans són més pròxims al xinesos del sud o als vietnamites que no pas als africans. Les diferències externes no impliquen, ni de lluny, diferències de parentiu.
Ha estat la indiferenciació fisiològica dels humans, és a dir, l’elevat grau d’adaptabilitat, el que els ha permès l’adaptació als diversos entorns climàtics. La capacitat instrumental i la intel·ligència reflexiva (amb capacitat d’aprenentatge, memòria i previsió) els han afavorit el desenvolupament d’una eina extrasomàtica: la cultura. Aquestes característiques particulars han permès l’adaptació a tots els ambients, i l’espècie humana ha esdevingut ubiqua en el planeta. En realitat el terme adaptació és un terme relatiu, però, en qualsevol cas, comporta millores en la qualitat de vida d’un individu dins un ambient determinat. En parlar d’adaptació, en antropologia, hom pot referir-se a l’adaptació d’una població a un determinat entorn, o bé de l’adaptació d’un individu després d’un canvi de situació; en aquest darrer cas, es prefereix el terme aclimatació.
En el cas de l’adaptació d’una població, intervé la selecció natural; la variabilitat genètica dels humans proporcionada per les mutacions antigues o recents ha permès la selecció de triar automàticament afavorint les mutacions avantatjoses i eliminant les desfavorables per a la vida d’una població en un determinant entorn; d’aquesta manera, s’han diversificat fenotips en biomes dissemblants. Les mutacions favorables en un ambient àrtic no ho són en zones tropicals. La selecció natural garanteix la supervivència del millor adaptat a unes condicions concretes: clima, fonts d’aliment i resistència a les malalties en són les més importants. Resumint, l’aspecte extern és fruit de la tendència de la vida a intentar reduir l’estrès.
Malgrat la flexibilitat pròpia de l’espècie, que ha proporcionat la possibilitat de sobreviure en molt diversos climes, els humans requereixen unes condicions peculiars de temperatura, insolació, humitat, etc. Si aquestes condicions s’allunyen molt dels valors òptims, es pot parlar d’estrès fisiològic, que desencadenarà tot un seguit de mecanismes per tal de mantenir l’homeòstasi. El conjunt de reaccions destinades a recuperar l’homeòstasi poden encabir-se en el concepte d’aclimatació, abans esbossat. L’aclimatació és la resposta d’un organisme davant de canvis més o menys sobtats, i no comporta canvi genètic, com és el cas de l’adaptació. El fet de ser una espècie homeoterma, és a dir, que regula fisiològicament la seva temperatura corporal, els humans han desenvolupat mecanismes de termòlisi per si la temperatura exterior augmenta molt, i de termogènesi en cas contrari. De la mateixa manera, també tenen mecanismes destinats a mantenir la humitat corporal i per aturar les radiacions ultraviolades que podrien provocar mutacions, però permetent alhora de rebre suficient llum per a finalitzar la síntesi de la vitamina D. Per totes aquestes raons els humans suen, tremolen, tenen nassos amples o estrets, o es posen bruns a l’estiu.
L’aclimatació, doncs, és una resposta fisiològica individual davant d’un canvi en les condicions externes. Però quan un grup humà ha romàs temps suficient en un lloc determinat, les condicions climatològiques han fet avantatjoses certes característiques, les més apropiades per a l’entorn, de manera que tindran més èxit els portadors de fenotips més adients per al clima, les fonts d’aliment o la resistència a malalties. Així doncs, ha estat el clima, i no cap raó de parentiu, el responsable de la morfologia externa dels grups humans; el “disseny” humà, no és més que el reflex d’on han viscut els ancessors. En el cas de les poblacions humanes establertes a les selves des d’antic, en haver estat sotmeses a la pressió de la selecció natural, han vist modificada la seva fesomia segons el clima. Les característiques més importants que s’han dibuixat en la morfologia humana d’aquestes poblacions han afectat el color de la pell, com a defensa davant la insolació; l’amplada del nas, com a conseqüència del grau d’humitat i l’escalfor ambiental; i l’estatura, que segons alguns autors és efecte també del clima de les selves.
El color de la pell
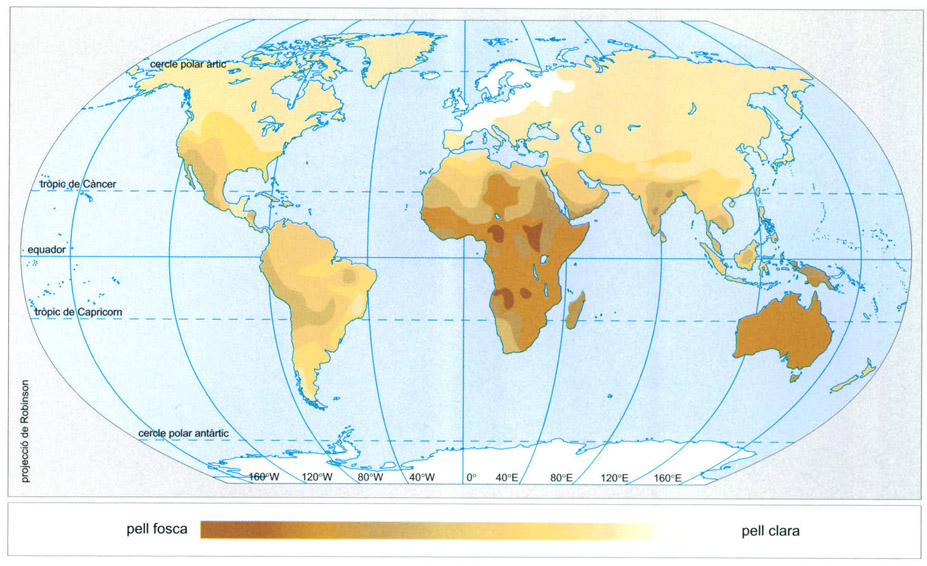
Distribució del color de la pell en els humans abans de les grans migracions dels darrers segles. Ha estat la selecció natural la responsable de la distribució del color de la pell abans dels grans viatges i de les migracions de grups humans esdevinguts els darrers 500 anys. Molts racistes de pell clara han de pensar que no gaire lluny en el seu arbre genealògic els seus progenitors tenen la pell fosca.
Editrònica, a partir de dades proporcionades pels autors
La coloració fosca de la pell en els humans és un caràcter vinculat amb les poblacions de les regions tropicals del planeta i s’associa a la protecció davant la radiació ultraviolada, més abundant a les latituds tropicals que no a les més septentrionals o meridionals. Tot i que el mecanisme pel qual s’ha seleccionat la pigmentació de la pell dels humans no ha estat plenament elucidat, i que s’especula que els ancestres dels humans podrien tenir una pell fosca sota el pèl, sembla que a les latituds equatorials la pressió de selecció ha actuat per la via d’afavorir els fenotips amb pells posseïdores d’una capa protectora de les radiacions ultraviolades capaç d’evitar danys fotoquímics com càncers de pell o hipervitaminosi per excés de producció de vitamina D. A mesura que grups d’humans abandonaven Àfrica i ascendien en latitud, els individus de pell més clara es veien afavorits d’alguna manera; la despigmentació relativa de les poblacions extratropicals seria un caràcter secundari resultat de l’avantatge selectiu de les pells clares per a la síntesi de vitamina D en latituds amb menor intensitat de radiació ultraviolada; s’haurien seleccionat les pells clares per no patir hipovitaminosi D. De tota manera, la selecció continua actuant, ja que en casos de poblacions de pell clara retornades a les latituds tropicals en temps relativament recents a l’escala de la durada de l’evolució humana, com els amerindis o alguns polinesis, s’observa una coloració progressivament més fosca com més a prop de les latituds equatorials viuen i com més temps fa que s’hi han instal·lat.
Les vitamines i el color de la pell
La vitamina D, o factor antiraquític, és molt escassa a la majoria dels aliments usuals (el peix, sobretot el greix de peix, n’és excepcionalment ric), però pot ser sintetitzada per l’acció de la radiació ultraviolada a partir d’un esteroide present a les cèl·lules de les capes inferiors de la pell humana: el 7-dehidrocolesterol. La vitamina D és essencial per al metabolisme del calci i del fòsfor: controla l’absorció del calci pels budells, en regula l’excreció pels ronyons i ajuda a la mineralització de l’esquelet i, en conseqüència, al creixement normal dels infants. Els infants que no disposen de la quantitat adient de vitamina D (hipovitamínics) no calcifiquen adequadament l’esquelet en creixement, de la qual cosa resulten deformacions, principalment de cames i pelvis, que són conegudes amb el nom de raquitisme i que poden arribar a ser causa de mort. D’altra banda, en casos de raquitisme lleu en dones s’han donat problemes a l’hora de parir per reducció del canal del part. Tanmateix la hipervitaminosi també és perillosa, perquè causa la calcificació de teixits tous i pertorba la funció renal.
Si bé necessària per a la síntesi de la vitamina D, la radiació ultraviolada també representa un risc de cremades i de càncers de pell; els humans de pell fosca molt rarament en pateixen, de càncer de pell, mentre que la incidència entre les poblacions de pell despigmentada és més elevada com més a prop de l’equador viuen. Així, per exemple, a la població de les illes Britàniques (al voltant dels 55° de latitud N) la taxa d’incidència dels càncers de pell és de 28 casos per 100 000 entre els homes i de 15 per 100 000 entre les dones, mentre que a la població d’origen britànic de Sud-àfrica (al voltant dels 30° de latitud S) és de 133 i 72 per 100 000 respectivament. A la població blanca de Queensland, a Austràlia (al voltant dels 20° de latitud S), també majoritàriament d’origen britànic, arriba a ser de 265 i 156 per 100 000, respectivament.
En el cas dels habitants de les selves plujoses s’ha argumentat que el suposat valor adaptador de la pell fosca enfront de l’excés de radiació no existiria en unes poblacions que ocupen un hàbitat on la radiació que efectivament arriba a terra resulta molt disminuïda pel filtre que representa el gruix de la vegetació. La realitat és que la majoria dels humans de les selves plujoses tropicals viuen exposats a una radiació solar incident molt considerable. Tret d’alguns pobles exclusivament caçadors-recol·lectors que viuen, efectivament, dins de la selva, en constant desplaçament, la majoria de les poblacions de les selves plujoses practiquen una o altra forma d’agricultura i, amb aquest objecte i amb el d’establir les seves habitacions, obren clarianes on passen la major part del temps de treball i de descans. Fins i tot la majoria dels qui no practiquen l’agricultura busquen més aviat les vores dels rius o les clarianes o les vorades de boscos per a les seves activitats i els seus assentaments, per provisionals que siguin.
Les poblacions de pell més fosca del planeta són al continent africà. La pigmentació dels melanoafricans és determinada per la dimensió dels melanosomes (doble que la dels melanosomes dels blancs europeus), però les diferències de la pell no s’acaben amb el color: el derma és més coherent, la capa còrnia més gruixuda i les glàndules sudorípares més actives que en la pell dels blancs.
Les melanines i altres pigments epidèrmics
La coloració de la pell humana depèn principalment de la producció de melanines (eumelanines i feomelanines), pigments de color bru fosc produïts pels melanòcits, unes cèl·lules especials, amb prolongacions ramificades, situades al límit entre derma i epidermis. Són substàncies químicament derivades de la tirosina, com alguns neuro-transmissors, i la seva producció és estimulada per l’acció d’algunes hormones específiques secretades per la hipòfisi, especialment l’hormona melanocito-estimulant (MSH).
El nombre de melanòcits és força constant d’uns individus a uns altres i també d’unes poblacions a unes altres (uns 1 500 per mm2), però és força variable d’unes parts del cos a unes altres dins d’un mateix individu, amb màxims a la cara, al front i al dors i mínims a la part ventral del tronc i a la cara palmar de mans i peus, i és lleugerament inferior en les dones que en els homes d’una mateixa població. De tota manera, els individus de poblacions de pell fosca poden tenir melanòcits al peritoneu i la seva retina és més fosca que la dels individus de pell clara. També hi ha variacions de la quantitat de melanines segons l’edat: els infants sempre neixen més despigmentats que els adults del seu mateix grup; en arribar la fase pre-puberal, hi ha un increment en la secreció d’hormona melano-estimulant. Les melanines són sintetitzades als melanòcits en forma de grànuls que són transferits a les cèl·lules de les capes profundes de l’epidermis, el nombre, la mida i la forma d’agrupament dels quals determina la coloració de la pell.
Altres pigments, com ara carotens o hemoglobines, col·laboren també en la coloració final de la pell dels individus, que es veu afectada també per factors com l’estat de salut (en casos d’anèmia l’aparença de la pell és força més pàl·lida, en casos d’icterícia la pell grogueja, i en casos de greus transtorns hepàtics o biliars, arriba a prendre un to brunenc) o en l’embaràs, quan sol haver-hi un increment en els dipòsits de melanina especialment en l’arèola del mugró i en la línia mitjana abdominal. També influeixen en la coloració de la pell la temperatura ambiental, les condicions de pH i la concentració de les vitamines A, B i C, i de metalls (coure i zinc) en l’organisme.
La qüestió de les mides
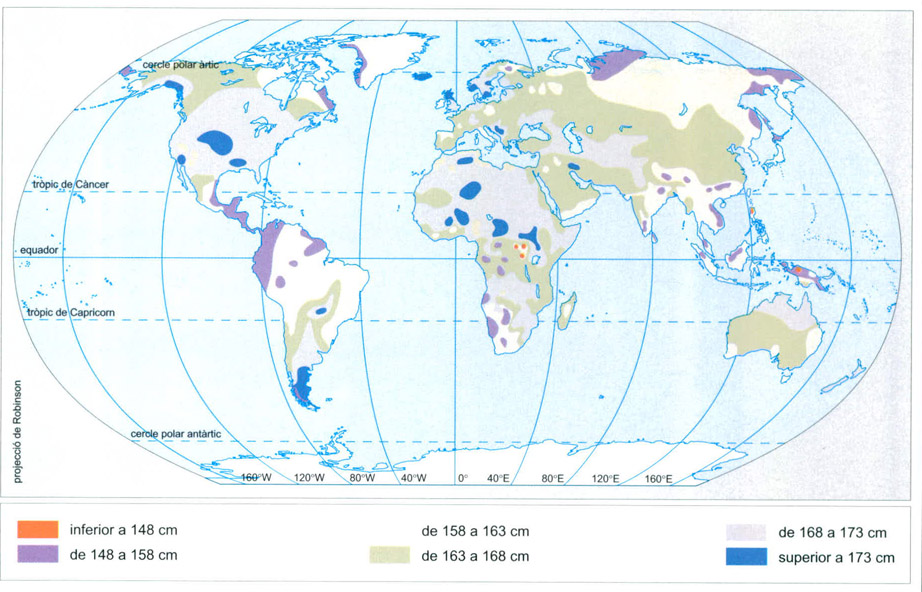
Distribució dels humans segons l’alçada mitjana abans de les grans migracions dels darrers segles. La relació morfològica entre les poblacions de menys alçada (taques vermelles) reflecteix un parentiu filogenètic: negritos de Filipines i poblacions de baixa alçada que habiten les muntanyes de Nova Guinea són parents dels pigmeus africans. Els grups de baixa alçada del SE asiàtic són descendents de la primera onada d’humans moderns que sortí d’Àfrica, i guarden parentiu proper amb els pigmeus de la selva africana i, més llunyà, amb els kung, o boiximans, del Kalahari.
Editrònica, a partir de dades proporcionades pels autors
Podem observar que els pobles que viuen a les selves tropicals, on el clima és molt humit, en general són sempre de mida petita; això succeeix tant entre els pigmeus africans com a l’Índia, al sud d’Indonèsia, Filipines o Nova Guinea; entre els maies d’Amèrica central o els habitants de la selva tropical brasilera. En un ambient on la humitat és del 100% (nivell de saturació), la sudoració, que és el principal mecanisme dels humans per a eliminar la calor excessiva, no és gaire eficaç o no ho és en absolut, ja que la suor no s’evapora fàcilment, sinó que roman líquida: no refreda l’organisme. Sense que hi hagi lligams genètics entre totes aquestes poblacions, comparteixen les seves peculiars dimensions corporals, el tipus d’hàbitat —la selva plujosa—, i les formes de vida associades a la cacera i la recol·lecció. Comparteixen també, probablement, el fet de ser els descendents de les més antigues poblacions d’humans moderns que sortiren d’Àfrica i arribaren a les diferents regions en què viuen, relegades ara a àrees marginals per successives expansions d’altres poblacions més poderoses .Els pigmeus i els altres humans diminuts tenen, a més, un altre avantatge: en ser minúsculs tenen menor massa muscular, especialment a les cames i els braços, el seu organisme produeix menys calor interna i, per tant, la despesa per a mantenir-se en un equilibri homeostàtic tèrmic és menor: l’esforç del pigmeu en moure’s és més petit que el d’un individu més gran que ell, ja que el pes a transportar és menor. No sabem si aquestes poblacions s’han tornat petites amb el temps o si ho han estat sempre. Sembla que el fenomen hauria pogut esdevenir per manca de resposta dels teixits a l’hormona de creixement (GH) després de la pubertat, quan hi ha l’estirada forta en altres poblacions humanes.
Alguns autors no estan d’acord amb l’explicació abans exposada per a la mida dels petits humans de la selva ja que, argumenten, no totes les poblacions d’humans diminuts viuen a la selva i, a més, els pigmeus de l’Àfrica equatorial foren desplaçats cap a la selva durant l’expansió dels bantus 3 000 anys enrere, massa poc temps per a canviar tant; abans explotaven l’ecotò selva-sabana. Aquestes paradoxes, diuen, semblen indicar l’existència d’una altra explicació evolutiva, no únicament la tèrmica, ni, per descomptat, la teoria que justificava la baixa estatura dels habitants de la selva per tal de moure’s amb més facilitat entre el brancatge. L’explicació evolutiva exposada diu que és la manca d’aliment disponible per als humans en les selves el que ha fet mantenir la pressió selectiva sobre els grups pigmoides (poblacions humanes de baixa estatura, però no clarament pigmees): els individus petits requereixen menys quantitat d’aliment per a mantenir-se; explicació que serviria també per als khoisan del desert del Kalahari, humans de mida petita i no habitants de zones selvàtiques.
La humitat de la selva
Les poblacions que viuen des d’antic a les selves —pot tornar-se a fer servir l’exemple dels pigmeus— tenen els nassos més amples del món. Això torna a ser una adaptació a les condicions climàtiques de la selva; l’aire calent i humit de la selva no ha de ser canviat de condicions en entrar en el cos en el moment de la inspiració, en tot cas, l’única cosa que pot succeir és que l’aire que entra a través de l’àmplia base d’implantació del nas per sobre un llavi superior que evapora suor, el refresqui.
L’índex nasal, una relació entre l’amplada i la longitud del nas d’una persona, és condicionat principalment per factors selectius imposats per la variació en el grau d’humitat de l’ambient. A l’Àfrica hi ha els índexs nasals més elevats —i també el major grau de variabilitat; recordem el cas dels beduins del desert—, especialment entre grups de pigmeus.
Els aborígens australians també posseeixen un dels índexs nasals més elevats del planeta, de la mateixa manera que a Amèrica, les poblacions de les selves tropicals són les de major índex nasal de tot el continent. Aquest fenomen de convergència morfològica per causa del clima il·lustra el fet que l’amplada del nas té tendència a ser superior en els llocs en què l’aire és més calent i més humit.
Altres diferències morfològiques
Els individus de coloració fosca, especialment els melanoafricans, tenen una pell més gruixuda, més vascularitzada i amb un nombre superior de glàndules sudorípares que els leucoderms. Les pells fosques resisteixen millor la insolació, l’agressió de paràsits i les temperatures altes i, en conseqüència, en els casos de congelació (per exemple, els esdevinguts entre els soldats dels Estats Units d’origen africà a Alaska) pateixen més dany que la pell dels leucoderms. L’engruiximent de la pell és conseqüència del desenvolupament del teixit conjuntiu dèrmic, fenomen que es manifesta en cicatritzar (les cicatrius solen ser hipertròfiques i queloides). Aquesta mena de cicatrització ha estat aprofitada culturalment per alguns grups africans per tal de tenir marques tribals identificatives: amb instruments punxants realitzen incisions de determinada forma geomètrica que, a l’hora de cicatritzar, quedaran evidents i serviran per a distingir uns grups dels altres.
Els cabells dels melanoafricans i d’altres poblacions de color de les selves plujoses són cresps i curts, amb el resultat que formen petites tofes arrapades al cap. En secció, són de forma oval aplanada o reniforme i han estat intensament pigmentats; fora dels albins, són sempre ben negres. Hom ho atribueix a una resposta de protecció del crani davant de possibles lesions d’origen actínic.
Salut i malaltia a les selves plujoses
Els problemes de salut de les poblacions humanes a l’àmbit de les selves plujoses equatorials són de naturalesa i origen molt diferents. No sols estan relacionats amb les condicions ambientals generals específiques d’aquest bioma, sinó més aviat amb les condicions socials i higièniques que caracteritzen les poblacions dels països del Tercer Món, en general. Tanmateix, l’extrema biodiversitat del bioma es manifesta també en un conjunt ampli i complex d’agents patogènics, de vegades molt localitzats, que afecten tant els humans com altres organismes. Així, doncs, existeix un cert nombre de malalties infeccioses o transmissibles que són en alguns casos exclusives i en altres molt preferentment implantades a l’àmbit de les selves plujoses.
Les principals infeccions i les vies de transmissió
Moltes de les malalties tropicals es transmeten per via digestiva, gairebé sempre a través d’aigües o aliments infectats. L’amebiasi (causada majorment pel protoctist Entamoeba histolytica), les anquilostomiasis (produïdes pels nematodes Necator americanus i Ancylostoma duodenale, també transmissibles a través de la pell), l’ascaridiasi (ocasionada pel nematode Ascaris lumbricoides), la gnatostomiasi (generada pel nematode Gnathostoma spinigerum), la giardiasi (produïda pel protoctist Giardia lamblia), etc., sense ser exclusives de l’àmbit de les selves plujoses, hi tenen una elevada incidència, com l’hi tenen també la febre tifoide, la disenteria bacil·lar i diferents hepatitis. Un cas ben particular és el de les malalties causades per trematodes ingerits en consumir cargols o peixos infectats, concretament les angiostrongiloidosis (provocades per Angiostrongylus cantonensis i A. costaricensis), pròximes a les esquistosomiasis de transmissió cutània. Hi ha també casos de còlera, sobretot al sud-est asiàtic, importantíssima malaltia tractada al volum 3 d’aquesta mateixa obra.
Perquè encara hi ha, en efecte, malalties que es transmeten a través de la pell o les mucoses. Potser les que tenen més incidència a l’àmbit de la selva plujosa són les esquistosomiasis o bilharziosis, que es contreuen en contacte amb aigües dolces on visquin cargols dels gèneres Bulinus, Biomphalaria i Oncomelania infectats amb els trematodes causants (Schistosoma haematobium, S. japonicum, etc.). També són molt importants algunes anquilostomiasis ja esmentades, l’estrongiloidosi (produïda pel trematode Strongyloides stercoralis), les malalties de transmissió sexual, algunes malalties víriques greus que cursen generalment amb febre hemorràgica, i les treponematosis no venèries, concretament el pian (ocasionat per l’espiroqueta Treponema pertenue) i la pinta (produïda per T. carateum).
Però les més importants d’aquestes malaties infeccioses es transmeten per la intervenció com a vectors d’alguns insectes o altres artròpodes, principalment diferents menes de mosquits. De molt, la més estesa d’aquestes malalties a l’àmbit de les selves plujoses és la malària o paludisme, però n’hi ha d’altres: les tripanosomiasis, la febre groga, el dengue i altres malalties víriques que cursen amb febre, i algunes filariosis en foren exemples. També les leishmaniosis (ocasionades per diversos protoctists del gènere Leishmania), una de les quals, el kala-azar, és tractat a l’àmbit dels boscos monsònics d’aquest mateix volum.
La malària o paludisme
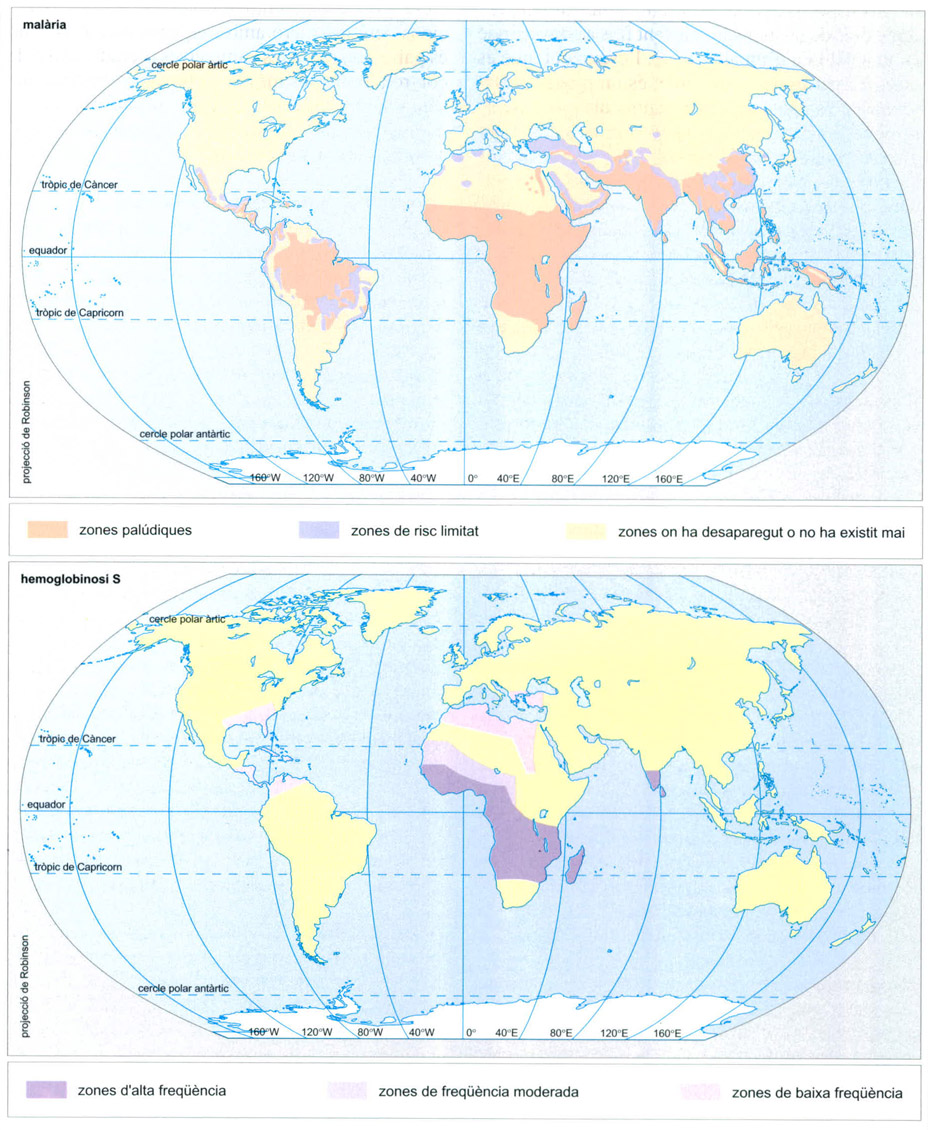
Distribució de la malària i l’hemoglobinosi S, dues malalties freqüents al tròpic. La primera és infecciosa, mentre que la segona és genètica. Des de mitjan Plistocè, durant el desenvolupament de les darreres glaciacions, els humans pateixen malària o paludisme, la malaltia causada per protoctists del gènere Plasmodium i transmesa per mosquits del gènere Anopheles. El mosquit pot reproduir-se a qualsevol zona humida (llacs, bassiots o un recipient abandonat) i passar el paràsit d’un individu malalt a un altre per les picades de les femelles. La malària, que pot ser mortal encara avui dia, sens dubte ha estat una causa de mortalitat elevada en el passat, especialment després del Neolític. Algunes mutacions en els gens de l’hemoglobina no permeten el desenvolupament normal del plasmodi en el eritròcits, amb la qual cosa els individus portadors d’aquests gens s’han vist seleccionats positivament a les àrees endèmiques de malària.
Editrònica, a partir de dades proporcionades pels autors
La malària és una malaltia parasitària que afecta els eritròcits de la sang i que es manifesta per característics accessos de calfreds, febre i suor, esplenomegàlia (creixement del volum de la melsa) i anèmia. Els paràsits que la causen són els plasmodis (Plasmodium), uns protoctists de l’embrancament dels apicomplexs (anomenats, fins a 1984, esporozous) amb cicles vitals complexos que inclouen una fase asexuada com a paràsits de vertebrats i una altra etapa, que inclou la seva fase sexuada i l’esporògena, com a paràsits d’artròpodes (principalment mosquits i paparres), els quals actuen com a vectors de transmissió de la malaltia. Prototipus actualment de malaltia tropical, el paludisme ha estat estès fins a dates ben recents per totes les zones humides de clima càlid o temperat. Era endèmic a tota la Mediterrània fins a mitjan segle XX i encara en resten alguns focus aïllats a l’orient de la conca i al Magrib. Encara al final dels anys quaranta en subsistien alguns focus al delta del Rin, als Països Baixos. Avui, però, deixant a banda focus aïllats en alguns països de clima temperat, el paludisme és una malaltia tropical que afecta primordialment les àrees de l’àmbit de les selves plujoses en les quals és una malaltia hiperendèmica (uns 100 milions de casos anuals).
El cicle malàric
En el cas dels plasmodis que causen la malària els dos hostes són els humans i els mosquits del gènere Anopheles. La fase asexuada que desenvolupen en els humans comença amb la picada d’un mosquit infectat (una femella d’Anopheles) i la inoculació d’una multitud d’esporozoïts (unes cèl·lules allargades amb unes peculiars estructures en un dels àpexs anomenades “complex apical”) del plasmodi. Els esporozous penetren en alguna cèl·lula del fetge dins de la qual van creixent fins a convertir-se en una cèl·lula d’un altre tipus, l’esquizont, que es divideix per escissió múltiple; és un procés característic dels apicomplexs i alguns altres grups de protoctists, pel qual l’esquizont dóna origen per gemmació a una colla de cèl·lules, semblants als esporozous, que s’anomenen merozoïts. Aquestes són les que, en el cas dels plasmodis de la malària, envaeixen els glòbuls rojos, a costa dels quals creixen fins a convertir-se també en esquizonts que poden repetir el procés descrit unes quantes vegades i alliberar a la sang de l’infectat successives generacions de merozoïts. Alguns dels merozoïts, però, no es desenvolupen com a esquizonts sinó que donen lloc a unes formes sexuades que romanen al si dels eritròcits humans. Si una femella d’Anopheles ingereix, en una picada, algun d’aquests eritròcits, pot tenir lloc, al seu estómac, la fase sexual del cicle vital del plasmodi que conduirà a la formació de nous esporozous.
Els plasmodis i les febres
Els accessos de calfred, febre i suor característics de la malària corresponen precisament a l’alliberament dels merozoïts per ruptura dels eritròcits afectats i es presenten amb un ritme característic de cada espècie de Plasmodium: cada 48 hores en el cas de P. ovale i de P. vivax (febres terçanes), cada 72 en el de P. malariae (febres quartanes) i amb intervals breus i indefinits però més sovint també de 48 hores en el de P. falciparum (febres terçanes malignes). La quinina ha estat el tractament antimalàric d’elecció durant segles, però cap vacuna no havia estat posada a punt fins que un equip de metges colombians reeixí a preparar-ne la primera l’any 1993, de caràcter sintètic, actualment (1994), en fase d’experimentació.
De les quatre espècies principals de Plasmodium que afecten els humans, la més estesa i més perillosa és P. falciparum, que és l’espècie predominant a Àfrica, al sud-est d’Àsia i en alguns països d’Amèrica, i és l’única que pot resultar mortal en ocasionar complicacions neurològiques, renals o càrdio-vasculars agudes i greus. Es considera que és l’espècie que ha evolucionat més tardanament com a paràsit humà per tal com tot i la seva virulència no és capaç de romandre latent en diferents teixits del malalt per rebrotar al cap d’uns anys com poden fer altres espècies (P. ovale i P. vivax al fetge i P. malariae als glòbuls rojos), de manera que, paradoxalment, en àrees intensament infestades per P. falciparum, sobretot a Àsia i a Amèrica, pot ser causa d’afeccions menys greus en adults supervivents d’infeccions infantils amb aquest paràsit que no P. vivax, de fet menys virulent. P. ovale, una espècie exclusivament africana i característica d’àrees de selva, sembla representar el millor ajustament hoste-paràsit entre els plasmodis; causa afeccions benignes però amb freqüents recidives, i no és gaire sensible a les campanyes d’eradicació o de control. També és més aviat benigne el paludisme causat per P. malariae. Altres plasmodis causen malalties en primats, ocells i altres animals, també amb mosquits com a vectors.
Les anomalies genètiques associades
La llarga convivència d’algunes poblacions humanes amb la malària ha estat ocasió per a la selecció d’algunes anomalies genètiques aparegudes espontàniament que donen lloc a la síntesi d’hemoglobines anormals. Tot i que per elles mateixes poden ser causa de malalties congènites greus en alguns individus, representen per a la població en conjunt una protecció enfront de la malària. Aquest és el cas de les anomalies que són causa de l’anèmia falciforme o de la talassèmia.
L’anèmia falciforme afecta principalment poblacions de l’Àfrica occidental i central, l’àrea mediterrània, l’est d’Anatòlia, la península Aràbiga i l’Índia, i consisteix en la presència en els eritròcits d’una hemoglobina, l’hemoglobina S, amb un canvi d’aminoàcid (valina en comptes d’àcid glutàmic) de la seqüència de la cadena β. Aquesta alteració aparentment insignificant és causa d’una característica deformació dels eritròcits que prenen una forma de falç i que, en cas d’esforç físic o de qualsevol altra circumstància que ocasioni requeriments elevats d’oxigen, les molècules d’hemoglobina precipitin, i els eritròcits falciformes esdevinguin allargassats, cosa que els pot dur a obstruir els capil·lars i a ser destruïts en la resposta immunitària de l’organisme, fins a ocasionar un quadre d’anèmia amb totes les seves seqüeles.
Aquesta alteració és el resultat d’una mutació recessiva en un gen precís d’un cromosoma autosòmic; és, doncs, de transmissió hereditària no lligada al sexe. Un de cada quatre descendents d’una parella de portadors d’aquest gen recessiu pot ser un homozigot, cas en què es manifesta la malaltia; dos dels quatre, heterozigots, potencials transmissors però amb una bona resistència enfront del paludisme; el quart, ni malalt ni portador del gen. Quan els heterozigots resulten infectats pel plasmodi, els eritròcits infectats esdevenen fàcilment falciformes, cosa que fa difícil l’intercanvi d’ions potassi, i el paràsit mor. Això explica que una anomalia genètica tan letal es conservi en determinades poblacions: representa una defensa enfront d’un paràsit encara més perillós per dos de cada quatre descendents d’una parella de portadors, amb la taxa d’un malalt. Aquesta pressió de selecció explica també la diferent presència d’aquest gen en les poblacions africanes (de prop d’un 40%) i les afroamericanes (entorn un 9%); les primeres es troben en contacte permanent amb el plasmodi, les segones no, i l’efecte letal de la mutació preval sobre la inútil protecció envers un paràsit absent, en el cas de les segones.
Una anomalia semblant, pel que fa a la resistència enfront de la malària, però que afecta un altre punt de la seqüència d’aminoàcids de la cadena ß i que ocasiona la presència a la sang de l’anomenada hemoglobina E, és freqüent en poblacions de la conca mediterrània, el sud de l’Índia, el sud-est d’Àsia i Nova Guinea. Respecte a la talassèmia, vegeu La talassèmia, ja que la conca mediterrània n’ha estat afectada.
La febre groga i el dengue
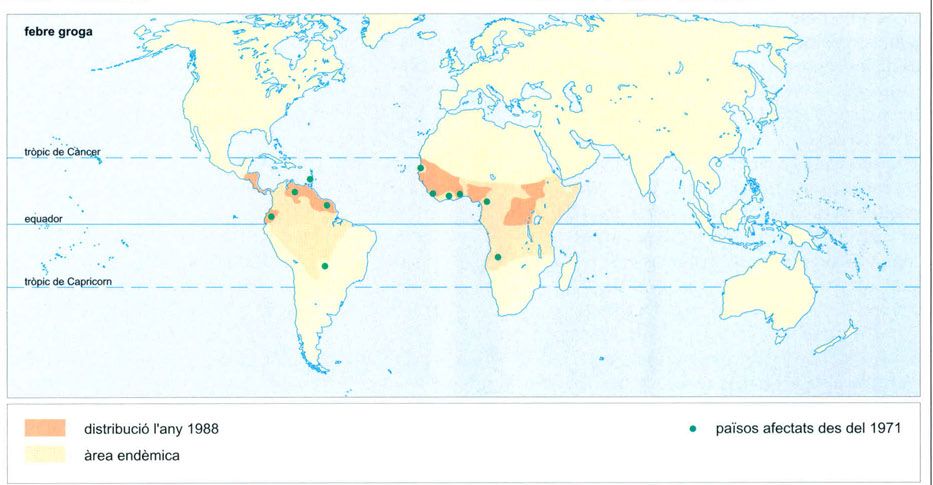
Distribució al món de la febre groga. El desenvolupament de les malalties infeccioses al tròpic té a veure amb molts factors, i no sols els ecològics. Els greus problemes socials, la pobresa i les guerres no permeten l’establiment efectiu de campanyes de conscienciació i vacunació als països on precisament hi ha més incidència.
Editrònica, a partir de dades proporcionades pels autors
La febre groga és una altra malaltia tropical transmesa per picada de mosquit. Es manifesta per l’aparició sobtada de febre, intens mal de cap i cruiximent general. La febre baixa al cap de tres o quatre dies per tornar a pujar a continuació acompanyada d’icterícia, conseqüència de necrosi hepàtica, i sovint d’un vòmit negre de sang i bilis. La mortalitat és elevada entre els adults que la contreuen, mentre que és més benigna en els nens, els quals, en les regions endèmiques, sovint tenen anticossos sense haver manifestat la malaltia i només per haver estat en contacte amb el patògen.
Originàriament, la febre groga és una malaltia pròpia dels simis (i, a Amèrica, sembla que també dels didèlfids) que esporàdicament pot infectar alguns humans com a conseqüència de picades dels vectors del virus, que són mosquits del gènere Haemagogus (H. spegazzinii, H. equinus) a les selves americanes, i del gènere Aedes (A. africanus, A. simpsoni) a les africanes. Tanmateix quan alguna persona portadora de febre groga arriba a un nucli de població on hi ha mosquits de l’espècie Aedes aegypti, molt vinculada a les aglomeracions humanes, la febre groga pot entrar en fase epidèmica. Tot i el seu nom, encara que sembla tenir un origen africà, A. aegypti és una espècie que viu a totes les regions tropicals i subtropicals del món, principalment a l’Àfrica i a Amèrica, però també a l’Índia, a Malaca i al nord d’Austràlia.
Les grans epidèmies de febre groga havien estat particularment devastadores a l’Amèrica tropical i havien arribat a afectar al segle XIX ciutats tan allunyades del tròpic com Filadèlfia i Boston. Si bé en la seva forma epidèmica pràcticament no té incidència des de fa molts anys, la seva forma selvàtica és endèmica a moltes regions de selva plujosa, principalment africanes, i no es poden descartar rebrots en poblacions humanes que convisquin amb poblacions d’A. aegypti. Això no obstant, d’ençà que l’any 1985 el mosquit asiàtic A. albopictus —que és vector del dengue, però també pot ser-ne de la febre groga— passà accidentalment del Japó als Estats Units, s’ha produït una recrudescència de la malaltia a ciutats intertropicals americanes. I també a l’Àfrica, afectada per una renovada activitat d’A. aegypti i per la recent presència d’A. albopictus, que també hi ha arribat: d’uns centenars de casos durant els anys setanta, s’ha passat a molts milers en els noranta.
El dengue és una altra malaltia vírica que és transmesa per diversos mosquits del gènere Aedes, principalment A. aegypti (i per A. albopictus, com acabem de veure). Es manifesta amb un quadre febril, dolors articulars i musculars generalitzats, erupció cutània a la cara, els braços i el pit al cinquè dia de la malaltia. En alguns casos, principalment en infants i molt particularment al sud-est d’Àsia i a Nova Guinea, pot ocasionar també febre hemorràgica.
Altres febres hemorràgiques d’origen víric de diversa gravetat, com l’anomenada febre “chicungunya” i diverses encefalitis víriques, també poden ser transmeses per picades de mosquits.
Les filariosis
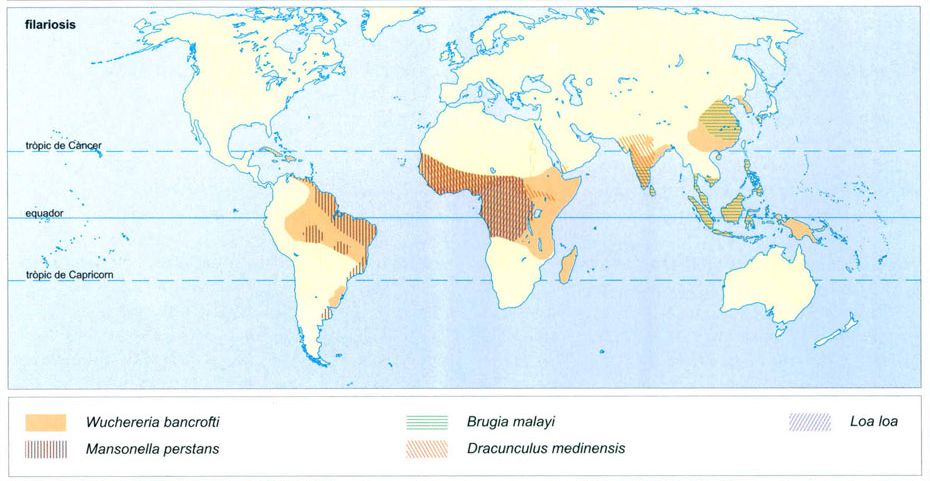
Distribució al món de les filariosis.
Editrònica, a partir de dades proporcionades pels autors
Però, a més de plasmodis i de virus, els mosquits poden també ser, a l’àmbit de les selves plujoses, vectors d’altres paràsits importants, com per exemple algunes filàries (Wuchereria bancrofti, Dracunculus medinensis, Brugia malayi, Loa loa, Onchocerca volvulus, Mansonella perstans, etc.). En el cas de W. bancrofti, els mosquits susceptibles de servir-li de vector són molt nombrosos (Aedes polinesiensis, A. fijiensis i Culex quinquefasciatus a les illes del Pacífic tropical més orientals, Anopheles punctulatus a Nova Guinea, els arxipèlags melanesis i els més occidentals de Polinèsia, A. gambiae, A. funestus i A. melas a Àfrica i A. darlingi a Amèrica del Sud). Pel que fa a Brugia malayi, la transmeten principalment mosquits del gènere Mansonia (M. annulifera, M. uniformis, M. longipalpis) i alguns del gènere Anopheles (A. barbirostris).
Les filàries són nematodes paràsits les larves dels quals, les anomenades microfilàries, són transmeses per artròpodes (mosquits, tàvecs i simúlids). Quan creixen i esdevenen adults poden envair els vasos i les glàndules limfàtiques i ocasionar-hi inflamacions, principalment a les extremitats i als òrgans sexuals masculins, que poden anar acompanyades de calfreds, febre, dolors i malestar general. Si la malaltia esdevé crònica, l’obstrucció dels vasos limfàtics deguda a l’amuntegament dels paràsits la pot fer evolucionar en una elefantiasi també localitzada principalment a les extremitats inferiors i, en els homes, a l’escrot.
Les tripanosomiasis
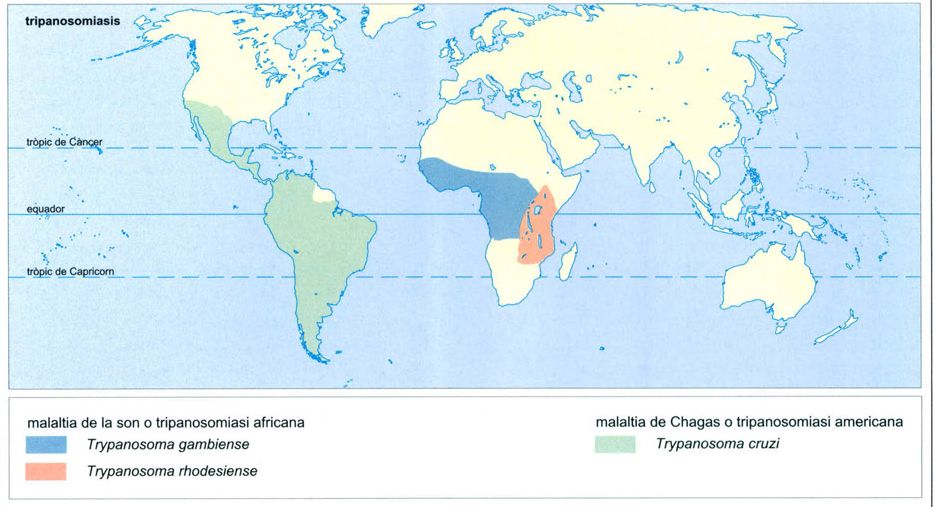
Distribució al món de les tripanosomiasis.
Editrònica, a partir de dades proporcionades pels autors
Les tripanosomiasis són malalties tropicals causades per protoctists del gènere Trypanosoma. Afecten sobretot el bestiar, però algunes provoquen malalties als humans, per bé que sempre hi ha un hoste intermediari, domèstic o silvestre, en el complicat cicle tripanosomiàsic.
La tripanosomiasi africana
A l’àrea intertropical africana té una incidència forta una forma de tripanosomiasi humana coneguda amb el nom de malaltia de la son. De fet, es tracta de dues malalties diferents, una pròpia de les àrees seques de sabana, i l’altra de les àrees humides més o menys compreses dins l’àmbit de la selva plujosa. En tots dos casos, el cicle comporta un hoste intermediari portador (ruminants silvestres i porcs domèstics, respectivament) i un dípter vector, la mosca tse-tse (Glossina morsitans i G. palpalis, també respectivament).
La malaltia de la son oriental o de la sabana, causada per Trypanosoma brucei rhodesiense, és tractada al volum 3. A propòsit de la selva plujosa, cal centrar-se en la malaltia de la son occidental, menys greu, que és l’ocasionada per T. brucei gambiense, transmesa per Glossina palpalis a partir del porc domèstic i del gos. Un cop presents a la sang del malalt, havent estat picat per una mosca infectada, els tripanosomes n’alteren el sistema limfàtic, provoquen febres i erupcions cutànies, i acaben afectant el sistema nerviós central, fet que es tradueix en un estat d’abúlia i somnolència que acaba essent fatal en molts casos, però gairebé mai no ho fa de manera immediata. En definitiva, es tracta d’una malaltia zoonòtica seriosa que, en les seves dues formes, presenta uns 200 focus de permanent presència, amb uns 50 milions de persones en constant risc d’infecció.
La tripanosomiasi americana
La tripanosomiasi americana afecta les àrees rurals de tot el continent de Mèxic cap al sud, però té una especial incidència a les zones de selva plujosa mesoamericanes i colombianes. Hi és coneguda amb el nom de malaltia de Chagas, en honor del metge brasiler Carlos Justiniano Chagas (1879-1934), que l’any 1909 n’establí l’etiologia exacta.
La malaltia de Chagas és ocasionada per Trypanosoma [=Schizotrypanum] cruzi i transmesa als humans i a altres mamífers per l’heteròpter triatòmid conegut a quasi tota l’Amèrica llatina amb el nom de “vinchuca” (Triatoma infestans). De fet, també són vectors del protoctist altres triatòmids (T. dimidiata, Rhodnius prolixus, Panstrongylus megistus), però la seva incidència és menor, sobretot perquè a penes tenen hàbits domèstics. I és que la singularitat de la “vinchuca” és que s’ha adaptat a viure entre els humans, amb una especial preferència per les cabanes i barraques amb sostres de fibres vegetals, on nidifica. En tot cas, la “vinchuca” pica indistintament qualsevol mamífer, tots ells susceptibles de contreure la malaltia, i també ocells, rèptils i amfibis, que n’esdevenen portadors sense caure’n malalts. Els humans contreuen aquesta tripanosomiasi quan són picats per “vinchucas” infectades, quan reben transfusions sanguínies de persones igualment infectades o per via placentària. Això darrer és ben possible, ja que la permanència dels tripanosomes a la sang dels afectats pot ser molt llarga.
En efecte, si hom ha estat infectat, després d’una incubació d’1 a 3 setmanes, el torrent sanguini s’omple de paràsits i es manifesta la malaltia en la seva forma aguda (febre, malestar general), la qual, sobretot en els infants, cursa amb una morbiditat prou alta (fins d’un 13%). Si sobreviu, l’afectat esdevé un malalt crònic asimptomàtic durant 10-20 anys, període en què, per picada, pot anar infectant ell mateix noves “vinchucas”, i alhora desenvolupa una miocarditis, una meningo-encefalitis, una oftàlmia o diverses visceromegàlies: esdevé un malalt crònic simptomàtic de pronòstic greu. De fet, la majoria de les cardiopaties en l’àmbit de la selva plujosa americana són ocasionades per aquesta tripanosomiasi.
La síndrome d’immunodeficiència adquirida (SIDA)
Una malaltia de procedència incerta —com 500 anys enrere ho fou la sífilis, l’origen geogràfic de la qual és discutit encara avui— s’ha escampat en forma de pandèmia en aquest final de mil·lenni i ha posat terme a un llarg període durant el qual la humanitat havia perdut pràcticament la memòria de grans pandèmies a escala mundial. Es tracta de la síndrome d’immunodeficiència adquirida (SIDA), formalment descrita per primera vegada el 1981, bé que les primeres manifestacions de l’epidèmia, encara no identificada com a tal, sembla que es produïren a Califòrnia i a Nova York a mitjan dècada dels setanta.
Tot i que aquestes manifestacions primerenques de l’actual pandèmia de SIDA apareguessin a Amèrica del Nord, hi ha la quasi certesa de l’origen africà i selvàtic de la malaltia, que és transmesa pel retrovirus l’HIV-1 (Human Inmunodeficiency Virus, 1), semblant a d’altres del mateix grup aïllats en simis, principalment el STLV-III (Simian T Lymphotrophic Virus, III) identificat en poblacions salvatges de cercopitec verd (Cercopithecus sabaeus). S’han trobat anticossos de la SIDA a la sang d’un donador zairès extreta el 1959, i sembla que la malaltia ja era present en petites poblacions aïllades de les selves d’Àfrica molt abans. La màxima incidència de la SIDA, de fet, es dóna a l’Àfrica sud-sahariana, on s’estima que el nombre total d’infectats (portadors o malalts) ultrapassa els 8 milions, contra el mig milió d’Europa, més d’un milió a Amèrica del Nord i uns 3,5 milions a la resta del món. Bé que no tots els infectats arriben a desenvolupar la malaltia, la taxa de mortalitat entre ells és molt superior a la de la resta de la població. Molts epidemiòlegs opinen que les possibilitats de contagi romangueren limitades mentre el contacte amb l’exterior de les poblacions inicialment afectades fou mínim, però que quan els contactes es generalitzaren esdevingué possible la formació d’una reserva infectiva suficient per desfermar la pandèmia actual. Probablement iniciada a l’Àfrica, no fou reconeguda fins que es manifestà als Estats Units i d’allí passà a Europa, i a la resta del món. L’epidèmia s’expandí entre drogoaddictes intravenosos i homosexuals barons europeus i nord-americans, però actualment hi ha una tendència inquietant de creixement entre la població heterosexual.
Les manifestacions patològiques de la infecció per l’HIV són molt diverses. Molts infectats poden no manifestar cap mena d’afecció durant molts anys, fins i tot deu o més. En altres casos comencen per manifestar símptomes inespecífics, com febre, erupcions cutànies i malestar general des del moment que comencen a produir anticossos. Però el que caracteritza la malaltia és l’alteració del sistema immunològic del malalt —sobretot conseqüència de la destrucció d’un tipus particular de limfòcits, els T4 cooperadors-inductors—, el qual esdevé molt vulnerable a agents patogènics oportunistes que rarament afecten les persones amb un sistema immunitari normal (el rizòpode Pneumocystis carinii, coccidis dels gèneres Cryptosporidium i Toxoplasma, fongs com Candida albicans, herpesvirus del grup dels citomegalovirus, etc.). També el fa susceptible a algunes afeccions tumorals poc freqüents, com el sarcoma de Kaposi o el limfoma de Burkitt, particularment freqüents, sobretot el segon, entre els malalts de la SIDA africans.
En poc temps, la SIDA ha passat de ser una afecció desconeguda a ser, per una banda, la malaltia que suscita més temors i controvèrsies i, per una altra, la més estudiada de totes. En ser transmesa a través de les mucoses i dels líquids orgànics, el risc de contagi se centra en les transfusions de sang contaminada i en tot contacte amb intercanvi de fluids corporals. Això darrer la converteix, en part, en una malaltia de transmissió sexual. En tot cas, abans del 1981 la SIDA no “existia” per a la medicina. Al cap de dos anys almenys dos laboratoris, un a França i un altre als Estats Units ja havien aïllat l’HIV-1, i el 1985 hom aïllà a l’Àfrica occidental l’HIV-2, un virus relacionat amb la mateixa patogènia. Prop de deu anys més tard, la SIDA ha esdevingut una amenaça a la salut coneguda arreu del món, objecte de campanyes de prevenció, d’esforços de recerca i de debat social a una bona part del món, sobretot als Estats Units, a Europa i Àfrica. En alguns països africans, la infecció afecta del 20 al 40% de la població sexualment activa d’algunes ciutats, i en el medi rural també ha tingut una extraordinària expansió. A ciutats com Abidjan, on abans del 1983 no se n’havia enregistrat ni un sol cas, la SIDA fou la principal causa de mort de la població adulta, sense distinció de sexes, l’any 1993. Els factors socials i econòmics ajuden a l’acceleració del procés de propagació: els moviments de població provocats per les guerres i la recerca de feina distant de la llar faciliten el contacte freqüent amb prostitutes. El problema deixa de ser exclusivament mèdic, ja que, en tornar a casa temporalment poden infectar les dones, que al seu torn poden passar-ho als infants; o bé retornen a casa malalts per a morir, amb la qual cosa augmenten el nombre de vídues i orfes de manera alarmant.