Característiques del grup
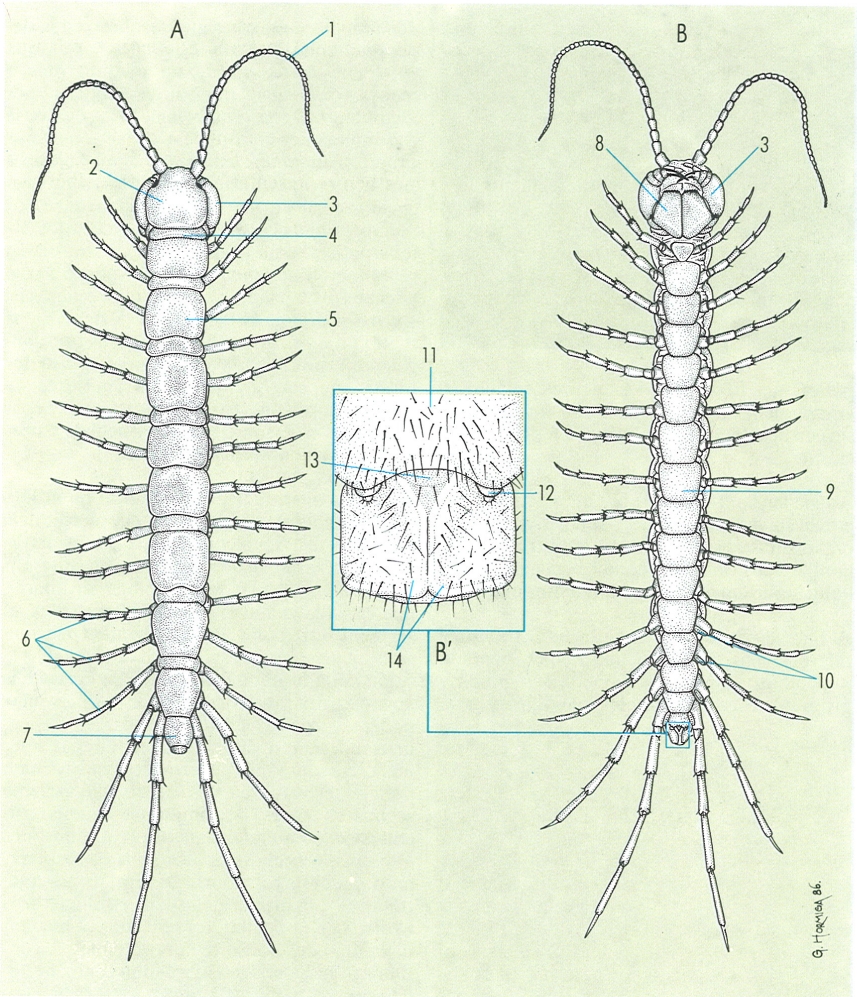
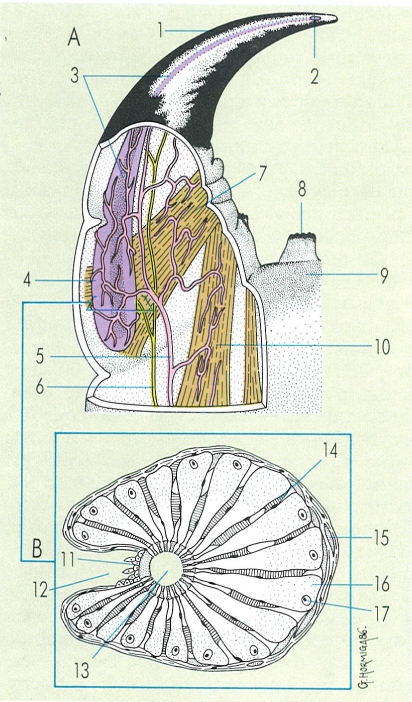
Aspecte general extern dels quilòpodes, en visió dorsal (A) i ventral (B), i detall de la regió posterior d’un mascle (B’). Hom hi distingeix: 1 antena, 2 càpsula cefàlica. 3 forcípula, 4 tergita del segment forcipular, 5 tergita del tercer segment del tronc, 6 pota, 7 tèlson, 8 coxostèrnum forcipular, 9 esternita del vuitè segment del tronc, 10 pors coxals, 11 esternita del primer segment genital, 12 gonopodi, 13 penis, 14 valva anal.
Gustavo Hormiga, original dels autors.
Els quilòpodes, que a casa nostra reben els noms populars de centcames, centpeus i escorçanes, són miriàpodes lucífugs i higròfils que normalment viuen sota les pedres. En el seu cos hom pot distingir el cap amb els òrgans bucals, el segment forcipular amb els seus apèndixs (la tranformació d’aquests apèndixs de potes locomotores en forcípules que intervenen en la masticació motiva el nom d’aquests artròpodes, ja que la paraula quilòpode prové dels mots grecs keilos, llavi, i podos, peu), el tronc constituït per un nombre variable de segments, cada un dels quals té només un parell de potes, i finalment un grup de segments terminals amb els seus apèndixs modificats, reduïts, i àdhuc completament atrofiats.
Morfologia
Malgrat que tots els quilòpodes presenten una estructura bàsica anàloga, hi ha un grup relativament petit que presenta un seguit de notables diferenciacions. Per això, hom distingeix dos tipus morfològics distints, dels quals exposarem a continuació les seves particularitats per separat.
Tipus morfològic dels geofilomorfs, els escolopendromorfs i els litobiomorfs
La gran majoria dels quilòpodes corresponen al primer tipus morfològic, ja que aquest és característic dels geofilomorfs, els escolopendromorfs i els litobiomorfs. El cos, format per un tegument tou sense impregnacions de sals minerals, és aplanat, allargat i estret en forma de cinta.
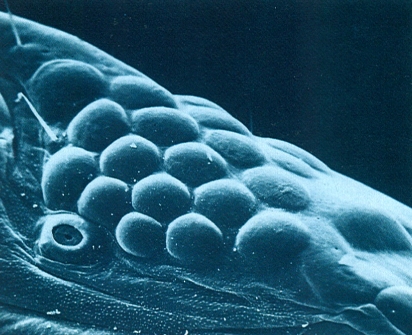
Camp ocel·lar del cap d’un litobiomorf (Lithobius validus) vist al microscopi electrònic d’escandallatge (× 130), al qual hom pot distingir l’ocel·le principal, a la dreta, i tota la resta d’ocel·les distribuïts en files més o menys regulars, i a sota d’aquests, l’òrgan de Tömöswary.
Antoni Serra
El cap, de forma lenticular o rectangular, és constituït per la fusió de l’àcron amb sis metàmers, corresponents als segments preantenal, antenal, postantenal, mandibular i els dos segments maxil·lars. Dorsalment, la càpsula cefàlica és bombada i formada per una peça única, encara que a alguns grups hi ha un solc frontal transvers situat al terç anterior. A les zones lateroanteriors del cap hi ha un parell d’antenes multiarticulades, que s’aprimen progressivament cap a la part distal. A la banda de darrere de l’articulació de l’artell basal de les antenes es localitza l’òrgan de Tömösvary, anomenat també òrgan temporal, petita fossa rodona o ovalada amb la vora fortament quitinitzada. A continuació d’aquests òrgans hi ha els camps ocel·lars o grups d’ocel·les independents. Els ocel·les falten completament en els geofilomorfs, en alguns grups d’escolopendromorfs i en espècies cavernícoles de litobiomorfs.
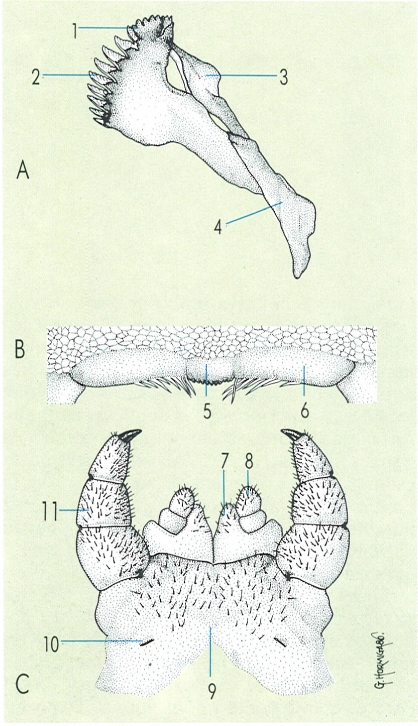
Peces bucals dels quilòpodes: mandíbula (A), labre (B) i segon parell de maxil·les (C). Hom hi distingeix les estructures següents: 1 làmina dentada, 2 làmina pectinada, 3 còndil articular, 4 tronc mandibular, 5 peça mitjana, 6 peça lateral, 7 prolongació coxal, 8 telopodi biarticular, 9 sincoxostèrnum, 10 obertura de la glàndula salival, 11 telopodi triarticulat.
Gustavo Hormiga, a partir d’originals diversos.
A la cara ventral, la càpsula cefàlica presenta una zona prelabial més o menys ben delimitada, amb una curvatura còncava a la part posterior, on se situa el labre. Aquesta estructura és constituïda per una peça mitjana imparella i dues peces laterals simètriques; normalment la peça mitjana és més petita que les laterals i pot presentar denticles curts, en forma de tubercle, o llargs i prims semblants a sedes filamentoses. A vegades les tres peces estan soldades formant-ne una de sola, la qual presenta una escotadura ampla a la part mitjana de la vora posterior. A banda i banda del labre hi ha una placa, les pleures cefàliques, que se situa a continuació de la vora lateral del cap. Damunt les pleures cefàliques es recolzen els braços externs dels fulcres, poderosos apodemes fortament quitinitzats, els quals, a més, tenen un braç anterior, que se situa sota el labre, i un braç posterior dirigit enrere, que delimita i dona suport al con hipofaríngic. El labre i les pleures cefàliques emmarquen latero-anteriorment una depressió, on se situen la hipofaringe i les peces bucals.
Aplicades lateralment sobre el con hipofaríngic, hi ha les mandíbules. Aquests poderosos òrgans mastegadors consten d’un tronc mandibular llarg amb un còndil articular i perllongat per darrere per una tija arquejada.
A l’aresta apical del tronc hi ha estructures diferenciades, tals com una làmina dentada molt dura, làmines pectinades, papil·les i grups de sedes. Damunt les mandíbules hi ha dos parells de maxil·les que recobreixen la depressió ventral del cap. El primer parell, que és el més petit, forma un conjunt més o menys triangular dorsalment còncau per adaptar-se al con hipofaríngic. La seva base és formada per la fusió de les coxes que constitueixen un sincoxostèrnum, ample i trapezoidal, del qual neixen dues petites prolongacions coxals orientades cap endavant i dos membres biarticulats, encara que els dos artells poden ésser fusionats en un de sol. El segon parell de maxil·les queda sobreposat al primer i és bastant més gros que aquest. El sincoxostèrnum és de forma i dimensions molt variables, gairebé sempre sense prolongacions i, prop de la vora posterior, porta dos forats que corresponen a les obertures de glàndules salivals. Els membres, molt separats entre ells, són llargs, triarticulats i normalment tenen una ungla apical.

Detall del cap d’un escutigeromorf (Scutigera coleoptrata), mostrant la forma gairebé cúbica que és característica d’aquest tipus morfològic. Noteu l’espectacularitat dels ulls, constituïts per un nombre elevat de petits ocel·les, i la forma de la zona prelabial, que delimita la regió anterior de la càpsula cefàlica.
Antoni Serra.
Immediatament darrere del cap hi ha el segment forcipular, notablement diferenciat dels segments del tronc. La seva tergita és de dimensions variables i és perfectament diferenciada (llevat de la dels escolopendromorfs, que és soldada a la següent). Ventralment, el segment forcipular és representat per una gran placa trapezoïdal, el coxostèrnum, resultat de la fusió de l’esternita i les coxes dels apèndixs. La seva vora anterior pot presentar poderoses dents, espines i sedes, o bé ser inerme, i sempre és dividida en dues parts per una escotadura més o menys profunda. En un punt de les vores laterals, de situació variable segons els grups, destaquen els còndils articulars coxofemorals, zones molt quitinitzades que engalzen els telopodis amb el coxostèrnum. Aquests telopodis són formats per quatre artells, el primer dels quals és el més gros i robust, i es pot interpretar com el resultat de la unió del trocànter i el prefèmur. A continuació hi ha dos anells que internament poden ser incomplets. El darrer artell és una poderosa ungla, fortament quitinitzada, llarga, cònica, arquejada, que gradualment es fa més fina i acaba en punxa. Per transparència s’observa un canal que prové d’una glàndula de verí i que s’obre a l’exterior una mica abans de la punta.
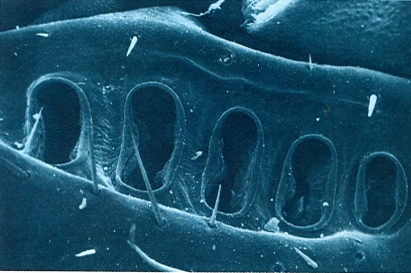
Porus coxals d’un litobiomorf (Lithobius validus), disposats en una fila, vistos al microscopi electrònic d’escandallatge (× 130). Aquests porus se situen en una depressió de la cara ventral de les coxes dels quatre darrers parells de potes del cos.
Antoni Serra.
Darrere el segment forcipular comencen els segments del tronc, cada un dels quals consta d’una tergita i una esternita unides mitjançant pleurites formades per una membrana tova que inclou plaques quitinitzades, l’estigma respiratori i un parell de potes. Les tergites, de forma gairebé sempre rectangular, poden ser totes semblants o bé alternar-se de grans i de petites, i en els geofilomorfs són formades per dues plaques, una d’anterior, la pretergita i una de posterior, la metatergita. Les esternites, de perfil pentagonal o rectangular, estan dividides també en una pre-esternita i una metaesternita. A la seva superfície hi ha estructures com ara solcs i fossetes, i en els geofílids hi ha també pors glandulars aïllats o formant agrupacions.
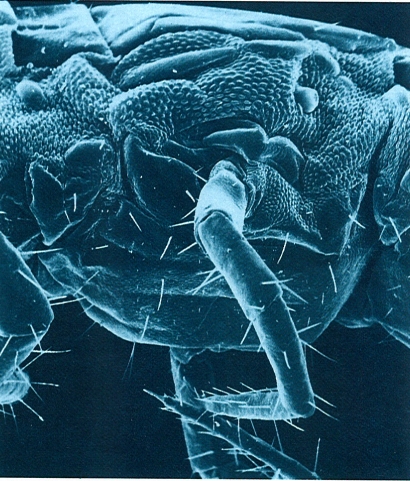
Segment del tronc d’un escolopendromorf (Cryptops trisulcatus), vist al microscopi electrònic d’escandallatge (× 50), en una visió lateroventral. Les zones amb petits tubercles corresponen a les esclerites pleurals, enmig de les guals se situa l’estigma respiratori, que té forma arrodonida. La regió. ventral, entre les potes, correspon a l’esternita.
Antoni Serra.
El conjunt d’esclerites que hi ha a les membranes laterals rep el nom d’eupleri. Aquestes plaques són molt variables d’uns grups a altres, i són els geofilomorfs els qui les presenten més ben caracteritzades. De totes, són quatre les que es troben d’una manera més constant a l’eupleri. La primera és l’esclerita estigmatífera, és a dir, on desemboquen les tràquees respiratòries. Les altres tres estan relacionades amb l’articulació de la coxa de les potes amb la paret lateral del cos i són la precoxa, la metacoxa i la catopleura. Entre totes tres delimiten un triangle anterodorsiposterior on s’allotja la coxa de la pota. Les potes, en nombre d’un parell per segment, s’articulen lateralment a l’eupleri i consten d’una coxa i un telopodi format per cinc o sis artells i acabat en una ungla. Les coxes d’algunes potes, principalment les dels darrers parells, poden tenir glàndules, els pors de les quals, anomenats pors coxals, tenen importància sistemàtica pel seu nombre i disposició. El telopodi consta d’un trocànter, de petites dimensions i que fins i tot pot ésser atrofiat, un prefèmur, un fèmur, una tíbia, un tars i un metatars; a vegades, el tars i el metatars estan fusionats i constitueixen un artell únic. El metatars va proveït normalment d’una espina apical, acompanyada o no d’una o dues espines suplementàries. Tots els artells de les potes poden presentar espines seguint disposicions característiques per a les diferents espècies. El darrer parell de potes és sempre més llarg i robust que els altres; en alguns grups aquestes potes presenten estructures especials, com ara serres dentades a la tíbia i al tars, apòfisis coxals i, encara, en algunes espècies, s’hi troben caràcters sexuals secundaris, tals com tubercles, grups de sedes, depressions, solcs o simplement el darrer parell de potes molt més gruixut en els mascles que a les femelles, de manera que la seva morfologia permet distingir-los.
Després del darrer segment proveït de potes, hi ha encara dos segments genitals, poc aparents dorsalment, i un segment terminal. El primer segment genital té una esternita molt gran, de la qual surten un parell d’apèndixs, els gonopodis, rudimentaris o pluriarticulats. Les femelles dels litobiomorfs són les que presenten uns gonopodis més evidents, cadascun fet d’un artell basal, ample i amb esperons a la vora posterior interna, i un artell distal més petit, seguit d’una ungla en forma de cullera. En el segon segment genital, de dimensions més reduïdes que les del primer, s’obre l’orifici genital, que en els mascles és envoltat per un penis; excepció feta dels escutigeromorfs, aquest segment genital no té apèndixs. El cos dels quilòpodes s’acaba amb un segment bastant particular, el qual s’homologa a un tèlson. La placa tergal pot ser gran o més o menys reduïda i recobreix les valves anals, peces lateroventrals entre les quals s’obre l’anus.
Tipus morfològic dels dels escutigeromorfs
El segon tipus morfològic correspon només a l’ordre dels escutigeromorfs, en els quals el cos no és pas aplanat i llarg, sinó bombat i curt. El cap és gros, gairebé cúbic, lleugerament deprimit a la part mitjana de la regió dorsal. La zona prelabial no s’amaga sota el cap, sinó que es dirigeix perpendicularment cap avall. Les antenes són filiformes, desmesuradament llargues i estan formades per una gran quantitat d’artells petits. Els ulls són globosos, grossos, formats per un gran nombre de petits ocel·les juxtaposats i no n’hi ha mai d’aïllats. Les peces del labre estan fusionades i formen una peça única. Les mandíbules són més curtes que les del primer tipus, però presenten pràcticament les mateixes estructures. El primer parell de maxil·les té un sincoxostèrnum gros, dividit longitudinalment per un solc fi i superficial. A banda i banda d’aquest solc hi ha dos òrgans sensorials, constituïts per voluminosos fascicles de tricomes fusiformes. Els telopodis són biarticulats; l’artell proximal és llarg i truncat obliquament, mentre que l’artell apical és molt més curt. El sincoxostèrnum del segon parell de maxil·les és curt i ample i porta dos telopodis molt llargs i gràcils, formats per quatre artells amb espines llargues, el darrer dels quals no porta ungla apical.
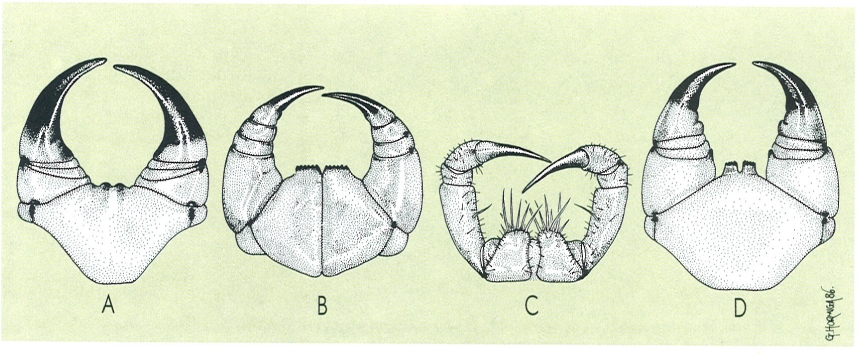
Diferents models de forcípules i coxostèrnums (segments als quals s’insereixen les forcípules), corresponents a: A geofilomorf, B litobiomorf, C escutigeromorf, D escolopendromorf.
Gustavo Hormiga, a partir d’originals diversos.
El segment forcipular té una tergita molt ample i curta que pot quedar dissimulada sota la vora posterior del cap. Les coxites de les forcípules, a diferència del que passava a l’altre tipus morfològic, són dissociades, encara que es toquen per la línia mitjana i posteriorment s’uneixen per un petit pont esternal. A la vora rostral de cada coxita hi ha una filera marginal d’espines molt llargues. En els escutigeromorfs, els telopodis són llargs i gràcils i estan formats per quatre artells, el darrer dels quals té una ungla menys robusta que la dels altres ordres de quilòpodes.
El tronc és format per quinze segments proveïts de potes, cada un dels quals té la seva corresponent tergita. En canvi, de tergites visibles només n’hi ha set, situades entre la tergita forcipular i les dues posteriors; les vuit tergites restants són molt reduïdes i estan amagades sota les grans. Aquestes plaques tergals tenen a la meitat de la seva vora posterior un estigma respiratori imparell, resultat de l’emigració i la fusió dels estigmes de l’eupleri de cada segment. Les potes són extraordinàriament llargues i gràcils, sobretot el darrer parell, pel fet que el tars i el metatars són formats per un gran nombre de petits artells.
Pel que fa als segments terminals, no hi ha cap diferència significativa respecte al primer tipus morfològic, llevat de la presència d’un parell de petits membres que es troben al segon segment genital.
Organització interna
Els sistemes de relació i control
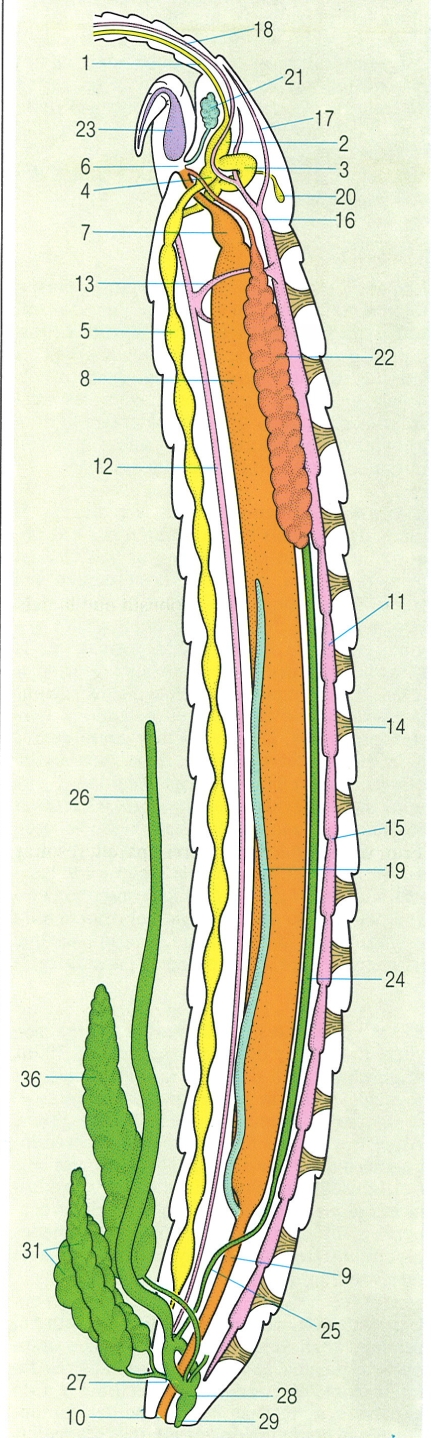
Anatomia d’un quilòpode, en un tall longitudinal del cos. Hom hi distingeix: 1 nervi antenal, 2 deutocervell, 3 protocervell, 4 tritocervell, 5 cadena ganglionar, 6 boca, 7 esòfag, 8 intestí mitjà, 9 intestí posterior, 10 anus, 11 vas dorsal, 12 vas ventral, 13 anell aòrtic, 14 brides musculars, 15 ostíol, 16 aorta cefàlica, 17 artèria cefàlica, 18 artèria antenal, 19 tub de Malpighi, 20 glàndula cefàlica, 21 glàndula maxil·lar, 22 glàndula salival, 23 glàndula del verí, 24 testicle, 25 canal deferent, 26 vesícula seminal, 27 canal ejaculador, 28 atri genital, 29 penis, 30 glàndula accessòria dorsal, 31 glàndules accessòries ventrals.
Gustavo Hormiga, a partir d’originals diversos.
El cervell dels quilòpodes presenta un deutocervell format per dos lòbuls, dels quals neixen dos nervis antenals. Posteriorment es localitzen dos lòbuls tritocerebrals que es perllonguen cap al darrere i formen la cadena nerviosa ventral. El protocervell forma una comissura que uneix els lòbuls tritocerebrals i està situat damunt de l’esòfag. Anteriorment, s’hi distingeixen dos lòbuls frontals; lateralment, dos lòbuls òptics que innerven els ocel·les, i lateroposteriorment, els nervis de les dues glàndules cerebrals. La cadena nerviosa ventral presenta dos ganglis per segment, ben individualitzats i units per connectius longitudinals. Si bé s’han detectat reaccions dermòptiques a algunes espècies anoftalmes, els principals òrgans fotoceptors són els ocel·les.
Els ocel·les, en els litobiomorfs consten d’una còrnia que té forma de lent biconvexa i és segregada per cèl·lules epidèrmiques diferenciades, les cèl·lules corneògenes. A sota la còrnia, a part de les cèl·lules corneògenes, hi ha dos tipus cel·lulars diferents; a la regió més externa se situen les cèl·lules cobertores, que per la seva estructura i situació hom pensa que intervenen en la secreció de la part endocuticular de la còrnia i que serveixen per a diferenciar morfològicament i mecànica un ocel·le dels altres; l’altre tipus cel·lular correspon a les cèl·lules retinals, de les quals n’hi ha de vuit a deu de laterals i de sis a vuit de basals. Les cèl·lules retinals constitueixen la cúpula ocular; són allargades i tenen dues regions diferenciades: una regió interna fotoceptora, amb microvil·lositats que constitueixen els rabdòmers, amb la zona central ocupada pel citoplasma, on es distingeixen vesícules de reticle endoplàsmic, sàculs gòlgics, cossos multivesiculars, ribosomes i mitocondris; i una part externa, que correspon al pol basal i que porta el nucli i una gran quantitat de grànuls pigmentaris. El límit entre les regions externa i interna de les cèl·lules retinals és configurat per l’existència de grans vacúols i rep el nom de zona intercalar. A sota de les cèl·lules retinals basals hi ha una gran quantitat d’àxons, que, juntament amb els que hi ha entre les cèl·lules laterals, formen els nervis òptics. Els nervis òptics són innervats pel protocervell.
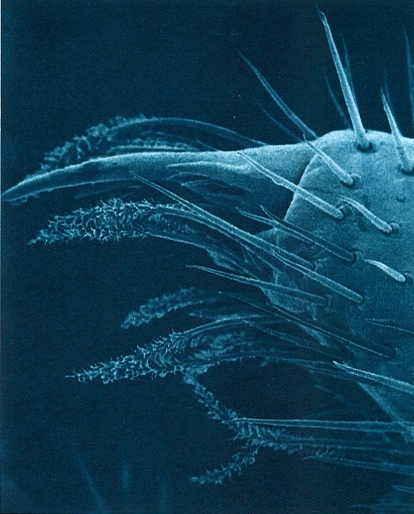
Extrem apical del telopodi del segon parell de maxil·les d’un litobiomorf (Lithobius variegatus rubriceps) vist al microscopi electrònic d’escandallatge (× 250). A més de l’ungla apical hom hi pot distingir dues menes de pèls sensitius: els uns en forma de plomall, possiblement de funció quimioceptora, i els altres llisos, mecanoceptors.
Antoni Serra.
El sentit del tacte és molt desenvolupat en els quilòpodes. Les sensacions tàctils són captades per mecanoceptors, sedes sensorials repartides per tot el cos, potes i antenes. Aquestes sedes presenten una inserció en el tegument, caracteritzada per una peça en forma de mitja lluna que les orienta en un sentit determinat. Les sensacions gustatives són percebudes per quimioceptors situats als palps de les maxil·les i a l’àpex de les antenes. Els escutigeromorfs, a les antenes, tenen uns òrgans especials que semblen actuar com a receptors audititus.
L’aparell digestiu
La boca és representada per l’obertura de la hipofaringe, delimitada anteriorment pel labre i lateralment per les mandíbules. A continuació hi ha l’esòfag, tub curt amb plecs longitudinals i sinuositats internes que actua com una mena de filtre pels aliments. Al seu extrem posterior hi ha una vàlvula associada a un esfínter muscular que determina l’inici de l’intestí mitjà. Aquesta porció del tub digestiu és la més llarga i té una secció transversal notablement més ample que la resta. És al seu nivell que té lloc la digestió i l’absorció. Finalment hi ha l’intestí posterior, que és curt, estret, en forma d’essa, i s’obre a l’exterior a través de l’orifici anal situat al darrer segment del cos.
L’aparell respiratori
Els quilòpodes respiren mitjançant un sistema traqueal repartit més o menys regularment per tot el cos. Les tràquees tenen forma de tub i es mantenen obertes gràcies a uns engruiximents quitinosos de les parets, que els dona un aspecte anellat. Desemboquen en cambres que s’obren a l’exterior mitjançant els estigmes, els quals se situen a les pleures de determinats segments del tronc. Al gènere Lithobius, els segments estigmatífers són els situats en posició 3, 5, 8, 10, 12 i 14 i en altres litobiomorfs també hi ha estigmes en el primer segment del tronc. A la majoria dels escolopendromorfs els estigmes se situen als metàmers 3, 5, 8, 10, 12, 14, 16, 18 i 20. Tots els geofilomorfs presenten estigmes respiratoris a tots els segments del tronc i és característic d’aquest ordre el fet que les tràquees s’anastomosen i formen esquames.
L’aparell respiratori dels escutigeromorfs és completament diferent al de la resta de quilòpodes. Consta de cambres dividides en dos lòbuls i formades per la reunió de gran nombre de tràquees fines banyades per l’hemolimfa, els moviments intermitents de la qual provoca febles inspiracions i expiracions. Aquestes cambres comuniquen amb l’exterior mitjançant estomes imparells en forma de trau que se situen a la part mitjana de la vora posterior de les tergites grosses del tronc. Aquest tipus d’aparell respiratori recorda una mica els pulmons dels aranèids i dels escorpins.
L’aparell circulatori
L’aparell circulatori dels quilòpodes consta fonamentalment d’un vas dorsal i un vas ventral units mitjançant un anell aòrtic. El vas dorsal, o cor, és situat sota els teguments i els músculs dorsals i damunt el tub digestiu i els òrgans genitals. El sostenen unes brides musculars, de les quals unes són laterals i limiten uns sins o cavitats aliformes i unes altres són mitjanodorsals i configuren un si dorsal.
El cor és dividit en una sèrie de cambres, els límits de les quals es determinen per uns engruiximents que corresponen als ostíols. La primera cambra, situada al segon segment del tronc, es perllonga cap endavant mitjançant l’aorta cefàlica. Molt a prop del lloc on s’origina, configura l’anell aòrtic, format per la soldadura de vasos laterodorsals i lateroventrals, al qual envolta l’esòfag. A les regions lateroventrals d’aquest anell neixen les artèries forcipulars. L’aorta cefàlica continua fins a la meitat del cap, on origina una petita branca imparella que irriga l’esòfag i una mica més endavant dona dues grosses artèries cefàliques. Quan ateny el cervell, se separen dues noves artèries que, passant per sota la massa nerviosa, es dirigeixen cap a les antenes. Finalment l’aorta cefàlica travessa el cervell passant per la comissura del tritocervell i es bifurca en dues branques terminals que irriguen la vora anterior de la càpsula cefàlica.
El vas ventral se situa damunt la cadena nerviosa i, a partir del segon segment pedífer, origina un parell de vasos lateroventrals per segment, que són perpendiculars a l’eix del cos, i que esdevenen les artèries de les potes. Aquests vasos, a més, proporcionen ramificacions que irriguen els ganglis, els teguments i els músculs inferiors. En els escolopendromorfs algunes d’aquestes ramificacions acaben en forma de petites boles anomenades corpúsculs de Kowalevsky, que falten a la resta dels quilòpodes.
Cap endavant, el vas ventral s’acaba sobre el primer gangli ventral mitjançant dues branques, cada una de les quals origina tres ramificacions, que van a les peces bucals. Cap endarrere, aquest vas central finalitza damunt del gangli genital i dona dos parells d’artèries, una pel recte i l’altre per les glàndules accessòries dels òrgans genitals.
L’aparell excretor
Esquema d’una glàndula de verí d’un quilòpode, vista en un tall longitudinal (A) i transversal (B) de la forcípula. Hom hi distingeix: 1 forcípula, 2 orifici del canal de la glàndula verinosa, 3 canal excretor de la glàndula, 4 glàndula de verí, 5 artèria forcipular, 6 nervi, 7 múscul adductor curt, 8 placa dentària, 9 coxostèrnum, 10 múscul adductor llarg, 11 epiteli, 12 solc de la glàndula, 13 canal excretor, 14 alvèol conjuntivomuscular, 15 túnica conjuntivomuscular, 16 cèl·lula excretora, 17 nucli de la cèl·lula excretora.
Gustavo Hormiga, a partir de C. Duboscq 1899.
L’excreció es fa mitjançant un parell de tubs de Malpighi, conductes llargs que se situen sota l’intestí mitjà i desemboquen just al límit entre aquesta porció intestinal i la posterior. A més dels tubs de Malpighi, a alguns grups de quilòpodes hi ha un o dos parells de glàndules cefàliques. Al cap hi ha tambè un parell de glàndules maxil·lars, el conducte excretor de les quals s’obre en la cavitat bucal. Situades als segments anteriors del tronc, hi ha tres parells de glàndules que genèricament reben el nom de salivals. El parell de glàndules anteriors és unit íntimament al parell mitjà. Els canals excretors de les glàndules anteriors passen pel mig dels lòbuls de les posteriors, de manera que els canals d’uns i altres són contigus i paral·lels des del seu origen. En arribar al nivell del segment forcipular se separen i els canals excretors de les glàndules anteriors van a parar a la cavitat bucal i els de les glàndules mitjanes als orificis del sincoxostèrnum del segon parell de maxil·les. Les glàndules posteriors són més petites i perfectament ben individualitzades respecte de les altres.
Els seus canals, molt fins, es dirigeixen sense donar voltes a les pleurites del primer segment amb potes i s’obren a l’exterior en un punt situat damunt de la inserció de les potes. En els escolopendromorfs hi ha unes estructures globulars anomenades corpúsculs de Kowalevsky que sembla que tinguin una funció fagocitària i que també podrien produir cèl·lules saguínies. Es localitzen al damunt dels vasos sanguinis lateroventrals del cor des del quart segment pedífer fins al penúltim.

Els centpeus (Scolopendra cingulata) són potser els miriàpodes més coneguts de tothom, gràcies a les dimensions que alguns arriben a atènyer (fins a 17 cm) i a la pretesa perillositat del seu verí. Entre els miriàpodes hi ha, però, organismes diminuts i, de fet, la majoria són absolutament inofensius per a l’home.
Ramon Torres
Les glàndules del verí dels quilòpodes mereixen una atenció especial, tant perquè els són exclusives i característiques com per la important funció alimentària que duen a terme. A causa de les dimensions considerablement grans de les forcípules dels escolopendromorfs, els miriapodòlegs han estudiat preferentment les glàndules verinoses d’aquest grup i concretament de diverses espècies del gènere Scolopendra. Per això, la descripció que tot seguit s’exposa fa referència a aquest gènere. L’aparell verinós del quilòpodes es compon d’un parell de glàndules allotjades a l’interior de les forcípules. Aquest aparell es localitza, doncs, en el segment que segueix immediatament al cap i els seus apèndixs, els quals primitivament eren potes locomotores; a més, estan modificats i adaptats per a funcionar com a òrgans inoculadors del verí. Cada glàndula ocupa gairebé tot el telopodi, però no arriba al coxostèrnum de la forcípula. De color blanquinós, la seva regió posterior té forma cilíndrica i cap al davant es torna cònica fins al terç anterior de l’ungla, on el canal excretor s’individualitza i va a desembocar en un orifici ovalat situat a la cara superior de l’ungla, prop de l’àpex. A la vora externa de la glàndula hi ha un solc profund que ateny gairebé el canal excretor. La glàndula és innervada per tres branques del nervi forcipular, el qual s’origina en el gangli subesofàgic. Un vas arterial, provinent del collar esofàgic, acompanya el nervi i forma una ramificació que penetra al solc i es distribueix sobre les parets. Les tràquees tenen el mateix model de distribució que l’artèria.
Un tall transversal de la glàndula del verí, fet a qualsevol nivell, mostra perfectament la seva estructura interna. El perfil del tall és gairebé circular, amb un solc ample i profund que quasi arriba al canal excretor quitinós. Aquest canal té tres quartes parts de la seva paret amb perforacions on desemboquen les cèl·lules glandulars. La quarta part restant és llisa, recoberta només d’un epiteli i correspon a la regió pròxima al solc. Les cèl·lules glandulars es disposen radialment al voltant del canal; són piriformes, amb un citoplasma granulós i amb el nucli situat a prop de l’extrem extern. Cada cèl·lula glandular és envoltada per un alvèol conjuntivomuscular i els diferents alvèols tenen les seves parets comunes; aquesta íntima unió dels alvèols i l’existència d’una túnica conjuntivomuscular al voltant de tota la glàndula, dona una gran compacitat a tota l’estructura.
L’aparell reproductor
Les glàndules genitals dels quilòpodes se situen dorsalment al cos, entre el cor i el tub digestiu. Aboquen el seu contingut a l’exterior a través d’un orifici situat al segon segment genital.
L’aparell reproductor masculí consta dels testicles, amb el canal deferent que posteriorment configura l’epidídim, les bosses dels espermatòfors i finalment el canal ejaculador. A més, hi ha diversos parells de glàndules accessòries o annexes. La forma i nombre d’aquests elements varia segons els diferents ordres. En els geofilomorfs, l’aparell testicular és format per dos testicles grans, allargats i fusiformes, situats a nivells diferents; de cadascun surt un canal deferent superior i un d’inferior que van a parar a un canal deferent mitjà únic. La seva regió terminal és molt dilatada i actua com una vesícula seminal. Posteriorment, el canal deferent es bifurca en dos canals ejaculadors que envolten l’intestí i s’uneixen per sota, fent un canal ejaculador únic que penetra a l’atri genital, la porció distal del qual és evaginable, té peces quitinitzades i constitueix el penis. A la part anterior de l’atri genital desemboquen un parell de glàndules annexes dorsals de forma tubular i relativament llargues i a l’extrem posterior, prop de l’orifici genital, hi va a parar un canal format per la fusió dels canals de dues glàndules annexes ventrals. Els escolopendromorfs tenen un nombre de testicles que pot variar de quatre fins a vint-i-sis, distribuïts per parelles o desaparellats. De cadascun dels extrems de cada testicle neix un canal deferent que va a parar a un canal deferent axial, la part anterior del qual s’uneix a l’esòfag mitjançant un lligament suspensor. Els mascles de l’ordre dels litobiomorfs tenen un testicle imparell filiforme que es continua amb un canal deferent. Pel que fa als escutigeromorfs, els mascles tenen dos testicles; cada testicle presenta una regió anterior dilatada o macrotestis i una regió posterior estreta, en forma de canal llarg i enrotllat, que s’anomena microtestis.
L’aparell reproductor femení és molt menys variable. Fonamentalment consta d’un ovari, un o dos oviductes, un parell de receptacles seminals i un, dos o tres parells de glàndules accessòries. Els geofilomorfs presenten un ovari imparell molt llarg, situat dorsalment al damunt de l’intestí. El seu extrem posterior es perllonga mitjançant un parell d’oviductes que envolten l’intestí posterior i després es reuneixen en un conducte comú que va a parar a la regió anterior de l’atri genital. Hi ha un parell de receptacles seminals cilindricovoides, els canals dels quals poden desembocar separadament a l’atri, o bé unir-se prèviament en un canal comú. La majoria de geofilomorfs presenten dos parells de glàndules annexes, però alguna família en té tres. Hi ha un parell de glàndules dorsals que són llargues, tubulars i ben desenvolupades; els seus canals s’obren a la zona dorsal anterior de l’atri genital. Les dues glàndules ventrals normalment són molt més petites i els seus canals desemboquen a les zones lateroventrals de l’atri. El tercer parell correspon a unes glàndules suplementàries minúscules, situades damunt del diverticle dorsal de l’atri genital.
Biologia i ecologia
Reproducció
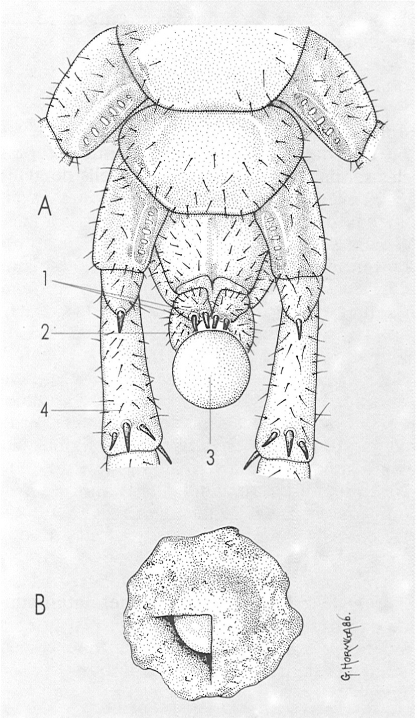
Esquema de la posició de l’ooteca en els darrers segments del cos (A) d’una femella de quilòpode durant la seva fabricació, i visió general d’una ooteca (B). Hom hi consigna: 1 esperó, 2 gonopodi, 3 ou, 4 darrer parell de potes.
Gustavo Hormiga, a partir de J. M. Demange 1981.
La gran majoria de quilòpodes, llevat d’algunes espècies de litobiomorfs, com per exemple el Lithobius tricuspis, que forma autèntiques colònies estables a sota de les pedres, viuen sols durant gairebé tot l’any. Quan arriba l’època de la reproducció, que als Països Catalans correspon fonamentalment als mesos de febrer, març i abril, és freqüent trobar parelles que es mantenen fins que ha tingut lloc la transferència d’espermatozoides del mascle a la femella. Acomplert aquest objectiu, normalment la parella es desfà, i la posta i la seva cura, quan es realitza, és a càrrec exclusiu de la femella. En tots els quilòpodes, la transferència dels espermatozoides a les vies genitals de la femella es realitza d’una forma indirecta, mitjançant espermatòfors.
Els espermatòfors, que contenen els gàmetes masculins, són fabricats a les vies genitals del mascle. La seva estructura és molt variable; en els litobiomorfs, geofilomorfs i escutigeromorfs, és una simple gota d’esperma protegida per una coberta molt minsa; en els escolopendromorfs són estructures molt complexes que tenen diverses cobertes, de les quals la més externa és la més gruixuda i contràctil i la més interna, molt fina, s’enfonsa profundament en la massa dels espermatozoides formant una invaginació que s’obre a l’exterior a través de la capa externa. Quan la femella pren l’espermatòfor, aquesta capa externa exerceix una pressió sobre la interna que fa que la invaginació surti cap a fora i penetri les vies genitals, on esclata i allibera l’esperma.
Tots els mascles dels quilòpodes, tret dels escutigeromorfs, que dipositen directament els espermatòfors sobre el sòl, teixeixen una tela més o menys complexa damunt de la qual deixen els espermatòfors. Després, el mascle fa un seguit de moviments per tal d’orientar la femella fins que aquesta agafa un espermatòfor mitjançant els seus gonopodis, que esdevenen turgents. Una vegada ha atrapat l’espermatòfor, la femella segrega un líquid espès que envolta totalment l’aparell espermàtic; quan hi ha un excés d’aquest líquid, la femella es gira i n’absorbeix una part. Hom pensa que és en aquest període, que pot durar una hora aproximadament, quan l’esperma penetra les vies genitals de la femella i té lloc la fecundació. Per això, hom pot afirmar que, si bé cal la presència dels dos sexes i hi ha estimulació mútua durant les diferents fases de la transferència espermàtica, no hi ha mai una còpula entre ells i la fecundació dels òvuls és interna. Al cap d’un temps, que varia d’uns grups a altres, després de la fecundació la femella fa la posta (al gènere Scolopendra té lloc al cap de quatre setmanes de la fecundació). El nombre d’ous i la seva grandària varien molt i no hi ha una relació directa entre la quantitat i la grossària. La talla dels individus es relaciona més amb el nombre d’ous de la posta que no pas amb les dimensions d’aquests.
La posta és protegida per la femella de dues maneres ben diferents. Les femelles dels geofilomorfs i des escolopendromorfs busquen o excaven elles mateixes una cavitat en el sòl, sota una pedra, sota la molsa, etc., hi fan la posta i immediatament l’envolten amb el seu cos. Quan té lloc l’eclosió dels ous i neixen les larves, les femelles també tenen cura dels nounats durant un cert temps. Ous i larves són protegits dels atacs de depredadors i de les floridures que tenen tendència a desenvolupar-se damunt de la seva superfície. Les femelles dels litobiomorfs i dels escutigeromorfs no fan mai una posta massiva, sinó que ponen els ous d’un en un i els van abandonant al sòl. Les femelles del gènere Lithobius, durant un cert temps, porten cada ou agafat mitjançant les ungles i els esperons dels seus gonopodis i li imprimeixen un moviment de rotació, al mateix temps que el van impregnant de partícules de terra que s’adhereixen gràcies a una mucositat que segreguen; el resultat final d’aquesta operació és la fabricació d’una ooteca de forma lenticular, bombada del mig, la qual és transportada durant un cert temps per les femelles, bo i tenint cura d’evitar-li qualsevol topada, i finalment la dipositen en algun lloc del sòl on hi hagi una certa humitat i no se n’ocupen més.
Desenvolupament postembrionari

Larves de quilòpodes corresponents a un litobiomorf (Lithobius variegatus rubriceps, a dalt) i a un escutigeromorf (Scutigera coleoptrata, a baix). En el primer cas, l’exemplar correspon a l’estadi de larva II del període de desenvolupament anamorf; hom hi pot comptar només vuit parells de potes i un nombre de segments del tronc inferior al de l’adult. En el segon cas, l’exemplar correspon a l’estadi immatur del període epimorf, ja que disposa de 15 parells de potes i segments, és a dir, el nombre definitiu, tot i que vist per sobre només es veuen aquells segments que tenen la tergita dorsalment desenvolupada. L’absència de segments genitals ben formats el diferencia de l’adult.
Antoni Serra.
Des del moment de la desclosa de l’ou fins que atenyen la maduresa, els quilòpodes sofreixen una sèrie de canvis i transformacions morfològiques, més o menys profundes, que es designen de manera general com a desenvolupament postembrionari. Aquest desenvolupament pot ser de dues menes, que separen els quilòpodes en dos grans grups, els epimorfs i els anamorfs. S’anomenen epimorfs aquells quilòpodes (geofilomorfs i escolopendromorfs) que en el moment de la desclosa ja posseeixen el nombre de segments i de parells de potes característic de l’adult. Contràriament, els quilòpodes anamorfs (litobiomorfs i escutigeromorfs) desclouen amb un nombre incomplet de segments i d’apèndixs i no és sinó al cap d’una sèrie de mudes que assoleixen el nombre definitiu de metàmers, és a dir, el de l’adult.
Els embrions dels geofilomorfs trenquen l’ou amb la dent de la seva cutícula embrionària i neixen amb el nombre definitiu d’apèndixs, la qual cosa indica que el seu creixement embrionari és molt llarg. Aquest primer estadi, que rep el nom de peripatoide, es caracteritza per tenir els teguments llisos, sense segmentació, els apèndixs no articulats, i per la manca d’estigmes, tràquees, glàndules coxals a les potes terminals i glàndules anals; a més, té la regió genital amorfa i el tub digestiu ple de reserves vitel·lines. A continuació, se succeeixen dos estadis anomenats fetus b i fetus a, en el decurs dels quals els apèndixs desenvolupen les seves articulacions i apareixen la segmentació del cos i les tràquees. L’estadi següent és el d’adolescent I i en ell els apèndixs, les tràquees, els estigmes i les glàndules anals inauguren el seu funcionament; la reserva vitel·lina ha desaparegut totalment i la regió genital s’ha desenvolupat, tot i que encara no presenta apèndixs. El nombre d’estadis que separen l’adolescent I de l’adult varia d’uns grups a altres.
En el primer estadi postembrionari, els escolopendromorfs ja tenen el nombre definitiu de parells de potes. Malgrat això, no pot considerar-se com a peripatoide, ja que la segmentació del cos és acabada i les potes ja són dividides en artells. Així, aquest estadi correspon aproximadament al segon dels geofilomorfs i, per tant, el desenvolupament està més avançat als escolopendromorfs. El gènere Scolopendra presenta després d’aquest estadi un estadi fetus immòbil, sense làmina dentada a la sincoxita forcipular i amb les fissures mandibulars simplement indicades. Segueixen tres estadis adolescents, a continuació dels quals l’animal ateny l’estat adult.
El desenvolupament postembrionari dels litobiomorfs es divideix en dos períodes, un d’anamorf i un d’epimorf, i per això a vegades se l’anomena hemianamorfosi. Cada període es compon de diversos estadis, que per a l’espècie Lithobius forficatus són les que tot seguit exposem. El període anamorf presenta sis estadis: fetus, amb 7 parells de potes i un parell de monyons; larva I, amb 7 parells de potes, un parell a mig desenvolupar, 2 parells de monyons i dos parells d’estigmes; larva II, amb 8 parells de potes, 2 parells de monyons i 2 parells d’estigmes; larva III, amb 10 parells de potes, 2 parells de monyons i 3 parells d’estigmes; larva IV, amb 12 parells de potes, 3 parells de monyons i 4 parells d’estigmes; larva mitjana, amb 12 parells de potes, 1 parell a mig desenvolupar, i 2 parells de monyons. A partir de l’estadi següent comença el període epimorf, que ja presenta el nombre definitiu de potes (15) i d’estigmes (6). Els estadis que el componen són: agenital, amb la regió genital completament glabra i amb els seus apèndixs amb prou feines esbossats; immatur, amb 1 + 1 esperons als apèndixs genitals de la femella, els artells terminals indiferenciats acabats en una ungla minúscula, i els mascles amb els apèndixs molt rudimentaris; pseudomatur I, amb els apèndixs genitals més desenvolupats; pseudomatur II, caracteritzat pel nombre d’ocel·les, de glàndules coxals i de dents forcipulars; madur júnior, amb l’aparell genital del mascle acabat de desenvolupar, encara que el nombre d’artells antenals, d’ocel·les i de glàndules coxals és encara inferior al de l’adult; madur sènior, que ja ha assolit el seu creixement complet i és, per tant, adult.
En els escutigeromorfs passa el mateix que en els litobiomorfs, ja que existeix un període anamorf seguit d’un epimorf. Analitzarem tot seguit els estadis de cada període tal com els presenta Scutigera coleoptrata. El període anamorf consta dels estadis: larva I, amb 7 parells de potes; larva II, amb 9 parells de potes, 2 parells de monyons i 5 tergites; larva in, amb 11 parells de potes, 2 parells de monyons i 6 tergites; larva IV, amb 13 parells de potes, 2 parells de monyons, 7 tergites ben formades i una tergita esbossada. A continuació segueixen els estadis epimòrfics, amb 15 parells de potes i 8 tergites: agenital, amb una zona genital embrionària, amb apèndixs glabres i encara independents a la femella; immatur, amb la regió genital diferenciada, els apèndixs de la femella fusionats i els del mascle molt curts; prematur, amb els apèndixs genitals més desenvolupats (els del mascle atenyen la meitat de la seva llargària); pseudomatur, amb la regió i els apèndixs genitals diferenciats de l’adult només per una quitinització menys accentuada; madur, és a dir que ja ha aconseguit el desenvolupament complet.
Alimentació i òrgans vulnerants

Els hàbits depredadors dels quilòpodes queden ben il·lustrats en aquesta fotografia, d’un litobiomorf (Lithobius variegatus rubriceps) en el moment de caçar un insecte, en aquest cas, un microcorifi. L’ha capturat amb les forcípules, amb les quals li ha injectat el verí, i seguidament procedeix a trencar-li els teguments i a menjar-se’n, amb l’ajut de les maxil·les, les parts internes, més toves.
Antoni Serra.
Tots els quilòpodes són estrictament depredadors i, per tant, cacen i s’alimenten de petits animals. Normalment les seves preses són altres artròpodes, però també invertebrats com ara oligoquets; els geofilomorfs són especialistes en perseguir i capturar aquests cucs en les seves pròpies galeries del sòl. És notable la capacitat dels escutigeromorfs per a caçar dípters, mosques i mosquits, gràcies a la seva gran rapidesa de moviments. Un sistema ben particular de caça és l’emprat pels escolopendromorfs, sobretot per les grans espècies d’Scolopendra i Theatops, quilòpodes que tenen el darrer parell de potes molt robust i proveït de fortes espines i ungles, que el converteixen en un poderós òrgan prènsil: quan detecten la presència d’una presa de dimensions regulars, per exemple, de la grandària d’un grill, l’agafen i l’immobilitzen amb el darrer parell de potes i després, girant el cos, li claven les forcípules i li injecten el verí necessari per a paralitzar-la o, més normalment, matar-la. Sigui quin sigui el sistema de caça, tots utilitzen les forcípules per tal d’injectar el verí de les seves glàndules. La composició d’aquest verí varia d’unes espècies a altres, però normalment és fet de petites molecules de substàncies farmacològicament actives, com ara la histamina i l’acetilcolina, i molècules grans de caràcter proteic que actuen com a toxines; juntament amb aquestes substàncies hi ha enzims que afavoreixen la difusió de les toxines i que són els responsables del dolor que provoca immediatament la picada.
S’ha parlat a bastament de l’efecte del verí dels quilòpodes, sobretot de les escolopendres, sobre l’home, i normalment hi ha una clara tendència a exagerar-ne els efectes. Als Països Catalans l’espècie que és més gran és Scolopendra cingulata. Algunes versions populars atribueixen a aquesta espècie alguns casos en els quals s’ha produït la mort d’una persona o d’algun animal domèstic. Ara bé, experiències realitzades per miriapodòlegs dignes del crèdit més absolut i incidents comprovats directament demostren que la picada de Scolopendra cingulata a una persona adulta normal produeix un dolor intens a partir del moment en què l’animal injecta el verí, seguit d’una inflamació de la regió on s’ha produït la picada (si l’escolopendra ha picat en un dit es pot arribar a inflamar tota la mà fins al puny) i un augment de la temperatura d’aquesta zona, que acaba produint un edema i una necrosi superficial al punt de la picada; tots aquests efectes desapareixen al cap de pocs dies. D’altra banda, les conseqüències d’una picada depenen de l’època en què es produeixi; el cas que acabem d’exposar correspondria a un dia de primavera, que és quan l’escolopendra és totalment activa; si, contràriament, la picada es produeix a l’hivern, època en la qual l’escolopendra es troba gairebé en un estat de letargia, els efectes queden reduïts a una petita butllofa urticant, poc dolorosa, que desapareix al cap d’una o dues hores. Un altre aspecte que s’ha constatat és que una picada no produeix cap mena d’immunització, ja que picades successives realitzades en el decurs d’un mes produeixen totes els mateixos efectes que la primera.
Les conseqüències de la picada d’una escolopendra en els artròpodes són ben diferents. En general, tots són molt sensibles al verí d’aquests quilòpodes, fins a produir-los una mort ràpida. Entre els quilòpodes mateixos, Scutigera coleoptrata és extraordinàriament sensible al verí de Scolopendra cingulata, la qual mor al cap de vuit o deu minuts de rebre’n la picada. En canvi, l’escolopendra posseeix una gran immunitat al seu verí i pot ser picada impunement per un altre individu de la mateixa espècie.
Tota la resta de quilòpodes no representen cap perill per a l’home ja que, en primer lloc, llevat d’alguns Lithobius relativament grans, no manifesten cap mena d’agressivitat envers una persona, a la qual no identifiquen com a presa; en segon lloc, són tan petits que no aconsegueixen travessar l’epidermis humana; i, finalment, la poca quantitat de verí que inoculen i la seva composició no causen més que una lleugera coïssor.
Algunes espècies de quilòpodes, sobretot de geofilomorfs, en certes ocasions ocupen temporalment les fosses nasals, les cavitats auditives i fins i tot les vies digestives. Aquests casos, cal considerar-los més com a presències accidentals i temporals que no pas com a veritables qüestions de parasitisme; normalment només provoquen a l’hoste algunes pertorbacions i molèsties no massa importants. Els geofilomorfs tenen unes forcípules molt petites i febles i, per tant, les seves glàndules ventrals juguen un paper molt important. Diverses experiències han demostrat que quan toquen un petit artròpode, per exemple, un tisanur, l’impregnen d’un líquid que segreguen les glàndules ventrals i li provoquen la mort; un insecte d’una talla més gran, per exemple, una mosca, també és impregnat d’aquesta secreció i, si bé no el mata, com a mínim, el paralitza. Aquest sistema de caça esdevé molt eficaç per a capturar els cucs de terra; els geofilomorfs els abracen i segreguen el líquid necessari per a enverinar-los. Aquest fet justifica l’assimilació fisiològica de les glàndules forcipulars amb les glàndules ventrals; les primeres només difereixen de les segones per la seva adaptació especial a un parell de potes modificades morfològicament i fisiològica, com són les forcípules.
Quan els quilòpodes han agafat una presa i l’han morta o immobilitzada amb el seu verí, o simplement li han produït una ferida més o menys important amb les forcípules, comencen a devorar-la. Excepte les escolopendres més grans, que poden menjar-se íntegrament tot el cos de la presa, inclòs l’exosquelet dels artròpodes, normalment els quilòpodes en trenquen la cutícula a nivell d’una articulació mitjançant les seves poderoses mandíbules i procedeixen a buidar-la del seu contingut. Les forcípules tenen una funció prou important en l’absorció de l’aliment; s’enfonsen a la massa alimentària, l’arrenquen i ajuden el segon parell de maxil·les a dur-la cap a la boca. El resultat final és que de la presa només queda pràcticament l’exosquelet, totalment buit del seu contingut.
Els depredadors dels quilòpodes són diversos: ocells, rèptils, mamífers i, fins i tot, insectes grans. Per defensar-se dels seus atacs, utilitzen dos sistemes, un d’actiu, mitjançant les forcípules, i un altre que es pot considerar com a passiu. Aquest sistema passiu és molt evident en diverses espècies de Lithobius, en les quals el darrer parell de potes és més llarg i robust que la resta i normalment no l’utilitzen per a caminar, sinó que el mantenen una mica enlairat, sense tocar a terra; quan es troben davant d’un perill, es queden immòbils o fugen, però en tots dos casos aixequen exageradament el darrer parell de potes per tal que si el depredador els ataca, els pinci per una d’aquestes potes; aleshores el quilòpode s’autotomitza aquest apèndix, és a dir, se’n desprèn voluntàriament mitjançant una forta contracció dels músculs situats a nivell de l’articulació del trocànter i el prefèmur. Si aconsegueix fugir, en les successives mudes es regenera progressivament l’apèndix autotomitzat. A vegades en aquesta regeneració es produeix un error genètic i en lloc de la pota corresponent es forma un gonopodi o una antena.
És ben freqüent que els quilòpodes tinguin paràsits, externs com ara àcars o bé interns com per exemple gregarines; és interessant assenyalar que aquests endoparàsits són específics, és a dir, que una determinada espècie de gregarina parasita una determinada espècie de quilòpode.
Els hàbitats: els biòtops epigeus
Habitualment hom diu que els quilòpodes són higròfils i lucífugs. Aquestes característiques ecològiques semblen confirmades pel fet que es troben pràcticament arreu on hi ha un cert grau d’humitat i no hi ha una exposició directa a les radiacions solars.

El medi lapidícola ofereix un microclima en el qual les condicions d’humitat i temperatura són òptimes per a la majoria dels quilòpodes. La fotografia mostra un centpeus (Theatops erytrocephala) posat al descobert en aixecar una pedra, situada al costat d’un tronc.
Antoni Serra.
De tots el biòtops epigeus, el medi lapidícola és el preferit per una gran majoria d’espècies de quilòpodes. El nom del gènere Lithobius, el més abundant als Països Catalans, indica ben clarament l’afecció dels seus representants a aquest medi. Dessota les pedres, es crea un microclima humit, amb pocs corrents d’aire i amb una temperatura notablement més estable que a l’exterior; a aquestes condicions ambientals, cal afegir un altre factor per explicar el delit que tenen els quilòpodes de refugiar-se sota les pedres. Tots presenten un elevat estereotropisme, és a dir, una gran necessitat que el seu cos estigui en contacte amb un objecte dur.
En general s’accepta com un fet demostrat que els quilòpodes fugen de la llum del sol. Experiències realitzades amb algunes espècies de Lithobius, però, demostren que aquest fototropisme negatiu no sempre es manifesta clarament. Si en un terrari, amb una atmosfera ben humida, situat a l’ombra, és a dir, sense aguantar una insolació directa, hom posa diversos trossos de vidre de diferents colors (transparents i negres, per exemple), es veurà que els Lithobius es refugien indistintament sota els vidres clars o sota els foscos. Això pot ésser degut al fet que l’efecte de dessecació produït per l’energia tèrmica de les radiacions solars és contrarestat per la humitat ambiental, i demostra que altres possibles accions són molt menys importants i no semblen influir en l’elecció del color del vidre que hipotèticament podria protegir-los millor.
Els medis muscícola i humícola representen biòtops del domini epigeu caracteritzats per la seva foscor relativa, una temperatura més aviat baixa i poc variable, un grau d’humitat que arriba gairebé a la saturació i una gran quantitat de matèria orgànica en diferents graus de descomposicó. A les molses, la fauna dels quilòpodes és representada per formes petites corresponents a algunes espècies de Lithobius, Cryptops i geofilomorfs de dimensions petites. A l’humus no passa el mateix, ja que la fullaraca del sòl dels boscos caducifolis permet el desenvolupament d’una fauna humícola important que s’alimenta dels detritus vegetals; els quilòpodes, a part de les condicions favorables d’aquest medi, hi són ben freqüents, pel fet que, en ser depredadors estrictes, utilitzen la fauna humícola com a base de la seva alimentació.
Els hàbitats: els biòtops endogeus
Pel que fa al medi endogeu, dos grups de quilòpodes tenen representants que es poden considerar com a veritables habitants del sòl, els geofilomorfs i els escolopendromorfs, tots de cos comprimit i estilitzat, amb teguments prims, poc pigmentats, els apèndixs curts i molts anoftalms (tots els del primer i el gènere Cryptops del segon).
Els quilòpodes del sòl estan sotmesos a una migració vertical i hi ha un trànsit del medi endogeu cap als altres medis exposats. Durant les èpoques seques, els horitzons més superficials del sòl perden humitat i els quilòpodes, igual que la resta de fauna, davallen envers capes més profundes bo i cercant condicions més òptimes i aprofitant les galeries excavades per altres éssers d’aquest medi. Contràriament, quan, per excés de pluges o perquè puja el nivell freàtic de les aigües de les vores d’un riu o d’un llac, el sòl s’inunda, els trobem a les capes més superficials, al dessota de les pedres.
Si bé no podem parlar d’autèntiques espècies foladòfiles de quilòpodes, no és gens estrany que als caus i als nius d’animals superiors sovint es trobin aquests artròpodes. Aquest fet, hom el pot explicar ben fàcilment, ja que aquests hàbitats són com microcavernes, en les quals les condicions ambientals són pràcticament constants i favorables durant l’any.
Els hàbitats: els biòtops hipogeus
Finalment, considerarem els biòtops hipogeus. El medi cavernícola és constituït per les cavitats del sòl, és a dir, les coves, els avencs, les esquerdes, les fissures i el medi subterrani superficial. També s’hi poden afegir les cavitats artificials com ara els pous, els túnels, les mines, etc. Sigui quin sigui el seu origen, tots aquests biòtops tenen unes condicions ambientals molt semblants i configuren les característiques peculiars del món subterrani. Els quilòpodes, igual que tots els animals que viuen en el medi hipogeu, hi estan adaptats en un grau més o menys elevat. Segons les seves característiques morfològiques i fisiològiques, podem establir una classificació ecològica dels quilòpodes cavernícoles.
En primer lloc, hi ha els quilòpodes trogloxens, és a dir, els que són forasters de les coves. Poden ésser-ne regulars, quan hi entren voluntàriament, atrets per la humitat, la temperatura i els recursos alimentaris, o bé accidentals, quan la seva presència a les coves és fortuïta i involuntària. Lògicament, no presenten cap signe d’adaptació al medi subterrani i no hi fan mai el seu cicle biològic complet. A aquesta categoria pertanyen alguns escolopendromorfs i geofilomorfs, però tal vegada l’exemple més clar el constitueix Scutigera coleoptrata, un escutigeromorf habitualment epigeu.
Les espècies troglòfiles són aquelles que viuen i es reprodueixen normalment a les coves, però que també es troben en els ambients epigeus. Un bon exemple d’aquesta categoria ecològica el constitueixen nombroses espècies de geofilomorfs i, sobretot, de litobiomorfs, que formen comunitats estables prop de l’entrada de les coves, on es reprodueixen amb tota normalitat.
La categoria de quilòpodes més interessant des d’aquest punt de vista ecològic és la dels troglobis, animals estrictament adaptats al medi hipogeu i que no es troben mai a l’exterior. Són gairebé tots litobiomorfs, i tenen una sèrie de caràcters morfològics que poden considerar-se com a autèntiques respostes adaptatives al medi hipogeu. Molts litobiomorfs troglobis mostren un grau de despigmentació més o menys accentuat segons les espècies; les seves antenes presenten un gran desenvolupament, que és resultat tant de l’elevat nombre d’artells que les componen, com de l’augment de la llargada de cadascun; paral·lelament a l’allargament de les antenes, els litobiomorfs cavernícoles sofreixen un notable increment de la longitud de les potes, especialment les terminals; la regressió de l’aparell ocular és també molt freqüent i sovint condueix a una anoftàlmia absoluta; paral·lelament a aquesta regressió, hom constata un augment progressiu de la grandària de l’òrgan de Tömösvary, el qual ateny les seves dimensions més grans quan la falta d’ocel·les és absoluta.
Sistemàtica i filogènia
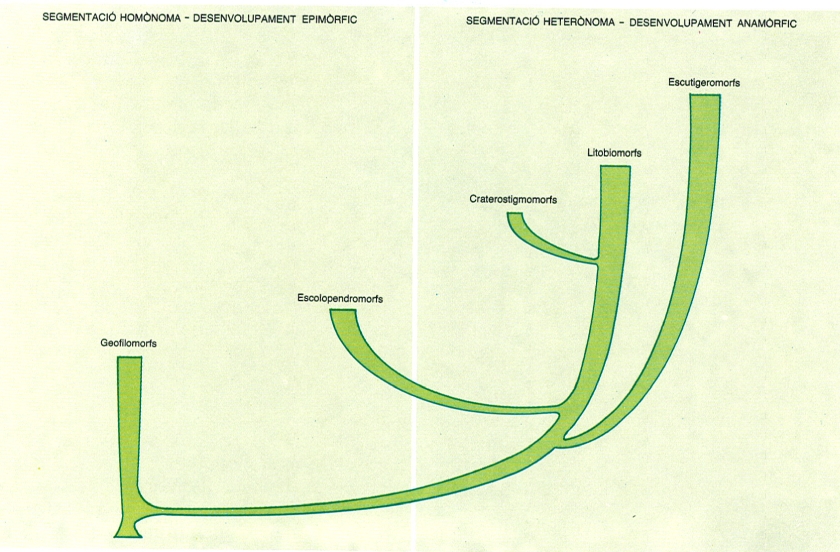
Esquema de la filogènia dels quilòpodes que exposa la manera com, hipotèticament, s’han originat els cinc ordres actuals a partir d’un ancestre comú. A l’esquema hom pot veure com els grups més primitius presenten una segmentació del tronc homònoma i un desenvolupament de tipus epimòrfic, mentre que els més evolucionats, tenen la segmentació heterònoma i el desenvolupament anamòrfic.
Maber, a partir d’originals diversos
La classe dels quilòpodes (Chilopoda) comprèn cinc ordres: els litobiomorfs (Lithobiomorpha), els escutigeromorfs (Scutigeromorpha), els escolopendromorfs (Scolopendromorpha), els geofilomorfs (Geophilomorpha), i els craterostigmomorfs (Craterostigmomorpha). Els quatre primers ordres es troben representats als Països Catalans, mentre que el darrer, el dels craterostigmomorfs, és propi de la regió australiana. Abans d’entrar en les característiques de cadascun dels grups de quilòpodes, farem algunes consideracions sobre la seva evolució i filogènia.
Tota l’evolució dels quilòpodes és presidida per dos processos, els efectes dels quals són ben diferents. D’una banda, hi ha una clara tendència a la reducció del nombre de segments, i a una simplificació de les seves estructures. Simultàniament, té lloc una constant adaptació a les petites variacions del seu medi, que provoca una diferenciació i complicació d’algunes característiques morfològiques.
La taquigènesi, és a dir, la tendència a la condensació del desenvolupament, és un procés que domina l’evolució dels quilòpodes. Aquesta contracció taquigenètica provoca una maduració cada cop més precoç dels elements sexuals i abreuja la durada del desenvolupament del cos, de tal manera que fixa el nou organisme en un estat que pot qualificar-se de larvari, ja que no té ni la talla ni el nombre de segments dels seus antecessors. Aquesta reducció metamèrica pot ser de dues menes, una d’ordre evolutiu, fixada en el patrimoni hereditari, i una altra d’ordre ecològic, lligada al creixement postembrionari. Hom pot fer evident la reducció metamèrica evolutiva en estudiar la segmentació dels diversos ordres de quilòpodes; els uns amb els segments homònoms, de volums iguals i de superfícies tergals idèntiques (geofilomorfs), i els altres amb els segments heterònoms més o menys voluminosos, amb tergites grans i petites (escolopendromorfs, litobiomorfs i escutigeromorfs). Tots els metàmers dels geofilomorfs tenen una gran tergita dividida en dues plaques, pretergita i metatergita, mentre que en els altres tres ordres els metàmers són alternativament de tergita gran i de tergita petita, o sigui, hi ha una alternança de metàmers macrotergals i metàmers microtergals. Amb l’estudi d’aquesta segmentació i de diverses estructures morfològiques i anatòmiques, hom ha demostrat que els metàmers microtergals deriven de metàmers macrotergals per amputació del territori metamèric posterior i això implica la desaparició dels estigmes, dels troncs nerviosos sensorials tergoposteriors i una modificació de la musculatura macrotergal primitiva.
Aquestes constatacions permeten precisar el que és la reducció metamèrica i comprendre que en un primer moment la reducció tendeix a modificar certs metàmers no a l’atzar, sinó regularment un de cada dos, reduint el seu volum i, en un segon, a fer desaparèixer el metàmer mateix amb els seus apèndixs. Hi ha, doncs, una reducció metamèrica que interessa els elements d’un segment i una reducció metamèrica que afecta el nombre de metàmers. Aquest procés evolutiu, els efectes del qual són fixats en el patrimoni hereditari, implica una heteronomia secundària derivada de l’homonomia segmentària primitiva.
A part d’aquests fenòmens hereditàriament establerts n’existeixen d’altres que són accidentals i que estan relacionats principalment amb les condicions ecològiques que actuen sobre el desenvolupament. El resultat de la seva acció és una reducció metamèrica ecològica o oligomeria ecològica. Aquest tipus de reducció només té lloc en els geofilomorfs.
Les larves d’aquests quilòpodes tenen davant del tèlson una zona àpoda que pot originar, després d’una muda, nous segments amb potes. Condicions ecològiques particulars determinen una disminució o un atur del creixement segmentari d’aquesta zona i dónen com a resultat una gran diversitat de fórmules segmentàries individuals i específiques.
Conjuntament amb aquests processos de contracció, se’n manifesta un altre d’adaptació, de complicació, que hom considera com el model evolutiu per excel·lència i que es denomina diferenciació. És en virtut d’aquest procés que apareixen i es desenvolupen diverses variacions progressives que es manifesten quan una línia filètica es perpetua a través de nombroses generacions, en unes condicions que no comporten greus alteracions de l’equilibri morfològic dels seus representants. Hi ha, per tant, un antagonisme entre els dos processos de contracció i de diferenciació, i és en el resultat d’aquest antagonisme on podem trobar l’origen de les associacions entre estructures contradictòries, unes d’aparença arcaica i d’altres de caràcter especialitzat. A continuació exposarem els principals caràcters evolutius del diversos ordres de quilòpodes, tot analitzant les relacions filogenètiques que hi ha entre ells.

Quilòpodes (× 2). 1 Pachymerium ferrugineum, 2 Cryptops trisulcatus, 3 Scolopendra cingulata, 4 Lithobius piceus, 5 Scutigera coleoptrata.
Marisa Bendala
De tots els quilòpodes, els geofilomorfs són els més pròxims al tronc original primitiu, tal com ho indiquen els nombrosos caràcters arcaics que s’hi poden reconèixer. La seva desclosa és tardana i el cos ja té la seva segmentació completa molt abans que aquesta es produeixi. El cos és molt allargat i està format per un elevat nombre de segments, de 30 a 173, que pot variar d’uns individus a altres de la mateixa espècie. El cap és, proporcionalment, molt petit. Els segments del tronc són homònoms; tergites i esternites són precedides per esclerites intercalars, entre les quals destaquen les pretergites de grandària aproximadament igual a la meitat de les metatergites. És freqüent trobar entre la tergita forcipular i el cap les restes d’una esclerita tergal que correspon a un metàmer maxil·lar. Existeixen encara òrgans segmentaris, tals com les estigmes i les glàndules esternals, disposats en una sèrie ininterrompuda d’un extrem a l’altre del cos. El darrer parell de potes és separat pels segments terminals. El nombre d’artells de les antenes és baix, de 14. El tronc de les mandíbules és homogeni, sense fissures i el còndil dorsal mediocre. Les esclerites dels segments terminals són nombroses i els apèndixs genitals són simples en els dos sexes. Els òrgans dels sentits estan poc desenvolupats. Al costat d’aquests caràcters primitius, trobem algunes estructures que tenen la marca de la diferenciació: teguments més gruixuts a les formes més grans; maxil·les i forcípules soldades dues a dues per la base; vora externa del fèmur, de la tíbia i de les forcípules atrofiada; paratergites i pleurites nombroses; sistema traqueal complex.
En els escolopendromorfs el nombre de caràcters arcaics disminueix. Si bé l’animal surt de l’ou completament format, aquesta formació no ha precedit gaire a la desclosa. El cos és ja molt més condensat, el nombre de segments pedífers disminueix a 21 o 23 i és fix. Els segments són homònoms; les pretergites i les presternites desapareixen, encara que es poden trobar restes de l’esclerita postcefàlica. El nombre d’òrgans segmentaris disminueix (9 o 10 estigmes amb les pleurites reduïdes) o bé desapareixen totalment (glàndules ventrals). En definitiva, són poques les estructures primitives que han estat respectades per la contracció taquigenètica o la reducció metamèrica, mentre que la diferenciació n’ha desenvolupat d’altres: antenes amb artells més nombrosos (de 15 a 23); mandíbules amb fissures complicades i amb un gran còndil articular; forcípules amb làmines dentades marginals; el darrer parell de potes es toca i és més pròxim a l’eix del cos; la major part de les esclerites terminals ha desaparegut, etc.
Els mateixos fenòmens són encara més evidents en els litobiomorfs, en els quals la desclosa té lloc molt abans que s’acabi la formació del cos, i es redueix la talla i el nombre de segments proveïts de potes, que queda fixat en 15; el cap és gran i representa del 9 al 10% de la longitud del cos; la segmentació dorsal és clarament heterònoma; el nombre d’estigmes es redueix a 6 o 7; i les potes terminals tornen a estar separades, tot ressorgint la disposició primitiva per efecte de la contracció. Ressorgiments anàlegs s’observen al fèmur i a la tíbia de les forcípules, que són cilíndrics, amb la vora externa no atrofiada, a les mandíbules, les fissures de les quals són incompletes, i en el sistema traqueal, més simple fins i tot que en els geofilomorfs. En contraposició, progressen certes diferenciacions: els artells de les antenes i els ocel·les són més nombrosos, el labre presenta làmines petites i els apèndixs genitals es compliquen, almenys en la femella.
En els escutigeromorfs predominen les diferenciacions i són més acusades que en els ordres anteriors. Hi ha una migració dels estigmes, que es tornen dorsals i imparells, amb un sistema traqueal complex. El cap presenta un notable desenvolupament i hi ha una complicado de les estructures de la faringe. Les antenes i els tarsos de les potes presenten un nombre molt elevat d’artells. Al costat d’aquestes diferenciacions tornen a aparèixer caràcters vells, tals com la dissociació dels membres de les maxil·les i de les forcípules, caràcter desconegut en els altres grups i que representa la reaparició d’estructures larvals.
És imprescindible parlar de l’ordre dels craterostigmomorfs, encara que no es trobi als Països Catalans (és propi d’Austràlia), per a poder tenir una visió completa de la filogènia d’aquesta classe. Aquest ordre participa, d’una banda, de les característiques dels escolopendromorfs, com ara l’aspecte general, el nombre d’esclerites tergals (21) i les diferenciacions de la sincoxita forcipular; de l’altra, s’aproxima als litobiomorfs pel nombre de segments pedífers (15) i d’estigmes (6 parells) i per la conformació del fèmur i de la tíbia forcipular, complets externament. Finalment, la forma del cap i la composició de l’aresta mandibular fan pensar en els òrgans corresponents de certs geofilomorfs. Segons tot això, sembla que aquest quilòpode s’ha de concebre com a escolopendromorf, en procés de contracció, que encara no ha assolit completament el tipus litobiomorf i que presenta alguns caràcters que hom pot considerar com a autèntiques reminiscències ancestrals.
Si analitzem els diferents grups segons l’ordenació en què els hem esmentat, veiem que les estructures simples, primitives, del primer desapareixen progressivament per deixar lloc a associacions de caràcters que es podrien considerar contradictòries; malgrat tot, ho són només en aparença i la seva associació s’explica pels efectes combinats de la contracció taquigenètica o reducció metamèrica i de la diferenciació. Així, doncs, hi ha un tronc primitiu d’afinitats geofilonianes, del qual se separa un tronc secundari; aquest ha originat les branques dels escolopendromorfs, dels craterostigmomorfs, dels litobiomorfs i dels escutigeromorfs. Aquests darrers sembla que es van separar aviat del tronc comú, segons que indiquen les acusades diferències que presenten. Els litobiomorfs es van separar més tardanament i no han tingut temps d’especialitzar-se tant com els escutigeromorfs.
Els grups de quilòpodes
Els geofilomorfs


Dels coneguts centpeus, l’exemplar de la fotografia, un geofilomorf típic de llocs una mica secs (Dignathodon microcephalum), es caracteritza per la forma general que pren el cos, que s’aprima fins arribar al cap, que és molt petit. Destaca, també, la robustesa especial del darrer parell de potes, caràcter que és present tant en els mascles com en les femelles.
Un dels caràcters que defineix els geofilomorfs, la presència de dues regions — pretergita i metatergita— al tegument endurit que cobreix cada segment, és clarament visible en aquest exemplar (Geophilus carpophagus), una espècie que, tot i tenir 55 parells de potes, no és de les més grosses que podem trobar al nostre país, on en coneixem d’altres que arriben a tenir-ne 160 parells. Noteu la semblança de gruix del darrer parell de potes amb totes les altres, caràcter que en aquesta espècie separa els mascles de les femelles.
Antoni Serra
Els quilòpodes geofilomorfs es caracteritzen, sobretot, pel fet de tenir més de vint-i-cinc parells de potes. Tenen el cos molt llarg, una mica deprimit dorsiventralment, como si fos una cinta. Els segments del tronc estan recoberts per una tergita dividida en dues regions, una anterior o pretergita, i una posterior, més gran, o metatergita. Cada segment del tronc té un parell de potes, excepte els quatre darrers, que són àpodes. El nombre d’estigmes és igual al de parells de potes menys un. El darrer parell de potes queda molt separat pels segments àpodes i es disposa lateralment respecte a l’eix longitudinal del cos. De les diverses famílies que componen l’ordre dels geofilomorfs, n’hi ha quatre que tenen representants als Països Catalans: els himantàrids (Himantariidae), els esquendílids (Schendylidae), els orídids (Orydae) i els geofílids (Geophilidae).
Els himantàrids són grossos i amb un nombre elevat de segments. El labre és format per una sola peça transversa, amb la vora bucal profundament escotada i dentada. L’aresta apical de les mandíbules és formada per una placa dentada robusta i molt pigmentada i diverses làmines pectinades en forma de pinta. Diversos gèneres, com Himantariella, Pseudohimantarium, Haplophilus, Nesoporogaster, etc., es troben als Països Catalans, i les espècies Pseudohimantarium mediterraneum i Himantariella scutellaris són les més freqüents a les nostres contrades.
Contràriament al que passa a la família anterior, els esquendílids són petits i filiformes. Tenen el labre fet de dues peces laterals, unides per una peça mitjana en forma d’arca dividida en dents o callositats; a la vora apical de les mandíbules hi ha una placa dentada i una sola làmina pectinada. Com a gèneres d’aquesta família presents als Països Catalans cal mencionar Hydroschendyla, Schendyla i Brachyschendyla. És notable l’espècie Hydroschendyla submarina que viu a la costa, entre la grava i les algues, suportant immersions temporals degudes als moviments del mar.
La família dels orídids és caracteritza per tenir el labre format per una sola peça. La vora apical de les mandíbules presenta diverses làmines en forma de pinta i no té mai una placa dentada. Les forcípules són curtes i petites, de tal manera que no són visibles des de la banda dorsal. L’únic representant als Països Catalans d’aquesta família correspon a l’espècie Orya barbarica que ha estat trobada a l’Altet (l’Alacantí) i Santa Pola (el Baix Vinalopó).
Finalment, els geofílids presenten un labre format sempre per tres peces, de les quals la mitjana és molt variable, ja que pot ser reduïda, o bé pot ser fins i tot més gran que les laterals. La vora apical de la mandíbula és ocupada por una sola làmina pectinada contínua; no hi ha cap placa dentada. De tots els geofilomorfs, aquesta família és la que té més espècies als Països Catalans, pertanyents als gèneres Henia, Dignathodon, Chaetechelyne, Strigamia, Clinopodes, Pachymerium, Geophilus, etc.
Els escolopendromorfs
Els components d’aquest ordre es caracteritzen per presentar 21 parells de potes, excepte alguns grups, que en tenen 23. El segment forcipular no té tergita pròpia, ja que aquesta s’ha fusionat amb la del primer segment pedífer; d’aquesta manera, a continuació del cap es troben 21 tergites subiguals, les quals no presenten cap divisió transversal en dues regions. Els estigmes respiratoris són laterals i n’hi ha nou i deu parells. El darrer parell de potes és molt robust, prènsil i s’orienta gairebé en la mateixa direcció que l’eix longitudinal del cos.

Les escolopendres o centpeus (Scolopendra cingulata) en estadi de desenvolupament incomplet, com la de la fotografia, tot i tenir el nombre definitiu de potes i de segments (que és de 21), es distingeixen pel color ataronjat viu del darrer parell de potes, la darrera tergita, el cap i les antenes, color que desapareix en els adults. A les nostres contrades, l’espècie il·lustrada pot arribar a fer 10 cm de llargada.
Antoni Serra
Els escolopendromorfs comprenen dues famílies, la dels escolopèndrids (Scolopendridae) i la dels criptòpids (Crytopidae).
La família dels escolopèndrids se subdivideix en dues subfamílies, la dels escolopendrins (Scolopendrinae) i la dels otostigmins (Otostigminae), aquesta darrera no representada als Països Catalans.
Els escolopèndrids es caracteritzen per la presència constant de quatre ocel·les a cada banda del cap, i pel fet de tenir els tarsos de totes les potes fets de dos artells. Les esternites normalment tenen dos solcs longitudinals paral·lels, excepcionalment només un o fins i tot cap, però mai no presenten solcs transversals. A la subfamilia dels escolopendrins pertanyen Scolopendra cingulata i Scolopendra canidens oraniensis. S. cingulata es caracteritza per tenir l’apòfisi coxal del darrer parell de potes curta, subcònica i amb cinc espines com a màxim; el prefèmur d’aquestes potes és curt i té com a màxim deu espines. Els caràcters que defineixen S. canidens oraniensis són principalment l’apòfisi coxal del darrer parell de potes que és llarga, subcilíndrica, amb almenys set espines, i el prefèmur, llarg, amb vint o més espines.

Els criptòpids, dels quals veiem un representant a la fotografia (Cryptops parisi) són probablement els menys coneguts de tots els quilòpodes. Les petites dimensions del seu cos i les característiques dels biòtops on viuen (en el sòl, sota les pedres, sota les escorces dels arbres o entre la fullaraca) fan que passin desapercebuts.
Antoni Serra
La família dels criptòpids comprèn tres subfamílies: la dels escolocriptopins (Scolocryptopinae), la dels criptopins (Cryptopinae) i la dels teatopsins (Theatopsinae), de les quals únicament coneixem al nostre país les dues darreres. A la família dels criptòpids, els quilòpodes no tenen mai ulls. Els tarsos de les potes que van del primer parell fins al dinovè tenen un sol artell (excepte a Trigonocryptops, que en tenen dos) i les esternites tenen solcs transversals o un solc longitudinal, molt rarament dos de longitudinals. La subfamilia dels teatopsins es caracteritza pel fet de presentar taques oculars, zones despigmentades de les zones laterals del cap, la sincoxita forcipular amb plaques llargues i dentades i les potes del darrer parell curtes i molt gruixudes. L’única espècie d’aquest grup que es troba a casa nostra és Theatops erytrocephala.
A la subfamilia dels criptopins, diferentment de l’anterior, no hi ha taques oculars. La vora anterior de la sincoxita forcipular no té plaques dentades, les potes terminals són llargues i primes, i a la majoria de gèneres hi ha una placa dentada con una serra, molt característica. Dels diversos gèneres que componen aquest grup, només Cryptops té representants als Països Catalans: C. trisulcatus, C. parisi, C. hortensis i C. savignyi.
Els litobiomorfs
Els litobiomorfs es caracteritzen sobretot per tenir 15 parells de potes. D’altra banda, després de la tergita forcipular hom troba 16 tergites (la número 15 queda amagada sota l’anterior), de les quals les que ocupen els llocs 2, 4, 6, 9, 11, 13, i 15 són més petites que les altres. Els segments 3, 5, 8, 10, 12 i 14 tenen un parell d’estigmes laterals i el darrer parell de potes és llarg, més desenvolupat que la resta. Ni les antenes ni els tarsos de les potes no es divideixen en nombrosos artells.

D’entre els litobiomorfs, el de la fotografia (Lithobius variegatus subespècie rubriceps) és, sens dubte, el més freqüent als Països Catalans i el més gros del seu grup. Normalment viu sota les pedres, en llocs no massa humits, i, fins i tot, força assolellats, i té una notable capacitat d’autotomització del darrer parell de potes (és a dir, d’arrencar-se’I voluntàriament) quan es troba en perill.
Antoni Serra.
Els litobiomorfs comprenen dos subordres, el dels litobiomorfins (Lithobiomorphinae) i el dels craterostigmomorfins (Craterostigmomorphinae). Ja hem comentat que aquest darrer és considerat com un ordre a part per alguns miriapodòlegs. Tots els seus representants als Països Catalans pertanyen a la família dels litòbids (Lithobiidae), concretament als gèneres Lithobius i Eupolybothrus. Les dues diferències morfològiques fonamentals que separen aquests dos gèneres són que en el primer els pors coxals dels quatre darrers parells de potes són poc nombrosos i es disposen en una sola filera, i els gonopodis dels mascles són curts, mentre que a Eupolybothrus els pors coxals són nombrosos i es distribueixen en diverses fileres irregulars, i els gonopodis dels mascles són molt llargs. Al gènere Lithobius pertanyen gairebé totes les espècies dels Països Catalans. Com a més freqüents hom pot citar L. rubriceps, L. castaneus, L. pilicornis, L. tricuspis, L. piceus, L. pyrenaicus, etc., i interessants endemismes cavernícoles com L. ambulotentus d’una cova dels Pirineus orientals, L. jorbai d’una cavitat de Castelló i L. vivesi de diverses coves i avencs de Mallorca. Pel que fa al gènere Eupolybothrus, només es coneix una citació de E. leptopus, a una cova de Tarragona.
Els escutigeromorfs

L’escutígera (Scutigera coleoptrata, l’única espècie que viu al nostre país), és inconfusible per l’exagerada llargada de les seves potes i antenes. La rapidesa dels seus moviments li permet caçar fins i tot petits dípters, com ara mosques i mosquits. Cal destacar, també, que és un quilòpode força freqüent a les cases, encara que actualment és molt rar en els medis urbans.
Antoni Serra
Per moltes característiques morfològiques, els escutigeromorfs poden ésser considerats com els quilòpodes més evolucionats. El seu tronc es compon de 15 segments pedífers, cada un dels quals té un parell de potes i una esternita; contràriament, només hi ha set tergites entre la tergita del segment forcipular i les dues dels dos segments terminals (intermediari i anal). A la part mitjana de la vora posterior d’aquestes 7 tergites hi ha una escotadura ben marcada, al mig de la qual s’hi troba un solc longitudinal curt que correspon a un estigma respiratori que, per tant, és dorsal i imparell. El cap és globulós i, a banda i banda, s’hi troben els ulls, grosses masses subtriangulars formades per gran quantitat d’ocel·les agrupats. Les antenes són llargues, filiformes i estan formades per una gran quantitat de petits artells. Igualment, els dos artells tarsals de les potes estan dividits en anells molt nombrosos.
Comprenen una sola família, la dels escutigèrids (Scutigeridae), que, al seu torn, engloba dues subfamílies, la dels escutigerins (Scutigerinae) i la dels tereuonemins (Thereuoneminae), aquesta darrera, exòtica. A la primera pertany Scutigera coleoptrata, espècie comuna als Països Catalans.