Un bioma petit i fragmentat
Què és una selva? La majoria de les persones entenen per selva una selva equatorial plujosa, és a dir, una pluviïsilva intertropical. I, tanmateix, en regions més allunyades de l’equador també hi ha altres boscos d’estructura exuberant i amb aspecte de selva. Són les selves plujoses temperades, que, tot i que presenten una riquesa menor en espècies que les selves tropicals, per la seva estructura, fisiognomia, complexitat, diversitat, com també per l’alta especialització de les adaptacions de les seves espècies i per les seves interaccions, s’assemblen més a aquestes que no pas a altres boscos.
La fragmentació i la dispersió actuals
Les característiques dels diferents biomes del planeta depenen principalment de les condicions climàtiques i edàfiques, però també, en bona part, de la història de cadascuna. Sense oblidar, és clar, altres factors ecològics que sovint tenen una importància decisiva per al desenvolupament, la dinàmica i les heterogeneïtats del bioma, tant en l’espai com en el temps. Entre aquests factors cal citar les pertorbacions, ja siguin naturals o d’origen antròpic, el relleu i les interaccions entre els individus de les diferents espècies. Així, en cada regió del planeta els factors ecològics actuen com a filtre, seleccionant, d’entre totes les disponibles localment, només unes espècies determinades; les que aconsegueixen sobreviure al llarg de la història integraran la flora i la fauna de cada zona concreta. La història, doncs, és un factor ecològic major, i es pot dir que en pocs biomes es manifesta de manera més eloqüent que en el de les selves temperades.
Per la seva distribució geogràfica, les selves temperades constitueixen un bioma disjunt, molt fragmentat i distribuït per regions molt allunyades entre elles. En comparació amb altres biomes, com les selves tropicals o el bosc boreal, l’àrea total ocupada per les selves temperades no és gaire gran. Tot i això, estan repartides per tots els continents i en cada un han experimentat històries evolutives diferents. Tots aquests factors han contribuït a configurar un bioma amb una gran riquesa d’espècies i una gran diversitat de formacions.
Les selves temperades prosperen sota climes de temperatures suaus, sense grans oscil·lacions tèrmiques anuals, i amb precipitacions poc o molt abundants i uniformement distribuïdes al llarg de l’any. Evidentment, fora de l’àmbit equatorial, aquestes condicions només es produeixen en regions molt concretes del planeta, sotmeses a la influència de determinants règims de vents que porten pluges i suavitzen les temperatures extremes. El seu àmbit geogràfic comprèn les zones subtropicals humides influïdes pels vents alisis i monsònics, com també les zones temperades plujoses condicionades pels vents marítims de l’W. Totes aquestes selves temperades es localitzen en illes i a les façanes litorals dels continents —les orientals en el cas de les àrees subtropicals afectades per alisis i monsons, les occidentals en el de les temperades pròpiament dites—, sotmeses al règim dels vents marítims de ponent. La seva característica climàtica fonamental és l’absència gairebé total de glaçades i de períodes de sequera, si bé el ritme estacional pot ser marcat.
L’origen tropical remot
Encara que, generalment, les espècies integrants d’una flora selvàtica concreta es corresponen amb les de la regió florística a la qual pertanyen, en la composició de les selves temperades és remarcable la presència d’espècies d’origen tropical. Es poden esmentar com a exemple algunes falgueres arborescents a Nova Zelanda, palmeres i bromeliàcies epífites (Tillandsia usneoides) al SE dels Estats Units i al SE del Brasil, bambús del gènere Chusquea a Xile i al Brasil,i lauràcies del gènere Persea a la Macaronèsia, a l’Amèrica del Nord i a les selves sinojaponeses. Aquesta és una característica que, com altres que es comentaran, evoca l’origen tropical o subtropical comú d’aquestes selves. Certament, d’una manera genèrica, es pot considerar que tots els boscos actuals deriven de la selva plujosa tropical primigènia, però les laurisilves criden l’atenció perquè pràcticament constitueixen una selva tropical fora del tròpic. Al llarg de la història, factors com les altes temperatures i l’absència de grans oscil·lacions climàtiques han tingut un paper molt important en la redistribució de les espècies vegetals i en l’evolució de les seves adaptacions. No és casual que els boscos actuals que més s’assemblen als que existiren en el Terciari mitjà siguin precisament els que es localitzen en regions de clima suau i plujós; la persistència d’aquestes condicions climàtiques, que en certes àrees s’han mantingut al llarg de milions d’anys, ha afavorit la subsistència de flores antigues en àrees disjuntes on, d’altra banda, també han trobat refugi algunes espècies de coníferes relictes.
Molts dels gèneres representats en la flora de les selves temperades actuals ja eren importants en els boscos del Terciari. El registre fòssil documenta de manera molt evident les afinitats florístiques existents entre aquestes plantes que, sobretot a nivell de família i de gènere, són notables i justifiquen la distribució disjunta de les diferents espècies. A més, si es comparen entre elles les flores de les diferents selves, aquestes afinitats indiquen clarament un origen geogràficament pròxim, però alhora posen de manifest que el procés evolutiu ha estat diferent.
A les selves temperades hi ha un nombre elevat de paleoendemismes. Són espècies relictes, que solen tenir una àrea de distribució restringida, i que han quedat aïllades filogenèticament a causa de l’extinció dels tàxons més pròxims. Aquest seria el cas d’espècies relictes de podocarpàcies i de coníferes de les selves temperades de l’hemisferi sud, o concretament del taxodi de Virgínia o d’aiguamoll (Taxodium distichum) al SE dels Estats Units.
La influència limitada de les glaciacions
Algunes característiques fisiognòmiques de les selves temperades posen de manifest que es formaren sota les condicions climàtiques tropicals imperants fins la meitat del Terciari, de manera que les actuals selves temperades es poden considerar relictes o residus de les selves d’aquell període, molt més riques en espècies. D’altra banda, la gran biodiversitat i l’alt grau d’especialització de les interaccions interespecífiques existents indiquen que aquestes selves han evolucionat ininterrompudament durant un llarg període de temps i en unes condicions climàtiques que, pel fet d’haver-se mantingut sense grans oscil·lacions al llarg de milions d’anys, també es podrien considerar relictes.
En efecte, a excepció de les situades en zones de latitud i altitud més elevades, les selves temperades no es veieren afectades per les glaciacions quaternàries. La selva valdiviana, per exemple, no experimentà de manera dràstica els efectes de les glaciacions, i per això moltes espècies s’hi refugiaren. Tot i que els casquets de glaç cobrien la totalitat dels Andes fins al paral·lel 44°S, i malgrat que arribaren a cobrir la meitat oriental de l’illa de Chiloé, a partir d’aquella latitud cap al N les glaceres només s’estenien per les àrees muntanyoses més altes. Els Andes, amb la seva orientació N-S, facilitaren les vies de migració de moltes espècies que, escapant de les glaciacions del Plistocè, migraren cap a latituds més càlides. La zona valdiviana es convertí així en un refugi segur per a moltes d’aquestes espècies.
Un cas similar seria el de les selves temperades del NW de l’Amèrica del Nord, les anomenades selves vancouverianes. Les glaciacions han estat un procés geològic important a les cotes més elevades dels arrengleraments muntanyosos de l’àrea que va d’Alaska a Oregon. Al Plistocè, la capa de glaç continental arribava pel S fins a la badia de Puget Sound, on avui es troba la ciutat de Seattle. Tanmateix, el bosc plujós d’aquesta regió existeix com a mínim des de fa dos milions d’anys i s’hi troben plantes pertanyents a gèneres que han canviat molt poc des del Mesozoic. L’última glaciació, que al S d’Alaska s’acabà fa tot just 4 000 anys, amb prou feines reduí la biodiversitat d’aquests boscos gràcies a l’existència de corredors migratoris i de nombrosos refugis. Fins i tot hi ha espècies vegetals que encara es troben en estadi de dispersió a partir d’aquests refugis, i que fins fa poc no havien arribat als seus actuals límits septentrionals de distribució, com és el cas de la tuia gegant o “western red-cedar” (Thuja plicata) o de l’“Alaska yellow-cedar” (Chamaecyparisnootkatensis).
En moltes regions de Nova Zelanda, les terrasses dels rius, les valls glacials en forma de U i les planes al·luvials són testimonis de l’últim període de glaciació, que assolí el seu punt àlgid fa tan sols 25 000 anys i acabà fa menys de 15 000 anys. Les glaceres encara són molt evidents al llarg dels Alps Meridionals neozelandesos (la més gran és la glacera Tasman, que té 29 km de longitud). Tanmateix, sembla que en aquest arxipèlag aïllat del Pacífic meridional el descens de les temperatures durant les glaciacions fou tan sols d’entre 4 i 6°C.
Un descens semblant (entre 4 i 5°C) s’enregistrà a la Macaronèsia. Gràcies a això, a les Canàries, les Açores i Madeira s’han conservat espècies i comunitats senceres del Terciari; de fet, molts fòssils trobats a l’Àfrica septentrional i al S d’Europa corresponen a espècies ja desaparegudes d’ambdós continents i que, actualment, només sobreviuen en aquests arxipèlags. No hi ha cap àrea, ni al continent africà ni a l’europeu, que reuneixi unes característiques anàlogues, i és per això que la flora d’aquestes illes té un enorme interès biogeogràfic i ecològic.
L’abundància d’endemismes (que, per posar alguns exemples, constitueixen el 81,1% de la flora de Nova Zelanda, el 46,6% de la de les illes Canàries i el 34,1% de la del Japó) posa de manifest l’evolució autònoma que han experimentat les selves temperades. D’altra banda, però, en funció de l’hemisferi i de l’àrea geogràfica que ocupen, en aquestes selves hi ha representats elements molt diversos. És evident que, després de les repetides fragmentacions de Lauràsia i Gondwana i un cop iniciada la deriva dels diferents continents i illes, la flora i la vegetació de cada espai del bioma s’han desenvolupant de manera independent al llarg de la història. Els faigs australs (Nothofagus), per exemple, són característics de les selves temperades de l’hemisferi sud, mentre que alzines i roures (Quercus) i altres gèneres de fagàcies són típics de les de l’hemisferi nord. Així i tot, els components majoritaris d’algunes selves temperades pertanyen a famílies i gèneres que, a primer cop d’ull, no semblen ser del tot coincidents amb el bioclima. Potser el cas més sorprenent d’aquesta discordança sigui el de la selva plujosa de coníferes de la façana nord-occidental de l’Amèrica del Nord, sobretot si es compara amb la resta de selves temperades. (Vegeu també El clima dels darrers deu mil anys)
La tectònica de plaques, motor de la història
La complexitat del bioma de les selves temperades és extrema i la contribució dels factors geogràfics i històrics a aquesta complexitat ha fet difícil i polèmica la seva interpretació. Ja a mitjan segle passat Joseph D. Hooker, Charles Darwin i Alfred R. Wallace havien polemitzat en la interpretació de la distribució actual de les espècies: per què aquestes són on són i com hi han arribat. Les ciències naturals disposen de dues grans teories unificadores: la de la tectònica de plaques i la de l’evolució per selecció natural (entre altres mecanismes evolutius) que ajuden a organitzar les observacions fetes sobre el terreny i a plantejar-se noves preguntes. (Vegeu també La formació dels continents)
La deriva dels continents i la resiliència florística
Quan el gran continent de Pangea començà a fragmentar-se, a l’inici del Juràssic, les gimnospermes dominaven els boscos: les araucàries (Araucaria) eren cosmopolites, els ginkgos (Ginkgo) abundaven a l’hemisferi boreal i els podocarps (Podocarpus) a l’austral. De la primera fragmentació, en resultaren dos grans continents: Lauràsia, format pel que avui són l’Amèrica del Nord i Euràsia (tret de l’Índia), i Gondwana, constituït per les terres dels actuals continents d’Àfrica, l’Amèrica del Sud, l’Índia, Austràlia i l’Antàrtida. Posteriorment, aquests dos supercontinents es dividiren novament i els fragments s’anaren separant.
El clima del Cretaci i de la primeria del Terciari era força càlid. El gradient de temperatura a les latituds més altes era suau i la zonació climàtica no era gens acusada, tot i que s’hi distingien una zona tropical (entre els 30°N i els 30°S), una de temperada i, finalment, una altra de més petita, de clima relativament sec, incrustada entre les dues primeres i situada a la part occidental d’un i altre continent, entre els 30°N i els 30°S. Els registres d’isòtops d’oxigen, de pol·len i de restes vegetals fòssils indiquen que, durant aquest període, la humitat era molt elevada; no existien les estepes, ni els deserts, ni els casquets polars, el relleu dels continents era molt poc accidentat i tenia una altitud mitjana molt inferior a l’actual. No hi havia barreres geogràfiques importants, de manera que la biota podia dispersar-se fins a latituds elevades, on el clima es mantenia lliure de glaçades.
En aquestes condicions ambientals, durant el Cretaci inferior i mitjà (fa uns 118-105 milions d’anys) les angiospermes iniciaren la seva evolució fulgurant als dos grans continents, Lauràsia i Gondwana. El registre fòssil testimonia també que les flores de les latituds superiors de tots dos hemisferis ja eren força diferents en el moment de la seva aparició, durant el Cretaci. A la primeria del Terciari (fa uns 65 milions d’anys), en un període de tan sols uns 50 milions d’anys, la riquesa en plantes vasculars assolí nivells extraordinaris. Les angiospermes acabaren arraconant les gimnospermes i els pteridòfits, i és interessant constatar que aquest auge i diversificació no estan relacionats amb canvis climàtics acusats.
Fins ben entrat el Miocè, el clima temperat i humit dominant afavorí la formació d’extenses selves tropicals i subtropicals d’angiospermes. En tots dos hemisferis hi havia representants d’una flora pantropical arctoterciària en la qual abundaven diferents famílies, com les lauràcies (Laurus, Persea, Cinnamomum), les anacardiàcies (Rhus, Pistacia), les teàcies (Camellia), les moràcies (Celtis, Morus, Broussonetia), les hamamelidàcies (Hamamelis, Liquidambar), les aquifoliàcies (Ilex), les ericàcies (Arbutus, Gaultheria), les fagàcies (Castanea, Fagus, Nothofagus, Castanopsis, Lithocarpus, Quercus), les mimosàcies (Albizia, Acacia), les fabàcies (Robinia), les cesalpiniàcies (Cercis) o les saxifragàcies (Ribes, Hydrangea). Altres famílies tropicals que atenyien latituds temperades extratropicals a l’hemisferi boreal eren les magnoliàcies (Liriodendron, Magnolia, Tetracentron), les juglandàcies (Carya, Engelhardia [=Engelhardtia], Juglans), les hipocastanàcies (Aesculus) i encara d’altres. També hi havia famílies d’origen extratropical que s’estenien per les zones temperades i temperades fredes de l’hemisferi boreal, com ara les betulàcies, les platanàcies i les trocodendràcies. Altres tenien una distribució principalment tropical i temperada a l’hemisferi sud, com les mirtàcies, les winteràcies i les proteàcies. A l’hemisferi austral, les podocarpàcies (Podocarpus, Saxegothaea) i les araucariàcies (Araucaria, Agathis) també eren components habituals dels boscos.
Però per causes geològiques i climàtiques, tota aquesta flora començà a quedar fragmentada i arraconada. A mitjan Terciari s’inicià un refredament, gradual i generalitzat, que culminà amb les glaciacions del Quaternari. El descens de la temperatura fou més acusat a partir del final de l’Oligocè, fa uns 25 milions d’anys. Durant el Miocè, el Pliocè i el Plistocè es completaren les grans orogènesis alpines (Alps, Pirineus, Himàlaia, Andes, Muntanyes Rocalloses), que, en modificar sensiblement la circulació atmosfèrica, provocaren també canvis notables en el clima de les seves àrees d’influència. Paral·lelament, es dessecaren grans masses d’aigua (la Mediterrània, per exemple) i es produí una continentalització del clima. Tot aquest llarg procés de transformacions tingué repercussions enormes per a la flora: l’aparició de barreres climàtiques i geològiques afavorí la vicariançageogràfica i l’aparició d’àrees de distribució disjuntes per a nombrosos elements de la flora arctoterciària.
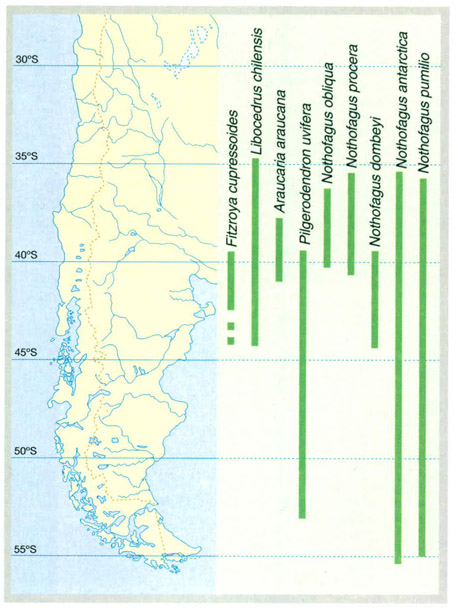
La distribució latitudinal d’algunes espècies arbòries de l’extrem meridional d’Amèrica ajuda a caracteritzar els diferents tipus de bosc d’aquesta àrea, però sobretot reflecteix les vicissituds climàtiques de finals del Terciari i del Quaternari. Coníferes com Fitzroya cupressoides o Araucaria araucana han quedat confinades en àrees restringides. Alguns Nothofagus abasten una amplitud latitudinal més gran, encara que a les latituds més septentrionals els faigs australs solen trobar-se a més altitud que a les latituds més meridionals, on poden aparèixer pràcticament al nivell de la mar.
Idem, a partir de Hueck, 1978
Es produí una reculada de la flora cap a latituds més baixes i quasi totes les espècies tropicals i moltes de termòfiles de la vegetació arctoterciària que no trobaren refugi s’extingiren. A Europa, la disposició de les muntanyes i de la conca mediterrània en sentit E-W posà obstacles a la migració, i aquesta és una de les causes principals que la flora d’aquest continent sigui més pobra en elements arctoterciaris que la de l’Amèrica del Nord o la de l’Àsia oriental. El fet que nombrosos elements florístics, com els faigs (Fagus), les magnòlies (Magnolia), les tsugues i hemlocks (Tsuga), els avellaners de bruixa (Hamamelis) o les dues úniques espècies existents del gènere Apollonias, presentin àrees de distribució disjuntes és un senyal de l’arraconament progressiu que patí la flora arctoterciària i que culminaria durant el Plistocè. A més, la presència d’endemismes relictes, com el ginkgo (Ginkgo biloba) o els taxodis (Taxodium) també és indicadora d’aquest procés.
Al llarg del Quaternari (els últims 1,6 milions d’anys) s’han produït 24 glaciacions, i de mitjana, la temperatura ha estat molt més baixa que durant el Terciari. Les fases glacials, en què s’acumulaven grans masses de gel a les regions de latituds altes i a les muntanyes, acabaven de sobte i eren succeïdes per períodes interglacials de menys durada, d’entre 10 000 i 20 000 anys. D’altra banda, al compàs de les glaciacions es començaren a formar, a tots dos hemisferis, les grans zones desèrtiques situades al límit dels tròpics, i d’això en resultà la fragmentació definitiva dels boscos primigenis en sentit latitudinal.
Pel que fa a la distribució de la vegetació, els canvis més dràstics es produïren durant el Plistocè. Les selves tropicals quedaren definitivament comprimides en direcció a l’equador; a les regions que no havien experimentat els canvis climàtics de manera tan acusada (refugis) persistí la vegetació subtropical dominada per elements arctoterciaris, i es produí una expansió de la tundra i un retrocés generalitzat dels boscos. Moltes espècies del Terciari s’extingiren, i també s’originaren noves espècies per hibridació i per poliploïdia. En general, durant els períodes glacials les espècies migraren en direcció a l’equador, i durant els períodes interglacials ho feren en sentit invers; i així, al compàs dels períodes d’avançament i retrocés del gel, es produïren grans retallades en les àrees de distribució dels components de la flora, que no estava adaptada a temperatures tan baixes.
El paper decisiu de l’orogènia
A Europa, com ja s’ha assenyalat, la disposició transversal de les muntanyes i de la conca mediterrània impedí la migració de les espècies i, de manera semblant, les regions àrides del Sàhara també actuaren de barrera. Alguns elements florístics, tanmateix, com roures i alzines (Quercus) ampliaren la seva àrea de distribució i s’estengueren fins als tròpics, encara que no han arribat a formar part de les selves temperades de l’hemisferi austral. La flora arctoterciària quedà arraconada en refugis. Així, per exemple, algunes espècies d’Europa i el SW d’Àsia, com la pícea de Sèrbia (Picea omorika), quedaren restringides als Balcans, i altres a les ribes de la mar Negra i la mar Càspia. La Macaronèsia constituí un dels refugis més apropiats per a molts elements d’aquesta flora arctoterciària; gràcies a això, s’hi conserven comunitats relictes que combinen espècies d’aquest període i espècies pròpies d’altres regions florístiques i que actualment presenten àrees de distribució disjuntes, com el gènere Apollonias, que té una espècie, el “barbusano” o banús canari (A. barbujana), que viu a les Canàries i una altra que creix a l’Índia. A l’altre extrem de la conca mediterrània (costa sud-occidental de la mar Càspia i costa nord-oriental de la mar Negra) subsisteixen també altres romanalles arctoterciàries, si bé formant part d’una flora molt més modificada. (Vegeu també Els processos orogènics)
A l’Índia, les muntanyes de l’Himàlaia, que començaren a formar-se a la primeria del Cenozoic, fa 65 milions d’anys, posseïen una vegetació tropical. El continent indi, després de separar-se del SE d’Àfrica, anà fent el seu viatge cap al S de la Xina a una velocitat d’uns 10 cm cada any, i s’hi uní ja entrat el Terciari. Els processos orogènics que desencadenà aquesta unió donaren lloc a la formació de les muntanyes de l’Himàlaia, que no es completà fins al Pliocè. Els processos orogènics provocaren canvis dràstics en la vegetació, i la major part de la flora originària fou substituïda. Actualment, els boscos estan formats per espècies que s’hi establiren i s’hi han mantingut des del Plistocè, i la vegetació tropical està restringida a la zona oriental.
A l’hemisferi austral la flora quedà fragmentada per causes geològiques i climàtiques. El procés de refredament culminà amb la glaciació total de l’Antàrtida i la destrucció de la seva flora, que probablement era temperada. S’hi han trobat restes fòssils (fulles, pol·len) de faig austral (Nothofagus). En canvi, no s’han trobat restes fòssils d’aquest gènere ni a l’Àfrica ni a l’Índia. A l’hemisferi austral, el gènere Nothofagus evolucionà independentment dels altres gèneres de fagàcies, propis de l’hemisferi boreal, i tot sembla indicar que el seu centre de dispersió se situà en àrees de latituds altes, coincidint amb una gran activitat tectònica i volcànica durant períodes freds. Les rutes de migració passaren necessàriament per l’Antàrtida, que durant el Cretaci superior, fa uns 100 milions d’anys, encara mantenia connexions amb Austràlia i l’Amèrica del Sud.
El registre fòssil demostra que l’Antàrtida tingué un paper clau en la redistribució de la flora de l’hemisferi sud. Austràlia, Nova Zelanda i l’Antàrtida acabaren de trencar les connexions encara existents durant la darreria del Paleocè, i des d’aleshores els seus boscos han evolucionat per separat. Aquesta circumstància es posa de manifest en l’especiació de nombrosos gèneres que els són comuns i en el gran nombre d’endemismes que inclouen. A l’hemisferi austral, a part l’Antàrtida, l’efecte de les glaciacions fou més acusat a les muntanyes, però en general la temperatura només baixà uns quants graus. Es produïren migracions i extincions d’espècies, sobretot de coníferes, però aquestes extincions no foren tan massives com a l’hemisferi boreal. A Austràlia, durant el Terciari les selves s’estenien per tot el territori i, concretament durant l’Oligocè, la prevalença d’un clima més sec afavorí també l’expansió dels eucaliptus (Eucalyptus) arreu del continent. La selva plujosa tropical quedà arraconada a la part nord-oriental, i les selves temperades, tant les plujoses com les més seques, a la part sud-oriental de l’illa continent i a Tasmània.
La dominància de les coníferes al NW dels Estats Units i el Canadà es deu a la història particular de la regió i al seu relleu. La flora arctoterciària també s’estenia per aquesta regió fins a l’Eocè, època en què formava selves temperades mixtes de planifolis i coníferes. El refredament progressiu del clima i l’orogènesi dels sistemes muntanyosos que es produí durant el període que va de l’Oligocè fins al Pliocè provocaren l’extinció de gairebé tota la flora arctoterciària i l’expansió d’una flora més xeròfila. Aquesta, però, fou desplaçada més tard per les coníferes, que en poc temps tornaren a ocupar les àrees de les quals havien estat excloses durant les glaciacions del Plistocè; els sistemes muntanyosos, disposats en direcció N-S, proporcionaren vies de migració per a les espècies, obligades a migrar a latituds més baixes pel refredament i la continentalitat del clima durant el Terciari. Espècies com ara les del gènere Sequoia es quedaren en aquestes latituds més baixes cap on havien migrat, mentre que altres foren bandejades durant el Pliocè i el Plistocè. Per contra, les pinàcies, alguns gèneres de les quals són components destacats a les selves vancouverianes (Picea i Abies, per exemple), esdevingueren dominants durant el Miocè i continuaren expandint-se durant el Pliocè, coincidint amb un nou refredament del clima.