La dominància de l’estrat arbori
Totes les boscanes decídues tenen una flora i una vegetació similars i, dins d’un cert marge de variació, comparteixen una mateixa estructura. En estat natural, aquesta estructura comporta un cert nombre d’estrats: al capdamunt, un estrat de grans arbres, els veritables protagonistes de les boscanes; ocasionalment, un estrat d’arbres petits o mitjans; per sota dels arbres, un estrat arbustiu que conforma el sotabosc, i, en darrer lloc, un estrat herbaci integrat generalment per una multitud de petites plantes anuals. També hi pot haver, però no sempre, un estrat lianoide format per plantes enfiladisses que, des de terra, es recolzen als troncs i les branques d’arbres i arbustos per pujar fins a diferents alçades, i que poden arribar a ser localment una part important de la biomassa del bosc. En algunes boscanes, l’estrat arbori és compartit per un cert nombre d’espècies diferents, mentre que en altres zones només hi ha una o dues espècies dominants. A les fagedes, per exemple, els faigs quasi sempre senyoregen en solitari, mentre que les rouredes presenten una diversitat més gran, caracteritzada per diferents espècies de roures que acostumen a barrejar-se amb arbres d’altres gèneres. Determinades boscanes complexes tenen nombroses espècies d’arbres la dominància relativa dels quals varia sovint d’un punt a un altre o segons gradients ambientals.
La diversitat de les espècies d’arbres
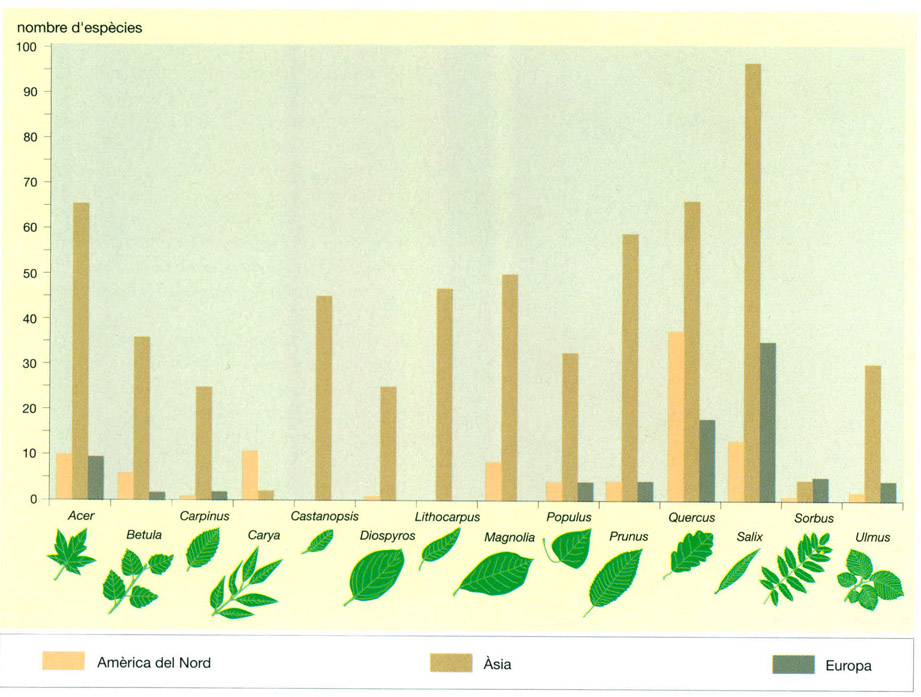
La major part dels gèneres d’arbres més comuns a les boscanes decídues estan més ben representats, en nombre d’espècies, al continent asiàtic que no pas a l’Amèrica del Nord o a Europa. Alguns, com Castanopsis i Lithocarpus fins i tot són exclusius de l’Àsia oriental. Els gèneres Quercus i Salix, especialment rics en espècies, són dels pocs que es troben en tots tres continents, amb 66 espècies asiàtiques, 37 d’americanes i 18 d’europees el primer, i 97, 13 i 35 respectivament el segon. La majoria de gèneres, però, només estan ben diversificats als boscos caducifolis de l’orient asiàtic, amb uns pocs representants a l’Amèrica del Nord i a Europa. Com mostra el diagrama de barres, els boscos d’aquest darrer continent són els més pobres en nombre d’espècies.
IDEM, a partir de dades proporcionades per l’autor
Aquesta imatge d’una flora rica i variada i d’una estructura complexa descriu una situació natural que ja no és fàcil de trobar en les boscanes decídues que subsisteixen als diferents països temperats, i encara menys a les parts més transformades del bioma. Hi ha hagut enormes canvis resultat de l’acció humana i només algunes àrees de boscanes decídues temperades mantenen l’esplèndida complexitat de la vegetació original.
Pel que fa a les principals espècies d’arbres i arbustos, la composició florística és prou homogènia en tot el bioma. La majoria dels arbres que creixen a les boscanes decídues són planifolis de fulla caduca, amb una proporció molt petita de coníferes de fulla caduca com els làrixs (Larix), que són en canvi un element important dels boscos boreals, sobretot a Sibèria, o els taxodis (Taxodium), que ho són als boscos perennifolis del SE dels Estats Units. No totes les espècies arbòries que creixen a les boscanes decídues són, però, de fulla caduca; hi ha àrees importants de boscos mixtos de planifolis decidus i coníferes perennifòlies i també, principalment a altituds elevades, a les latituds més fredes o bé en plantacions, algunes àrees on predominen aquestes coníferes.

Els arbres dels boscos temperats es distribueixen en uns 65 gèneres, pertanyents a una trentena de famílies diferents. Tots són caducifolis, però el seu aspecte és ben diferent, tant la mida i el port com la morfologia de la fulla i del fruit. Els tells (Tilia), per exemple, es distingeixen per les seves capçades altes i arrodonides i les flors fragants. En canvi, el tret més característic dels faigs australs (Nothofagus), de l’Àmèrica del Sud, són les seves petites fulles. Les càries (Carya) són arbres grans, emparentats amb les nogueres (Juglans), que es troben a les parts americana i asiàtica del bioma, on formen extensos boscos acompanyant els roures. En canvi, els castanyers falsos (Castanopsis), com el seu nom indica, estan emparentats amb els veritables castanyers (Castanea), als quals s’assemblen molt. Les seves castanyes també es mengen torrades. Els carpinus (Carpinus) són arbres de mida molt variable, de 13 a 30 m d’alçada, que presenten un tronc característicament acanalat i fruits en aments penjants, semblants als dels bedolls (Betula), uns caducifolis de vida curta que creixen en sòls pobres a les vores dels boscos d’Amèrica, Europa i Àsia. En llocs humits també dels tres continents són comuns els verns (Alnus), uns arbres molt resistents que treuen flors abans que fulles i fan fruits en forma de cons llenyosos. Els oms (Ulmus), de fulles simples i petits fruits alats, no formen poblacions denses sinó que creixen escampats pels boscos. Les zelkoves (Zelkova), del Caucas i l’Àsia oriental, i els lledoners (Celtis), que tenen representants també a les parts europea i americana del bioma, hi estan emparentats.
Jordi Corbera
No obstant aquesta relativa homogeneïtat, hi ha diferències notables en la diversitat d’espècies que hom troba a les boscanes de les diferents regions del bioma. Generalment la diversitat és més gran a l’Amèrica del Nord que a Europa i encara ho és més a l’Àsia, sobretot a l’oriental. Prenent només els 10 gèneres d’arbres representats a les boscanes decídues per un major nombre d’espècies (Salix, amb més d’un centenar d’espècies; Acer, Quercus, Prunus i Magnolia, que ultrapassen la cinquantena, i Lithocarpus, Castanopsis, Betula, Populus i Ulmus que ultrapassen la trentena), resulta palès que les boscanes decídues de l’Àsia oriental són molt més riques en espècies que la resta, ja que s’hi troben més de 500 espècies d’arbres pertanyents a aquests 10 gèneres, mentre que a les americanes només n’hi ha una vuitantena de pertanyents a 8 d’aquests gèneres (Lithocarpus i Castanopsis en són absents) i a les europees una setantena pertanyents a 7 gèneres (en són absents Lithocarpus, Castanopsis i Magnolia). Fins i tot si es mira gènere per gènere la desproporció és gran: enfront de les 97 espècies de salzes (Salix) que creixen a les boscanes asiàtiques, només se’n fan 35 a les europees i 11 a les nord-americanes, i enfront de les 66 espècies d’aurons, blades i erables (Acer) i altres tantes de roures (Quercus) que hi ha a les primeres, se’n troben respectivament 9 i 18 a les europees i 10 i 37 a les nord-americanes. Només comptats gèneres, més aviat secundaris, com les càries (Carya) a l’Amèrica del Nord, o les moixeres i servers (Sorbus) a Europa, es troben més diversificats a les boscanes d’aquests continents que a les de l’extrem orient asiàtic.
La causa d’aquestes diferències rau probablement en l’orientació de les grans barreres muntanyoses d’Europa —sobretot els Pirineus i els Alps—, que s’allargassen d’E a W i que impediren la dispersió de la flora meridional cap al N en l’època postglacial. Les serralades de l’Amèrica del Nord i de l’Àsia oriental s’estenen més aviat de N a S i per aquesta raó no impediren que les plantes provinents de zones més meridionals recolonitzessin el bioma. A més, l’existència de grans valls fluvials contribuí a la dispersió. En alguns casos, fins i tot espècies subtropicals han aconseguit penetrar en àmbits climàtics que no són els seus i s’hi mantenen estrictament acantonades en enclavaments favorables.
Les fagàcies: els faigs, els roures i els castanyers
Les espècies d’arbres dominants dels boscos caducifolis temperats pertanyen totes a un petit nombre de famílies. La més remarcable de totes, atès el nombre d’espècies arbòries de les boscanes que hi pertanyen i les extensions que dominen, és la de les fagàcies, a la qual pertanyen els faigs típics (Fagus), els faigs australs (Nothofagus caducifolis), els roures (Quercus) i els castanyers (Castanea). Les fagàcies són plantes llenyoses, la majoria arborescents de fulles esparses i simples, quasi sempre decídues. Són característicament monoiques, és a dir, amb flors masculines i femenines separades però presents unes i altres en tots els peus, i es pol·linitzen anemofílicament, és a dir, que el pollen, molt lleuger, és transportat pel vent. Les llavors (fages, glans, castanyes) són grosses i riques en substàncies de reserva, aprofitables per molts animals dels boscos on viuen.
Els faigs típics (Fagus) no són un dels gèneres de fagàcies més nombrosos, però, en canvi, són importants pel caràcter dominant que tenen la majoria dels faigs a les boscanes on viuen. Així, la desena d’espècies de faigs que hi ha al món tenen totes un paper dominant, cadascuna a l’àrea on viu: el faig comú (F. sylvatica) a l’Europa central i occidental; el faig oriental (F. orientalis) dels Balcans al Caucas; el faig americà (F. grandifolia) a l’Amèrica del Nord; els faigs japonesos (F. crenata, F. japonica) al Japó, etc. A l’Amèrica austral hi ha diverses espècies del gènere Nothofagus, algunes de les quals són perennifòlies i característiques de les laurisilves valdivianes, però altres són de fulla caduca, com l’imposant raulí (N. procera) i el “roble” (N. obliqua) de les zones litorals del N del bosc valdivià, o com la lenga (N. pumilio) i el “ñirre” (N. antarctica) dels raiguers andins patagònics i de la Terra del Foc.
El faig comú (Fagus sylvatica) és un arbre remarcable. La seva dominància conspícua i el seu vigor competitiu no són igualats per cap altra espècie arbòria de fulla caduca de l’hemisferi nord. Això es deu en part al seu ampli espectre climàtic i edàfic, a la seva longevitat i a la capacitat competitiva que li proporciona la seva tolerància a l’ombra. En condicions òptimes pot arribar a assolir alçades de 40 m, tot i que les seves arrels solen romandre superficials. Les fulles són d’un verd brillant, amb una vora suaument ondulada i, quan són joves, un xic pubescents; a la tardor es tenyeixen amb tonalitats daurades i donen a l’arbre el seu aspecte més bell. Els fruits, les fages, s’alliberen en obrir-se la cúpula eriçada de punxes on s’apleguen de tres en tres; són aliment d’elecció de molts animals del bosc, però el faig només en produeix regularment cada any a la regió meridional de la seva àrea de distribució (més cap al N aquesta producció es limita als anys favorables).
Si no hi hagués hagut cap interferència humana d’ençà dels primers poblaments, el faig seria actualment l’espècie dominant a la major part de l’àrea de distribució dels boscos caducifolis europeus. Actualment, s’estén per quasi tota l’àrea europea del bioma tret de la sud-oriental, on el faig oriental (Fagus orientalis) ocupa el seu lloc, i de les regions europees de l’anomenada Comunitat d’Estats Independents, on és pràcticament absent. A les muntanyes de Crimea, és l’endèmic faig de Crimea (F. taurica) el que pobla les boscanes muntanyenques. El faig americà (F. grandifolia) fa el mateix paper a les boscanes de l’Amèrica del Nord, des d’Ontàrio fins al golf de Mèxic però amb preferència pels indrets frescals i humits, on esdevé fàcilment dominant, sobretot a la regió dels Grans Llacs. Al Japó, F. crenata i F. japonica es reparteixen el domini de les boscanes més extenses, el primer a les regions més septentrionals i a la zona central muntanyosa del país i el segon més al S o en cotes més baixes. A la Xina es dóna una situació equivalent amb F. englerana i F. longipetiolata.
Les espècies del gènere Quercus, nom que els romans donaven als roures, dominen la major part dels boscos caducifolis temperats, sobretot a l’hemisferi nord. Hi ha aproximadament 600 espècies d’aquest gènere a tot el món, des del S de Suècia i l’extrem oriental de Sibèria fins a Indonèsia i el Perú. Moltes d’aquestes espècies s’hibriden fàcilment entre elles, cosa que fa d’aquest gènere tot un repte per als taxònoms vegetals. Aquesta capacitat d’encreuament les fa molt adaptables i els ha permès colonitzar entorns molt diversos, entre ells les terres temperades on es fan les boscanes. Hi ha moltes espècies del gènere que són xeròfites i fins piròfites i, a les àrees més seques que colonitzen, per exemple a les mediterrànies, poden competir amb els pins gràcies al fet que posseeixen arrels profundes i escorces gruixudes resistents al foc. Per això, encara que el tronc mori, el sistema radicular sobreviu i la planta pot rebrotar. També hi ha espècies adaptades a condicions molt més humides, com ara les de planes inundables mal drenades i fins i tot d’aiguamolls. No totes són decídues, encara que les que sí que ho són, és a dir, els roures, constitueixen el grup de planifolis de fulla caduca de clima temperat amb més diversitat geogràfica i ecològica. Són arbres de dimensions considerables, d’escorça fosca i rugosa i capçada menys densa que la del faig; de fulles característicament sinuades, encara que amb grans variacions d’una espècie a una altra i fins i tot dins de cada espècie. Els seus fruits, els aglans, també són característics dels Quercus.
A Europa es troben una trentena d’espècies del gènere Quercus. Unes quantes, no decídues, es fan a la regió mediterrània i queden fora de l’abast del bioma de les boscanes, però n’hi ha almenys dues, el roure pènol (Q. robur) i el roure de fulla gran (Q. petraea), que són dominants a la major part de les boscanes decídues europees, tret de les regions més orientals del bioma. El roure cerris o roure turc (Q. cerris) es troba a gran part de la regió sud-oriental de l’àrea, des del SE de França fins a la costa septentrional de Turquia. El roure martinenc (Q. humilis [= Q. pubescens]), força més estès, està distribuït per bona part de la regió meridional de l’àrea, des de la Península Ibèrica fins a Anatòlia.
A l’Amèrica del Nord hi ha una major varietat d’espècies del gènere Quercus, encara que no totes decídues. Solament al Canadà i als Estats Units se’n poden comptar 60, i Mèxic, que en té aproximadament 115, és considerat el centre mundial de dispersió del gènere. Tot i que hi ha moltes espècies de roure, i també molta hibridació i variabilitat genètica entre elles, la totalitat dels roures de l’Amèrica del Nord es pot dividir en dos grans grups: els roures blancs i els roures rojos. Els roures blancs tenen fulles de vores arrodonides i glans de gust dolç, que maduren cada any i que tenen l’interior de la cúpula llis. El roure blanc americà (Q. alba), l’espècie més típica del grup, es pot trobar a tota la meitat oriental dels Estats Units, des dels Grans Llacs fins al golf de Mèxic. És un arbre alt que creix més de 30 m i el seu tronc pot arribar a tenir un diàmetre de més d’1 m. Treu branques a un nivell molt baix que formen una volta molt oberta, rodona i d’amplada impressionant. Els glans neixen directament dels branquillons, sense peduncle. No és pas una espècie exigent; ocupa terrenys molt diversos i de vegades pot formar boscos quasi purs. Als Apalatxes, hom troba una forma arbustiva d’aquesta espècie que viu a més de 1 500 m. Els roures rojos tenen fulles de lòbuls punxeguts i vores llises amb espines (com les de l’alzina de la conca mediterrània); llurs glans maduren cada dos anys, són amargants i tenen l’interior de la cúpula pubescent. Dins d’aquest grup, l’espècie més característica és el roure roig comú (Q. rubra), conegut a Europa amb el nom de roure americà, que avui creix en plantacions a les regions temperades d’arreu del món i es troba a latituds més septentrionals que el roure blanc. Penetra dins les províncies marítimes del Canadà i forma part del bosc caracterís-tic de la regió dels Grans Llacs. Altres roures comuns d’aquests boscos són, entre els blancs, el de glans grosses (Q. macrocarpa) i el roure “chinquapin” (Q. muehlenbergii), i entre els roures rojos, el negre (Q. velutina) i l’escarlata (Q. coccinea).
La major part de les seves espècies es troben a Àsia, però moltes creixen en àrees tropicals o subtropicals i no són caducifòlies. Tanmateix, roures com el “kunigi” (Quercus acutissima) i el “kashiwagi” o roure dàimio (Q. dentata) tenen un paper de primer ordre a les boscanes japoneses, mentre que altres, afavorits pels canvis en la coberta vegetal provocats per l’agricultura o els incendis, es poden considerar característics dels boscos secundaris; és el cas, per exemple, del roure mongol (Q. mongolica), que mostra preferència per les clarianes obertes en solells eixuts i carenes ventades.
Els castanyers (Castanea) són tots caducifolis. Són arbres d’escorça llisa i grisenca que poden assolir 35 m d’alt i 10 m de perímetre, amb una capçada cònica quan són joves i globular en els exemplars vells i amb fulles grans i lanceolades amb el marge serrat i una nervadura regular. Tenen una gran capacitat de regeneració a partir de la soca, fet que n’ha facilitat l’explotació en bagues i perxades. Els seus fruits, les castanyes, són comestibles i en alguns casos força estimats, tant pels humans com pels animals. El castanyer comú (C. sativa) s’estén pràcticament per tota l’àrea meridional de les boscanes decídues europees i per les anatòliques, fins al Caucas, mentre que en el seu límit septentrional arriba fins al S d’Anglaterra (on, com a tota la part més occidental de la seva àrea de distribució actual, es creu que fou introduït pels romans) i a diferents punts de l’Europa central. També a l’Àsia oriental creixen castanyers com ara C. seguinii a la Xina i C. crenata al Japó, sense comptar els falsos castanyers del gènere Castanopsis com el “shii” japonès (C. cuspidata). A l’Amèrica del Nord el castanyer americà (C. dentata) havia estat una espècie dominant, juntament amb els roures, a moltes boscanes dels Apalatxes i altres regions, però fou pràcticament eliminat pel xancre del castanyer, una malaltia introduïda.
Les betulàcies: els bedolls, els verns, els avellaners i els carpinus
Les betulàcies, la família que inclou bedolls, verns, avellaners i carpinus, estan molt emparentades amb les fagàcies, fins al punt que gairebé només es diferencien per característiques molt poc conspícues de les flors. Les betulàcies són, com les fagàcies, típicament monoiques. Bedolls i verns tenen una presència gens menystenible a les boscanes i sovint són dominants en determinades condicions ambientals extremes. Els avellaners figuren entre les espècies més importants del sotabosc arbustiu encara que, de fet, poden assolir dimensions de petits arbres. A les boscanes europees, per exemple, els bedolls (Betula) apareixen, bé a les comunitats de ribera, on s’hi fa el bedoll pubescent (B. pubescens), bé als límits de l’àrea de les boscanes amb els boscos muntanyencs de coníferes o amb la taigà boreal, on s’hi fa primordialment el bedoll comú (B. pendula). A les boscanes americanes els bedolls tenen una presència relativament més important, sobretot el bedoll groc (B. alleghaniensis), que es presenta sovint a les boscanes madures; el bedoll d’escorça de paper (B. papyrifera) i el bedoll gris (B. populifolia) apareixen més aviat en estadis inicials de la successió.
Els verns (Alnus) solen anar associats a sòls sotmesos a inundació, sigui a les riberes, sovint formant vernedes on són l’únic arbre dominant, sigui en sòls entollats o torberes. No és infreqüent que apareguin associats amb bedolls i freixes o fins i tot com a arbres del sotabosc d’algunes fagedes. El vern comú (A. glutinosa) és un arbre de capçada oblonga i escorça fosca i llisa, amb fulles de forma obovada característica, dentades i una mica viscoses. Pot fixar nitrogen gràcies a una relació simbiòtica amb un actinomicet. Forma vernedes a les riberes de les boscanes europees i fins i tot en l’àmbit mediterrani. El vern pilós (A. incana) és molt semblant (potser una mica més alt) però amb el revers de les fulles pubescent i, a diferència d’aquell, característic de les terrasses inferiors dels rius, és propi d’ambients montans i frescals. El vern manxú (A. mandshurica) forma part del sotabosc de les fagedes de la regió de l’Ussuri, a la frontera sino-russa.
Els avellaners, en particular l’avellaner europeu (Corylus avellana), són petits arbres d’entre 6 i 10 m d’alt que desenvolupen nombrosos rebrots a partir de la mateixa soca, cosa que els confereix un caràcter més aviat arbustiu. Tenen fulles grans, doblement dentades, amples, rugoses i arrodonides, amb un àpex punxegut. El seu fruit, l’avellana, és gustós i té un gran valor nutritiu. Molts animals de les boscanes cullen avellanes i se les mengen. Les nombroses branques de la planta, sovint amb l’ajuda de la mà dels humans, s’entrecreuen formant denses i impenetrables tanques vives només practicables pels animals de la boscana. A Anatòlia es fa l’avellaner turc (C. colurna), més gran que l’europeu, de port més arbori, amb una capçada piramidal i fruits també més grossos, i a la Xina l’avellaner xinès (C. chinensis), de dimensions veritablement arbòries, ja que pot assolir 40 m, i l’avellaner manxú (C. mandshurica), de distribució més septentrional. Tots tres produeixen fruits comestibles. Els avellaners americans (C. americana i C. rostrata) són similars als europeus, però la seva presència a les boscanes no és tan freqüent ni destacada.
Els carpinus (Carpinus) són, com els avellaners, petits arbres del sotabosc de rouredes i altres boscanes que, en algunes zones, poden arribar a competir en abundància amb els roures i que, ben il·luminats, poden assolir veritables dimensions arbòries. El carpinus comú (C. betulus) es troba a la major part de les boscanes decídues d’Europa, des de la França atlàntica i el SE d’Anglaterra fins al centre de Rússia i Anatòlia i des del S de Suècia fins als Apenins i els Balcans, però és absent a les boscanes de tot el N i l’W de les illes Britàniques i a les d’Ucraïna. Amb bones condicions d’il·luminació, per exemple a la vorada d’un bosc, pot arribar a fer 20 m d’alçada. Sol tenir una capçada densa i irregular, àmpliament cònica, i un tronc sinuós amb l’escorça d’un color gris platejat, finament ratllada i profundament acanalada. Les fulles, alternes, són semblants a les de l’avellaner però més allargades, ovato-oblongues. Els aments, tant masculins com femenins, pengen a l’extremitat de branques diferents i els fruits, en madurar, es presenten en raïms penjants de 7 o 8 parells de núcules protegides per unes bràctees verdes que semblen petites fulles. Al SE d’Europa i a Anatòlia es fa el carpinus oriental (C. orientalis), més petit i generalment arbustiu; a l’Àsia oriental creixen, entre altres, el carpinus manxú (C. cordata) i el japonès (C. japonica), mentre que a l’Amèrica del Nord la presència del gènere és menys significativa, pràcticament limitada al carpinus de Carolina (C. caroliniana).
Les platanàcies i les cercidifil·làcies: els plàtans i els katsures
Una altra família relacionada amb fagàcies i betulàcies són les platanàcies, amb un únic gènere al qual pertanyen els plàtans (Platanus), amb representants a les boscanes de l’Europa oriental i Anatòlia (P. orientalis) i a les de l’Amèrica del Nord (P. occidentalis). Són arbres que poden superar els 30 m d’alt, de tronc dret i llarg que es ramifica força amunt, amb una escorça llisa que es va desprenent a grans plaques donant-li un aspecte característic i una gran varietat de colors, del gris al groc verdós passant pels torrats. La fulla és gran, palmada i de forma característica, de color verd intens a la primavera i estiu i bronzejat a la tardor. El plàtan comú (P. x hybrida), àmpliament cultivat com a arbre de carrer a l’Europa occidental i meridional, és un híbrid de plàtan oriental (P. orientalis) i plàtan americà (P. occidentalis).
El katsura (Cercidiphyllum japonicum) és l’únic representant vivent de la família de les cercidifil·làcies, també afí a la de les fagàcies encara que amb caràcters que semblen relacionar-la amb les magnoliàcies i altres famílies d’angiospermes primitives. Té els dos sexes separats en peus diferents però en uns i altres les flors se situen als nusos de les branques. Són arbres alts, fins d’uns 25 o 30 m, d’aspecte gràcil i delicat, amb troncs d’escorça clara i fissurada i capçades esclarissades de branques primes i arquejades i fulles arrodonides que, quan surten a la primavera, són d’un color rosat brillant, a l’estiu d’un verd profund pel dret i blavisses pel revers i a la tardor de colors molt variats.
Les magnoliàcies: les magnòlies caducifòlies i els tulipers
La família de les magnoliàcies és considerada per molts autors com la més primitiva de totes les famílies d’angiospermes, en particular per la morfologia de les flors, que disposen les seves nombroses peces helicoïdalment entorn de l’eix floral, força llarg. Té una àmplia distribució per les regions càlides i temperades d’Amèrica i d’Àsia, i només una part dels seus representants són caducifolis, entre altres unes quantes magnòlies (Magnolia), si bé a les zones més meridionals de les seves àrees respectives algunes de les espècies caducifòlies no perden la fulla fins que no ha aparegut el nou fullam. En general, les magnòlies tenen fulles grans i lluents i flors blanques o de colors pàl·lids, i produeixen un fruit peculiar semblant a una pinya feta de folíols endurits entre els quals sobresurten lleugerament les llavors de color carmí. Les espècies de magnoliàcies caducifòlies més característiques de les boscanes americanes són l’arbre de cogombres (M. acuminata), que rep aquest nom a causa de la forma dels seus fruits, molt semblant a la dels petits cogombres als quals són tan afeccionats els nord-americans, i que es troba des de les pinedes de la costa SE dels Estats Units fins als Grans Llacs, la magnòlia de paraigües (M. tripetala) i la magnòlia de fulla gran (M. macrophylla).
El tuliper de Virgínia (Liriodendron tulipifera) és una magnoliàcia caducifòlia de les boscanes decídues americanes que es caracteritza per ser una espècie pionera molt agressiva, amb prou longevitat per a compartir una posició dominant en estadis avançats de successió dels boscos mesofítics. Un paper equivalent té a les boscanes xineses més meridionals el tuliper xinès (L. chinense).
Les salicàcies, les ulmàcies i les tiliàcies: els salzes, els oms i els tells
Les salicàcies i les ulmàcies solen anar associades als medis més humits, en particular els riberencs, i tenen un paper important en tots els boscos de ribera i indrets humits de les regions temperades. A la família de les salicàcies pertanyen salzes (Salix), d’una banda, i pollancres i trèmols (Populus), d’una altra; a la de les ulmàcies, oms (Ulmus), lledoners (Celtis) i zelkoves o falsos oms (Zelkova).
El gènere Salix és el més diversificat a totes les boscanes del món, encara que la majoria dels seus representants no són arbres de gran port, sinó més aviat arbrissons (o fins i tot arbustos, com en el cas de les sargues) de branques i llucs llargs, fins i flexibles, que els han fet molt apreciats per a treballs de cistelleria. Hi pertanyen els salzes pròpiament dits i les vimeteres, propis de les riberes, i els gatsaules, propis de clarianes i altres ambients oberts del bioma. Entre els salzes pròpiament dits es poden esmentar el salze blanc (S. alba) i el salze cendrós (S. cinerea) de les boscanes atlàntiques europees, o el salze negre (S. nigra) de les nord-americanes; entre les vimeteres, la vimetera comuna (S. fragilis) o la vimetera blanca (S. viminalis) de les boscanes europees, i entre els gatsaules, el gatsaule comú europeu (S. caprea).
De més port que els salzes són els pollancres i els trèmols (Populus), tots ells grans arbres de creixement ràpid i fusta blana, però mentre que els pollancres, com el pollancre ver (P. nigra) europeu o la carolina (P. deltoides) nord-americana, són arbres de ribera igual que els salzes, els trèmols, com el trèmol (P. tremula) eurasiàtic o el trèmol americà (P. tremuloides), són propis de clarianes o de boscanes molt obertes igual que els gatsaules. La majoria són espècies dioiques, és a dir amb peus masculins i femenins separats, amb flors masculines i femenines en ament que es desclouen abans que apareguin les fulles. Aquestes, que solen tenir el limbe tan ample com llarg, són de formes i grandàries molt variades; si bé la forma més general per als pollancres sol ser ampla a la base i apuntada a l’àpex, els trèmols acostumen a tenir les fulles orbiculars i fistonades.
Els oms són arbres esvelts, de capçada poc densa, amb fulles caduques relativament petites i característicament asimètriques a la base. També és molt característic el fruit, que és una sàmara, és a dir, un fruit dotat d’un pericarp membranós que ajuda a la dispersió de les llavors pel vent (anemocòria). En el cas dels oms la sàmara és com una làmina fina i lleugera, de color verdós, que duu al seu centre la grana. Creixen sobretot en llocs humits i frescals, sense estar necessàriament vinculats als boscos de ribera; quan les omedes creixen al llarg del curs d’un riu sempre ho fan a la banda més externa i més allunyada de l’aigua. L’om comú (Ulmus minor) és propi de les àrees europees meridionals del bioma, des del SE d’Anglaterra fins al Caucas, mentre que l’om pedunculat (U. laevis) viu a gran part de l’Europa central i oriental de la regió però no a l’Europa atlàntica ni a la Península Ibèrica; l’oma (U. glabra) era comuna a totes les boscanes europees tret de les de la zona sud-oriental. Tant l’om americà (U. americana) com el roig (U. rubra) estan molt estesos per totes les boscanes nord-americanes i fins i tot excedeixen l’àrea del bioma al llarg dels cursos d’aigua. Oms europeus i americans han estat dramàticament delmats al llarg de gran part del segle XX (d’ençà del 1917 a Europa i des del 1930 a l’Amèrica del Nord) per la grafiosi o mal holandès, una malaltia provocada pel fong ascomicet d’origen asiàtic Ceratocystis ulmi.
Els lledoners (Celtis) s’assemblen molt als oms però tenen la fulla més llarga i més ampla a la base, d’una forma semblant a la de les ortigues, fet que els ha valgut el nom anglès de “nettle trees” o arbres ortiga, i un fruit en drupa, semblant a una petita cirera de la mida d’un pèsol. El lledoner comú (C. australis), originari de les boscanes balcàniques i anatòliques, ha estat molt difós com a arbre cultivat i sovint subspontani a tota l’Europa meridional, fins i tot en indrets frescals de la conca mediterrània, i és l’únic representant del gènere a les boscanes europees. A les americanes n’hi ha mitja dotzena, entre els quals destaquen el lledoner de rossí (C. occidentalis), que creix bàsicament a la meitat septentrional de l’E dels Estats Units, des de Tennessee i el N d’Arkansas i Oklahoma cap al N, i també es cultiva de vegades en jardins i parcs d’Europa occidental, i el lledoner dolç (C. laevigata), que es fa més aviat a la meitat S de l’E dels Estats Units, des del SE de Virgínia i el centre d’Illinois fins a Florida i la costa texana.
Les zelkoves o falsos oms (Zelkova) s’assemblen als oms per la fulla, però tenen un fruit que s’assembla al del lledoner, bé que encara és més petit. Tenen el seu centre de diferenciació a les boscanes decídues de l’Àsia oriental, sobretot a les xineses, però hi ha un representant del gènere, la zelkova del Caucas (Z. carpinifolia), a les boscanes caucàsiques i hircanianes. El “keyaki” o zelkova japonesa (Z. serrata) és comú a les fagedes de la part septentrional de Honsh¯u i l’extrem meridional de Hokkaidō, al Japó.
Afí a les salicàcies i les ulmàcies és la família de les tiliàcies, a la qual pertanyen els tells (Tilia). Són arbres caducifolis que poden arribar a 25 m d’alçada, de capçades altes i arrodonides i fulles en forma de cor i de marge serrat. Les flors, poc acolorides però de fragància característica, són hermafrodites i generalment aplegades en petites inflorescències que de vegades, com en el cas del tell de fulla gran (T. platyphyllos), surten al mig d’una bràctea d’un verd pàl·lid. El tell de fulla gran es troba a la major part de la regió submediterrània i a les zones meridionals medioeuropees. El tell de fulla petita (T. cordata) s’estén, bé que localitzat, per tota l’àrea de les boscanes decídues europees tret de les de Turquia, Grècia i el S d’Itàlia. El tell argentat (T. tomentosa) està confinat al SE, des d’Eslovàquia, Hongria i Sèrbia fins a Turquia i Ucraïna. El tell americà (T. americana) i el blanc (T. heterophylla) es troben a les boscanes de roure i cària i a les complexes dels Apalatxes.
Les juglandàcies i les hipocastanàcies: les nogueres i els castanyers d’Índia
Entre les famílies amb gèneres importants a les boscanes figura també la de les juglandàcies, a la qual pertanyen nogueres (Juglans), càries (Carya) i falsos freixes (Pterocarya), i la de les hipocastanàcies, a la qual pertanyen els castanyers d’Índia (Aesculus).
De nogueres (Juglans) se’n troben a les boscanes de tots els continents, encara que no solen ser arbres dominants. Són arbres robustos i de capçada ampla, que sovint poden assolir dimensions considerables, amb fulles compostes imparipinnades, de folíols grans i llisos, i fruits grans i comestibles. A les boscanes de terra baixa del SE d’Europa i d’Anatòlia, la noguera vera (J. regia) sol viure a les fagedes però és un arbre que, a causa del seu fruit apreciat, fou introduït a tota la resta d’Europa des de molt antic i es troba a molts indrets, tant cultivat com subspontani. A les boscanes de l’Àsia oriental s’hi fan la noguera manxú (J. mandshurica) i la noguera japonesa (J. ailanthifolia), i a les americanes la noguera negra (J. nigra). Les càries (Carya) són, com els roures, un grup d’arbres de gran amplitud ecològica però, a diferència d’aquells, estan restringits geogràficament a l’Amèrica del Nord excepte alguna espècie, com la cària de Tonquín (C. tonkinensis), de l’Àsia oriental. Són parentes properes de les nogueres (Juglans); tenen, com elles, fulles compostes i produeixen una nou generalment comestible. Es troben sobretot en indrets secs, associades amb roures, però la diferenciació genètica ha donat lloc a espècies adaptades a hàbitats molt diversos. Tant la cària amarga (C. cordiformis), com la cària vera (C. glabra) o la cària blanca (C. ovata) tenen una distribució molt àmplia a l’Amèrica del Nord, tant a les boscanes decídues pròpiament com fora. Altres, com la cària aquàtica (C. aquatica), tenen una distribució més restringida, especialment cap al S. Un altre grup d’espècies, entre altres el pacaner o cària d’Illinois (C. illinoensis), molt apreciat per la seva nou, penetra fins al Mèxic subtropical. A l’inrevés de les càries, els falsos freixes (Pterocarya), també anomenats de vegades càries alades, són exclusivament asiàtics, sobretot de les boscanes de l’Àsia oriental, encara que el fals freixe del Caucas (P. fraxinifolia) és un endemisme de les boscanes caucàsiques.
Els castanyers d’Índia (Aesculus) han rebut aquest nom a causa de la procedència oriental del castanyer d’Índia ver (A. hippocastanum), el més corrent en jardins i parcs europeus, originari dels Balcans i les muntanyes de l’Àsia occidental fins a l’Himàlaia però ja conegut d’antic a l’Europa occidental. Tanmateix, el centre de diversificació dels castanyers d’Índia sembla que és més aviat a l’Amèrica del Nord, on es coneixen una vintena d’espècies d’arbres i arbustos del gènere entre les quals destaca el castanyer d’Índia groc (A. octandra). També n’hi ha a l’Àsia oriental, com el castanyer d’Índia japonès o “tochinoki” (A. turbinata). Són arbres d’uns 20 m d’alt, d’escorça fosca i capçada alta, densa i en forma de cúpula, de fulles oposades i compostes, amb els folíols (que són de 5 a 7 i recorden les fulles del castanyer) palmats i de grandària creixent des dels més externs fins al central. Les flors, amb els sexes separats, s’apleguen en inflorescències vistoses amb flors masculines al capdamunt i femenines a la base. Els fruits, semblants a les castanyes però més arrodonits, són amargants i no comestibles.
Les aceràcies: els aurons i els erables
A la família de les aceràcies pertanyen algunes de les espècies de coloració tardoral més espectacular de les boscanes decídues de tots els continents, però molt especialment de les boscanes mixtes de la regió oriental d’Àsia i de l’Amèrica del Nord. Aurons, blades i erables s’inclouen en el gènere Acer, que constitueix ell tot sol gairebé la totalitat de la família (només dues espècies d’arbres caducifolis de la Xina central i meridional es classifiquen en un segon gènere d’aceràcies). El centre de diferenciació del gènere se situa a les boscanes de l’Àsia oriental (prop d’un centenar d’espècies, sense comptar les perennifòlies de les selves temperades i tropicals del S de la Xina) i, encara que generalment no ocupen una posició dominant, a la tardor no passa desapercebuda la contribució paisatgística dels canvis de color de les seves capçades. Alguns, els anomenats aurons d’escorça de serp, són remarcables també per la peculiar coloració de les seves escorces, verdoses i ratllades longitudinalment de blanc, argentat o taronja, com l’auró del pare David (A. davidii) de la Xina o l’auró roig d’escorça de serp (A. capillipes) del Japó. Entre les espècies d’auró més característiques d’aquestes boscanes figuren l’auró de Her (A. hersii), l’auró d’escorça de paper (A. griseum) i l’auró coreà (A. pseudosieboldianum).
Tret tal vegada de l’erable de sucre (Acer saccharum), que, a la regió dels Grans Llacs, pot ser codominant amb el faig americà (Fagus grandifolia), els aurons i erables també solen ocupar una posició subordinada a les boscanes nord-americanes, però la seva aportació colorista a la imatge tardoral d’aquestes boscanes no és menys remarcable que a les asiàtiques. A les boscanes americanes, l’erable de sucre és una espècie de gran longevitat, molt tolerant a les condicions de vida del sotabosc i que, en indrets frescals i humits, pot aparèixer en qualsevol estadi de la successió, mentre que l’erable argentat (A. saccharinum) prefereix els indrets inundables.
És als boscos de la regió dels Grans Llacs, ben coneguts pels colors vius que presenten a la tardor, on els aurons americans es manifesten amb la seva paleta més variada. Un viatge en tren des de Montreal fins a Nova York vorejant el riu Hudson és una excel·lent manera d’apreciar aquesta varietat. A banda del faig americà i de l’erable de sucre, que són codominants, en aquesta regió es troben, entre altres, l’auró roig (Acer rubrum) i l’auró ratllat de Pennsilvània (A. pensylvanicum), a més de diversos roures.
A les boscanes europees la varietat no és tan gran, bé que creix cap al SE i torna a ser considerable al Caucas. L’erable o auró noruec (A. platanoides) creix a la major part de les boscanes decídues europees tret de les àrees nord-occidentals (el NW de França, illes Britàniques i Benelux) i també es troba a les boscanes euxíniques i caucàsiques; el plàtan fals (A. pseudoplatanus) té una àrea de distribució semblant però que inclou també l’Europa nord-occidental; l’auró blanc (A. campestre), tot i estar també molt estès per les boscanes europees, no és tan comú. L’auró insigne (A. insigne) és una espècie pròpia de les fagedes caucàsiques.
Les rosàcies: les moixeres i els fruiters de pinyol
A la família de les rosàcies pertanyen una colla d’arbres i arbustos de les boscanes decídues els fruits carnosos dels quals, que són un dels aliments preferits de molts ocells i mamífers, han desvetllat també de molt antic l’interès dels humans per les seves qualitats organolèptiques i alimentàries. Per això molts d’aquests arbres de la família de les rosàcies han estat cultivats durant tants segles i els seus límits naturals són avui impossibles de determinar. La majoria dels fruiters de fruita dolça que abasteixen els mercats de les regions temperades, en efecte, són originaris de les boscanes, principalment de les asiàtiques, on figuren entre les espècies més significatives del sotabosc. Els gèneres Malus (les pomeres), Pyrus (les pereres) i Prunus (al qual pertanyen totes les fruites de pinyol: pruneres, cirerers, albercoquers, presseguers, etc.) concentren la majoria dels arbres o arbustos de la família de les rosàcies del sotabosc de les boscanes.
Les pereres silvestres (Pyrus) són presents sobretot a les boscanes submediterrànies, anatòliques i caucàsiques, de les quals és originària la perera comuna (P. communis); a les boscanes de l’Àsia oriental s’hi troba la perera japonesa o “nasi” (P. serotina). Tot i que les pomeres silvestres són més nombroses i diverses a les boscanes xineses i a les muntanyes de l’interior d’Àsia, també se’n fan a les boscanes dels Balcans, Anatòlia i el Caucas, moltes transformades pel conreu. Els fruiters de pinyol (Prunus) es troben a totes les boscanes però amb un gran centre de diferenciació a l’Àsia oriental (d’un total de 400 espècies només 17 —i no totes de port arbori— són europees) i una bona varietat a les boscanes de l’Amèrica del Nord. Tampoc no són exclusius d’aquest bioma; hi ha espècies d’aquest gènere a les mediterrànies i a les selves temperades. De les boscanes xineses són originaris el presseguer (P. persica), l’albercoquer (P. armeniaca) i molts altres de menor difusió.
També són rosàcies les moixeres i els servers (Sorbus), de fruits menys apreciats però que comparteixen amb els aurons algunes de les coloracions tardorals més espectaculars del món dels caducifolis. Les moixeres tenen fulles de color contrastat entre anvers i revers i les flors disposades en inflorescències corimbiformes, que tot i ser força vistoses no ho són tant com els seus fruits, de colors molt vius. La moixera comuna (S. aria) i la moixera de pastor (S. torminalis) tenen la fulla simple, la primera ovada i la segona lobulada, d’una forma que recorda la d’alguns aurons; la majoria de moixeres, però, tenen la fulla composta, pinnada. La moixera i la moixera de pastor són freqüents a les boscanes europees i la segona estén la seva àrea de distribució fins al Caucas. La moixera de guilla (S. aucuparia) a Europa, la moixera americana (S. americana) a l’Amèrica del Nord i la moixera de l’Amur (S. amurensis) i la japonesa (S. commixta) a l’Àsia oriental, són més muntanyenques i només davallen a la plana al N de l’àrea de distribució de les boscanes, allà on entra en contacte amb els boscos boreals.
Les oleàcies: els freixes
Finalment cal fer referència a la família de les oleàcies, present a les boscanes tant del Vell com del Nou Món, a la qual pertanyen els freixes (Fraxinus) i també diferents gèneres d’arbustos i lianes, com forsíties (Forsythia), gessamins (Jasminum) i lilàs (Syringa).
Els freixes són arbres grans, que poden ultrapassar els 20 m d’alçada de tronc dret, amb l’escorça grisa i capçada alta, en forma de cúpula irregular, amb fulles imparipinnades, amb 5 a 11 folíols sèssils de forma lanceolada i marge dentat. Sense ser pròpiament de ribera, són arbres més aviat higròfils, encara que són freqüents pels riberals cap al S de les seves àrees de distribució. El freixe de fulla gran (Fraxinus excelsior) creix a totes les boscanes decídues europees, tret de les de l’extrem nord-occidental (Escòcia i N d’Irlanda) i a les anatòliques. El freixe de flor o freixe del mannà (F. ornus) es troba a la meitat oriental de la regió submediterrània; excepcionalment es localitza de forma aïllada a la muntanya mediterrània, concretament al S del País Valencià, que representa l’extrem occidental de la seva àrea de distribució. A les boscanes americanes es fan el freixe comú (F. nigra), el blanc (F. americana) i el roig (F. pennsylvanica). A les de l’orient asiàtic destaquen el freixe de fulla ampla (F. rhynchophylla) i el freixe manxú (F. mandschurica).
L’esclarissada diversitat del sotabosc
Pel que fa al nombre d’espècies, el sotabosc dels boscos caducifolis temperats és molt més divers que l’estrat arbori. La presència i el funcionalisme del sotabosc depenen, en gran mesura, de si el cobricel de les capçades dels arbres permet o no el pas de la llum, fet que determina quines espècies hi poden prosperar. Les espècies esciòfiles o d’ombra necessiten una ombra densa i creixen sota la volta ombrívola de les capçades dels grans arbres, els quals de joves també són esciòfils. Les plantes heliòfiles necessiten claror i busquen les clarianes assolellades que els arbres caiguts hagin pogut deixar o bé les ombres més precàries d’arbres, com per exemple els roures, que a l’estiu són menys ombrívols que els faigs.
L’aparença canviant dels arbustos i de les herbes
A l’estrat arbustiu, arbustos i arbrissons ocupen diferents nínxols ecològics i afegeixen encara més varietat a les formacions boscoses; només les fagedes solen tenir un sotabosc pobre en arbustos en el qual de vegades únicament els nabius (Vaccinium) arriben a ultrapassar el port herbaci. L’estrat herbaci és encara més ric, divers i complex que el sotabosc arbustiu i és constituït per un nombre molt elevat de petites plantes que han desenvolupat diferents estratègies per sobreviure (vegeu també vol. 4, pàg. 98): teròfits que moren cada any i deixen llavors que germinen uns mesos més tard; geòfits que mantenen viva una part de la planta sota terra, generalment un bulb o rizoma; hemicriptòfits que preserven algunes parts externes com la tija o les fulles unides a un sistema radicular molt desenvolupat que sobreviu sota terra. Cada any, la part aèria de la planta reapareix i, sovint, com s’esdevé a les fagedes de l’Europa central, és aquest estrat herbaci vernal, divers encara que fugaç, el que separa un tipus de bosc d’un altre encara que l’espècie dominant sigui la mateixa. De les plantes que integren aquest estrat herbaci se’n poden destacar la família de les poàcies o gramínies, la representada per un nombre més crescut d’espècies, i la de les orquidàcies per la seva vistositat, que a la primavera i l’estiu ajuda, amb un toc de color espectacular, a embellir encara més les boscanes.
Moltes plantes floreixen abans que els arbres i els arbustos que hi ha per sobre seu hagin tret la fulla. Aquesta és l’anomenada estratègia pre-vernal i l’empren les plantes que necessiten créixer a ple sol, cosa que només poden fer abans que el fullam tanqui el cobricel de la boscana. A les boscanes de l’Europa atlàntica, moltes espècies precoces com la primavera (Primula vulgaris), la maduixera (Fragaria vesca), diverses violes (Viola), el buixol (Anemone nemorosa) i l’heura de terra (Glechoma hederacea) floreixen fins i tot abans no comenci la primavera. Aquestes espècies assenyalen el despertar del bosc després de la seva llarga letàrgia hivernal i formen una catifa vivament acolorida. Sovint apareixen tímidament entremig de la darrera neu convidant els primers insectes que gosen fer sentir el seu brunzit a gaudir del seu nèctar, i els caminants a fruir de la seva bellesa i el seu acoloriment.
Altres espècies són típicament de primavera o vernals. Floreixen justament quan els arbres i els arbustos comencen a treure les primeres fulles. Precisament el moment en què el cobricel es va tancant de manera progressiva pel creixement del fullam dels arbres i va limitant la llum que penetra a l’interior del bosc. Són característicament vernals les veròniques (Veronica), la lleteresa de bosc (Euphorbia amygdaloides), la rapa tacada (Arum maculatum), la búgula (Ajuga reptans), l’escil·la atlàntica (Hyacinthoides non-scripta) i moltes altres no tan comunes.
Finalment, hi ha un tercer grup de plantes herbàcies, les estivals, que s’han adaptat a florir precisament a l’estiu, quan la quantitat de llum que arriba a la superfície és severament limitada pel fullam ja ben desenvolupat d’arbres i arbustos. Entre aquestes plantes estivals es poden destacar l’ortiga pudent (Stachys sylvatica), la campaneta traqueli (Campanula trachelium), l’herba de les encantades (Circaea lutetiana), la didalera purpúria (Digitalis purpurea) i moltes altres.
A aquesta catifa de plantes vasculars o superiors, s’hi pot afegir una capa de molses, líquens i fongs que, sobretot a les àrees més humides, fan augmentar considerablement el nombre total d’espècies presents.
Les rosàcies i les araliàcies: els esbarzers i els bastons del diable
Entre els arbustos de les boscanes destaquen els de dues famílies, les rosàcies i les araliàcies, que es caracteritzen, entre altres coses, per tenir nombrosos representants amb espines, agullons i altres estructures vulnerants. No és, tanmateix, un caràcter general en aquestes famílies, que també tenen representants arbustius o lianoides inermes i fins i tot herbacis.
Entre els arbustos heliòfils de les boscanes decídues d’arreu del món destaca precisament una profusió de rosàcies arbustives del gènere Crataegus, que inclou els arços, els espinalbs i els atzerolers. El seu centre de diferenciació se situa a les boscanes nord-americanes, però a Europa i a l’Àsia oriental també hi ha algunes espècies importants d’aquests arbustos caducifolis. L’espècie més freqüent a les boscanes europees és l’arç blanc (C. monogyna), que és molt important des del punt de vista ecològic, ja que els seus fruits, rics en vitamina C, són l’aliment de molts ocells i d’altres animals durant la tardor i l’hivern. D’altra banda, les seves espines punxegudes són una protecció excel·lent per als ocells que hi nien. Les seves flors, com les de totes les rosàcies, nodreixen tota una multitud d’insectes a la primavera i a l’estiu. Els arços han estat respectats i utilitzats pels humans des de temps immemorials amb l’objecte de fer-ne tanques vives que avui fan inconfusibles moltes àrees, en altre temps forestals, d’arreu d’Europa. L’aparició de les seves flors marca el començament de la primavera; d’aquí ve el nom de “may” (‘maig’, el mes en què l’arç floreix a la Gran Bretanya) amb què es coneix aquest arbust en anglès. Els arços, juntament amb els avellaners, també predominen als sotaboscos de les boscanes de l’Àsia oriental; els més importants són l’arç de fulla gran (C. pinnatifida), que dóna un fruit voluminós, lleugerament piriforme, de color vermell brillant amb punts blancs, i l’arç laciniat (C. laciniata), de fulles profundament lobades, grisenques i cotonoses, que dóna un fruit de color taronja. A l’Amèrica del Nord, entre les nombroses espècies d’arços destaquen l’arç escarlata (C. pedicellata), que és molt fàcil de reconèixer per les nombroses flors de cada un dels seus raïms, de distribució més aviat septentrional; l’arç gebrat (C. pruinosa) i l’arç d’esperó (C. crus-galli), difosos per tota l’àrea de les boscanes nord-americanes; i, als sectors més meridionals tres espècies importants: l’arç espatulat o “littlehip hawthorn” (C. spathulata), que té les fulles trilobades i profundament dentades; l’arç nabiu (C. brachycantha), distribuït més aviat per l’interior del continent; i l’arç julivert (C. marshallii), el nom del qual deriva de les seves fulles inconfusibles i que pot créixer, al contrari que les espècies precedents, tot al llarg de la costa sud-oriental dels Estats Units.
Els arços no són les úniques rosàcies del sotabosc de les boscanes; esbarzers (Rubus), rosers boscans (Rosa), alguns fruiters de pinyol de petites dimensions com l’aranyoner (Prunus spinosa) o plantes herbàcies com les maduixeres (Fragaria) o l’espirea de jardí (Spiraea cantonensis) són també rosàcies. Els esbarzers són de les rosàcies més esteses i conegudes arreu de les boscanes decídues (i també fora, ja que s’estenen pels biomes veïns allà on hi ha contacte entre bosc i espai obert i prou humitat), on es presenten associats amb altres arbustos i lianes més o menys heliòfils del sotabosc. Sarmentosos, de ràpid creixement i coberts d’agullons, formen densos poblaments, gairebé impenetrables: les bardisses. Igualment carregades d’agullons són les branques dels rosers boscans o gavarreres, però les seves flors són més grans i aromàtiques que les dels esbarzers i els fruits respectius no s’assemblen de res. Els esbarzers tenen un fruit globós, la móra, format de fet per una colla de petites drupetes esfèriques aglomerades en un fruit col·lectiu; els rosers, en canvi, fan un fruit piriforme que en realitat és el receptacle carnós i acolorit que suporta uns fruitets (aquenis) poc conspicus i coberts de setes rígides.
No tots els fruiters de pinyol, ni les pomeres i pereres són arbres. Alguns, com l’aranyoner de les boscanes europees, el cirerer de Virgínia (Prunus virginiana), la pomera borda blanca (Malus angustifolia) o la pomera borda dolça (M. coronaria) són arbustos o, com a molt, petits arbres del sotabosc arbustiu de les boscanes. El mateix passa amb la majoria dels corners (Amelanchier), arbustos alts, de branques llargues i primes, fulles escasses i flors molt vistoses de pètals llargs i estrets; el corner europeu (A. ovalis) es troba a les rouredes medioeuropees i submediterrànies en companyia de gavarreres i de boixos; el corner americà comú (A. lamarckii [= A. laevis]) viu al sotabosc de boscanes molt diverses dels Apalatxes fins a Terranova; el corner arbori o “downy serviceberry” (A. arborea) té fulles més grans que el seu parent, fruits més secs i és un component comú del sotabosc de les boscanes de roure dels Apalatxes, encara que la seva àrea de distribució és força dilatada.
Ben diferent de la de les rosàcies és la família de les araliàcies, predominantment tropical però ben representada a les boscanes asiàtiques i americanes i, en molt menor grau, a les europees. A les boscanes asiàtiques viuen nombroses araliàcies arbustives del gènere Aralia, les aràlies, molt diferents de les seves parentes europees, les heures (Hedera). Les aràlies són arbustos caducifolis vigorosos i amb nombroses espines, amb les flors i els fruits sovint negres, aplegats en umbel·les semblants a les de les heures. Tenen fulles compostes, de vegades bipinnades i fins i tot tripinnades, d’un verd grisenc fosc, amb folíols ovalats i mucronats. L’aràlia manxú (A. elata), coneguda també com a bastó del diable per les seves branques nuoses i espinoses, és una espècie comuna que creix habitualment fins a 15 m d’alçada en condicions favorables i es cultiva com a ornamental a totes les regions temperades, igual que l’aràlia japonesa o “udo” (A. cordata). També pertanyen a la família de les araliàcies l’enfiladís ginseng siberià (Eleutherococcus [= Acanthopanax] senticosus) i el ginseng ver (Panax ginseng). Les araliàcies estan gairebé tan ben representades a l’Amèrica del Nord com a l’Àsia oriental (40 espècies d’aràlies entre les dues regions, mentre que a Europa no n’hi ha cap). El bastó del diable comú (Aralia spinosa) és un arbust que deu el nom a les seves branques, que són d’allò més espinoses i semblants a les de les seves congèneres asiàtiques i que, com aquestes, produeix umbel·les de fruits negres. Fins i tot s’hi fa el ginseng americà (Panax quinquefolius), succedani de l’asiàtic i objecte com aquell de conreu i d’un comerç força actiu.
Les cornàcies i les caprifoliàcies: els sanguinyols, els saücs i els xuclamels
Cornàcies i caprifoliàcies són dues famílies gairebé cosmopolites, però amb una sòlida representació a les boscanes decídues de tots els continents. Les cornàcies són representades principalment pels sanguinyols (Cornus), particularment abundants a les boscanes de l’Àsia oriental però presents també a l’Amèrica del Nord i a Europa, i que deuen el seu nom a la particular coloració vermella, com de sang, que prenen els brots joves. Són arbustos de fulles simples, oposades, amb les flors petites aplegades en inflorescències força denses. Cornàcies com el sanguinyol comú (C. sanguinea) o el corneller mascle (C. mas) són corrents a les boscanes europees. El kousa (C. kousa), que creix al Japó i a Corea, pot assolir 7 m d’alçada i presenta branques ben dretes; a la tardor, les seves fulles es tornen d’un carmí bronzejat i produeix uns fruits semblants a les maduixes. El sanguinyol gegant xinès (C. controversa), que pot assolir un port arbori (més de 20 m d’alt), és el representant més gran del gènere i presenta branques horitzontals escalonades. El sanguinyol xinès comú (C. chinensis) té una talla intermèdia entre les dues espècies precedents i un port més obert que el del kousa. El sanguinyol florit (C. florida) pot assolir 15 m d’alçada. A l’Amèrica del Nord el sanguinyol alternifoli (C. alternifolia) es fa fàcilment en sòls humits, mentre el sanguinyol de vímets (C. stolonifera) viu pràcticament a tot el subcontinent nord-americà.
Tot i que poc nombrosa, la família de les caprifoliàcies està força diversificada a les regions temperades i comprèn tant arbustos com lianes àmpliament distribuïts a tots els biomes temperats i, en particular, a les boscanes decídues. Entre els arbustos destaquen els saücs (Sambucus) i els viburns (Viburnum), i entre les lianes diferents xuclamels o lligaboscos (Lonicera), alguns dels quals també són arbustius.
Els saücs són arbustos o petits arbres que es fan principalment en llocs humits de fondalades o vores de rierols. Fan unes grans fulles compostes de 5 a 7 folíols lanceolats i de marges dentats, i uns corimbes de flors blanques que deixen pas a uns fruits en drupa ben negres (o, si més no, de colors foscos) quan són madurs; les branques solen tenir una medul·la blanca i blana. Estan particularment ben representats a les boscanes nord-americanes, ja que de la trentena d’espècies que hi ha al món, 12 creixen en aquesta àrea; el més estès és el saüc americà comú (Sambucus canadensis). El saüc europeu comú (S. nigra) es fa a les boscanes higròfiles de tot Europa. A les clarianes de moltes fagedes s’hi fa el saüc racemós (S. racemosa).
Entre els viburns hi ha tant espècies perennifòlies com caducifòlies, però les que es fan a les boscanes generalment pertanyen a les segones. És el cas del tortellatge (Viburnum lantana) i de l’aliguer (V. opulus), dos viburns de les boscanes europees, o de l’arç negre americà (V. prunifolium), del viburn adormit o “possumhaw viburnum” (V. nudum) i del viburn d’ovella o “sheepberry” (V. lentago) de les boscanes de l’Amèrica del Nord.
Els lligaboscos o xuclamels són principalment lianes de fulles oposades i flors grans, molt aromàtiques (generalment pol·linitzades per papallones nocturnes), característiques de les boscanes decídues, sobretot de les de l’Àsia oriental, encara que també se’n fan en altres ambients forestals temperats. A les boscanes mixtes de la regió de l’Ussuri i de l’Amur, però, els xuclamels no són pas les úniques lianes; precisament aquestes boscanes presenten una extraordinària riquesa d’espècies lianoides, menys abundants a les boscanes més meridionals, tal vegada per la pressió mil·lenària d’una elevada densitat humana. Entre aquestes plantes enfiladisses destaquen algunes vitàcies com la parra de l’Amur (Vitis amurensis), esquisandràcies com la magnòlia enfiladissa xinesa (Schisandra chinensis) i actinidiàcies com la kolomikta (Actinidia kolomikta) i altres congèneres.
Les aceràcies arbustives, les aquifoliàcies i les celastràcies: els aurons arbustius, els grèvols i els evònims
Encara que la majoria dels aurons són arbres considerables, tal com ja s’ha exposat, n’hi ha també molts, principalment de les boscanes asiàtiques i americanes, que no excedeixen el port arbustiu o que, encara que el tinguin arbori, no arriben a fer més de 8 o 10 m. Seria el cas, per exemple, de dos aurons característics de les boscanes més muntanyenques dels Apalatxes: l’auró de muntanya (Acer spicatum), de fulla marcadament trilobada i escorça llisa d’un bru rogenc, i l’auró ratllat de Pennsilvània (A. pensylvanicum), un petit auró de pell de serp i de fullatge semblant però escorça longitudinalment ratllada de blanc. També és el cas d’un altre auró arbustiu, igualment muntanyenc però d’àrea de distribució més meridional, emparentat amb l’erable de sucre però de dimensions i fulles molt més petites: l’erable de guix (A. leucoderme), que deu el nom a la seva escorça de color blanc i es fa a les boscanes de l’altiplà del Piedmont, des de Carolina del Nord fins a Alabama. A les boscanes de l’Àsia oriental seria també el cas, entre altres, de l’auró groc manxú (A. ukurunduense), propi sobretot de la vall de l’Ussuri.
També poden assolir port arbori algunes aquifoliàcies com els grèvols (Ilex), en particular el grèvol europeu (I. aquifolium), freqüent al sotabosc de moltes fagedes i lligat tradicionalment a les decoracions nadalenques, però en general creixen més aviat de forma arbustiva. Al contrari que l’europeu, que és perennifoli, el grèvol americà decidu o “possomhaw” (I. decidua) de les boscanes meridionals nord-americanes és caducifoli, tot i que una colla de congèneres seus de les mateixes boscanes, com el grèvol americà comú (I. opaca), el grèvol americà muntanyenc (I. montana) o el grèvol verticil·lat (I. verticillata), són de fulla perenne.
Una família molt propera a la de les aquifoliàcies és la de les celastràcies, a la qual pertanyen els evònims (Euonymus), arbustos presents a les boscanes de tots els continents. A les boscanes europees, principalment a rouredes esclarissades i bardisses de clariana o vorada de bosc, es fa l’evònim europeu (Euonymus europaea), de fulla caduca, un arbust de tija quadrangular i fulles oposades, àmpliament lanceolades, que dóna uns fruits vistosos de color rogenc. També és caducifoli l’evònim de foc (E. atropurpurea), d’espectacular coloració tardoral i fruits vistosos i persistents.
Les grossulariàcies i les ericàcies: els ribers, els nabius i els rododèndrons
Ribers (Ribes) i nabius (Vaccinium) tenen en comú que són productors de petits fruits comestibles molt apreciats pels animals de les boscanes, però pertanyen a dues famílies ben diferents, una i altra amb representants a les boscanes i també fora d’elles: grossulariàcies i ericàcies. També tenen en comú el fet d’estendre la seva àrea de distribució tant a les boscanes com als boscos boreals. A les boscanes eurasiàtiques, des de les illes Britàniques i Escandinàvia fins a la conca de l’Amur, són corrents el riber vermell o grosseller (Ribes rubrum) i el riber negre o cassís (R. nigrum), també cultivats pels seus fruits, mentre que a les americanes es troba el riber negre americà (R. americanum) i altres de menys apreciats. Pel que fa als nabius resulta particularment remarcable el nabiu arbori o “sparkleberry” (Vaccinium arboreum) de les boscanes nord-americanes meridionals, ja que té la característica de ser caducifoli a la part més septentrional i alterosa de la seva àrea de distribució i perennifoli a la més meridional i de cota més baixa.
A més a més dels nabius, a les boscanes es fan també altres ericàcies, i això sense comptar el singular paper de brucs i brugueroles a les landes secundàries que es fan a molts indrets del bioma. Es tracta, principalment a les boscanes nord-americanes i a les asiàtiques, de rododèndron (Rhododendron), un gènere molt difós per les muntanyes subtropicals d’Àsia i l’Amèrica del Nord. El rododèndron laurifoli rosa (R. maximum), que pot assolir 12 m d’alçada, malgrat el seu origen subtropical, estén la seva àrea de distribució fins a l’estat de Maine, a l’extrem nord-oriental dels Estats Units; el rododèndron purpuri o rododèndron de Catawba (R. catawbiense) dels Apalatxes meridionals no és tan alt com el seu parent del N i té fulles més amples. El llorer de muntanya (Kalmia latifolia), una altra ericàcia, és un clar exemple d’intrusió del món subtropical a les boscanes decídues nord-americanes més meridionals; la seva fulla perenne recorda, com indica el seu nom, la del llorer i forma masses denses quasi impenetrables als sòls àcids de gran part de les boscanes més orientals dels Estats Units, de Maine fins a Geòrgia.
Les hamamelidàcies i altres famílies: els avellaners de bruixa i altres arbrissons
La família de les hamamelidàcies inclou un centenar d’espècies d’arbres i arbustos de les regions temperades d’Àsia i Amèrica i també de les regions tropicals d’Àsia, Austràlia i Àfrica oriental. De fet és una relíquia de la flora terciària de les latituds mitjanes. A les boscanes complexes xineses i americanes es troben alguns liquidàmbars (Liquidambar) veritablement arboris; fins i tot a les euxínico-caucàsico-hircanianes s’hi fa el liquidàmbar oriental (L. orientalis) que pot arribar a fer 8 m. Aquesta és també la condició de la majoria del anomenats avellaners de bruixa (Hamamelis). El nom d’avellaners de bruixa (“witch-hazel”) correspon sobretot als americans, en particular a l’avellaner de bruixa comú o hamamelis de Virgínia (H. virginiana) les enforcadures del qual són usades pels saurís a l’Amèrica del Nord per a buscar aigua. Com el seu nom indica, són arbustos que pel fullam i els fruits s’assemblen força als avellaners, encara que el seu parentiu és molt remot. A banda de l’avellaner de bruixa comú, que sempre es localitza en indrets on l’aigua al sòl és abundant i l’àrea de distribució del qual penetra cap al N fins a les Províncies Marítimes del Canadà i cap al S fins a Florida, a l’Amèrica del Nord també es fan l’avellaner de bruixa de fulla gran (H. macrophylla), de fulles cobertes de diminuts tubercles, cosa que les fa aspres al tacte, i l’avellaner de bruixa vernal (H. vernalis), de distribució més meridional. A les boscanes de l’Àsia oriental es troben H. japonica i H. mollis.
Les relíquies de la flora terciària són particularment nombroses als sotaboscos de les boscanes de l’Àsia oriental i de l’E de l’Amèrica del Nord: pitosporàcies com el pitòspor japonès o “tobira” (Pittosporum tobira), un arbust perennifoli, de caràcter quasi subtropical que s’ha fet omnipresent als jardins europeus però que és originari de les boscanes xineses i japoneses; hidrangeàcies com el fals taronger xinès (Philadelphus schrenkii), conegut a Europa com a planta ornamental apreciada per l’olor de tarongina de les seves flors, o la dèutzia de l’Amur (Deutzia amurensis); cesalpiniàcies com l’arbre de l’amor americà o “redbud” (Cercis canadensis), un arbrissó de flors roses espectaculars que pot arribar a mesurar 15 m d’alçada; anacardiàcies com el sumac brillant (Rhus copallina), un arbrissó de llargues fulles pinnades, caduques, amb densos raïms de flors blanques aplegats a l’extrem de cada tija, propi de les boscanes nord-americanes però que estén la seva àrea de distribució cap al S fins a Florida i l’E de Texas.
Les comunitats forestals
El bioma de les boscanes decídues és essencialment forestal. Abans que comencés l’explotació agro-silvo-pastoral dels espais d’aquest bioma, la major part del terreny era boscat, tret del de condicions més extremes, com ara els llocs massa inundats o les carenes rocoses i ventades. S’ha de remarcar que, a diferència del que passa a les mediterrànies, on la degradació de les comunitats forestals condueix a l’establiment de formacions arbustives, la degradació de les boscanes decídues sol conduir a la formació de prats. Només sobre substrats que afavoreixen l’acidificació a causa del rentat continuat de les pluges s’instal·len unes bosquines particulars: les landes. L’heterogeneïtat de les condicions del medi i les diferències en les respectives flores, fruit d’històries geològiques divergents, introdueixen una variabilitat considerable en la tipologia de comunitats forestals que es poden trobar als diferents punts del bioma.
Els tipus de boscana decídua
Dins d’aquesta variabilitat es poden reconèixer en conjunt quatre tipus generals de boscanes, que es donen a la major part del bioma, bé que amb diferents combinacions d’espècies a cada regió.
En primer lloc les boscanes mesofítiques, que ocupen la major part del territori en tots els emplaçaments que es poden qualificar de mesofítics, és a dir, sense condicions extremes d’inundació ni de sequera, amb sòls més aviat profunds i ben desenvolupats, amb un pH moderadament bàsic i que només a intervals molt llargs pateixen pertorbacions importants, ja sigui en el règim climàtic o ocasionades per meteors extrems o bé per la intervenció humana.
A les altituds i latituds més elevades del bioma, de clima més fresc, en contacte amb els boscos de coníferes boreals o subalpins, es fan boscanes amb coníferes. En molts països sobreviuen molts exemples de boscanes d’aquest tipus gairebé verges, ja que, considerant les condicions de clima i sòl més desfavorables en què es troben, han estat menys alterades que les mesofítiques per la intervenció humana al llarg dels mil·lennis.
En emplaçaments sotmesos a estrès (per exemple, amb tendència a la sequera, sotmesos a incendis més o menys periòdics o alterats pels humans) apareixen boscanes seques, fredes o secundàries, principalment amb roures, pins i bedolls com a espècies dominants, però que a l’orient asiàtic i nord-americà també presenten una àmplia gamma d’altres gèneres.
Finalment, en sòls humits o inundats de manera més o menys permanent, a les planes al·luvials, es donen les boscanes higrofítiques (vegeu també pàg. 170-171). Les riberes són els emplaçaments d’elecció de pollancredes, vernedes, salzedes, etc, que sovint penetren profundament, al llarg dels cursos d’aigua, als biomes veïns més secs. En emplaçaments lleugerament més elevats: freixenedes, omedes i algunes rouredes asseguren la transició cap a les boscanes mesofítiques.
Les boscanes mesofítiques típiques: les fagedes i les freixenedes
Les boscanes mesofítiques (fagedes, rouredes humides, boscanes mixtes de l’orient asiàtic i americà, etc.) constitueixen el màxim exponent de la vegetació del bioma. El llegendari de tots els pobles que han viscut en contacte amb aquests boscos els han idealitzat i fins de vegades sacralitzat, però aquest respecte no ha impedit que fossin eliminats en una gran part per posar els seus sòls en conreu. Les velles llegendes germàniques conserven la memòria dels temps en què el Danubi i el Rin naixien enmig d’intrincades boscanes (fagedes i rouredes), els límits de les quals no havia albirat mai cap home per més jornades que hi esmercés. Un dels temes clàssics de tardor per als haiku a la literatura japonesa és l’avenç de N cap a S o dels cims cap a la plana al final de la tardor del “momiji zensen”, el front dels fullatges rojos dels aurons. Avui, però, cal buscar aquestes boscanes justament en llocs que no són exactament els òptims, per exemple en estatges montans dels Alps, els Pirineus o els Apalatxes, perquè aquelles que creixien en els millors sòls (sigui a la vall del Rin o a la del riu Groc, a la plana de Tòquio o a les costes de Nova Anglaterra, a la desembocadura del Hudson o a les riberes del Tàmesi) han estat massivament desplaçades, en molts llocs des de fa segles, pels conreus, els habitatges, les indústries i les vies de comunicació.
Per als europeus la fageda és la boscana per excel·lència, la “Hallewald” (que en alemany significa ‘bosc-sala’ o sigui ‘boscana’), anomenat així per la seva disposició pràcticament en dos únics estrats: l’arbori, alt i vigorós, representat pels faigs i algun altre caducifoli escadusser, i l’herbaci, dens i tendre, amb un estrat arbustiu desdibuixat i discontinu. De fet, però, sota el nom de fageda s’engloben boscos força variats que el que tenen en comú és primordialment la dominància del faig a l’estrat arbori. Depenent dels sòls es poden trobar fagedes amb un sotabosc més o menys ric i divers. Depenent de la seva situació geogràfica se’n poden trobar amb diferents espècies de faig com a arbre dominant i amb companyies més o menys variades a l’estrat arbori. Així, si a Europa el faig és pràcticament l’únic arbre de les fagedes, a l’Àsia occidental ja comença a trobar-se amb altres caducifolis, que es fan més nombrosos a l’Àsia oriental; finalment, a Amèrica comparteix en molts casos la dominància amb l’erable de sucre i altres aceràcies.
Les fagedes europees
Les fagedes de faig comú (Fagus sylvatica) es distribueixen molt àmpliament per tota l’Europa central, fins als Urals a l’E, fins al SW de Noruega pel N i fins a les muntanyes més elevades del centre de la Península Ibèrica i de Sicília pel S. Mentre que cap al seu límit septentrional, a les costes del SW d’Escandinàvia, la fageda és una boscana de terra baixa que es dóna al nivell de la mar, a mesura que davalla en latitud s’enfila en altitud, fins a superar els 1 800 m a les regions més meridionals de la seva distribució, a l’Etna. El seu òptim, però, se situa principalment a les regions més humides de l’Europa central, tot i que també n’hi ha una esplèndida representació a l’Europa atlàntica. Nombrosos autors consideren la fageda la comunitat forestal climàcica de la major part de l’espai del bioma de les boscanes decídues a l’Europa continental. A l’estrat arbori, el faig creix generalment sense companyia d’altres arbres, ja que l’ombra densa de la fageda inhibeix el desenvolupament de moltes plantes, tant arbòries com arbustives, però també a causa dels usos forestals que, en moltes zones, han eliminat els arbres de menor interès fuster i han implantat plantacions de faig fora dels límits naturals de la fageda. Així i tot, ocasionalment, a les boscanes medioeuropees i atlàntiques el faig pot créixer associat amb altres espècies arbòries, com l’avet blanc (Abies alba), la pícea o avet roig (Picea abies), el roure de fulla gran (Quercus petraea), l’oma (Ulmus glabra), el freixe de fulla gran (Fraxinus excelsior), etc.
Generalment, els sotaboscos de les fagedes europees són molt pobres en arbustos, però així i tot s’hi fan algunes espècies arbustives o subarbustives, moltes compartides amb les rouredes. Els saüquers (Sambucus), per exemple, o els avellaners (Corylus) són molt més corrents a les rouredes. En algunes fagedes que creixen sobre substrat calcari, sobretot a la part meridional de la seva àrea de distribució, apareix el boix (Buxus sempervirens), un arbust de fulla perenne de creixement molt lent i olor característica. En canvi, a les més frescals, i generalment sobre substrat més aviat àcid, apareixen les matetes dels nabius (Vaccinium); a Europa principalment el nabiu comú (V. myrtillus) i el nabiu vermell (V. vitis-idaea), que entapissen el terra i, de vegades, són l’única planta no herbàcia del sotabosc; les seves baies comestibles són molt apreciades pels animals de les boscanes i també pels humans. En algunes fagedes centreuropees s’hi troben espècies enfiladisses, com les heures (Hedera) o algun lligabosc o xuclamel (Lonicera) i arbustos, com el lloreret (Daphne laureola).
En canvi, a la majoria de fagedes europees l’estrat herbaci és relativament ric i divers, només en algunes el terra apareix desproveït de plantes i cobert exclusivament per la capa de fullaraca. Hom pot distingir diferents tipus de fagedes d’acord amb la presència o l’absència de determinades plantes, la majoria de brotada pre-primaveral o primaveral, d’entre les quals només algunes —poques, tanmateix— són capaces de florir sota l’ombra densa de la volta dels arbres a ple estiu. Entre les plantes característiques d’aquestes fagedes es poden esmentar l’el·lèbor verd (Helleborus viridis), el corniol vulgar (Aquilegia vulgaris) i el lloreret (Daphne laureola). Algunes poàcies, com la poa de boscana (Poa nemoralis) i Bromus ramosus, també són freqüents, com també moltes altres herbes més característiques de les rouredes. Algunes fagedes es caracteritzen per la presència d’orquídies i ciperàcies; destaquen entre les primeres diferents espècies de curraià (Cephalanthera) i la neòtia (Neottia nidus-avis), i entre les segones abunden el càrex blanc (Carex alba) i el càrex digitat (C. digitata). El ranuncle aconitifoli (Ranunculus aconitifolius), diferents cardàmines (Cardamine) i la verònica de fulla d’ortiga (Veronica urticifolia) són característiques de les fagedes amb avet. La mèlica uniflora (Melica uniflora) acompanya la fageda des de la Suècia meridional fins al centre d’Itàlia i amb ella es troba habitualment l’all de bruixa (Allium ursinum).
Les fagedes que creixen en sòls calcaris també tenen les seves plantes herbàcies característiques, com el buixol (Anemone nemorosa), l’espunyidella d’olor (Galium odoratum) i l’herba de les encantades (Circaea lutetiana). Les típiques dels sòls àcids, en canvi, són la lúzula boscana (Luzula sylvatica) i el nabiu comú (Vaccinium myrtillus). A l’estatge subalpí, acompanyen la fageda la paradella arifòlia (Rumex arifolius), les vídues bordes boscanes (Knautia dipsacifolia), el ranuncle aconitifoli i la lletuga alpina (Cicerbita alpina) entre d’altres, i també algunes falgueres com ara la falguera mascle (Dryopteris filix-mas).
Les freixenedes
A l’Europa atlàntica, en contacte amb les fagedes apareixen les freixenedes de freixe de fulla gran (Fraxinus excelsior), sigui perquè aquest substitueix totalment el faig a les regions més elevades, sigui perquè localment marquen una etapa de la successió natural cap a les boscanes mesofítiques (fagedes o rouredes) en el trànsit entre els boscos de ribera i aquestes. A les freixenedes, al costat dels freixes de fulla gran, es pot trobar també l’avellaner europeu (Corylus avellana), dominant al sotabosc, i altres espècies arbòries com l’oma (Ulmus glabra), la moixera comuna (Sorbus aria) o el tell de fulla petita (Tilia cordata) i arbustives com la vidalba (Clematis vitalba) i el tortellatge (Viburnum lantana). A l’estrat herbaci de les més meridionals d’aquestes freixenedes s’hi fan l’espunyidella d’olor, l’all de bruixa, que els dies humits impregna l’atmosfera del bosc de la seva olor penetrant, i el melcoratge de bosc (Mercurialis perennis), que forma, com el precedent, denses catifes; en les àrees més septentrionals, en canvi, s’hi fan el rovell d’ou (Trollius europaeus) i el polemoni blau o escala de Jacob (Polemonium caeruleum), que a moltes zones ha esdevingut una raresa.
Les fagedes euxíniques i hircanianes
Al SE d’Europa el faig oriental (Fagus orientalis) desplaça el faig comú (F. sylvatica). Les fagedes de faig oriental, en efecte, s’estenen cap a l’E per les muntanyes d’Anatòlia fins al Caucas i les muntanyes hircanianes, on ocupen els estatges altimontans, entre els 1 000 i els 2 000 m, i constitueixen la massa forestal més ben conservada i econòmicament més significativa de la regió. El faig oriental és una espècie íntimament relacionada amb el faig comú, fins al punt que, a les zones de contacte entre les àrees de distribució d’una i altra, poden hibridar-se.
Les fagedes de faig oriental són tan esplèndides com les de faig comú. A les zones més elevades l’arbre apareix com a dominant acompanyat d’avets; a les regions més baixes s’associa amb la noguera (Juglans regia), el roure cerris (Quercus cerris), el roure hongarès (Q. frainetto), el roure castanyer comú (Q. castaneifolia), el freixe de fulla estreta (Fraxinus angustifolia), el freixe de flor (F. ornus), l’auró insigne (Acer insigne), el vern del Caucas (Alnus subcordata), el carpinus oriental (Carpinus orientalis), la moixera de pastor (Sorbus torminalis), l’oma (Ulmus glabra) o el teix europeu (Taxus baccata). Entre les espècies de l’estrat herbaci es poden esmentar el ranuncle de Constantinoble (Ranunculus constantinopolitanus), la periploca grega (Periploca graeca) i la fritil·lària pòntica (Fritillaria pontica).
Les fagedes asiàtiques
A l’Àsia oriental només es pot parlar pròpiament de fagedes a Corea i al Japó. En les àrees continentals del bioma, els faigs queden marfosos en tot el complex de les boscanes decídues mesòfiles mixtes, encara que espècies com Fagus englerana o F. longipetiolata puguin conformar localment poblaments més o menys purs, sovint a la zona de transició entre les boscanes mixtes amb coníferes dels confins amb la taigà i les purament decídues.
A les fagedes més septentrionals i montanes del Japó, tant a l’extrem meridional de Hokkaidō com a la part septentrional de Honsh¯u o a les muntanyes de més cap al S, domina Fagus crenata però també hi són comuns altres arbres, com l’auró “itayakaede” (Acer mono), el castanyer d’Índia japonès (Aesculus turbinata), el freixe Fraxinus spaerhiama, la noguera japonesa o “onigurumi” (Juglans ailanthifolia), diferents roures (Quercus) i el “keyaki” o zelkova japonesa (Zelkova serrata). A diferència de les boscanes europees, el sotabosc està força poblat d’arbustos, entre els quals destaquen diferents espècies japoneses de viburn (Viburnum). També apareixen fagedes semblants a aquestes a Corea, si bé allí el faig dominant és F. multinervis.
Més cap al S apareix encara un altre faig, l’“inubuna” (F. japonica), que conviu amb una gran varietat d’espècies caducifòlies (roures, castanyers, aurons, til·lers, freixes, zelkoves), l’abundància de les quals varia segons l’àrea considerada. Aquest bosc majestuós s’enriqueix, a més, amb la presència d’algunes coníferes gegantines com ara el sugi o cedre roig japonès (Cryptomeria japonica), que localment pot formar poblacions pures i, en realitat, no és un cedre sinó un representant de la família de les taxodàcies, és a dir, un parent de les sequoies i de la hiba o “asuwahinoki” (Thujopsis dolabrata) que, localment, pot formar poblacions pures.
Les fagedes nord-americanes
A les boscanes decídues nord-americanes les fagedes apareixen sobretot cap al N, sobre sediments d’origen glacial (“tills”) de les planes d’Indiana i Wisconsin i de la part oriental de la regió dels Grans Llacs. Tot i que en aquestes comunitats hi ha altres espècies de fulla ampla, el faig americà (Fagus grandifolia) i l’erable de sucre (Acer saccharum) hi són codominants, en part gràcies a la seva tolerància a l’ombra, i en alguns rodals arriben a constituir fins al 90% de la població total d’arbres. Les comunitats de faig americà i erable de sucre se solen trobar, en aquesta zona, sobre sòls de textura densa i compacta. El faig americà tolera indrets més humits i més mal drenats i sòls més àcids que l’erable de sucre, mentre que aquest exigeix sòls més ben drenats però tolera un marge de variació de pH més ampli que va de molt àcid a lleugerament alcalí. Totes dues espècies posseeixen sistemes radiculars relativament superficials que són afectats fàcilment pels incendis. Cap a l’W, el tell americà (Tilia americana) desplaça gradualment el faig d’aquestes boscanes, mentre que als indrets més secs apareixen les rouredes amb cària, característiques del marge occidental de les boscanes decídues nord-americanes.
La colonització d’aquesta regió pels nord-americans d’origen europeu en els darrers segles ha alterat considerablement la composició originària d’espècies, en part a causa de les rompudes per establir-hi conreus i en part a causa de la tala selectiva que féu desaparèixer de l’estrat arbori espècies com l’esmentat tell americà, el tuliper de Virgínia (Liriodendron tulipifera), el roure blanc (Quercus alba) i el cirerer americà (Prunus serotina), deixant espècies menys valorades, com el mateix faig americà, les capçades del qual han substituït els buits deixats pels altres arbres.
Les fagedes austroamericanes
Les fagedes de l’extrem meridional d’Amèrica es distribueixen entre una latitud de 35 i 55°S i una altitud que va des del nivell de la mar fins a 1 800 o 1 900 m; per tant no és pas sorprenent que, tot i l’extensió relativament limitada que tenen, siguin d’una gran variabilitat. La confusió comença perquè la fageda per excel·lència d’aquesta àrea (el “Sur Chico” dels geògrafs xilens) és, si hom s’ateny a la nomenclatura local, una roureda! Es tracta, en efecte, dels boscos del “roble” Nothofagus obliqua, també conegut com a “roble pellín” i “pellín”, de la província xilena de La Araucanía i àrees limítrofes de Bío-Bío. L’explicació és que aquest faig austral té les fulles retallades i l’escorça fosca, més semblant a la d’un roure de l’hemisferi boreal que a un faig. Els boscos de “roble” ocupen els estatges inferiors, fins a uns 1 000 m d’altitud, a la part septentrional de la seva àrea de distribució (entre els 36 i els 37°S, més al N els estatges més baixos són ocupats per la mediterrània xilena); cap al S es barreja progressivament amb el raulí (N. procera), que té una amplitud ecològica més gran i que ocupa els marges altitudinals i meridionals del “Sur Chico”. Són boscos densos i alts, amb cobricels que s’enfilen fins a 40 m, amb un estrat arbori dominant de “roble” i un estrat arbori més baix que, a les regions on el raulí entra en aquestes fagedes, està ocupat principalment per arbres perennifolis més propis de la laurisilva valdiviana, com el lingue (Persea lingue) i el llorer xilè (Laurelia aromatica).
El raulí domina precisament les latituds més meridionals del “Sur Chico”, entre els 38 i els 40°S aproximadament, encara que a la regió dels llacs de la vall longitudinal xilena es prolonga fins als 41°S i, localment, fins a latituds més meridionals. Encara que s’assembla extraordinàriament al faig comú, el raulí no s’arriba a fer tan alt com el “roble” (amb cobricels d’uns 35 m d’alçada) però també apareix associat tant a arbres caducifolis com a perennifolis. Entre els primers destaca un altre faig austral: el “ñirre” (Nothofagus antarctica) i entre els segons el “mañío“ (Podocarpus salignus) i l’ulmo (Eucryphia cordifolia) i un segon faig austral, molt important més al S, a les selves temperades magallàniques: el “coihue” (Nothofagus dombeyi).
Més al S, l’únic caducifoli important és la lenga (Nothofagus pumilio). Els boscos de lenga formen una franja forestal discontínua al vessant oriental dels Andes, on constitueixen el límit de la vegetació forestal, tant amb relació a l’altitud com amb relació a l’aridesa del subdesert patagònic. De fet, remunten al N fins a la latitud del llac Aluminé (38°55’S), on el límit superior del bosc se situa a 1 700 m, i arriben pel S fins a la Terra del Foc, on se situa entre 500 i 600 m, i fins i tot a les illes més meridionals i extremes, on en llocs arrecerats i en cotes que no superen els 450 m d’altitud (200 a l’illa de Los Estados) encara es poden trobar claps d’aquests boscos. El seu límit inferior se situa habitualment a 400 o 500 m per sota del límit superior excepte a les latituds més meridionals, on poden arribar fins al nivell de la mar.
La lenga és un arbre més baix que el “roble” o el raulí, en els casos més favorables pot arribar a assolir alçades fins de 30 m, però és més usual que no passi dels 20 i al seu límit superior sempre apareix agarrigat a causa de l’acció combinada del vent, la neu i la solifluxió. Generalment forma rodals purs, però a les localitzacions més desfavorables es troba associada amb el “ñirre” i, a les més afavorides del N de l’àrea de la lenga, amb el “coihue” i l’araucària xilena (Araucaria araucana).
Les boscanes submesofítiques: les rouredes
Conjuntament amb les fagedes, les rouredes són el màxim exponent de les boscanes decídues, tant al centre i l’orient d’Europa com a l’Europa atlàntica. Dues espècies, el roure pènol (Quercus robur) i el roure de fulla gran (Q. petraea), es reparteixen el protagonisme, amb una certa preferència atlàntica del segon i medioeuropea del primer. Així i tot, ambdós arbres poden compartir dominància en determinades localitats, com passa al famós bosc de Sherwood, prop de Nottingham, a Anglaterra, en altre temps refugi de Robin Hood i els seus arquers i avui quasi desaparegut, en el qual totes dues espècies conviuen barrejades. Les rouredes, encara més que les fagedes, han estat l’ambient pristi dels humans que han conformat la majoria dels pobles medioeuropeus. Els boscos primerament roturats pels agricultors neolítics foren rouredes, i de les rouredes han extret històricament els recursos complementaris que necessitaven, alguns de gran importància com a base del desenvolupament econòmic dels temps moderns. Potser per això el roure pènol és, en moltes regions europees, un arbre emblemàtic i alhora sobreexplotat, de manera que moltes rouredes de roure pènol estan, avui, en una situació molt precària.
Les rouredes atlàntiques i medioeuropees
La roureda de roure pènol (Quercus robur) és una boscana d’arbres robustos i esvelts (que poden assolir més de 40 m) i capçades denses, bé que no tant com les de les fagedes. Es fa en sòls rics, profunds, preferiblement humits i sense calci, en àrees de clima humit sense secades a l’estiu. Com que no forma un cobricel tan dens com el de les fagedes, altres arbres petits poden créixer a redós dels roures, i es desenvolupa un estrat arbustiu més dens i ric. El roure pènol és un arbre de creixement lent i llarga vida (pot viure un miler d’anys), fins al punt que es considera que cada centímetre de circumferència del tronc representa un any de vida, i no floreix fins a l’edat de 40 o 50 anys. Ho fa entre l’abril i el maig, al mateix temps que comencen a aparèixer les seves fulles; la gla madura al setembre i cau de l’arbre a l’octubre. La roureda de roure de fulla gran (Q. petraea) és menys exigent pel que fa a sòls. N’admet de més secs i menys profunds que la roureda de roure pènol i suporta més bé els calcaris. En conjunt, però, la distribució de tots dos roures és quasi coincident.
Les rouredes atlàntiques, siguin de roure pènol, de roure de fulla gran o de tots dos, poden acollir altres arbres, principalment bedolls (Betula), pollancres (Populus), moixera de guilla (Sorbus aucuparia), vern comú (Alnus glutinosa), carpinus comú (Carpinus betulus), oma (Ulmus glabra), pomera borda (Malus sylvestris), teix europeu (Taxus baccata) i grèvol (Ilex aquifolium). La presència i l’abundància de cada un d’aquests arbres és determinada pel tipus de sòl i per la situació geogràfica. El bedoll i la moixera de guilla, per exemple, es desenvolupen a les terres altes, mentre que el vern creix més aviat en condicions humides.
Pel que fa al sotabosc, les rouredes posseeixen un estrat arbustiu força ric que varia d’acord amb les condicions locals, però en el qual alguns gèneres i espècies particularment significatius són quasi sempre presents. Els avellaners (Corylus) en són un exemple; l’avellaner europeu (C. avellana) es fa pràcticament a totes les boscanes europees i, en vorades de bosc i al llarg del costat més allunyat de l’aigua de molts boscos de ribera, fins i tot forma avellanoses gairebé monospecífiques. Fan el mateix altres espècies del gènere a les regions on creixen. Al costat dels avellaners els sanguinyols i cornellers (Cornus) també apareixen a les boscanes decídues de tot Europa; el sanguinyol comú (C. sanguinea) creix a tots els boscos de roure, mentre que el corneller mascle (C. mas) és més característic dels de roure hongarès (Quercus frainetto) i de roure cerris (Q. cerris). Un altre grup d’arbustos de les rouredes, variat i abundant, és el dels arços (Crataegus), gènere que s’estén arreu del món i mostra una gran adaptabilitat a diferents condicions ambientals. Al costat d’aquests, a les rouredes europees sovint creixen encara altres arbustos: el saüc europeu comú (Sambucus nigra), l’aranyoner (Prunus spinosa), l’aliguer (Viburnum opulus), l’evònim (Euonymus europaea), el cambró (Rhamnus cathartica), l’olivereta (Ligustrum vulgare) i molts altres.
L’estrat herbaci és ric en espècies pre-vernals com la prímula vera (Primula vulgaris), la maduixera boscana (Fragaria vesca), el buixol (Anemone nemorosa) o l’heura de terra (Glechoma hederacea), i vernals, com diferents veròniques (Veronica) o la búgula (Ajuga reptans), però també en espècies plenament estivals que accepten bé l’ombra densa de les capçades ben carregades de fullam, com l’ortiga pudent (Stachys sylvatica), la campaneta traqueli (Campanula trachelium), les didaleres (Digitalis) o els epilobis (Epilobium). Hom hi troba igualment nombroses espècies de poàcies i de ciperàcies i força molses. Les falgueres, com la falguera comuna (Pteridium aquilinum), la falguera de rei (Osmunda regalis), la falguera femella (Athyrium filix-femina) o la falguera dels pantans (Thelypteris palustris) abunden sovint. Freqüentment, les rouredes dels boscos de roure pènol de l’Europa central tenen al seu estrat herbaci una delicada planta de flors grogues, mancada de clorofil·la perquè és paràsita, el xuclador (Melampyrum pratense), però principalment hi ha poàcies com la lúzula blanca (Luzula luzuloides) o la festuca ovina (Festuca ovina), i molses com Dicranum scoparium i Leucobryum glaucum. En les àrees on el roure de fulla gran és codominant amb el pi roig (Pinus sylvestris), per exemple en les àrees continentals de terra baixa, apareixen el nabiu, el xuclador i ciperàcies com Carex pilulifera.
Cap al N de la seva àrea de distribució, a les rouredes de roure pènol també es troben associats el trèmol (Populus tremula) i la moixera de guilla (Sorbus aucuparia). En algunes àrees de clima continental, en contacte amb la taigà, el pi roig (Pinus sylvestris) és codominant amb el roure de fulla gran i es troba associat amb el bedoll, el trèmol i el tell de fulla petita (Tilia cordata). Més cap al S, el roure de fulla gran duu associat el castanyer (Castanea sativa) i comencen a aparèixer el freixe de fulla gran (Fraxinus excelsior), el pollancre ver (Populus nigra), el tell de fulla gran (Tilia platyphyllos) i l’om (Ulmus minor). Cap al SE, als Balcans, la diversitat és més gran; hom pot trobar el roure hongarès (Quercus frainetto) i el roure cerris (Q. cerris) al costat del roure martinenc (Q. humilis [=Q. pubescens]), del carpinus oriental (Carpinus orientalis), del server (Sorbus domestica), de la moixera de pastor (S. torminalis), del freixe de flor (Fraxinus ornus), de la perera silvestre (Pyrus pyraster), del tell argentat (Tilia tomentosa) i d’altres espècies secundàries més petites, com l’avellaner turc (Corylus colurna), més gran que l’europeu, amb fruits també més grossos i de port més arbori, amb una capçada piramidal.
Molt característica de l’Europa central, a altituds compreses entre 200 i 500 m, és la boscana de carpinus comú (Carpinus betulus) i roure (sigui roure pènol o roure de fulla gran). Cap a l’W, a la costa atlàntica, aquesta boscana prefereix àrees relativament seques, mentre que cap a l’E, a les proximitats de la mar Negra, en condicions més continentals, necessita més humitat. La proporció de cada espècie és variable i, de tant en tant, en boscanes d’aquest tipus poden aparèixer altres espècies com el plàtan fals (Acer pseudoplatanus), el tell de fulla petita (Tilia cordata) i de vegades l’avet blanc (Abies alba) o la pícea (Picea abies) i, a les regions atlàntiques, fins i tot el faig (Fagus); en canvi, a les àrees més meridionals on domina el roure pènol, les espècies que hi viuen associades són l’auró blanc (Acer campestre), l’auró de Tartària (A. tataricum), l’om comú (Ulmus minor) i el cirerer silvestre (Prunus avium). Arreu d’Europa, la gran varietat dels boscos de roure i carpinus es reflecteix també en les plantes que creixen a l’estrat herbaci. El tipus nord-occidental de boscana de roure i carpinus el caracteritzen el narcís de muntanya (Narcissus pseudonarcissus), la rèvola (Stellaria holostea) i diferents campànules (Campanula) i lligaboscos com, per exemple, el lligabosc atlàntic (Lonicera periclymenum). En canvi, a les de tipus oriental hi sovintegen Viola mirabilis i Festuca heterophylla. Les de tipus subatlàntic de les terres baixes càlides acullen la maduixera (Fragaria vesca), l’espunyidella boscana (Galium sylvaticum), el muguet (Convallaria majalis), la poa de boscana (Poa nemoralis) i el càrex boscà (Carex sylvatica).
Les rouredes euxíniques i caucàsiques
Al N d’Anatòlia, les rouredes ocupen els espais moderadament humits i van deixant pas a les estepes a mesura que el clima esdevé més sec. Cap a l’W, a la major part de Turquia, una gran part de les rouredes són constituïdes per roure cerris (Quercus cerris) i roure hongarès (Q. frainetto), amb els quals es barregen el carpinus oriental (Carpinus orientalis), el castanyer (Castanea sativa) i el trèmol (Populus tremula). Gairebé totes aquestes rouredes han estat molt degradades per un aprofitament i un pasturatge abusius i, normalment, s’exploten com a bosc baix. La meitat occidental del SE d’Anatòlia, entre 1 000 i 2 000 m d’altitud, és una mica més boscosa, probablement perquè està exposada als vents carregats d’humitat procedents de la Mediterrània, però els boscos estan igual de degradats que a la resta de la regió. Les rouredes esclarissades s’estenen encara del SE d’Anatòlia cap a les muntanyes del Zagros, a més altitud com més al S.
També es troben rouredes a les zones muntanyoses del N de Crimea i als raiguers de la serralada del Caucas. A la costa sud-occidental de la mar Càspia encara hi ha extensions força grans de roureda de roure castanyer comú (Quercus castaneifolia), en indrets amb temperatures moderades i una precipitació una mica més alta.
Les rouredes asiàtiques
A l’Àsia oriental no hi ha pròpiament rouredes, si no és com a boscana secundària resultant d’una pertorbació o d’una plantació. Tanmateix, com a Europa i a l’Amèrica del Nord, existeix una àrea en la qual els roures tenen una presència particularment destacada a les boscanes decídues, que aquí són d’una heterogeneïtat, riquesa i magnificència només comparables amb les de les selves tropicals. En particular s’hi troben el roure dàimio (Quercus dentata), que és el roure de fulles més grans (poden arribar a superar els 30 cm de llarg) i està àmpliament distribuït a la Xina, a Corea i al Japó, l’omnipresent roure mongol (Q. mongolica) i també Q. aliena. Al costat dels roures destaquen les nombroses espècies d’aurons, com l’auró de Her (Acer hersii) o l’auró d’escorça de paper (A. griseum), de verns (Alnus), de moixeres i servers (Sorbus), de magnòlies (Magnolia), de moreres (Morus), de pereres (Pyrus), de pomeres (Malus) i de pruneres i altres arbres amb fruits de pinyol (Prunus). Al Japó, si l’avenç del front dels fullatges rojos dels aurons, el “momiji zensen”, indica la plenitud de la tardor, el “hana zensen”, el front de les flors que es van obrint progressivament de SW cap a NE tot al llarg del país, és el tema central dels haikus de la primavera.
Completa l’esplendor d’aquestes boscanes una gran varietat d’altres espècies d’arbres: fagàcies com el castanyer comú (Castanea sativa), el castanyer xinès (C. mollisima) o diferents espècies de Castanopsis; ulmàcies com els lledoners (Celtis biondii, C. labilis, C. julianae) o la zelkova de Schneider (Zelkova schneideriana); oleàcies com el freixe de fulla ampla (Fraxinus rhynchophylla); araliàcies com el “sen” (Eleutherococcus [=Kalopanax] pictus); rutàcies com el surer de l’Amur (Phellodendron amurense); fabàcies o papilionàcies com Maackia sinensis, i fins i tot teixos com el teix japonès (Taxus cuspidata). Aquests boscos han estat talats a gran escala i en algunes zones apareixen substituïts per boscos secundaris de roures, pollancres i bedolls, amb l’afegitó, cap al S, de plantacions de pi xinès (Pinus tabulaeformis).
Les rouredes nord-americanes
A les rouredes de l’E de l’Amèrica del Nord es dóna una situació intermèdia entre la de les d’Europa i la de les de l’Àsia oriental. No és tan freqüent la dominància exclusiva d’una sola espècie de roure o de dues, com a Europa, però la complexitat de les boscanes tampoc acostuma a ser tan gran com la de les de l’Àsia oriental. Es troben rouredes amb diferent composició de l’estrat arbori i del sotabosc a la majoria de les regions en què poden dividir-se les boscanes.
A la regió més meridional del bioma, la de les boscanes de roures, càries i pins, d’estius calorosos, hiverns relativament suaus, amb una estació de creixement llarga i una humitat elevada, s’hi troba principalment una rica barreja de roures, entre els quals el roure blanc (Quercus alba), el roure negre (Q. velutina), el roure eriçó o “bur oak” (Q. macrocarpa), el roure “chinquapin” (Q. muehlenbergii), el roure de Maryland (Q. marilandica), etc.; també abunden les càries, com la cària amarga (Carya cordiformis), la cària vera (C. glabra), la cària tomentosa (C. tomentosa), la cària blanca (C. ovata), etc. Al S, les boscanes d’aquesta regió reben la influència dels boscos perennifolis més meridionals, cosa que es tradueix en la presència de magnòlies, tulipers i pins.
El foc ha estat un factor determinant per a la composició d’aquestes boscanes i ha afavorit els pins i els roures piròfits. A banda dels focs espontanis, sembla que antigament els indis ja se servien del foc per crear hàbitats favorables als animals que caçaven. Els pins, sobretot el pi “loblolly” (Pinus taeda) i el pi americà de fulla curta (P. echinata), són particularment abundants a la part meridional de l’altiplà de Piedmont, des de les Carolines cap al SW. En el passat, les pinedes pirofítiques havien estat un tret característic de la regió, però, actualment, les pinedes amb sotabosc de planifolis o els rodals mixtos de pins i planifolis en camps abandonats posen de manifest la gran influència que tingué l’acció humana, amb els seus conreus itinerants, en l’establiment d’aquella vegetació permanent controlada pel foc (piroclímax). El bosc caducifoli, generalment una roureda, tendeix a substituir les pinedes als indrets més fèrtils i humits i als llocs on els incendis són relativament poc freqüents; però en condicions extremes, amb incendis freqüents i sòls eixuts, també hi ha roures que tendeixen a substituir fins i tot els pins més resistents al foc.
Al NW d’aquesta regió s’estenen els Apalatxes, serralada que constitueix per ella mateixa una regió, la de les boscanes apalatxianes de roures i castanyers, i que es prolonga més enllà de l’extrem muntanyós septentrional fins a la costa que va de Nova Jersey a Massachusetts. En conjunt, és una regió de clima una mica més fred que el de la precedent, marcat per l’altitud i el relleu, amb una gran varietat de sistemes naturals, des de boscanes mesofítiques complexes situades als característics “coves” (valls engorjades i arrecerades), fins a boscos de tipus boreal a les carenes més alteroses; les rouredes són pròpies sobretot de terra baixa i muntanya mitjana. Fins al començament del segle XX, el castanyer americà (Castanea dentata) dominava en molts indrets al costat dels roures, però fou pràcticament eliminat de l’estrat arbori pel xancre, una greu malaltia que afecta l’escorça de l’arbre i és causada per Endothia parasitica, un fong paràsit originari de Xina introduït accidentalment als boscos americans cap el 1909.
Els principals factors que han afectat la composició dels boscos d’aquesta zona, però, han estat els incendis i les glaçades. A la primeria del segle XVIII, amb l’avenç de la colonització europea, la major part del bosc fou estassat i cremat, i sovint es deixava que els incendis cremessin sense control grans extensions de terreny. Els incendis també foren habituals durant els anys de màxima activitat de l’explotació forestal, entre el final del segle XIX i el començament del XX. Les glaçades també han estat sempre una pertorbació d’importància, però no tan evident. Tenen efectes diferents sobre les diferents espècies arbòries de fulla caduca; algunes les resisteixen, com la majoria d’espècies de roure i de cària, altres no, com el tuliper de Vírginia (Liriodendron tulipifera) i els pins, potser perquè són perennifolis. Tot i aquestes pertorbacions, el model de distribució d’espècies no sembla haver canviat significativament des dels temps pre-colonials.
Finalment, cal considerar una tercera regió de rouredes nord-americanes, la de les boscanes occidentals de roures i càries. Aquesta zona, la més occidental de les regions de boscanes decídues de l’E de l’Amèrica del Nord, s’estén a l’W del Mississipí, des de la plana litoral de Texas, al golf de Mèxic, al SW, fins a les praderies de Manitoba al N, encara que alguns autors adjudiquen el seu extrem meridional, en terres de Texas i Oklahoma, a la regió de les boscanes meridionals de roures, càries i pins. Es tracta dels anomenats “cross timbers”, unes rouredes esclarissades que fan la transició cap a les praderies i on predominen el roure de Maryland (Q. marilandica), conegut a la regió com “blackjack oak” per la seva escorça negrosa i la seva fusta tova i flexible, i el Q. stellata, un roure blanc conegut com “post oak” per la qualitat de la seva fusta com a material de construcció. Una falca penetra cap a l’E del Mississipí, entre la vall baixa de l’Ohio i les ribes meridionals del llac Michigan. Cap a l’W, les boscanes van quedant confinades a les valls encaixades que solquen les planes cobertes que havien ocupat anteriorment les praderies del centre de l’Amèrica del Nord, avui cultivades pràcticament pertot.
Les rouredes i les rouredes amb càries (aquestes rarament són dominants) d’aquestes regions més occidentals són probablement les boscanes amb més variabilitat geogràfica i ecològica de tota l’Amèrica del Nord, cosa que no és estranya atès el nombre i la varietat d’espècies que hi ha, tant d’un gènere com de l’altre, i les diferències de latitud entre els extrems septentrional i meridional. Les diferències més marcades es manifesten al llarg del límit de les glaciacions quaternàries. A les àrees meridionals el clima és continental, amb estius càlids i hiverns freds i un gradient de pluviositat descendent d’E a W que produeix un canvi gradual en la vegetació, però sense que es marquin límits precisos entre els diferents tipus; aquests límits més o menys indeterminats no sols depenen de la pluja sinó també de les condicions climàtiques locals, el nivell de la capa freàtica i el tipus de sòl. A l’extrem meridional, les rouredes es veuen sovint acompanyades de pins o de xiprers calbs, mentre que a les àrees septentrionals, les rouredes amb càries ocupen una àmplia zona modelada per les glaciacions i de clima més continental que el de les àrees meridionals. Als llocs favorables dels obacs de les muntanyes, a l’estrat arbori també apareixen aurons, tells, freixes, nogueres i fins i tot magnòlies caducifòlies.
Les boscanes mesofítiques complexes
Ja a la regió euxino-caucàsico-hircaniana es poden trobar boscanes d’elevada diversitat a diferents indrets de les zones al·luvials càlides i humides. Les platanedes de plàtan oriental (Platanus orientalis), que es desenvolupen a les parts baixes de les valls de la costa turca de la mar Negra, en són un exponent. Entre les mars Negra i Càspia, on el clima suau i les abundants precipitacions afavoreixen un bon desenvolupament del bosc, es troben boscanes de gran interès florístic, amb una varietat d’espècies elevada, bé que, per damunt dels 1 000 m la proporció de faig augmenta i el bosc es transforma progressivament en rodals mixtos al si de la fageda.
Els nivells de biodiversitat s’incrementen encara més en la secció oriental del bioma. Allí, a la part meridional de l’àrea de les boscanes, les espècies de fulla caduca es barregen amb les espècies laurifòlies per formar una boscana de transició cap a la laurisilva. Tanmateix, les espècies caducifòlies tendeixen a desplaçar les perennifòlies perquè la regeneració espontània d’aquestes pateix més les interferències humanes, molt importants a causa de la densitat demogràfica de la regió. També és en aquesta àrea on apareixen algunes espècies rares, relictes de la flora del Terciari, com el ginkgo (Ginkgo biloba) o la sequoia xinesa (Metasequoia glyptostroboides), una conífera de fulla caduca que fins el 1941 només era coneguda en estat fòssil. També s’hi troben diferents menes de roures i de tells, i altres espècies entre les quals anardiàcies com el pistatxo xinès (Pistacia chinensis) o el sumac xinès (Rhus chinesis), la betulàcia Carpinus fargesii, la cornàcia Cornus controversa o sanguinyol gegant xinès, el caqui (Diospyros kaki), que és una ebenàcia, la fagàcia Castanea seguinii, el liquidàmbar xinès (Liquidambar formosana), que és una hamamelidàcia, la juglandàcia Platycarya strobilacea, la lauràcia Sassafras tzumu, la mimosàcia Albizia kalkora o la fabàcia Dalbergia hypheana.
Al Japó hi ha boscanes mesofítiques complexes similars a les descrites que s’estenen a través de les illes de Ky¯ush¯u i Shikoku i a l’extrem meridional de Honsh¯u. Una de les espècies dominants és Persea [=Machilus] thunbergii, característica de les laurisilves del S del Japó. Aquest arbre creix fins a la part central de l’illa de Honsh¯u associat amb l’alzina japonesa de fulla estreta (Quercus acuta), que es distingeix perquè les seves glans s’agrupen dins una coberta vellutada. Entre els aurons, abunda l’auró japonès (Acer palmatum), que presenta moltes formes diferents de colors vistosos. També hi ha castanyers, com Castanea crenata, i, cap al S, sovintegen diverses espècies de falsos castanyers (Castanopsis). Encara que no sigui exclusiva del Japó, el katsura (Cercidiphyllum japonicum) és una espècie molt característica perquè és omnipresent en la imatgeria japonesa, amb les seves fines branques poblades de fulles arrodonides i d’alçada mitjana (uns 15 m), sovint creix amb port arbustiu, ramificat des de la soca i és l’únic representant de la família de les cercidifil·làcies.
Als Estats Units, les boscanes mesofítiques complexes, sense assolir la formidable diversitat de les de l’Àsia oriental, tenen també una presència destacada. Es distribueixen del SW de Pennsilvània i la capçalera de l’Ohio fins al NE d’Alabama, un espai que coincideix a grans trets amb els sectors dels altiplans interiors dels Apalatxes que no van ser afectats directament per les glaceres wisconsinianes. Cap al S es marfonen amb les boscanes de roures, càries i pins, i cap a l’E i el NE amb les rouredes apalatxianes. El clima és temperat càlid, amb estius calorosos i plujosos, amb precipitacions i humitat relativament elevades; la vegetació es caracteritza per la presència d’un variat mosaic de comunitats mesofítiques complexes amb un bon desenvolupament tant de l’estrat arbori com del sotabosc arbustiu i de l’herbaci. L’estació de creixement, que és llarga i càlida, juntament amb uns hiverns suaus, crea les condicions adequades perquè el nombre de comunitats i d’espècies arbòries mesofítiques sigui més gran que a cap altra zona de l’Amèrica del Nord. Les espècies més comunes són molt nombroses: el faig americà (Fagus grandifolia), el tuliper de Virgínia (Liriodendron tulipifera), el tell blanc (Tilia heterophylla) i el tell americà comú (T. americana), l’erable de sucre (Acer saccharum) i l’auró negre (A. nigrum), el castanyer d’Índia groc (Aesculus octandra), el roure blanc (Quercus alba) i el roure roig (Q. rubra), el bedoll groc (Betula alleghaniensis) i el bedoll dolç (B. lenta), el cirerer americà (Prunus serotina) i el cirerer de Pennsilvània (P. pensylvanica), la magnòlia de fulla gran (Magnolia macrophylla) i la magnòlia de paraigües (M. tripetala), l’arbre de cogombres (M. acuminata), el freixe blanc (Fraxinus americana), la cària amarga (C. cordiformis) i la cària blanca (C. ovata), la noguera negra (Juglans nigra), i fins i tot el vistós tupelo negre (Nyssa sylvatica), un dels rars representants de la família de les nissàcies.
A l’W de la part meridional de la zona de les boscanes mesofítiques, des de la vall baixa de l’Ohio cap al S, fins al N de Mississipí i Alabama, es troba una regió accidentada que combina vessants abruptes, peneplans erosionats, altiplans dissecats i cursos d’aigua divagants amb àmplies conques al·luvials. S’hi troba un mosaic de formacions vegetals no menys complex que el descrit, el qual fa de transició entre les boscanes de més a l’E, més riques en espècies, i les extenses rouredes i comunitats de roure amb cària de més al S i de més a l’W. Aquestes boscanes mesofítiques occidentals complexes sovint són considerades com una zona diferenciada de la resta de boscanes mesofítiques complexes americanes.
Les boscanes mixtes amb coníferes
Les boscanes amb coníferes es troben a les altituds i latituds més elevades del bioma, de clima més fresc, en contacte amb els boscos de coníferes boreals o subalpins. Tenen la seva màxima extensió a les boscanes asiàtiques i nord-americanes, mentre que a Europa només tenen alguna significació a les boscanes més nord-orientals, a la Rússia centre-septentrional, on l’estrat superior de les rouredes de roure pènol (Quercus robur) amb tell de fulla petita (Tilia cordata) es veu envaït de manera progressiva per les coníferes de la taigà immediata, principalment per la pícea o avet roig (Picea abies), mentre que l’estrat arbori inferior continua dominat per caducifolis com el mateix tell de fulla petita, l’erable (Acer platanoides) o el trèmol (Populus tremula). Més a l’E, als contraforts dels Urals, entre els 52° i els 56°N, on la roureda és substituïda per la telleda de tell de fulla petita, boscana i taigà també s’interpenetren.
A l’Àsia oriental, a la regió dels rius Ussuri i Amur, a la frontera russo-xinesa (una de les zones més ben conservades de tot el macrocontinent eurasiàtic) també convergeixen els dos biomes. En el que pot ben bé qualificar-se de paradís botànic, aquesta regió acull un gran nombre d’espècies endèmiques al costat d’espècies d’afinitats boreals, provinents de la taigà, i d’espècies d’afinitats subtropicals que han anat avançant de mica en mica al llarg de la costa xinesa cap al N.
Allà, la taigà siberiana es transforma a poc a poc en boscos mixtos de coníferes i planifolis, principalment bedolls com el bedoll d’Erman (Betula ermanii) i el bedoll blanc asiàtic (B. platyphylla), a més a més d’alguns trèmols (Populus tremula). La part més meridional de la regió és dominada per l’avet manxú o avet gegant negre (Abies holophylla), amb un estrat arbori secundari de carpinus manxú (Carpinus cordata) i moltes espècies d’arbres i d’arbustos típics d’àrees més meridionals de les boscanes de l’Àsia oriental, com el “sen” (Eleutherococcus [=Kalopanax] pictus) o l’auró coreà (Acer pseudosieboldianum). També són abundants les pinedes, sobretot de pi coreà (Pinus koraiensis), barrejades amb diverses espècies de planifolis decidus com el freixe manxú (Fraxinus mandschurica), l’om d’escorça blanca (Ulmus propincua), la noguera manxú (Juglans mandshurica), l’auró manxú (Acer mandshuricum) o l’auró coreà, el tell de l’Amur (Tilia amurense) o el tell manxú (T. mandshurica) o el característic i interessant surer de l’Amur (Phellodendron amurense), un arbre de la família de les rutàcies que, com el freixe manxú o el teix japonès, constitueix un vestigi de la flora terciària. El surer de l’Amur és un arbre d’escorça grisa i suberosa, profundament clivellada com la dels suros mediterranis, que pot arribar a fer fins a 15 m d’alçada, amb les branques disposades en una àmplia corona, no excessivament densa; les seves fulles, decídues, es tornen roges a la tardor, i alhora l’arbre es carrega de raïms de fruits d’un negre brillant, com petites cireres fosques.
L’estrat arbustiu d’aquests boscos comprèn un seguit d’espècies endèmiques, com el lilà de l’Amur (Syringa amurensis) i la falsa acàcia de l’Amur (Maackia amurensis). La flora herbàcia no és menys rica i peculiar. Entre les espècies més representatives es poden esmentar la campanulàcia Codonopsis ussuriensis, que té unes flors que recorden les de les gencianes però d’un verd pàl·lid i tubercles comestibles; la petita papaveràcia de flors blanques Corydalis repens; el cipripedi de flor gran (Cypripedium macranthum), que és una orquidàcia; l’adonis de l’Amur (Adonis amurensis), ranunculàcia amb denses inflorescències grogues que en la seva florida pre-vernal llueixen espectacularment enmig de la neu, la cariofil·làcia Lychnis fulgens, de flors d’un vermell esclatant, i moltes altres espècies que reparteixen la seva activitat entre primavera i estiu. També hi arriben, encara que es tracta d’espècies d’àrea més meridional, ranunculàcies com el corniol de l’Amur (Aquilegia amurensis), liliàcies com l’hemerocal·lis groc o flor d’un dia groga (Hemerocallis lilioasphodelus [=H. flava]), diverses peoniàcies del gènere Paeonia i nombroses cariofil·làcies del gènere Stellaria.
Hi ha boscos mixtos semblants a aquests que s’estenen cap a Manxúria i Corea. Aquí el pi coreà encara és la conífera dominant, per bé que hom hi pot trobar igualment el làrix manxú (Larix olgiensis), la pícea manxú (Picea mandshurica) i la pícea coreana (P. obovata koraiensis). Gradualment, les espècies caducifòlies passen a fer-se dominants. S’hi troba el roure mongol (Quercus mongolica), sobretot en solells eixuts i carenes ventades, i el freixe manxú (Fraxinus mandschurica), però també hi són ben representades les pomeres silvestres (Malus), més nombroses a la Xina que en cap altre país del món; entre les més comunes cal esmentar la pomera de Hall (M. halliana) i la pomera borda siberiana (M. baccata), un arbre molt gran de florida molt primerenca, com també M. toringoides. Més cap al S, les espècies característiques d’aquests boscos mixtos són el freixe de fulla ampla (Fraxinus rhynchophylla), diversos aurons (Acer), el “sen” (Eleutherococcus [=Kalopanax] pictus), el teix japonès (Taxus cuspidata) i d’altres espècies d’àrea més àmplia com el castanyer comú (Castanea sativa) i la noguera comuna (Juglans regia), dues espècies de fusta i fruit molt apreciats que han arribat, de la mà dels humans, fins a l’extrem occidental d’Euràsia (Península Ibèrica i illes Britàniques) després d’haver migrat a través de tot el continent asiàtic i l’Europa meridional. L’estrat arbustiu també és ric en espècies; entre les més corrents hi ha l’avellaner manxú (Corylus mandshurica) i l’araliàcia Eleutherococcus [=Acanthopanax] senticosus.
Un tipus de bosc mixt semblant al descrit és el que cobreix l’illa japonesa de Hokkaidō, tret de la península meridional d’ōshima. Les coníferes dominants en aquest cas, però, són unes altres, principalment l’avet japonès (Abies sachalinesis) i la pícea de Hokkaidō (Picea jezoensis). Les espècies caducifòlies més freqüents, en canvi, són gairebé totes compartides amb el continent, com el roure mongol (Quercus mongolica), el roure dàimio (Q. dentata), l’om del pare David (Ulmus davidiana), el freixe manxú (Fraxinus mandschurica) i l’“itayakaede” (Acer mono).
A les boscanes nord-americanes, la zona septentrional de transició entre boscanes i bosc boreal amb presència de coníferes s’estén des del SE de Manitoba i el N de Minnesota, a l’W, fins a Maine i Nova Escòcia, a l’E, passant pel N dels Estats Units i el S del Canadà. És la regió de boscanes decídues més gran de l’E de l’Amèrica del Nord. Aquesta zona es caracteritza per una estació de creixement relativament curta, uns hiverns freds i nivosos i una geomorfologia d’origen glacial. Als indrets de característiques encara mesofítiques, la base del poblament arbori la formen espècies pròpies dels estadis avançats de la successió: erable de sucre (Acer saccharum), faig americà (Fagus grandifolia), bedoll groc (Betula alleghaniensis), tell americà (Tilia americana) i auró roig (Acer rubrum), i com a acompanyants l’om americà (Ulmus americana), el freixe blanc (Fraxinus americana), el freixe comú (F. nigra) i el roure roig comú (Quercus rubra). Entre les espècies pròpies dels estadis inicials de la successió destaquen el trèmol americà (Populus tremuloides) i el bedoll d’escorça de paper (Betula papyrifera) i, més localment, el trèmol de fulles dentades (Populus grandidentata), el bedoll gris (Betula populifolia) i el cirerer de Pennsilvània (Prunus pensylvanica). Les comunitats on dominen les coníferes corresponen als ambients de clima més extrem de la regió, que generalment coincideixen amb les latituds més septentrionals i les cotes més elevades.
Les boscanes atípiques
En condicions que s’allunyen de les òptimes del bioma, a causa del clima o de les pertorbacions, apareixen encara sistemes forestals que es poden conceptuar com a boscanes. Moltes rouredes secundàries, en particular les de roure mongol de l’Àsia oriental, es podrien assignar a aquest grup, però fins i tot algunes comunitats primàries, com la majoria dels bedollars, en serien representatives. Quan les boscanes mixtes del N de la Xina i de Corea es destrueixen i l’espai que ocupen s’abandona, s’hi desenvolupen diversos tipus de boscos secundaris. El més comú, sobretot en indrets relativament eixuts, és el bosc de roure mongol (Quercus mongolica), dominat per espècies pioneres com el pollancre del pare David (Populus davidiana) i el bedoll blanc asiàtic (Betula platyphylla), dues espècies que germinen fàcilment a les clarianes assolellades.
Al N del Japó, semblantment, les rouredes de roure mongol o els rodals de pi roig japonès o akamatsu (Pinus densiflora) poden barrejar-se amb les fagedes en les exposicions o sobre els substrats menys favorables als faigs, o bé poden ocupar les àrees pertorbades.
El bedollar és un tipus de boscana que ocupa grans extensions de les àrees de clima temperat fred, per sobre del límit de les rouredes i per sota del bosc de coníferes boreal o muntanyenc, a gairebé totes les regions del bioma. Els bedollars acostumen a estar formats per rodals purs d’una o més espècies de bedoll (Betula), però de vegades els acompanyen altres arbres, principalment trèmols, salzes o moixeres. També apareixen en forma de rodals dins dels boscos boreals, tant a Euràsia com a Amèrica.
Les formacions arbustives i herbàcies
A causa del clima particularment humit i del substrat àcid i pobre, en moltes àrees del bioma de les boscanes decídues, principalment a les costes atlàntiques europees, es donen grans extensions mancades de vegetació forestal i amb predomini d’una vegetació arbustiva, les landes; hi predominen les ericàcies, principalment brucs (Erica) i bruguerola (Calluna vulgaris). Cap a l’interior del continent, sobretot a muntanya i sobre substrats àcids, també es donen comunitats equivalents i matolls on predominen genísties i falgueres. A l’altra banda de l’Atlàntic, a les costes nord-americanes, també apareixen comunitats parcialment similars, però diferenciades per la presència escadussera d’algun arbre, principalment de pins; a l’Amèrica del Nord, tal i com passa també a l’Àsia oriental, les comunitats arbustives amb predomini d’ericàcies són més muntanyenques.
Quan els sòls no són tan pobres, els espais desforestats del bioma de les boscanes decídues acostumen a sostenir, tret que siguin sòls cultivats o transformats més pregonament, formacions herbàcies d’elevada diversitat, amb predomini de poàcies i fabàcies. Com en el cas de les landes, només l’explotació permanent per part dels humans o dels seus ramats permet mantenir aquest tipus de vegetació que, abandonada, evoluciona força ràpidament cap a la reconstrucció de la boscana, seguint un procés de successió natural. El tipus d’explotació que n’assegura el manteniment permet diferenciar dues menes principals de formacions herbàcies: els prats de dall i els prats de pastura.
Les landes atlàntiques
Des del S d’Escandinàvia fins al N de la Península Ibèrica, incloses les illes Britàniques, seguint la costa i penetrant cap a l’interior sobretot a les illes Britàniques, al N d’Alemanya i als Països Baixos, s’estenen paisatges oberts i ondulats ocupats primordialment per una vegetació arbustiva baixa (d’alçada no superior a 1 m) o nana (alçada que no supera els 25 cm), dominada principalment per la bruguerola. És la “Heide” germànica, el “heath” britànic o la “lande” francesa.
La bruguerola (Calluna vulgaris) és una mata baixa de fulles petites i imbricades, disposades en quatre rengles al llarg de les branques i amb flors petites, rosades o violàcies, que apareixen entre la fi de l’estiu i el principi de la tardor. En els sòls ben drenats, al costat de la bruguerola creixen altres ericàcies de port similar com el bruc vermell (Erica cinerea) o el nabiu comú (Vaccinium myrtillus), i s’hi troben també, més o menys esparsos, arbustos més grans com el ginebre (Juniperus communis), la gatosa atlàntica (Ulex europaea), el bàlec (Genista balansae [=G. purgans]) o la ginesta d’escombres o gòdua (Cytisus [=Sarothamnus] scoparius); per sota de l’estrat d’arbustos baixos dominants encara poden créixer algunes mates rastreres parcialment tolerants a l’ombra com el nabiu vermell (Vaccinium vitis-idaea), l’èmpetrum negre (Empetrum nigrum) o la boixerola (Arctostaphylos uva-ursi).
En sòls més humits però superficialment airejats, almenys a l’estiu, s’hi fa una landa de bruguerola i bruc d’aiguamoll (Erica tetralix); quan la humitat és permanent, el bruc d’aiguamoll i el romaní de torbera (Ledum palustre) desplacen la bruguerola i la vegetació es fa més oberta, incorporant diferents poàcies com les molínies (Molinia), ciperàcies com les cotoneres (Eriophorum), i fins torberes d’esfagnes. Aquestes brugueres o landes higròfiles són els “moor” de les illes Britàniques, tan característics del paisatge atlàntic desforestat de vora els “heath”.
L’origen d’aquestes landes ha estat objecte de controvèrsia per a botànics i fitogeògrafs al llarg de més d’un segle, però sembla que tret d’algunes torberes humides molt mal drenades, totes ocupen espais que podrien estar ocupats per boscanes; han estat originades per la intervenció humana i mantingudes per la pastura continuada i els incendis repetits en àrees desforestades pobres en nutrients. L’abandó de les antigues pràctiques pastorals i la introducció de noves tècniques agrícoles ha reduït dràsticament l’extensió de landes de la majoria de països d’Europa occidental. Només a Escòcia, Irlanda i el N d’Anglaterra se’n conserven extensions importants, principalment per interessos cinegètics.
A la regió nord-americana de les boscanes, les formacions que presenten més similituds amb les landes atlàntiques europees són els “heath balds” dels Apalatxes, formacions d’ericàcies que creixen sobre substrats de roques àcides als vessants dels Apalatxes exposats als vents de l’W. Encara que les espècies dominants també pertanyin a la família de les ericàcies, són molt diferents dels brucs i brugueroles predominants a les landes atlàntiques europees. A baixa altitud, als “heath balds” dominen el rododèndron laurifoli rosa (Rhododendron maximum) i el llorer de muntanya (Kalmia latifolia), mentre que a cotes superiors predomina el rododèndron purpuri (R. catawbiense). A baixa altitud, les formacions que serien més semblants a les landes atlàntiques europees són les pinedes esclarissades conegudes com a “pine barrens” (literalment, ‘erms de pins’) de les costes de Nova Jersey, com també altres formacions semblants de Long Island i de les costes de Massachusetts i Nova Escòcia. Es tracta més aviat de rouredes i pinedes esclarissades amb un sotabosc baix d’ericàcies que creixen sobre sòls sorrencs i pobres, però també aquí les espècies i fins els gèneres dominants són ben diferents dels de les landes europees: diferents nabius (Vaccinium vacillans, V. angustifolium), Kalmia angustifolia, Gaylussacia baccata, Myrica pensylvanica i fins i tot pins com el “pitch pine” o pi de brea (Pinus rigida), i roures com el roure grèvol (Quercus ilicifolia) que es presenten agarrigats assolint escassament una alçada d’un parell de metres, als sòls més pobres de les anomenades “pine plains” de Nova Jersey. Hi ha formacions arbustives d’aquest tipus que, ja sense cap cobertura arbòria, apareixen encara a l’E i el S de Terranova.
Els matolls acidòfils de genísties i falgueres
Cap a l’interior del continent europeu les landes no penetren gaire, ja que la majoria d’espècies de les landes atlàntiques són poc resistents a les gelades. Només apareixen formant claps als vessants exposats als vents atlàntics de les muntanyes mitjanes de l’W d’Europa, des de les Ardenes fins als Pirineus i les àrees muntanyoses de l’interior de Catalunya. En aquestes àrees, talment com a les més meridionals de les landes, més que la bruguerola, predominen gairebé sempre fabàcies del grup de les genísties (Genista, Ulex, Sarothamnus) i sovint porten associats diferents brucs i falgueres.
El matoll de gòdua i falguera, resultat de la roturació de rouredes i fagedes seguida de pastura continuada i incendis repetits, es presenta com una formació arbustiva més alta que la landa pròpiament dita (uns 2 m). Aquesta és una formació dominada per la gòdua (Cytisus [=Sarothamnus] scoparius), que amb les seves tiges dretes i nombroses, sempre verdes, forma una estructura perenne per sota de la qual la falguera comuna (Pteridium aquilinum) constitueix grans tofes que, en indrets particularment maltractats, poden convertir-se en dominants i esdevenir un falgar. En alçada, ja en contacte amb l’estatge subalpí, es pot donar el matoll de bàlec (Genistapurgans), més baix però sovint més dens que el de gòdua.
Els prats de dall
La fisiognomia dels prats de dall és força homogènia tot i les variacions florístiques, ecològiques i geogràfiques que s’hi poden apreciar. Normalment, es tracta de formacions denses i esponeroses, exclusivament herbàcies, amb una forta dominància de poàcies, que en el moment de màxim desenvolupament assoleixen una alçada de prop d’1 m. Els prats de dall més típics són els de fromental (Arrhenatherum elatius), en els quals, al costat d’aquesta poàcia, en predominen també d’altres com el dactilis comú (Dactylis glomerata), i diferents poes (Poa pratensis, P. trivialis), festuques (Festuca pratensis, F. arundinacea, F. rubra) o raigrassos (Lolium perenne, L. multiflorum), com també una colla d’espècies herbàcies d’altres famílies: fabàcies com els trèvols (Trifolium pratense, T. repens) o les guixes de prat (Lathyrus pratensis); asteràcies o compostes com la margarida de camp (Chrysanthemum leucanthemum) o la barbeta (Tragopogon orientalis); umbel·líferes com la pastanaga borda (Daucus carota) o les pampes (Heracleum sphondylium); cariofil·làcies com la flor de cucut (Lychnis flos-cuculi); rubiàcies com les espunyidelles (Galium lucidum, G. pratense), etc. No és estrany, a més de dallats, aquests prats són objecte d’altres pràctiques agronòmiques, raó per la qual se’n tornarà a parlar a la part 3 d’aquest volum.
Cap a muntanya, i en punts particularment humits de la terra baixa, algunes ciperàcies (Carex verna, C. hirta) poden desplaçar parcialment les poàcies. Per altra banda, gràcies a la successió de floracions els prats de dall montans d’Europa són plens de flors de vistositat diversa durant més de les dues terceres parts de l’any, des de les molt primerenques de la grandalla (Narcissus poeticus), dels safrans de muntanya (Crocus), de les orquídies, etc., fins a les molt tardanes dels còlquics (Colchicum autumnale, C. speciosum) i altres espècies tardorals.
Els prats humits de pastura
En sòls més prims o menys humits, els prats que s’hi fan són menys esponerosos i no creixen prou per a permetre dues dallades l’any. Es tracta en aquest cas de prats rasos, bé que densos, que es mantenen com a tals amb una explotació que consisteix a sotmetre’ls a pastura. El predomini de les poàcies és menys palès que en els prats de dall i sempre es tracta d’espècies de menys alçada que el fromental. Són prats, per exemple, de plantatge mitjà (Plantago media) i càrexs (Carex glauca, C. verna) amb margall erecte (Bromus erectus), algun trèvol i les característiques eufràsies (Euphrasia), o bé de plantatge mitjà i festuca rogenca (Festuca nigrescens). Aquestes plantes dominants, sempre poc vistents, solen anar acompanyades d’un seguici d’esplèndida floració vernal en la qual apareixen representants d’innombrables famílies, fabàcies com l’astràgal dànic (Astragalus danicus) i diferents trèvols (Trifolium), rosàcies com les alquemil·les (Alchemilla) o les potentil·les (Potentilla), asteràcies com el milfulles (Achillea millefolium), calcides i cards (Cirsium), labiades com les prunel·les (Prunella), etc.