Les aigües continentals al domini de les boscanes
Els ecosistemes d’aigües continentals no existeixen aïllats de l’entorn. Encara que no hi hagi una correspondència exacta entre els trets que caracteritzarien una regió limnològica i la vegetació decídua, l’ecologia de llacs i rius del bioma de les boscanes decídues està íntimament lligada a les característiques generals d’aquest. Per això, alguns dels trets característics que es descriuen aquí serien igualment aplicables a altres biomes de clima temperat, encara que hi predomini una vegetació d’aciculifolis de fulla perenne com a la taigà (vegeu també vol. 8), o en determinades selves temperades (vegeu també vol. 6). Així, per exemple, el clima es caracteritza pel pas repetit, encara que no totalment regular, de pertorbacions atmosfèriques amb influència de masses d’aire polars, que aporten una quantitat d’aigua suficient (al voltant de la mitjana continental o per damunt) i ben repartida; aquesta aigua, però, només és utilitzable per la vegetació durant una part de l’any a causa de la important oscil·lació tèrmica anual. Els sòls, per la seva banda, en general són recents, de formació postglacial (entre 10 i 20 000 anys), cosa que també explica que, amb relació a altres biomes, la fertilitat sigui més elevada i la biodiversitat més baixa, aspectes aquests igualment aplicables a la vida aquàtica. Per altra banda, les precipitacions relativament copioses i el terreny desigual que caracteritzen la major part del bioma de les boscanes decídues accentuen la importància de les aigües continentals.
Els grans cursos fluvials
Alguns dels rius de més gran notorietat del món travessen el bioma de les boscanes o el seu curs hi transcorre en la seva totalitat. És el cas del Rin, el Danubi, el Vístula, el Sena, el Loira o el Tàmesi a Europa; o el Sant Llorenç, el Delaware o el Hudson a l’Amèrica del Nord; a la Xina, l’Amur recorre els confins septentrionals del bioma, i l’immens Iang- Tsé (Chang) els meridionals, mentre que altres grans rius com el Groc o el Yalu el travessen.
Molts dels rius del bioma de les boscanes decídues són relativament joves, amb una considerable acció erosiva; encara no han arribat a dissenyar configuracions tan complicades com les que hom troba en els antics rius tropicals, de poc pendent, que s’entretenen a jugar amb la selva i obtenen, pel que fa a la generació i el manteniment de la biodiversitat (vegeu també vol. 2, pàg. 144-166), resultats molt positius tant per al riu com per a la vegetació perifèrica. En general, els rius més joves de les regions temperades i glacials no s’entretenen amb meandres complicats o no han tingut temps per a generar canals d’evacuació de traçat molt sinuós. Els seus recorreguts corresponen a una situació relativament més jove, quan el riu encara té major pendent i major capacitat d’erosió i de transport. Això fa doblement interessant l’estudi dels materials dissolts, en suspensió o arrossegats per l’aigua, perquè són testimonis del que pot sortir de la conca i, indirectament, indicadors de la seva ‘salut’ i de tot allò que reflecteix les propietats dels sòls, de la vegetació i de la vida en general.
L’interès inicial es convertí aviat en tot un programa d’estudi. Cap a la dècada dels seixanta, Bormann i Likens iniciaren recerques en aquest sentit a l’Amèrica del Nord, a la conca del Hubbard Brook, un riuet a Nou Hamsphire. Aquests estudis, que posen en relleu el significat de l’íntima interacció entre aigua i sòl sota la influència dels microorganismes, bacteris i fongs i de la vegetació superior, no han estat gaire imitats en altres llocs, probablement perquè demanen molta paciència i constància. És escaient, però, que hagin començat en latituts temperades, de marcada fluctuació durant l’any, perquè això pot ajudar a entendre més directament la forta variació estacional que pateixen rius de les regions de latitud mitjana, bàsicament induïda per l’acumulació de neu durant l’hivern i la crescuda que es produeix quan es fon. Aquest ritme imposat és característic dels sistemes fluvials de la regió temperada i, a més, cal afegir que la conservació d’una reserva suficient de biodiversitat en els sòls de lleres i ribes i damunt els palets i les roques és essencial per al funcionament biològic normal del sistema fluvial. La llera d’un riu és un exemple perfecte de les relacions entre l’arxiu d’una immensa biodiversitat i l’expressió, en cada moment i en cada lloc precís, de la diversitat existent.
El fluir de l’aigua és el factor més important per a la vida als rius i l’evolució natural en els organismes fluvials tendeix a afavorir aquelles característiques que permetin disminuir la taxa de multiplicació però mantenint l’ocupació d’un determinat tram de riu. És cert que la velocitat de l’aigua condiciona fortament la vida als rius de manera que en la massa d’aigua en moviment, on viuen en suspensió organismes de vida breu i multiplicació ràpida, les condicions canvien més de pressa; per gran que sigui la velocitat de l’aigua, però, cap riu no està mancat de vida. Alguns peixos fins i tot es mouen contracorrent cap a les aigües de capçalera on hi ha menys competidors i, per tant, aquests trams inicials dels rius són ecosistemes molt productius on la competència és baixa. Per altra banda, el mateix substrat està subjecte a l’operació selectiva del corrent, que s’emporta aigües avall tots els palets que són massa petits per a resistir la riuada. Hi ha una gradació dels materials i una selecció positiva de les característiques dels pobladors que faciliten l’adherència a les pedres i, al mateix temps, l’obtenció d’aliment. Madeixes d’algues fluctuant en el corrent, animals que tracten de mantenir el seu lloc de maneres molt diverses (per exemple, adherint-se al substrat mitjançant coixinets de seda, o dotant-se d’un llast en forma de tubs fets de pedretes enganxades també amb seda, etc.), xarxes o ventalls estesos en el corrent per diferents organismes, etc; aquestes són adaptacions que permeten una supervivència parcial, però sempre acaben fallant en algun moment, com ho prova l’anomenada deriva, que és aquella fracció d’organismes que l’aigua s’emporta avall sense remei. La deriva creix enormement amb les riuades i el manteniment del poblament requereix que sigui compensada a nivells més alts per una capacitat de multiplicació suficient.
Així com el plàncton de mars i llacs ha evolucionat sota la pressió d’haver de compensar el risc de sedimentació, les poblacions suspeses en el riu, l’anomenat potamoplancton, han de compensar l’efecte de la deriva o arrossegament pel corrent. Es pot utilitzar la mateixa aproximació quantitativa, substituint la dimensió vertical per una altra de més o menys horitzontal. Si hi troben l’aliment adient, una de les possibilitats que tenen alguns organismes, sense que això exclogui del tot el risc d’arrossegament en cas de riuades particularment intenses, és introduir-se en el si del sediment del riu. De fet, el riu conté una vida paral·lela en els intersticis entre els components sòlids del seu llit; aquí es troben animals de menes molt diverses que fan de pont entre la vida d’aigües superficials i la pròpia d’aqüífers i aigües de coves. Aquesta fauna, no cal dir-ho, pateix molt les conseqüències de la contaminació, que esgota l’oxigen que hi havia en aquest hàbitat intersticial.
Els llacs i els estanys
Al bioma de les boscanes també hi ha incomptables rius, rierols i estanyols més petits, juntament amb uns quants llacs enormes que són veritables mars interiors d’aigua dolça, com ara els Grans Llacs nord-americans. De fet, la limnologia, la ciència que s’ocupa de l’estudi de les aigües continentals, nasqué en bona mesura a partir de recerques sobre els llacs del domini de les boscanes.
Aquests llacs estan molt influïts pel ritme estacional. La forta oscil·lació tèrmica anual implica la formació de glaç a l’hivern, si més no a la superfície. Atès que la màxima densitat de l’aigua es dóna a 4°C, és obvi que en les masses d’aigua profundes l’oscil·lació tèrmica anual a l’entorn d’aquest valor determina dos períodes d’estratificació anual en els llacs de les regions temperades, un a l’estiu, durant el qual la temperatura en superfície és poc o molt superior als 4°C, i un altre a l’hivern, en què és inferior i sovint es forma glaç. Per tant, hi ha dos períodes de transició, a la primavera i a la tardor, durant els quals es produeix la circulació i la barreja vertical de l’aigua; això possibilita el retorn corresponent de nodriment mineral per dues vies: per ascensió d’aigua del fons en què està dissolt o per barreja vertical; aquesta repetició de la barreja vertical anual en les aigües lliures fa que la fertilitat dels llacs de latituds temperades pugui ser més elevada que en localitzacions polars o tropicals.
L’erosió glacial ha excavat les conques de molts llacs, més aviat petits, la fondària efectiva dels quals acostuma a ser inferior a 100 m. En valls anteriorment ocupades per antigues geleres, amb la sortida tancada a causa de les morenes glacials, se situen actualment un nombre considerable de llacs més extensos. Aquesta mena de llacs són més profunds (fins a 400 m o més) i són nombrosos als Alps, a Escandinàvia, en alguns àrees de les illes Britàniques (el Lake District anglès), la Península Ibèrica (llac de Sanàbria) o l’Amèrica del Nord, i en entorns equivalents d’altres serralades, per exemple, a la regió araucana dels Andes, entre Argentina i Xile, a les latituds compreses entre 39 i 42°S. Val a dir que en cadenes muntanyoses més antigues, com els Pirineus, l’erosió ha anat més enllà: les morenes terminals de les valls majors han estat arranades i els llacs principals han desaparegut al seu darrere; queden, malgrat tot, petites depressions lacustres en les parts més altes, entre 1 800 i 2 600 m d’altitud, l’origen de les quals es troba en altres formes d’erosió glacial. Dintre d’aquesta mateixa regió climàtica existeixen llacs en grans depressions tectòniques, com els vuit Grans Llacs nord-americans, que acullen el respectable volum de prop de 32 000 km3.
Els sòls relativament immadurs, molt comuns al bioma, són un factor d’aportació de nodriment i d’equilibri de l’alcalinitat en les conques per la seva notable capacitat de retenció de nutrients durant l’hivern. Aquest fet afavoreix el manteniment d’unes condicions que durant molts anys s’han considerat estàndard en la tipologia dels llacs. Això, però, es deu a la senzilla raó que la ciència limnològica es començà a desenvolupar a la regió temperada boreal i que, per tant, la limnologia arrossega encara en molts aspectes aquest biaix incorporat dels seus orígens. El concepte d’eutrofització, l’augment de la fertilitat de l’aigua que en fa disminuir la qualitat per a diferents usos, que s’acostuma a formular sobre la base del que s’esdevé en els llacs marginals alpins, també forma part de la mateixa tradició eurocèntrica. S’ha criticat que la imatge plàcida dels llacs centreuropeus, popularitzada en tantes belles fotografies de calendari, s’hagi erigit gairebé en model universal del funcionament dels llacs, però això era inevitable si es pensa que fou a l’Europa continental (principalment a Alemanya, Suïssa i Escandinàvia) on, com tantes altres coses més o menys bones, nasqué la limnologia. Això sense comptar que problemes com l’eutrofització, que nasqueren a Europa, s’han anat manifestant després en altres llocs.
La manifestació externa, més o menys estereotipada, de l’eutrofització resulta d’una entrada intensificada de nodriment, que es tradueix en un increment de la fertilitat i de la productivitat de plàncton de l’aigua. La descomposició d’aquest plàncton en el cicle de retorn en aigües profundes, ja fora de l’abast de la llum, consumeix una quantitat indesitjablement elevada d’oxigen; de vegades el consum és tan elevat que la quantitat que en conté l’aigua resulta insuficient. Això fa que l’aigua continuï retenint substàncies pudents i estimuli el desenvolupament, excessiu, o si més no indesitjable, de microorganismes. Al mateix temps, un material orgànic reduït, negre, va a parar al sediment, on primer s’intercala en forma d’estrats foscos, i acaba després per ennegrir tota la massa dipositada. En cilindres de sediment extrets amb un mostrejador tubular es veuen els progressos de l’eutrofització; comença amb estrats prims i foscos, estivals, que cada any augmenten de gruix fins que el sediment acaba presentant-se totalment ennegrit.
L’eutrofització correspon també a un efecte de vàlvula que es produeix en els estrats superficials del llac. La forta assimilació i producció vegetal que es dóna en aquests estrats té com a conseqüència una sobresaturació d’oxigen, prou persistent per a impedir el pas gradual i progressiu d’oxigen de l’atmosfera cap avall que es produiria si no hi hagués aquesta barrera a les aigües superficials. La pèrdua d’oxigen de l’aigua del fons fa minvar la seva qualitat per als usos que tenia. L’únic remei —costós i parcial— consisteix a injectar oxigen o aire a pressió. La barreja vertical hivernal és turbulenta i destrueix aquest efecte de vàlvula, però sol ser parcial i la fondària fins a la qual arriben els seus efectes depèn de la temperatura mínima de l’hivern corresponent. Els hiverns més freds aconsegueixen que la barreja es propagui fins a les proximitats dels fons o fins al mateix fons, mobilitzant el nodriment (principalment compostos de nitrogen i fòsfor) eventualment acumulat en fondàries a les quals l’aigua no arribà a barrejar-se en temporades anteriors. Això es fa palès en llacs italians del S dels Alps (Maggiore) o en altres llacs dels confins meridionals de les boscanes europees (Ohrid); al voltant de la Mediterrània, els hiverns del anys 1951, 1957, 1963 i 1970 foren freds i, en aquests llacs, l’aigua circulà fins al fons ocasionant produccions primàries excepcionalment elevades.
El cicle normal de la vida en un llac es pot descriure a partir del moment de la fusió del glaç. El desglaç fa augmentar la densitat de població d’organismes i, seguint l’augment de la temperatura en superfície fins a 4°C, la densitat de l’aigua també augmenta i la circulació vertical s’intensifica, aportant nodriment als estrats il·luminats. S’hi multipliquen moltes diatomees, com Asterionella, Fragillaria, Synedra, Aulacosira, Cyclotella, Stephanodiscus i altres. A poc a poc s’incrementa la multiplicació d’organismes fotosintetitzadors d’altres grups; cloròfits, dinòfits i euglenòfits i, sovint, cap a la tardor, cianobacteris, que són fixadors potencials de nitrogen gasós en forma orgànica. Mentrestant, l’aigua s’ha escalfat per damunt de 4°C, s’ha estratificat per densitats i els estrats més superficials van exhaurint el seu contingut original en nutrients essencials (sobretot nitrogen i fòsfor). Paral·lelament, té lloc la multiplicació del zooplàncton, començant pels rotífers que filtren i retenen organismes i partícules orgàniques petites en general, després els cladòcers, igualment filtradors, i, finalment, els copèpodes; aquest ordre de dominància consecutiva constant és correlatiu amb la duració creixent de la vida individual en els diversos grups, des de pocs dies en els rotífers fins a unes quantes setmanes, o fins i tot mesos, en els copèpodes. Sovint, la ràpida multiplicació de les Daphnia (cladòcers) en primavera fa, o feia, una bona funció de neteja de fitoplàncton, i dóna lloc a un interval de temps durant el qual l’aigua del llac és particularment transparent.
La formació hivernal de glaç en superfície limita seriosament la composició i l’expansió de la flora vascular litoral, aturant pràcticament la seva activitat vegetativa durant l’hivern. Aquesta flora queda destruïda en bona part i, en qualsevol cas, el glaç limita la diversitat d’espècies en comparació amb el nombre de les que viuen en vores de llacs i zones humides de zones més càlides de la biosfera. En compensació, el desenvolupament de torberes, dominades per molses, pot ser considerable en aigües molt poc mineralitzades, encara que siguin molt fredes.
Molts dels organismes que han estat esmentats es difongueren amb especiació ràpida al final de l’època glacial; alguns inclouen copèpodes del grup Cyclops estrenuus i peixos com els corègons (Coregonus), i salmònids molt variats propis de llacs europeus i nord-americans. Tanmateix, tot i l’innegable caràcter recent o jovenívol de l’ocupació de la major part dels llacs de les regions temperades, no falten els que, per unes o altres circumstàncies històriques, generalment per una llarga persistència de la massa d’aigua dolça, han permès la supervivència de formes de vida prou antigues i suposadament extingides en altres àrees geogràfiques, i acumulen, per tant, un gran nombre d’endemismes. El paradigma d’aquests llacs seria el llac Baikal, però també cal esmentar el llac d’Ohrid, als Balcans, que és una mena de museu entre els llacs europeus per la conservació de molts organismes de nissagues pre-glacials, o el llac Biwa, del Japó, que, si bé ha experimentat una recolonització postglacial, té un registre sedimentari que es remunta a fa més de 2 milions d’anys.
El llac Biwa és excepcionalment paradigmàtic pel que fa a la capacitat d’acumulació de testimonis del passat que tenen els llacs, ja que, com resulta fàcilment imaginable, en una zona com la del bioma de les boscanes decídues, les alteracions provocades per les glaciacions han esborrat generalment els testimonis que s’haguessin pogut dipositar abans. Tot i això, fins i tot en llocs on fa molts milers d’anys que no hi ha aigua, els sediments corresponents a fons de llacs antics tenen sempre una història per contar. Un bon exemple seria el paquet d’estrats d’origen lacustre que omple bona part de la depressió de la Cerdanya, als Pirineus catalans, que conté testimonis de la vida i vicissituds del llac que ocupà aquella conca fa uns 5 milions d’anys. Parlar-ne aquí es justifica perquè la seva flora fòssil, de la qual es conserven restes en els sediments que corresponen a les ribes de l’antic llac, mostra una bona representació de caducifolis. El sediment de l’antic llac està perfectament estratificat i en bona part consisteix en valves silícies de diatomees, comparables a les dels dipòsits d’alguns llacs africans recents. Això permet deduir que el fons no contenia animals que pertorbessin la deposició de subtils estrats anuals i, per tant, el fons devia ser anòxic i amíctic, és a dir, les aigües del llac no es barrejaven gaire en sentit vertical i, per tant, en superfície devien ser relativament i persistentment càlides. En els mateixos sediments queda reflectit un interessant episodi de deposició massiva de fosfats, tal vegada induït per un ingrés sobtat de grans quantitats de calci en el llac; talment com si les aigües d’alimentació haguessin començat a erosionar sobtadament roca calcària en alguna part de la conca que drenaven. Es pot avaluar la velocitat de sedimentació en el llac fòssil de la Cerdanya en 1 m de gruix per cada 6 000 anys.
L’estudi de sediments lacustres ha ajudat a reconstruir el passat i s’ha descobert que és molt interessant per a precisar el moment d’ejeccions de volcans propers, o, entre altres coses, la data de construcció de calçades romanes prop de llacs italians. Restes de diatomees, altres algues, closques de rizòpodes i grans de pol·len en sediments lacustres i en torberes il·lustren sobre canvis recents en el país, siguin glacials o postglacials. Ara s’ha trobat una altra utilitat a l’examen minuciós d’aquests canvis ecològics contemporanis a petita escala, amb relació a la distribució i la intensitat dels efectes de les pluges àcides. Amb aquest propòsit de control, recentment s’ha organitzat una xarxa de seguiment d’un conjunt de llacs de muntanya sobretot d’Europa.
La interacció entre les boscanes i els sistemes límnics
Les boscanes decídues, com tots els altres boscos, actuen com a reguladors naturals dels sistemes d’aigua dolça. La coberta arbòria redueix l’impacte físic de la pluja i la neu, mentre que les arrels extreuen una determinada proporció d’aigua del sòl i eviten l’erosió en cas que es produeixin precipitacions extremes. Així, les entrades d’aigua a rius i rierols són regulades amb molta més eficàcia que en el cas que no hi hagués arbres. Sovint, les inundacions i l’erosió catastròfiques són el resultat de la pèrdua de bosc; catàstrofes d’aquesta mena han impulsat la reforestació de diferents àrees de països com Àustria, Suïssa, Xina o Japó.
Tant els arbres vius com els morts proporcionen un hàbitat als animals d’aigua dolça. Les arrels dels arbres que s’estenen dins de l’aigua proporcionen refugi a alguns peixos, com la truita (Salmo trutta) a Europa, mentre que la part aèria dels arbres proporciona un emplaçament segur per al niu a diferents ocells aquàtics, com el bernat pescaire (Ardea cinerea) a Europa, l’ànec mandarí (Aix galericulata) a l’Àsia oriental i l’ànec de Carolina o ànec boscà nord-americà (A. sponsa). Potser encara més crucial, tot i que sovint menys reconegut, és el paper dels branquillons i branques que, en caure a l’aigua, creen una àmplia diversitat d’hàbitats per a animals aquàtics més petits i per a la fresa dels peixos. Semblantment, els grans troncs caiguts en llacs o rius proporcionen una àmplia gamma d’hàbitats a plantes i animals, entre altres, per exemple, els diminuts briozous, que formen colònies cespitoses (Cristatella) o arborescents (Plumatella) sobre les superfícies d’aquests troncs i les de tota mena de materials morts d’origen vegetal. Altres animals aprofiten les restes d’arbres i fulles com a aliment, i les larves de moltes frigànies (tricòpters) prenen fragments de branquillons i altres materials per a la construcció dels seus estoigs de protecció característics; a l’Europa occidental, per exemple, la larva de Limnephilus rombicus es construeix l’estoig amb petits trossos d’arrel que talla de la mateixa llargada, mentre que la de Glyphotaelius pelludicus fa servir trossos de fulles mortes. L’ombra que proporcionen els arbres també pot ser crucial, sobretot per als estanyols més petits quan, a l’estiu, l’escalfament excessiu els podria causar problemes.
De vegades, els arbres tenen l’efecte contrari, és a dir, el de reduir la varietat d’éssers vius en algunes masses d’aigua dolça, convertint-les en hàbitats exclusivament adequats per a espècies altament especialitzades. La caiguda massiva de fulles en estanyols petits pot comportar una pèrdua d’oxigen de l’aigua i, per tant, convertir-la en inadequada per a la vida de moltes plantes i animals, excepte per a aquells que viuen a l’aigua però respiren l’aire de la superfície. Aquest seria el cas d’alguns heteròpters com els barquers (Corixa) i les vespes d’aigua (Notonecta), o el de larves com els cucs de comuna o cucs de cua de rata (larves d’Eristalis), que tenen un tub llarg de respiració que arriba fins a la superfície i els permet viure en aigües molt contaminades (i també en petites acumulacions d’aigua amb poc oxigen o gens, en troncs d’arbres i troncs caiguts).
Les aigües dolces, al seu torn, també proporcionen recursos a les boscanes, fent variar radicalment la seva composició específica. Fins i tot estanyols força petits poden resultar vitals com a punt d’abeurada per a alguns animals, i encara resulta més important el paper de les aigües estancades i corrents, que ajuden a emmagatzemar una aigua que assegura la supervivència dels arbres en cas d’una sequera prolongada. Molts animals de les boscanes cacen en aigües dolces, per exemple l’àguila pescadora (Pandion halietus), un rapinyaire que s’alimenta de peixos i està molt estès tant per l’Amèrica del Nord com per Europa. Falciots (Apus), orenetes (Hirundo) i avions (Delichon) s’alimenten d’insectes voladors que surten de nimfes i larves aquàtiques, fent vol rasant amb la boca oberta per damunt de l’aigua.
La fauna aquàtica
En la fauna de les aigües dolces del bioma de les boscanes hi ha representats la majoria de fílums d’animals i de protoctists consumidors (protozous), alguns mai establerts en el medi aeri. És una fauna rica i abundant, en alguns casos àmpliament evocada per l’imaginari col·lectiu i, per tant, reflectida en la creació literària.
Els protoctists i els invertebrats dulciaqüícoles
La major part de les espècies són invisibles a ull nu, com ara els protozous, que són unicel·lulars, i els microscòpics rotífers; tant els uns com els altres són predadors o s’alimenten de matèria orgànica morta i inclouen tant representants mòbils com sedentaris. Més grans, però no pas per això gaire més sofisticades, són les esponges o porífers, els briozous i les hidres. Les hidres, que són celenterats d’aigua dolça, tenen una base que els permet fixar-se al substrat i disposen també d’entre 6 i 8 braços que utilitzen a tall d’arpó per a disparar unes cèl·lules especials, carregades d’una substància tòxica, que s’anomenen nematocists i que paralitzen i atrapen la presa; davant de qualsevol perill reculen ràpidament fent capgirells.
També cal esmentar els platihelmints (cucs plans) i els anèl·lids (cucs segmentats). Aquests grups, essencialment marítims, tenen representants adaptats a les condicions de vida de les aigües dolces, però no hi assoleixen els mateixos nivells de diversitat que a la mar. Normalment, els platihelmints de vida lliure de les aigües dolces (n’hi ha una colla que són paràsits d’organismes aquàtics o que hi passen almenys una part del seu cicle vital) fan com a màxim de 20 a 30 mm de llargada i tenen una gran capacitat de regeneració. La majoria dels anèl·lids d’aigua dolça són oligoquets o hirudinis (sangoneres); entre els primers destaquen els tubífexs (Tubifex i gèneres afins), que es caracteritzen per ser dels pocs invertebrats que tenen hemoglobina; els segons, malgrat la seva mala reputació com a paràsits i xucladors de sang, poden ser pares modèlics, i algunes espècies protegeixen els joves penjant-los sota els cossos dels adults. Uns quants platihelmints i sangoneres colonitzen àrees terrestres humides, incloent-hi els boscos. Tot i que són més comunes als tròpics, es troben diverses sangoneres terrestres als boscos temperats, on s’enganxen als animals que passen.
En un nivell més elevat de l’escala evolutiva hi ha diversos grups d’artròpodes, en particular els crustacis, la mida dels quals oscil·la des dels minúsculs cladòcers o puces d’aigua (Daphnia i gèneres afins) i copèpodes com els Cyclops, fins als gàmmars (amfípodes) i crancs de riu (decàpodes). El cranc xinès d’aigua dolça (Eriocheir sinensis), àmpliament distribuït, viu als rius, però a l’estiu es trasllada a aigües salabroses per fresar. Els gàmmars (Gammarus, Echinogammarus), els únics amfípodes amb representants veritablement dulciaqüícoles, tenen espècies ben adaptades a tota mena de graus de salinitat de les aigües; en un mateix estuari poden conviure diferents espècies, cada una amb la seva particular adaptació a la salinitat, que es traslladen amb els moviments de les marees per mantenir-se sempre en aigües amb nivells de salinitat apropiats.
La majoria dels insectes d’aigua dolça només passen les seves etapes juvenils sota les aigües o, si hi romanen d’adults, ho fan canviant el seu sistema de respiració de l’aire atmosfèric. Entre les nimfes de vida aquàtica més conegudes es troben les de libèl·lules i espiadimonis (odonats), efímeres (efemeròpters) i perles (plecòpters). Els adults, força colorits, són només la culminació breu d’una vida que en comptats casos pot durar un parell d’anys o més. Les efímeres adultes no tenen ni tan sols òrgans bucals per menjar, ja que la breu durada de la seva vida i les urgències de la reproducció no els donen ocasió d’alimentar-se; el cicle embrionari, en canvi, normalment dura sis mesos o un any. Les libèl·lules adultes, magníficament adaptades a caçar al vol, tampoc no viuen més d’unes quantes setmanes, que sovint passen voltant per clarianes dels boscos a poca distància de l’aigua. La majoria de les nimfes d’efímeres viuen en rierols de corrent ràpid, on s’arrosseguen entre les pedres mirant d’evitar que se les emporti el corrent, però algunes l’aprofiten en les seves migracions riu avall. Algunes espècies d’efímera tenen nimfes de vida lacustre, com ara les del gènere Cloeon, que són nedadores ràpides i elegants. Els insectes plenament aquàtics inclouen coleòpters (escarabats d’aigua) i heteròpters (xinxes d’aigua, teixidors, barquers, sabaters, escorpins d’aigua). La majoria d’espècies conserven la capacitat de volar i, a la nit, sovint es mouen d’una bassa o rierol a un altre.
Dins de la classe dels aràcnids, algunes aranyes s’han adaptat a la vida aquàtica, sia caminant sobre la superfície o, en el cas de l’aranya d’aigua (Argyroneta aquatica), àmpliament distribuïda, emportant-se bombolles d’aire al seu niu submergit. Més comuns són els àcars, minúsculs i sovint de colors llampants, que normalment tenen larves paràsites.
El darrer dels grans grups d’invertebrats dulciaqüícoles és el dels mol·luscs, únicament representat a les aigües dolces per bivalves i gastròpodes. A causa de les condicions de vida menys extremes, la majoria dels uns i dels altres tenen una closca més prima que la de les espècies marines corresponents (o fins i tot, en el cas dels cargols, que les terrestres).
Els vertebrats de vida aquàtica o amfíbia

Diverses espècies de peixos habiten les aigües dolces del bioma de les boscanes decídues. Entre els més abundants hi ha les perques (Perca), força comunes als rius nord-americans, però amb representants també als europeus, com Perca fluviatilis, que habita tant en aigües estancades com en rius de corrent ràpid. La família dels salmònids també hi està ben representada, amb el salmó (Salmo salar) com a espècie més característica. El salmó és, de fet, un peix anàdrom: viu a la mar però va a reproduir-se a les aigües dolces remuntant els rius (s’ha dibuixat un mascle fèrtil, amb la mandíbula característicament deformada en aquest període). La seva extraordinària habilitat per a salvar les cascades i altres accidents durant l’ascens riu amunt és ben coneguda. Un salmó gran, si disposa de profunditat suficient per agafar impuls, pot saltar fins a 3 m fora de l’aigua. La truita de rierol (Salvelinus fontinalis) és un altre salmònid dels rius de les regions temperades, que, procedent de l’Amèrica del Nord, ha estat introduït a les aigües dolces de pràcticament tot el món. A Europa hi arribà a final del segle XIX, juntament amb un altre salmònid, la truita arc iris (Salmo gairdneri), aquesta originària de la costa occidental del continent nord-americà. Emparentat amb els salmònids hi ha el luci (Esox lucius), distribuït per les aigües calmades i fredes de tot l’hemisferi septentrional. La família dels ictalúrids, a la qual pertany el peix gat Ameiurus melas, i la dels centràrquids són exclusives de les regions temperades nord-americanes. La segona inclou unes 160 espècies repartides en 9 gèneres. La perca americana de boca grossa (Micropterus salmoides), de fins a 83 cm, és el representant més gran de la família. Els ciprínids es troben a les tres regions del bioma, i la tenca (Tinca tinca) és un dels seus representants europeus. Altres peixos comuns són la lota (Lota lota), l’únic representant d’aigua dolça de la família dels gàdids, i el silur (Silurus glanis, vegeu també la figura 112).
Jordi Corbera
Entre els vertebrats aquàtics, els peixos són de bon tros els més comuns de tots. Quasi tots els peixos passen tota la seva vida a sota l’aigua, tot i que alguns fan breus incursions en el medi terrestre: les anguiles joves (Anguilla anguilla a Europa; A. rostrata a l’Amèrica del Nord; A. japonica a l’Àsia oriental), en la seva migració des de la mar fins als gorgs i rabeigs en els quals passen la major part de la vida (vegeu també vol. 10, pàg. 108-109), poden reptar ocasionalment pel sòl humitejat per la pluja o la rosada. En condicions naturals, la major part de les aigües dolces estarien envoltades de bosc, i molts peixos s’han adaptat a la presència d’arbres, utilitzant troncs morts i restes vegetals per a refugiar-se i reproduir-se, o aprofitant l’ombra dels arbres durant els estius calorosos. Els silurs (Silurus glanis) dels rius eurasiàtics, com els seus parents, els peixos gat americans (Ictalurus [=Ameiurus] nebulosus i espècies afins) dels rius de la zona oriental de l’Amèrica del Nord, quan s’aparellen construeixen un niu sota les arrels dels arbres i, després de la fresa, el mascle el vigila i defensa els ous fins a la desclosa.
Amfibis i rèptils viuen a cavall entre el terra i l’aigua; crien i passen la seva infantesa a l’aigua però sovint es traslladen a terra ferma —sempre a llocs humits— quan són adults. Alguns estan íntimament associats amb els hàbitats boscans, com ara les reinetes (Hyla), que solen viure en àrees boscoses i sovint s’enfilen molt amunt de les capçades dels arbres. A banda d’alguns rèptils, com les tortugues d’aigua europees (Emys orbicularis) o les nord-americanes (Pseudemys concinna i altres congèneres), completament aquàtics, moltes espècies de serps també són bones nedadores; tant la colobra de collar europea (Natrix natrix), com la serp de garrotera comuna nord-americana (Thamnophis sirtalis) viuen bàsicament en llocs humits, entre altres els boscos, i normalment se situen a prop de l’aigua.
Molts ocells tenen hàbitats que s’encavalquen entre boscos, rius i llacs. Diverses espècies d’ànecs, com ara el bec de serra gros (Mergus merganser) d’Europa i l’Amèrica del Nord, s’alimenten a l’aigua i es reprodueixen en forats dels arbres. El bernat pescaire europeu (Ardea cinerea) i diversos agrons nord-americans, com ara el bernat blau (A. herodias), s’alimenten pescant peixos i invertebrats en aigües poc profundes i es reprodueixen en colònies que instal·len en fragments de bosquines riberenques. La valona (Tringa glareola) hiverna als marges de l’aigua dolça però sovint es reprodueix als boscos, mentre que les becades, tant l’americana (Scolopax [=Philohela] minor) com l’europea (Scolopax rusticola), són essencialment ocells boscans que s’alimenten en àrees pantanoses. L’espectacular vol de parada de la becada és un tret característic que anima la vida de les boscanes decídues europees els vespres de primavera. Altres ocells d’aigua dolça del bioma o bé limiten els seus moviments gairebé totalment a rius i rierols, com ara les diverses merles d’aigua (Cinclus), que caminen al llarg dels llits dels rierols capturant minúsculs invertebrats, o bé migren entre hàbitats humits temperats i la tundra o les torberes àrtiques, com fan, per exemple, tètols (Limosa) i pòlits (Numenius).
Finalment, un grup relativament petit de mamífers del bioma temperat caducifoli s’ha adaptat a passar-se tota la vida, o una part, a l’aigua. Els més destacats d’aquests són els castors europeu (Castor fiber) i americà (C. canadensis), que són els que combinen més clarament la vida aquàtica amb un hàbitat forestat: tallen els arbres que creixen a les riberes i construeixen dics per mantenir un nivell d’aigua òptim per a protegir el niu que es fan enmig de l’aigua amb branques i branquillons. Els castors són comuns en llocs amb grans àrees de bosc pantanós, com les que es troben normalment a la part oriental de l’Amèrica del Nord i als estats bàltics de Letònia i Lituània. El visó europeu (Mustela lutreola) també es passa la major part de la vida dins l’aigua i al seu voltant, però sovint estableix el seu niu en un forat d’un arbre. Les llúdries (Lutra lutra), per la seva banda, se senten més atretes per emplaçaments on els arbres no arriben ben bé fins al marge del riu. L’ós bru (Ursus arctos), que viu al bosc, es banya regularment ja que és un bon nedador i fins i tot captura peixos amb les urpes.
La vegetació aquàtica i de ribera
Les plantes —i les algues, els protoctists fotosintetitzadors tradicionalment identificats com a plantes— també són versàtils a l’hora d’adaptar-se a l’aigua dolça. Innombrables espècies d’algues, en efecte, es troben a les aigües del bioma temperat caducifoli, i sovint semblen formar una massa verda al si de l’aigua. Entre les més atractives hi ha les diatomees, un grup d’algues unicel·lulars que tenen una coberta silicificada molt variable en estructura i coloració (vegeu també vol. 10, pàg. 61-63). Les cobertes de diatomees es conserven als sediments dels llacs per temps gairebé indefinit i, com que la composició de les espècies de diatomees varia amb les condicions de l’aigua, l’anàlisi d’aquests sediments permet fer-se una idea de la història del llac i ha estat utilitzada a Europa per a esbrinar l’impacte de la contaminació de l’aire en l’acidificació dels llacs.
La vegetació hidrofítica o helofítica
Moltes espècies de plantes fanerògames també s’han adaptat a les condicions de vida a les aigües dolces, tant en total immersió o flotant sobre la superfície de l’aigua (hidròfits), com romanent mig submergides a les ribes (helòfits). La imatge de les boscanes és indissociable de la pinzellada de verdor particularment higròfila pròpia dels corrents d’aigua de curs tranquil, dels rabeigs i dels llacs o estanyols. Des de les llentilles d’aigua (Lemna) fins als nenúfars (Nuphar, Nymphaea, Nelumbo), hi ha tota una cohort d’espècies interessants i bellíssimes.
A les aigües quietes de tolls i basses hom troba poblaments flotants de llentilles d’aigua (Lemna), d’azol·la (Azolla) i els petits briòfits del gènere Riccia, extraordinàriament semblants a les mateixes llentilles d’aigua. Aquestes presenten una morfologia molt peculiar, amb una làmina suradora de forma lenticular i una arreleta penjant que serveix no sols per a la funció nutrícia característica d’aquest òrgan sinó també per a mantenir la posició de la planta quan l’aigua es mou.
Les plantes que viuen en aigües corrents, en canvi, presenten adaptacions morfològiques molt diferents. A Europa, per exemple, el ranuncle aquàtic comú (Ranunculus aquatilis), que prefereix els estanyols, presenta dos tipus de fulles, les superiors, laminars, que suren sobre l’aigua al voltant de les flors, i les de la resta de la planta pràcticament reduïdes a la nervadura, finament dividides, i que es mouen de manera ondulant en la direcció del corrent; el ranuncle fluviàtil (R. fluitans), propi de les aigües ràpides, manca de fulles flotants. També hi ha diferències segons la composició química de l’aigua, amb plantes adaptades a condicions bàsiques i altres a condicions àcides: el ranuncle aquàtic de fulla rodona (R. omiophyllus) prefereix els estanyols àcids de les torberes, el ranuncle aquàtic calcífil (R. pencillatus var. calcareus) és exclusiu dels rierols calcaris, mentre que la creixenassa (R. baudotii) creix en aiguamolls litorals.
Macròfits de les aigües continentals
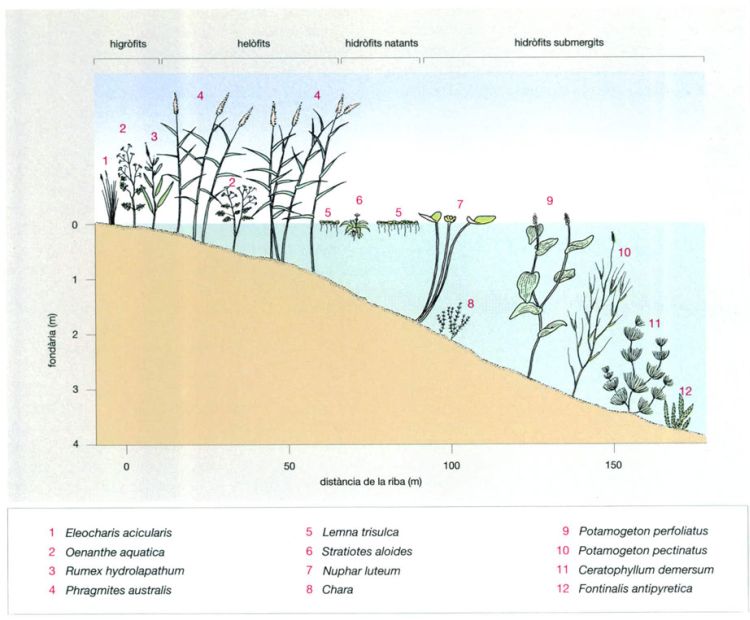
Jordi Corbera, a partir de fonts diverses
Les diferents comunitats de macròfits que creixen a les vores dels llacs de les boscanes decídues marquen la transició dels ambients de terra ferma als de l’aigua lliure. A alguns macròfits els cal estar arrelats al fons, i per tant creixen només a la zona litoral, on les aigües són someres. Normalment es tracta de plantes altes que surten fora de l’aigua, com els canyissos (Phragmites). Se’ls anomena macròfits emergents, per a diferenciar-los de les espècies flotants i submergides característiques de les aigües més profundes. La major part de macròfits són angiospermes, però també s’hi troben algunes grans algues (per exemple Chara) i briòfits. Més cap a l’interior del llac apareixen fulles flotants; unes pertanyen a plantes que suren lliurement, com Stratiotes, mentre que altres són de plantes arrelades al fons però amb llargues tiges que arriben a la superfície, com per exemple Nuphar. Algunes espècies tenen també les fulles submergides, i només les flors treuen el cap fora de l’aigua. Encara més endins hi ha espècies que viuen totalment submergides, bé arrelades al fons, com els potamogètons (Potamogeton) i els ranuncles (Ranunculus), bé surant lliurement, com per exemple les llentilles d’aigua (Lemna).
Les comunitats helofítiques, és a dir, de plantes herbàcies que arrelen a l’aigua però que tenen parts aèries ben dreçades, caracteritzen les ribes d’estanys i llacs. Canyissars, jonqueres i herbassars esponerosos poblen en anelles concèntriques aquestes ribes i, allí on les aigües són àcides i pobres en nutrients, dominen les torberes. A les regions més humides de l’Europa atlàntica, en particular Irlanda i algunes àrees de Gran Bretanya, en medis més rics en nutrients i fins i tot sobre materials calcaris, també es donen torberes.
Les boscanes de ribera
Les boscanes higrofítiques caracteritzen els sòls humits o inundats, de manera més o menys permanent, de les planes al·luvials de les regions temperades. Les més importants són els boscos de ribera (pollancredes, vernedes, salzedes), estretament vinculats als cursos d’aigua. La transició entre aquests i les boscanes mesofítiques pròpiament dites la constitueixen comunitats d’arbres decidus (freixenedes, omedes i fins i tot algunes rouredes). Tots plegats són elements importants de la vegetació del bioma de les boscanes decídues i, en els boscos de ribera, ultrapassen els seus límits seguint els cursos d’aigua que penetren els biomes veïns, més eixuts, de les estepes i praderies o de les mediterrànies.
El cicle vital d’algunes espècies riberenques és influït pels alts i baixos del nivell de les aigües. A la zona oriental dels Estats Units, per exemple, les llavors del bedoll negre americà (Betula nigra) maduren al final de la primavera, just en el moment en què el retrocés de les aigües d’inundació deixa al descobert els marges de la ribera. Hi ha arbres, com els salzes, que creixen arran mateix de l’aigua i altres, com els verns (Alnus), que ho arriben a fer en àrees inundades, fins i tot als “fens” o torberes inundades per aigües riques en calcària, on la majoria d’arbres no poden sobreviure. Molts pollancres (Populus) de la zona temperada, com ara el “cottonwood” dels pantans nord-americans (P. heterophylla), també creixen en condicions força humides. A Itàlia, en àrees inundables de la vall del riu Po, el cultiu del pollancre gavatx (P. nigra var. italica) és una forma d’aprofitament tradicional.
De vegades, en el mateix espai, coexisteixen formacions forestals i medi aquàtic. Això passa, per exemple, a les boscanes inundades, molt riques biològicament, de Letònia i Lituània, a les quals, durant una gran part de l’any, els arbres emergeixen per sobre d’un gruix d’aigua d’alguns centímetres. Altres boscanes depenen per a la seva subsistència de les inundacions periòdiques de l’hivern; aquest és el cas dels singulars boscos de les illes del riu Potomac, que travessa la ciutat i el districte federal de Washington, la capital dels Estats Units. Diferents arbres, entre els quals nombroses espècies de salzes i algunes de vern i de pollancre, creixen preferentment en les condicions d’humitat de les riberes. Si el bosc és malmès, o si els rius són desviats o contaminats, ambdós ecosistemes pateixen les conseqüències de les malvestats infligides a l’altre.
Les salzedes i les pollancredes
Els boscos de ribera, dominats pels salzes i els pollancres, constitueixen les més importants i difoses de les boscanes higròfiles. Al llarg dels cursos d’aigua, amb diferents combinacions d’espècies dominants i acompanyants segons el risc d’inundació, la fondària i l’abundor del mantell freàtic, els boscos de ribera solen disposar-se a les ribes en bandes paral·leles.
Arran d’aigua, i de vegades dins i tot de la llera, hi ha les salzedes, molt variades pel que fa a composició específica, ja que les espècies de salze (Salix) són nombroses i la majoria s’hibriden sovint entre elles. Es tracta de bosquetons primordialment arbustius que formen estrets poblaments d’arbustos i arbrissons, principalment d’aquest gènere, al llarg dels cursos d’aigua de tot el bioma, tant a l’Europa occidental com a Àsia o a Amèrica. Fan un paper fonamental en l’estabilització de marges i riberes i en la protecció dels altres boscos de ribera menys soferts enfront de les riuades que les salzedes.
Una altra formació ben diferenciada són les pollancredes que es troben arreu d’Europa, a la vora de llacs i rius de règim regular; el pollancre (Populus nigra) també prospera molt bé en sòls pantanosos que, si són drenats, permeten el desenvolupament de boscos humits de roures. Freixes de fulla gran (Fraxinus excelsior) i bedolls pubescents (Betula pubescens) poden acompanyar els pollancres en aquests boscos de ribera i, en el cas dels bedolls pubescents, arribar a formar boscos monospecífics muntanyencs per damunt dels límits de rouredes i pinedes.
Les vernedes i les omedes
A les zones humides de sòls fèrtils al·luvials periòdicament inundats durant llargues temporades, es poden formar boscanes complexes constituïdes per una gran diversitat d’espècies (roures, oms, aurons, tells, freixes) sense que cap no sigui clarament dominant. També es poden formar, com passa a l’Europa atlàntica, boscanes, del tipus de vernedes i omedes, amb estrat arbori dominat per una sola espècie.
Les vernedes són un element important de la vegetació medioeuropea i atlàntica. Es troben íntimament lligades, no sols a les riberes de rius i llacs, sinó també als sòls entollats, on poden ocupar grans superfícies, i, sovint, a les torberes. Juntament amb el vern (Alnus glutinosa), solen aparèixer, a l’estrat arbori, el freixe de fulla gran (Fraxinus excelsior) i, a l’estrat arbustiu, l’avellaner europeu (Corylus avellana), el saüc europeu comú (Sambucus nigra), el sanguinyol comú (Cornus sanguinea), la fràngula (Rhamnus frangula), diferents salzes i gatsaules (Salix), com el salze cendrós (S. cinerea), i diverses espècies de riber (Ribes). A l’estrat herbaci es desenvolupen l’ortiga gran (Urtica dioica), la viola palustre (Viola palustris), la dolçamara (Solanum dulcamara), la cal·la o lliri d’aigua europeu (Calla palustris), la falguera de rei (Osmunda regalis) i la falguera cartoixana (Dryopteris carthusiana).
Les omedes no són tan exigents pel que fa a les condicions d’humitat edàfica, però tampoc no tenen problemes per a viure en terrenys periòdicament inundats. Així, generalment ocupen una banda més separada de les lleres dels rius que les salzedes i les pollancredes. A l’Amèrica del Nord, per exemple, a la zona de les boscanes de faig i erable de sucre, l’extrem més humit del gradient de boscanes de la regió correspon a un tipus de comunitat amb om americà (Ulmus americana), freixe comú (Fraxinus nigra) i erable argentat (Acer saccharinum), encara que l’om americà ha estat molt delmat, fins i tot en el seu hàbitat natural, per la malaltia holandesa.