El metabolisme de les boscanes
Arreu del món i a totes les latituds, els boscos són els ecosistemes terrestres on la vida es manifesta amb una diversitat més gran. Les boscanes decídues no són una excepció. Tanmateix, el manteniment d’aquesta diversitat exigeix una complexa trama de relacions entre les espècies, i d’aquestes amb els factors ambientals: producció i manteniment de matèria viva, circulació d’energia, aigua i nutrients, estratègies d’obtenció d’aliment o de defensa enfront els depredadors, reciclatge dels residus, etc. Les plantes, principalment els arbres, que són els que a les boscanes controlen tot el sistema, prenen energia de la radiació solar, aigua i nutrients del sòl i diòxid de carboni de l’aire. Amb aquests recursos sintetitzen els seus teixits i mantenen les seves funcions vitals. Com en tots els ecosistemes terrestres, part de la biomassa vegetal és consumida pels animals fitòfags o productors secundaris que, al seu torn, són consumits, almenys en part, pels carnívors. Les parts no consumides de la biomassa de productors i consumidors i les restes de les excrecions d’aquests darrers acaben tornant al sòl, on els descomponedors les mineralitzen i, així, tornen a posar els nutrients a disposició de les plantes.
El cicle dels nutrients

Els canvis estacionals de la vegetació de les boscanes decídues vénen acompanyats de canvis en l’ocupació dels hàbitats per molts dels animals que hi viuen i de canvis en el cicle dels nutrients i en el flux d’energia. A la tardor, la caiguda de les fulles aporta un volum més gran de matèria orgànica morta al sòl, que els organismes detritívors i descomponedors començaran a descompondre. És al final d’aquesta estació i principi de l’hivern que una part de la fauna es retira als hàbitats subterranis, de manera que alterna períodes de vida epigea amb períodes de vida hipogea. Durant tot l’hivern els animals segueixen enterrats, i el flux de nutrients s’alenteix. Quan arriba la primavera, els organismes tornen a pujar a la superfície i reprenen les seves activitats. El cicle de nutrients s’accelera, ja que els arbres comencen a treure fulla i els cal reutilitzar els nutrients que els descomponedors del sòl s’han encarregat de mineralitzar. El bosc experimenta aquests canvis de manera cíclica, i cada any a la mateixa època recupera el seu aspecte típic.
Jordi Corbera, a partir de Walter & Breckle, 1989
Els aliments de què es nodreixen els éssers vius de les boscanes decídues es poden trobar en dos estats: immobilitzats en diferents compartiments de l’ecosistema (sòl, sediments, fusta) o circulant entre aquests diferents compartiments. En general, es pot dir que cada àtom dels elements que constitueixen els nutrients alterna entre un estat i l’altre, i que el temps que passa en cada estat depèn de les característiques intrínseques de cada boscana. Aquest cicle és continu, és a dir, no té inici ni final. Tot i això, sembla enraonat prendre com a punt de partida els nutrients que es troben al sòl en forma d’ions i la seva absorció per les plantes, que els acumularan de manera duradora —en forma de fusta— o els incorporaran temporalment en materials caducs de vida més curta com ara fulles, flors, llavors o fruits. Al final, després de la descomposició i de la mineralització d’aquells materials, tots els nutrients acaben tornant al sòl, on queden a disposició de les arrels de les plantes per a ser assimilats.
Carboni, nitrogen, fòsfor, potassi, calci i magnesi són els principals elements de la biomassa vegetal i també els més importants dels nutrients. La seva absorció és una de les pedres angulars de la vida vegetal. A diferència del que passa a les selves equatorials, on el reciclatge és molt ràpid i sovint la major part dels nutrients es troben en la vegetació i no pas al sòl, a les boscanes decídues la importància del sòl en la distribució de nutrients és força més gran, encara que la vegetació mateixa en conté quantitats molt considerables. El total absorbit es reparteix entre la part que es queda retinguda a l’arbre com a material llenyós i la que l’arbre restitueix en forma de fulles i altres òrgans o produccions de renovació més ràpida (flors, fruits, arrels fines, etc.). L’absorció varia per a cada element en funció de l’espècie i l’edat de cada arbre; en general es pot dir que els arbres de creixement més ràpid (que a les boscanes decídues no són necessàriament els més joves) retenen molts més nutrients, mentre que els arbres vells, que ja acumulen poca biomassa nova, en retenen menys.
La quantitat de cada element procedent de la virosta que retorna al sòl és variable; en general, a la biomassa vegetal s’acumulen quantitats més elevades de calci, nitrogen i potassi que de magnesi, fòsfor i altres elements, però a les fagedes l’acumulació de nitrogen sol superar la de calci. En canvi, la velocitat d’alliberament del nutrient (en relació amb la seva quantitat) en el procés de la mineralització decreix segons un altre ordre: potassi, magnesi, calci, fòsfor i nitrogen. Consumada la mineralització, el retorn de nitrogen al sòl, que en general és el nutrient més important, supera els 50 kg/ha, prop d’un 5% del nitrogen total.
A banda d’aquest cicle pràcticament tancat hi ha entrades de nutrients de l’exterior, com el nitrogen atmosfèric fixat per determinats bacteris i fongs associats a les arrels d’algunes plantes, o productes d’alteració dels minerals del sòl. També hi ha pèrdues, la principal de les quals és la deguda a la lixiviació, procés en el qual l’aigua percola en profunditat fins més enllà dels horitzons del sòl accessibles a les arrels de les plantes. Amb relació a aquest cicle extern les entrades més importants són les de nitrogen (74,6 kg/ha en mesures fetes en boscanes centreuropees) i les de calci (71,5 kg/ha). Una part important d’aquests nutrients arriben per deposició aèria, ja sigui arrossegats per les precipitacions, ja com a resultat de la sedimentació de la pols, però en el cas del nitrogen també s’ha de tenir en compte la fixació del nitrogen atmosfèric per bacteris i fongs, tant simbiòtics de les plantes com no, font que pot ser particularment rellevant en determinades boscanes, com les vernedes.
El cicle de l’aigua
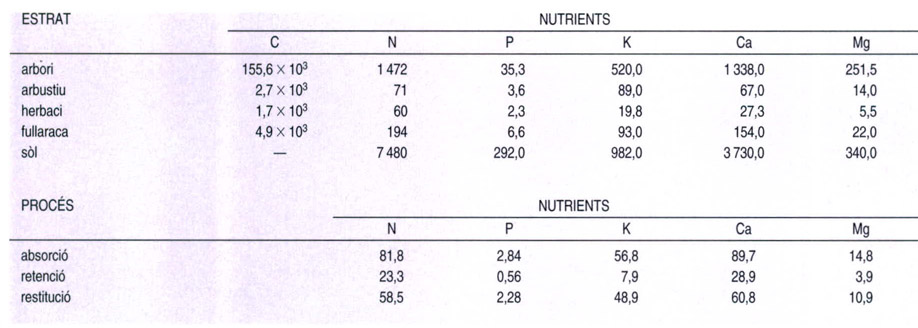
Abundància (en kg/ha) dels nutrients principals del sòl i als diferents estrats d’una boscana centreuropea (a dalt) i quantitats anuals absorbides de cadascun a nivell de l’estrat arbori, amb indicació de la part realment retinguda i la que es restitueix al sòl (a baix). La major part dels nutrients es troben a la matèria orgànica morta (al sòl i a la fullaraca), i dels que formen part de la matèria viva, la majoria es troben a l’estrat arbori, el més ben desenvolupat a les boscanes decídues. Cal remarcar la diferència entre les quantitats de carboni (C), de nitrogen (N) i les dels oligoelements fòsfor (P), potassi (K), calci (Ca) i magnesi (Mg), i com els arbres retenen menys de la meitat del total de nutrients que absorbeixen.
A partir de dades proporcionades per l’autor
La vida de les boscanes decídues, com la de tots els altres ecosistemes terrestres, no es pot entendre sense una activa circulació d’aigua, un factor limitador molt important, no tan sols per a les plantes dels boscos caducifolis, sinó també per a tots els organismes i els processos que en depenen. L’aigua, com altres components de les boscanes decídues, es comporta cíclicament, amb entrades, sortides i una circulació interna en l’ecosistema. L’entrada d’aigua al bosc és conseqüència de la precipitació natural que hi cau, sigui directament a terra o bé sobre la vegetació. L’aigua que és interceptada per les capçades dels successius estrats de la vegetació s’evapora parcialment abans d’arribar al sòl (en un percentatge que és inversament proporcional a l’abundor i la intensitat de la pluja), mentre que la resta circula cap al sòl lliscant per les branques i els troncs o hi degota directament des del fullatge. La major part de l’aigua que arriba a terra és absorbida pels horitzons superficials del sòl, gruixuts i flonjos, capaços d’absorbir l’aigua molt ràpidament, retenir-la i posteriorment alliberar-la segons les demandes del medi. Part de l’aigua que arriba al sòl, sobretot si aquest està saturat o no és porós i per tant és incapaç de retenir-la, s’escorre superficialment pendent avall i abandona el sistema; és una aigua que, al final, tancarà el cicle arribant a la mar a través de la xarxa fluvial. Normalment, però, és molt més important l’entrada d’aigua sòl endins, és a dir, la infiltració.
L’aigua d’infiltració es pot quedar en els horitzons superficials del sòl o bé penetrar més avall (aigua de percolació), constituint la reserva hídrica del sòl. Les plantes, tant en els horitzons superficials com en els profunds, n’absorbeixen una part, en una nova etapa del cicle de circulació interna de l’aigua a la boscana. Una gran part de l’aigua absorbida per les plantes, sobretot pels arbres i els arbustos, torna a l’atmosfera a través de la transpiració, que juntament amb l’evaporació directa del sòl i les capçades constitueix la sortida d’aigua més important del sistema. Aquesta aigua s’afegeix a la reserva d’aigua atmosfèrica procedent de l’evaporació de la mar, els llacs i altres superfícies d’aigua que acabarà tornant a entrar en el cicle de les boscanes i dels altres ecosistemes terrestres en forma de precipitació.
La transpiració de les fulles a través dels estomes és important perquè garanteix el flux d’aigua rica en nutrients a través de les cèl·lules de la planta, i perquè a més a més acompleix la funció de regular la temperatura. L’aigua, absorbida per les arrels, en efecte, s’utilitza per a transportar nutrients a la resta de la planta, i constitueix més de la meitat del pes de les cèl·lules vegetals. El transport és possible gràcies a la pressió osmòtica: la concentració més elevada de sals a les arrels fa que es desplacin a les parts més altes de la planta, on aquesta concentració és més baixa. La transpiració, però, i en conseqüència el moviment de la saba depenen igualment d’una sèrie de factors climàtics.
La temperatura és, potser, el més important d’aquests factors. En un dia càlid d’estiu una fulla pot transpirar en una hora el seu propi pes en aigua. A l’estiu, una fageda transpira diàriament l’equivalent d’una capa d’aigua de 4 mm de gruix per unitat de superfície, mentre els roures, que transpiren menys, ho fan a raó de l’equivalent d’una capa d’aigua de 2,3 mm per unitat de superfície. La transpiració, com a mitjà per regular la temperatura, varia evidentment segons si és de dia o de nit (la llum regula l’obertura i el tancament dels estomes) i, com és natural, segons les estacions, ja que cessa amb la caiguda de la fulla a la tardor i torna a començar a la primavera. La humitat de l’aire i el vent també són factors importants en la regulació de l’obertura o el tancament dels estomes a través dels quals es produeix la transpiració. Com més lluny és l’aire del punt de saturació, més elevada és la transpiració; el vent, en renovar l’aire que envolta les fulles, impedeix que aquest se saturi amb el vapor d’aigua provinent de la transpiració i per tant l’estimula.
L’economia de la producció
La producció primària d’un ecosistema depèn de la llum solar que rep, i aquesta depèn de la seva situació latitudinal. Les boscanes decídues reben menys llum que les selves tropicals o fins i tot que les sabanes llenyoses dels tròpics, on la llum incideix quasi perpendicularment. L’eficiència fotosintetitzadora, però, tant en boscanes com en selves o sabanes, és molt baixa, i en cap cas no s’arriba a acumular més de l’1,5% de l’energia lumínica en forma d’energia química de la matèria orgànica.
En termes generals, sembla que la fotosíntesi hauria de ser més eficaç a plena llum, però en realitat hi ha plantes, com ara les falgueres, que no toleren la il·luminació directa i encara menys la intensa radiació solar que hi ha al pic de l’estiu i que, en canvi, poden fer la fotosíntesi amb nivells de llum molt baixos. Són les plantes anomenades esciòfiles, o amants de l’ombra, a diferència de les heliòfiles, o amants de la llum. Es tracta de diferents respostes a la quantitat variable de llum de què es pot disposar dintre el bosc o, en general, de qualsevol comunitat vegetal i poden variar al llarg de la vida de l’arbre. Faigs (Fagus), aurons (Acer) i tells (Tilia), per exemple, són esciòfils durant els primers estadis de la seva vida, de manera que els plançons poden créixer a l’ombra dels arbres adults; en canvi, un cop han assolit una talla mitjana comencen a tolerar més bé nivells elevats d’insolació. Les clarianes assolellades de les fagedes de l’Europa atlàntica s’omplen aviat de plantes heliòfiles com els avellaners (Corylus avellana, C. americana), diferents roures (Quercus robur, Q. petraea) i alguns pins (Pinus sylvestris), però quan aquestes espècies ja han crescut, els plançons esciòfils dels faigs poden créixer a la seva ombra. A les boscanes decídues, la quantitat de llum que rep cada estrat varia segons les estacions. A l’hivern, per exemple, un 25% de la llum que incideix sobre els cimals d’una fageda arriba al sòl; quan hi ha fulla, en canvi, aquest percentatge es redueix al 2%.
La majoria dels arbres de les boscanes decídues no creixen gaire quan són joves però, progressivament, van creixent més en termes d’increment del seu pes sec. Els faigs, per exemple, presenten les taxes de creixement anual més altes entre els 35 i els 40 anys i, quan són més vells, aquesta decreix. Una fageda o una roureda de 120 anys té una biomassa d’unes 275 t/ha (en pes sec), mentre que a les selves tropicals aquest valor pot arribar a les 1 000 t/ha. El balanç de matèria orgànica d’un ecosistema, en aquest cas d’una boscana decídua, s’expressa habitualment en termes de quantitat de carboni que les plantes assimilen i respiren. El carboni, en efecte, és el component més important de la biomassa de la planta i s’assimila de manera força homogènia a tots els estrats de qualsevol boscana. Un tret característic d’aquestes, però, és l’elevat percentatge de biomassa que concentren els arbres i, concretament, les seves parts estructurals, sobretot en el cas de les fagedes, on l’estrat arbustiu és molt limitat i l’herbaci gairebé inexistent. En l’estrat arbori, les parts perennes (troncs, branques i arrels), és a dir, la fusta, representen més del 98% de la biomassa. A les rouredes, per exemple, els mateixos arbres constitueixen un nínxol ecològic particularment diversificat: centenars d’espècies d’invertebrats, sense comptar els animals de més envergadura, poden viure, sense anar més lluny, en un únic roure pènol (Quercus robur).
La necromassa, és a dir, la biomassa morta però encara no reciclada, és també molt important en les boscanes decídues, en particular la fusta morta, molt abundant, que té un paper essencial en l’ecologia del bosc a l’hora de reciclar nutrients. La taxa mitjana de mortalitat anual en les boscanes decídues del centre d’Europa és del 5,35% per als arbustos i de l’1% per als arbres. En algunes fagedes s’han observat densitats de fins a 36 arbres morts, però encara en peu, per hectàrea, la majoria d’edats compreses entre els 220 i els 270 anys. I no cal dir que, a més dels que romanen drets, n’hi ha molts de caiguts per terra en diferents estadis de descomposició. Hi influeixen molts factors, però una de les causes naturals de mort dels arbres més comunes, sobretot quan són vells, són els vents forts. En les boscanes també és molt corrent la caiguda contínua de branques i branquillons morts, que entapissen el terra, com també la fusta morta en arbres que encara són vius però han estat parcialment necrosats per l’acció de fongs paràsits.
La fusta morta es recicla gràcies a l’acció dels fongs lignícoles, que la utilitzen com a base alimentària de la qual obtenen energia i nutrients per a créixer i reproduir-se. La major part d’aquests fongs (un 85%) disposen d’un sistema enzimàtic capaç de descompondre tant la lignina com la cellulosa de la fusta. Hi produeixen l’anomenada ‘podridura blanca’: la fusta pren un color esblanqueït i un aspecte esponjós però manté la seva estructura fibrosa fins a estadis de descomposició molt avançats. El 15% restant disposa d’enzims que ataquen exclusivament la cel·lulosa i altres polisacàrids de la fusta produint l’anomenada ‘podridura bruna’: la fusta perd ràpidament resistència i s’esquerda i s’esmicola en un residu de fragments de forma sensiblement cúbica i de color bru que no són altra cosa que lignina lleugerament modificada.
La virosta és un altre component molt important de la matèria orgànica d’una boscana decídua. Les quantitats més grans es donen després de la caiguda de la fulla, i al final de l’estiu és quan n’hi ha menys, just abans que les fulles comencin a caure. A les fagedes la proporció entre els valors més alts i els valors més baixos és de 3 a 2. La matèria orgànica del sòl, però, no és sols la que aporten la fusta morta i la virosta sinó que inclou també la matèria orgànica viva que representen les arrels (la principal forma de matèria orgànica del subsòl) i els animals de la fauna edàfica. Per això els horitzons profunds del sòl contenen molta més matèria orgànica (prop de tres vegades més) que els horitzons húmics superficials. Això s’explica per l’activitat mineralitzadora que hi ha al sòl i pel gruix dels horitzons minerals. Les arrels es distribueixen amb força homogeneïtat i constitueixen una part important de la massa del sòl. Alguns autors creuen que representen entre 15 i 25 vegades la matèria acumulada en la virosta, i entre 1/3 i 2/3 de tota la matèria viva.
La diversitat dels consumidors

L’abundància i el nombre d’espècies de gastròpodes en els boscos centreuropeus estan determinats, entre altres factors, pel tipus de sòl de cada bosc en concret. Les fagedes —els boscos més característics de les boscanes decídues de l’Europa continental—, en aquest cas les que es desenvolupen sobre sòls calcaris, presenten una biodiversitat de gastròpodes més gran que els boscos mixts de roure i faig, que es desenvolupen sobre sòls menys exigents i menys calcaris, i que tenen un estrat arbori menys dens. Aquesta abundància es confirma tant pel que fa al nombre d’individus d’espècies grans com al d’espècies més petites.
Dades proporcionades per l’autor
Les relacions entre els organismes vius de qualsevol ecosistema, que solen pertànyer a una gran varietat de grups diferents, són determinades per la naturalesa de l’ambient on viuen. Les relacions que s’estableixen entre ells no són cadenes tròfiques simples ni lineals, sinó que més aviat constitueixen xarxes interconnectades extremament complexes. D’altra banda, l’acusada estacionalitat climàtica de les boscanes decídues fa que aquestes xarxes complexes no siguin constants al llarg de l’any; els cicles de la matèria i de l’energia tenen fases estacionals durant les quals la vida que hi ha sobre i a l’interior del sòl canvia de forma. En el cas dels animals, o consumidors, els processos de renovació segueixen uns patrons de temps diferents dels dels productors primaris, és a dir dels de les plantes. Els boscos caducifolis experimenten canvis cíclics que els permeten acollir un gran nombre d’espècies animals de tota mena.
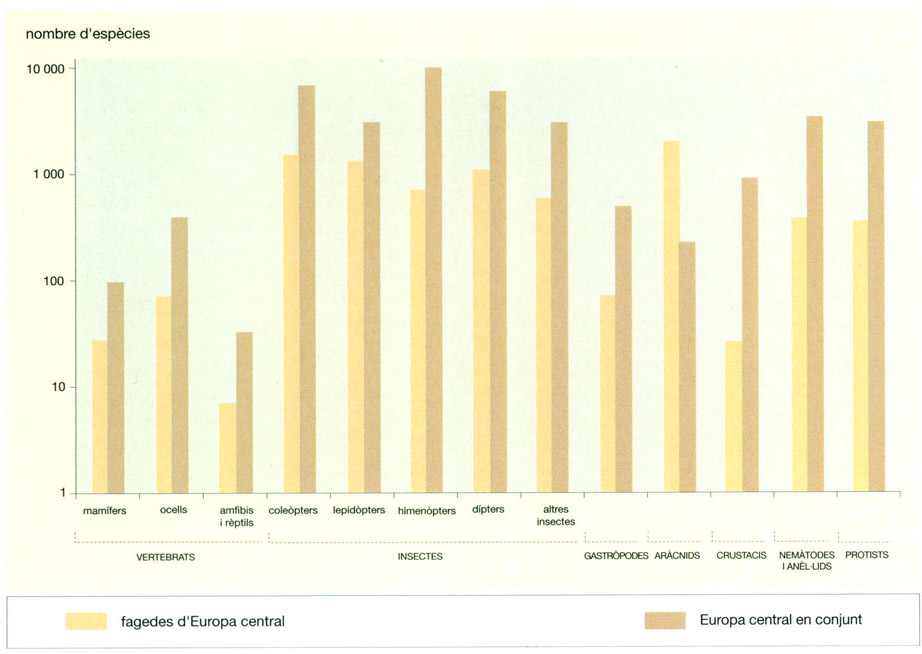
Biodiversitat zoològica en una fageda de l’Europa central (el nombre d’espècies està en escala logarítmica) comparada amb la del conjunt d’aquesta part del continent. Sobre un total d’unes 40 000 espècies dels grups aquí citats, a la fageda objecte d’estudi se’n troben unes 6 700, cosa que representa un 17%. Alguns grups zoològics es troben més ben representats al bosc de faigs que no pas d’altres. Així, per exemple, aproximadament el 43% dels lepidòpters de l’Europa central habiten la fageda (unes 1 300 espècies sobre un total de 3 000), mentre que els himenòpters hi tenen un 7% dels seus representants europeus (700 espècies sobre 10 000, en nombres rodons) i els crustacis només el 3% (unes 25 espècies de les aproximadament 900).
IDEM, a partir de dades proporcionades per l’autor
Els mamífers, tant si són herbívors com carnívors o omnívors, s’han adaptat a tots els biòtops de les boscanes. Els cérvols (Cervus elaphus, C. nippon) són grans consumidors de tota mena de matèria vegetal, però sobretot de plantes herbàcies, fulles verdes i brots d’arbres i arbustos. Els omnipresents ratolins (Apodemus, Peromyscus), pel seu gran nombre i perquè consumeixen grans quantitats de fruits i de llavors, també acompleixen una important funció dins les boscanes. A la fageda de Fontainebleau, prop de París, s’ha comprovat que la densitat del ratolí de camp ratllat (Apodemus agrarius) pot fluctuar entre 20 individus/ha fins a gairebé cap, fluctuació relacionada amb el fet que els faigs són anyívols i no fan fruit cada any.
Gràcies a la gran varietat d’aliments que poden explotar, les diferents espècies d’ocells ocupen, com els mamífers, una àmplia varietat de biòtops. A diferència d’aquests, però, la part més important de la seva dieta són els insectes, si bé no poden treure profit de la fauna edàfica tal com fan alguns mamífers durant l’hivern. La majoria dels ocells són territorials i defensen un espai entorn del seu niu, dins del qual disposen de prou aliment. Per això hi ha una relació clara entre les dimensions del territori, les seves necessitats tròfiques i la disponibilitat d’aliment en el medi.
En un estudi sobre la biomassa de les sis espècies de picot que viuen a les fagedes de Fontainebleau, es trobaren en conjunt 20 ocells/km2, amb una biomassa total de 4 330 g/km2. La biomassa individual més elevada correspon al picot negre (Dryocopus martius), amb un pes de 300 g i una presència de 0,4 individus/km2. La biomassa més gran per espècie corresponia al picot verd (Picus viridis) amb 1 830 g/km2, gràcies a la seva considerable biomassa individual (180 g), inferior només a la del picot negre, i al fet de ser l’espècie més abundant, amb una presència de 10 individus/km2. Un estudi dels hàbits alimentaris del picot negre en una fageda ha revelat que pot arribar a consumir fins a 150 000 exemplars de formigues (en el cas estudiat, del gènere Lasius) durant l’època de cria, amb una proporció d’ingesta de dues larves per cada tres insectes adults.
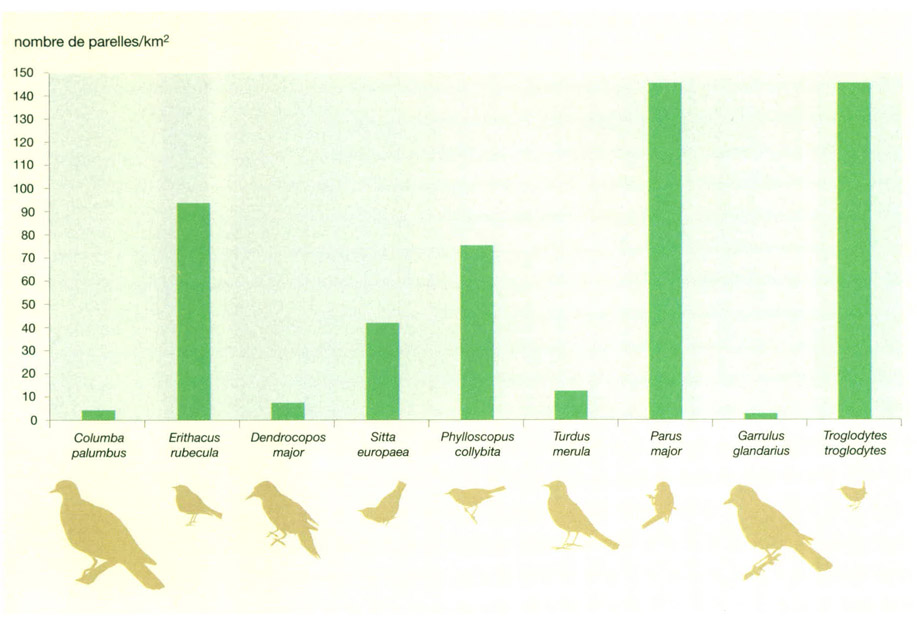
En una fageda es poden trobar una gran varietat d’ocells ocupant estrats molt diversos, des de les capçades dels arbres fins als matolls més baixos. El nombre de parelles/km2 de cada espècie, i per tant la seva biomassa total, depèn d’una sèrie de factors relacionats sobretot amb la vegetació. Algunes de les espècies més petites són també les més abundants; en són bons exemples el pit-roig (Erithacus rubecula) o el cargolet (Troglodytes troglodytes) dues espècies que es mouen en els estrats més baixos de vegetació. A més, molts d’aquests ocells menuts, incloent-hi la mallerenga carbonera (Parus major), són migradors parcials en les zones al N de les seves àrees de distribució. Altres ocells de mida reduïda, però de costums més arborícoles, com el pica-soques blau (Sitta europaea) o el mosquiter groc petit (Phylloscopus collybita), són relativament menys abundants que les espècies anteriors, si bé mostren una densitat notablement més alta que els ocells de més biomassa individual. Molts d’aquests últims, per exemple el gaig (Garrulus glandarius) o el picot garser gros (Dendrocopos major), prefereixen l’estrat arbori per a alimentar-se i niuar.
IDEM, a partir de dades proporcionades per l’autor
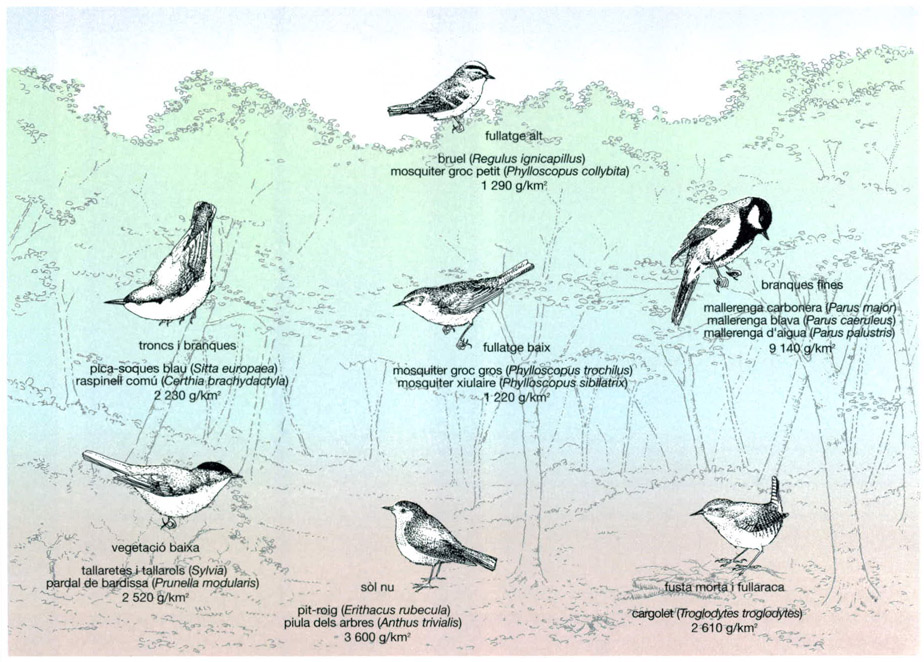
La comparació de la biomassa total d’adults de les diferents poblacions de passeriformes d’una fageda posa de manifest que les espècies que ocupen un nínxol alimentari en els estrats inferiors de la vegetació representen una biomassa més gran que les espècies que s’alimenten al fullatge, fet directament relacionat amb el nombre d’ocells de cada espècie present i amb la seva mida individual. Si hom es fixa en alguns dels ocells que ocupen les branques fines, per exemple, és remarcable que les tres espècies citades representin el 40% de la biomassa total de tots els estrats. Els ocells que s’alimenten a nivell del sòl, entre la fullaraca i la fusta morta o en l’estrat de vegetació més baixa representen percentatges d’entre un 11 i un 16% de la biomassa total dels passeriformes considerats. Les espècies que ocupen els dos estrats superiors, en canvi, presenten percentatges força inferiors, al voltant del 6%.
Jordi Corbera, a partir de dades proporcionades per l’autor
Els ocells més comuns, tant pel que fa al nombre d’espècies com al d’individus, són moixons que ocupen petits territoris. Hi ha estudis que han demostrat que en una fageda centreuropea es poden trobar, per hectàrea, un centenar de parelles de la vintena d’espècies que hi poden viure. Aquestes espècies ocupen nínxols diferents, i cada una d’elles posseeix una distribució de biomassa també diferent. L’anàlisi de l’abundància relativa de cada espècie revela que els factors que afecten més la distribució són la composició de la vegetació i el tipus de sòl, seguits, sobretot a l’estrat arbori, de la densitat de les plantes llenyoses.
Els lepidòpters, és a dir, les papallones tant diürnes com nocturnes, i en particular les seves larves, les erugues, són grans consumidores de matèria vegetal, sobretot les larves de les arnes. De les 1 300 espècies de falenes o papallones nocturnes que viuen a l’Europa central, 550 ho fan a les fagedes. Erugues de 75 espècies diferents s’alimenten de la fulla de faig (Fagus) i les de 223 espècies més s’alimenten de la del roure pènol (Quercus robur). Els lepidòpters han coevolucionat en estreta harmonia amb les plantes, fins al punt que la diversitat de lepidòpters en un ecosistema és determinada pel nombre i la varietat de plantes que hi viuen. Les arnes, i sobretot les seves larves i pupes, serveixen d’aliment a molts animals, sobretot ocells, formigues, aranyes i rates-pinyades. També hi ha altres insectes, com algunes vespes i mosques, que els parasiten. De fet, els lepidòpters ponen molts ous, però molt pocs individus arriben a la fase adulta. Una investigació referent a una població del noctúid Panolis flammea posà de manifest que, de les 127 pupes/m2 que hi havia d’aquest lepidòpter, només un 8% arribava a aparellar-se i, per tant, només un 4% arribava a pondre ous. Molts individus, ja des del mateix moment de la posta, moren per raons diverses, primordialment la depredació. Només el 0,05% dels ous sobreviuen i formen la crisàlide; en total, doncs, se’n perd el 99,95%.
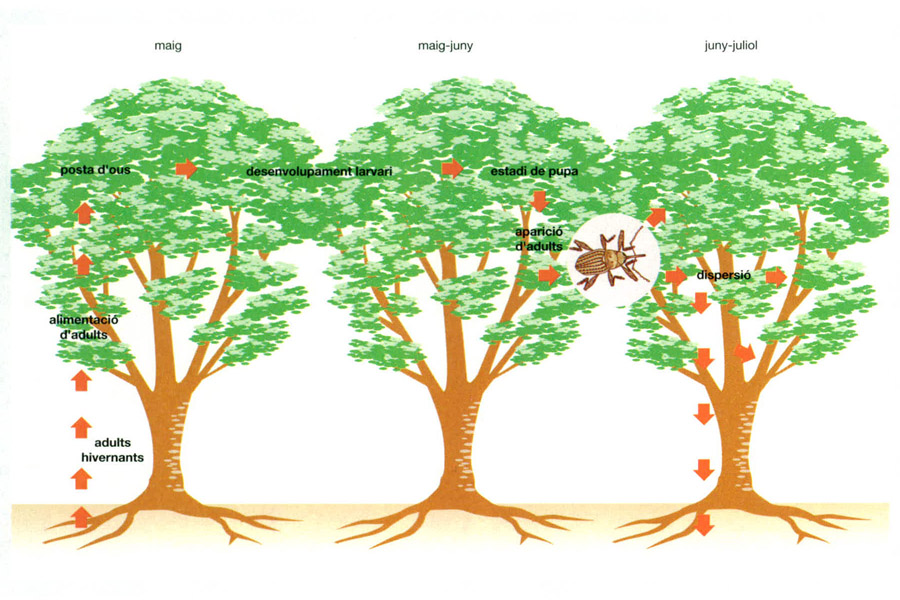
El cicle vital del curculiònid Rhynchaenus fagi transcorre tot sobre el faig comú (Fagus sylvatica). En fagedes de Dinamarca, al principi de maig els adults d’aquest insecte fil·lòfag que hibernaven al sòl emergeixen, s’encreuen i realitzen la posta. Cada femella deixa entre 30 i 35 ous vora el nervi central de les fulles del faig. Al cap de 16-19 dies, cap al final de maig i principi de juny, les larves entren a l’estadi de pupa, que s’allarga 17 o 18 dies, i a la segona meitat de juny comencen a aparèixer els adults, que durant aquest mes i el següent es dispersen i roseguen la part verda de les fulles, deixant tan sols l’esquelet dels nervis. Les densitats més elevades d’aquest curculiònid s’han observat després de la posta, al final de juny.
IDEM, a partir de C. Ascaso Terren, a Terradas, 1984
Els coleòpters o escarbats són arreu el grup animal més nombrós, i també ho són a les boscanes decídues. De les 6 800 espècies que es coneixen a l’Europa central, 1 500 es troben a les fagedes, i d’aquestes, 200 estan específicament lligades a aquesta mena de bosc. En estadi de larva, els coleòpters solen ser consumidors insaciables de matèria vegetal, però també n’hi ha que tenen larves carnívores. Alguns tenen cicles vitals molt llargs, amb fases larvàries d’uns quants anys de durada, fet que respon sobretot al baix contingut energètic d’alguns dels seus principals aliments com, per exemple, la fusta. Altres, com les marietes, acompleixen una funció important dins l’ecosistema alimentant-se dels pugons, insectes que viuen de la saba de les plantes.
Els himenòpters (abelles, vespes i formigues principalment) són importants tant pels hàbits alimentaris d’algunes de les seves larves com per la depredació a què estan sotmeses certes espècies. Molts són paràsits o fins i tot hiperparàsits, és a dir, organismes que parasiten altres paràsits. També n’hi ha molts que tenen importància com a pollinitzadors. Les formigues practiquen una explotació intensiva dels recursos tròfics disponibles al voltant dels formiguers, que de vegades són enormes. La formiga roja (Formica rufa) es mou normalment dins d’un radi de 40 o 50 m al voltant del niu, 100 m com a màxim. En un formiguer hi poden viure entre 500 000 i 800 000 individus, i fins a 60 reines, que tenen una esperança de vida de 15 anys. En un any s’hi poden moure de 250 a 500 kg de melassa de pugó, de la qual s’alimenten aquestes formigues abans d’acabar menjant-se el pugó, encara que llimacs, cucs i altres insectes, entre ells moltes larves de papallones nocturnes, són també part important de la seva nutrició. Els himenòpters són un dels grups animals més importants als boscos, perquè són pol·linitzadors importants i regulen les poblacions d’altres animals (per exemple arnes que poden constituir flagells per a determinades espècies vegetals) dels quals són paràsits o depredadors. Altres grups importants d’insectes consumidors a les boscanes són els dípters (mosques i mosquits) i els afidoïdeus (els pugons). S’ha calculat que, en teoria, un afidoïdeu pot tenir 1030 descendents en un sol estiu.
Les aranyes són un altre dels grups de més significació ecològica a les boscanes decídues, ja que són predadors que s’alimenten sobretot d’insectes vius. Una aranya pot arribar a consumir uns 2 000 insectes al llarg de la seva vida, que pot tenir una durada d’un any o dos. Atès que hi pot haver unes 150 aranyes/m2, poden arribar a eliminar 300 000 insectes en dos anys i controlar així amb eficàcia alguns flagells. Amb el verí que posseeixen aconsegueixen dissuadir molts depredadors potencials. És un grup molt divers que ocupa tots els nínxols ecològics; fins i tot n’hi ha d’aigua. En alguna fageda concreta, s’han arribat a trobar prop d’un centenar d’espècies d’aranyes grosses i unes 500 de petites, pertanyents a una trentena de famílies diferents.
La fauna del sòl i els descomponedors
La descomposició, la mineralització i el reciclatge de tota la matèria orgànica morta que generen productors i consumidors de la boscana es desenvolupen primordialment sota terra. Aquests processos tenen lloc sobretot al final de la primavera i a l’estiu, i és per això que al començament de la tardor hi ha molt poca matèria per descompondre al sòl del bosc.
A la fauna del sòl hi ha dos grans grups d’organismes que transformen la matèria orgànica morta. Un grup, compost principalment per artròpodes i cucs, tritura i digereix la fullaraca, la fusta i els animals morts. L’altre grup, compost per fongs i bacteris, es dedica després a descompondre pròpiament els productes de l’activitat dels organismes del primer grup. La funció dels primers és essencialment ingerir la virosta, esmicolar-la i trencar les parets cel·lulars de les plantes. Un cop les fulles han quedat reduïdes a partícules fecals, constituents essencials de l’humus, els bacteris completen el procés de mineralització. Així mateix, una part de l’aliment de la fauna del sòl procedeix de la producció secundària (animals morts i defecacions) i dels mateixos fongs i bacteris responsables de la descomposició pròpiament dita. Els productes d’aquest últim pas de la cadena, que consisteixen en restes animals i vegetals ja mineralitzades o reduïdes a compostos senzills, poden ser metabolitzats novament per les plantes.
Fauna edàfica

Jordi Corbera, a partir de Walter & Breckle, 1989
La categorització de la fauna del sòl en funció de la grandària es mostra de forma reduïda en l’esquema, on s’inclouen exemples dels grups més importants. La franja blava corresponent a cada grup indica la mida a què pertanyen els seus representants en relació amb els valors de l’escala en mil·límetres que hi ha a la part inferior de la figura. La xifra entre parèntesis al costat de cada grup taxonòmic és una estimació mitjana d’individus en un volum de sòl d’una àrea superficial d’1 m2 i profunditat 30 cm. La varietat de grups presents és enorme i si s’analitzés la manera com s’interrelacionen entre ells dins d’aquest subsistema es veuria que la xarxa és força complexa i que hi ha una relació directa entre les dimensions de les diferents espècies pertanyents a un grup i les diverses funcions que els corresponen en la degradació de la materia orgànica. Els grups pertanyents a la microfauna, bacteris, fongs i protozous, són sempre els més nombrosos. Entre la resta dels grups n’hi ha de paràsits, d’hiperparàsits, de sapròfags, etc., una gran diversitat funcional, com ja s’ha dit, que contribueix a modificar i caracteritzar les propietats del sòl i l’existència de la qual és possible gràcies a la riquesa estructural del sòl, a la presència d’aigua i a l’entrada total d’energia.
Les primeres transformacions de la matèria orgànica del sòl de la boscana produeixen l’humus, el complex de substàncies orgàniques parcialment mineralitzades (sobretot lignina de la fusta i excrements de cucs de terra) que representa una fase intermèdia en la descomposició de la matèria orgànica morta del sòl. A les boscanes decídues, l’humus s’acumula sota la virosta i constitueix l’horitzó húmic del sòl. La fauna del sòl esmicola i mineralitza tant la virosta com l’humus. Aquesta activitat, en la qual es consumeix oxigen i es produeix diòxid de carboni perquè hi ha implicats fenòmens de respiració i fermentació, decreix a l’hivern, però no s’atura, perquè sota la virosta i els detrits la temperatura del sòl varia menys que a la superfície externa i la humitat es manté constant al 100%. L’equilibri de l’atmosfera del sòl es restableix per difusió gràcies a la porositat del sòl, però, tot i això, els nivells de diòxid de carboni i d’oxigen solen ser respectivament més alt i més baix que a la superfície. La ‘respiració del sòl’, mesurada en termes de quantitat de diòxid de carboni difós per la superfície del sòl per unitat de temps, determina el grau d’activitat de les arrels i dels organismes. També permet estimar quanta matèria orgànica es mineralitza en un any, una xifra aproximadament equivalent a la producció primària de l’ecosistema.

Les larves dels dípters ocupen gran diversitat de nínxols ecològics, com mostra aquesta representació d’una soca de faig (Fagus) d’entre 6 i 8 anys. De fet, només a l’Europa central existeixen 340 espècies de dípters adaptades exclusivament a la vida a les fagedes, i les larves de moltes d’elles constitueixen una part important de la fauna edàfica. Bé que la biologia i l’ecologia d’aquestes larves no s’han estudiat amb detall, són tan abundants que la seva activitat alimentària és d’importància cabdal per a la descomposició de la matèria orgànica morta. Pertanyen a famílies molt diverses (els erínnids actualment pertanyen a la família dels xilofàgids) i ocupen gairebé qualsevol hàbitat edàfic, tant a la superfície del sòl com en el seu interior. La seva alimentació està també molt diversificada; consumeixen des de cadàvers de petits animals morts i excrements fins a restes vegetals i fusta. Al seu torn, constitueixen una part important de la dieta de molts altres animals.
Jordi Corbera, a partir de Walter & Breckle, 1989
La fauna del sòl posseeix una sèrie de característiques derivades de la necessitat d’adaptar-se a les condicions especials d’aquest ambient. En total, 1 litre de sòl d’una fageda d’Europa central sol contenir uns 22 g de pes fresc de biomassa animal, encara que en condicions ideals aquesta xifra pot arribar a ser deu vegades superior. Es tracta d’animals que no són sensibles a la llum, però en canvi responen a estímuls químics i mecànics. Són molt petits, sovint vermiformes, i poden ser excavadors o minadors segons sigui la porositat del sòl. També estan ben adaptats a condicions parcialment anaeròbies. La seva distribució en els diferents horitzons mostra una clara estratificació, presentant una densitat més gran en la virosta i en els horitzons superiors.
Entre els cucs de la fauna edàfica, els nematodes són particularment nombrosos; en 1 litre de sòl d’una boscana decídua se’n poden arribar a trobar 30 000. Són un dels grups de diversitat més alta que hi ha al bioma. Formen part de diferents xarxes tròfiques, ja que es poden alimentar de la saba de les plantes, de fongs, de protoctists —tant productors (algues) com consumidors (protozous)— i de bacteris; alguns també poden parasitar arrels. Els anèl·lids, entre els quals hi ha els llambrics o cucs de terra (lumbrícids), viuen entre la virosta i a l’interior del sòl, però a les boscanes decídues la seva activitat no és gaire elevada, ni tampoc la seva presència, que amb prou feines arriba a representar, en pes fresc, uns 20g/m2.
Entre els microartròpodes del sòl hi ha insectes (col·lèmbols, per exemple) i molts organismes d’altres grups. Els àcars, sobretot els oribàtids, figuren entre els microartròpodes més abundants i són molt actius en el sòl. A les fagedes centreuropees se n’han arribat a comptar fins a 16 500 individus/m2 de mitjana anual; són més nombrosos a l’hivern que a l’estiu i contribueixen a la formació d’humus consumint virosta, fusta i fongs. Els collèmbols també són molt nombrosos, amb mitjanes anuals de 17 500 individus/m2, i se solen trobar sobre les restes de plantes i altres matèries en descomposició. Als oribàtids i col·lèmbols pertanyen les espècies dominants de la fauna de la virosta, que és on es troben els dos terços dels organismes del sòl.
Els macroartròpodes més abundants del sòl són els porquets de sant Antoni (isòpodes), els milpeus (diplòpodes), les escolopendres (quilòpodes) i els opilions. Els dos primers grups s’alimenten sobretot de matèria vegetal en descomposició, mentre que els altres dos s’alimenten d’altres animals del sòl, com ara col·lèmbols i àcars, i no tenen tanta importància com a descomponedors. Tret dels quilòpodes, tots aquests grups tenen cicles d’activitat ben definits: els isòpodes són comuns a partir de l’abril, amb un màxim al juny i al juliol; els milpeus són més actius de febrer-març fins a juliol-agost, amb un pic entre març i juny; els opilions assoleixen els seus màxims de població entre juliol i desembre. En qualsevol cas, tots són abundants a tot arreu on hi hagi matèria orgànica en descomposició, i la seva distribució, sobretot la d’isòpodes i diplòpodes, que plegats representen unes tres quartes parts de la fauna de macroartròpodes de qualsevol sòl de boscana decídua, depèn en bona part de l’existència d’humitat i del tipus de sòl.
Un cop la macrofauna del sòl ha acabat la seva feina entren en escena els descomponedors pròpiament dits, que són els fongs i els bacteris. Entre ells són especialment importants els bacteris dels nòduls de les arrels de les plantes lleguminoses i certs fongs simbiòtics, ja que fixen el nitrogen atmosfèric i el posen a disposició de les arrels de la planta. Aquesta darrera etapa de la descomposició és la mineralització, un procés complex, amb diferents fases, algunes dutes a terme per bacteris molt especialitzats, que dóna com a productes finals molècules minerals senzilles (aigua, diòxid de carboni, amoníac, nitrats, fosfats, sulfats) i elements nutrients. Normalment comencen a actuar al mateix temps diferents descomponedors; primer de tot els fongs fan penetrar les hifes entre la virosta o al si de la fusta morta mentre els animals de la fauna edàfica en processen una part al seu sistema digestiu. Més tard els bacteris ataquen la virosta processada pels animals i convertida en dejeccions. La virosta de verns, freixes i oms triga un any a descompondre’s del tot; la d’arços i cirerers en triga un i mig; la de tells i aurons, dos anys; la de roures, bedolls i pollancres, dos i mig, i la de faigs, tres anys. Encara que la biota bacteriana del sòl de les boscanes decídues no és ben coneguda, sí que se sap que, així com els fongs es desenvolupen millor en sòls àcids, els bacteris ho fan més bé en sòls bàsics. Tant les poblacions de fongs com les de bacteris són controlades per protoctists consumidors (protozous) i per la microfauna del sòl.
El ritme de les estacions
La gran diversitat de la vida a les boscanes decídues no es manifesta únicament en les pautes espacials sinó també en les temporals. A les boscanes decídues, en efecte, l’estacionalitat és un factor que condiciona profundament totes les formes de vida. L’activitat dels éssers vius i les seves relacions entre ells i amb els diferents factors ambientals són determinades per l’existència d’estacions ben diferenciades, cada una amb unes característiques climàtiques ben definides.
La primavera freturosa
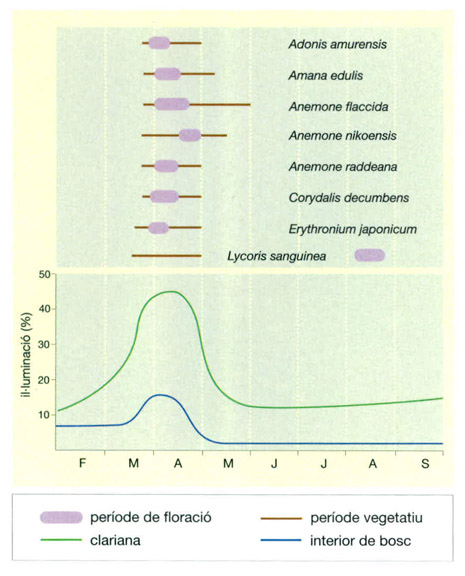
Relació entre l’aparició de les plantes efímeres (a dalt) i la intensitat lumínica (a baix) a nivell del sòl en un bosc caducifoli. A partir del solstici d’hivern l’allargament del dia i l’elevació creixent del Sol sobre l’horitzó fan créixer la intensitat lumínica a nivell de terra. Al llarg de la primavera, però, l’aparició de les fulles compensa amb escreix aquest increment i la intensitat lumínica davalla fins a un mínim el mes de maig, en el qual es manté fins a la caiguda tardoral de les fulles. A partir del mes de març la intensitat lumínica és suficient perquè les plantes efímeres treguin fulles i aprofitin ràpidament per a florir. Gairebé totes treuen flor al cap de poc d’haver tret fulles, però algunes ho fan més tard, com per exemple Anemone nikoensis, i diverses espècies bulboses, com la liliàcia Lycoris sanguinea, que trauran flor a partir del bulb al cap d’uns quants mesos.
IDEM, a partir de M. Numata & M. Ohno, 1985
L’arribada de la primavera assenyala la fi de la letargia hivernal. La neu quasi ha desaparegut, el dia es comença a allargar i la temperatura comença a pujar significativament. Tanmateix, la primavera és una estació irregular en què les onades de fred alternen amb períodes càlids i assolellats. Encara hi pot haver algunes glaçades ocasionals, que són les més perilloses per a arbres i arbustos, perquè els poden afectar quan tot just comencen a brotar. Així i tot, un cop el sòl s’ha alliberat de la capa de neu es comença a escalfar, i sota la virosta l’activitat augmenta notablement. Així que la temperatura supera els 5°C durant unes quantes hores al dia, l’activitat que s’havia aturat a l’hivern es reprèn i el bosc torna a l’efervescència.
Els arbres i els arbustos viuen un procés de rejoveniment; hi torna a haver activitat des de les arrels fins als borrons, on les noves fulles i les noves flors es desensonyen a punt per a la desclosa. La saba comença a circular pels vasos de troncs i branques i transporta aigua carregada amb sals minerals fins a les extremitats, on les fulles i les flors es comencen a desenvolupar. A la majoria d’arbres i arbustos de les boscanes decídues la pol·linització és anemòfila, és a dir, que el pol·len ha de ser transportat pel vent de les flors masculines a les femenines perquè es pugui formar el fruit amb les llavors. És important, doncs, que el vent no trobi obstacles al seu pas (com ara les fulles d’arbres i arbustos) a l’hora de transportar el pol·len en suspensió a través del bosc. Per això, de fet, moltes espècies floreixen abans de treure la fulla. A Europa, per exemple, els avellaners (Corylus) són els primers anemòfils que despengen els seus llargs aments de flors masculines i que ofereixen el seu pol·len a les característiques rauxes de vent, típiques de les primeres setmanes primaverals.
Els roures (Quercus), les nogueres (Juglans) i moltes altres espècies no triguen gaire a imitar-los, tot i el risc que una glaçada tardana durant la floració pugui inhibir la formació del fruit. De vegades, les flors es poden desenvolupar abans que les fulles en funció de la llargada del dia; hi ha espècies que responen a l’allargament del dia, n’hi ha que reaccionen a l’escurçament i n’hi ha que hi són indiferents. Als faigs (Fagus), per exemple, la floració i la foliació poden tenir lloc al mateix temps, i les petites dimensions de les fulles joves no impedeixen que el vent transporti el pol·len lliurement. El sòl, d’altra banda, es comença a omplir de flors, sovint molt vistoses, que també es desclouen amb l’arribada de la primavera. Durant l’hivern aquestes plantes sobreviuen en forma de bulbs, rizomes o altres òrgans modificats en teixits de reserva i responen amb immediatesa a l’escalfament del sòl degut a la tebior dels primers raigs de sol de la primavera. Les prímules (Primula), la fetgera (Hepatica nobilis), el buixol (Anemone nemorosa) i altres congèneres, la gatassa (Ranunculus ficaria) i moltes altres treuen el cap per entre la virosta, i fins i tot per entre la neu, i encatifen el terra del bosc abans que la fulla dels arbres faci disminuir la disponibilitat de llum.
La formació de la fulla exigeix que la maquinària viva de l’arbre hagi assolit el seu ple funcionament i el sotmet a un gran esforç. Però, a més, un cop les fulles s’han desenvolupat i són funcionals, l’arbre els ha de proporcionar l’aigua, les sals minerals, el diòxid de carboni i l’energia solar que necessiten per a fotosintetitzar les substàncies que són essencials a la planta per a viure, créixer, reproduir-se i que seran acumulades als teixits. La transpiració, que és el mecanisme de pèrdua d’aigua a través de les fulles en forma de vapor, bombeja una gran quantitat de saba des de les arrels cap a la resta de la planta.
Les llavors també abandonen l’estat de latència que en retarda la germinació. Els embolcalls impermeables que les protegeixen s’esqueixen, l’aigua s’emporta les substàncies inhibidores que hi havia a l’ametlla i els efectes de la llum i la temperatura (fred o calor) estimulen el desenvolupament de l’embrió. Quan la llavor està fisiològicament a punt, és a dir, quan les condicions causants de l’estat de latència han desaparegut, una presència suficient d’aigua, escalfor i oxigen estimula el metabolisme latent de la llavor i l’embrió es comença a desenvolupar. Les llavors grosses, com aglans, castanyes i nous, posseeixen grans endospermes que proporcionen aliment a la plàntula mentre el seu potencial fotosintètic es desenvolupa.
Els animals responen també de manera immediata a les noves condicions ambientals. Els ocells que havien emigrat a la tardor tornen, i el bosc s’omple dels cants territorials dels mascles, que anuncien la seva presència i reclamen com a seus els recursos alimentaris de l’àrea que consideren necessària per a dur a terme amb èxit la cria dels pollets. Les poblacions d’invertebrats experimenten de cop veritables explosions demogràfiques quan els ous que han superat l’hivern eclosionen i quan emergeixen els insectes i altres artròpodes adults que han hibernat com a pupes o nimfes. L’escalfor fa reviure els habitants del sòl i milers d’espècies comencen a engendrar compulsivament milions de diminuts descendents, la gran majoria dels quals seran consumits per animals més grossos. Però també n’hi haurà uns quants que sobreviuran i completaran el seu cicle vital per tal d’assegurar la supervivència de l’espècie.
Els mamífers hibernants s’acaben despertant, sovint desnodrits i necessitats de grans quantitats d’aliment, i s’afegeixen a l’enrenou general del bosc. Els rosegadors i altres petits mamífers amb taxes metabòliques molt elevades necessiten alimentar-se gairebé constantment, i encara més al principi de l’època de cria. Com que a la primavera els arbres i els arbustos no tenen fruit, els lirons (glírids), els ratolins (múrids) i els talpons (micròtids) s’han d’alimentar principalment de petits animals, com per exemple els insectes, que ja són abundants. Els grans mamífers ja es troben també en plena activitat, i alguns, com l’ós bru (Ursus arctos), es desperten afamats i inquiets. Aquest és especialment el cas de la femella, que durant l’hivern ha parit una, dues o tres cries dintre el cau hivernal i que, amb l’alletament, ha esgotat les seves reserves de greix de la tardor i ha quedat desnodrida.
La calor ambiental creixent desperta els amfibis i els rèptils i a mesura que el cos se’ls escalfa esdevenen actius una altra vegada. Els gripaus fan petits moviments migratoris cap a les basses on van néixer per iniciar el cicle de cria i a la nit obsequien el bosc amb els seus raucs rítmics. El sòl i la virosta cobren vida pròpia; els processos de descomposició es reprenen per tal de retornar al sòl els nutrients i l’energia que els arbres acabaran convertint novament en els recursos alimentaris que donen vida al bosc.
L’estiu generós
Després de l’esclat vital de la primavera, a l’estiu l’activitat animal i vegetal s’estabilitza, però sense decréixer. L’objectiu fonamental per a cada espècie és assegurar que la producció garanteixi la supervivència d’un nombre suficient de descendents. A tots els estrats de les boscanes la vida es manifesta en tot el seu esplendor i revela el seu perfecte ajustament a una màxima eficiència en l’aprofitament dels recursos. Les innombrables interaccions entre els milers d’espècies existents es tradueixen en un nombre gairebé il·limitat d’estratègies de supervivència.
L’estiu és l’època d’un veritable esclat reproductor; cal generar moltíssims descendents perquè algun pugui arribar a sobreviure. Els nous tanys broten generosament, tant en arbres com en arbustos; la producció de fruits és molt elevada, i moltes plantes joves es marceixen a l’ombra dels grans arbres o són consumides pels animals. És un temps de sobreabundància de producció, no sols genèsica, per assegurar la supervivència de l’espècie, sinó també de reserves adreçades a garantir la supervivència individual. La saba arriba en abundància al fullatge dels arbres, que és el centre de producció de totes les substàncies que l’arbre necessita per a créixer i viure. En arbres i arbustos la producció s’aprecia principalment en fulles i fruits, encara que moltes de les espècies de les boscanes decídues, com ara els roures i els faigs, són anyívoles, és a dir, que no fan fruit cada any sinó sols un cop cada dos o tres.
Per als arbres, sobretot a l’estiu, l’aigua és un factor limitador important, no tant pel fet que no plogui (com passa a les mediterrànies) com per l’alt índex d’evapo-transpiració. Les diferents espècies de faig que hi ha als boscos caducifolis d’arreu del món, per exemple, són molt sensibles a la sequera i necessiten volums d’aigua relativament grans. El faig comú (Fagus sylvatica) vol humitat ambiental i un mínim de 200 mm de pluja a l’estiu. Altres espècies, com els roures (Quercus), els freixes (Fraxinus) i els oms (Ulmus), necessiten que hi hagi aigua al sòl i encara més a l’estiu. La calor, malgrat que la pèrdua d’aigua també serveix com a mecanisme termoregulador, ocasiona el tancament dels estomes de les fulles per tal d’evitar una transpiració excessiva.
De la capçada del roure més alt fins al subsòl on arrela, la vida bull de manera incessant al llarg de tot l’estiu. Les erugues minadores de diferents microlepidòpters nocturns s’alimenten del parènquima de les fulles; els escanyapolls (Lucanus cervus) i altres lucànids xuclen la saba que vessa de les ferides dels troncs dels arbres, mentre les seves larves s’alimenten de la fusta de l’arbre mateix. Milions de pugons (afidoïdeus) s’alimenten també de la saba que xuclen de les branques tendres i són devorats al seu torn per les marietes (coccinèlids). Les aranyes, les vespes i els ocells mengen grans quantitats d’insectes. Els pica-soques (Sitta) i els raspinells (Certhia) corren amunt i avall dels troncs dels arbres grossos buscant insectes a l’escorça, mentre mallerengues (Parus), mosquiters (Phylloscopus) i tallarols (Sylvia) cerquen les seves preses per les fulles i el brancatge. Aquests ocells són encalçats al seu torn per mustèlids i per rapinyaires boscans com ara l’esparver vulgar (Accipiter nisus), i fins i tot es poden veure atacats en els seus nius per les serps. Els esquirols arborícoles (Sciurus i gèneres afins) tenen preferència per les capçades dels arbres, però poden baixar a terra cercant aliment o per canviar d’arbre, a desgrat del risc de ser atacats pels seus enemics naturals, martes i fagines (Martes).
I al capdavall, tot aquest joc impressionant de menjar i defugir ser menjat té per objectiu principal obtenir el màxim èxit possible en la reproducció de la pròpia espècie. Els arbres i els arbustos produeixen milers i fins i tot milions de fruits i de llavors, dels quals potser cap no arribarà a sobreviure, ja que, en un bosc, una planta només deixa espai per a una altra de semblant quan es mor, però no abans i, en tot cas, mai per a més d’una. Tant els insectes com altres invertebrats engendren també milers de descendents cada any. D’aquests, molt pocs arriben a la maduresa, però ajuden amb els seus nutrients i la seva energia a la supervivència de moltes altres espècies predadores, generalment més grans. La vida al sòl també desplega una gran activitat, tant per part dels petits descomponedors com dels talps i les musaranyes que els encalcen, o fins i tot dels artròpodes predadors com ara les aranyes, molt abundants als sòls forestals.
Les primeres tempestes de final d’estiu i un lleuger descens de la temperatura són l’anunci dels canvis espectaculars que afectaran les boscanes a la tardor, durant la qual els preparatius del llarg parèntesi de repòs hivernal substitueixen gradualment el brogit de l’estiu.
La tardor espectacular
A les boscanes decídues la tardor es caracteritza principalment per la caiguda de la fulla dels arbres, que al final queden del tot despullats, i pel canvi dràstic que això provoca en la vida forestal. A més, mentre té lloc aquest procés, la boscana es revesteix d’una gamma impressionant de colors que assoleix el seu màxim esplendor en alguns punts de l’E de l’Amèrica del Nord i d’Àsia i és protagonitzat principalment pels aurons, blades i erables (Acer), de variadíssimes tonalitats que van del vermell al groc, sense que els vermells dels faigs (Fagus) i els grocs i bruns dels roures, les nogueres i les càries (Quercus, Juglans, Carya) i els oms (Ulmus), per dir-ne només uns quants, desdiguin gens de l’espectacle.
El fenomen de la caiguda de la fulla encara no ha estat del tot aclarit per la ciència. Fins no fa gaire es pensava que es devia a l’acció de l’àcid abscíssic, que rebé aquest nom precisament perquè se’l tenia per responsable de l’abscissió, és a dir, el trencament de la base del pecíol que provoca la caiguda de la fulla. Actualment, però, es creu que és l’etilè qui actua com a principal hormona de la senescència de la fulla i en causa l’abscissió estimulant la síntesi de cel·lulosa a la base del pecíol. Entre la fulla i el seu punt d’inserció a la branca es forma així una capa de cèl·lules suberoses anomenada ‘zona d’abscissió’; aquestes cèl·lules van entorpint les connexions entre el pecíol i la branca, de manera que el transport de nutrients a la fulla es redueix i aquesta no pot sintetitzar eficaçment la clorofil·la. La fulla perd la seva coloració verda i adopta altres coloracions degudes a pigments més duradors i menys exigents en nutrients. Finalment, les connexions entre la fulla i la branca s’interrompen del tot, la fulla mor i és enduta pel vent o bé cau simplement per la força de la gravetat. Els processos fisiològics que provoquen la caiguda de la fulla són estimulats pels canvis en la durada del dia, fenomen que s’anomena fotoperíode. Amb experiments d’hivernacle s’ha demostrat que, si es manté el fotoperíode natural, les fulles cauen igualment, independentment de la temperatura i dels altres factors ambientals que puguin modificar-se a voluntat en l’hivernacle.
Des d’un punt de vista evolutiu, la caiguda tardoral de les fulles planes i relativament grans dels arbres i els arbustos caducifolis es pot interpretar com un compromís entre l’eficàcia deguda al fet de disposar d’una gran superfície per a captar la llum i efectuar la fotosíntesi durant l’estiu càlid i plujós, i la necessitat d’evitar una pèrdua excessiva d’aigua a través d’aquesta gran superfície durant l’hivern, fred i eixut. Amb la caiguda de la fulla, es redueix a la meitat el volum d’aigua als teixits de l’arbre, i el risc de glaçades disminueix en gran mesura. La transpiració a través de les branques i les gemmes és molt baixa, i pràcticament s’atura la circulació de líquids. Totes les activitats metabòliques, de fet, disminueixen enormement i la planta entra en una mena de letargia. La tardor, en efecte, amb els grans canvis que comporta en la fisiognomia del paisatge, és primordialment una premonició de l’hivern que s’acosta. Els dies s’escurcen, les temperatures baixen, arriben les primeres glaçades i nevades i tornen les ventades fortes i ratxoses. Són aquests els fenòmens que causen els grans canvis que es verifiquen en la vegetació al llarg d’aquesta estació, i aquests canvis són, al seu torn, adaptacions necessàries per a poder superar l’estació més desfavorable de l’any i aconseguir sobreviure fins que arribi la primavera.
L’abscissió provoca la caiguda de les fulles, però també la de fruits i llavors. Alguns fruits, principalment els carnosos, com ara les drupes de les rosàcies del gènere Prunus, o els poms de pomeres (Malus) i pereres (Pyrus) silvestres, ja han caigut abans. Els fruits van madurant al llarg de la tardor, i així els altres habitants del bosc disposen d’una quantitat molt important d’aliment. Herbívors i omnívors, com ara el porc senglar (Sus scrofa) —que s’alimenta d’aglans, fages, nous, pacanes i tot allò comestible que pugui trobar—, són els principals consumidors de la producció de fruits. Però no en són els únics, com demostra l’estudi tardoral dels excrements d’altres animals normalment considerats carnívors com ara la guineu (Vulpes vulpes), l’ós (Ursus), el teixó (Meles meles), tots els mustèlids i fins i tot el llop (Canis lupus). Tots són consumidors fervents dels fruits que produeixen els arbres i arbustos de les boscanes decídues i els aprecien d’allò més pel fet que se’n troben en abundància i amb facilitat.
També hi ha fruits que es mantenen a la branca durant molts mesos, de vegades fins a la primavera següent, com és el cas dels dels freixes (Fraxinus). Els fruits rics en vitamines de les moixeres (Sorbus) pengen fent raïms a l’hivern, però els ocells i els petits mamífers se’ls mengen aviat i contribueixen així a la seva dispersió, ja que les llavors que contenen, deslliurades de la coberta carnosa un cop han travessat el sistema digestiu de l’animal, són excretades en un altre indret allunyat de l’arbre progenitor; d’aquesta manera l’animal ajuda la planta a colonitzar noves àrees. Aquest és precisament el significat evolutiu d’un fruit carnós, saborós, nutritiu i acolorit: els consumidors potencials acompleixen la funció d’escampar-ne les llavors per la regió. Aquest fenomen s’anomena zoocòria (dispersió duta a terme per animals). Altres espècies d’arbres i arbustos han adoptat sistemes de dispersió diferents; els aurons (Acer) i els freixes (Fraxinus), per exemple, posseeixen llavors alades que són transportades pel vent (anemocòria) i no han d’esmerçar energia en la producció d’un fruit carnós; els verns (Alnus), que creixen als marges dels rius, disseminen les seves petites llavors per mitjà de l’aigua (hidrocòria).
La major part de la producció primària cau a terra i queda a disposició dels habitants del sòl, on es va descomponent lentament. La fauna es limita als estrats inferiors perquè la falta de fulla impedeix que hi prosperin espècies fitòfagues arborícoles. La majoria dels ocells insectívors han migrat i molts mamífers es preparen per a l’hivern construint caus i emmagatzemant menjar. El vent dispersa les aranyes joves que pengen dels seus llargs fils, i creixen bolets pertot arreu. Els fongs, que tot i ser-hi presents també a la primavera són eminentment tardorals, són un component essencial en la producció secundària del bosc; creixen abans de les primeres glaçades sempre que troben un cert grau d’humitat a l’aire i al sòl. Hi ha una gran varietat de fongs i de bolets (que no són sinó el cos fructífer de determinats fongs), que creixen al sòl de les boscanes o sobre la fusta morta, i constitueixen una font nutritiva gens menystenible per a molts dels seus habitants.
L’hivern silenciós
A les boscanes decídues l’hivern és l’estació més tranquil·la, l’època de la quietud i el silenci. El bosc descansa en molts sentits i de diferents maneres; la majoria d’ocells han migrat i la seva xerradissa sorollosa s’ha aturat fins a la primavera. Molts mamífers, amfibis i rèptils entren en una fase de letargia o hibernació i pràcticament sembla que hagin desaparegut del bosc. Els insectes i altres artròpodes moren, o bé passen l’hivern en estat letàrgic o en forma de larves o pupes immòbils. Fins i tot les llavors entren en una fase de letargia: tot i que podrien haver germinat en el moment de madurar i caure, retarden la germinació fins a la primavera per diferents mètodes.
Els arbres i els arbustos es troben inevitablement exposats al fred i a altres condicions adverses. Les plantes herbàcies, en canvi, passen l’hivern en forma de llavors, de bulbs o de rizomes i d’aquesta manera eviten els rigors de l’estació: les llavors sota la virosta, i bulbs i rizomes sota terra. Per a arbres i arbustos, l’estació freda comporta el risc addicional de la possible formació de glaç als teixits de les seves parts aèries, que pot causar la mort del vegetal, bé per deshidratació i danys físics causats per la formació de cristalls de glaç, bé per alteracions en l’estat col·loïdal del citoplasma cel·lular o en la permeabilitat de les membranes plasmàtiques a baixa temperatura.
Els mecanismes destinats a prevenir aquests danys constitueixen un procés anomenat enduriment, l’única manera de resistir amb eficàcia les glaçades. En aquest procés se succeeixen diferents fases: en primer lloc un pre-enduriment, que es dóna quan la temperatura encara és per sobre dels 0°C, i comporta un ràpid increment de la concentració de sucres en els sucs cel·lulars. L’enduriment pròpiament dit es produeix durant la primera setmana de glaçades, al final de la tardor, quan la temperatura se situa entre els -3 i els -5°C. Comporta canvis en l’estructura col·loïdal del citoplasma cel·lular (principalment reduint el contingut d’aigua) i en la permeabilitat de la membrana plasmàtica, cosa que assegura que no es produeixin danys cel·lulars per formació de cristalls de glaç i permet a la planta suportar temperatures de fins a -30°C. La fase d’enduriment final incrementa la resistència a la congelació i les parts aèries de la planta esdevenen pràcticament inactives. En condicions de laboratori, s’ha comprovat que es pot sotmetre una planta a congelació amb nitrogen líquid a -196°C sense que es formin cristalls de glaç i es pot invertir el procés de vitrificació o congelació absoluta sense dany si es descongela la planta de manera adequada, pas a pas. Les arrels, que mai no es veuen exposades a l’acció del fred com les parts aèries de la planta ni han de resistir inclemències tan extremes, sempre es mantenen poc o molt actives i constitueixen una reserva important de saba i de substàncies nutritives que es podran utilitzar durant la següent estació de creixement fins que les fulles puguin reprendre la seva activitat sintetitzadora. Les espècies acomoden els seus processos d’enduriment a la temperatura de les regions on viuen. El roure pènol (Quercus robur) i el plàtan fals (Acer pseudoplatanus), per exemple, són molt més resistents a la congelació que altres espècies dels gèneres respectius pròpies de zones més occidentals d’Europa, de clima menys continental.
L’absència de vida a les boscanes decídues a l’hivern només és aparent. N’hi ha prou que caigui una fina capa de neu i es mantingui unes hores o uns dies per descobrir el traç de molts dels secrets hivernals del bosc. La vida i la mort no es deturen; els rastres damunt la neu tant poden revelar la persecució d’un animal herbívor per part d’un de carnívor com les cerimònies nupcials i d’aparellament de molts dels habitants del bosc. Molts mamífers s’aparellen pel gener, i per tant en aquesta època són més actius i es desplacen molt més del que és habitual.
Més avall, sota la neu, l’activitat al sòl s’ha reduït però no s’ha aturat del tot. Les fulles caigudes van desapareixent, consumides i digerides per cucs, bacteris, fongs i una miríade d’artròpodes, que al seu torn serveixen d’aliment a musaranyes (Sorex, Crocidura, Blarina) i talps (Talpa, Mogera, Parascalops, Condylura). Moltes de les llavors que han quedat mig colgades a la virosta són desenterrades, una darrere l’altra, pels ocells que es passen l’hivern al bosc i per petits mamífers com ara els talpons (Microtus, Eothenomys). Hi ha ocells que emmagatzemen determinats tipus de llavors, com és el cas del gaig europeu (Garrulus glandarius), que té el costum d’enterrar aglans per menjar-se’ls a l’hivern i contribueix així, sense voler però ben activament, a la regeneració de les rouredes.
Els ocells i els mamífers de bosc que es mantenen actius durant l’hivern pateixen taxes de mortalitat elevades, sobretot en hiverns rigorosos durant els quals molts moren de fred o de gana. Per a aquests animals algunes espècies d’arbres i arbustos de les boscanes decídues que no perden la fulla a l’hivern, com ara els grèvols (Ilex) i el teix europeu (Taxus baccata), són particularment importants; els proporcionen refugi, sobretot als ocells i, en el cas dels grèvols, les seves fulles i llavors són una font d’aliment essencial.
Les pertorbacions i els processos de successió
A primera vista sembla que les boscanes decídues siguin sistemes estables, immutables, en els quals les condicions arrecerades pròpies de l’interior del bosc mitiguen els extrems de la calor, el fred, la sequera, la inundació, el foc i la tempesta. No obstant això, la seva estructura i la seva composició canvien constantment a petita escala, i, ocasionalment, estan sotmeses a pertorbacions ràpides i catastròfiques, com ara els efectes dels focs i les tempestes. Quan això passa, es pot observar com evoluciona la boscana en la seva recuperació.
Els agents de pertorbació
Entre els agents de pertorbació més importants que poden afectar les boscanes decídues temperades figuren el vent, la sequera, els atacs de paràsits, els incendis, la neu i el glaç i la possible inestabilitat de l’emplaçament.
Els vents violents poden desarrelar els arbres o trencar-ne els troncs, deixant-los reduïts al monyó d’una soca. Els arbres més grans són més vulnerables que els més petits, de manera que, per a una mateixa espècie, els arbres més joves resulten relativament menys afectats que els més vells. En boscos madurs, amb arbres de totes les edats, els arbres més petits i els plançons poden sobreviure enmig de les restes abatudes dels arbres més grans.
La sequera és inusual al bioma de les boscanes decídues. Per això una sequera estival prolongada unida a la calor pròpia de l’estació pot causar la mort d’exemplars adults de faigs, bedolls i altres espècies arbòries, deixant dempeus els arbres morts, que es podreixen i es desintegren a poc a poc. Habitualment, els efectes immediats d’una sequera no són tan dràstics sinó que no passen de ser subletals, però deixen els arbres afectats més exposats i vulnerables als atacs de fongs i altres paràsits.
Les agressions biòtiques per part de paràsits o de consumidors són una altra causa de pertorbació; els arbres adults poden morir a causa d’una infecció fúngica. Les erugues defoliants redueixen el creixement i poden debilitar de manera fatal arbres que ja estiguin estressats per altres factors. Les fluctuacions en les poblacions de cérvols i d’altres herbívors poden perjudicar la regeneració en moments de màxima superpoblació. Els sanguinyols (Cornus) i altres arbustos pateixen els atacs de virus mentre oms i castanyers europeus i americans han estat delmats a causa d’infeccions fúngiques d’origen exòtic introduïdes pels escarabats de l’escorça, com per exemple la malaltia holandesa de l’om, que ha destruït oms a la major part d’Europa, o el xancre del castanyer, que ha reduït dràsticament les poblacions nord-americanes de castanyers.
A diferència dels boscos de coníferes, que solen cremar-se a intervals de dècades, les boscanes decídues són poc propenses a encendre’s. Les més seques, com ara les rouredes, es cremen ocasionalment i, en tot cas, el foc s’hi pot introduir des dels boscos de coníferes, a través de les franges de contacte amb les boscanes. Normalment, el foc crema els arbres més petits i els plançons, però només socarrima els arbres més grans.
A l’hivern, la neu i glaç són habituals a les boscanes i les espècies que hi viuen estan adaptades a aquestes condicions. Tanmateix, les nevades fortes i tardanes poden trencar branques i arranar les tiges més primes del sotabosc. Això sense comptar amb l’efecte combinat de neu i vent propi de les turbonades hivernals que poden afectar algunes àrees del bioma.
La inestabilitat de l’emplaçament és un factor més de pertorbació de les boscanes, sobretot de les que es fan en terrenys de pendent fort. Els arbres que creixen en un terreny en pendent acostumen a desenvolupar capçades asimètriques i a tenir un arrelament superficial. Quan assoleixen el seu ple creixement tendeixen a caure cap avall, aixafant els arbres que creixen per sota d’ells al llarg del pendent.
La intervenció humana és possiblement el factor de pertorbació més important de les boscanes decídues. Pot ser directa, en forma de tala o artigada, o indirecta, en forma de contaminació atmosfèrica o de freqüentació desconsiderada. La mateixa gestió forestal dels boscos, per conservadora que sigui, implica també una intervenció humana en la boscana.
La successió recuperadora
Les pertorbacions causades per aquests agents poden ser més o menys importants, fins i tot es pot parlar de pertorbació catastròfica si hi ha destrucció completa o quasi completa de l’estrat arbori. Les boscanes, tanmateix, són capaces de recuperar-se de les pertorbacions que experimenten i ho fan passant per una successió regular de quatre etapes: regeneració, embosquinament, maduració i bosc adult.
En l’etapa de regeneració, les llavors germinen, donen plançons i creixen rebrots nous de les soques conservades d’arbres i arbustos. L’arbreda és oberta, hi predominen normalment els arbres heliòfils i els arbustos. La biomassa és reduïda i la diversitat elevada. Aquesta etapa rarament dura més de 30 anys, i el normal és que sigui molt més curta.
L’etapa d’embosquinament comença quan els plançons que creixen, i sovint també les soques que broten, es combinen per formar una coberta completa. La competència es fa molt intensa i la densitat d’arbres disminueix ràpidament a mesura que els supervivents es fan més grans. Alguns arbustos moren i el nombre d’arbres de creixement ràpid, generalment exigents pel que fa a la llum disponible (heliòfils), disminueix ràpidament, mentre que les espècies d’arbres que toleren l’ombra es poden desenvolupar millor ocupant la franja d’estrat arbori intermedi o de sotabosc arbustiu. La diversitat disminueix, però la biomassa augmenta ràpidament. La quantitat de fusta morta continua essent mínima.
En la fase de maduració, l’estrat arbori que forma el cobricel ha assolit la seva alçada màxima, però la densitat i la diversitat d’aquest estrat es minimitzen (en el cas de moltes boscanes europees es pot arribar a una situació en què el cobricel és pràcticament monospecífic). A mesura que declina el vigor dels arbres més alts, sota seu continua la regeneració dels arbres que toleren bé l’ombra i la dels arbustos esciòfils. Finalment, la boscana assoleix una biomassa màxima amb un cobricel tancat, un estrat arbori intermedi d’arbres resistents a l’ombra i un sotabosc d’arbustos i plançons.
La fase de bosc adult es desenvolupa després de 150 a 200 anys, segons la composició de l’estrat arbori. S’hi arriba gradualment a partir de la mort d’alguns arbres vells i la consegüent formació de clarianes, i un cop els arbres esciòfils han crescut fins a ocupar amb les seves capçades el lloc que els correspon en el cobricel. Es desenvolupa una estructura pluristratificada a mesura que els arbustos i els plançons que es regeneren, incloent-hi els dels arbres heliòfils, creixen en els buits. La biomassa és més escassa que al final de la fase de maduració, però les estructures vertical i horitzontal són més complexes. Es desenvolupen alguns arbres de grans dimensions i la quantitat de fusta morta augmenta.
Un tipus diferent de successió és la que es produeix en un terreny prèviament desforestat. La major part de la superfície forestal natural del bioma ha estat talada per poder satisfer les necessitats de terres agrícoles i d’espais per a la construcció, deixant un escampall de claps de boscos i bosquines de forma i mida molt diverses. Però els canvis en la propietat o en el tipus d’explotació del territori poden invertir la tendència i permetre que el sòl desforestat es reconverteixi en bosc de manera natural o fins i tot mitjançant la plantació de nous arbres. La successió natural a partir d’un sòl desforestat (successió secundària) passa per les etapes prèviament descrites per a un bosc que es recupera d’una pertorbació, però amb diferències significatives. Normalment, la fase de regeneració és més llarga, perquè no hi ha soques a partir de les quals puguin créixer immediatament nous rebrots i les fonts de llavors poden ser llunyanes. Les espècies colonitzadores solen ser les que tenen uns mecanismes de dispersió eficients a llarga distància, per exemple, arbres pioners com ara els bedolls i els verns. Els nous boscos que es desenvolupen solen ser fragmentats, en part perquè els primers colonitzadors creixen ràpidament i formen perxades des de les quals els ocells deixen caure llavors fresques. La forma dels arbres sol ser més variable, perquè els primers colonitzadors tenen espai per a formar capçades que es poden estendre molt. Malgrat tot, quan s’ha format una bosquina, les successions secundàries es desenvolupen cap a la regeneració de boscos madurs, que poden ser difícils de distingir de les successions posteriors a la pertorbació del bosc primari.
Plantar té l’efecte d’escurçar l’etapa de regeneració i genera boscos dominats per les espècies plantades. Les plantacions antigues, sobretot aquelles en les quals s’ha descurat el manteniment, desenvolupen una estructura en estrats múltiples, perquè alguns arbres moren i altres espècies envaeixen l’espai lliure. Finalment, aquestes plantacions desenvolupen una estructura madura molt similar a la d’origen natural.
Quan una pertorbació catastròfica destrueix completament l’estrat arbori s’inicia la successió descrita anteriorment. A la major part del bioma, aquestes pertorbacions són esdeveniments poc freqüents separats per intervals ben bé de centenars d’anys. Les pertorbacions parcials o no catastròfiques, és a dir, esdeveniments que són causa de la mort d’arbres concrets o de petits grups d’arbres, però que deixen intacta la major part del bosc, són força més freqüents en la majoria de les boscanes, com ara una ventada que fa caure uns quants arbres o trenca unes quantes branques gairebé cada any, però es produeixen molt rarament huracans o turbonades que arranin completament un bosc.
De fet, és gairebé impossible que hi hagi dues pertorbacions catastròfiques pròximes en el temps, perquè els arbres triguen uns 50 anys a poder créixer prou per a ser vulnerables al vent o la sequera. Les pertorbacions més petites, en canvi, creen anualment en el cobricel espais buits d’una extensió que normalment equival a entre el 0,5 i el 2% de tota la superfície arbrada, amb una mitjana d’aproximadament un 1%. La proporció varia enormement d’un any a l’altre fins a assolir un màxim de vora el 6% després d’una sequera greu, però sovint és nul·la o insignificant durant molts anys seguits. Els espais buits també solen estar distribuïts de manera força irregular, segons la naturalesa de la pertorbació.
En boscos que mostren una mitjana de creació de buits d’un 1% anual, cada arbre ocupa el seu espai en el cobricel durant una mitjana d’uns 100 anys. Les capçades de les espècies heliòfiles poden trigar 50 anys a arribar al cobricel i normalment els arbres més tolerants amb l’ombra triguen molt més, potser fins a uns 150 anys, de manera que la major part dels arbres del cobricel de les boscanes decídues no solen viure més enllà d’uns 150 a 300 anys. Tanmateix, aquestes mitjanes amaguen variacions importants segons els gèneres o les espècies. De fet, molts exemplars de roures, de tulipers, de tsugues, de tells i d’altres arbres viuen molt més, fins a 500 o 600 anys, mentre que freixes i carpinus, per exemple, rarament viuen més de 250 anys, i algunes espècies de bedolls, pollancres i salzes viuen menys de 100 anys.
La successió i l’estructura
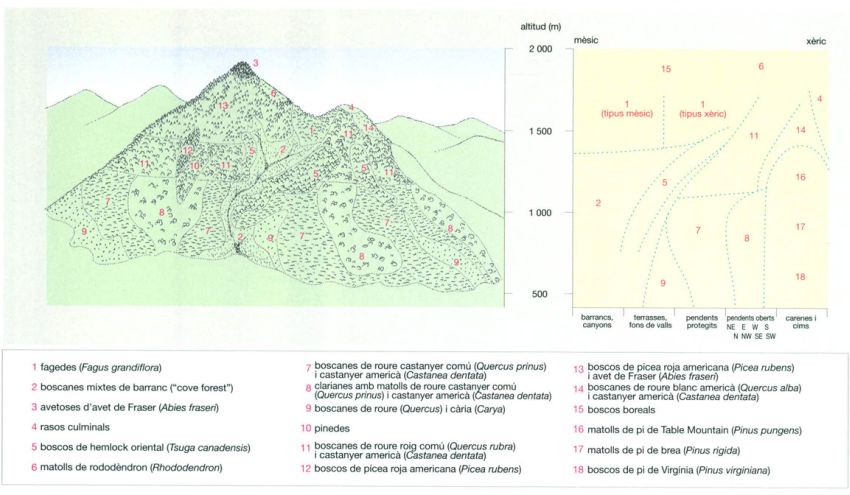
La distribució dels diferents tipus de vegetació segons la topografia a les muntanyes Great Smoky, als Apalatxes meridionals (EUA), es pot esquematitzar sobre aquesta vall muntanyosa ideal encarada a l’E. El pic més alt té 1 981 m d’altitud i el més baix 1 676 m. L’estudi de les Great Smoky per Robert H. Whittaker (1920-80) ha esdevingut un clàssic dels estudis sobre vegetació ja que va servir al seu autor per posar les bases de la teoria del continuum, inspirada en la concepció individualística de les comunitats vegetals. D’acord amb aquesta concepció, oposada a la més organicista de les escoles fitosociològiques europees (com la sigmatista, dominant entre els botànics catalans), una comunitat vegetal no és més que un grup de plantes que tenen requeriments ambientals (altitud, insolació, temperatura, etc.) similars, i, que, per tant, tenen la mateixa distribució espacial, sense que això signifiqui, com defensen aquestes altres escoles, que formin veritables associacions vegetals.
Jordi Corbera, a partir de Whittaker, 1956
Una boscana decídua és, doncs, una barreja d’espècies d’arbres amb una longevitat, una mitjana de creixement i una adaptació a les condicions d’ombra molt diferents. La seva estructura és determinada per la interacció de les pertorbacions amb el creixement, la longevitat, la periodicitat de la producció de fruits i la capacitat de regeneració de cada una de les espècies arbòries que la constitueixen. El bosc que es desenvolupa després d’una pertorbació catastròfica tindrà, inevitablement, una edat gairebé uniforme en una gran àrea.
El bosc en procés de recuperació creix ràpidament a partir de les llavors presents a terra i dels rebrots produïts per les soques i els troncs caiguts. Els únics arbres més vells són alguns de grans, aïllats o en petits grups, salvats accidentalment de la pertorbació, i els del sotabosc, que, juntament amb plançons i arbustos són menys vulnerables a la destrucció. A primera vista, la densa flota d’arbres de capçada estreta que resulta sembla una plantació, bé que els arbres no estan arrenglerats i les espècies s’hi barregen. Quan la darrera pertorbació catastròfica data d’uns quants segles i la boscana ha assolit una situació de bosc adult, la seva estructura és determinada pel patró de formació de buits. Quan es forma un espai buit, s’omple de plançons (rarament rebrots de soques) que es converteixen en una perxada i finalment es redueixen a només un o dos arbres que creixen fins a arribar a omplir el cobricel amb la seva capçada. En un bosc on la majoria d’anys es formen petits buits, hi ha un assortiment complet d’edats i es presenten tots els estadis possibles de creixement. El bosc és, per tant, un mosaic de petits grups d’arbres d’edat uniforme barrejats íntimament; de grans arbres de l’estrat superior situats a tocar dels buits més recents i de rodals de plançons i perxades esprimatxades que intenten abastar el cobricel.
Una estructura així, propera a la d’un bosc gestionat per selecció de grups d’edat, és típica de les boscanes decídues complexes, on les pertorbacions catastròfiques són poc freqüents i la major part de les ordinàries consisteixen en la mort d’uns arbres concrets a causa de l’edat, els cops de vent i els atacs fúngics. Allà on són freqüents altres tipus de pertorbacions, es desenvolupen, en conseqüència, estructures diferents. Sobre pendents escarpats, on la inestabilitat de l’emplaçament causa la caiguda primerenca dels arbres, els buits s’allarguen pendent avall (ja que tots els arbres cauen cap al mateix cantó) i l’estructura és oberta, amb un dens estrat arbustiu. En aquells llocs on els incendis són freqüents, els arbres més grans sobreviuen millor que els plançons i els arbres més petits (que només sobreviuen en grups salvats de les flames) i el resultat és un bosc alt, irregular i biestratificat. En planes al·luvials, on el moviment natural de les lleres dels rius destrueix els boscos però diposita bancs de sorra on en poden créixer de nous, el resultat és una sèrie d’arbredes d’edat uniforme que reflecteixen fidelment la història dels desplaçaments de la llera.
Les espècies poc tolerants amb l’ombra només prosperen en buits de dimensions més grans, però les més tolerants creixen en buits de qualsevol mida. De fet, molts buits s’omplen amb plançons que anteriorment creixien a l’ombra. Aquesta ‘regeneració avançada’ implica invariablement espècies esciòfiles. Per tant, en intervals prolongats entre pertorbacions catastròfiques, la composició del bosc tendeix cap a la dominància d’aquestes espècies i així, en regions menys alterades, fagedes i telledes esdevenen característiques dels sòls mèsics. Els arbres heliòfils (roures, tulipers) tenen la seva oportunitat després de pertorbacions catastròfiques o bé quan poden ocupar grans buits formats com a conseqüència de la mort simultània de grups d’arbres. En boscanes de creixement antic és comú trobar-hi una generació de roures vells, relíquia d’un incendi o una destrucció important de tres o quatre segles enrere, que creixen sense successors enmig d’una bosquina més jove dominada per espècies esciòfiles.
La composició dels boscos canvia de manera natural al llarg de dècades i segles. Després d’una pertorbació important, els arbres efímers (sovint exigents pel que fa a la llum) i els arbustos moren primer, deixant els arbres més longeus. En el bosc adult, els buits s’omplen amb una gran varietat d’espècies, segons l’arbre que hi havia abans al cobricel o el que doni bons fruits en un any determinat. En molts boscos caducifolis temperats, alguns arbres són incapaços de regenerar-se sota un cobricel d’arbres de la mateixa espècie, és a dir, que l’espècie principal alterna la dominància en qualsevol punt particular del bosc. Exemples d’aquestes espècies alternants inclouen els faigs i els avets, els roures i els bedolls, i el fals plàtan i el freixe de fulla ampla. Inevitablement, l’atzar juga un paper molt important com a factor determinant del buit que ocuparà cada espècie. Pel que fa als buits, la composició canvia de manera calidoscòpica, i cada espècie individualment només es pot regenerar fortament durant uns quants anys de cada segle, quan hi ha una conjunció afortunada entre dos factors: abundància de fruits i emplaçaments de regeneració adequats. A un nivell més ampli, la composició del bosc és un estat estable-fluctuant que canvia lentament com a resultat de modificacions en el clima i en els sòls.
La variabilitat vinculada al substrat i al relleu
A una escala territorial prou àmplia, la major part de les boscanes decídues passarien en un moment o altre per una etapa de bosc adult. De fet, la modelització dels índexs de pertorbacions i creixement suggereix que, en estat natural, el bosc madur ocuparia un 75% de la superfície del bioma. En les discontinuïtats naturals de les valls d’inundació, aiguamolls, pendents escarpats i cingles, on la inclinació és massa pronunciada o els sòls massa prims per a sostenir arbres, i en el límit altitudinal del bosc, on les acumulacions de neu i els solcs dels allaus impedeixen el creixement d’arbres, es fragmentaria de manera permanent la coberta natural. Els espais oberts temporals serien transitoris, excepte en sòls pobres o humits on els efectes combinats del creixement lent i dels animals brostejadors impedirien la regeneració dels arbres durant dècades. Naturalment, les coníferes formen part del bosc en sòls secs i pobres, en emplaçaments contínuament alterats i en el límit altitudinal del bosc, és a dir, en els límits geogràfics i edàfics de les boscanes decídues.
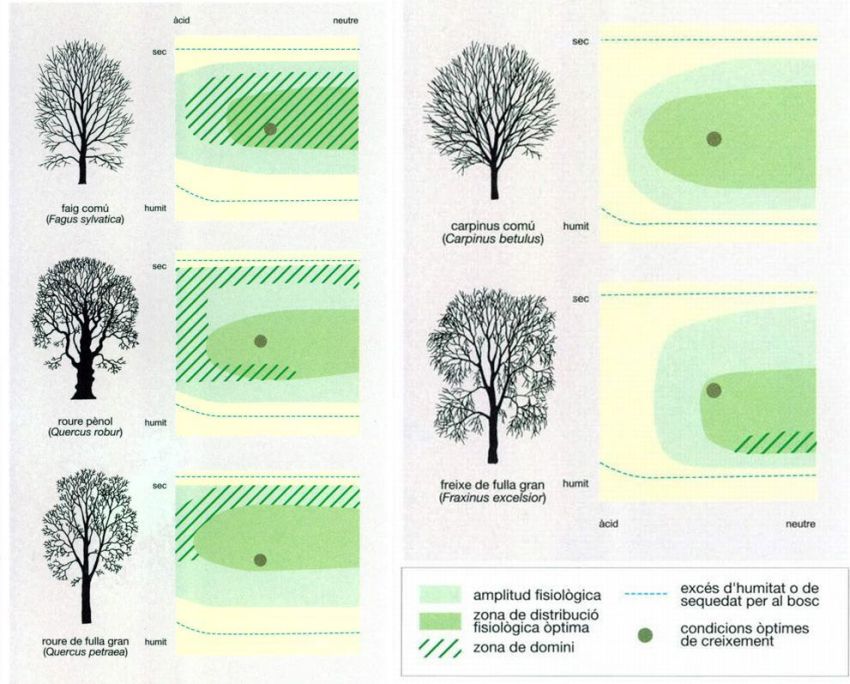
Les característiques edàfiques determinen la dominància d’unes o altres espècies arbòries, segons el grau d’exigència de cadascuna i la seva habilitat competitiva, tal com exemplifica aquest gràfic dels requeriments edàfics i la dominància de cinc arbres caducifolis europeus a la plana de la Baixa Saxònia (Alemanya). Cada espècie arbòria es desenvolupa dins uns límits determinats d’humitat i d’acidesa del sòl, i, bé que poden ser molt amplis, sempre hi ha una zona òptima per al creixement. Les espècies que no són gaire exigents pel que fa als requeriments edàfics poden créixer a pràcticament tots els ambients aptes per al bosc (representats al gràfic per les línies blaves discontínues), i per tant són les que dominen, com passa a Europa amb el faig (Fagus sylvatica) i els roures (Quercus). Les espècies més exigents perden competitivitat, i només poden créixer en zones amb condicions edàfiques encara acceptables però no ocupades per altres arbres.
Jordi Corbera, a partir de G. Jahn, a Röhrig & Ulrich, 1991
En qualsevol cas, la composició i la dinàmica de les boscanes decídues varien segons el substrat i l’altitud. Un exemple clàssic seria el de les muntanyes Great Smoky, al S dels Apalatxes, a l’orient nord-americà. Aquesta regió conté una gamma molt àmplia de tipus de bosc, des de les molt productives boscanes de les fondalades encaixades i ombrívoles (“cove forests”) de sòls humits, profunds i bàsics a baixa altitud, fins als boscos subalpins de pícea roja americana (Picea rubens) i d’avets de Fraser (Abies fraseri) dels cims. El patró de distribució dels tipus de bosc és determinat tant per la geomorfologia com per l’altitud. Adopta la forma d’estatges altitudinals poc definits, interromputs per claps que s’allargassen en el sentit de l’altitud i de corredors estrets definits per barrancs i carenes. L’esquema d’aquesta zonació també posa en relleu les tres circumstàncies sota les quals els boscos de coníferes de fulla perenne es troben associats amb boscanes decídues: en estadis ombrívols i tardans de la successió (per exemple el bosc de tsuga); en emplaçaments secs i de baixa fertilitat (per exemple les pinedes); i a gran altitud, on sovint es troba una forma o altra de bosc boreal o assimilable.
Els boscos de l’Europa central presenten una zonació molt similar. A una escala territorial àmplia, els estatges altitudinals canvien amb la latitud i l’exposició. Al bell mig de la zona temperada de l’Europa central, en altituds baixes, la zonació adopta forma de boscanes complexes amb roures i castanyers o roures i carpinus i va ascendint cap a les fagedes i els boscos d’avet blanc i de pícea fins a arribar, al límit de l’estatge alpí supraforestal, als boscos de làrix, pícea o pi. Dins dels boscos caducifolis complexos de les terres baixes, la major part dels arbres grans creixen millor en aquells sòls que tenen una humitat moderada i un pH bàsic, però el bedoll tendeix a dominar en emplaçaments mèsics, deixant que altres espècies dominin en tipus de sòls marginals.
Les pertorbacions i la gestió forestal poden desvirtuar temporalment aquestes relacions, forçant l’admissió d’una proporció molt superior de gèneres heliòfils, alguns efímers, com ara els bedolls, mentre que d’altres, com els roures i els tulipers, poden viure fins a 500 anys. Les pertorbacions també són responsables, fins a un cert punt, dels patrons altitudinals i topogràfics, perquè alguns emplaçaments sofreixen més pertorbacions que d’altres i per tant faciliten la regeneració i la presència permanent d’arbres heliòfils, per exemple, en boscos que creixen en pendents forts, a gran altitud i a les riberes.
Estatges de vegetació als Alps suïssos
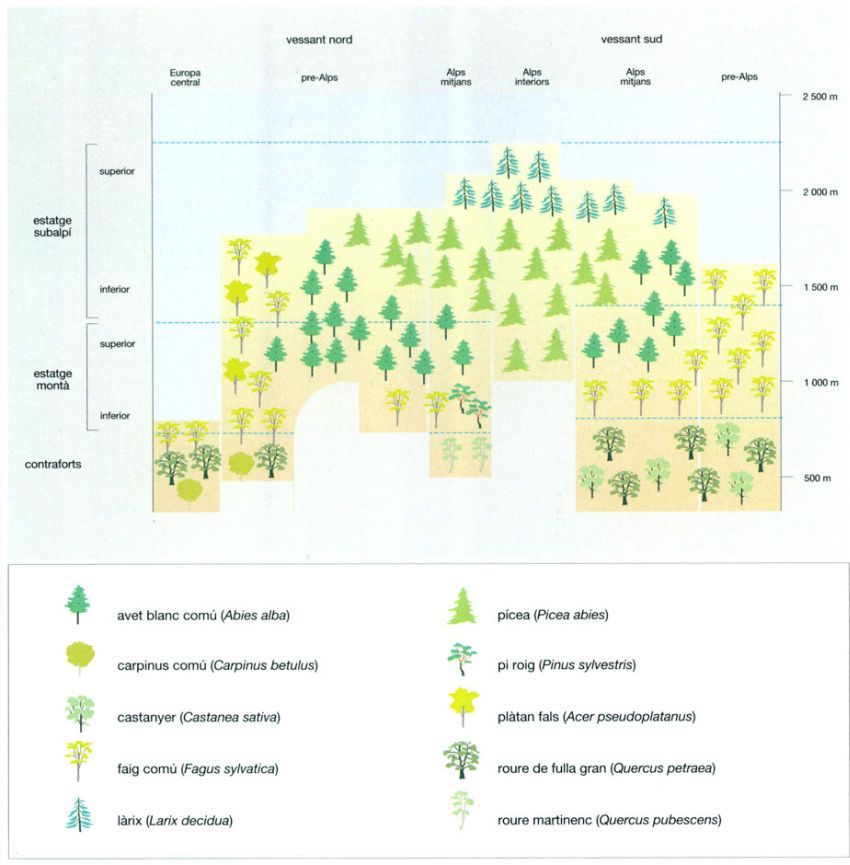
Jordi Corbera, a partir de G. Jahn, a Röhrig & Ulrich, 1991
La distribució de la vegetació als Alps suïssos és resultat del clima general, les condicions ambientals particulars de cada àrea i la influència humana. Als contraforts de les muntanyes, entre els 400 i els 600 m d’altitud, dominen els boscos de roure (Quercus), que al vessant N es barregen amb carpinus (Carpinus) i al vessant S amb castanyers (Castanea). Segurament aquesta predominància del roure i no del faig (Fagus) és deguda a la gestió del boscos pels humans. L’estatge montà (800-1 300 m) es caracteritza per boscos mixtos de faig (Fagus sylvatica) i avet blanc (Abies alba), que, segons les condicions edàfiques, van acompanyats de plàtans falsos (Acer pseudoplatanus), freixes de fulla gran (Fraxinus excelsior) o, als vessants calcaris, tells (Tilia). Aquestes formacions són substituïdes a l’estatge subalpí (1 300-2 200 m) per fagedes amb pícea (Picea abies), que als Alps interiors apareixen ja a alçades inferiors, a partir de l’estatge montà superior. Finalment, l’estatge subalpí superior és dominat per làrixs (Larix).