Els tipus biològics
“La tundra és un bosc sense arbres”, diu un aforisme rus. Un aforisme encertat perquè posa de relleu la similitud fisiognòmica entre la tundra i el sotabosc dels boscos boreals. Arbustos nans (nabius i gerderes) i un dens estrat de molses i líquens són trets comuns prou significatius.
Els líquens
Juntament amb els briòfits, els líquens són components indefectibles dels paisatges de la tundra. Els líquens de la tundra mostren una sorprenent diversitat d’espècies, sobretot si es pensa en les severes condicions climàtiques en què s’han de desenvolupar. A la tundra de la península de Taimir, a l’extrem septentrional d’Euràsia, se n’ha identificat 226 espècies, a l’illa de Devon, a l’arxipèlag àrtic canadenc, 260 més, i només a les zones de tundra i de tundra arbrada de la part oriental d’Europa se n’hi fan unes 500. Aquest índex de biodiversitat relativament elevat s’explica perquè els líquens, com les molses, tenen uns requeriments molt limitats pel que fa a l’ambient i el clima en què viuen. És per això que són distribuïts a tota la zona de la tundra, de les regions més septentrionals a les més meridionals, incloent-hi la tundra arbrada dels confins de la taigà.
D’acord amb la sistemàtica adoptada en aquesta obra, els fongs i els líquens, organismes àmpliament presents en gran part de la tundra, no són plantes, sinó que constitueixen un regne a part: el dels fongs. Atesa la importància d’aquest regne (en particular dels fongs liquenificats o líquens) en el poblament i el paisatge de la tundra, a diferència d’altres biomes, en els quals, o no es tractaven o se’n feia una menció escadussera al capítol de flora i poblament vegetal, en el títol i en el contingut del present capítol es fa esment exprés de la micota, és a dir, del poblament fúngic, representat bàsicament pels líquens en el cas de referència.
Per la morfologia del seu tal·lus, s’han diferenciat tradicionalment tres tipus de líquens: els crustacis, que formen crostes sobre el substrat colonitzat, del qual esdevenen inseparables; els foliacis, que formen estructures laminars unides només al substrat per alguns punts o mitjançant alguna mena d’òrgans de fixació; i els fruticulosos, que es fixen al substrat per un sol punt, l’anomenat disc basal, i són poc o molt ramificats. A la tundra es poden trobar líquens de totes tres menes. Els líquens crustacis colonitzen les pedres grans i les superfícies de les roques; la seva aparença és molt diversa: taques brillants de diferents colors (taronja, groc llimona, oliva, verd poma, etc.), punts separats o agrupats de color negre, marró o gris; fins i tot, de vegades arriben a presentar l’aspecte d’un jeroglífic. Els líquens fruticulosos es troben entre les espècies dominants de la tundra (juntament amb algunes molses i plantes vasculars) i poden existir en forma de densos poblaments exclusivament liquènics. En canvi, els líquens foliacis no tenen un paper tan apreciable en la formació de la biocenosi; així i tot, els seus tal·lus relativament grans (de fins a 5 cm de diàmetre o més), amb les seves diferents tonalitats de gris i de verd, són força freqüents a les tundres de molses i líquens, a les tundres arbustives i a les mulleres embosquinades.
El creixement dels líquens, en particular dels crustacis, és característicament lent, però constant. Si es comparen les dimensions del tal·lus amb la seva edat, en líquens crustacis de la tundra es poden observar índexs de creixement de prop de 0,2 a 0,3 mm anuals. Fins i tot n’hi ha una espècie en la qual s’ha mesurat un creixement de només 0,004 mm l’any: tot un rècord de lentitud, ja que això significa que en un miler d’anys aquest liquen només arribaria a mesurar 4 mm més! Tanmateix, n’hi ha que han assolit creixements que fan pensar en longevitats que es poden comptar per segles. Datacions radiocronològiques han confirmat que, contràriament al que s’havia cregut durant molt de temps, són nans (els líquens) i no gegants (els grans arbres) els éssers vius que assoleixen edats més provectes. Així, com que els líquens crustacis colonitzen els substrats de pedra abans que cap planta, és possible determinar l’edat de molts materials (si més no, el temps que fa que estan exposats a la colonització pels éssers vius) a partir de la mida dels líquens que els han colonitzat. Mitjançant aquesta tècnica, anomenada liquenometria, s’ha pogut saber, per exemple, que les morenes dels Urals septentrionals s’havien format força recentment, fa poc més de 700 anys, al voltant dels anys 1230 a 1270 de l’era cristiana.
Els líquens fruticulosos del gènere Cladina (els anomenats líquens dels rens, ja que són un excel·lent aliment a l’hivern per a aquests animals) renoven la seva plena activitat vital així que arriba la primavera, tan bon punt s’ha fos la neu; en aquell moment, la temperatura del seu tal·lus és de només 5°C. Els líquens dels rens (Cladina rangiferina, C. alpestris, C. sylvatica, etc.) mai no arriben a fer enllà de 6 o 7 cm de llargada encara que visquin uns quants decennis, i en el seu cicle vital es poden distingir tres fases de creixement. La primera, de creixement accelerat, pot durar de 5 a 25 anys (de mitjana uns 10) i acaba quan el tal·lus assoleix la seva llargada definitiva a raó, al començament, d’entre 0,5 i 0,7 mm anuals i, al final, fins a deu vegades més, d’uns 5 o 7 mm per any. La segona fase, de creixement estabilitzat, dura uns quants decennis, de vegades fins i tot un segle, i, si bé el creixement roman estable, la planta ja no augmenta de llargada. Com que els líquens dels rens requereixen molta llum per al seu creixement, la seva part inferior va morint com a resultat de l’ombra que li fan les branquetes més altes; aquest fenomen és especialment palès quan el substrat és humit o en els anys de molta precipitació estiuenca. La tercera fase, de progressiva inhibició del creixement, es caracteritza per un alentiment del creixement, fins a la seva total inhibició, i la mort gradual del liquen.
El creixement dels líquens foliacis és comparable al de les formes fruticuloses i, a més, els uns i els altres responen als canvis d’humitat ambiental de manera més aparent; els seus tal·lus, extremament secs i retrets als períodes de sequera, es fan elàstics, suaus i fins i tot desprenen una olor peculiar i fresca després de les pluges fortes. Les branquetes dels líquens dels rens, per exemple, s’estiren i els tal·lus de Peltigera aphthosa, més propers a la superfície del sòl, prenen una coloració verda brillant. En aquest estat de saturació d’aigua, els líquens perden la seva habitual fragilitat i sofreixen menys els efectes mecànics, del transport o de cops, per exemple.
En ser “lents però constants”, els líquens són uns testimonis vius, encara que silenciosos, dels canvis climàtics, els increments de la contaminació ambiental i determinades catàstrofes ecològiques sobtades. Hi ha tota una escala d’espècies de líquens que tenen requeriments i sensibilitats diferents, de manera que el mal causat pels diferents contaminants sobre cadascuna és selectiu i diferenciat. Això ha permès desenvolupar tècniques que, partint de les variacions en l’índex de creixement i el coneixement de la diferent capacitat d’acumulació no selectiva d’unes i altres espècies de líquens, donen indicacions i mesures referents al grau de contaminació ambiental i als seus agents causals.
Les molses típiques i els esfagnes
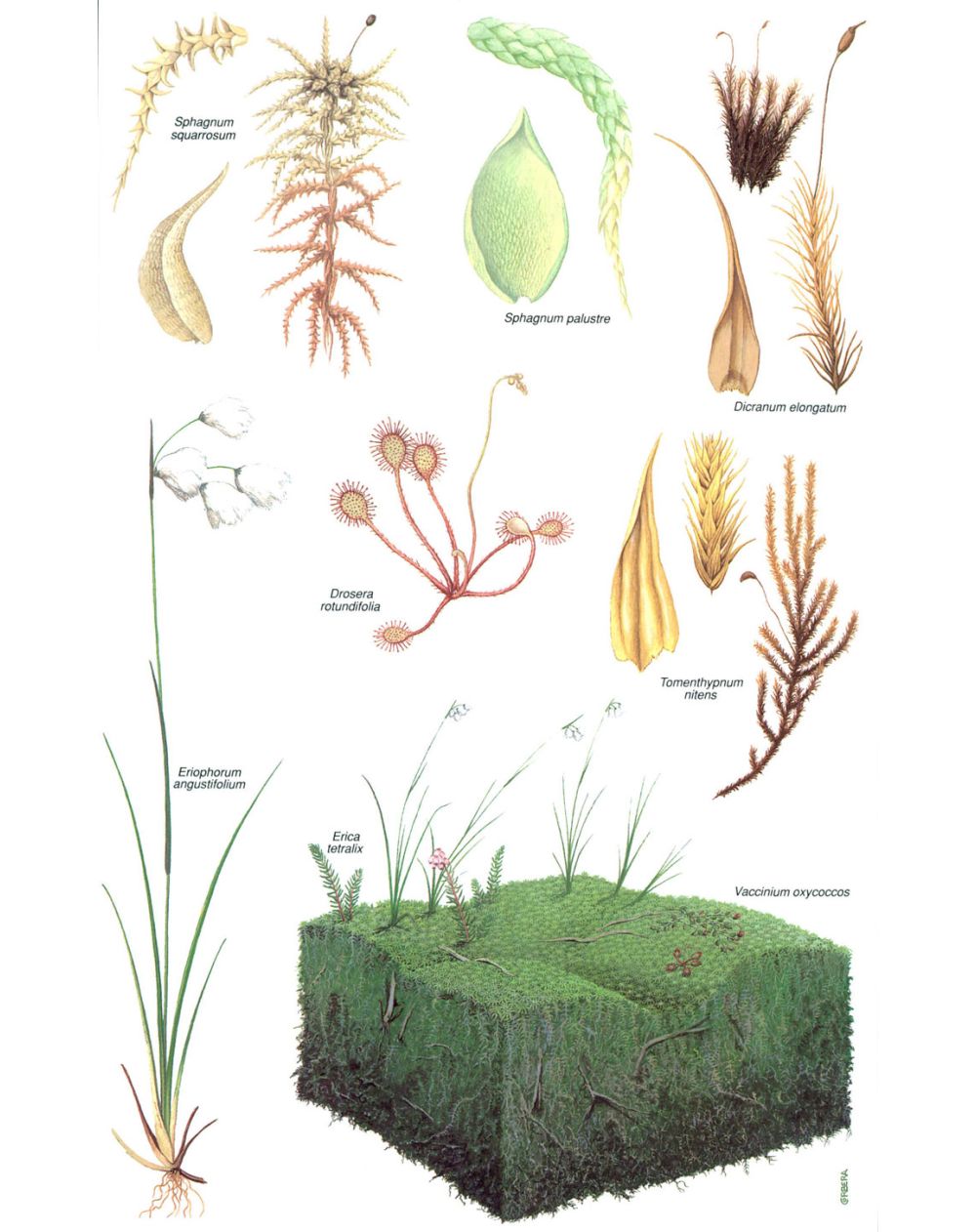
La vegetació de les torberes és formada, bàsicament, per molses del grup de les esfagnes. Les esfagnes acidifiquen el medi, ja que tenen un elevat contingut en àcids orgànics, i les plantes que creixen a les torberes han d’estar adaptades a un ambient àcid. Quan les molses (com Dicranum elongatum, Sphagnum squarrosum, S. palustre i Tomenthypnum nitens) han originat un sòl més o menys estable, algunes plantes superiors són capaces d’arrelar a les torberes, com per exemple la dròsera (Drosera rotundifolia), el bruc d’aiguamoll (Erica tetralix), la cotonera de fulla estreta (Eriophorum angustifolium) i el Vaccinium oxycoccos. En canvi, els líquens, tan abundants a la tundra, no apareixen a les torberes, ja que hi ha massa aigua perquè hi puguin créixer.
Jordi Corbera, a partir de fonts diverses
Les molses són plantes molt difoses a tota la tundra i, a més, també n’hi ha força diversitat d’espècies, de vegades algunes dotzenes per metre quadrat. La coberta de molses té una funció termoreguladora molt important. A l’hivern, juntament amb la capa de neu, protegeixen del fred les tiges repents perennes, els rizomes, les arrels i els borrons regeneradors (fins i tot els que són en estat latent); a l’estiu, amb la seva intensa evapo-transpiració, crea un microclima molt més favorable.
Com més humit és un hàbitat, més augmenta als teixits de les molses el contingut de nitrogen i de reserves. Així, els hàbitats amb elevada disponibilitat de nitrogen afavoreixen tant el creixement de les molses com el de les plantes vasculars. En conjunt, però, el creixement de les molses tampoc no és gaire ràpid, i representa d’un 30 a un 40% de la producció total anual de la fitomassa aèria de les comunitats vegetals de la tundra. Així i tot, en condicions molt favorables de disponibilitat d’aigua, les brials, és a dir, les molses típiques, es poden allargar de manera inusualment ràpida. Per exemple, a l’illa de Devon, a l’arxipèlag àrtic canadenc, s’ha observat un creixement de quasi 3 cm en un exemplar de Meesia triquetra que creixia en una situació favorable (al costat d’un curs d’aigua), mentre que el creixement d’una mota de jónçares en una mullera era deu vegades inferior.
A les mulleres, àmpliament distribuïdes per tota la tundra, predominen els esfagnes (Sphagnum), i més com més al sud, fins al punt que les torberes d’esfagnes desplacen les de molses hipnobrials, habituals a les latituds més septentrionals. Els esfagnes són una família monogenèrica de molses molt especialitzades i de característiques peculiars, entre les quals el sobtat canvi de color en funció dels canvis estacionals d’intensitat de llum. La síntesi de pigments vermells i bruns per part d’aquestes plantes es considerà inicialment com una adaptació per a protegir-se de la reverberació de la llum, però en realitat resulta estar més relacionada amb el cicle diürn de les temperatures ambientals. S’ha observat que l’enrogiment dels esfagnes va lligat a un retardament del creixement, sempre que la temperatura nocturna passi de manera sobtada de 2°C positius a prop de –3°C. En produir-se aquest descens de temperatures, el contingut de clorofil·la de l’esfagne disminueix, de manera que el color rogenc reflecteix el seu estat fisiològic. Els repetits canvis de color són indicatius d’una inestabilitat en els processos metabòlics als teixits d’aquestes plantes. El fet que, al final del període vegetatiu estival, els esfagnes de les torberes es tornin vermells (de fet prenen diverses tonalitats que van del rosa al porpra, sovint força atractives visualment) es pot comparar amb l’acoloriment que prenen les fulles de les plantes vasculars caducifòlies en arribar la tardor, abans de desprendre’s.
Segons uns estudis de la dinàmica estacional de l’activitat fotosintetitzadora dels esfagnes fets a Barrow, a l’extrem septentrional d’Alaska, la fotosíntesi neta augmenta, des del juny fins al començament d’agost, en una proporció d’1,5 a 2 vegades per unitat de massa seca, i de 2,5 a 3 vegades per unitat de superfície. En el cas de les molses típiques és a l’inrevés: s’observa una minva de la fotosíntesi neta durant el mateix període; en el cas de la molsa Dicranum elongatum, per exemple, la diferència era d’1,3 vegades per unitat de massa i de 2,6 vegades per unitat de superfície. La conclusió que se’n pot treure és que les molses típiques i els esfagnes responen de manera diferent als canvis d’intensitat de la llum i la duració del fotoperíode.
Els esfagnes viuen sempre en aigües àcides i poc mineralitzades, però ells mateixos tenen la capacitat d’acidificar, mitjançant processos d’intercanvi iònic, el medi aigualós que poblen. Aquesta capacitat és determinada pels àcids orgànics (principalment màlic i cítric) i els àcids poliurònics, els quals constitueixen d’un 10 a un 25% de pes sec dels esfagnes.
Les plantes vasculars
A més de les torberes, amb el tou de molses que les caracteritza, hi ha altres comunitats higròfiles en les quals les plantes vasculars tenen el seu paper: mulleres amb càrexs i cotoneres i altres herbeis, fonamentalment els pradells llacosos d’arctòfila (Arctophila fulva), freqüents a les ribes dels rius i dels llacs. L’arctòfila, una gramínia típicament circumpolar, és coneguda pel seu polimorfisme polivalent: es tracta d’una planta molt valuosa com a aliment per als rens i per als ocells aquàtics, ben adaptada a hàbitats inestables, negats temporalment o contínuament. Els seus rizomes disposen d’unes cavitats especials plenes d’aire per tal que la limitada aeració dels terrenys inundats no impedeixi la respiració de la planta.
Les formacions d’arbustos (habitualment salzes, verns o bedolls) de la tundra solen trobar-se a les planes inundables dels rius. L’efecte tèrmic determina que en aquestes zones hi hagi un alt índex de creixement. El creixement anual del salze Salix dasyclados, característic de les tundres meridionals, pot ultrapassar els 40 cm. El bedoll nan (Betula nana), el “yernik” dels russos, és una espècie arbustiva típica de la tundra i d’àmplia distribució també a la tundra arbrada. Segons les condicions ambientals, el “yernik” pot créixer en forma semiprostrada (fins a 70 o 80 cm d’alçada), procumbent (de 30 a 35 cm) o completament repent i arrelat a la capa de molses de les torberes (de 10 a 30 cm per sobre del tou d’esfagnes). El creixement del “yernik” és fonamentalment vegetatiu: als 60 o 70 anys d’edat la tija principal i el seu sistema d’arrels moren, mentre que les branques de l’esquelet que resta treuen arrels i formen nous i petits arbustos, cosa que sol ocórrer més d’una vegada.
Les herbes graminoides tenen un paper important a la tundra. De càrex ciclocarp (Carex globularis), per exemple, n’hi ha gairebé pertot arreu; té rizomes subterranis perennes que emeten cada any brots vegetatius epigeus que duren un sol estiu. Els brots del càrex de tija arrodonida (C. rotundata), per contra, poden viure tres o quatre anys, però només una o dues de les fulles superiors resisteixen el període hivernal; durant els primers anys creix un brot epigeu vegetatiu, que després floreix, fructifica i mor. Aquesta espècie pertany al grup de plantes verdes durant l’hivern amb brots de diferents edats. El càrex aquàtic (C. aquatilis) i la cotonera de fulla estreta (Eriophorum polystachyon) tenen el mateix tipus de desenvolupament de brots epigeus que C. rotundata, però amb la diferència que poden durar d’un a dos anys més.
Les estratègies adaptatives
En unes latituds on l’hivern dura prop de tres quartes parts de l’any, els organismes, especialment els vegetals, es veuen obligats a adaptar-se a les baixes temperatures i a la limitada disponibilitat d’aigua durant una temporada. D’aigua n’hi ha tanmateix, però a efectes de la vegetació només resulta utilitzable la que es troba en estat líquid, que escasseja. Les plantes de la tundra, víctimes de sequera funcional en un món ple d’aigua sòlida, han de passar la major part de la seva vida en estat latent.
La defensa enfront del fred i el vent
Els vegetals de la tundra protegeixen la majoria de les arrels, dels rizomes (en el cas de les herbes) i dels brots i les branques subterrànies (en el dels arbustos i les mates) mantenint-los als horitzons subsuperficials del sòl, a menys de 20 cm de fondària, on el règim tèrmic és menys hostil. És una manera d’assegurar-se un nou període vegetatiu, ja que és en aquests òrgans on es concentra la reserva alimentària produïda durant l’estiu precedent. Així i tot, els rizomes subterranis i les tiges amb borrons en estat latent de vegades necessiten una protecció addicional enfront del fred: és per això que les cotoneres i els càrexs formen nombrosos borrons, molts dels quals no s’utilitzen, i mantenen les bases de les fulles mortes unides a la tija.
Una altra manifestació d’aquestes adaptacions al fred és la producció d’un nombre excessiu de gemmes, tant axil·lars com adventícies, per part dels arbustos, que també produeixen, d’altra banda, un nombre considerable de gemmes foliars o de botons florals. Aquesta mena de “reassegurança” posa de manifest la seva eficàcia, sobretot en aquells casos en què, a causa del fred o de danys mecànics o zoogènics, moren algunes branques o fins i tot una part considerable de la planta. Aquesta forma de protecció, pràcticament universal entre les plantes llenyoses de la tundra, afavoreix el restabliment de la coberta vegetal després d’una alteració. Certament no totes les gemmes foliars es desenvolupen formant una fulla: una gemma del bedoll nan o “yernik” (Betula nana), amb 21 o 22 primordis foliars, pot produir, per exemple, de 7 a 8 fulles, mentre que en el romaní de torbera (Ledum palustre) només es desenvolupen de 12 a 14 fulles de les 26 o 28 que podrien fer-ho, i la resta de primordis foliars moren.
L’adaptació de les plantes a les baixes temperatures de l’hivern també es manifesta a nivells anatòmics i bioquímics. La gran concentració de monosacàrids als òrgans perennants de la planta ocasiona la minva de l’activitat respiratòria (molt important per a estalviar la despesa de les reserves nutritives). A més, els monosacàrids eviten la coagulació de proteïnes a temperatures per sota de 0°C, inhibint la cristal·lització de l’aigua. Aquests sucres solubles en aigua s’acumulen, a temperatures inferiors a 0°C, com a resultat de la despolimerització del midó insoluble produït en la fotosíntesi. La transició del fred a la calor intensifica els processos d’oxidació-reducció, cosa que fa augmentar als teixits de les plantes el contingut d’hemicel·lulosa, nitrogen d’origen proteic, vitamines i pigments; simultàniament, hi ha una producció activa d’enzims (catalases, peroxidases, oxigenases, etc.). En el cas de les plantes que no presenten cap procés bioquímic de resistència al fred, s’observa un altre tipus d’adaptació: una oxidació exhaustiva de la reserva d’aliment.
Les plantes de la tundra necessiten protegir-se del vent, i, en aquest cas, la selecció ecotòpica ha de jugar al seu favor: les petites matetes en coixinet, com les saxífragues (Saxifraga) i algunes minuàrties (Minuartia arctica, M. macrocarpa, etc.) es distribueixen especialment als hàbitats pedregosos. S’arrapen així a la superfície de la roca i amb prou feines en sobresurten. D’aquesta manera no pateixen tant la força de l’impacte del vent.
L’economia hídrica
El creixement de les plantes de la tundra tan bon punt es fon la neu és afavorit per la capacitat de les arrels de funcionar normalment quan la temperatura del sòl oscil·la al voltant dels 0°C (en el cas de la gerdera herbàcia Rubus chamaemorus, per exemple, a partir dels –2°C). El fet que per sota d’aquestes temperatures l’aigua congelada resulti inaccessible per a les plantes donà lloc a la idea d’un “eixut fisiològic” que afectaria els hàbitats de la tundra i a la idea que hi havia d’haver una rica varietat de plantes amb diferents adaptacions al xerofitisme. Actualment, tanmateix, aquestes idees s’han abandonat perquè no s’han pogut provar experimentalment. El percentatge real d’espècies de tundra amb una estructura xeromòrfica és sorprenentment baix; al centre de Taimir, per exemple, és de només el 9,6% dels vegetals (15 espècies). En canvi, s’ha trobat que el 90% de les espècies tenia una estructura adaptada a una humitat moderada o excessiva (mesomòrfica, hidromesomòrfica o mesohidromòrfica), que correspon al règim hídric dels tipus bàsics d’ecotops més àmpliament representats a les regions septentrionals d’Amèrica i d’Euràsia.
Les diferències en l’anatomia i la morfologia de les fulles, com també en la seva longevitat, determinen les característiques funcionals de les plantes amb diferents tipus de ritmes fenològics. Així, la mitjana de contingut d’aigua a les fulles de les plantes perennifòlies durant l’estació de creixement se situa entre el 51% i el 66%, mentre que ho fa entre el 65 i el 79% a les de les espècies caducifòlies, amb valors màxims durant el període de rebrotada més intensa. L’índex de transpiració d’aquests dos tipus d’espècies és de 0,1 a 0,3 grams d’aigua per gram de pes fresc i de 0,4 a 0,8, respectivament. El valor de l’índex per al grup intermedi, les plantes semiperennifòlies, és de 0,3 a 0,4 grams per gram de pes fresc, amb un contingut d’aigua a les fulles comprès entre el 60% i el 75% del pes fresc.
La proporció entre les diverses formes d’aigua a les fulles també és diferent entre les espècies perennifòlies i les caducifòlies, per bé que en unes i altres preval l’aigua combinada. Quan es detura el creixement intensiu, la relació entre aigua lliure i aigua combinada se situa entre 0,3 i 0,5 a les plantes perennifòlies i per sota de 0,2 a les plantes caducifòlies; quan arriba la tardor, aquests valors augmenten fins a 0,6-0,9 en les primeres i fins a 0,3-0,4 en les segones. La pressió osmòtica dels sucs cel·lulars sol ser més elevada a les plantes perennifòlies (de 29 a 40 atm) que a les caducifòlies (de 14 a 29 atm). Tot i així, durant el període de creixement intensiu s’enregistren valors comparables (de 16 a 18 atm), si s’examinen fulles de la mateixa generació.
Les adaptacions del cicle vital
La brevetat del període vegetatiu produeix la famosa “explosió vital” característica de la primavera de la tundra. És tan breu aquest esclat vital, que la majoria de les fanerògames no poden florir i fructificar el mateix any, sinó que han de posposar la fructificació l’any següent al de la seva floració. A la tundra no es podria reproduir ni podria sobreviure cap planta estrictament anual; les festuques (Festuca brachyphylla, F. hyperborea i F. lenensis), els càrexs (Carex), els nabius (Vaccinium), les poligonàcies, les ericàcies i les altres plantes vasculars de la tundra són totes plantes perennes que viuen el temps més llarg possible per tal de produir, alternativament, un any flors i un altre llavors.
La primavera a la tundra és més aviat una “primavera de llum” que no pas una “primavera de calor”. Les plantes dels hàbitats més càlids són les que floreixen abans, com és el cas de les superfícies rocoses que queden lliures de neu una o dues setmanes abans que les àrees adjacents. Encara no han passat tres o quatre dies que les roques de l’illa de Devon, per exemple, comencen a despuntar entre la neu al començament de la primavera, que Saxifraga oppositifolia comença a florir. De set a vint dies més tard ho fan altres espècies com la dríade de fulla entera (Dryas integrifolia), el bruc àrtic (Cassiope tetragona), el pedicularis llanós (Pedicularis lanata) i la silene acaule (Silene acaulis). Les condicions de temperatura a les esquerdes de les roques i a les pedrusques són molt favorables per a l’activació de la fenologia de les plantes i per a la seva florida breu, però abundosa. Aquesta varietat en les condicions fa que un percentatge d’individus en flor sigui molt variat. A la ja esmentada illa de Devon, per exemple, s’han observat simultàniament els casos de set espècies, de les quals només d’un 30% a un 50% dels exemplars eren en flor, mentre que cinc altres espècies tenien en floració al mateix moment d’un 70% a un 100% dels exemplars.
Els dies de sol sense núvols les plantes poden arribar a assolir una temperatura superior a l’ambiental. Tant és així, que s’ha trobat en diverses espècies de plantes una relació entre el color de la flor i la diferència de temperatures assolida respecte a l’aire que l’envolta. La diferència més petita (prop d’1-1,5°C) correspon a les plantes amb la corol·la blanca, com ara les drabes (Draba) o el bruc àrtic (Cassiope tetragona). La diferència més gran (de 4,2°C) s’aprecia a l’oxítrop negre (Oxytropis nigrescens), una papilionàcia les flors de la qual tenen un bonic estendard de color violaci.
La fase fenològica de florida s’allarga quan fa fred. Així, en el cas de l’esmentat oxítrop negre, la duració del desenvolupament de cada flor dura de tres a cinc dies a temperatura òptima (més de 8°C), mentre que en dura sis a temperatures més baixes (de 2 a 3°C). La saxífraga opositifòlia (Saxifraga oppositifolia) floreix normalment durant tres o quatre dies, però manté les inflorescències durant deu dies a temperatures fredes. Els valors respectius per a altres espècies són d’un a dos dies de floració en condicions normals i cinc dies en cas de fred per a l’astràgal umbel·lat (Astralagus umbellatus), de dos a tres i set per a la dríade puntada (Dryas punctata), i de quatre a cinc i vuit per a alguna genciana (Gentiana algida). Durant la maduració de les llavors es fa palès l’efecte tèrmic de la pubescència dels lòbuls del calze.
El creixement i la reproducció
El ritme de desenvolupament de les plantes és determinat per les condicions climàtiques, i alhora depèn del termini d’aparició dels brots i del seu total desenvolupament. El fet que les gemmes foliars i els botons florals es formin completament a mitjan juliol o al final d’aquell mes representa una adaptació important a les severes condicions climàtiques de la tundra. La maduresa fisiològica dels botons florals es fa palesa quan hi ha una segona florida, que es dóna en condicions climàtiques anormals (una tardor inusualment càlida, per exemple), mentre que la de les gemmes foliars es manifesta amb l’aparició primerenca de noves gemmacions després que la planta hagi sofert danys prou greus produïts pel pasturatge, de tipus mecànic, etc.
Moltes plantes herbàcies perennes reprenen el creixement tan bon punt la suma de temperatures diàries que excedeixen dels 0°C arriba a 20-30°C. Aquest és el cas de la campaneta de fulla rodona (Campanula rotundifolia), dels rovells d’ou (Trollius europaeus i T. apertus), dels epilobis (Epilobium (=Chamaerion) angustifolium i E. latifolium), etc., totes elles espècies molt vistoses. Els borrons creixen o bé a les branques (inclosos els rebrots subterranis horitzontals) o bé als rizomes, i determinen el creixement la respectiva posició i la temperatura de l’aire (en el cas dels arbustos), la de la superfície del tou de molses (en el dels arbustos nans, especialment els prostrats) o la de l’interior d’aquest o, fins i tot, la de la capa superior del sòl (en el de les herbàcies perennes).
Diferents espècies d’un mateix gènere poden requerir, en una mateixa regió, diferents sumes de temperatures per damunt de 0°C. Aquest és el cas dels nabius (Vaccinium); enfront dels valors de 205 a 230°C que requereix el nabiu uliginós o de torbera (V. uliginosum), el nabiu comú (V. myrtillus) en requereix de 230 a 260°C i el nabiu vermell (V. vitis-idaea) entre 310 i 400°C. Aquest darrer no renova les fulles cada any, sinó que, en una mateixa planta, n’hi ha de diferents edats. Malgrat que la longevitat d’una mateta d’aquesta espècie és relativament limitada (9 a 12 anys), el seu sistema vegetatiu pot subsistir diversos decennis i anar-se estenent gradualment a noves àrees; s’han arribat a mesurar rizomes d’1 m a 1,5 m de llarg. Als Urals septentrionals, V. vitis-idaea prepara la floració durant 9 o 14 dies, passen de 5 a 9 dies entre l’aparició de les primeres flors i el moment de màxima florida, i aquesta dura en total de 14 a 20 dies; de 50 a 55 dies més tard maduren els fruits. Si es produeix una onada de fred en l’època de màxima florida, el més probable és que morin totes les flors.
En el cas del nabiu uliginós o de torbera (Vaccinium uliginosum) a la península de Iamal, el creixement anual dels rebrots (de 0,3 a 3 cm) dura de 14 a 17 dies; en cada un d’ells broten de 5 a 9 fulles, que cauen a la tardor. Als hàbitats més humits les fulles i els fruits són més grans, els rebrots més llargs i el període de creixement dura més que als hàbitats secs. El nabiu comú (V. myrtillus), que no resisteix tant el fred com les altres dues espècies del mateix gènere abans esmentades, és una planta curiosa per la seva capacitat de limitar el període anual de rebrotada tant per sobre com per sota de la superfície del sòl. El temps necessari perquè els fruits madurin depèn de les condicions climàtiques de cada any, cosa que pot variar considerablement. Als Urals septentrionals aquest període és determinat per una suma de temperatures positives de 760 a 910°C.
Altres mates prostrades de fulles petites, com l’èmpetrum (Empetrum hermaphroditum), l’èmpetrum negre (E. nigrum) i el nabiu de grua (Vaccinium oxycoccos) tenen una morfologia semblant, però el medi on pot viure aquesta darrera és considerablement més restringit que el dels èmpetrums: viu a les torberes i als boscos oberts inundats de pícees. Tant l’èmpetrum negre com la gerdera de pantà hibernen amb fulles verdes vives. En arribar la primavera, aquestes fulles “velles” inicien la seva activitat fotosintètica així que es fon la neu i, alhora, els borrons comencen a obrir-se i es desenvolupen les fulles noves. En el cas de la gerdera de pantà, les gemmes axil·lars poden romandre en estat vegetatiu un màxim de tres anys; si al final del segon any no s’han pogut desenvolupar, la planta produeix botons florals apicals que, l’any següent, donen lloc a brots fèrtils. Així, la tija principal té brots curts d’entre un i tres anys i quan un mor, en surt un altre. El nabiu de grua floreix relativament tard, la segona desena de juliol, cosa que normalment no permet els seus fruits de completar la maduració fins a la primavera següent. Per això aquests fruits, a la tundra, només es poden menjar a la primavera.