
Entre els cnidaris trobem organismes que viuen fixats al substrat durant tota la seva vida adulta (encara que en alguna fase del seu cicle poden ésser nedadors i planctònics), com ara els coneguts coralls, les gorgònies, les actínies i les anemones. Però també n’hi ha que viuen durant tot el seu cicle com a formes nedadores, típicament planctòniques, com ara les meduses. La de la fotografia, de l’espècie Cotylorhiza tuberculata, arriba a atènyer mig metre de diàmetre, i no és rar veure-la durant la primavera i la tardor prop de la costa, en aigües superficials; els adults neden solitaris i, curiosament, els acompanya un petit eixam de peixos que no es mouen del voltant dels seus tentacles.
Xavier Safont / M. Alba Camprubí
Els cnidaris i els ctenòfors són morfològicament semblants, amb el cos format essencialment per un sac amb tentacles; la cavitat interna, ampla i única, rep el nom genèric de celènteron Aquesta cavitat general permet la respiració dels teixits, la disponibilitat d’aliment a tot el cos, i l’excreció directa dels residus del metabolisme cel·lular. Encara que els celenterats foren considerats durant molt de temps com un grup natural, les diferències que hi ha entre cnidaris i ctenòfors en llur anatomia, histologia i desenvolupament demostren que es tracta de dos llinatges diferents i sense un avantpassat comú i deslligat de la resta dels metazous. Són, doncs, un grau estructural, no pas una entitat filogenètica.
Introducció al coneixement dels cnidaris
Els cnidaris són de simetria radial primària, malgrat que les formes més complexes poden tendir a ésser bilaterals. L’estructura general del cos és molt senzilla i es manté bàsicament uniforme en tot el fílum. L’eficàcia funcional d’aquesta anatomia tan poc sofisticada ha fet possible que els cnidaris siguin, fins i tot en l’actualitat, un dels grups zoològics més abundants dels medis marins: amb més de 9000 espècies, es troben a pràcticament tots els hàbitats. El millor exemple d’aquest èxit ecològic són els esculls de corall de les mars tropicals, que formen un dels ecosistemes més complexos que hom pot trobar al planeta. Tampoc no hem d’oblidar que constitueixen una part important del zooplàncton i que, fins i tot, se’n poden trobar a les aigües continentals.
Morfologia
La paret que envolta l’única cavitat del cos dels cnidaris, anomenada cavitat gastrovascular, és formada per dos epitelis vertaders separats per una tercera capa molt variable però que no constitueix un teixit, encara que pot contenir elements cel·lulars. Aquestes tres capes s’anomenen, de dins a fora, endoderma o gastroderma (amb funcions gàstriques), mesoglea (amb funcions de teixit connectiu) i epidermis. La boca és envoltada per uns tentacles característics, en una o més corones. Els diferents teixits donen lloc a sistemes no centralitzats d’estructura difosa: nerviós, muscular, digestiu i sensorial. D’altra banda, no hi ha sistemes respiratori, circulatori ni excretor, i el reproductor es limita a una sèrie de cèl·lules sexuals agrupades, que fan la funció de gònades. Presenten una certa especialització cel·lular, a conseqüència de la qual es produeix un agrupament i una organització dels diferents tipus de cèl·lules, que dóna lloc a veritables teixits. Així, per exemple, el teixit epitelial dels cnidaris és complex: a part de formar capes i cutícules i de contenir cèl·lules sensorials i glandulars, és el responsable de les funcions musculars, i, fins i tot, pot contribuir a la conducció d’impulsos nerviosos.
Una característica dels cnidaris és la possessió dels cnidocists, d’atac o defensa, continguts en una càpsula que inclou un llarg filament buit, enrotllat, acabat en un arpó (cnidòcil), connectat a un saquet de líquid tòxic que s’injecta a través d’aquest quan un canvi de pressió externa fa saltar una diminuta tapadora i el cnidocist es dispara. Les cèl·lules formadores dels cnidocists, anomenades cnidoblasts, quan contenen l’orgànul totalment format reben el nom de cnidòcits. Aquestes cèl·lules són especialment abundants als tentacles, tot i que es poden trobar per tota la superfície del cos d’una manera més ocasional.
Són dues les formes principals en què es poden trobar els cnidaris: el pòlip, o fase polipoide, i la medusa, o fase medusoide. Poden existir nombroses especialitzacions d’una i altra, però, en general, hom pot dir que el pòlip és la fase sèssil i la medusa la fase pelàgica. Una i altra poden constituir diferents fases del cicle vital d’una mateixa espècie, tot i que una d’ambdues es pot veure molt reduïda o, fins i tot, pot arribar a desaparèixer.
La fase de pòlip

En la fase de pòlip, els cnidaris són generalment diminuts i de morfologia senzilla: no són més que una mena de sac, amb una obertura envoltada de tentacles. Però els pòlips viuen reunits formant colònies que, gràcies als esquelets o estolons, adquireixen formes característiques. En el cas de la fotografia, els pòlips de Clava multicornis formen una colònia damunt una base laminar formada per un entreteniment de tubs corns. Espècies com aquesta viuen a poca fondària (menys de 10 m), en llocs exposats a l’onatge, i tenen una vida bentònica bastant efímera, ja que no dura més d’unes setmanes.
Josep M. Gili / Antoni Garcia
Pel que fa als pòlips, el seu cos presenta normalment una forma cilíndrica i més o menys allargada, anomenada tronc o columna. L’extrem oral se situa al bell mig d’un disc oral en els antozous, o bé a l’àpex d’un allargament o hipostoma en els hidrozous. En els hidrozous, la boca és rodona i la simetria radial que caracteritza el grup és, doncs, estricta. Contràriament, en els antozous la boca s’allarga i presenta un o dos solcs a un o ambdós dels seus extrems; per tant, la simetria radial es converteix en radiobilateral, encara que tant l’aspecte extern com el comportament continuen essent els d’un animal típicament radiat. La boca dels pòlips és envoltada de tentacles; aquests poden trobar-se repartits per tot el cos, o bé, com és més normal, agrupats en una o dues corones. Segons els diferents grups o les diferents espècies, els tentacles poden ésser buits o sòlids per dins, però, en tot cas, sempre es troben fortament armats de cnidocists. En alguns casos, rars, els tentacles han desaparegut, com per exemple en colònies polimorfiques, que tenen pòlips molt modificats per a dur a terme activitats molt concretes, o bé en espècies molts petites o paràsites. El pol aboral dels pòlips és molt variable i depèn, en gran part, de si són solitaris o colonials. En el primer cas, la base pot formar un disc podial adherent, que es fixa al substrat, o pot descansar sobre una base esquelètica, secretada pel mateix pòlip. Si es tracta de formes colonials, la base del pòlip és contínua i forma un tub o estoló que manté l’estructura trilaminar del pòlip i que comunica tots els membres de la colònia: és l’anomenat sistema estolonífer.
L’estructura bàsica dels pòlips és la següent: una epidermis externa, un gastroderma intern i la mesoglea que separa ambdós epitelis. La capa de mesoglea no acostuma a ésser gaire gruixuda; de fet, en els hidrozous no és més que una fina làmina desproveïda d’elements cel·lulars, que rep el nom de mesolamel·la. En els antozous, però, en lloc de la mesoglea hi ha un veritable mesènquima, amb les mateixes funcions que el teixit connectiu fibrós. La cavitat gastrovascular pot ésser des d’una cambra senzilla (en els hidrozous) fins a cambres complexes proveïdes de divisions internes longitudinals, separades per septes o envans. En la fase polipoide dels escifozous, la cavitat és dividida per quatre septes; en els antozous, els septes s’estenen des de la paret del cos fins a la faringe d’origen endodèrmic, que es prolonga des de la boca fins a l’interior de la cavitat gastrovascular. La compartimentació interna d’aquesta cavitat incrementa notablement la superfície d’intercanvi sense que augmentin les dimensions de l’animal.
La fase de medusa
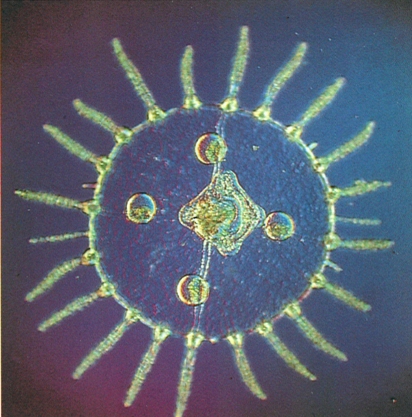
La fase de medusa dels cnidaris és, morfològicament, bastant uniforme: normalment és un cos en forma de campana tota envoltada de tentacles. La fotografia correspon a una hidromesusa vista des de sota, cosa que permet observar el marge de la campana, l’estómac, en posició central, i les quatre gònades penjant. Pertany al gènere Obelia, les diferents espècies del qual adopten un aspecte quasi idèntic durant la fase de medusa, de manera que hom les ha de reconèixer en la fase de pòlip. Les meduses d’aquest gènere són molt comunes en aigües costaneres i poc profundes, on acostumen a formar grans eixams.
Claude Carré
Externament, la fase medusoide és molt diferent de la polipoide. Això respon tant a la posició funcional de l’animal com a un increment espectacular de la capa de mesoglea, que arriba a constituir pràcticament la totalitat del volum de l’animal. Per això, les meduses s’assemblen a una mena de bol gelatinós en forma de paraigua o ombrel·la, que és precisament com s’anomena el seu cos. De la part còncava o oral se’n diu subombrel·la, i de la part aboral i convexa, exombrel·la. La boca se situa al bell mig de la subombrel·la, al final d’un tub més o menys llarg anomenat manubri, que la connecta amb l’estómac o cavitat gàstrica, homòloga a la cavitat gastrovascular dels pòlips. L’estómac, que és senzill en les hidromeduses, es divideix en les escifomeduses en quatre cavitats separades per septes. De l’estómac surten quatre canals radials (o un nombre múltiple de quatre), que determinen els radis principals o perradis; els sectors que queden entre ells són els interradis. Els radis que divideixen en dues parts iguals cadascun dels interradis s’anomenen adradis. Aquesta terminologia serveix per a situar correctament les diverses estructures que es troben al marge ombrel·lar (tentacles, òrgans de l’equilibri i fotoceptors). La capa externa, que envolta l’exombrel·la, la subombrel·la i el manubri, és epidèrmica. L’endoderma o gastroderma recobreix la part interna del manubri, els canals i l’estómac. Entre ambdós epitelis hi ha una capa de mesoglea (acel·lular en les hidromeduses, cel·lular en la resta). En les hidromeduses, una banda circular, el vel, surt del marge inferior de la subombrel·la i divideix l’espai subombrel·lar en dues parts; de funció impulsora, presenta una potent banda muscular. Les meduses amb vel reben el nom de craspedotes, i les que no en tenen, d’acraspedotes.
La medusa és la fase nedadora del grup però també és la que es reprodueix sexualment. Una d’aquestes dues funcions pot convertirse en la dominant, i llavors pot haver-hi un cert polimorfisme. Per exemple, en els sifonòfors hi ha meduses que perden els tentacles i la capacitat reproductora per a esdevenir campanes natatòries o nectòfors, i d’altres que es converteixen en suradors o pneumatòfors. Contràriament, en molts hidroïdeus les mesuses esdevenen simples recipients de les cèl·lules sexuals.
L’especialització cel·lular i els teixits
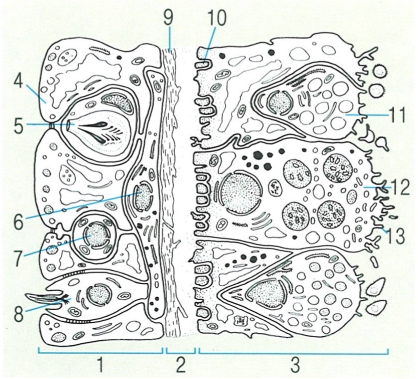
Secció transversal de la paret corporal d’una Hydra, on s’observa la composició cel·lular bàsica dels cnidaris. 1 Epidermis o ectoderma, 2 mesoglea, 3 gastroderma o endoderma, 4 cèl·lula epiteliomuscular (epidermis), 5 cnidoblast, 6 neurona, 7 cèl·lula intersticial, 8 cèl·lula neurosensorial, 9 fibra muscular longitudinal, 10 fibra muscular circular, 11 cèl·lula ganglionar, 12 cèl·lula epiteliomuscular (gastroderma), 13 cilis.
Jordi Corbera, a partir de fonts diverses
L’epidermis dels cnidaris és formada bàsicament per cèl·lules epitelials o cèl·lules de suport, normalment cuboides o columnars, però també aplanades (en l’exombrel·la de les meduses) o extremament primes i altes (en la columna de les anemones). Aquestes cèl·lules poden tenir un o més miomes (fibril·les contràctils) a la base i aleshores reben el nom de cèl·lules epiteliomusculars o mioepitelials; això és particularment evident en els hidrozous. En els escifozous i els antozous, el sistema muscular és format per cèl·lules que han perdut la funció epitelial primitiva; de fet, hom pot observar tota la gradació que va des de les cèl·lules epitelials pures fins a aquelles en què la funció muscular és pràcticament la única. Entre aquests dos tipus extrems hi ha les cèl·lules epiteliomusculars típiques, amb una (les més nombroses) o més fibril·les contràctils, que es poden fins i tot ramificar o anastomosar entre elles. També hi ha cèl·lules molt aplanades, en forma d’escata, que contenen diversos mionemes disposats en una sola direcció o bé perpendicularment els uns als altres.
El sistema muscular és força uniforme. En les formes polipoides dels hidrozous, la musculatura forma dues capes, una d’externa, amb les fibril·les disposades longitudinalment, i una d’interna, circular, a la base del gastroderma.
En contraure’s la musculatura externa provoca la contracció del pòlip, mentre que la interna permet l’elongació de l’animal. Contraccions parcials d’una i altra poden provocar la torsió del pòlip, ajudar-lo a la deglució de les preses i, en el cas de les hidres d’aigua dolça i d’algunes anemones, poden ésser responsables de la locomoció de l’animal. En les formes medusoides dels hidrozous, la musculatura del gastroderma és quasi absent, com també l’externa, que es limita a unes fibres longitudinals als tentacles i al manubri, i es disposa en feixos radials a la subombrel·la. La musculatura principal de les meduses és formada per cèl·lules musculars independents que formen bandes musculars extraepidèrmiques, disposades en cercle al voltant del vel (en les craspedotes) o de la subombrel·la. En els antozous, la musculatura epidèrmica és també molt reduïda: es limita als tentacles, al disc oral i al disc del peu. La musculatura gastrodèrmica forma una malla ben desenvolupada, amb bandes longitudinals. Tant en els antozous com en els escifozous, les cèl·lules musculars poden formar feixos i endinsar-se en la mesoglea.

La gran capacitat de retracció dels tentacles és patent en aquestes fotografies de l’antomedusa Neoturris pileata, abans (a la dreta) i després (a l’esquerra) d’alimentar-se; l’epidermis dels tentacles i del voltant de la zona de la boca té una gran quantitat de cnidòcits, que faciliten la captura de les preses. Aquesta espècie, que pot arribar a fer més de 3 cm d’envergadura, durant la primavera és comuna a les costes catalanes, sempre en aigües costaneres i sornes. Es desplaça, com moltes altres antomeduses, fent moviments ondulatorís característics. Noteu, en les fotografies, la massa rogenca central, que correspon a l’estómac i les gònades, i la nitidesa de la campana.
Claude Carré
Els cnidaris són els únics animals d’organització més senzilla que presenten un sistema nerviós organitzat, format per cèl·lules nervioses bipolars o multipolars (cèl·lules ganglionars). Generalment, és constituït per dos plexes o xarxes nervioses disposats just per sota l’epíteli, tant en l’epidermis com en el gastroderma (generalment més ben desenvolupat en la primera), poc connectats entre ells; però en la mesoglea del pennatulaci Veretillum cynomorium s’han trobat cèl·lules nervioses del que podria ésser un tercer plexe. En les hidromeduses, el plexe nerviós és limitat a la superfície de la subombrel·la i es connecta amb dos anells nerviosos que voregen l’ombrel·la pel marge; el superior innerva els òrgans sensorials i l’inferior, l’anell muscular del vel. En els escifozous, en lloc d’aquests anells hi ha ganglis nerviosos a la vora de cada òrgan sensorial o ropàlia, que connecten amb el plexe subombrel·lar. Pel que fa als pòlips, sembla que les hidres tenen un anell nerviós a l’hipostoma, i les anemones i els coralls, plexes nerviosos localment engruixits. La conducció dels impulsos nerviosos és simètrica, és a dir, pot anar en dos sentits, i la sinapsi no és terminal sinó que es produeix sempre que es troben dues expansions de les cèl·lules ganglionars; fins i tot pot haver-hi més d’una sinapsi entre dues cèl·lules nervioses, i també és normal que una neurona tingui més d’un efector. Els sistemes conductors dels impulsos nerviosos han estat objecte d’una recerca activa. S’ha demostrat l’existència de fins a tres sistemes conductors independents: un de ràpid (neurones grosses i bipolars), un de lent (neurones petites i multipolars) i, a més, en els hidrozous, una conducció nerviosa en absència de qualsevol element pròpiament nerviós (sembla que conduït per cèl·lules epitelials). A més, hi ha, en els tentacles i la regió de l’hipostoma, cèl·lules neurosecretores, cèl·lules ganglionars modificades, amb nombroses vesícules plenes d’una substància peptídica. En l’epidermis també hi són comunes les cèl·lules glandulars secretores de mucus, especialment prop de la boca i a la faringe dels antozous. Els pòlips solen tenir cèl·lules glandulomusculars al disc del peu, secretores d’un mucus que pot arribar a ésser molt abundant en espècies de coralls, i que constitueix una font important de matèria orgànica per a nombrosos animals dels esculls coral·lins. Entremig de les cèl·lules epitelials, hi ha cèl·lules sensorials, de funcions diverses (mecanoceptores, quimioceptores o fotoceptores, que poden ésser disperses per l’epidermis, o bé, com en el cas de les meduses, arribar a formar grups densos i constituir òrgans fotoceptors o de l’equilibri (estatocists).
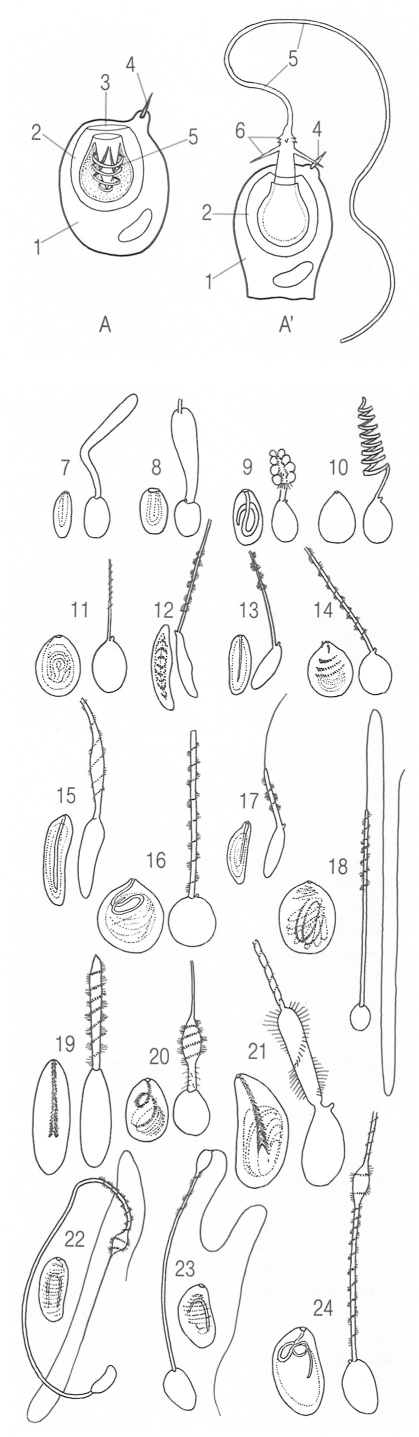
Cnidòcits o cnidoblasts dels cnidaris. A dalt, esquema general d’un cnidòcit. amb el nematocist carregat (A) i descarregat (A’). S’hi ha indicat: 1 cnidòcit o cnidoblast, 2 cnidocist o nematocist, 3 opérele, 4 cnidòcil, 5 filament i 6 espines. A baix, tipus de nematocists: 7 ropalonema anacròfer de sifonófor i 8 acròfer de sifonòfor; 9 desmonema d’hidromedusa i 10 d’hidra, 11 haplonema àtric d’hidropòlip, 12 basitric d’hidropòlip, 13 holòtric d’hidromedusa i 14 holòtric d’hidra; 15 anisoriza homòtric de sifonòfor i 16 heteròtric d’hidromedusa: 17 i 18 microbàsic mastigòfor d’hidromedusa, 19 microbàsic amastigòfor d’actínia; 20 microbàsic euritel homòtric d’hidromedusa i 21 heteròtric d’hidropòlip; 22 macrobàsic euritel telòtric d’hidromedusa, 23 meròtric d’hidromedusa i 24 holòtric d’hidromedusa.
Jordi Corbera, a partir de fonts diverses
A l’epidermis dels tentacles i als voltants de la regió oral són especialment abundants unes cèl·lules especialitzades, els cnidòcits, portadores dels cnidocists o nematocists, orgànuls propis i característics dels cnidaris. Aquests orgànuls són sorprenentment grans, ja que atenyen més de 100 µm. Els cnidòcits són cèl·lules arrodonides, que presenten el nucli en posició basal, el nematocist en posició apical, i, al seu costat, un flagel molt modificat, el cnidòcil, que fa de receptor dels estímuls que determinen la descàrrega. Quan els cnidòcits són joves i el nematocist és encara en formació, aquestes cèl·lules s’anomenen cnidoblasts (tot i que aquest nom s’empra sovint per a designar el cnidòcit madur). El nematocist consta d’una càpsula, constituïda per una paret doble que s’invagina en l’àpex i dóna lloc a un filament que roman enrotllat al seu interior. En el punt on s’inicia aquest filament, la càpsula presenta una tapadora, l’opercle. Quan el nematocist és estimulat, l’opercle s’obre i es produeix l’eversió del filament (moltes vegades eriçat d’espines), que es clava en la presa o el depredador i injecta el líquid verinós contingut en la càpsula. En molts casos, els nematocists no serveixen per a injectar verí sinó per a enganxar o envoltar les preses o les partícules alimentàries, o per ajudar a la fixació del pòlip al substrat, com és el cas dels espirocists dels antozous. El contingut de la càpsula dels nematocists és format per una barreja de substàncies que produeixen una acció hemolítica o miolítica, principalment; hi ha espècies en les quals és particularment virulenta: a les mars australianes, Chironex fleckeri causa anualment més víctimes humanes que no pas els tan temuts taurons.
A l’epidermis hi ha, a més, unes cèl·lules anomenades intersticials, que no tenen una funció evident; a partir d’aquestes es formen tant els cnidoblasts com la resta de cèl·lules, que es van malmetent amb el temps; també poden donar lloc a noves neurones i, en alguns casos, cèl·lules sexuals.
El gastroderma té una estructura molt semblant a la l’epidermis. La majoria de les seves cèl·lules, mioepitelials, presenten alhora una funció digestiva, i per això reben el nom de cèl·lules nutrício-musculars; sovint presenten un gran nombre de vacúols i un o dos flagels. També hi ha cèl·lules glandulars, de dos tipus bàsics: les mucoses, secretores de mucus, situades vora la boca, a la faringe, i les granulars, secretores d’enzims per a la digestió extracel·lular. Les cèl·lules sensorials que hi ha al gastroderma són similars a les de l’epidermis, però molts menys abundants; pràcticament no hi ha cnidòcits, però en els escifozous i els antozous n’hi ha denses agrupacions que formen part dels filaments gàstries, els filaments septals i els aconcis. Finalment, hi ha també cèl·lules indiferenciades. En alguns coralls i anemones és comú de trobar-hi protists vegetals simbionts (zooclorelles o, més comunament, zooxantel·les). Hom ha pensat que aquesta simbiosi és la responsable de l’existència dels esculls de corall, ja que permet d’accelerar la deposició de carbonat càlcic en els coralls que contenen aquestes algues.
| Tipus de nematocists en els cnidaris | ||||||
|---|---|---|---|---|---|---|
| ASTOMOCNIDI: filament tancat a l’extrem. | ||||||
| Ropalonema: filament acabat en forma de maça. | ||||||
| Espironema: filament uniforme i enrotllat en espiral. | ||||||
| Desmonema: filament estret a la base i sense formar un dard ben definit. | ||||||
| Euritela: filament ample a la base i amb un dard ben definit. | ||||||
| Espirotel microbàsic: filament curt i enrotllat. | ||||||
| Espirotel macrobàsic: filament molt curt i no enrotllat. | ||||||
| ESTOMOCNIDI: filament obert a l’extrem. | ||||||
| Haplonema: filament que no forma un dard definit. | ||||||
| Isorize: filament de diàmetre uniforme. | ||||||
| Àtric: filament sense espines. | ||||||
| Basítric: filament amb espines només a la base. | ||||||
| Holòtric: filament amb espines per tot arreu. | ||||||
| Anisorize: filament dilatat a la base. | ||||||
| Àtríc: filament sense espines. | ||||||
| Homòtric: filament amb totes les espines iguals. | ||||||
| Heteròtric: filament amb les espines de la base més grans que les altres. | ||||||
| Heteronema: filament amb un dard ben definit. | ||||||
| Rabdoïdeu: filament de diàmetre uniforme. | ||||||
| Mastigòfor: dard estret envoltat d’una filera espiral de petites espines. És microbàsic quan la llargada del filament és menys de 3 vegades la llargada de la càpsula ( microbàsic b mastigòfor quan el contorn entre el dard i el filament és arrodonit; microbàsic p mastigòfor quan el contorn és triangular). És macrobàsic quan la llargada del filament és 4 vegades superior a la de la càpsula. | ||||||
| Amastigòfor: sense filament a l’extrem del dard. És microbàsic quan la llargada del dard és inferior a 3 vegades a llargada de la càpsula. És macrobàsic quan el dard és 4 vegades més llarg que la càpsula. | ||||||
| Ropaloïdeu: dard de diàmetre irregular. | ||||||
| Euritel: dard engrossit a l’extrem i espines petites. És microbàsic quan el dard és tres vegades més curt que la càpsula i homòtric quan té les espines iguals i heteròtríc quan les té desiguals. És macrobàsic quan el dard és quatre vegades més llarg que la càpsula, i telòtric quan porta les espines distáls i holòtric quan les porta repartides per tot. | ||||||
| Estenotel: dard engrossit a la base, amb espines grans. | ||||||
| Biropaloïdeu: dard de diàmetre irregular a diferents zones. | ||||||
| ESPIROCIST: càpsula de paret molt fina i dard enrotllat en una espiral molt compacta dins de la càpsula; el dard és de diàmetre uniforme i no presenta cap mena d’espines | ||||||
Biologia i ecologia
La reproducció i el desenvolupament
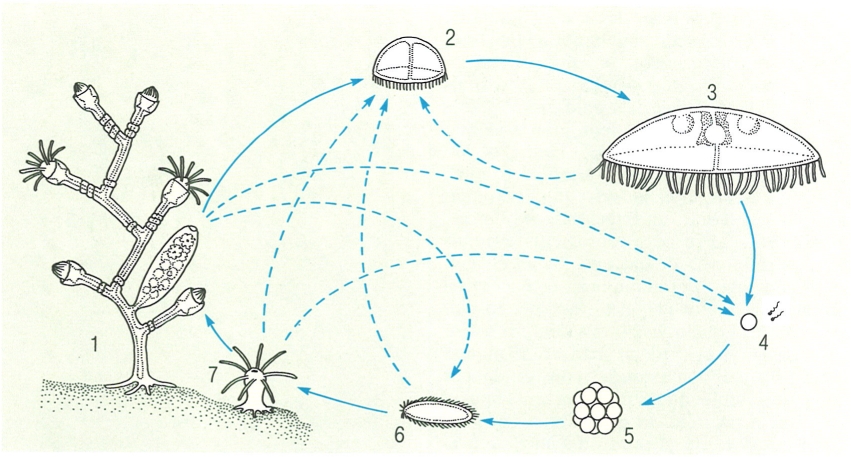
Cicle biològic dels hidrozous. 1 Colònia de pòlips, 2 hidromedusa jove, 3 hidromedusa adulta (fèrtil), 4 gàmetes, 5 embrió, 6 larva plànula, 7 pòlip primari. La línia contínua indica el procés més habitual, mentre que la discontínua indica les diferents variacions que el cicle pot seguir en les diferents espècies i segons les condicions ambientals. S’han indicat les fases sexuals en blanc i les asexuals en groc.
Jordi Corbera, a partir de fonts diverses
Una de les característiques més importants dels processos de reproducció en els cnidaris és la seva gran capacitat de reversibilitat morfogenètica, ja que poden transformar cèl·lules i teixits en les formes més senzilles que els van originar. D’una manera general, els cnidaris presenten reproducció asexual i sexual, i mentre que els medusozous poden presentar una fase de medusa i una altra de pòlip al llarg del seu cicle biològic, els antozous mai no passen per l’estat de medusa. Això porta a un fenomen peculiar del cicle de vida dels cnidaris, anomenat alternança de generacions, que palesa la diversitat de formes de reproducció del grup. Bastants cicles de vida alternen els dos tipus de reproducció i tenen formes reproductores diferents.
La reproducció sexual es duu a terme, en general, mitjançant les formes medusoides. Els ous i els espermatozoides produïts per les meduses formen un zigot que dóna lloc a una plànula, i aquesta a un pòlip. Els pòlips es reprodueixen asexualment mitjançant la formació de gemmes. Al llarg de tot aquest cicle, els organismes són sempre diploides, i les formes sexuals, medusoides, es formen sempre a partir d’un conjunt de cèl·lules, mai d’una única cèl·lula. D’aquest esquema bàsic hi ha dues variacions importants. D’una banda, es donen processos de reproducció asexual diferents de la simple formació de meduses per gemmació; molts pòlips i meduses produeixen per gemmació altres individus similars, i en força casos no se separen, sinó que formen colònies (hidropòlips i sifonòfors). D’altra banda, pot ésser que algun dels dos estats, el de medusa o el de pòlip, no existeixi en tot el cicle de vida, o bé que es doni en una forma reduïda o gairebé aberrant.
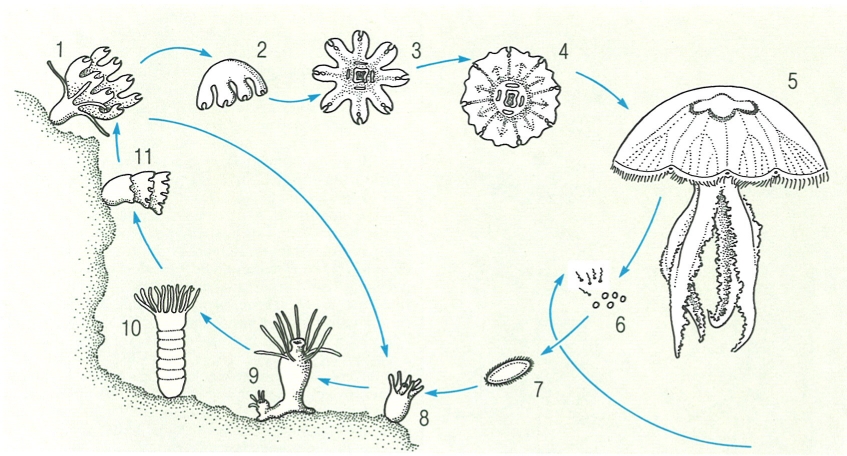
Cicle de vida de l’escifomedusa Aurelia aurita. 1, 2 Efírula, 3 èfira, 4 medusa jove, 5 medusa adulta (fèrtil), 6 gàmetes, 7 larva plànula, 8 fase de pòlip o escifistoma, 9, 10, 11 estrobilació. S’han indicat les fases sexuals en blanc i les asexuals en groc.
Jordi Corbera, a partir de fonts diverses
Pel que fa a la reproducció asexual, la més corrent és la gemmació, per la qual una transformació histològica local dóna lloc a una aglomeració de cèl·lules i a un engruiximent de teixits que, després, per invaginació, formarà un nou pòlip o medusa. Un altre mecanisme, menys important, és la fissió transversal, com en el cas de les actínies o de l’estrobilació dels escifopòlips; la fissió longitudinal, encara que també es pot produir, és rara en els cnidaris. Un sistema bastant estès entre les actínies i els zoantaris és la fragmentació de la colònia o del pòlip i la regeneració de cada tros fins a formar un individu complet, com passa amb les hidres d’aigua dolça. Finalment, la formació d’estructures de resistència és poc estesa entre els cnidaris, encara que formes com els estolons de resistència són comunes en algunes espècies d’hidropòlips. No es pot dir que hi hagi dimorfisme sexual somàtic, però en molts casos els dos sexes es poden reconèixer quan les gònades són madures, o quan s’estan desenvolupant; de totes maneres, en algunes hidromeduses es poden veure petits canvis morfològics, que corresponen a sexes diferents. En general, els hidrozous i els escifozous són dioics; els antozous, en canvi, no és rar que siguin monoics, amb un clar hermafroditisme funcional.
Les cèl·lulés germinals i les accessòries per al procés de la reproducció s’originen a partir de cèl·lules intersticials, pràcticament les mateixes que donen lloc a altres cèl·lules especialitzades (d’origen endodèrmic en els escifozous i els antozous, i majoritàriament ectodèrmic en els hidrozous); bastant grosses i amb nuclis ben visibles, es troben àmpliament repartides per tot el cos i es desplacen per formar agrupacions de tipus gonadal. La gametogènesi és força similar a la de la majoria dels invertebrats. Hi ha cicles gametogènics circadiaris o estacionals, i només unes quantes espècies alliberen cèl·lules sexuals al llarg de tot l’any. En les possibilitats i la periodicitat de la reproducció sexual intervenen factors associats a l’alimentació, el creixement, l’edat i la temperatura.
El desenvolupament embrionari està d’acord amb la senzillesa de la seva estructura i la seva gametogènesi. En els hidrozous, la segmentació de l’òvul fecundat tendeix a ésser total. Les dues primeres segmentacions són perpendiculars, la tercera és equatorial; al final, són de tipus radial, i s’arriben a formar fins a 64 blastòmers. L’adhesió parcial o total entre blastòmers dóna lloc, en molts casos, a una segmentació irregular (com en els sifonòfors). Els embrions de molts cnidaris presenten un tipus de segmentació asimètrica peculiar, que comença per un pol i va avançant progressivament cap a l’altre. La blástula pot ésser formada per un paquet compacte de cèl·lules (estereoblàstula) o per una mena d’embolcall extern que cobreix una massa interna (celoblàstulà). La gastrulació és generalment multipolar, amb diferents processos d’ingressió cel·lular (unipolar o multipolar, una delaminació multipolar o fins i tot una barreja de totes tres). En els escifozous i els antozous, a diferència dels hidrozous, la segmentació és més regular, encara que la diferència principal és que en aquests darrers hi ha una quantitat més gran de vitel en els embrions.
La formació de la larva s’inicia moltes vegades després de la migració de cèl·lules de l’interior cap a la perifèria. Les larves de gairebé tots els cnidaris són planctòniques i del tipus plànula. En aquesta, formada per un endoderma, un ectoderma i, moltes vegades, una cavitat gastrovascular interna, ja hi ha representats tots els tipus de cèl·lules del grup. En canvi, en els sifonòfors i les meduses traquilines hi ha uns estadis larvals intermedis procedents de la metamorfosi de la larva. En alguns hidropòlips gimnoblàstics o atècats es desenvolupa l’anomenada larva actínula, queté vuit tentacles. En alguns antozous es pot veure una postlarva ciliada, coneguda amb el nom de zoantel·la o ceriàntula, i en els escifozous es presenta l’escifistoma, similar a l’estadi de pòlip dels hidrozous.
El creixement
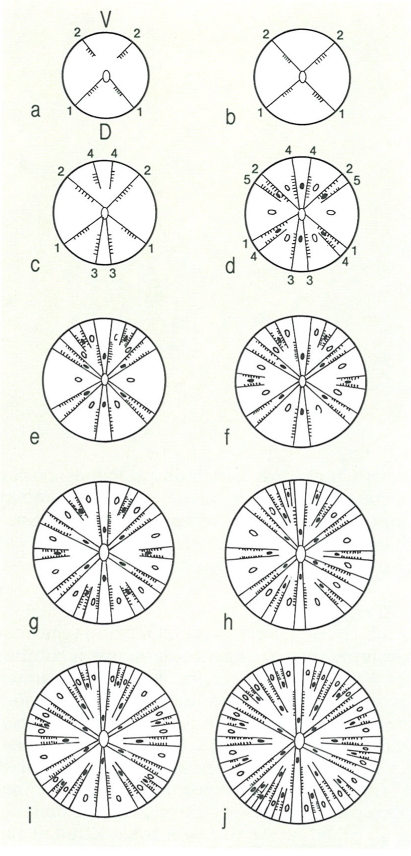
Esquema del desenvolupament de l’esquelet en els antozous madreporaris al llarg de les fases larval (a-d) juvenil (e-h) i adulta (i-j). Els números indiquen l’ordre d’aparició dels mesenteris. Els cercles representen la base dels tentacles (endotentacles en negre, exotentacles en blanc); V ventral, D dorsal.
Jordi Corbera, a partir de fonts diverses
En la major part dels cnidaris, especialment els colonials, el creixement està íntimament lligat a la reproducció asexual: reproducció i creixement s’ajunten en el moment que, per gemmació, comença a formar-se una colònia. La gemmació representa una transformació histològica i morfològica dels teixits que implica sobretot canvis d’estructura i de composició cel·lular. Aquest procés s’adequa al següent esquema bàsic: s’eixamplen un o dos epitelis en un lloc determinat; augmenta el nombre de cèl·lules ectodèrmiques; el teixit es comença a evaginar; l’endoderma forma quatre bandes que representen la simetria radial posterior; creix en forma de papil·la i es defineixen els caràcters morfològics primaris (els tentacles, etc.); finalment, el pòlip queda diferenciat, però resta unit al progenitor per l’estoló o per una connexió de mesoglea. Aquest procés mostra variacions importants, segons els grups i fins i tot en una mateixa colònia. En general les colònies es formen per gemmació i creixement tissular, sense que se’n separin els pòlips nous, que normalment són iguals (excepte en els sifonòfors).
Les taxes de creixement varien molt d’un grup a l’altre. Els hidropòlips tenen les taxes més altes i el creixement més ràpid, ja que poden doblar la seva biomassa en uns 2 dies per terme mitjà, 7 o 8 dies en les colònies més velles. Amb més temperatura i més aliment, la taxa de creixement augmenta, però no ho fa exponencialment, ja que els individus petits creixen molt més ràpid que els grans. L’eficiència neta en el creixement pot arribar a ésser molt gran en les hidres (fins un 80%), en els hidropòlips (fins un 40%) i no tant en les hidromeduses (fins un 25%). També es pot considerar que aquestes últimes presenten taxes de creixement ràpides, ja que poden do blar la seva biomassa en uns 15 dies. Entre els antozous, els que creixen més ràpid són els coralls, que, malgrat que tenen esquelets calcaris, poden doblar llur biomassa en menys de tres anys; en canvi, alguns alcionaris han de passar uns 12 o 15 anys per adquirir les dimensions de l’adult, bé que els dos primers anys ja assoleixen el 10% de la seva biomassa; una gorgònia d’1 m d’alçada viu uns 50 anys de mitjana, mentre que una colònia de corall vermell d’uns 20 cm creix com a màxim 2 mm per ramificació cada any (un 5% de la seva biomassa). Els organismes no colonials, com les actínies, poden doblar la biomassa en un, dos o tres anys, i poden assolir les dimensions de l’adult en uns 10 anys, encara que atenyin la maduresa sexual molt abans (fins i tot dins els dos primers anys); en canvi, quan es reprodueixen asexualment per bipartició, la biomassa es pot doblar en uns 10 dies. Igual que en els organismes colonials, el creixement no és mai continu i pot variar molt dins la mateixa espècie i fins i tot dins una mateixa població. La disponibilitat d’aliment és el factor primari, però s’hi associen l’hidrodinamisme i la competència per a la captura de preses. A més, es pot assegurar que el creixement s’atura parcialment (en els colonials) o totalment (en els solitaris) quan s’inicia la reproducció sexual. També hi ha molta incidència dels fenòmens ambientals (temperatura), que fan que el creixement sigui més lent o s’aturi momentàniament.
L’alimentació

Els cnidaris bentònics són suspensívors sèssils: és a dir, s’alimenten dels organismes o les partícules suspesos o que corren arrossegats pels corrents. Pera millorar la captació d’aliment, les colònies fan estructures perpendiculars al substrat i actuen com a xarxes capaces de retenir les preses i facilitar la feina de captació d’aliment als pòlips de la colònia. En la colònia de la fotografia, de l’espècie Nemertesía antennina, es veuen les brosses i restes de matèria orgànica que han quedat retingudes entre les seves branques. Aquesta estratègia de caça pot ésser contraproduent en zones de corrents forts i carregats de sediment, ja que la seva acció malmet les branques de les colònies.
Josep M. Gili/Antoni Garcia
En general els cnidaris són carnívors, capturen preses mitjançant tentacles o estructures tentaculars, encara que recentment hom els ha detectat altres fonts d’aliment. Les preses s’enganxen als tentacles gràcies a l’acció dels nematocists i els espirocists. A més dels diferents tipus de tentacles, també presenten altres estructures; per exemple, els representants planctònics tenen tractes ciliats que els permeten de capturar organismes petits o els ajuden a desplaçar-se. Els cnidaris bentònics són considerats organismes majoritàriament suspensívors, que viuen a costa de les preses que passen pel seu voltant o de les que són transportades pel corrent. Així, es pot parlar de filtradors passius, que capturen les preses de diferents maneres: o bé les atrapen amb els tentacles i les transporten cap a la boca, o recullen tot el que els cau al damunt (cadàvers o organismes ferits), o bé retenen els individus que se’ls apropen cercant refugi. S’alimenten de tota mena de preses, petites o grosses, i fins i tot més grosses que ells mateixos.
Les preses habituals de la majoria dels cnidaris són organismes del zooplàncton, i per això, durant la nit, quan la densitat de zooplàncton epibentònic augmenta, cobreixen les necessitats energètiques mínimes. No sembla que hi hagi una selecció gaire acurada de les dimensions o del tipus de presa, però en el cas dels cnidaris sèssils, predominen els petits crustacis i tota mena de larves. Les actínies són els més variables i poden capturar des de petits peixos fins a petites larves d’invertebrat. En canvi, els hidraris, mengen tota mena de presa que passi per la seva boca, tot i que algunes vegades s’han vist diversos individus d’una mateixa colònia compartint una presa grossa. En tots els casos sembla que és l’hidrodinamisme el factor que condiciona la captura, ja que es necessita una velocitat mínima del corrent per a capturar preses; però, al mateix temps, una velocitat massa elevada pot deformar la biomecánica del cnidari i impedir que estigui en condicions de caçar. En les espècies epipelàgiques, el tipus d’alimentació no sembla pas diferent, però la capacitat de moviment fa que, potencialment, puguin capturar més preses. Les hidromeduses i els sifonòfors s’alimenten de petits crustacis, altres cnidaris, ictioplàncton i altres organismes planctònics. Però la necessitat de recerca fa que hagin d’invertir més energia en la captura. Hi ha altres possibles fonts d’aliment, menys conegudes, com ara els bacteris i la matèria orgànica que aquests porten associada, i també detrits orgànics d’origen vegetal; és més discutible el fet que es puguin alimentar de fitoplancton (hom pensa que la cel·lulosa és indigerible pels teixits gastrodèrmics dels cnidaris). Una font important d’aliment és la matèria orgànica dissolta, ja que la superfície externa dels cnidaris té una densitat important de microvellositats; per l’epidermis s’absorbeixen substàncies dissoltes (aminoàcids) tant d’una manera regular (algunes actínies) com en condicions de mancança d’altres recursos alimentaris. L’eficiència de la ingestió d’aliment varia notablement entre les diferents espècies. Sembla molt alta en les hidres, i disminueix en els hidropòlips, les hidromeduses, els sifonòfors, els octocoralls i els hexacoralls; a tall d’exemple es pot dir que una colònia d’hidropòlips pot capturar gairebé l’equivalent de deu preses per pòlip i dia, mentre que les colònies de zoantaris rarament capturen més d’una presa per pòlip i dia. Alguns aminoàcids alliberats pel pòlip en contactar el tentacle amb la presa activen l’obertura de la boca i els mecanismes d’ingestió.
La respiració i l’excreció
Els cnidaris no tenen òrgans o estructures específics per a la respiració. Generalment, i sobretot les espècies marines, viuen en hàbitats on l’oxigen és abundant i són exclusivament aeròbics; l’intercanvi de gasos es fa per simple difusió a través de la superfície corporal, tant l’ectodèrmica com l’endodèrmica. Quan s’esgota l’oxigen del medi o quan en aquest s’acumulen gasos de rebuig, necessiten renovar la pel·lícula d’aigua que hi ha en contacte amb el cos; per això tenen zones de cilis i fan moviments peristàltics i musculars superficials. La majoria d’espècies capten l’oxigen amb l’aigua, que és bombada contínuament cap a l’interior de la cavitat gastrovascular. Les zones on es fa l’intercanvi de gasos són des de simples bandes de cilis en les hidromeduses fins a zones especialitzades i connectades amb el sifonòglif en els hexacoralls; en canvi, en els petits pòlips dels hidraris, la respiració és ectodèrmica.

El madreporari Cladocora caespitosa és el més comú dels pocs cnidaris mediterranis que tenen algues simbionts (solament detectades en alguns madreporaris, actiniaris i hidraris), responsables del color verdós que prenen els seus teixits. Aquesta associació és, en canvi, molt important en les espècies formadores d’esculls de corall, fins al punt que les algues intervenen en el metabolisme del madreporari, ja que li subministren oxigen i aliment i consumeixen substàncies residuals, producte de l’excreció de les cèl·lules de l’animal.
Josep M. Gili/Antoni Garcia
El consum d’oxigen per unitat de pes varia segons les condicions ambientals, que, per això, afavoreixen o limiten l’activitat de l’individu. La temperatura és potser el factor més important, bé que també depèn de l’edat de l’organisme o del seu estat. El que sembla general és que hi ha una relació positiva entre un consum elevat d’oxigen i els organismes més grossos, fins i tot dins una mateixa espècie. Els cnidaris que consumeixen relativament més oxigen són les hidromeduses, i després els hidropòlips: de 2 a 5 vegades més que una actínia, en les mateixes condicions metabòliques. Les actínies són les que tenen els valors relatius de consum més baixos, propers als d’alguns antozous grossos i colonials com les gorgònies i els alcionaris. En les actínies hom pot veure variacions importants en la respiració segons l’activitat i el comportament de l’individu: el consum és similar entre individus de la mateixa espècie a diferents graus d’il·luminació, tant si tenen algues simbionts com no; quan passen gana respiren molt més a poc a poc (un 25% menys), i la taxa de consum és també inferior (un 35% menys) quan estan contretes o es troben fora de l’aigua, en ambients intermareals. La major part de l’oxigen consumit s’inverteix en el catabolisme de les proteïnes (un 75%), mentre que els carbohidrats i els lípids representen valors molt més baixos (un 7% i un 10%, respectivament). Aquesta contribució al balanç metabòlic dels cnidaris, i més concretament de les actínies, s’ha donat quan les condicions aeròbiques (saturació d’oxigen) i l’aliment han estat suficients. A més, en condicions de deficiència d’oxigen, i com a resposta a la anòxia, algunes actínies tenen un cert metabolisme anaeròbic: aproximadament el 25% de l’energia necessària prové d’aquesta font.
Gairebé tots els cnidaris expulsen contínuament per la boca trossos d’organismes o partícules indigeribles. Aquesta és la forma més habitual d’excreció, ja que no tenen òrgans o glàndules especialitzats en aquesta funció. Els residus, especialment els resultants del catabolisme proteic, són llançats a l’exterior directament per les pròpies cèl·lules digestives, o bé cauen a la cavitat gastrovascular, i d’allà surten per bombeig a l’exterior. La major part de l’excreció líquida (més del 80%) es fa en forma d’amoni dissolt, un 5% en forma d’aminoàcids, i la resta com a urea i d’altres productes menys importants. El nitrogen excretat triga un dia o més que l’oxigen consumit a recuperar la concentració que tenia abans de l’alimentació. La quantitat de productes excretats depèn, no solament de la quantitat d’aliment ingerit, sinó també de l’activitat metabòlica de cada organisme; així, el potencial excretor per unitat de temps és superior en els hidraris i inferior en les actínies i els coralls. Paral·lelament, els estats prolongats de fam o de manca d’aliment fan disminuir tant l’excreció com la respiració, a fi d’augmentar la capacitat de supervivència de l’animal.
Els hàbitats
Les comunitats bentòniques

En l’habitat litoral, en els primers metres de fondària, s’instal·len cnidaris capaços de resistir l’embat de les onades, fixats fortament al substrat, com ara Aiptasia mutabilis (a dalt); o bé capaços de suportar èpoques de dessecació més o menys llargues, com Actinia equina (al mig); d’altra banda, els hidraris que hi viuen fan colònies petites, com Sertularella ellisi (a baix), proveïdes d’un pensare engruixit i molt flexible, que oscil·la sense trencar-se en una situació d’agitació continuada.
Xavier Safont / M. Alba Camprubí i Josep M. Gili / Antoni Garcia
L’ambient mediolitoral és poc propici per al desenvolupament dels cnidaris. Les poques espècies que s’hi troben, d’una manera estacional i en indrets molt concrets, són les actínies Actinio equina i A. cari, que mostren una gran capacitat d’adherència al substrat i són capaces de retenir molta aigua, la qual cosa els permet de resistir la dessecació diürna; d’altra banda, la reproducció es dóna en el millor moment d’humectació de cada zona, i com que incuben els embrions, els joves ja neixen desenvolupats. Hi ha pocs hidraris que visquin normalment en aquests ambients; l’hidrozou Sertularella ellisi creix just sota els voladissos litorals o en cubetes grans, on forma colònies de nombrosos petits hidrocaules.
En l’ambient infralitoral, la resposta dels cnidaris a la dominancia de les algues es fa a través de dues grans estratègies: l’ocupació del substrat dur i la colonització dels mateixos tal·lus algals. Per a ocupar el substrat rocós directament, s’han d’adaptar a les condicions hidrodinàmiques. Quan els corrents són molt forts i continuats, només hi creixen algunes espècies que, com Balanophyllia regia, presenten un esquelet ample i dur que s’adhereix molt fort a la roca; altres hidrozous, com Halecium mediterraneum, o Sertularella gaudichaudi, creixen al peu del tal·lus de les algues o d’altres organismes incrustants, on el corrent no és tan fort. Els organismes que hi tenen més èxit són els que formen colònies estoloníferes o incrustants, com els estolonífers Clavularia ochracea i Parerythropodium coralloides. Algunes espècies d’actínies o anemones, com Anemonia sulcata i Aiptasia mutabilis, de cos tou i flexible, s’instal·len i s’enganxen fortament a la roca per a resistir l’impacte del corrent. Pel que fa a l’estratègia consistent a utilitzar les algues com a substrat, cal dir que no totes les algues són igualment colonitzades: l’estructura i la forma del tal·lus (superfícies llises o rugoses, planes o abruptes, etc.) afavoreixen més o menys l’assentament de les larves que inicien la colonització; d’altra banda, moltes algues secreten pel tal·lus substàncies que inhibeixen aquest assentament. El caràcter estacional de moltes algues és un altre factor important per a l’epibiosi, de manera que els cnidaris que viuen damunt les algues han de tenir cicles d’acord amb els cicles d’aquestes. Allà on dominen les algues esciàfiles el grau d’epibiosi és més alt, ja que aquestes algues fan poblacions de tal·lus molt perdurables. La majoria dels hidraris que colonitzen aquest ambient viuen sobre tal·lus algals, però a la costa catalana n’hi ha d’especialment adaptats, com Coryne pusilla, Bougainvillia ramosa, Halecium petrosum (hidraris atècats), Campanularia assymetrica, Clytia hemisphaerica, Orthophyxs crenata, Obelia dichotoma, Dynamena cornicina, Sertularia distans, Antennella secundaris, Kirchenpaueria pinnata, Plumularia setacea i Aglaophenia pluma (hidraris tècats). Es poden colonitzar també substrats animals, però sempre a més fondària, on la dominancia algal disminueix; la tendència més peculiar dels cnidaris, i especialment dels hidraris, ha estat colonitzar altres hidraris, cas molt freqüent en les comunitats infralitorals, per exemple de les espècies Filellum serpens, Hebella parasítica i Halecium tenellum (hidraris tècats).

Els prats de gorgònies, com els de la fotografia, d’Eunicella singularis, són un dels elements més representatius de l’hàbitat circalitoral (per sota dels 10 o 20 m de fondària). Les colònies creixen verticalment sobre el substrat, formant una mena de ventalls que s’orienten en la direcció dels corrents dominants. Hom pensa que la intensitat i el règim de corrents són factors que limiten l’extensió dels prats de gorgònies en els fons marins.
Enric Ballesteros
En l’ambient circalitoral, els cnidaris bentònics, especialment els antozous, assoleixen la màxima representativitat. En l’ambient coral·ligen o en les coves submarines són representats per un gran nombre d’espècies, que constitueixen un percentatge molt elevat de la biomassa total de la comunitat. Aquí l’hidrodinamisme constitueix un vehicle de transport d’aliment i de dispersió de larves. Els antozous d’aquest ambient acostumen a ésser individus voluminosos, amb taxes de renovació baixes, amb reproducció asexual eficaç, capaç de controlar el creixement de les espècies veïnes. En molts casos les comunitats adopten la formació de fàcies, perquè les noves colònies, originades per l’assentament de larves, s’instal·len al voltant d’una colònia primigènia. Cal destacar, entre els octocoralls, les mans de mort (Alcyonium acaule), la gorgònia blanca (Eunicella singularis) i la gorgònia roja (Paramuricea clavata), tots tres de creixement lent i reproducció asexual molt desenvolupada (la reproducció sexual es limita a una sola emissió anual d’un gran nombre de larves o gàmetes). En aquestes comunitats, amb una concentració molt alta d’organismes, són importants factors biològics com la competència i la capacitat de captació d’aliment, més que no pas els factors físics. D’estratègia suspensívora, necessiten desenvolupar estructures i formes adequades per a obtenir gran quantitat de partícules alimentàries; és conegut el cas de les gorgònies, que formen ventalls amb ramificacions orientades perpendicularment al corrent dominant. De fet, es pot dir que la força de l’hidrodinamisme i la talla de l’organisme o la colònia tenen molta relació. Els hidraris s’hi troben àmpliament representats, però les espècies que representen més bé l’estratègia general dels cnidaris són les que formen grans colònies aïllades (per exemple, Eudendrium rameum, Halecium beanii, Synthecium evansi i Aglaophenia kirchenpaueri); d’altres creixen habitualment com a epibionts damunt gorgònies (Sertularella crassicaulis), esponges o altres invertebrats (Campanularia hincksi). Entre els antozous, s’hi troben les anemones que formen fàcies, com Parazoanthus axinellae (zoantari) i el corall vermell (Corallium rubrum, gorgoniaci), a més de les esmentades abans, que contrasten amb les que tendeixen a créixer formant petites agrupacions aïllades, com Epizoanthus arenaceus (zoantari) i Caryophyllia smithi (madreporari). També d’aquest ambient, cal destacar els escleractiniaris, especialment abundants a les coves submarines, com Leptopsammia pruvoti, Polycyathus muellerae, Hoplangia durotrix i Caryophyllia inornata (madreporaris), que poden formar poblacions molt denses.

Les fulles de posidònia (Posidònia oceànica) constitueixen un hàbitat marí peculiar, que ocupen solament organismes especialitzats, d’entre els quals destaquen els hidraris. Sertularia perpusilla, que veiem en la fotografia, formant estolons escassament ramificáts, és una de les espècies més esteses en l’alguer de posidònia, i viu exclusivament en aquest hàbitat. Les seves colònies creixen en les parts basats de les fulles de posidònia, allà on els teixits són nous, i això li permet de prescindir de la reproducció sexual (és a dir, de formar meduses o larves de vida lliure), ja que per estendre’s en té prou fent avançar els estolons al llarg de la fulla o fent estolons propagulars, que es desprenen de la colònia mare i s’instal·len en altres fulles de la mateixa planta.
Josep M. Gili / Antoni Garcia
Els ambients sedimentaris costaners són substrats sorrencs o detrítics, essent la densitat d’organismes que hi creixen al damunt o semienterrats (la major part, cnidaris) bastant baixa. Les possibilitats d’assentament dels cnidaris augmenten amb la fondària (amb menys onades, el sediment és menys remogut) i, en general, la presència d’obstacles aïllats (pedres, etc.) afavoreix la formació de clapes de colonització. En els fons de sorra i detrítics de la costa hi ha cnidaris que han desenvolupat parts basáls inertes de la colònia per a introduir-se al substrat, com les actínies Phymanthus pulcher i Bunodactis verrucosa, el ceriantari Cerianthus membranaceus, o el pennatulaci Veretillum cynomorium. Amb rizomes a la part basal, que els permeten de sustentar-se però bascular damunt el sediment, cal esmentar l’hidroïdeu Halecium halecinum i la gorgònia Lophogorgia sarmentosa, o, amb estructures semblants, els pòlips de Monomyces pigmaea.

Associació de mutualisme entre l’actinia Adamsia carciniopados, proveïda de filaments urticants. i el cranc Pagurus prideauxi. Aquest tipus d’associació permet que espècies sèssils puguin colonitzar fons marins tous, de sorra i fang, on manquen substrats inerts colonitzables.
Josep M. Gili/Antoni Moreno
D’altres s’assenten en substrats mòbils (conquilles d’alguns gasteròpodes transportades per crustacis pagúrids), en una associació moltes vegades de caire específic, com és característic de les actínies Calliactis parasítica i Adamsia carciniopados, que estableixen relacions de mutualisme amb el cranc; un cas similar és el dels zoantaris Epizoanthus mediterraneus i E. paguricola: creixen només sobre conquilles, les quals van diluint a mesura que desenvolupen un carcinoeci que arriba a substituir-les. Sovint s’hi troben espècies forànies de cnidaris, de presència ocasional, ja que en els fons detrítics que es troben prop de substrats rocosos, hi van a parar trossos de rocs o concrecions provinents de comunitats veïnes carregats d’organismes que, de fet, hi sobreviuen poc temps. Als alguers de fanerògames dels fons sorrencs, el gran desenvolupament dels rizomes i les rels confereix una gran estabilitat al substrat i això fa possible la instal·lació d’una comunitat animal rica i variada; entre els cnidaris dominen, a l’alguer de posidònia, els hidraris tècats ( Sertularia perpusilla, Campanularia asymetrica, Aglaophenia arpago i Plumularia obliqua posidoniaé), que creixen exclusivament damunt les fulles; les colònies estoloníferes d’aquestes espècies creixen en sentit contrari al del creixement de les fulles de posidònia, sempre cercant de colonitzar més substrat, i al mateix temps emeten estolons propagulars que els permeten de recolonitzar mates veïnes. Damunt els rizomes creixen cnidaris que prefereixen indrets menys il·luminats, normalment als fons circalitorals.

També en substrats tous, de sorra i fang, trobem pennatulacis que s’enterren parcialment mitjançant el peu estèril de la colònia. L’espècie de la fotografia, Virgularia mirabilis, fa una colònia en forma de ploma, i encara que pot arribara formar prats molt extensos, sovint les colònies es troben solitàries. Els pòlips s’alimenten dels organismes que viuen a la superficie del sediment, d’on són remoguts periòdicament per corrents de fons.
Lluís Dantart
Als fons de la plataforma i del talús continental, el grau i la freqüència de la sedimentació, que no és gradual sinó que depèn del pendent i del règim de corrents de fons locals, són els factors que limiten més la distribució dels cnidaris. Per exemple, en els canyons submarins, que es poden trobar prop de la costa, el fort pendent fa que hi hagi una sedimentació contínua, i això dóna molta inestabilitat al substrat, de manera que encara que el corrent dugui prou aliment, els organismes sèssils no s’hi poden instal·lar; però allà on sobresurten rocs grans, prop de la base del canyó, on la pluja de sediments minva, s’hi desenvolupen denses poblacions de pennatulacis, gorgònies o madreporaris, que donen lloc als coneguts fons de coralls blancs ( Dendrophyllia cornígera o Madrepora oculatà). Els cnidaris se situen, però, majoritàriament en indrets on la deposició és baixa i, per tant, hi ha un sediment fi. El desenvolupament de poblacions quasi monospecífiques enmig de grans extensions desèrtiques de sorra és el model normal de distribució en aquest fons. Són especialment característics els prats de pennatulacis de la zona mitjana superior de la plataforma, les anomenades plomes de mar ( Pennatula phosphorea i Pteroides griseum); al mig de les grans colònies poden créixer cnidaris individuals, com la mà de mort (Alcyonium palmatum), que s’enterra parcialment al substrat, el clavell de mar ( Caryophyllia smithi, madreporari), les actínies Actinauge richardi i Edwardsia claparedii, o la gorgònia Eunicella verrucosa, que creix preferentment sobre pedres petites o mitjanes, aïllades al mig d’àmplies extensions de fang i sorra. Entre les espècies que se situen preferentment damunt d’altres organismes o conquilles cal esmentar Hormathia coronata (actiniari). Hi ha cnidaris típicament colonitzadors d’aquests fons profunds, del talús o de la zona batial, però difícilment (només en llocs molt concrets, on no arriben els corrents de fons locals) arriben a formar-hi grans agrupacions; en generai, les colònies o els individus atenyen dimensions més grans que les espècies de menys fondària, i hi son comunes la gorgònia Isidella elongata, l’antipatari Parantipathes larix, el pennatulaci Funiculina quadrangularis i el madreporari Desmophyllum cristagalli. Als fons de la zona mitjana de la plataforma continental es formen els anomenats fons d’hidraris, amb unes quantes espècies d’hidraris tècats, especialment Nemertesia antennina, Lythocarpia myriophyllum i Schizotrica frutescens, que fan poblacions petites però denses, quasi monospecífiques, separades entre si, que en conjunt poden ocupar grans àrees; al seu damunt, s’hi poden instal·lar altres espècies de cnidaris epibionts.
Les comunitats planctàniques del medi costaner
Prop de la costa, a la regió nerítica, les conditions hidrogràfiques són heterogènies, en comparació amb la regió oceànica o de mar oberta. Aquí, la distribució i el desenvolupament de les poblacions de cnidaris és condicionada per fenòmens meteorològies i hidrogràfics, i per factors biològics, com la durada dels cicles, la capacitat de desplaçament dels organismes i les sèves necessitats tròfiques.
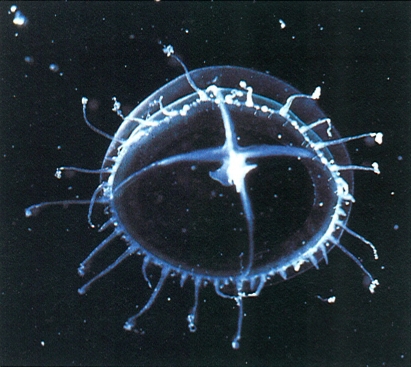
Un bon exemple del plàncton costaner és aquesta leptomedusa, Laodicea undulata, carnívora molt activa que forma eixams prop de la costa catalana, especialment freqüents al final de la primavera. Té una fase de pòlip (de tipus semblant al del gènere Cuspídella), el qual s’instal·la sobre algues i roques a molt poca fondària.
Claude Carré
La presèneia de les espècies en una massa determinada d’aigua depèn de la temperatura i la salinitat de l’aigua. En la majoria de les espècies costaneres cal una temperatura mínima de més de 12°C. perquè es produeixi la reproducció i es puguin formar poblacions d’hidromeduses o de sifonòfors. Per tant, es pot parlar de marcada estacionalitat en molts cnidaris planctônics, exceptuant-ne alguns, com la traquimedusa Rhopalonema velatum, la leptomedusa Eirene viridula la narcomedusa Solmundella bitentaculata, que es troben tot l’any, però amb una densitat moderada. Les espècies més abundants també son regulars tot l’any perô a més tenen una abundància màxima cap a mitj an primavera i al començament de l’estiu. L’espècie més abundant i més freqüent, amb diferèneia, és la tranquimedusa Aglaura hemistoma, que és epipelàgica, seguida de la també traquimedusa Persa incolorata, que neda en una capa no tan superficial d’aigua. La majoria de les meduses que es troben a la nostra costa són epipelàgiques. Entre les d’aigües més profundes, destaquen, per sota els primers 200 m, la narcomedusa Solmaris flavescens i l’escifomedusa Peryphylla peryphylla. Entre les hidromeduses hi ha d’altres espècies que presenten una abundància estacional molt marcada, i per això tenen cicles reproductors més curts, son més oportunistes i poden desenvolupar poblacions denses en espais petits, com per exemple les antomeduses Neoturris pileata, Euphysa aurata, Leuckartiara octona i Laodicea ondulata, la leptomedusa Lizzia blondina, que a la primavera forma grans concentracions prop de la costa. En les antomeduses i les leptomeduses de vida metagènica, el més important és que hi hagi condicions adequades per al desenvolupament de la fase pòlip; així, les denses poblacions d’Obelia o de Clytia hemisphaerica ( leptomeduses) o de Podocoryne carnea (antomedusa) fan suposar que hi ha un fons adequat per al desenvolupament dels pòlips.
L’aparició de grans concentracions de determinades espècies també pot ésser deguda a la influència de factors hidrografics locals; el ritme dels vents costaners que bufen de mar a terra durant el dia i, a l’inrevés, de terra a mar durant la nit, produeix un corrent horitzontal que moltes espècies epipelàgiques, com les meduses, utilitzen per a dispersar-se. Altres són transportades per corrents més forts, com els de la plataforma, originats tant per l’arribada d’aigües al·lòctones com per afloraments locals d’aigües profundes; és el cas, per exemple, de la presència, a la zona nord de la costa catalana, d’espècies al·lòctones (el sifonòfor Lensia meteori) i d’espècies de fondària (L. conoidea) prop de la costa, a la zona de Blanes. Un altre fenomen que explica la distribució dels cnidaris a la costa catalana és la presència de zones frontals, possiblement associades a processos biològics que afecten el conjunt del zooplàncton; el front plataforma-talús, al qual s’associen espècies de distribució més costanera juntament amb d’altres de mar oberta, sembla que dóna lloc a una zona excepcional, d’una gran diversitat, relativament lluny de la costa.

Les colònies del sifonòfor Halistema rubrum són bastant freqüents a la costa catalana a qualsevol època de l’any. Es tracta d’una espècie holoplanctònica, és a dir que roman durant tot el seu cicle de vida en el plàncton. Com altres fisonectes, es troba preferentment en aigües relativament profundes (per sota dels primers 50 m) i oceàniques, si bé tampoc no és rara a la zona nerítica.
Claude Carré
Els sifonòfors segueixen, en general, un model de distribució força semblant al de les meduses, tot i que hi ha diferències importants pel que fa a la composició quantitativa i qualitativa de la població d’una zona determinada. Els sifonòfors són més abundants i més presents perquè tenen més d’una generació anual, una taxa de creixement més elevada i una mobilitat superior. L’espècie més abundant a la costa catalana, la calicòfora Muggiaea atlàntica, presenta dues generacions consecutives, a la primavera i a l’estiu. Altres, també abundants, desenvolupen dues o tres generacions en poc temps (Lensia conoidea) o, al contrari, molt separades en el temps (Chelophyes appendiculata). N’hi ha d’eminentment epipelàgiques, que, de fet, són la majoria de les més abundants, i d’altres que mostren una distribució més profunda (Lensia conoidea). Aquest caràcter epiplanctònic dels sifonòfors de la costa catalana queda reflectit en l’àmplia distribució de moltes de les espècies, com Abulopsis tetragona i Lensia subtilis, que en fases poligàstrica i eudòxica es troben conjuntament en el plàncton. Aquesta associació dels sifonòfors a masses d’aigua de característiques diferents és compartida per moltes espècies; per exemple, la calicòfora Eudoxoides spiralis· prefereix aigües més càlides i es troba, especialment a la tardor i a l’hivern, a la meitat meridional de la costa catalana, on la temperatura de l’aigua és normalment superior a la de la meitat septentrional. Altres, com la també calicòfora Muggiaea atlàntica, s’imposen a la majoria de cnidaris planctònics en aigües més costaneres, de concentració salina més variable; la capacitat d’aquesta espècie d’adaptar-se a canvis de salinitat fa que sigui la més abundant a les zones més productives, relacionades amb aportacions d’aigües continentals. De tota manera, M. kochi pot substituir-la com a espècie més abundant. Com en el cas de les hidromeduses, hom pot parlar d’espècies meroplanctòniques i holoplanctòniques; els sifonòfors són quasi tots holoplanctònics, i els fisonectes són més oceànics. Les grans colònies dels fisonectes Agalma elegans, Halistema rubrum i Nanomia bijuda i de la calicòfora Hippopodius hippopus no es poden considerar rares en aigües de plataforma; sovint ofereixen poca resistència al corrent i per això solen ésser desplaçades cap a zones pròximes al talús.
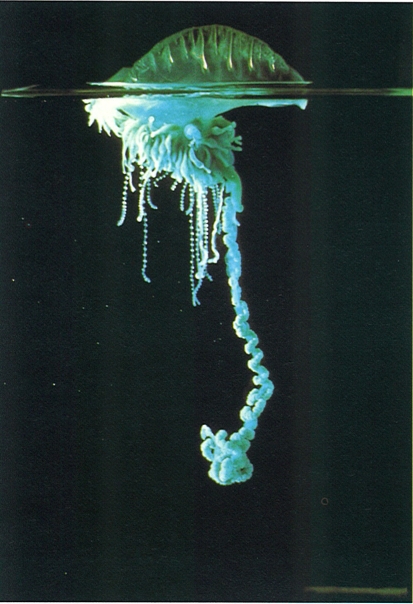
El borm de vela (Physalia physalis) és l’únic representant dels cistonectes a la Mediterrània i és potser l’espècie de cnidarí més perillosa, perquè els seus filaments urticants produeixen ferides d’una certa importància, que deixen cicatrius a la pell difícils de fer desaparèixer. Es tracta d’una espècie que sempre sura a la superfície, gràcies al gran desenvolupament del pneumatòfor (entre 30 i 35 cm). Encara que generalment es troba a alta mar, sovint és arrossegada pels vents locals i llançada, de vegades en grans quantitats, a la platja.
Claude Carré
Les escifomeduses i els sifonòfors cistonectes cal que es tractin a part dels cnidaris costaners; els més coneguts són la caravel·la o borm de vela (Physalia physalis) i l’anomenada barqueta (Velella velella). Aquests organismes formen colònies polimòrfiques d’hidropòlips i hidromeduses, que són arrossegades a la mateixa superfície de la mar i es concentren prop de les platges o en badies tancades, on apareixen formant grans eixams; en la majoria dels casos, l’orientació del seu flotador és un indicador de la seva procedència, ja que es desenvolupa d’acord amb la direcció del vent dominant. Les escifomeduses es desplacen lentament i formen part del nèuston; poden arribar ocasionalment a formar grans eixams, si es donen les condicions tròfiques ideals perquè les seves èfires es desenvolupin bé. Aquestes èfires es troben temporalment en el plàncton; les de l’acàlef luminiscent (Pelagia noctiluca) es veuen tot l’any menys a l’hivern. Nausithoe punctata és habitual en el plàncton de primavera i estiu, mentre que les grans meduses, com el borm radiat (Chrysaora hysoscella), l’acàlef cresp (Cotylorhiza tuberculata) i el borm blau (Rizostoma pulmo) són bastant ocasionals prop de la costa, on es poden veure nedant en solitari.
Les comunitats planctòniques de la mar oberta
La uniformitat ambiental que presenta la mar oberta, amb petits canvis que afecten només les aigües més superficials (excepte quan hi ha afloraments locals d’aigües profundes o sistemes frontals) afavoreix les formes holoplanctòniques, la reproducció asexual (especialment la dels sifonòfors, que inclou una fase poligàstrica i formació d’eudòxies) i les distribucions geogràfiques i batimètriques àmplies. En la mar oberta és on els cnidaris fan els desplaçaments verticals de migració més grans, ja que han de recercar l’aliment en un ambient on les preses són més aviat escasses; en el cas dels sifonòfors, tan sols la formació d’una termoclina estival pot impedir, parcialment, l’arribada d’individus de zones semiprofundes a la superfície durant la nit. Alhora, els seus depredadors són escassos; no obstant això, és corrent que els cnidaris formin grans colònies per augmentar les possibilitats de defensa, com és el cas de molts sifonòfors fisonectes ( Agalma elegans, A. okeni o Nanomia cara), o que tinguin la facultat de fugir amb rapidesa, com algunes calicòfores ( Lensia conoidea i Chelophyes appendiculata).
Moltes de les espècies de la mar oberta tenen una distribució batimètrica molt àmplia, fins al punt que hom hi pot trobar formes eminentment batipelàgiques, com la calicòfora Vogtia. La majoria dels sifonòfors i les meduses es troben en la zona nerítica, però n’hi ha de la mateixa espècie que tenen formes batipelàgiques i epipelàgiques una mica diferents. Entre les hidromeduses més comunes a la Mediterrània occidental destaquen les holoplanctòniques, com la majoria de les narcomeduses i traquimeduses, espècies amb cicles de vida llargs i amb sistemes de reproducció asexual. També les hidromeduses fan migracions verticals de llargues distàncies; l’amplitud dels moviments fa que es trobin formes i espècies mesopelàgiques i batipelàgiques, la qual cosa comporta un grau elevat d’especialització. És també característic dels cnidaris oceànics la formació d’agrupacions o eixams monospecífics, circumscrits a uns límits de fondària estrets; és el cas de força escifomeduses (excepte les coronades, que han de restar prop de la costa perquè passen per una fase de pòlip).
Biogeografia
En una visió global de la fauna de cnidaris de la costa catalana, el component més important és format per les espècies d’origen nordatlàntic (un 44%), seguit, en aquest ordre, per les cosmopolites (un 29%), les endèmiques (un 23%) i, finalment, les de distribució circumtropical (un 9%). Si considerem les espècies planctòniques, els percentatges de cosmopolites i de nord-atlàntiques són molt similars (un 40%), mentre que les endèmiques no superen el 10%. Entre les espècies bentòniques, el percentatge més alt correspon a les d’origen nord-atlàntic (45%), però el d’endèmiques també és alt (30%). Tant en els hidrozous com en els antozous, el percentatge de nord-atlàntiques se situa per sobre del 40%, però mentre que en els antozous el nombre d’espècies endèmiques és similar a l’esmentat, en el cas dels hidrozous les endèmiques i les cosmopolites són pels volts del 20%. Els hidrozous es poden dispersar tant per mitjà de les larves com de les meduses, però hom pensa que les primeres tenen més èxit, o que, almenys energèticament, són més rendibles; això justificaria un nombre més alt d’espècies d’origen atlàntic, tot i que les que poden desenvolupar meduses amb un temps de residència prou llarg al plàncton són les cosmopolites. Alhora, aquestes darreres són afavorides en el plàncton principalment pel seu caràcter mesopelàgic. En l’actualitat no es pensa que la fauna de cnidaris de la Mediterrània sigui pobra, comparada amb la de l’oceà Atlàntic.