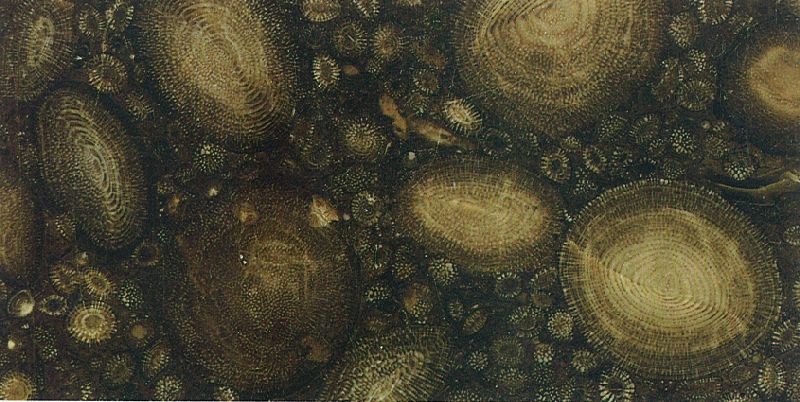
Secció polida d’una roca calcària amb nummulits de l’Eocè (× 3).
Jordi Vidal
Els foraminífers són protozous rizòpodes, és a dir, organismes unicel·lulars amb pseudopodis. Hom els considera pròxims al grup de les amebes i les tecamebes, però diferentment d’aquestes, tenen una closca calcària.
L’estructura cel·lular d’un foraminífer és realment excepcional per les seves dimensions i per la seva complexitat (hom ha dit que únicament són comparables als animals superiors, formats per òrgans i teixits, i no a les cèl·lules que els formen); recordem que els nummulits de l’Eocè o les loftúsies (Loftusia) del Cretaci superior poden atènyer una grandària superior als 10 cm, i que la seva closca és dividida en un gran nombre de compartiments. Els foraminífers formen un dels pocs grups de protists que fixen permanentment la seva superfície cel·lular per mitjà d’una closca mineralitzada, segregada totalment o parcial pel mateix animal. El protoplasma s’estén fora de la closca a través d’una o de diverses obertures i la recobreix exteriorment formant els pseudopodis. El seu protoplasma és incolor, però pot contenir petites quantitats de pigments orgànics, compostos de ferro i material lipidic, que li donen color, bé que aquest pot derivar també de la presència de simbionts. A l’interior del protoplasma hi ha el nucli o els nuclis, d’una grandària que varia d’una espècie a l’altra; inclou també les estructures pròpies dels protists, que hom anomena organel·les per tal de diferenciar-les dels veritables orgànuls dels metazous. Els foraminífers realitzen les funcions pròpies de qualsevol cèl·lula: per a la nutrició, els pseudopodis (formats per fins filaments d’una substància gelificada) creen un corrent de convecció que introdueix a l’interior del protoplasma les partícules alimentàries i que expel·leix les que excreta; alguns foraminífers (Bathysiphon) són filtradors.
La majoria dels foraminífers són bentònics i viuen sobre el fons marí, les plantes, els bucs de vaixells enfonsats, etc. Alguns han escollit la vida sèssil i viuen fixats durant tota la seva existència damunt d’un substrat, i generalment cimentats per mitjà de la closca. D’altres només es fixen temporalment i, per tant, poden desplaçar-se lliurement mitjançant els pseudopodis, que poden arribar a tenir una llargària superior a dues vegades el diàmetre de la closca; la velocitat de desplaçament és d’alguns centímetres per hora, i observacions fetes en el seu hàbitat natural indiquen que hi ha una termotaxi positiva i una fototaxi negativa en el seu desplaçament. Hom pensa que els individus sèssils que cauen al fons del mar des de les plantes marines on viuen són capaços de trobar el camí de tornada a les fulles que els suportaven; i, igualment, que els que queden enterrats en el sediment del fons per corrents de tempesta troben el camí de tornada cap a la superfície. Algunes espècies són planctòniques, i viuen surant en l’aigua marina, on es desplacen únicament pels moviments d’aquesta.
Si tenim en compte que els foraminífers poden atènyer dimensions considerables i que el tipus de creixement és lineal (una cambra després d’una altra), hem de pensar que el protoplasma ha de recórrer grans distàncies per sortir a l’exterior de la closca, cosa que dificulta la locomoció; la solució d’aquest problema és establir connexions tan curtes com sigui possible entre la primera cambra i la darrera. El tipus de creixement lineal fa que en closques amb obertures simples, quan el protoplasma es retreu, la locomoció quedi blocada; per tal de mantenir la mobilitat durant els estadis retrets, algunes espècies han desenvolupat tot un sistema de «portes d’emergència» (el sistema de canals) a cada cambra de la seqüència ontogenètica, la qual cosa permet que el protoplasma pseudopodial surti des de cada cambra quan les darreres estan blocades. En aquestes formes canaliculades, l’última cambra no té obertura primària sinó que aquesta es forma per reabsorció dels materials de la paret de la cara d’obertura en el moment de començar la construcció de la cambra següent. Els canals permeten als foraminífers d’escapar-se dels medis «tòxics», fins i tot amb el protoplasma retret.
La closca
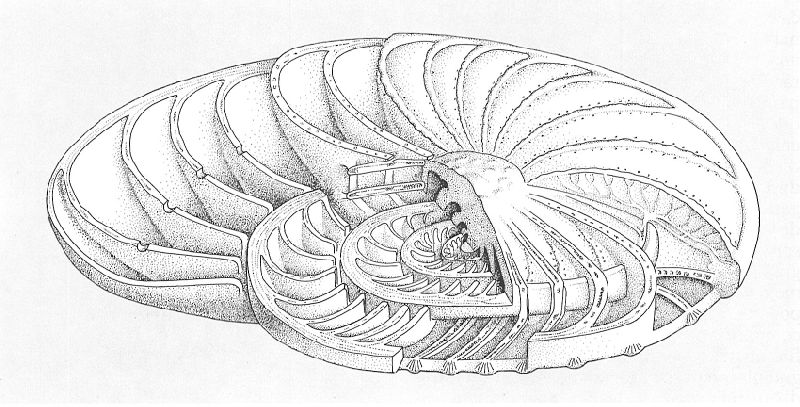
Estructura interna d’una operculina, segons la interpretació de Carpenter (1862) modificada per Reichel, per a mostrar tota la seva complexitat. L’organisme viu, unicel·lular, es recobreix d’aquesta closca que ell mateix secreta i n’ocupa cadascuna de les cambres; a més, emet pseudopodis a l’exterior a través de petites obertures, per mitjà dels quals es desplaça o és capaç de crear corrents que li portin aliment.
Amadeu Blasco, original de W. Windhauser
L’element bàsic que diferencia els foraminífers i l’únic que es conserva en estat fòssil és la closca, que pot tenir una complexitat molt variable. Pot tenir un sol compartiment, que forma l’anomenada cambra, que creix d’una manera continuada (foraminífers uniloculars) o bé diverses cambres (foraminífers pluriloculars), que es formen a partir del prolòcul o cambra embrionària en etapes successives de creixement (creixement discontinu). El protoplasma que participa en la formació de cada cambra surt per l’obertura o les obertures de la cambra precedent. Tots els elements d’una mateixa cambra corresponen a una única etapa de creixement.
La cambra és limitada per la paret mineralitzada que la separa de l’exterior, i en la qual hom distingeix tres parts (distal o perifèrica, umbilical o proximal i lateral), segons la seva posició respecte de l’eix de creixement. La part de la paret que conté l’obertura o les obertures s’anomena cara apertural; en formar-se una nova cambra, la cara apertural de l’anterior queda englobada al seu interior i constitueix l’àrea d’unió entre ambdues cambres veïnes (septe). El pas del protoplasma cel·lular d’una cambra a l’altra es fa a través de les obertures. L’espai interior de la cambra, el lumen, pot estar totalment o parcialment dividit. Si la divisió és completa, la cambra queda dividida en un conjunt de cambres més petites o cambretes. En un animal fortament compartimentat, els diferents espais són intercomunicats, de manera que el protoplasma els ocupa tots d’una manera simultània. Possiblement no hi ha cap diferenciació biològica entre cambres i cambretes.
Composició i estructura de la paret
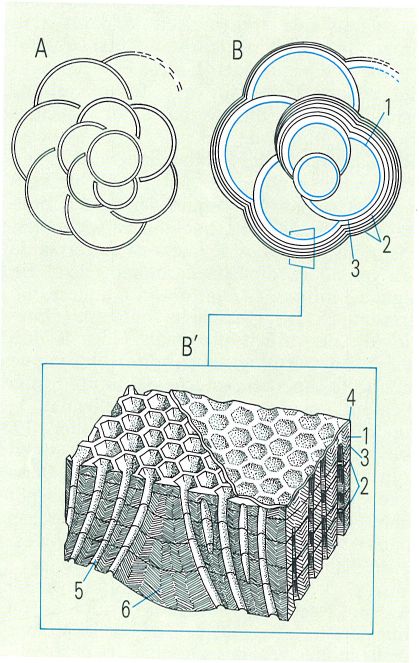
Esquema d’una closca no laminada (A) i d’una closca laminada (B) de foraminífer, i detall (B’) de l’ultrastructura de la paret lateral, perforada, d’una cambra. Totes les làmines són mineralitzades excepte la més interna, que s’ha representat puntejada, que és orgànica. La interpretació es deu a Hottinger i a Muller-Merz. Hom hi ha indicat: 1 làmina interna, 2 làmines externes, 3 làmina mitjana, 4 capa orgànica, 5 porus, 6 pilar ornamental.
Amadeu Blasco
El tipus de paret de la closca dels foraminífers varia segons el material que participa en la seva construcció i la seva estructura, i per això hom pot emprar-lo per a distingir els grups; malgrat que la paret pot ser modificada per processos diagenètics, l’especificitat de la modificació ajuda sovint a reconèixer la paret original. En els foraminífers actuals, cal distingir tres tipus fonamentals de paret mineral·litzada: aglutinada (o arenosa), aporcellanada i lamel·lar-perforada (hialina); totes són entapissades interiorment per una capa orgànica. Entre les formes fòssils, cal distingir, a més, les closques microgranulars.
La paret aglutinada és constituïda per material exogen (aglutinat) que es troba inclòs en un ciment segregat pel mateix individu. Si n’observem una secció amb llum incident se’ns presenta de color gris clar, mentre que amb llum transmesa és de color gris fosc. El material aglutinat és divers: grans de quars o altres minerals, restes d’organismes, etc. En alguns casos hi ha una veritable selecció i només s’aglutinen espícules d’esponja, closques d’altres foraminífers més petits, diatomees, plaques ambulacrals d’estrelles de mar o coccòlits i, llavors, la selecció no és lligada al medi i és considerada, per tant, específica. La major part de les vegades, però, s’aglutina material detrític de l’entorn: en fàcies calcàries, el material aglutinat pot ser exclusivament calcari (aleshores es fa difícil de distingir aquestes closques de les calcàries pròpiament dites), mentre que en sediments gresosos o margogresosos l’acumulació de material aglutinat és molt densa i arriba fins i tot a emmascarar els caràcters estructurals de l’interior de la closca.
La paret microgranular és característica d’un grup extingit de foraminífers que foren dominants durant el Paleozoic. La paret descrita clàssicament com a microgranular és constituïda per cristalls de calcita equidimensionals amb empaquetat dens. Aquesta microstructura ha estat objecte de moltes discussions a causa del probable emmascarament de la naturalesa real, ja que pot haver-se produït una recristal·lització posterior. Això no obstant, sembla haver-hi acord a considerar que la paret és formada per material segregat, encara que pot incloure petites proporcions de partícules aglutinades. En la majoria de les formes d’aquest grup poden distingir-se capes de diferent textura, que responen a dos models generals: d’una banda, capes microgranulars i, de l’altra, capes fibroses o pseudofibroses, que són el resultat d’una alineació regular dels grànuls de calcita. Algunes formes, especialment les de la superfamília dels fusulinacis, poden arribar a presentar un grau molt alt de complexitat: hom pot arribar a diferenciar-hi fins quatre capes diferents. Les formes microgranulars han estat esmentades en el Cambrià d’Austràlia, per la qual cosa la seva aparició se situa just després de la de les closques aglutinades.
La paret aporcel·lanada és d’origen endogen. És formada per grans de calcita sense una orientació òptica preferent. Són closques opaques, que es mostren blanques amb llum incident i negres amb llum transmesa. S’alteren fàcilment, i la recristal·lització comença al centre de la closca o bé modifica únicament l’anomenada capa basal (en els alveolínids i els fabulàrids); aquesta facilitat d’alteració és conseqüència de la porositat de la paret. Les primeres closques aporcellanades presents en el registre fòssil apareixen en materials del Carbonífer. Tant les closques aporcellanades com les aglutinades i les microgranulars que ja hem comentat no són lamel·lars. El terme no lamel·lar indica que, per a formar una cambra nova, no hi ha addició de material sobre la superfície exposada de les cambres ja formades. No obstant això, és interessant d’observar que en alguns orbitòlits (els Opertorbitolites) la paret lateral de les cambres és doblegada enrere bo i recobrint part del disc preexistent; de la mateixa manera, el botó umbilical que presenten alguns gèneres aporcellanats (com ara els meandropsínids), també és format per làmines superposades. L’estructura aporcellanada, bé que respon a la mateixa construcció que les closques aglutinades, és més complexa, ja que en ella la formació d’una cambra nova no sols implica la secreció d’un esquelet orgànic sinó també un procés que dirigeix la formació de cristalls de calcita.
La paret lamel·lar perforada és formada primàriament per dues làmines (interna i externa) i una zona intermèdia formada per grans cristalls i per intersticis reblerts de matèria orgànica, la qual cosa indica per on s’ha iniciat la mineralització en el moment de formar-se la nova cambra. En aquest sistema de creixement lamel·lar, la làmina externa creix de manera que recobreix la part de la closca ja formada. La major part de les closques lamel·lars són de calcita i les làmines es disposen en piles i de manera radial a la superfície de la closca. Això no obstant, n’hi ha alguns que construeixen la paret amb aragonita i són d’estructura columnar (Ceratobulimina, Epistomina). Un cas especial és el del gènere Carterina, que té una closca espicular, és a dir, formada per espícules monocristal·lines de calcita. La construcció dita lamel·lar sembla aparèixer només en foraminífers amb porus, per la qual cosa hom anomena aquestes closques perforades, diferentment de la resta, que són imperforades. Els porus són zones de la closca en forma de tub on no es diposita carbonat càlcic. En algunes formes (Amphistegina) aquest tub presenta envans. Quan hi ha simbionts (Operculina), els orificis proximals i distals dels porus són morfològicament diferents, ja que mentre la superfície interna del tub té forma d’embut envoltat d’una vora poligonal, la superfície externa és simple i circular. La presència de porus sempre s’ha considerat com un caràcter sistemàtic important. Quan l’individu és viu, els porus són tancats per matèria orgànica i no els pot travessar cap mena de protoplasma. La funció dels porus és, entre d’altres, d’establir l’intercanvi de gasos entre el protoplasma de la cambra i el medi extern. Les closques lamel·lars perforades són transparents o translúcides, bé que les formes fòssils moltes vegades són opaques per rebliment de sediment. Les seccions es mostren fosques amb llum incident i translúcides amb llum transmesa. Aquest grup de foraminífers comprèn les closques de morfologia més complexa, i la seva aparició en el registre fòssil és posterior a la dels altres grups.
La forma de la closca
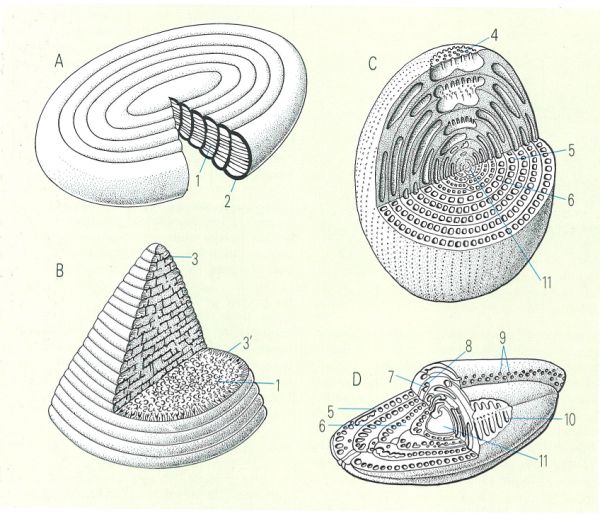
Disposició de les cambres i forma de la closca en diferents tipus de foraminífers: A discoïdal-evoluta, B cònico-uniseriada, C enrotllament miliòlid, D planispiralfusiforme. 1 pilars d’endosquelet, 2 exosquelet simple, 3 i 3’ bigues i llastes de l’exosquelet, 4 obertura en trematoporus, 5 cambretes, 6 capa basal, 7 canal preseptal, 8 canal postseptal, 9 obertures, 10 envans, 11 cambra inicial.
Amadeu Blasco, a partir de fonts diverses
D’una manera general, podem dir que la forma de la closca d’un foraminífer és condicionada pels factors següents: la forma i les dimensions de l’embrió, el nombre d’etapes de creixement fins a arribar a l’estat adult (és a dir, el nombre de cambres de què disposa un foraminífer), la forma de la cambra i les seves modificacions durant l’ontogènia, la disposició de les cambres, i el nombre i la disposició de les seves obertures.
Pel que fa a la forma i les dimensions de l’embrió, cal tenir en compte que el prolòcul (primera cambra en un foraminífer plurilocular) pot ser simple (ortostílic), o bé tenir una comunicació en forma de tub semienrotllat amb la cambra següent (flexostílic, com en els alveolínids) o, encara, ser format per més d’una cambra (normalment, protoconquillla i deuteroconquilla), i en aquest cas és extremament complexa, com en els orbitolínids. La presència de més d’una obertura a la cambra embrionària i la seva disposició condicionen la sortida del flux protoplasmàtic per més d’un lloc i, per tant, la formació simultània de més d’una cambra.
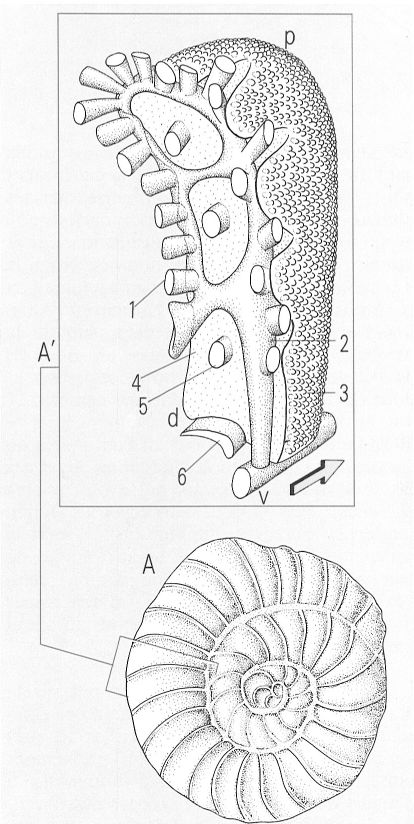
Closca sencera d’un foraminífer del tipus Daviesina operculiforme (A) i detalls de l’estructura d’una cambra (A’). 1 canals laterals suturals, 2 canal lateral del sistema intraseptal, 3 superfície lateral perforada de la cambra, 4 superfície septal de la cambra, 5 estoló, 6 obertura principal, d costat dorsal de la cambra, v costat ventral de la cambra, p costat perifèric de la cambra.
Amadeu Blasco, amb dades de M. Caus, L. Hottinger i Tambareau.
La forma de la cambra i les seves modificacions ocorregudes durant l’ontogènia són controlades per la longitud i la disposició dels corrents pseudopodials responsables de la formació de la capa orgànica de la nova cambra, que posteriorment serà calcificada.
Les cambres se situen en línia, una darrere l’altra, i segons un eix que determina que les closques siguin rectilínies (uniseriades), enrotllades en un pla (planispirals), helicoïdals (trocospirals) o fins i tot en diversos plans (enrotllament miliòlid). Tots aquests tipus, però, poden aparèixer combinats —algunes closques inicialment enrotllades poden esdevenir biseriades o uniseriades—. Alguns foraminífers presenten dos tipus diferents de cambres: les que es formen a partir de l’embrió (cambres equatorials) i les que ho fan a partir de les equatorials (cambres laterals), disposició que presenten, per exemple, els orbitolínids.
El nombre i la disposició de les obertures són responsables del flux del protoplasma a l’interior de les closques. Les obertures poden ser úniques o múltiples i poden canviar al llarg de l’ontogènia. Diem que una obertura és primària quan s’ha originat al mateix temps que la cambra (cas dels foraminífers no perforats i petits lamel·lars perforats), mentre que les obertures són considerades secundàries quan es formen per reabsorció localitzada de les diferents làmines del septe quan es forma la nova cambra (en alguns foraminífers lamel·lars perforats). Les obertures primàries estan lligades a la forma i a la disposició de la cambra. En els foraminífers en què els estolons es formen per reabsorció, és independent de la forma de la darrera cambra.
El ritme de creixement durant l’ontogènia no és constant i pot sofrir acceleracions. És el cas de les closques planispirals, que evolucionen a cícliques. Les closques trocospirals tenen tendència a crear un umbilic (espai obert al centre de la closca), que té l’avantatge de comunicar directament cada cambra amb l’exterior mitjançant la creació d’una o més obertures umbilicals suplementàries durant el creixement.
La forma final d’un foraminífer pot ser la mateixa encara que s’hagi generat per diferents formes de creixement. Així, les closques discoïdals poden derivar d’un creixement uniserial (Orbitolina), espiral (Nummulites) o d’anells de cambres (Orbitoides, Discocyclina). Contràriament, el mateix tipus de disposició de les cambres pot donar formes diferents; l’exemple més clar el tenim en els gèneres del Cretaci superior: Cuneolina, en forma de ventall, i Dicyclina, cíclica.
Estructura interna de la closca
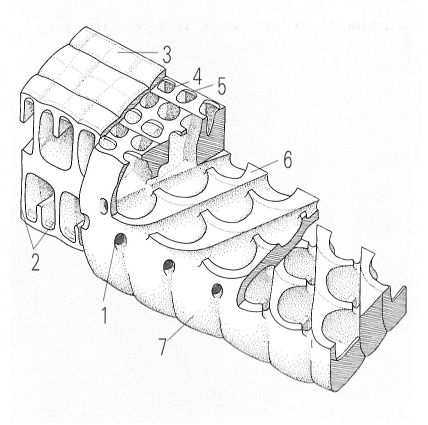
Detall dels elements estructurals d’un foraminífer aglutinat discoïdal, on es pot observar el tipus d’endosquelet format pel sistema d’estolons encreuats obliquament. 1 estoló, 2 punt d’unió entre l’exosquelet i l’endosquelet, 3 paret lateral, 4 biga (element de l’exosquelet), 5 llasta (element de l’exosquelet), 6 septe, 7 cara lateral.
Amadeu Blasco, a partir d’originals de L. Hottinger i M. Caus.
Tret del gènere aglutinat aberrant Julinella, tots els grans foraminífers tenen un gran nombre de compartiments. Durant l’ontogènia, els compartiments successius no excedeixen d’un cert volum sense ser dividits. En la filogènia d’espècies relacionades s’observa que el volum dels compartiments és més o menys constant, bé que la grandària és molt superior. En aquest cas, el volum del compartiment és constant i pot servir en la taxonomia de les espècies. Malgrat això, no hi ha una relació directa entre el volum i el nombre de compartiments; així, hi ha casos amb cambres relativament grans no dividides i d’altres de petites i dividides.
L’interior de la cambra pot ser dividit per elements estructurals que reben el nom d’exosquelet quan afecten només les parets externes, o bé d’endosquelet quan estan lligats al sistema d’obertures. La presència o absència d’aquests elements i la seva forma i disposició constitueixen un caràcter taxonòmic. L’exosquelet consta d’elements estructurals que, segons la seva posició i sempre en relació amb els sistemes arquitectònics, són anomenats bigues quan són perpendiculars al septe i llastes quan són transversals, és a dir, paral·lels al septe. L’endosquelet presenta un conjunt d’elements estructurals que s’insereixen entre els septes i la cara d’obertura; són els pilars o elements en forma de columna, i els envans o elements en forma de paret contínua. Aquests elements interns a la cambra s’ordenen de manera que no impedeixen el pas del protoplasma a través de les obertures de cambres successives. Hi ha grups que tenen un endosquelet particular: els miliòlids presenten una capa massissa en forma de «sol» (la capa basal), que cobreix les zones de la closca ja formades i exposades a l’hora de formar-se una cambra; l’endosquelet dels fusulínids és semblant, i és anomenat coma («chomata») quan es concentra en dues bandes en espiral que voregen l’obertura, o bé paracoma («parachoma») quan és més complex i presenta envans. En el cas dels foraminífers lamel·lars, les estructures endosquelètiques poden tenir forma de placa i cobrir només les obertures, com en els bulimínids i els bolivínids, o bé poden cobrir en part o totalment el terra de les cambres formades anteriorment, com en els rotàlids. Com ja hem dit, les closques lamel·lars presenten una diversificació morfològica molt més gran que les closques no lamel·lars, la qual cosa fa que en algunes la làmina interna esdevingui independent de l’externa i recobreixi la cara de l’obertura de la cambra precedent formant-hi l’anomenat plec septal.
En alguns foraminífers lamel·lars perforats, les unions de dues cambres consecutives són molt petites perquè tenen les sutures enfonsades cap a l’interior. A conseqüència d’aquest enfonsament, es genera un espai buit obert a l’exterior. Aquests espais, que es formen també per processos de reabsorció en diferents parts de la closca, són reblerts de protoplasma pseudopodial i, probablement tenen la funció de mantenir una diferenciació permanent del protoplasma. Hom distingeix tres sistemes fonamentals de canals, que es presenten en els elfídids, els nummulítids i els calcarínids, respectivament. Variacions d’a-quests tipus fonamentals donen els diferents sistemes de canals que hom pot trobar en el conjunt de foraminífers, segons el seu arranjament i la forma de les cambres.
Els principals grups de foraminífers
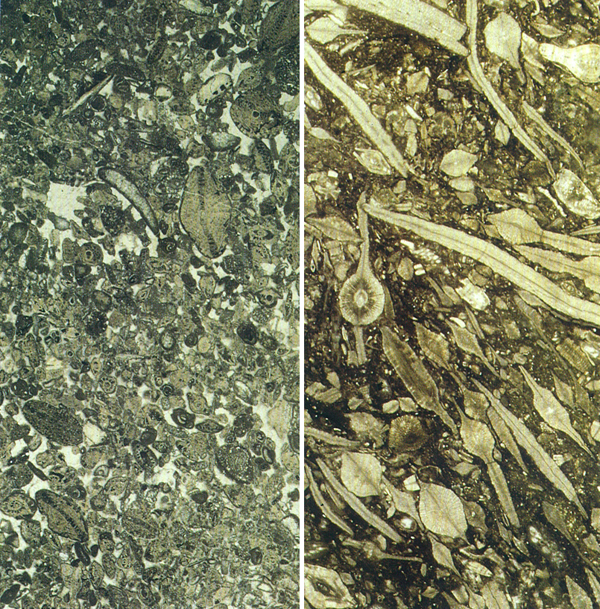
Microfàcies de microforaminífers. A l’esquerra, procedent del Cretaci superior de Sant Julià de Cerdanyola (Berguedà) on podem distingir principalment Orbitoides i Siderolites (× 8). A la dreta, procedent de l’Eocè de la Plana de Vic, amb nombroses seccions de discociclines i també algunes d’Heterostegines i Nummulites (× 7,5).
Maragda Caus
Hom utilitza normalment en els treballs micropaleontològics tres grups artificials: els macroforaminífers (foraminífers grans i d’estructura interna complexa), els microforaminífers i els foraminífers planctònics (caracteritzats pel seu hàbitat particular). Però dins l’àmbit de la sistemàtica, la classificació de l’ordre dels foraminífers més utilizada és la de Loeblich i Tappan (posteriorment modificada, en part, pels mateixos autors) tot i que encara hi ha alguna controvèrsia respecte dels caràcters d’importància sistemàtica en alguns dels grups. Aquest sistema de classificació considera l’ordre dels foraminífers subdividit en cins subordres: els al·logromins (Allogromiina), que són actuals i que per tant no tractarem en aquesta part de l’obra; els textulariins (Textulariina), els fusulinins (Fusulinina), els miliolins (Miliolina) i els rotalins (Rotaliina).
Els textularins
Es caracteritzen perquè tenen les closques aglutinades i no laminars, i entre ells hom en distingeix dues superfamílies: la dels ammodiscacis (Ammodiscacea), que reuneixen formes uniloculars, com Rhabdammina, Ammodiscus, i Bathisyphon, i la dels lituolacis (Lituolacea), on trobem formes simples com Textularia, Ammobaculites i Trochammina, al costat de formes molt complexes, com ara els representants de les famílies de lituòlids (Lituolidae) en sentit ampli (Spirocyclina, Choffatella, Ciclopsinella, etc.) i dels orbitolínids (Orbitolinidae), amb Orbitolina, Paleodictyoconus, Coskinolina, etc.
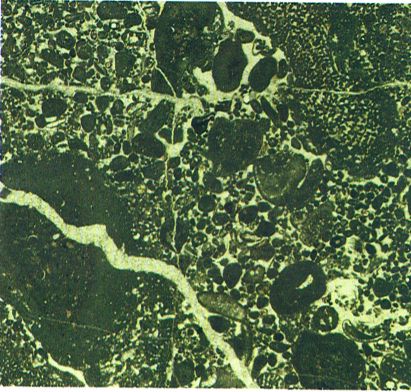
Microfàcies del Juràssic de So Na Moixa, a Mallorca, on podem veure orbitopsel·les i lituoseptes, foraminífers aglutinats, d’estructura complexa (× 10).
Maragda Caus.
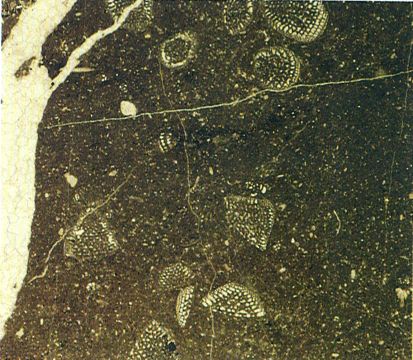
Microfàcies del Cretaci inferior de la serra de Prada (Alt Urgell) amb seccions de petits orbitolínids (× 10).
Maragda Caus.
Els lituòlids són formes grans, complexes, espiralades o cícliques que es troben de manera abundant en sediments soms del Cretaci. Els orbitolínids, al contrari, tenen forma de barret xinès. Ambdós grups són importants per a la datació dels sediments abans esmentats i són molt abundants als Països Catalans. Podem reconèixer-los a les calcàries de Prada (Alt Urgell), a les calcàries del Montsec i a les calcàries d’Aulet (Alta Ribagorça), per exemple.
Els fusulins
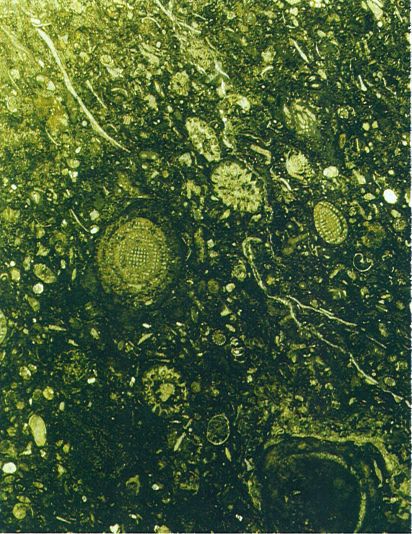
Microfàcies del paleozoic, d’una mostra procedent de Iugoslàvia. Podem distingir-hi, principalment, fusulines i algues calcàries (× 10).
Maragda Caus.
Dins aquest subordre hom inclou una gran varietat de foraminífers de closca formada per calcita microgranular, que en les formes més primitives és formada per una sola capa. Bé que apareixen en el Paleozoic inferior, tenen la màxima diversificació durant el Carbonífer i el Permià. Hom en distingeix tres superfamílies: la dels paratnuramminacis (Paratnuramminaceae), que reuneix formes d’origen problemàtic, moltes vegades sense relació entre elles, i de closca unilocular, que varia d’esfèrica fins a forma de tub; la dels endotiracis (Endothyracea), que agrupen petits foraminífers paleozoics amb closca bilocular o plurilocular, on les cambres poden estar enrotllades de diversa manera (Climacammina, Archaediscus, Bradyina, Endothyra); i la dels fusulinacis (Fusulinacea), que formen un grup del Carbonífer i el Permià que reuneix la major part de tots els fusulins.
Els fusulinacis són rars als Països Catalans, on el Paleozoic marí és poc desenvolupat, mentre que a la península Ibèrica són importants els fusulínids carbonífers de Cantàbria. Els fusulinacis tenen una gran complexitat estructural: són proveïts d’una closca enrotllada al voltant d’un eix, la forma de la qual varia des de lenticular fins a fusiforme. La paret calcària (espiroteca) és formada, en les formes més primitives, per una sola capa (proteca) mentre que les formes més evolucionades poden tenir-ne fins a quatre (tèctum, tectòrium superior, tectòrium inferior i diafanoteca). En el Carbonífer superior es troba una modificació de la paret que inclou un tèctum i una diafanoteca eixamplada en la qual s’observen porus gruixuts que de vegades es ramifiquen o es fusionen i donen lloc a cavitats més grans (els alvèols), que en conjunt formen una estructura anomenada kerioteca; l’espai intern de la closca és dividit en cambres que es comuniquen entre elles per una obertura situada a la base de cada septe, en la seva part central (el túnel). Durant el Permià, en lloc d’un túnel poden aparèixer a la base dels septes obertures múltiples (foramina). Els fusulínids, ultra els tectòriums, presenten altres dipòsits secundaris d’una gran importància sistemàtica, les comes i les paracomes. Altres dipòsits secundaris són els reompliments axials. La gran especialització d’aquest grup fa que s’utilitzin àmpliament en correlacions del Carbonífer i el Permià, i així hom ha establert zonacions per a aquests dos períodes.
Els miliolins
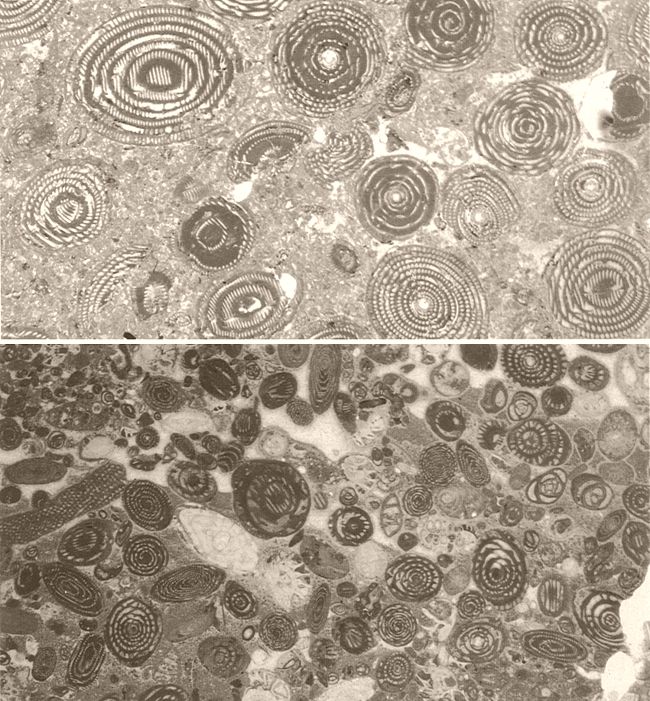
Microfàcies de l’Eocè de les serres marginals catalanes (a dalt), on podem veure nombroses seccions d’alveolines (× 13), i d’Ivars de Noguera (a baix), on destaquen fabulàries, orbitolines, alveolines i alguns macroforaminífers lamel·lars perforats (× 7,5).
Maragda Caus.
El grup dels miliolins és format per foraminífers de closca imperforada i d’aparença aporcellanada. És un grup heterogeni, que inclou formes simples com Vidalina, Ciclogyra o Fischerina, formades per un tub enrotllat, i formes complexes, com Sorites, Orbitolites i Lacazina. Inclou diversos grups de macroforaminífers que tenen una gran importància biostratigràfica perquè hom els ha emprats per a fonamentar bona part de la biostratigrafia del Cretaci superior, el Paleogen i el Neogen. Destaquen, per la seva importància, els meandropsínids (Meandropsinidae), els sorítids (Soritidae) i els alveolínids (Alveolinidae).
Els meandropsínids del Cretaci superior són formes de grans dimensions, molt planes i amb la superfície externa coberta de cambres d’aspecte meandriforme. Els trobem a les calcàries del Montsec (Meandropsina), al barranc de la Posa (Fascipira) o al massís de Bac Grillera (Larrazetia).
Els sorítids apareixen al Terciari i colonitzen la major part de les plataformes eocenes de les nostres terres (Orbitolites i Opertorbitolites). Són formes grosses, planes i que presenten interiorment una estructura interna on els estolons es disposen de manera encreuada. Podem observar-los a les formacions d’Orpí (Anoia), d’Àger (Noguera) o de Collbàs (Anoia). Els Sorites, Marginopora i Amphisorus del Miocè i els actuals tenen una estructura semblant als Orbitolites. El gènere miocè i l’actual Archaias és semblant al grup dels sorítids, però els seus estolons tenen disposició radial.

Aspecte de dues microfàcies del Cretaci superior: a dalt (× 5) procedent del congost de Terradets (serra del Montsec), on podem veure abundants lacazines i altres miliòlids i algunes rotalies i macroforaminífers aglutinats complexos; a baix en una mostra que procedeix dels voltants d’Orcau (Pallars Jussà), hi podem distingir lepidorbitoides i briozous (× 5).
Maragda Caus
Un altre grup important de macroforaminífers aporcellanats el constitueixen els miliòlids i els alveolínids. En el primer cas tenim formes com Lacazina, Fabularia o Periloculina, abundants a les plataformes carbonatades tant del Cretaci superior (calcàries del Montsec; calcàries d’Alinyà, Alt Urgell; calcàries de la Nou, Berguedà) com del Paleogen (calcàries d’Ivars de Noguera, Llorenç de Montgai, Noguera). Tenen com a característica principal la presència d’un endosquelet en forma de capa basal que pot arribar a formar envans i a individualitzar cambretes; l’obertura (trematoporus) també és un caràcter distintiu.
Els alveolínids constitueixen un grup homogeni, de forma des d’esfèrica fins a fusiforme i d’enrotllament planispiral involut. L’endosquelet és igual al dels miliònids (capa basal) i té envans que divideixen la cambra en nombroses cambretes. La capa basal és massissa a les formes esfèriques però és d’estructura irregular a les fusiformes. En alguns casos, la capa basal es pot engruixir (flosculinització). Les obertures principals es disposen en filera a la cara oral i poden alternar amb d’altres de diàmetre menor (obertures suplementàries). Les cambretes d’un mateix pas de creixement estan interconnectades per canals que corren paral·lels al septe (canal postseptal i preseptal). Entre els alveolínids, n’hi ha de cretacis que, comunament anomenats prealveolines, engloben diversos gèneres (Praealveolina, Ovalveolina, Cisalveolina), com els que apareixen a la formació de Santa Fe, de paleògens (Alveolina), comuns en tota la plataforma soma durant l’Eocè mitjà i inferior, i de neògens i actuals com Borelis (que apareix a Mallorca i també al Penedès), Fosculinella i Alveolinella. El gènere Bullalveolina és restringit a l’Oligocè.
Els rotalins

Diversos orbitoides seccionats és el que destaca en la microfàcies de la fotografia (× 7,5), del Cretaci superior de la serra del Montsec.
Maragda Caus.
Dins del subordre dels rotalins se situen tots els foraminífers laminats perforats. La presència, doncs, de dues làmines permet la construcció de closques molt complexes i d’estructures fins ara desconegudes en el registre fòssil, com els plecs septals o els sistemes de canals. Loeblich i Tappan diferenciaren en aquest ordre nombroses superfamílies segons l’orientació òptica dels elements càlcics, però aquesta divisió és encara controvertida. Ací utilitzarem la classificació artificial, però pràctica, que els separa en microforaminífers, macroforaminífers i foraminífers planctònics.
Dins dels microforaminífers, que inclouen rotalins simples i petits, hom engloba diverses superfamílies. Els nodosariacis (Nodosariacea), que són formes uniseriades (Nodosaria, Frondicularia), planispirals (Lenticulina) o bé modificacions d’aquestes formes (Marginulina), tenen com a característica principal la presència d’una obertura amb una placa radiada en posició terminal. Els buliminacis (Buliminacea) són formes principalment biseriades (Bolivina) o triseriades (Bulimina), que presenten una obertura terminal i tenen forma de coma. Els orbitoïdacis (Orbitoidacea) comprenen rotalins de closca trocospiral, entre elles les del gènere Discorbis, que pot adaptar-se a medis lacunars. Els espiril·linacis (Spirillinacea) són característics perquè tenen la closca formada per un únic cristall de calcita (Spirillina, Patellina). En els robertinacis (Robertinacea) la closca és constituïda per aragonita en lloc de calcita (Ceratobulimina, Robertina). Els cassidulinacis (Cassidulinacea) reuneixen formes trocospirals, amb obertura en forma de llàgrima (Cassidulina, Pleurostomella), mentre que els nonionacis (Nonionacea) tenen closques planispirals i bàsicament involutes, i tenen l’obertura basal amb un llavi (Nonion, Pullenia). Finalment, els carterinacis (Carterinacea) són representats pel gènere Carterina, una forma gens comuna pel fet de tenir una closca formada per espícules calcàries disposades en una matriu també de calcita.
Els macroforaminífers perforats pertanyen principalment a dues superfamílies, la dels orbitoïdacis (Orbitoidacea) i la dels rotaliacis (Rotaliacea). Els orbitoïdacis inclouen els macroforaminífers anomenats en general «orbitoïdals» pel tipus de construcció de les seves cambres; tenen cambres de dos tipus: les equatorials, amb creixement orbitoïdal, i les laterals, que les flanquegen. Cal distingir-ne diversos grups que, si bé tenen un aspecte similar, no tenen cap relació filètica entre ells: els orbitòidids del Cretaci superior (Orbitoides i Onphalocyclus) i els lepidorbitòidids, també del Cretaci superior (Helicorbitoides, Lepidorbitoides), tots presents als Prepirineus; els discociclínids del Paleogen, comuns a les margues d’Igualada (Anoia) i de Vic (Osona); i els lepidociclínids del Neogen. Els rotaliacis són més complexos i també més coneguts. Cal distingir els rotàlids, amb formes trocospirals (Ammonia, Rotalia), els miogipsínids, en els quals les cambres adultes tenen desenvolupament orbitoïdal (Miogypsina, Miogypsinoides) i els calcarínids, un grup molt important del Cretaci superior (Siderolites, Pseudosiderolites) que apareix en les mateixes localitats que els orbitòidids però que també té representants en el Paleogen (Miscellanea, Pellatispira) i a l’actualitat (Calcarina). Els nummulítids, que a més dels Nummulites inclouen altres gèneres com Operculina, Heterostegina i Cicloclypeus, foren extraordinàriament importants en les mars eocèniques, fins al punt que, durant molt temps, hom anomenà l’Eocè període nummulític. Es tracta de formes planispirals, evolutes o involutes, algunes de les quals poden esdevenir cícliques (Cicloclypeus); en aquestes hom ha fonamentat, en bona manera, la biostratigrafia de les plataformes somes del Paleogen. Els sediments que contenen nummulits i altres gèneres afins són molt abundants als Països Catalans; apareixen a tota la plana de Vic, a la rodalia d’Igualada (on, a la serra de Collbàs, hi ha un coll anomenat de la Llentilla, per l’abundància d’aquest microfòssil), al Gironès, al País Valencià, a Mallorca, etc. La pedra nummulítica s’ha utilitzat de temps antic per a la construcció (pensem, per exemple, en la catedral de Girona). Els elfídids són formes planispirals que presenten unes característiques pròpies en la closca, conegudes com a processos retrals.
Els foraminífers planctònics corresponen a la superfamília dels globigerinacis (Globigerinacea). Són formes amb enrotllament trocospiral, planispiral o biserial, però totes tenen com a distintiu la forma globosa de les cambres i perforacions molt gruixudes. Algunes presenten fines espines que ajuden en la flotació mentre que d’altres són carenades. Apareixen en el Juràssic amb formes molt primitives anomenades protoglobigerines, però la seva gran expansió tingué lloc al Cretaci. El seu mode de vida les fa menys dependents del medi, comparat amb els altres foraminífers, i per això hom els ha utilitzats per a establir correlacions i hi ha diverses zonacions que tenen aquests foraminífers com a base. Per al Cretaci hom utilitza principalment les diverses espècies dels gèneres Rotalipora, Marginotruncana, Dicarinella i Globotruncana, entre altres, i per al Paleogen els representants dels globigerínids, els globorotàlids i els hantkenínids. Al Neogen trobem globigerínids i globorotàlids.
Paleoecologia
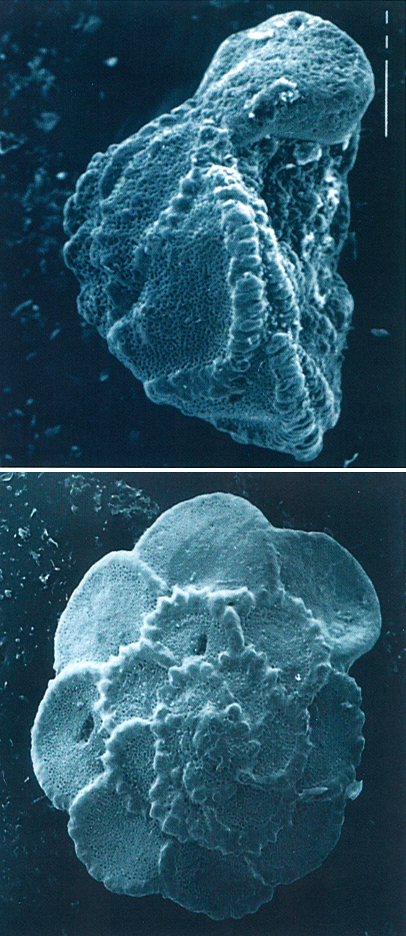
Foraminífers planctònics dels gèneres Rosita (a dalt) i Globotruncana (a baix), del Cretaci superior de l’Alta Ribagorça i del Pallars Jussà, respectivament, vistos al microscopi electrònic d’escandallatge (× 150).
Antoni Gómez
Els foraminífers constitueixen un dels pocs grups zoològics, tant fòssils com vivents, que es troben en tots els medis marins, des de les maresmes (hiposalins o hipersalins) fins als ambients pelàgics. La seva distribució és condicionada per diversos factors, tots interrelacionats. Un d’important és la profunditat, a causa de la seva incidència directa sobre la penetració de la llum, cosa que és especialment clara en els foraminífers que tenen simbionts. Pel que fa a la morfologia de la closca respecte del gradient de profunditat, en un gran nombre de foraminífers s’observa una tendència a l’aparició de formes aplatades en les closques discoïdals, trocospirals o còniques, a mesura que augmenta la fondària. Aquesta tendència es manifesta en la substitució d’unes espècies per unes altres o bé en una variabilitat intraspecífica. Entre les formes planctòniques, les espinoses generalment viuen a menys profunditat que les carenades. L’energia de l’aigua és important en el cas dels foraminífers bentònics de poca fondària perquè actua sobre el tipus de substrat (dur o tou), que influeix en la distribució d’alguns grups de foraminífers, principalment els de grans dimensions, i els nutrients. També hi ha una correlació entre la fondària, el tipus de substrat i l’energia de l’aigua, i el desenvolupament de grànuls o pústules sobre la closca dels macroforaminífers perforats. També el desenvolupament extrem del sistema de canals que es produeix en certes closques és lligat als factors ara esmentats. Hi ha, encara, altres factors, com la salinitat, la temperatura, la maduració de les poblacions i el grau de coordinació del cicle reproductor amb el cicle estacional, que influeixen d’una manera variable en la distribució dels macroforaminífers.