Les adaptacions dels animals a la vida a la sabana
L’estació seca sol provocar condicions difícils per a la vegetació que repercuteixen sobre les poblacions dels animals herbívors, l’alimentació dels quals perilla al mateix temps que disminueix la disponibilitat d’aigua per a abeurar-se. Les diverses espècies animals de sabana s’han d’adaptar a aquest cicle i ho fan de diferents maneres segons la seva anatomia, fisiologia, comportament i durada de la vida. L’escassesa de plantes llenyoses, altrament, planteja als animals problemes de cau i de protecció. En resposta a aquesta reducció, es desenvolupa la vida dins el sòl, que compensa, en part, la disminució de la dimensió vertical de la vegetació, i sorgeixen adaptacions especials per a camuflar-se entre les herbes. Les adaptacions dels animals a les particularitats de les sabanes es manifesta en tots els nivells d’organització, tant en la morfologia i en la fisiologia dels individus com en l’organització de la vida social i en la dinàmica de les poblacions.
Les adaptacions a escala individual
Cada organisme individual ha d’afrontar les diverses exigències del medi on viu. En el cas de les sabanes, aquestes traves deriven, en primer lloc, dels factors climàtics, sobretot durant l’estació seca, quan sorgeixen els problemes de trobar aigua per a beure i el material vegetal necessari per a l’alimentació. S’hi afegeix la necessitat de fugir dels depredadors en unes condicions en què és relativament difícil amagar-se.
El problema de l’aigua
Per a moltes espècies el problema de l’aigua és bàsic durant una part de l’any. Alguns carnívors de les zones subdesèrtiques es conformen amb la sang i els teixits de les seves preses i, entre els ungulats, alguns com l’òrix comú (Oryx dammah) i l’addax (Addax nasomaculatus) poden abstenir-se de beure durant moltes setmanes, però això són excepcions. Realment pocs mamífers aconsegueixen sobreviure sense beure i, com que durant l’estació seca l’aigua es troba a pocs llocs els animals han de desplaçar-se molt per a arribar-hi o bé concentrar-se al seu voltant, cosa que provoca una sobrepastura que augmenta la destrucció de la vegetació coincidint amb un període crític per a aquesta. A més, la necessitat de beure que tenen els animals ha conduït els depredadors a esperar les preses prop dels punts d’aigua. Aquest tipus de caça és una de les característiques importants en la vida de les feres, i el preu que en paguen els herbívors és un dels elements principals de la seva demografia. En el moment de beure fins i tot les girafes poden ser víctimes dels lleons, dels quals normalment s’escapen.
Per als animals de mida petita, com per exemple els rosegadors, poc capaços de desplaçaments de llarga distància, però també menys exigents pel que fa a les necessitats d’aigua, sovint la rosada és una font d’aigua suficient. La rosada és freqüent durant l’estació seca, a causa de les grans diferències de temperatura entre el dia i la nit provocades per la baixa higrometria de l’aire. La rosada també és utilitzada per alguns simis, com la mona vermella o “patas” (Erythrocebus patas). Altres espècies aprofiten l’aigua que hi ha en algunes plantes suculentes, i altres fins i tot s’abstenen de beure gràcies al seu metabolisme, que els permet mobilitzar quan cal els greixos de reserva i obtenir, així, aigua metabòlica.
Les altes temperatures no afavoreixen, a diferència de l’hivern a les contrades temperades, el pas a un estat de vida latent. Aquest estat només és possible a condició d’enterrar-se profundament en el sòl fins a arribar a horitzons menys càlids i més humits, tal com fan diferents organismes de la sabana d’altitud de Nimba, com el gripau vivípar Nectophrynoides occidentalis. En aquella mateixa sabana, per exemple, la closca ben desenvolupada dels cargols Gonaxis els dona una protecció eficaç mentre romanen colgats. La majoria de les espècies longeves d’amfibis de les diverses sabanes del món tenen un període de vida latent; només estan actius i es reprodueixen durant l’estació de les pluges que, de vegades, pot ser extremament curta, per contra, durant l’estació seca, s’enfonsen fins als horitzons més humits del sòl, sovint en el mateix emplaçament dels bassals temporals llavors dessecats. Els cucs de terra, en canvi, responen a l’estació seca entrant en quiescència i deshidratant-se. L’arribada de les pluges els permetrà, de seguida, reprendre la vida. Aquests organismes, però, es troben només a les sabanes relativament humides —on són abundants— mentre que són absents a les sabanes massa àrides.
El problema de l’alimentació: les migracions
La necessitat de trobar aliment planteja, de vegades, un problema difícil als herbívors, ja que en el cicle estacional de la vegetació a les sabanes generalment hi ha un període on aquesta pràcticament no existeix o, si més no, és poc favorable per a ser consumida. La recerca de l’aliment durant l’estació seca es pot traduir en migracions. La migració és la solució adoptada per molts ocells. Segons els casos es tracta d’un simple desplaçament intracontinental o bé d’una migració a gran escala. En el primer cas, l’espècie deixa una regió massa seca per arribar a una sabana més humida. A Lamto, per exemple, la sabana pre-forestal rep, durant l’estació eixuta, nombrosos migradors provinents de la zona saheliana. Quan arriba l’estació de les pluges, tornen per a niar en una sabana més àrida del Sahel. En el cas de les migracions a gran escala, els migradors de l’hemisferi septentrional es reprodueixen a les zones fredes o temperades d’Europa i retornen a l’Àfrica durant l’estació seca, generalment a sabanes relativament humides. Anteriorment ja s’ha citat el cas de les sabanes arbòries dels vessants del Nimba, a Guinea, que aixopluguen de desembre a març molts passeriformes nidificants a Europa. A la sabana d’altitud d’aquesta mateixa muntanya són orenetes i diversos rapinyaires les que aprofiten l’estació seca, però càlida, per a fugir dels freds de l’hivern europeu.
Alguns mamífers també fan migracions, però, naturalment, sempre dins un mateix continent. Així els elefants i diversos antílops poden desplaçarse desenes i, fins i tot, centenars de quilòmetres buscant ambients més favorables i una vegetació de millor qualitat. Malauradament aquestes migracions són limitades actualment per les instal·lacions humanes. És el cas, a l’Àfrica meridional, dels antílops saltadors o “springbok” (Antidorcas marsupialis), que abans realitzaven unes migracions que seguien milions d’individus. Les migracions del nyu de l’Àfrica central, en canvi, entre les sabanes baixes de la plana, pastura ideal durant l’estació de les pluges, i les sabanes boscoses dels altiplans, encara útils durant l’estació seca, continuen avui en dia atraient l’atenció dels naturalistes.
Els problemes de seguretat
Les sabanes són uns medis descoberts on, sovint, la visibilitat és molt bona. Això planteja un difícil problema de supervivència als animals que són preses potencials. Per resoldre aquest problema s’han adoptat solucions molt diferents d’acord amb la diversitat de la seva morfologia i les seves aptituds.
Una primera solució és la fugida. Els animals de les sabanes, especialment els mamífers, acostumen a ser corredors ràpids, de vegades fins i tot excepcionals. La zebra n’és l’exemple clàssic, però alguns antílops, com l’impala i les gaseles, encara són més notables ja que afegeixen salts a la velocitat. Les serps i els llangardaixos de les sabanes són també molt ràpids, com ho són també molts insectes i aràcnids. Aquesta aptitud de fugida ha determinat entre els depredadors adaptacions paral·leles. Alguns carnívors són també corredors remarcables, capaços d’esgotar les seves preses: és especialment el cas del guepard. Els licaons tenen, quan corren, una resistència excepcional.
A la tàctica de fugida es contraposa el camuflatge, que s’aconsegueix de diverses maneres: homocromia, homotípia i coloració disruptiva. Molts animals tenen una coloració propera a la que domina en el medi. Durant el període de creixement de l’herba, molts són verds. A les sabanes groguenques, sovint el seu color és ocre o bistre. Després dels incendis, alguns adopten una coloració negra. Les llagostes, els fàsmids i els pregadéus són els insectes que presenten més sovint aquest tipus d’adaptació. En algunes espècies el color de l’individu no varia al llarg de la seva vida; en altres es produeix una adaptació successiva als colors que corresponen a l’evolució del medi. Això s’observa per exemple en alguns pregadéus de les sabanes de l’Àfrica occidental. Per fer el canvi de coloració en funció dels colors del medi aprofiten el moment de la muda. Els coleòpters i els heteròpters són freqüentment homocroms amb el color del sòl, adoptant una coloració bruna o negra. A l’homocromia sovint s’afegeix una homotípia, que es manifesta a la sabana, per la forma allargada dels animals segons la forma de les gramínies. Els insectes, concretament les llagostes, els pregadéus i els fàsmids o insectes bastó proporcionen els exemples més característics d’aquestes semblances. Entre els cicadinis (petites cigales i insectes afins) s’observen homotípies sorprenents amb les llavors de les gramínies en les quals viuen. Per ser més eficaç, l’homotípia està pràcticament sempre associada a una homocromia.
En els animals de talla superior a la d’una tija o d’una fulla de gramínia són els dibuixos els que creen les semblances, gràcies a ratlles longitudinals de colors que van per exemple des del negre fins al groc. És el cas de molts amfibis de sabana, com Ptychadaena i alguns Hyperolius. Un tercer tipus de camuflatge està vinculat a la presència, sobre el cos, de dibuixos denominats de coloració disruptiva. Fent con si fossin taques d’ombra en un medi assolellat, modifiquen la imatge de la forma de l’animal, que és coneguda pels depredadors i per tant perillosa, per imatges no identificables i aparentment sense relació amb un ésser viu. Aquests fenòmens de camuflatge són innombrables en el món dels animals. Ja s’ha destacat el dels ous d’ocells, la closca dels quals és sovint coberta de diferents taques que fan passar desapercebuda la seva forma; els pollets de les espècies nidífugues també presenten taques o ratlles negres i clares que els fan pràcticament invisibles al niu.
També és el cas de molts ocells de les sabanes de colors apagats, i dels mamífers amb coloració tigrada, com per exemple la llebre, que és difícil de descobrir al seu cau. Les coloracions del pelatge dels antílops, amb ratlles blanques sobre fons ocre o rogenc també constitueixen excel·lents camuflatges traïts només pels moviments de l’animal. És conegut també el paper capital de les ratlles negres que arriben als ulls o fins i tot els inclouen, ja que aconsegueixen fer menys perceptible aquest òrgan que, si no, és molt fàcil d’identificar, o molts altres detalls del pelatge dels mamífers, del plomatge dels ocells o de les escates dels rèptils. Al camuflatge es vincula el fenomen d’immobilització reflexa: espantat, l’animal deixa de moure’s i d’aquesta manera fa més difícil que el depredador el vegi. Aprofita així al màxim el camuflatge per a passar desapercebut. Aquest fenomen ha estat adoptat per nombroses espècies, fins i tot de talla gran com els cérvols de l’Amèrica del Sud. També es troba en els pollets d’ocells i en alguns insectes o aranyes. L’homotípia i l’homocromia igual com passa amb l’aptitud per a córrer, també són presents entre els depredadors, possiblement perquè ells són també preses potencials d’altres depredadors, i també perquè aquesta similitud els permet no ser vistos per les seves preses.
Una conseqüència de l’homotípia en els medis de sabana, molt similars d’un a altre continent, és l’existència de notables convergències de forma: espècies que pertanyen a grups taxonòmics molt allunyats arriben a tenir aspectes extremament semblants com per una mena de mimetisme a distància. Aquests fenòmens no són propis dels medis de sabana, però aquí adquireixen modalitats diferents de les que es troben a la selva, ja que la il·luminació, el medi vegetal i, per tant l’aspecte general dels organismes animals són en ells mateixos molt diferents.
Les adaptacions etològiques i socials
Les possibilitats visuals que ofereixen els medis més o menys oberts, com són les sabanes, han afavorit en moltes espècies la vida en grups d’individus nombrosos. Certament, hi ha semblances amb les espècies de les selves, però el fenomen és menys freqüent si s’exclou el cas dels simis i els súids. De fet, la vida en grup facilita l’orientació dels animals quan es traslladen. També és un mitjà eficaç de defensa enfront els depredadors, molt especialment quan s’acompanya d’una estructura social sòlida.
La vida en grup
Algunes espècies de vida social, com les formigues i els tèrmits, estan preadaptades a la vida a les sabanes. És especialment el cas dels tèrmits, als quals les colònies populoses permeten la construcció al terra de nius voluminosos que asseguren una notable protecció davant la sequedat de l’aire i també enfront els depredadors. Moltes espècies de tèrmits tenen, doncs, a les sabanes un paper important que queda palès en la gran abundància de nius gegants.
Moltes espècies d’ocells de les sabanes viuen també formant grups. És el cas de l’estruç (Struthio camelus) i també, entre les espècies de talla gran, de les gallines de Guinea (Numida), i dels francolins (Francolinus i Pternistis). Les gangues (Pterodes) formen també estols nombrosos a les sabanes seques tant a l’Àfrica com a l’Àsia. Per a atènyer quotidianament els punts d’aigua, dels quals tenen necessitat, es desplacen en colles de molts centenars; les tórtores a l’Àfrica i els periquitos a Austràlia es comporten de la mateixa manera. Encara hi ha similituds infinitament més importants en els plocèids del grup dels teixidors; s’anomenen així perquè construeixen nius teixits d’herbes, generalment en forma d’una retorta amb l’obertura cap avall. La quelea de bec vermell, també anomenada menja-mill (Quelea quelea), constitueix immenses colònies de milers d’individus que nien en els arbres de la sabana, sovint en extensions de desenes d’hectàrees; consumidors de llavors, són un flagell temible dels conreus de mill, per això s’han provat tot els mitjans de lluita contra ells, fins i tot els gasos asfixiants i els llançaflames. En el teixidor social o republicà (Philetarius socius), es fa palesa una veritable vida social amb la construcció de nius col·lectius; aquests nius, reutilitzats i engrandits any rere any, poden ser de molts metres de diàmetre i tenen centenars de llotges en forma de retorta, cadascun dels quals serveix de niu a una femella que hi pon els ous i hi cria els pollets.
L’estructura social
Fora dels insectes socials (especialment formigues i tèrmits), és evidentment entre els mamífers on es troben els exemples més característics de la vida social que garanteix a l’espècie una seguretat i una eficàcia més grans. Així, la majoria dels ungulats, per exemple, viuen en grups familiars que comprenen un mascle dominant, femelles i les seves cries. En algunes espècies, els grups familiars s’apleguen de vegades formant grans ramats. És el nombre el que representa en tals casos una garantia de supervivència enfront de l’atac dels depredadors.
Els pècaris (Tayassu pecari, T. tajacu) i els facoquers (Phacochoerus aethiopicus) també formen grups que són un dels fonaments de la seva seguretat. En els elefants (Elephas maximus, Loxodonta africana), els ramats formen una veritable societat. Fora dels mascles solitaris, un ramat es compon, generalment, d’una colla de femelles, una de les quals adopta el paper dirigent tant per orientar el ramat com per evitar els perills i, eventualment, durant l’estació seca, per buscar els punts d’aigua. Aquesta vida social desenvolupada ha contribuït a desenvolupar la intel·ligència dels animals i els ha permès plantar cara, amb una màxima eficàcia, als problemes amb què es troben.
Entre els carnívors, les mangostes viuen en grups ben organitzats, sense que això impliqui l’organització de caceres en comú. Una veritable vida en societat és la que hi ha entre les suricates (Suricata suricatta) del S d’Àfrica, assegurada per caus associats en una veritable ciutat; l’interès més gran d’aquesta vida col·lectiva, amb associacions d’entre 10 i 30 individus, és assegurar una vigilància més eficaç davant dels depredadors terrestres, com el serval (Felis serval), el caracal (F. caracal) i els xacals (Canis), o aeris (rapinyaires) i, de vegades, també per escalfar-se mútuament durant les nits fredes. A diferència del lleopard (Panthera pardus) que caça en solitari, tal com fan també tots els fèlids petits, entre els guepards (Acinonyx jubatus) hi ha una cooperació estreta, però el grup té sempre un nombre molt reduït, sovint consisteix únicament en una parella o, de vegades, en dos o tres mascles. El lleó ( Panthera leo), en canvi, viu en grups petits d’uns pocs individus: un mascle, algunes femelles i les seves>cries. La caça és una ocupació on les femelles són protagonistes principals. La caça col·lectiva permet esperar i matar fàcilment preses grosses i ràpides com els nyus, els grans antílops o les zebres. En els licaons (Lycaon pictus), s’ha desenvolupat al màxim la cooperació social amb aquesta finalitat: el licaó, animal de talla mitjana, pot d’aquesta manera atacar preses força més grosses que ell, com zebres o nyus. La caça a l’aguait és freqüent, sobretot quan l’estació seca obliga els ungulats a reagrupar-se al voltant dels punts d’aigua.
És evidentment entre els simis on es troben els exemples d’una vida social més desenvolupada. En la mona vermella o “patas” (Erythrocebus patas), per exemple, espècie de talla petita que no disposa de mitjans de defensa importants, el que fa possible la supervivència de l’espècie és el comportament social, associat a un pelatge poc vistós i a un comportament discret i relativament silenciós. Per passar la nit, les mones vermelles fan servir els arbres i els arbustos dispersos per la sabana. Al contrari del que fan mentre és de dia, a la nit s’allunyen les unes de les altres, disminuint així els perills que representen els depredadors. Afegeixen a les altres precaucions una vida itinerant per un extens territori, evitant romandre gaires nits seguides en un mateix indret. La coherència del grup és assegurada conjuntament per l’acció del mascle dominant i per les activitats d’esplugatge habituals entre els simis. Enfront dels babuïns, els “patas” s’esquiven discretament.
Els cercopitecs de cara negra (Cercopithecus aethiops) són uns simis també fràgils i mal equipats per defensar-se, es protegeixen amb una vida arborícola i amb la seva organització social. Viuen en grups que poden ser de moltes desenes d’individus. Contràriament als formats per les mones vermelles, aquests grups són formats per molts mascles i moltes femelles, a més d’individus joves. Durant la nit, fan servir sempre els mateixos arbres d’un territori que coneixen bé. L’extensió d’aquest territori depèn principalment de les fonts alimentàries que contingui. Els cercopitecs de cara negra, pel fet que són arborícoles i per tant beneficiaris d’una tercera dimensió en l’espai, tenen un territori més reduït i es desplacen força menys que les mones vermelles o els babuïns grocs.
Els babuïns grocs (Papio cynocephalus) són, sens dubte, els simis de les sabanes que tenen l’organització social més perfeccionada. Viuen en grups sovint nombrosos amb una estructura social força definida i molt jerarquitzada. Al capdamunt hi ha una aristocràcia formada per uns pocs mascles, els més robustos i més emprenedors. Dominen els altres mascles i els joves. Entre les femelles, la dominància recau normalment en les que acaben de tenir una cria, i aconsegueixen així ser protegides pels mascles dominants. Perden aquest status quan els petits les deixen. Quan són a punt d’esdevenir adultes, les femelles, que no interessen els mascles dominants fins que realment són fecundes, solen ser inseminades per mascles joves. Aquest comportament, que limita la paternitat als mascles més forts, constitueix una mena de pràctica eugènica adreçada a una millora del grup. L’autoritat dels mascles dominants és reforçada per la solidaritat que hi ha entre ells. Es manifesta en actituds d’intimidació respecte als altres mascles, intimidació que de vegades acaba en algunes mossegades. L’individu punit practica aleshores un cert cerimonial d’humilitat amb relació al seu superior. Gràcies a aquesta sòlida estructura social, un grup de babuïns és capaç de resistir a un lleopard. En els desplaçaments, els joves mascles marxen davant, com a exploradors. Quan topen amb un lleopard són els mascles dominants que es llancen sobre d’ell i, sovint, el maten ferint-lo amb les seves potents dentadures o el fan fugir, no sense que, de vegades, en quedi algun d’ells estès. Durant el combat, les femelles i els joves es refugien als arbres o a les roques. Davant dels lleons el comportament és diferent: tot el grup corre a buscar refugi als arbres més alts. Els facilita la supervivència el bon coneixement del terreny que tenen, que els permet d’estar-se permanentment en un mateix territori.
L’estratègia de caça de les lleones
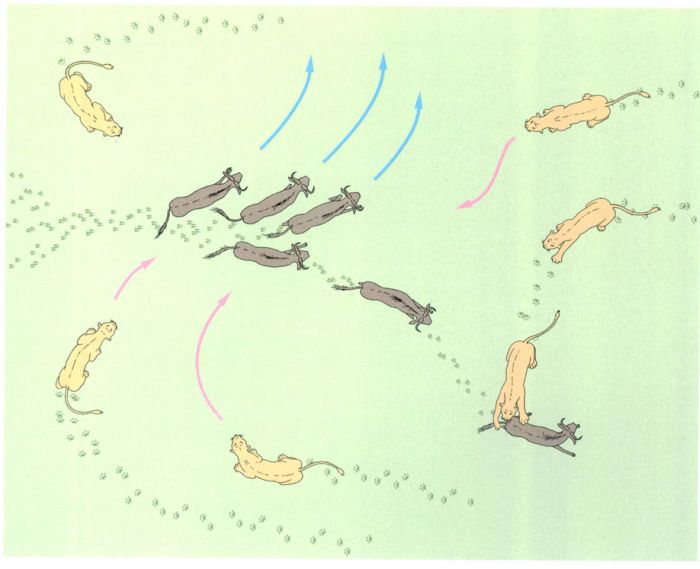
Jordi Corbera, a partir de fonts diverses
L’estratègia de caça de les lleones mostra el grau de sociabilitat d’aquest gran carnívor. Tot i que una lleona pot arribar als 55 km/h, no és capaç de mantenir aquesta velocitat durant un temps gaire llarg, de manera que li és més fàcil aconseguir un bon àpat si coopera amb les seves companyes. Un cop han localitzat les preses potencials (en el cas del dibuix, un ramat de nyus), les lleones es despleguen en ventall i les encerclen. Aleshores cada lleona tria la seva víctima, generalment l’animal que té més a prop. Els nyus, empesos per la por, es dispersen, i sempre n’hi ha un que acaba dirigint-se cap a una de les lleones que l’espera a l’aguait, a punt per saltar-li a la gropa. Un cop ha fet caure l’animal a terra, la lleona li clava una bona mossegada al coll o al morro i manté la pressió durant una breu estona. En no poder respirar, la presa es desploma gairebé sense debatre’s. Bé que són les lleones les que s’encarreguen totes soles de la caça, són els mascles els primers a menjar i, per tant, els que s’emporten les parts més bones. Fins que no s’han atipat, no donen permís a les femelles per alimentar-se. Si hi ha prou carn, els cadells podran menjar al costat de les seves mares, però si l’aliment escasseja, són els últims a tenir accés a la presa, i no sempre aconsegueixen apaivagar la gana.
La cooperació entre espècies
Sovint, igual com passa a la selva, es desenvolupa una cooperació entre diferents espècies. Ja s’ha fet esment dels fenòmens de simbiosi alimentària que permeten als herbívors utilitzar la cel·lulosa gràcies als microorganismes, però hi ha també associacions entre moltes espècies de vertebrats. Els ramats d’ungulats, per exemple, sovint són formats per diferents espècies com nyus, zebres i, de vegades, altres antílops o, fins i tot, estruços. Hi ha també, entre els babuïns i els ungulats, una associació freqüent que afavoreix la vigilància dels depredadors. L’olfacte molt ben desenvolupat dels ungulats unit a la vista excel·lent i a la possibilitat d’estar a l’aguait posant-se dalt d’un arbre dels babuïns, incrementen molt la seguretat.
Aquesta funció d’aguait també és garantida pels ocells que acompanyen sovint els ramats. El més conegut és l’esplugabous (Bubulcus ibis): aquest bell ocell blanc s’aprofita de l’agitació dels insectes, concretament dels ortòpters, que provoca la marxa dels grans ungulats, per a descobrir-los i menjar-se’ls. Una funció d’alarma semblant ha estat atribuïda als esplugabúfals (Buphagus), sempre presents a l’esquena de búfals i rinoceronts.
En conjunt, encara que la vida en grup també es dona en els animals selvàtics, molt especialment en els simis, està força més desenvolupada en les espècies que habiten a la sabana. Facilitada per la possibilitat de mantenir el contacte visual –cosa difícil a la selva, on predominen les relacions per senyals sonors–, assegura als individus una seguretat més gran enfront els depredadors. Recíprocament, de tota manera, la vida en grup també permet a aquests últims una eficàcia més gran en la caça.
La vida sota terra: els caus
Ja s’ha destacat la importància que té la vida en el sòl per a la fauna de sabana. La primera causa és que s’hi localitza una fracció important de la producció vegetal, superior a la de la vegetació epigea. La segona és l’existència de condicions menys extremes de temperatura i d’humitat, especialment apreciables a la sabana seca.
Els principals beneficiaris d’aquestes condicions són, en primer lloc, els insectes socials, formigues i sobretot tèrmits. Quan la humitat es manté al sòl durant un temps suficient, els cucs de terra són també una fracció essencial del poblament, però són eliminats per un clima massa sec. Aleshores són els tèrmits que esdevenen els animals dominants del sòl. La seva presència sol traduir-se en un gran nombre de construccions epigees que prolonguen el domini endogen i asseguren el manteniment de condicions de medis favorables fins i tot al punt més extrem de l’estació seca. Sembla, d’altra banda, que algunes espècies de tèrmits aconsegueixen baixar fins a fondàries de desenes de metres per trobar l’aigua que els és necessària. Els tèrmits dominen la fauna dels sòls de les sabanes seques, però també són presents a les humides. La categoria dels tèrmits humívors, que s’alimenten selectivament de les petites arrels en descomposició i no del sòl agafat sense triar, té més importància a les sabanes humides, mentre que a les seques predominen les espècies consumidores de gramínies, l’alimentació de les quals està amb relació al cultiu simbiòtic de fongs.
L’abundància dels tèrmits fa que s’iniciïn xarxes tròfiques en les quals intervenen, en particular, diferents espècies de formigues, com també nombrosos ocells en el moment de la dispersió dels individus sexuats. Diversos mamífers són també depredadors de tèrmits i adaptats a aquest règim. El més característic a l’Àfrica és el porc formiguer (Orycteropus afer), estès per tot el continent, però també el pangolí gegant (Manis gigantea), present a l’W africà, del Senegal, a ponent, fins a Uganda, a llevant i altres congèneres. Una mena de hiena, l’“aardwolf” o pròteles (Proteles cristatus), també és termitòfaga i consumeix especialment grans quantitats de Trinervitermes quan aquests tèrmits es desplacen de nit en busca d’herba per tallar. Un paper semblant fan, a les sabanes americanes, tatús i óssos formiguers.
A més d’aquests habitants permanents i els seus depredadors, el sòl també serveix de recer temporal, durant la nit o el dia segons els casos, per a moltes espècies epigees a les quals l’absència o l’escassetat d’arbres no permet trobar recer en un altre lloc. La majoria dels rosegadors, per exemple, excaven caus, fins i tot els de mida gran com la rata de Gàmbia (Cricetomys gambianus), la rata de canyar de Swinder o “aulacodi” (Thryonomys swinderianus), els esquirols terrestres (Xerus) i el porc espí (Hystrix cristata). També ho fan altres espècies, de mida encara més gran: a més dels porcs formiguers (Orycteropus afer), pangolins (Manis) i pròteles o “aardwolf” (Proteles cristatus), que sovint aprofiten termiters que prèviament han buidat; hi ha els senglars de riu (Potamochoerus porcus) i, sobretot, carnívors com les hienes o les mangostes, a més d’insectívors i macroscèlids.
Les adaptacions demogràfiques
L’adaptació de les espècies a les exigències de la vida a la sabana, especialment a les climàtiques i a les repercussions en la vegetació, no es basa únicament en els trets morfològics i fisiològics individuals o en un comportament social desenvolupat. Afecta també les diverses característiques dels seus cicles estacionals, és a dir, dels seus paràmetres demogràfics.
Entre els animals que viuen molts anys, una manera d’evitar les dificultats de l’estació seca és migrar a un altre territori més clement. Per poder migrar o restar al lloc, els mamífers i els ocells normalment ajusten el cicle reproductiu al cicle estacional. Els naixements i les eclosions es produeixen durant el període més favorable per a l’alimentació de les cries, és a dir, quan arriben les primeres pluges. En els cas dels ungulats la mare disposa aleshores d’una herba rica que afavoreix la lactància i, de seguida, el deslletament. Per als ocells, és el període en què els insectes són abundants i això els permet d’alimentar fàcilment els pollets.
L’adaptació demogràfica de les espècies de vida curta, com molts invertebrats i petits vertebrats, és d’un tipus molt diferent. El període d’activitat dels animals és limitat a una única estació de les pluges i la supervivència durant l’estació seca s’assegura de diferents maneres. Una de les més corrents és la persistència en estat d’ou. Molts insectes tenen aquesta adaptació, però també alguns rèptils: els ous es ponen al sòl, al començament de l’estació seca, i només eclosionen quan arriba el període de pluges, mentre que els adults moren poc després de la posta. Aquest cicle els permet, a més, evitar la destrucció pel foc. Aquest és el cas, a la sabana de Lamto, del llangardaix Mabuya buettneri del qual no es troba cap adult a la sabana quan aquesta s’incendia al gener. Les seves postes es troben en aquell moment profundament enterrades sota terra i eclosionaran amb l’arribada de les primeres pluges. L’animal només viu, així, una desena de mesos en total.
Hi ha també espècies xeròfiles per a les quals l’estació de les pluges és l’estació desfavorable. A la sabana d’altitud de Nimba, per exemple, la majoria de les llagostes (acrídids), que són animals heliòfils, es desenvolupen i es reprodueixen durant l’estació seca, a diferència de la majoria dels altres artròpodes. Hi ha altres casos curiosos, com el d’alguns gasteròpodes de closca molt reduïda (Gymnarion) de la sabana d’altitud del Nimba. Els ous es ponen cap a la meitat de l’estació de les pluges i el desenvolupament comença tot seguit. Quan arriba l’estació seca, els individus joves es fixen sobre una pedra i passen a un estat de vida ralentit, protegits per la closca que s’aferra fortament al substrat i per tant perfectament aïllades de l’atmosfera molt seca que hi ha a l’exterior. Reprenen l’activitat quan comencen les pluges de l’any següent, no sense que hi hagi hagut, tanmateix, una elevada mortalitat. El creixement és aleshores molt ràpid i esdevenen adults en menys d’un mes, cap a la meitat de l’estació de les pluges, quan cadascun pon moltes desenes d’ous que comencen el seu desenvolupament tot seguit. Quan arriba la següent època eixuta, els adults han crescut més que no pas la seva closca, que és molt rudimentària, i per tant no es poden aixoplugar al seu interior. Per això moren tots els adults, de manera que només els joves aconsegueixen sobreviure i continuar així el cicle vital de l’espècie.
Un exemple de coevolució: escarabats i grans herbívors
Quan hom pensa en evolució, acostuma a considerar el procés que té lloc dins d’una població, d’una espècie, entre organismes que mantenen l’intercanvi genètic i que, per tant, tenen una herència comuna. La variació i la seva herència són els mecanismes mitjançant els quals es presenten i transmeten les diferències que són la primera matèria de l’evolució. Però hi ha un altre aspecte fonamental del concepte darwinià d’evolució biològica: la selecció. Aquest aspecte depèn del medi ambient: en l’expressió de cada caràcter, les manifestacions més adequades per a sobreviure en un determinat medi són afavorides. Els individus que les mostren tenen més possibilitats d’arribar a l’edat reproductora, i per tant de multiplicar-se, que aquells que desenvolupen caràcters diferents. En el transcurs del temps tendeixen a predominar les manifestacions més favorables i a canviar les característiques de l’espècie. Tot i que l’evolució no succeeix sempre de la manera abans assenyalada, aquesta explicació darwiniana (seria millor dir neodarwiniana) continua essent la base per a comprendre l’evolució de la majoria de les espècies de plantes i animals.
L’explicació darwiniana implica una interacció contínua amb el medi, gran modelador del procés evolutiu. Però el medi ambient és molt ampli. Algunes de les seves característiques o components biològics són molt més importants que d’altres per a determinades espècies, o per al conjunt d’organismes: la comunitat. Així succeeix que un organisme és afavorit en la seva supervivència per l’activitat d’un altre organisme, que al seu torn obté avantatges de l’existència del primer. Aquests dos organismes o grups d’organismes tindran processos evolutius que s’influeixen recíprocament. Això és el que s’anomena coevolució. Hi ha exemples que il·lustren bé aquest fenomen, per exemple flors que només són pol·linitzades per un determinat insecte i en les quals evolutivament apareixen uns caràcters (forma, color) que asseguren l’atracció d’aquest insecte. Alhora, l’insecte troba avantatges en la flor i al llarg del temps desenvolupa peces bucals de forma adequada i pautes de comportament que li permeten detectar les flors. Aquests ajustos succeeixen al llarg del temps, de generacions, d’èxits (que sobreviuen) i fracassos (que desapareixen). Però l’evolució no acostuma a tenir pressa.
Un dels casos més interessants de coevolució és el que correspon als grans mamífers herbívors i els escarabats buiners (coleòpters de la subfamília dels escarabeïns). Els grans mamífers herbívors produeixen una quantitat impressionant d’excrements que és utilitzada per a l’alimentació i la nidificació (aliment de les larves) dels escarabats. Els escarabats depenen fins a tal punt dels excrements que han desenvolupat adaptacions morfològiques i complicats comportaments per manejar-lo amb eficiència. Sense els escarabats, el fem resta en superfície, els seus elements nutritius no s’integren al sòl, o ho fan molt lentament, i el que és encara més greu, les buines seques constitueixen un obstacle físic per al desenvolupament de les plantes que mengen els grans herbívors. Sense mamífers no hi ha escarabats, però sense escarabats les sabanes que sustenten els grans ramats (o les pastures on pasturen ramats de bovins) no mantenen la seva productivitat.
Aquesta relació de mútua dependència ecològica és present en totes les zones tropicals i temperades del món, però és més notable a les sabanes africanes: l’ecosistema que ha conservat fins pràcticament els nostres dies el nombre més elevat de grans mamífers. Amb formes diferents —fins i tot contràries— de com es presenta a les sabanes africanes, la relació mamífers-escarabats és igualment evident en les selves del tròpic americà, on el nombre de grans mamífers és molt menor (hi ha moltes espècies, però predominen els rosegadors i els rats-penats), cosa que ha obligat els escarabats a desenvolupar estratègies d’alimentació alternatives. A Austràlia no existien mamífers placentaris, per això la fauna d’escarabeïns, que era escassa, estava adaptada a les boletes produïdes per cangurs i altres marsupials. Però quan els europeus introduïren el bestiar boví es va plantejar un terrible problema, ja que els excrements no eren atacats pels escarabats aborígens. Això obligà les autoritats australianes a desenvolupar un complex i costós programa d’introducció d’escarabats africans i mediterranis.
La rica fauna de grans mamífers herbívors de les sabanes africanes va acompanyada d’una riquesa d’escarabats sense equivalent. Les sabanes africanes contenen 1 500 espècies d’escarabeïns, aproximadament una tercera part del total mundial. Per altra banda, a les sabanes africanes no sols hi ha moltes espècies com a ecosistema, també n’hi ha moltes dins d’una determinada sabana: fins a 150 en aquelles que encara conserven elefants, enfront d’un màxim de 50 espècies de les selves americanes, el segon tipus d’ecosistema en riquesa d’espècies d’escarabeïns. I també hi ha moltes espècies (i moltíssims individus) en cada buina: fins a 40 espècies i molts centenars d’individus en una buina d’elefant. S’ha arribat a estimar que, a les sabanes de l’Àfrica occidental, els escarabats enterren una tona mètrica d’excrement per any i per hectàrea.
Per tal de poder coexistir tantes espècies semblants en una mateixa àrea i a partir d’un mateix recurs (les buines dels grans mamífers), els escarabats han desenvolupat una diversitat de respostes. Des de les anatòmiques (espècies més grans o més petites que exploten l’excrement de manera diferent, potes adaptades a fer rodolar una bola o a excavar en profunditat) fins a les etològiques o de comportament, en les quals les diferències són més variades i sofisticades. Així, hi ha espècies que són actives durant les hores del dia, d’altres ho són al crepuscle, d’altres durant la nit. Al llarg de l’any, els pics d’abundància de les diferents espècies no acostumen a ser sincrònics. Aquest repartiment de la dimensió temps al llarg del dia, o al llarg de l’any, redueix la competència directa. Una altra manera de reduir la competència és l’especialització alimentària. Tot i que a les sabanes africanes dominen els copròfags generalistes, és a dir, escarabats que ataquen els excrements de diferents grans herbívors, existeixen molts especialistes. Les enormes buines dels elefants, amb moltes fibres no digerides, són l’aliment i el material de nidificació de més d’un centenar d’espècies que les utilitzen prioritàriament. Entre elles, els grans excavadors del gènere Heliocopris (els escarabats més grans de la subfamília escarabeïns), i un piloter força gros, Kheper platynotus.
La distribució d’activitats al llarg del temps i la separació d’algunes espècies com a especialistes alimentaris no és suficient per a compensar l’agregació d’escarabats a cada buina. Aquesta agregació es converteix en un factor limitador si es considera la nidificació. L’excrement no es troba uniformement distribuït, sinó que presenta una agregació en boles o buines. Això provoca que la distribució d’escarabats per unitat de superfície sigui molt desigual. D’espais on no n’hi ha cap, a la buina on se’n poden acumular centenars. Si es té en compte que els escarabats fan els nius a terra, acumulant per a l’alimentació de la larva una certa quantitat d’excrement, resulta totalment impossible que tots els escarabats que arriben a una buina cavin galeries i facin els nius a sota. Resoldre el problema ecològic de disposar d’un aliment que es troba concentrat en certs punts i que a més és efímer, és a dir, que resta poques hores o dies amb possibilitats de ser utilitzat, ha estat el gran repte que ha marcat l’evolució dels escarabeïns. Resoldre’l els ha permès el domini i la riquesa que avui presenten. La solució ha estat la relocalització de l’aliment, és a dir, el canvi de la seva distribució espacial: d’una distribució agregada (concentrada) a una altra de dispersa.
La relocalització es fa tant amb la porció d’aliment que menjarà l’adult com amb l’aliment per a les larves (nidificació). És en la nidificació, és a dir, en la tasca de separar, distanciar i protegir una porció d’aliment que permeti el desenvolupament de la nova generació, d’ou a pupa, on l’evolució del comportament assoleix els seus extrems en els escarabeïns. La relocalització pot ser horitzontal o vertical. L’horitzontal es realitza tallant i modelant una bola i enduent-se-la rodolant (d’aquí els noms d’escarabat piloter, boller o rodador), per després enterrar-la. La vertical fent galeries de forma, calibre i profunditats diferents (segons les espècies) en les quals es protegeix una porció d’excrements.
A les sabanes tropicals d’Amèrica, per exemple als “llanos” de Veneçuela, no es presenta aquest moviment. Uns sòls pobres sostenen pastures molt més raquítiques que les africanes. No hi ha abundància de grans mamífers herbívors (als “llanos”, els grans consumidors d’aliment vegetal són les formigues talladores de fulles) i el mamífer autòcton gran més abundant és la capibara (Hydrochaeris hydrochaeris), un rosegador. Tampoc no hi ha una gran producció d’excrement. Els escarabats existeixen, però són molt menys diversificats en espècies i menys nombrosos que a les sabanes africanes.
Els factors biogeogràfics i la fauna sabanícola africana
La principal dificultat que hom troba en intentar descriure el poblament animal dels immensos espais compresos en el bioma de les sabanes és la seva gran diversitat, paral·lela a la dels hàbitats que inclou. El conjunt de tots els factors del medi físic i de la vegetació influeix lògicament en el poblament animal i encara són molt insuficients els estudis que s’han fet per tenir-ne un coneixement exacte, per altra banda difícil d’assolir, tant perquè exigeix un treball de camp considerable de recol·lecció i d’identificació, com també per la manca d’especialistes, particularment d’invertebrats, que, al capdavall, representen la major part de la fauna. L’acció destructora de les poblacions humanes, que s’ha accelerat les darreres dècades, ha alterat profundament les poblacions, i especialment les de grans mamífers, víctimes de la caça intensa i d’una modificació important del seu hàbitat. Només alguns parcs nacionals i algunes reserves permeten encara tenir una idea del que foren aquestes poblacions al començament del segle XX.
La diferenciació faunística de la sabana africana
La distància entre els diversos territoris, doncs, fins i tot a l’interior d’un conjunt continental provoca un aïllament relatiu. Aquest aïllament sovint s’incrementa per la presència, almenys temporal, de diferents obstacles com pot ser per exemple pels animals de sabana un desert o una selva densa. Mentre aquest aïllament continua, fins i tot sense canvis climàtics importants, es desenvolupen desviacions dels tàxons. Aquestes desviacions poden ser importants en continents tan grans com Àfrica o l’Amèrica del Sud. Per a diferenciar les poblacions també cal tenir en compte les diferències en l’extinció de les espècies, amb relació a les variacions de les condicions ecològiques. El grau de diferenciació taxonòmica lligat a la distància geogràfica i als obstacles ecològics evidentment depèn molt del grup que s’estudiï. És fàcil entendre que sigui baix, per exemple, per als ocells, capaços de recórrer volant centenars de quilòmetres, o per a algunes aranyes que el vent pot transportar a distàncies considerables. La desviació serà gran, en canvi, per a organismes com els cucs de terra o els miriàpodes, que són poc mòbils i molt sensibles a les condicions hídriques del medi.
Amb un territori sotmès a climes tropicals particularment extens i en el qual la sabana és el tipus de vegetació dominant, Àfrica és el continent més adient per a l’anàlisi de la diferenciació de les poblacions pel que fa a l’aïllament causat per la distància. Aquesta qüestió queda il·lustrada per la fauna dels grans mamífers, tot i que, tal com ja s’ha remarcat, aquests animals només subsisteixen de forma més o menys intacta a les reserves i als parcs nacionals com els del riu Comoé, a la Costa d’Ivori, i de Niokolo-Koba, al Senegal, que serviran aquí d’exemples representatius de la fauna de sabana de l’Àfrica occidental en contraposició a la de l’Àfrica oriental. Tots dos parcs donen una imatge relativament fidel de les poblacions de les sabanes de l’W africà.
La fauna africana occidental: els casos del Comoé i Niokolo-Koba
Tant les sabanes del parc nacional del riu Comoé com les del parc nacional de Niokolo-Koba, corresponen a tipus molt diversos, bé que recorden les del Serengueti i d’altres sabanes de l’Àfrica oriental. Situat encara més a l’W, el parc nacional de Niokolo-Koba, al Senegal, pròxim a la frontera guineana, és un altre dels indrets que poden donar una imatge de la fauna primitiva d’aquesta regió occidental de la sabana africana. El fet que aquest territori s’hagi convertit en una reserva ha permès, alhora, la conservació d’una part important de la fauna i la realització d’estudis de prospecció sistemàtics de nombrosos grups d’animals.

Principals espècies de mamífers carnívors i herbívors dels parcs nacionals del riu Comoé, a la Costa d’Ivori, i del Serengueti, a Tanzània. Les sabanes del Serengueti presenten una diversitat d’espècies molt més gran que les de Comoé, sobretot pel que fa a grans herbívors. En canvi, Comoé és molt més ric en mamífers zoòfags, amb excepció de cànids i fèlids, força abundants en ambdós parcs nacionals. Els 27 ungulats (1 proboscidi, 3 perissodàctils, i 23 artiodàctils) i els 9 carnívors que integren la mastofauna del Serengueti es poden prendre com a referència de la mastofauna de les sabanes de l’Àfrica oriental, el poblament animal més espectacular d’aquest bioma.
Dades proporcionades per l’autor
A la zona del riu Comoé, situada al N de la Costa d’Ivori, l’any es divideix en dues estacions contrastades; encara que l’estació seca pot durar vuit mesos, la pluviositat total és elevada i varia, segons els anys, entre els 1 100 i els 1 700 mm. La regió és coberta, principalment, de sabanes llenyoses boscades, arbrades o arbustives, però també de sectors de bosc clar al N i fins i tot bosc sec dens al S, al llarg dels cursos d’aigua. Alguns sectors de la plana al·luvial que voreja el riu Comoé quasi no tenen arbres i només tenen herba baixa. Aquesta diversitat, òbviament, augmenta la riquesa específica, i en particular el poblament de mamífers grans i mitjans. La regió de Niokolo-Koba es caracteritza també per la gran diversitat de medis naturals que s’hi combinen, cosa que fa molt artificial qualsevol intent de definir comunitats vegetals de gran superfície. Hi ha, efectivament, relacionades les unes amb les altres, sabanes més o menys arbòries, gramenets baixos sobre d’una cuirassa ferruginosa, gramenets inundables, boscos clars, boscos en galeria, etc. I, si bé nombrosos organismes de talla petita i poca capacitat de desplaçament es localitzen de manera més o menys estricta dins d’un o un altre d’aquests medis, no és menys veritat que es pot pensar que molts altres organismes, més eclèctics i amb una mobilitat major, també es poden trobar per tot el territori o en una bona part. Això és el que passa amb la majoria dels mamífers grans i amb molts ocells, però també amb nombrosos rèptils, amfibis i insectes.
En tot cas, si bé hi ha diferències amb les poblacions de l’Àfrica oriental, el model de les quals seria Serengueti, aquestes diferències no modifiquen la imatge de conjunt. La similitud trobada entre les poblacions de carnívors, la dispersió dels quals és més fàcil i les exigències ecològiques menys estrictes, és molt gran ja que l’absència del guepard a la zona occidental és recent. Entre els grups d’herbívors, en canvi, es nota l’absència d’algunes espècies característiques i abundants d’Àfrica oriental, les zebres, el nyus i la girafa si bé l’absència d’aquesta darrera és un fenomen secundari causat pel seu extermini recent d’una zona que, possiblement, tampoc no li era gaire favorable.
Com que pel que fa als grans mamífers, l’inventari del parc de Niokolo-Koba i, sens dubte, també el del riu Comoé són més o menys exhaustius, es pot afirmar que, en conjunt, aquestes poblacions de l’W africà són més pobres, sobretot en ungulats, que les de l’Àfrica oriental. Més que el nombre d’espècies, el que és inferior són els respectius efectius. Sens dubte s’hi ha de veure un efecte de la caça i del furtivisme però alguns indicis permeten no excloure una densitat baixa originària, tal com passa amb la sabana de Lamto.
Els poblaments de grans mamífers
Els grans mamífers —és a dir tots els mamífers exceptuant els insectívors, els rosegadors i els quiròpters— són representats, almenys, per una quarantena d’espècies, entre les quals 16 d’ungulats i 5 de primats.
L’elefant africà (Loxodonta africana) i l’hipopòtam (Hippopotamus amphibius), comú a tots els cursos d’aigua de mida suficient, es troben a totes les sabanes africanes i principalment a Gàmbia. Dels dos súids, només el facoquer (Phacochoerus aethiopicus), que és més sabanícola, és abundant mentre que el senglar de riu (Potamochoerus porcus), molt més escàs, només es troba als boscos en galeria i per tant no es pot considerar una forma de la sabana.
El búfal africà (Syncerus caffer) també és comú encara que no gaire abundant. Pel nombre, la població principal d’ungulats és constituïda pels antílops. Alguns són molt escassos, com el gran èland (Taurotragus derbianus) i l’antílop equí (Hippotragus equinus). Altres, en canvi, són més comuns i ho serien encara més si no fos pel furtivisme. Aquestes espècies són: l’antílop jeroglífic (Tragelaphus scriptus), l’oribí (Ourebia ourebi), observat sovint en parella, el duiker de flanc ros (Cephalophus rufilatus), el duiker comú (Sylvicapra grimmia), el redunca comú o “nagor” (Redunca redunca) i el cob defassa (Kobus defassa), el búbal (Alcelaphus buselaphus) i sobretot el cob comú (K. kob), que és l’espècie més abundant i fàcil d’observar; es tracta d’una espècie que cerca, principalment, les gramínies baixes de les sabanes herboses i per això es troba a les zones de la plana al llarg dels cursos d’aigua; així i tot, també freqüenta les sabanes llenyoses i fins i tot els boscos clars quan les seves pastures preferides han desaparegut amb l’estació seca. Els cobs viuen en ramats, a vegades molt nombrosos, i tenen tendència a degradar les zones on es concentren per sobrepasturatge i trepig.
Cal remarcar l’absència de gaseles (Gazella) i d’òrixs (Oryx), formes de zones més àrides, i de la girafa (Giraffa camelopardalis), una espècie que sembla que havia viscut a la regió i que hom ha intentat reintroduir sense èxit al parc nacional de Niokolo-Koba. El sassabi (Damaliscus lunatus korrigum), en altre temps abundant durant l’estació seca, també ha desaparegut.
Els carnívors són representats pel cap baix per 18 espècies. Entre els cànids abundants hi ha el xacal ratllat (Canis adustus) i el licaó (Lycaon pictus). Els vivèrrids són les diferents genetes (Genetta pardina, G. thierryi), la civeta de palmera (Nandinia binotata) i quatre espècies de mangosta, l’icnèumon (Herpestes ichneumon), la mangosta vermella (H. sanguineus), la de cua blanca (Ichneumia albicauda) i la de Gàmbia (Mungos gambianus). La hiena tacada (Crocuta crocuta) és comuna mentre que és dubtosa la presència de la hiena ratllada (Hyaena hyaena). Entre els fèlids es compten el lleó (Panthera leo), el lleopard (P. pardus), el serval (Felis serval) i el caracal (F. caracal), però no el guepard (Acinonyx jubatus).
Entre els primats, el més sabanoide —deixant a banda els humans— és el babuí comú (Papio papio). Absent sempre de les selves denses, busca més aviat sectors rocallosos. És un simi robust, rabassut i de color bru olivaci. Té el musell allargat, cosa que li dona aspecte de gos, d’aquí prové el nom de cinocèfal que també se li ha aplicat. Entre el mascle i la femella hi ha un dimorfisme important pel que fa al pes: el mascle pot pesar més de 30 kg, mentre que la femella no arriba a 10 kg. Els mascles tenen també els ullals més llargs i la crinera més espessa. El seu règim alimentari és omnívor i inclou bulbs, rizomes, llavors i fruits i també insectes i petits vertebrats. L’animal es porta el menjar a la boca amb les mans. La mona vermella o “patas” (Erythrocebus patas) és una espècie estesa a totes les sabanes africanes, des del N de l’equador fins al Sàhara, sempre fora de la selva. Les mones vermelles viuen en grups que poden ser d’una trentena d’individus. La seva alimentació és principalment herbívora però també mengen fruits, com els del tamarinde, llavors i fins i tot bolets o, encara ous, insectes i llangardaixos. Per a satisfer la set generalment en tenen prou amb l’aigua que obtenen de la pluja, de la rosada o fins i tot la que hi ha dins els vegetals, i això els permet viure en zones relativament àrides a les quals no poden anar els babuïns. A diferència dels babuïns i de les mones vermelles, els cercopitecs de cara negra (Cercopithecus aethiops) no són terrícoles. Passen la vida essencialment als arbres, tant de la sabana llenyosa boscada com de l’arbrada. El bosc en galeria, que els permet tenir alhora un refugi i una sabana a prop on completar la seva dieta, és el seu hàbitat predilecte. És en els arbres on troben l’aliment, sobretot, fruits i llavors, però també insectes. Aquest menjar se l’entatxonen a les galtes dilatables, anomenats sacs bucals, i el masteguen quan són a recer. Aquesta tècnica recorda l’emmagatzemament d’herba al rumen dels remugadors. Els ximpanzés, finalment, es troben en aquesta zona al límit septentrional de la seva àrea de distribució i per tant són poc nombrosos.
El pangolí gegant (Manis gigantea) i el porc formiguer o “aardvark” (Orycteropus afer) són dos consumidors de formigues i tèrmits més aviat escassos.
Els poblaments d’ocells
La població d’ocells és rica no sols en espècies sinó també en efectius. En total s’han assenyalat més de 320 espècies, pertanyents a 17 ordres diferents. Entre elles, alguns pelecaniformes, com el corb marí de cua llarga (Phalacrocorax africanus), l’anhinga africana (Anhinga rufa) i el pelicà blanc (Pelecanus onocrotalus) estan vinculats al medi aquàtic i, en el parc de Niokolo-Koba, viuen només a la vora del riu Gàmbia. La gallina de Guinea comuna (Numida meleagris) i el francolí comú (Francolinus bicalcaratus) són dues espècies de galliformes molt abundants.
Els rapinyaires són nombrosos i són representats per 25 falconiformes, en particular l’àguila marcenca (Circaetus gallicus), l’àguila saltimbanqui (Terathopius ecaudatus), l’àguila pescadora africana (Haliaeetus vocifer) i sis o set espècies de voltors, com també per set estrigiformes, entre els quals el duc africà (Bubo africanus) i el duc gegant (B. lacteus).
Hi ha dos psitaciformes abundants, el lloro del Senegal (Poicephalus senegalus) i la cotorra de cua llarga o de Kramer (Psittacula krameri), com també 7 espècies de columbiformes (gangues, coloms i tórtores), 10 de cuculiformes (cucuts, cucals i turacs), 9 de piciformes i 22 de coracidiformes entre les quals hi ha 3 gaigs, 5 abellerols, 3 càlaos i 8 blauets. Els passeriformes són l’ordre més ben representat, amb més de 100 espècies, de les quals 23 són de sílvids i 13 plocèids (teixidors i vídues).
Els poblaments de rèptils i d’amfibis
Els rèptils són nombrosos i representats per una trentena d’espècies, entre les quals una vintena d’espècies de serps. Al parc del riu Comoé s’han citat el varà de terra (Varanus exanthematicus), l’agama (Agama agama), el camaleó del Senegal (Chamaeleo senegalensis). D’entre les serps cal esmentar els pitons (Python sebae i P. regius), la cobra escopidora (Naja nigricollis), la mamba verda occidental (Dendroaspis viridis), la mamba negra (D. polylepis) i quatre escurçons: el bufador (Bitis arietans), Echis carinatus, Atractaspis dahomeyensis i Causus maculatus, que és el més comú.
Els amfibis són sobretot espècies adaptades als rigors d’una estació seca. El gripau Bufo regularis, molt terrícola igual que l’espècie excavadora Hemisus marmoratus, és el més estès. Una forma subaquàtica present a tot Àfrica és Discoglossus occipitalis. Altres amfibis són arborícoles com Leptopelis viridis, Hyperolius nitidulus o Afrixalus fulvovittatus. Moltes espècies de Ptychadaena són comunes a les vores dels cursos d’aigua.
Els poblaments d’invertebrats
Les sabanes de Niokolo-Koba són dels pocs llocs d’Àfrica on s’han dut a terme recerques sobre el conjunt de la fauna d’invertebrats i en particular de la d’insectes. Els estudis realitzats mostren la gran riquesa d’aquestes poblacions. De tota aquesta fauna, la població de tèrmits mereix una especial atenció pel seu gran nombre d’individus i també per la importància que tenen en el paisatge els grans termiters en pinacle construïts per Macrotermes bellicosus. Els termiters fungiformes de Cubitermes es desenvolupen sobre les superfícies nues de les cuirasses. Els altres grups d’insectes dominants pel nombre d’individus són les formigues, els acrídids, nombrosos i variats, els lepidòpters i els dípters, com també els coleòpters copròfags. Els aràcnids també són freqüents, amb aranyes, escorpins, solífugs i àcars, mentre que els miriàpodes són representats per milpeus (júlides i polidesmides), escolopendres (escolopendromorfs), etc.
La gran fauna africana oriental: el cas del Serengueti
Totes les zones de l’Àfrica oriental amb una pluviositat compresa entre 700 i 1 200 mm anuals estan cobertes de formacions vegetals que corresponen perfectament a la denominació clàssica de sabana. Abans de les matances que començaren al final del segle XIX, les poblacions de grans mamífers eren extraodinàriament abundants i tota la fauna era, en general, rica. Actualment aquesta fauna ha desaparegut d’una gran part de la regió i només alguns territoris, transformats en parcs nacionals, contenen encara mostres relativament representatives de les poblacions originals.
Les sabanes del Serengueti són situades a l’E de l’Àfrica central, just per sota de l’equador, entre les latituds 1°15’ i 3°30’ S. La característica més notable del clima és la seva irregularitat en l’espai i en particular la considerable variació que afecta el règim de pluges en menys de 200 km. El total de precipitacions oscil·la des dels 900 mm anuals que cauen al N i a l’W fins als 500 mm que es comptabilitzen a la zona meridional. Paral·lelament, l’estació seca esdevé més i més rigorosa, ja que les precipitacions disminueixen de 300 mm a menys de 100 mm, i la variabilitat interanual augmenta del 25% al 100%. En resposta a aquesta heterogeneïtat climàtica i a les irregularitats del relleu i dels sòls, les comunitats vegetals també són variades. Un dels seus trets principals és la presència de grans extensions planes de sabanes amb herba baixa en alguns sectors i més alta i boscosa en altres, de sectors rocallosos amb bardisses, de boscos en galeria i de bosquets dispersos.
La fauna d’invertebrats de les sabanes del Serengueti no ha estat objecte d’un estudi detallat. En canvi, el parc inclou, en unes condicions sens dubte pròximes al seu estat original, una població molt rica de grans mamífers que ha estat estudiada acuradament. Aquesta riquesa es fa palesa en la presència de 30 espècies d’ungulats i 13 de grans carnívors, a les quals s’afegeixen molts rosegadors i insectívors. Una primera característica que destaca de l’estudi dels animals de les sabanes del Serengueti és la poca importància quantitativa dels carnívors amb relació als herbívors, dels quals, per tant, no poden controlar l’expansió. Pel que fa als herbívors, cal destacar que hi ha una predominància considerable dels grans mamífers respecte dels petits (275 kg/ha enfront d’1 kg/ha) igual que respecte els acrídids (2 kg/ha). La resta de grups tenen una representació força limitada a la biomassa total, a excepció dels ocells, que són representats per nombroses espècies i es pot dir, malgrat no haver-se fet cap estimació precisa, que representen molts quilos per hectàrea, i, principalment l’estruç, amb valors d’1 kg/ha.
S’ha provat de determinar el consum de les diferents categories d’herbívors en les tres principals fàcies de vegetació: la sabana llenyosa d’herba alta, la sabana herbosa qualificada de pastura i el medi, molt peculiar, dels afloraments rocallosos dispersos a la regió anomenada dels “kopjes”. També cal subratllar l’existència de variacions importants dels nombres d’individus de cada espècie al llarg dels anys; així, per exemple, el nombre de nyus o el de búfals pot variar fàcilment del senzill al triple en el termini de pocs anys; mentre que el 1961 s’havien observat uns efectius de 263 000 nyus blaus (Connochaetes taurinus) i 29 500 búfals africans (Syncerus caffer), deu anys més tard eren respectivament 693 000 i 58 000 i, només en un any, passaren a ser 840 000 i 75 000, repectivament.
En conjunt, les sabanes del Serengueti donen una visió clara dels elements principals del poblament animal en allò que fa referència a les espècies de mida mitjana o gran, aquí molt àmpliament dominants. Els estudis més profunds referents als petits vertebrats i al conjunt dels invertebrats s’han realitzat a la sabana de Lamto i a la Costa d’Ivori.
Els grans herbívors
L’elefant africà (Loxodonta africana) és massa conegut per merèixer una extensa descripció. Els elefants viuen en grups familiars d’una femella i els seus petits, però aquests grups sovint es reuneixen en ramats més o menys grans que, de vegades, poden tenir diversos centenars d’individus. Els mascles joves deixen el grup familiar quan arriben a la pubertat —cap als 15 anys— i s’uneixen a altres mascles de la seva edat. El problema més gran d’aquests ramats és la recerca d’aliment i d’aigua, fet que comporta un cert vagareig. També és prou coneguda l’afició dels elefants al bany. Un aspecte important de la vida social de l’elefant és el prolongat aprenentatge dels joves al llarg d’un bon nombre d’anys. Aquest aprenentatge i la seva longevitat —més de 60 anys— contribueix, evidentment, a desenvolupar la intel·ligència dels elefants. La seva gran talla els protegeix de l’atac de les grans feres, fins i tot del lleó (Panthera leo). Tenint en compte aquesta absència d’enemics naturals, els elefants es multiplicarien si els humans no els matessin. La seva superabundància té efectes nocius notables sobre la vegetació. Els elefants destrueixen els arbres i transformen les sabanes en estepes molt degradades, tal com s’ha constatat en algunes reserves on abunden, com a la plana de Semliki.
El rinoceront negre (Diceros bicornis), que actualment viu en una àrea molt reduïda, a causa de la cacera massiva que ha sofert i del furtivisme que encara avui en dia el sotja, és un animal molt estrany que sembla sortit d’una col·lecció de fòssils. Dos ulls petits, dues grans banyes sobre el front, unes orelles acabades en una tofa de pèls i una coloració fosca són els seus caràcters distintius. Un llavi superior acabat en punta, molt mòbil, que li permet tastar les fulles d’arbustos i les branques de què s’alimenta, sobrepuja la boca. És un folívor brostador que aprecia particularment les fulles d’acàcia. El rinoceront negre generalment és solitari des del moment que arriba a l’edat adulta. La seva talla el protegeix dels depredadors, excepte dels humans, que se senten atrets per les virtuts afrodisíaques que s’atribueixen a la pols extreta de les seves banyes. Sovint es troba acompanyat d’esplugabúfals (Buphagus), que s’alimenten dels seus paràsits i, a més, l’alerten de possibles perills. El rinoceront blanc (Ceratotherium simum), que també té dues banyes al cap, pertany a un gènere diferent. Tanmateix, ambdues espècies presenten una convergència de forma, bé que no d’hàbits. Les respectives boques, justament, difereixen força, adaptades com estan a règims alimentaris també força diferents. El rinoceront blanc, en efecte, és un pasturador que s’alimenta d’herba curta i no és, com el negre, un brostejador de fulles.
Les zebres, i aquí, més especialment, la zebra de Burchell (Equus burchelli), tenen la morfologia general d’un cavall, amb una gropa robusta. Són conegudes sobretot pel seu pelatge ratllat, que alterna bandes horitzontals fosques i clares a les extremitats i fins als unglots, perpendiculars a l’eix del cos sobre els flancs i l’esquena, i més fines i més irregulars al cap i a les galtes. Són animals robustos i ràpids, amb un pes que pot superar els 300 kg. Viuen en ramats a les sabanes obertes on l’herba no és gaire alta, i la seva activitat és diürna. Les zebres viuen en grups familiars petits que comprenen un mascle, algunes femelles i els seus poltres i, de vegades, alguns mascles joves. La gestació dura una mica més d’un any. Sociables, sovint formen grups mixtos amb altres espècies. Els seus principals enemics són el lleó (Panthera leo), el licaó (Lycaon pictus) i la hiena tacada ( Crocuta crocuta). Els poltres són evidentment els principals amenaçats, fins i tot pels guepards (Acinonyx jubatus).
L’hipopòtam (Hippopotamus amphibius) també s’alimenta d’herba curta de la sabana. Per alimentar-se, de nit surt del riu o de la bassa on roman tot el dia i pot recórrer molts quilòmetres abans de trobar el terreny més adequat. No menja pràcticament mai dins de l’aigua. Tot i que el seu cos és molt pesant —el mascle pot arribar als 3 000 kg i la femella als 2 000 kg— i que les seves extremitats són molt curtes, pot córrer ràpidament quan se sent amenaçat o atacat. Un hipopòtam adult és massa voluminós per ser atacat pels depredadors, encara que no és impossible que un jove, si se’l sorprèn sol, pugui ser devorat per un lleó. Per això, la densitat de les seves poblacions assoleix, a les zones protegides, valors elevats. Els mascles grans són fàcilment agressius i poden atacar les petites embarcacions. En el moment del naixement, que es produeix al cap de 240 dies de gestació, les cries fan entre 40 i 50 kg de pes. Sembla que assoleixen la maduresa als 4 anys.
El facoquer (Phacochoerus aethiopicus) és un súid de talla gran, amb el cap voluminós i dos parells d’ullals allargats que formen unes llargues defenses, de vegades de més de 50 cm. El pes dels mascles pot superar els 120 kg i el de les femelles els 70 kg. El pelatge és reduït, fora d’una crinera que recorre des del front fins al mig de l’esquena. És un animal sedentari, estès a totes les sabanes africanes. S’alimenta d’herba tendra, rizomes, tubercles, escorces, fruits, però també, si té ocasió, de petits animals. A excepció dels vells mascles solitaris, l’estructura habitual és la de grups familiars susceptibles de reunir-se en colles nombroses. Les ventrades són de dues a quatre cries que, quan neixen, no pesen gaire més d’un quilo. El facoquer té com a depredadors habituals el lleó (Panthera leo) i el lleopard (P. pardus).

Les variacions en les taques del pelatge de les girafes (Giraffa camelopardalis) i de les zebres (Equus) no estan explicades del tot, però ben segur que tenen un valor adaptatiu. A més a més, presenten també una variació individualment, de manera que els serveixen per a reconèixer-se, ja que no canvien al llarg de la vida de l’animal. Les taques de les diferents subspècies de girafa varien molt tant en color com en mida i forma. En Giraffa camelopardalis reticulata, per exemple, són grans, regulars i són separades per una fina línia blanca, mentre que en G. c. tippelskirchi són més petites i de contorns irregulars. G. c. camelopardalis presenta un patró intermedi entre els dos anteriors. Tanmateix, no es donen tan sols tres o quatre patrons concrets, sinó que al llarg de tota l’àrea de distribució d’aquest artiodàctil s’observa una àmplia gamma de variació. Les diferents espècies de zebres també presenten pelatges diferents. A part un sistema de reconeixement individual, sembla que les ratlles del llom d’aquest èquid podrien tenir també altres funcions. Hi ha autors que creuen que creen un efecte desconcertant sobre els depredadors, mentre que altres opinen que simplement serveixen per a crear un efecte tranquil·litzant sobre les mateixes zebres, que se senten atretes per aquest particular dibuix. Això ajudaria a mantenir el ramat més unit, cosa que no deixa de ser una protecció davant dels depredadors. Les explicacions sobre el valor adaptatiu de les ratlles de les zebres, però, no s’acaben aquí. Encara altres autors opinen que representen una defensa contra la mosca tse-tse i consegüentment contra la nagana. Les ratlles del pelatge de la zebra confondrien aquest insecte, que detecta visualment les seves preses com un objecte fosc que es mou a poc a poc i es destaca contra el fons de la sabana.
Jordi Corbera
La girafa (Giraffa camelopardalis) és coneguda pel seu coll desmesurat, tant, que el cap dels mascles es pot situar a 6 m d’alçada i el de les femelles a 5 m. Tot i aquesta llargada, té només 7 vèrtebres cervicals, com la majoria dels mamífers. El seu sistema circulatori està adaptat per resoldre el problema de fer arribar la sang al cap gràcies a un cor excepcionalment poderós, que pesa prop de 10 kg. Una adequada regulació de la pressió en els vasos del cervell li permet controlar la circulació tant si el cap està arran de terra per beure com si es troba a 5 m d’alçada per menjar. Gràcies al seu llarg coll, la girafa té accés a una alimentació a la qual els altres mamífers herbívors no poden accedir. S’alimenta de fulles d’acàcia que agafa amb la seva llarga llengua (la longitud d’aquesta fora de la boca pot assolir 50 cm) sense témer ni les dures espines ni les formigues del gènere Crematogaster, agressives i verinoses, que viuen en aquests arbres. La girafa habita preferentment a les sabanes humides amb herba alta. Deixant de banda els humans, el seu principal enemic és el lleó, però aquest no l’ataca mai quan està sol. La girafa, d’altra banda, sap defensar-se enèrgicament amb els seus unglots, a cop de temibles guitzes.
L’èland comú o èland del Cap (Taurotragus oryx) és molt gran, els mascles més grossos poden arribar als 1 000 kg i les femelles als 600 kg. El seu pèl és bru amb algunes ratlles fines verticals de color blanc, més clares cap al coll, sota del qual hi ha una mena de bossa que penja. L’èland es troba fins al Senegal, però amb una àrea de distribució àmpliament interrompuda al Sudan. Per a alguns autors, la forma de l’Àfrica occidental, sovint anomenada èland de Derby i que es diferencia per la coloració, la grandària i la forma de les banyes, és considerada com una espècie diferent, el gran èland (Taurotragus derbianus). És una espècie que durant l’estació seca es desplaça molt a la recerca de menjar i d’aigua. L’èland pastura i alhora és brostejador, el seu règim alimentari depèn de l’estació i del biòtop. La maduresa sembla tardana i la longevitat pot superar els 20 anys. Els ramats són de mida variable i s’han reduït considerablement, igual que el nombre d’individus de l’espècie, massa perseguida. Com per a tots els antílops, els enemics són principalment el lleó (Panthera leo), el lleopard (P. pardus), el licaó (Lycaon pictus) i la hiena tacada (Crocuta crocuta).
L’antílop jeroglífic, “guib” o “bushbuck” (Tragelaphus scriptus), té a l’Àfrica una vasta àrea de distribució, ja que freqüenta les sabanes d’herba alta però també els boscos en galeria i les selves secundàries. És un antílop amb un aspecte que recorda un cabirol, amb un pes comprès entre 30 i 80 kg. El color del seu pèl és variable segons les races locals i només els mascles estan proveïts de banyes. És un brostejador de fulles i borrons, però també pot consumir herbes joves i tendres. El “guib” habita llocs més aviat humits, no té un cicle estacional precís i es pot reproduir al llarg de tot l’any. Viu solitari.
Per la seva forma elegant i la capa de color bru rogenc, els impales (Aepyceros melampus) són dels habitants més destacables de les sabanes arbustives, allà on s’alternen gramenets d’herba baixa i mates més o menys espinoses. Només els mascles tenen banyes, fines i netament anellades. La seva dieta és mixta, tant mengen herbes com fullam, cosa que constitueix una bona adaptació al cicle estacional de la vegetació. Són animals molt gregaris que viuen en ramats nombrosos que apleguen unes desenes de famílies formades cadascuna per un mascle dominant i diverses femelles amb les seves cries. També hi ha ramats de mascles sols. Animals molt àgils, capaços de fer salts que superen els 10 m de llarg i dotats de bona vista, bon olfacte i oïda excel·lent, els impales s’han de defensar de nombrosos enemics. Entre els seus predadors figuren els lleons (Panthera leo), els lleopards (P. pardus), els guepards (Acinonyx jubatus), els licaons ( Lycaon pictus) i la hiena tacada (Crocuta crocuta) i, pel que fa als individus més joves, els xacals (Canis adustus, el xacal ratllat; C. aureus, el xacal comú; C. mesomelas, el xacal de llom negre) i els grans rapinyaires.
L’antílop equí (Hippotragus equinus) és també un antílop molt gran, la seva talla és aproximadament la d’un cavall, amb un pes que pot arribar als 300 kg en el cas dels mascles. Les banyes, presents en ambdós sexes, són força desenvolupades, anellades i recorbades enrere. Viu a diversos tipus de sabanes, més o menys boscades o arbrades, consumint sobretot herba tendra i eventualment fulles d’arbres o d’arbustos. Ha de beure regularment cada dia.
El sassabi o topi (Damaliscus lunatus) és un antílop de 80 a 150 kg de pes, té el pèl rogenc a l’esquena i blanc al ventre i unes banyes ben desenvolupades que sobrepassen els 50 cm. Viu a les sabanes obertes en una àrea de distribució molt gran, que abasta gairebé Àfrica sencera, però molt fragmentada. És un pasturador que s’alimenta principalment d’herba i viu en grups familiars petits que comprenen un mascle i diverses femelles. Les famílies s’apleguen sovint en ramats més importants i s’associen de grat amb altres espècies.
La majoria dels duikers viuen a les selves. Alguns, tanmateix, també es troben a la sabana i més especialment a les sabanes humides d’herba alta, als límits amb la selva. És el cas del duiker de flanc ros (Cephalophus rufilatus), que té el pelatge d’un bru ataronjat tirant cap a ros amb la gorja blanca. Ambdós sexes tenen banyes fines i dretes. El seu pes és, només, d’uns 10 kg. La seva àrea de distribució s’estén des de Guinea i Gàmbia fins al N del Zaire.
El nyu, més concretament el nyu blau (Connochaetes taurinus), és un bòvid de talla gran amb un pes que pot superar els 200 kg. Té una silueta estranya que no recorda la de cap altre animal, un pelatge fosc, una alçada a la creu superior a la de la gropa, una gran crinera que recorre el coll i la part alta de l’esquena i unes banyes amb una forma molt particular. Tot i ser un animal robust, és una de les preses preferides dels grans carnívors, en particular del lleó (Panthera leo). Els joves encara estan especialment exposats als atacs dels predadors i tenen una alta mortalitat; molts són devorats tot just nascuts. Els nyus viuen en grans ramats formats per la reunió de petits grups familiars aplegats al voltant d’un mascle dominant, d’algunes femelles i dels petits nascuts aquell any. També hi ha grups que són formats per mascles joves. Pasturen principalment herba molt curta, és a dir tendra, i quan surten a buscar pastura tendra es produeixen les migracions massives que mouen centenars de milers de nyus de les sabanes herboses de la plana, que esdevenen inaprofitables quan arriba l’estació seca, a les llenyoses, aleshores menys inhòspites i el seu retorn quan comencen les pluges, al novembre.
El búfal africà (Syncerus caffer) és el més potent dels ungulats de la sabana. La base de les banyes amaga gairebé tot el front en els mascles, mentre que les banyes de les femelles, en canvi, estan molt menys desenvolupades. La seva talla és semblant a la d’un brau i el seu pes pot superar els 500 kg, sobretot els mascles de sabana, diferents de les formes que es troben a la selva plujosa. La seva àrea de distribució és extremament gran i cobreix totes les selves, les zones pantanoses i, sempre que no estiguin massa seques, també les sabanes, d’Àfrica. Freqüenta de grat les vores dels cursos d’aigua. És sobretot un pasturador que consumeix gramínies i que beu molt. Tot i la seva força, els lleons el poden abatre, però també sap resistir-los i a vegades ferir-los. Els vedells estan més exposats als predadors, en particular al caracal (Felis caracal) i a les hienes.
L’oribí (Ourebia ourebi), molt estès a l’Àfrica, des del Senegal fins al SE del continent, és una espècie de talla petita —no supera els 20 kg— de color rossenc amb el ventre blanc, i banyes fines i dretes en el cas del mascle. Freqüenta les sabanes relativament seques, on l’herba no és gaire alta. Generalment viu en parella, sense desplaçar-se gaire. S’alimenta sovint d’herba curta, més rarament de les fulles de les mates.
A les sabanes del Serengueti hi ha dues espècies de gaseles, totes dues amb nombrosos predadors. La gasela de Thomson (Gazella thomsoni) s’alimenta principalment d’herba curta i, en una proporció molt més petita, de fulles d’arbustos i arbres petits. Habita un territori de sabanes amb herba curta, clapada de mates disperses, en una àrea molt limitada de l’Àfrica centroriental. La gasela de Grant ( Gazella granti) té les banyes més llargues. El pes dels mascles pot assolir els 80 kg, i el de les femelles els 50. És una espècie que habita a les sabanes molt seques, fins i tot subdesèrtiques, però que també es troba en hàbitats més densos, però mai amb herba alta. La seva àrea de distribució és limitada a una petita part de l’Àfrica oriental.
El salta-roques o “klipspringer” (Oreotragus oreotragus) viu quasi exclusivament als afloraments rocosos, els “kopjes”. D’una alçada de 50 a 60 cm i un pes inferior als 20 kg, aquest antílop és particularment àgil. Es desplaça amb facilitat entremig de les roques gràcies a uns unglots molt petits i adherents. El pelatge dens li permet resistir les espines de la vegetació embosquinada. Generalment viu solitari, limitat al seu medi rocallós i s’alimenta tant d’herba com de fulles d’arbustos. Els seus enemics són principalment els lleopards (Panthera pardus) però també grans àguiles com la de Verreaux (Aquila verreauxii).
El dic-dic de Serengueti (Madoqua kirki) és un petit antílop amb les extremitats molt fines i el musell prominent. Normalment viu sol o en parella. La seva activitat és sobretot nocturna i la seva talla petita li permet protegir-se, entre les bosquines denses, dels nombrosos enemics com ara els lleopards, els caracals, les hienes i les àguiles.
A aquestes espècies herbívores cal afegir el damà roquer (Procavia capensis) que, com el salta-roques (Oreotragus oreotragus), freqüenta principalment els llocs rocallosos i accidentats. Encara que s’assemblen a les marmotes, tant per la mida com pel color, els damans roquers pertanyen a l’ordre dels hiracoïdeus, pròxim al dels proboscidis, és a dir els elefants. Tot i que freqüenten hàbitats força diversos, presenten una clara preferència per les zones rocalloses que els ofereixen un recer més segur. Encara que plantígrads, són extremament àgils per grimpar ja que la planta dels seus peus és coberta per una mena de coixinet que els assegura una bona adhesió sobre les roques. Les seves extremitats posteriors tenen 3 dits, mentre que les anteriors en tenen 4, amb petites urpes que encara els fan més bons grimpadors. Una coloració agutí, com la d’alguns gats, els dona una bona homocromia amb el medi herbaci, i la vida en grup, amb guaites que fan crits d’alarma en cas de perill, els dona una certa seguretat davant l’amenaça permanent que constitueixen per a ells les àguiles i els diferents depredadors terrestres com el lleopard. La gestació, que dura més de 200 dies, és molt llarga per tractar-se d’un animal d’aquesta mida i representa una fecunditat limitada per a l’espècie atès que el nombre de cries per ventrada generalment no passa de dues o tres. Dotats d’uns ronyons susceptibles de reabsorbir l’aigua, els damans roquers poden viure en medis molt àrids.
L’estruç (Struthio camelus) és un ocell corredor del grup dels ratites que, per la seva dieta alimentària predominantment herbívora, es pot relacionar amb els grans mamífers, particularment zebres i nyus, entremig dels quals no és estrany veure pasturar estruços. A aquest règim alimentari afegeix petits animals com ara llangardaixos, insectes i, fins i tot, algun rosegador. El seu aparell digestiu és, doncs, d’una eficàcia remarcable. L’alçada de l’estruç adult pot superar els 2,5 m, i el seu pes els 100 kg. Incapaç de volar, té unes potes ben fortes, que reposen tan sols sobre 2 dits i li permeten ser un gran corredor, que pot arribar a superar els 60 km per hora. Els combats entre mascles es fan a cops de pota i no de bec, encara que aquest és molt robust. L’estruç pon els ous sota terra o a la sorra, després d’haver netejat d’herba una petita superfície. Cada posta és de sis a vuit ous, d’un quilo i mig de pes, que són covats durant una mica més d’un mes. Quan eclosionen, els petits, naturalment molt indefensos enfront dels predadors, són defensats enèrgicament pels adults.
Els grans carnívors
El lleó (Panthera leo) és el depredador més gran de la sabana i el més poderós, amb un pes que pot arribar als 200 kg. La densitat de la seva població a Serengueti ha estat estimada en un individu per cada 10 km2, una biomassa d’uns 140 kg, ben pobra si es compara amb els 275 000 kg d’ungulats que hi pot haver per la mateixa superfície en aquesta regió. La proporció és encara més elevada en altres sabanes veïnes com la de Ngorongoro, on la densitat d’herbívors és molt elevada durant tot l’any. Els lleons, que assoleixen la maduresa als dos anys i no acostumen a viure’n més de quinze, viuen generalment en grups petits, d’una desena d’individus, entre els quals hi pot haver diversos mascles. De cada ventrada neixen normalment dues o tres cries que fan menys d’1,5 kg. La mortalitat juvenil és força important, als joves sovint els costa accedir a l’aliment i estan exposats a ser víctimes d’altres depredadors com ara les hienes o els lleopards. Els adults també poden morir víctimes d’un ramat de hienes, com també els mascles, de combats entre ells. Tenen com a preses tots els ungulats de la sabana, amb una preferència pels nyus, les zebres i les gaseles. La seva activitat és en part diürna, o si més no crepuscular.
La hiena tacada (Crocuta crocuta) és un animal de cap massís i forta dentadura, amb les potes anteriors més altes que les posteriors, i el pelatge rossenc, amb taques de color més fosc. El pes pot superar els 60 kg en els mascles. Viu en ramats, de vegades nombrosos, cada una amb un territori ben definit. Per criar, la hiena s’aixopluga dins un cau on poden néixer les cries —una o dues per ventrada— en qualsevol període de l’any, després d’una gestació de tres mesos. Sovint cooperen diferents femelles en la cria dels joves; així, mentre les unes guarden els cadells, les altres tenen la possibilitat de caçar. Encara que la hiena tacada s’alimenta de grat de carronya, en particular de restes d’animals morts pel lleó, és també un animal depredador, el més abundant de la sabana del Serengueti, notable i perillós per als ramats d’herbívors. El seu principal enemic és el lleó.
El lleopard (Panthera pardus) és ben conegut pel seu pelatge tacat. Els seus hàbits són solitaris i nocturns. El lleopard està estès per tots els hàbitats però principalment per les selves, per això no té a les sabanes del Serengueti un paper tan important com la hiena tacada o el lleó.
El guepard (Acinonyx jubatus) és un dels felins més elegants, que recorda una mica amb les seves línies esveltes les d’un llebrer. Amb 40 o 50 kg, és molt ràpid quan corre, la seva velocitat pot superar els 100 km per hora, molt més que qualsevol antílop. La seva forma de caça es basa en aquesta gran velocitat punta, que li permet fatigar la presa després d’haver-s’hi acostat reptant entre les herbes, la seva resistència és, en canvi, molt limitada. Les seves preses preferides són les gaseles, però també pot matar antílops més grans i fins i tot nyus quan s’apleguen uns quants guepards per caçar conjuntament. També captura llebres i dic-dics. El seu hàbitat són les sabanes d’herba d’alçada mitjana, que li permeten passar desapercebut i córrer a la vegada sense grans obstacles. El guepard, que també es troba a diverses parts de l’Àsia, especialment a Aràbia, a Iran i a l’Índia, ha estat utilitzat per a la munteria, però el seu ensinistrament és difícil. Per altra banda, tant a l’Àsia com fins i tot a l’Àfrica, ha esdevingut rar i pràcticament només es troba als parcs nacionals.
El licaó (Lycaon pictus) té una biologia molt notable lligada a la seva vida social. En efecte, viuen i cacen en gossades que els permeten, malgrat la seva poca grandària —un licaó pot pesar de 20 a 25 kg—, atacar preses tan grosses com els nyus o les zebres, i, per descomptat, els antílops de talla mitjana, les gaseles i les llebres. Els seus enemics, sobretot per als més joves, són les hienes ratllades i les àguiles. Tot i que en el passat les gossades havien estat força nombroses, actualment són només d’uns quants individus. La cooperació entre ells és estreta, quan no cacen s’arreceren, com les hienes, entre les mates, però també en caus, generalment manllevats als facoquers o als porcs formiguers (Orycteropus afer).
Els xacals es divideixen en tres espècies que tenen una mida força similar, però amb característiques ecològiques i àrees de distribució diferents. El xacal ratllat (Canis adustus) —el més estès a Àfrica— es troba a totes les zones de sabana, sobretot seques, entre el Senegal, Etiòpia i l’Àfrica meridional; té hàbits nocturns i les seves preses són animals petits molt diversos, carronya i fins i tot fruits. El xacal de llom negre (Canis mesomelas) només es troba a l’Àfrica oriental i a l’Àfrica meridional a les sabanes altes i arbòries; viu en parella i s’alimenta de preses diferents, com també de les restes de preses de grans feres. Finalment el xacal comú (Canis aureus), de pelatge groguenc, té una distribució sobretot septentrional que cobreix el N d’Àfrica, Aràbia, Àsia, Egipte i l’Àfrica oriental; com el de llom negre, viu en parella però la seva alimentació és molt més variada, ja que també menja vegetals.
Altres mamífers
A més d’aquests grans carnívors, al Serengueti també hi ha un cert nombre de petits carnívors però passen força desapercebuts al costat dels grossos, que han atret tota l’atenció dels investigadors. Com els petits rosegadors, quasi inexistents davant dels grans herbívors, aquests petits carnívors tenen un lloc molt reduït dins els ecosistemes d’aquestes sabanes.
Aquesta llista de grans mamífers zoòfags ha de ser completada pel porc formiguer o “aardvark” (Orycteropus afer), de l’ordre dels tubulidentats (anteriorment considerat dintre de l’ordre dels edentats). Aquest estrany animal s’alimenta de tèrmits dels quals fa un gran consum, els captura amb ajut de la llengua, que és llarga i viscosa com la dels pangolins (Manis). Té el musell prominent i cilíndric i les orelles llargues i dretes com les d’un conill. Dotat de potes potents i urpes fortes, obre sense esforç els termiters i hi excava fàcilment un cau profund, on passa els dies. És, en efecte, a la nit, quan surt a alimentar-se. Entre els seus predadors figuren especialment les hienes i el licaó.
El poblament de petits vertebrats i d’invertebrats: el cas de Lamto
Situada a l’Àfrica occidental, cap a 6°N, és el confí meridional de les grans extensions de sabana herbosa de la regió i fa com un gran tascó en forma de V al si del gran massís de selva densa de la Costa d’Ivori meridional. La sabana, per la seva banda, és penetrada per múltiples diverticles de bosc en galeria que voregen els cursos d’aigua tant grans com petits, fins i tot els temporals. El clima es pot considerar com a relativament humit ja que les precipitacions anuals són d’uns 1 300 mm, això sí, amb fortes variacions d’un any a un altre que poden oscil·lar entre valors extrems de 800 i 1 600 mm. Una estació seca que dura 5 mesos, variable segons els anys, imposa tanmateix considerables limitacions a la vegetació i a la fauna.
Els estudis climàtics posen de manifest la constància remarcable de les mitjanes mensuals de temperatura (25-26°C) en oposició a les fortes oscillacions diàries, accentuades durant l’estació seca, assuaujades durant la de les pluges, i acompanyades de variacions importants, en sentit oposat, de la humitat atmosfèrica. També han permès, al llarg dels anys, d’establir amb claredat la irregularitat de les variacions estacionals de pluviositat d’un any a un altre. Són aquestes variacions —i, per tant, les que resulten d’insolació i d’evapo-transpiració— les que marquen el ritme d’activitat de la vida vegetal i animal, influït per la intervenció dels focs, naturals o provocats, cada estació eixuta.
Sobre un substrat geològic divers on dominen els granits i els gneis, que afloren arreu, i les amfibolites, més localitzades, s’han format uns sòls força diversos. La sabana de Lamto ha estat atribuïda, des del punt de vista fitosociològic, a l’associació amb Brachiaria brachylopha, subassociació amb Loudetia simplexi la palmera Borassus aethiopum, atribució que de fet amaga la gran heterogeneïtat que es manifesta també en la vegetació d’aquesta sabana. A la pràctica, des d’un punt de vista fisiognòmic, es troben moltes fàcies que difereixen entre elles per la densitat dels arbres que sobresurten de l’estrat herbaci, fàcies que van des de la sabana herbosa amb tot just algunes palmeres disperses fins a una veritable sabana llenyosa amb un estrat herbaci poc abundant. Per altra banda, independentment de les fàcies que determina la densitat dels elements llenyosos, el tapís herbaci presenta, sota una aparent uniformitat, una diversitat important de les composicions específiques. Aquesta diversitat està vinculada especialment a la hidromorfia del sòl i al grau de recobriment de les plantes llenyoses. De forma progressiva se succeeixen, des de les zones més hidromorfes i més descobertes, sis espècies diferents de gramínies com a dominants: Loudetia simplex, Andropogon schirensis, Hyparrhenia diplandra, H. chrysargyrea, Sorghastrum bipennatum i Imperata cylindrica. Els únics arbres, sempre de mida petita, amb una freqüència notable, són la rubiàcia Crossopteryx febrifuga, l’euforbiàcia Bridelia ferruginea, l’araliàcia Cussonia arborea i Bauhinia (=Piliostigma) thonningii. Només el “rônier” o palmera de Palmira africana (Borassus aethiopum) supera sovint els 12 m d’alçada.
Els diversos inventaris realitzats en diferents grups animals donen una imatge quantitativa que completa, pels grups que no havien estat estudiats, els resultats assenyalats per a les sabanes de l’Àfrica oriental.
La relativa raresa dels grans mamífers
Tot i que a la regió de Lamto, on coexisteixen tipus de vegetació molt diferents, des de la selva densa fins a la sabana herbosa, i on, en distàncies petites, es manifesten diferències sensibles de clima, aixopluga una mastofauna rica i variada, a les sabanes pròpiament dites el poblament de mamífers grans i mitjans hi és força reduït. Una causa important d’aquesta limitació és la intensa pressió de caça que hi ha en aquesta regió i contra la qual la conversió en reserva d’una superfície massa restringida i sense suficient vigilància no ha pogut tenir l’eficàcia que caldria. Tanmateix, una altra causa, molt més fonamental, és l’escàs valor alimentari que té l’herba a Lamto, tal com passa a totes les sabanes relativament humides; sembla, en efecte, que en aquest sector la població de grans ungulats no ha estat mai important.
Així, a la sabana de Lamto no s’ha vist cap grup d’elefants africans (Loxodonta africana) des de fa més de 50 anys. D’hipopòtams (Hippopotamus amphibius), que abans hi entraven venint del riu Bandama, actualment ja no n’hi ha. Els facoquers (Phacochoerus aethiopicus), possiblement abundants en temps passats, també han desaparegut, igual que els oribís (Ourebia ourebi). Els antílops equins (Hippotragus equinus) i els búbals (Alcelaphus buselaphus) de les sabanes més septentrionals no sembla que hagin davallat tant cap al S. Només el cob comú (Kobus kob) subsisteix a la sabana de Lamto, on sens dubte havia estat abundant. També hi penetra, venint dels boscos en galeria, el búfal roig de selva (Syncerus caffer nana) i sobretot l’antílop jeroglífic o “guib” (Tragelaphus scriptus), i també alguns duikers com el de flanc ros (Cephalophus rufilatus) i el roig (C. dorsalis). Entre els primats típicament de sabana, encara s’hi troba el babuí anubis (Papio anubis) i potser també la mona vermella o “patas” (Erythrocebus patas). El “potto” (Perodicticus potto) entra sovint a la sabana al límit amb els boscos en galeria, igual com ho fa el cercopitec de cara negra (Cercopithecus aethiops).
Pel que fa a carnívors de talla mitjana o gran, manca el lleó (Panthera leo) mentre que la hiena tacada (Crocuta crocuta) pràcticament ha desaparegut. La civeta africana (Civettictis civetta) és sobretot forestal, com la civeta de palmera (Nandinia binotata). Hi ha, en canvi, diferents genetes (Genetta pardina i G. thierryi) i sobretot mangostes, en particular la vermella (Herpestes sanguineus) i, sobretot, la bruna o cusimansa (Crossarchus obscurus), que viu en grups de fins a una vintena d’individus tant a la sabana com al bosc en galeria. Tot plegat, una llista d’espècies evidentment d’interès biogeogràfic, però que, a causa dels seus febles efectius, tenen un paper mínim en el funcionament de la sabana de Lamto.
Els rosegadors
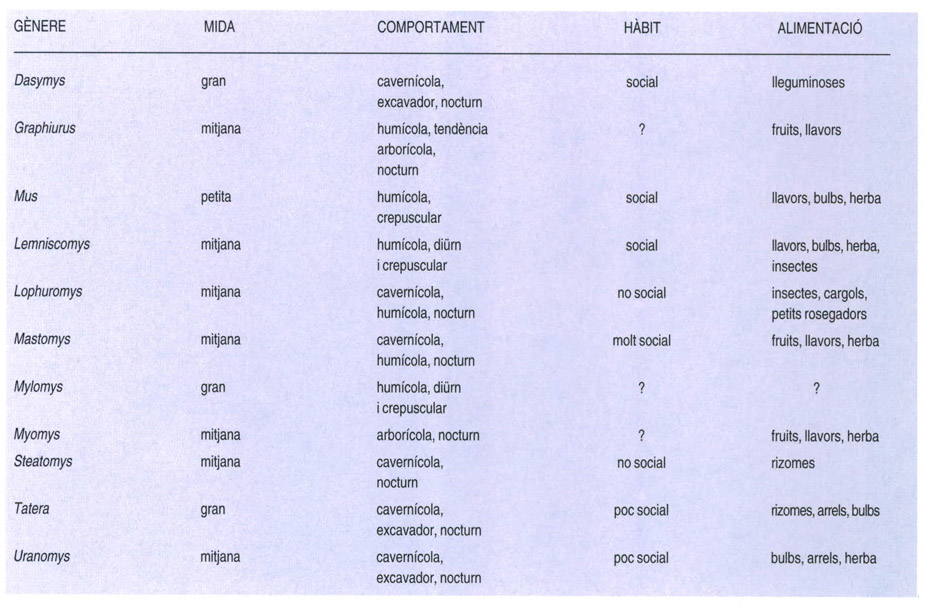
Característiques morfològiques, etològiques i dietètiques dels principals gèneres de rosegadors (Mus, Dasymys, Graphiurus, Lemniscomys, Lophuromys, Mastomys, Mylomys, Myomys, Steatomys, Tatera i Uranomys) de les sabanes de Lamto, a la Costa d’Ivori. Els rosegadors consumeixen principalment aliments vegetals, bé que algunes espècies de mida més gran tenen una dieta a base d’insectes i altres invertebrats. Tot i que a les sabanes africanes el nombre d’espècies de rosegadors és molt elevat, la seva biomassa generalment és més baixa que la dels herbívors més grans, i per tant tenen un efecte menor sobre la vegetació d’aquests ecosistemes.
Dades proporcionades per l’autor
La multitud de fàcies contribueix a diversificar les poblacions de petits rosegadors, com de tots els altres grups taxonòmics, i algunes espècies es mostren àmpliament dominants. Aquestes espècies són la rata de ventre blanc (Uranomys ruddi), el ratolí de sabana ratllat (Lemniscomys striatus), els ratolins nans (Mus) del subgènere Leggada i algun jerbu descalç com Tatera valida. Es troben també, en una abundància variable segons els medis, la rata de Dalton (Myomys daltoni) i una rata dormidora africana, Graphiurus murinus. En conjunt s’han observat una vintena d’espècies, però moltes provenen del medi forestal veí; les densitats, mai gaire elevades, presenten variacions estacionals i interanuals prou importants.
A aquestes petites espècies cal afegir la rata de Gàmbia (Cricetomys gambianus); d’un pes mitjà proper als 1 000 g, la densitat de la seva població pot ser estimada de mitjana en 0,1 individus/ha, és a dir, uns 100 g/ha en termes de biomassa. Una densitat semblant pot atribuir-se a l’esquirol terrestre africà (Xerus erythropus), que pesa aproximadament 350 g, cosa que representa una biomassa de 35 g/ha. La rata de canyar de Swinder o “aulacodi” (Thryonomys swinderianus), un animal d’hàbits nocturns que viu en grups petits, el pes del qual pot ser de 3 kg per individu i la densitat de 0,1 individus/ha, assoleix una biomassa de 300 g/ha. Alguns esquirols arborícoles com per exemple Paraxerus [=Aethosciurus] poensis i l’esquirol planador Anomalurus derbianus fraseri, dels boscos en galeria penetren també a la sabana i alguns hi fan niu a les palmeres mortes.
Els ocells
El poblament d’ocells comprèn moltes espècies diferents, algunes de les quals tenen una representació abundant. En una parcel·la d’estudi de 50 ha s’han observat més de 180 espècies diferents i, d’aquestes, 23 de forma constant i en gran nombre. Només tres espècies, la vídua d’esquena groga (Euplectes macrourus), el bulbul de ventre blanc (Pycnonotus barbatus) i el cistícola alacurt (Cisticola brachypterus), representen quasi un terç de tots els ocells, i amb cinc més, en completen la meitat. Les deu famílies principals són, per ordre decreixent dels seus efectius: la dels sílvids (sobretot els cistícoles de diferents espècies) i la dels plocèids, que suposen més de la meitat del poblament; els picnonòtids (bulbuls) amb el 10% del conjunt; i els colúmbids, fasiànids, coràcids, fenicoptèrids, psitàcids (lloros), burínids i apòdids. Pel seu pes individual elevat, els fasiànids (francolins i gallines de Guinea) representen més de la meitat de la biomassa del conjunt, seguits pels colúmbids (coloms i tórtores) i els coràcids (gaigs blaus).
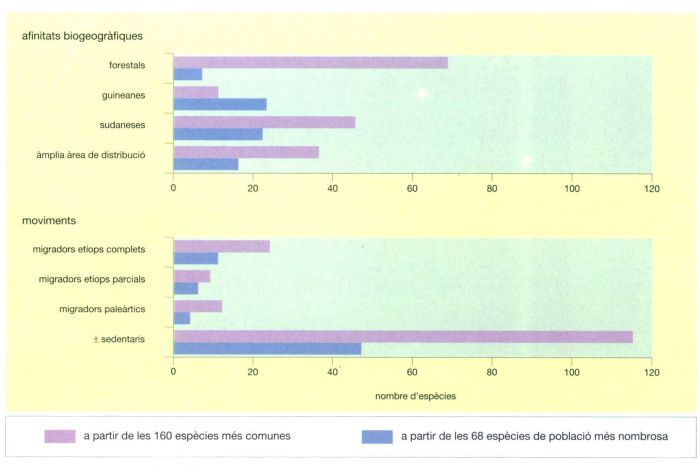
L’estructura del poblament de l’avifauna de la sabana de Lamto, a la Costa d’Ivori, es posa de manifest amb la classificació de les espècies d’ocells segons el seu origen biogeogràfic (a dalt) i el règim de migració (a baix). Atesa la proximitat de la selva, les espècies forestals són les més comunes o freqüentment observades, ja que sobre un total de 160 espècies, 68, és a dir el 43%, són forestals. En canvi, els ocells de bosc formen poblacions poc nombroses, i són els guineans i sudanesos els més ben representats pel que fa al nombre d’individus. D’altra banda, hi ha força espècies que són migradores temporals, bé d’origen etiòpic, bé d’origen paleàrtic, i tan sols es troben a Lamto durant l’estació seca.
Jordi Corbera, a partir de dades proporcionades per l’autor
Atesa la proximitat de les selves, el nombre d’espècies forestals observades és alt, però el nombre d’individus és escàs i per això no n’hi ha cap que figuri entre les 20 espècies dominants. Altrament moltes espècies d’origen sudanès es troben a Lamto en el límit de la seva àrea de distribució; només hi són presents a l’estació seca com a migradores temporals. De fet, dins del conjunt de l’avifauna de Lamto, un terç de les espècies sabanícoles són migradores, absents durant l’estació de les pluges, bé d’origen paleàrtic (petits passeriformes), bé d’origen etiòpic (principalment accipítrids, coràcids i meròpids) que provenen en molt més gran nombre de les regions veïnes d’Àfrica.
Només la quarta part de les espècies d’ocells tenen una densitat màxima al si de la sabana clara, mentre que més de la meitat depenen principalment dels bosquetons i boscos en galeria, i entren a la sabana només de forma accidental. A l’inrevés, les espècies de sabana no entren mai a la selva, en particular en el cas de les espècies vinculades a l’estrat herbaci com plocèids, sílvids i fasiànids. Certament el poblament no roman constant al llarg de l’any. És mínim a l’estació seca a la sabana herbosa, però és màxim a la mateixa època, prop de les vorades dels bosquetons o dels fondals boscats, més arrecerats i més rics en aliment. Les espècies es tornen a escampar pertot durant l’estació de les pluges, fins i tot algunes espècies vingudes de la selva com alguns frugívors. L’efectiu total mitjà és d’una desena d’individus per hectàrea, amb un pes d’1,2 kg, amb un màxim al final de l’estació seca, cap al mes d’abril, quan la presència de migradors és màxima, i un altre en plena estació de les pluges, quan els migradors han marxat, però els sedentaris estan en període de reproducció i una part dels frugívors de la selva exploten els fruits dels arbres de la sabana.
Els amfibis i els rèptils

Segons el règim alimentari, les serps de la sabana de Lamto, a la Costa d’Ivori, es poden agrupar en consumidors d’invertebrats, d’amfibis, de petits rèptils o de petits mamífers, segons tinguin com a preses exclusives (+++) o preferents (++) uns o altres d’aquests animals. Això no vol dir, però, que no cacin altres preses ocasionalment (+). Si bé hi ha serps que mostren una especialització alimentària molt marcada, d’altres —principalment de les famílies dels vipèrids i dels bòids— tenen una dieta molt més variada, i cacen preses molt diverses. D’altra banda, la distribució de les diferents espècies de serps està en relació amb la topografia, cosa que es pot explicar per les seves preferències alimentàries. Les serps que s’alimenten d’amfibis són especialment nombroses a les valls pantanoses i a les zones que queden periòdicament inundades, ja que les seves preses necessiten tolls d’aigua permanents per a completar el cicle vital. En canvi, als planells elevats abunden més els ofidis que s’alimenten de petits rèptils o mamífers, que no depenen tant de l’aigua.
Dades proporcionades per l’autor
Els amfibis i els rèptils tenen una importància gens negligible a les sabanes de Lamto. El nombre d’espècies és alt: 24 espècies d’amfibis, 8 de llangardaixos i 26 de serps, bé que amb diferències notòries de densitat segons els tipus de vegetació. Per als amfibis el factor principal és, evidentment, la presència més o menys abundant d’aigua; així la zona de fondals, al límit amb els boscos en galeria, està densament poblada, mentre que les zones més seques, de planell, en general són més pobres i amb una població de tipus diferent.
L’abundància de gripaus com Bufo regularis depèn de la presència de superfícies pelades que són el seu terreny de caça predilecte; la densitat disminueix als sectors massa plens d’herba i virosta. Algunes granotes terrícoles com Arthroleptis poecilonotus tenen la particularitat de tenir un desenvolupament directe a partir d’ous postos al sòl, per això és relativament independent dels cursos d’aigua de les fondalades; ben al contrari de les formes que es desenvolupen dins de l’aigua com Phrynobatrachus i Ptychadaena. En conjunt, la biomassa per hectàrea supera, durant una bona part de l’any, els 60 individus i els 200 g/ha.
Entre els llangardaixos, hi ha tres espècies d’escíncids diürns que dominen el poblament, Panaspis nimbaensis, Mabuya maculilabris i, sobretot, M. buettneri. Amb densitats que sovint superen els 50 individus per hectàrea, ocupen dins la població de vertebrats un lloc tan important com els amfibis, sobretot a les sabanes de tipus arbustiu. Aquest nombre disminueix de manera notable a les sabanes cremades i augmenta a les sabanes protegides del foc.
Les serps, supercarnívores, tenen evidentment densitats i biomasses menors que els llangardaixos i els amfibis, però la seva diversitat específica és gran i els seus règims alimentaris són variats. N’hi ha que mengen invertebrats, altres mengen llangardaixos, i fins i tot ocells o petits mamífers, en general de manera força especialitzada. Hi ha també una espècie, Dasypeltis scabra, que s’alimenta exclusivament d’ous d’ocells: té dos diverticles vertebrals que entren dins l’esòfag en forma de dues dents de serra que tallen la closca dels ous, que de seguida és escopida. Cal remarcar que la riquesa d’aquesta població sabanícola de rèptils i d’amfibis es deu, en part, a la immediatesa dels boscos en galeria a partir dels quals algunes espècies penetren en llocs descoberts durant l’estació de les pluges.
Els invertebrats epigeus no socials
La població d’invertebrats epigeus no socials de la sabana de Lamto és representada principalment per aranyes i insectes, mentre que els mol·luscs són pràcticament absents. El nombre d’espècies és elevat, si bé en termes de biomassa, acrídids i aranyes són els grups dominants, seguits dels heteròpters, les erugues de lepidòpters, les llagostes, els grills, les paneroles, els pregadéus i els coleòpters.
Les aranyes són especialment nombroses a la sabana i representen quasi la meitat dels artròpodes (fora dels insectes socials) i prop de la quarta part de la biomassa. Són, a més, molt diversificades, amb una vintena de famílies que comprenen més d’un centenar d’espècies que cohabiten regularment a l’estrat herbaci i a la superfície del sòl. Els licòsids dominen àmpliament aquest poblament, seguits dels saltícids i dels tomísids. Més de la meitat de les aranyes de l’estrat herbaci pertanyen a una d’aquestes tres famílies. Entre el fullatge d’arbres i arbustos, els saltícids i els tomísids són importants, però els licòsids, que viuen quasi exclusivament a terra, deixen lloc als argiòpids.
Els efectius i les biomasses del conjunt dels artròpodes augmenten durant l’estació de les pluges, però les fluctuacions només van del senzill al doble, ben poc pel que és usual en un poblament de sabana, cosa que cal relacionar amb el limitat rigor de l’estació eixuta. La densitat dels acrídids, per exemple, tampoc no arriba més que a triplicar-se. Aquesta estabilitat reflecteix la relativa constància de les principals característiques abiòtiques del medi: fluctuacions del fotoperíode inferiors a una hora al llarg de l’any i fluctuacions de temperatures mensuals mitjanes inferiors a quatre graus (força menors que les variacions entre el dia i la nit) i curta durada de l’estació eixuta, durant la qual les rosades són freqüents. La renovació parcial dels grups i de les espècies al llarg de les estacions atenua l’amplitud de les fluctuacions en el conjunt del poblament animal.
Els copròfags
Els cossos dels animals epigeus no són més que una petita aportació a la biomassa del sòl. El seu elevat contingut en proteïnes en fa, però, un aliment immediatament disponible i consumit gairebé sempre pels carnívors. No passa el mateix amb els excrements dels grans mamífers, en particular amb els dels ungulats: amb una proporció de l’ordre del 40% de matèria no assimilada en el menjar ingerit, els excrements dels ungulats representen una aportació quotidiana molt important a les sabanes. Aquells que se n’aprofiten també són, al seu torn, un element del poblament animal que cal tenir en compte.
Els copròfags són, primordialment, escarabèids i larves de dípters i de coleòpters hidrofíl·lids. Un olfacte molt desenvolupat lligat a la gran facilitat per a desplaçar-se que tenen els adults permet detectar la matèria fecal i arribar-hi de seguida que ha estat dipositada. Aquesta gran mobilitat fa molt delicada la determinació dels seus efectius i de la seva biomassa. Tanmateix s’ha provat d’avaluar-la.
A la sabana de Lamto, que no ha estat explotada ni pels humans ni pel seu bestiar, s’han classificat 77 espècies d’escarabèids copròfags i les sabanes més riques en grans mamífers herbívors encara tenen una fauna de copròfags més abundosa i diversa. Com a tots els medis, aquests escarabèids tenen biologies diverses. Més de la meitat de les espècies són diürnes; algunes, les més nombroses, colguen les matèries fecals per pondre-hi. D’altres, menys nombroses, les fan rodar a una certa distància en forma de pilotes, mentre que n’hi ha tres que són cleptoparàsits que fan servir les matèries preparades o transformades per altres espècies. Entre les espècies nocturnes també s’hi compten pilotaires però la gran majoria són excavadors.
Les formigues
Les formigues són, amb els tèrmits, un dels components principals de les poblacions d’insectes de la sabana de Lamto. Són representades per espècies molt diferents i per un nombre molt elevat d’individus. La majoria tenen el niu sota terra però realitzen l’activitat a la superfície. Els règims alimentaris van des d’una depredació pura i simple fins al consum de llavors, passant per l’aprofitament de matèria orgànica morta i de la secreció ensucrada que excreten les cotxinilles i els pugons.
L’espècie carnívora més abundant, i la més remarcable, és la formiga guerrera Anomma nigrans, que forma columnes de molts centenars de milers d’individus que solquen la sabana i la selva sense parar. Devoren tot el que troben al seu camí, des de cucs de terra fins als petits rosegadors que es deixin agafar, també tots els cadàvers. Les fileres corresponen tant a expedicions de caça com a trasllats de nius, sempre temporals. Hi ha un polimorfisme important que depèn de l’existència de nou categories d’obreres que es diferencien per la mida. Les més grans, qualificades de soldats, tenen les mandíbules molt desenvolupades. Una altra espècie de mida gran, Megaponera foetens, depredadora exclusiva de tèrmits, forma columnes força menys nombroses que les d’Anomma; ataquen els termiters i cada formiga s’enduu un tèrmit entre les mandíbules. Aquesta no és, però, una forma dominant a Lamto, ben al contrari d’una espècie propera, la formiga cadàver (Paltothyreus tarsatus), anomenada així per la fetor que desprèn, que consumeix preses molt diverses: tota mena d’insectes, particularment tèrmits, però també milpeus i cucs de terra. També és molt comú el gènere Camponotus, amb nombroses espècies que abunden sobre totes les espècies de gramínies i a l’interior de les seves tiges buides. Igualment nombroses i abundants són les espècies del gènere Crematogaster, presents tant als arbres com a les canyes de les gramínies, sobretot Crematogaster impressa. Altres espècies, per contra, viuen més aviat sota terra, per exemple les Amblyopone, que habitualment s’alimenten de miriàpodes quilòpodes (centpeus i escolopendres).
Entre les formigues principalment fitòfagues hi ha les del gènere Platythyrea i algunes del gènere Crematogaster amb nius epigeus. A Lamto, però, les formigues de règim vegetarià són poc nombroses. El consum de llavors és molt restringit, al contrari del que passa a les sabanes més seques; el practiquen sobretot espècies amb un règim més ampli, les de gèneres com Tetramorium, Pheidole o Atopomyrmex. L’ús de les secrecions ensucrades de les cotxinilles i de les mosques blanques és especialment patrimoni de les dels gèneres Crematogaster, Atopomyrmex i Platythyrea.
La fauna endogea
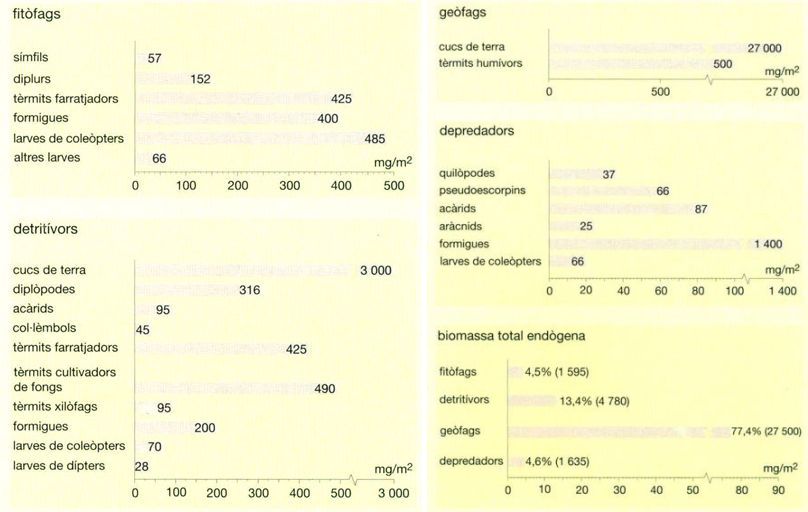
Biomasses mitjanes (en pes de matèria fresca) dels principals grups d’animals endogeus d’una sabana de Lamto, a la Costa d’Ivori. Cal destacar l’elevada biomassa dels cucs de terra, ja siguin detritívors o geòfags, que troben als sòls de Lamto les condicions adequades per a viure. De tota manera, a les sabanes amb pluges més irregulars els cucs de terra són molt menys abundants, i fins arriben a mancar completament a les més seques, ja que aquests invertebrats de cos tou són molt dependents de la humitat del sòl. L’altre grup dominant del poblament endogeu són els tèrmits, uns insectes socials confinats gairebé exclusivament a les regions tropicals i subtropicals. Els seus termiters en turonet sovint constitueixen un tret conspicu del paisatge de la sabana. Tanmateix, hi ha espècies de tèrmits que són arborícoles i fan el niu dins el tronc d’un arbre o un arbust, o bé a la seva superfície, i encara altres de subterrànies que construeixen el niu sota terra. Els tèrmits xilòfags, i els cultivadors de fongs, bé que representen una part gens menyspreable de la biomassa, ja que es tracta d’espècies de mida gran, no són especialment abundants a les sabanes obertes, ja que prefereixen hàbitats més densament forestats. En canvi, les espècies humívores i farratjadores, tant fitòfagues com detritívores, són les predominants.
IDEM, a partir d’Athias et al., 1974
Una de les característiques dels sòls de sabana és que acull un poblament animal abundós. Aquest medi té, en efecte, un microclima en el qual les variacions diàries de temperatura i humitat queden amortides i alhora ofereix als animals recer i refugi. Generalment els representants d’aquest poblament es classifiquen, en funció de la mida, en mesofauna (els animals de mida inferior a 3 mm) i macrofauna (de mida superior a 3 mm). La mesofauna comprèn oligoquets enquitrèids i el conjunt dels microartròpodes format, en particular, pels àcars i els col·lèmbols. A la macrofauna pertanyen els oligoquets no enquitrèids, les larves de coleòpters i els tèrmits.
Els cucs de terra, grup de particular importància al sòl de la sabana, tenen una densitat de més de 200 individus/m2, cosa que representa aproximadament 30 g per m2 en pes fresc. Hi ha 13 espècies, 3 de les quals es troben a la virosta suspesa de les fulles de les palmeres. Alguns viuen prop de la superfície i s’alimenten de la fullaraca; estan molt pigmentats i són ràpids. Els cucs de fondària són geòfags, menys pigmentats, més allargats, poc àgils i menys fecunds. La categoria intermèdia, qualificada d’epiendògena, és formada per cucs que s’alimenten de la terra remoguda prop de la superfície i expulsen els excrements en forma de petites muntanyetes molt característiques; una de les espècies d’aquest tipus, Millsonia anomala, ha estat estudiada molt acuradament i s’ha vist que el consum de terra, en període d’activitat, podia superar, cada dia, 15 vegades el seu propi pes. Es pot imaginar la quantitat de terra absorbida pel conjunt de la població. Els cucs de terra, molt sensibles a la humitat del sòl, tenen un cicle estacional molt marcat, amb una fase inactiva —la quiescència— durant l’estació seca. Els cucs, hermafrodites, ponen pocs ous i relativament resistents. La durada de la seva vida és, segons les espècies, d’un, dos o tres anys.
Els tèrmits són, amb els cucs de terra, l’element dominant del poblament animal endogeu. A Lamto n’hi ha més de 50 espècies, 36 a les sabanes, 15 de les quals hi tenen un paper important. En funció del règim alimentari es poden repartir en tres categories: farratjadors, cultivadors de fongs i humívors. Els tèrmits farratjadors col·lecten i consumeixen gramínies, mortes o vives, que tallen en petits fragments per portar-les al seu niu. Són representades principalment per 4 espècies del gènere Trinervitermes: T. togoensis, T. oeconomus, T. occidentalis i sobretot T. geminatus amb el 70% dels termiters. Aquests nius, en forma de cúpules irregulars, constel·len la superfície de la sabana amb una densitat de 50 a 75 per hectàrea; no se’n troba més que a la sabana. Els tèrmits cultivadors de fongs són recol·lectors de virosta de gramínies i de fusta morta. Amb els vegetals mastegats i pastats amb saliva fabriquen el que s’anomena “mola” on creixen els fongs dels quals s’alimenten. Les principals espècies de les sabanes de Lamto són: Ancistrotermes cavithorax, Microtermes toumodiensis, Odontotermes i Pseudacanthotermes militaris; curiosament, no s’hi troben Macrotermes. Els tèrmits humívors ingereixen terra, de la qual extreuen la matèria orgànica; la terra que consumeixen fa ben identificable l’abdomen negre de les obreres. Molt abundants a la major part de tipus de sabanes, totes les espècies de Lamto tenen nius hipogeus i sorprèn l’absència entre elles de Cubitermes, els constructors de ‘termiters bolet’, tan nombrosos arreu de les grans extensions de cuirasses ferruginoses més o menys pelades de les sabanes africanes. Una quarta categoria de tèrmits seria la dels xilòfags no cultivadors de fongs que consumeixen sobretot fusta parcialment degradada, però als ecosistemes de sabanes el seu paper és força restringit.
En conjunt els tèrmits de la sabana de Lamto tenen una densitat aproximada de 600 a 1 000 individus per m2, i representen una biomassa mitjana de més de 20 000 g per hectàrea (en pes fresc), és a dir, 5 500 g per hectàrea en pes sec, del qual uns 1 500 g corresponen als tèrmits farratjadors, 1 200 g als cultivadors de fongs i 2 500 g als humívors. El consum anual del conjunt de la població s’ha avaluat en més de 1 000 kg per hectàrea (en pes sec) de matèria vegetal i la producció de sexuats alats, en ocasió de les eixamenades, en més de 1 500 g (en pes fresc). Encara que amb densitats molt inferiors a les dels medis herbacis i forestals dels països temperats, els organismes de mida petita que constitueixen la mesofauna són nombrosos i corresponen a un gran nombre d’espècies.
Les àrees de repartiment de la fauna sabanícola africana
Les diferències observades en la composició de les poblacions animals de les diverses sabanes africanes estan causades per diferències de les àrees de distribució de les diferents espècies. Aquestes àrees són diferents perquè estan vinculades a les exigències ecològiques de les espècies i a les diferenciacions taxonòmiques que ha pogut provocar l’aïllament de les poblacions. El seu examen complementa la informació sobre la diversitat observada entre les poblacions de les diferents regions.
La relació de les àrees de distribució amb clima i vegetació
Una manera eficaç de determinar la relació entre el clima i la vegetació d’una banda i les espècies de l’altra és comparar les àrees de distribució respectives. Una aproximació com aquesta implica, però, que prèviament s’hagin establert aquestes àrees de distribució, cosa que representa un treball de camp important que, a les zones tropicals, només s’ha fet per a algunes espècies, ocells i grans mamífers principalment i, en una mesura força menor, rèptils i amfibis. Així i tot, encara falta molt perquè totes les regions de sabana siguin investigades d’una manera suficient. A l’Àfrica, la relativa regularitat de les fàcies de vegetació facilita la interpretació, encara que cal malfiar-se sempre de l’existència de variacions locals del medi —un aiguamoll, un illot de selva, una zona rocallosa— d’extensió massa petita per a sortir als mapes.
Les grans opcions ambientals
Algunes espècies localitzades a la regió somali i al corn d’Àfrica, com el “gerenuk” o antílop girafa (Litocranius walleri), la gasela de Clarke o dibatag (Ammodorcas clarkei), la gasela de Speke (Gazella spekei), la gasela de Soemmering (G. soemmeringi, el dic-dic de Phillips (Madoqua phillipsi), el de Swayne (M. swaynei) i el de Gunther (M. guentheri), només freqüenten sabanes àrides o semiàrides. Entre els carnívors, es troba també el cas de la mangosta d’Abissínia (Herpestes ochracea) i de la guineu orelluda (Otocyon megalotis). Així mateix, algunes espècies com la hiena bruna (Hyaena brunnea), la guineu del Cap (Vulpes chama), la mangosta grisa del Cap (Herpestes pulverulentus), la suricata (Suricata suricatta) i la musaranya elefant groga (Elephantulus intufi) es troben a les zones seques d’Àfrica meridional. Altres espècies tenen una àrea de distribució més estesa, però que encara correspon únicament a les sabanes seques, excloent les sabanes pre-forestals d’herba alta. És el cas de la gasela de front roig (Gazella rufifrons), del búbal (Alcelaphus buselaphus), del sassabi (Damaliscus lunatus), com també del guepard (Acinonyx jubatus) i de l’“aardwolf” o pròteles (Proteles cristatus).
Una situació inversa, és a dir, una preferència per les sabanes humides, correspon a les espècies amb àrees de distribució que recobreixen zones on hi ha boscos en galeria o fins i tot selves. Les següents espècies pertanyen a aquesta categoria, l’antílop jeroglífic o “bushbuck” (Tragelaphus scriptus), el duiker de flanc ros (Cephalophus rufilatus), el duiker comú ( Sylvicapra grimmia), el pangolí gegant (Manis gigantea), la rata de Gàmbia (Cricetomys gambianus), la rata de canyar de Swinder o “aulacodi” (Thryonomys swinderianus), el porc espí ( Hystrix cristata), la geneta Genetta tigrina, la civeta africana (Civettictis civetta), l’icnèumon (Herpestes ichneumon), la mangosta vermella (H. sanguineus), la mangosta de cua blanca (Ichneumia albicauda), el lleopard (Panthera pardus), el serval (Felis serval), el cercopitec de cara negra (Cercopithecus aethiops) i fins i tot els babuïns (Papio).
Les conclusions que es poden treure d’aquestes àrees de distribució són força imprecises, com ho són els seus límits. A més, només es refereixen a un nombre restringit d’espècies. Per tant, calen altres aproximacions per a aprofundir les relacions entre les espècies i el seu medi. S’han de basar en la comparació de biocenosis definides amb precisió en un biòtop molt limitat o recolzar-se en l’anàlisi de les poblacions que coexisteixen dins un territori heterogeni restringit.
La localització de les espècies en el medi
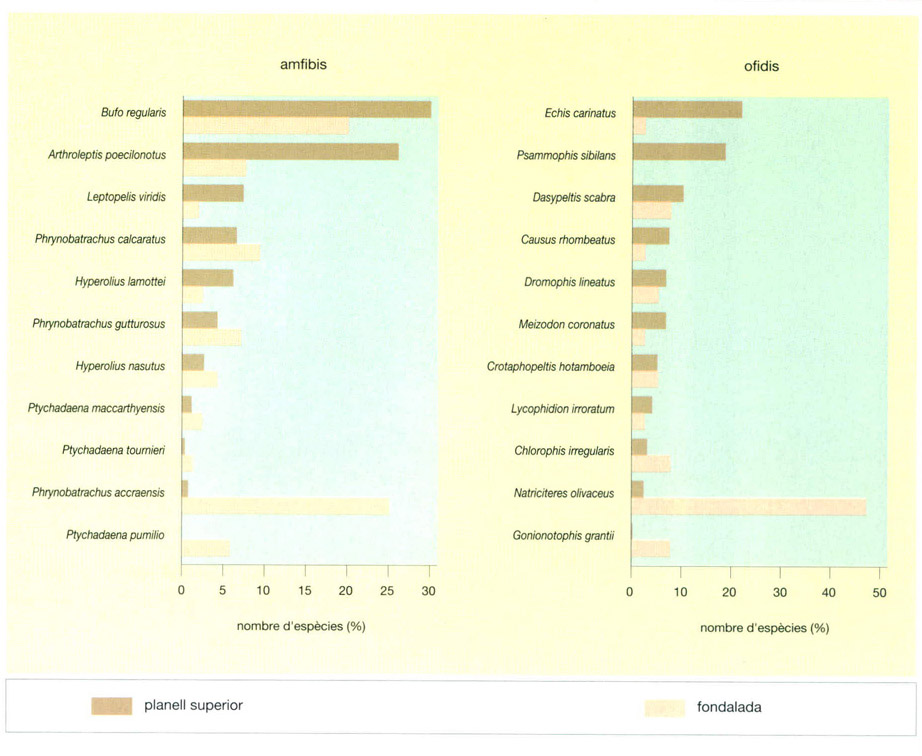
El poblament d’amfibis i el de rèptils ofidis de les sabanes de la reserva de Lamto, a la Costa d’Ivori, mostra una diferent composició taxonòmica a les fondalades i el planell superior, ja que la situació topogràfica determina en gran manera les condicions microambientals, la vegetació i el tipus d’aliments que els animals poden trobar. Els amfibis són molt sensibles al grau d’humitat i a la presència d’aigües permanents, dos factors que als ecosistemes secs de les sabanes sovint són determinants. Les fondalades solen ser molt més humides que no pas els vessants i els planells enlairats, que tenen un millor drenatge. Així, al planell superior de la reserva de Lamto són més abundants els amfibis terrestres, que no depenen tant de l’aigua, com per exemple Bufo regularis i Arthroleptis poecilonotus, que no pas els que en tenen una dependència més gran, com per exemple Ptychadaena pumilio o Phrynobatrachus accraensis, que abunden molt més a les fondalades. Amb els ofidis o serps passa una cosa semblant, bé que en aquest cas l’aigua no és un factor tan important, i agafa més relleu el règim alimentari de cada espècie.
Jordi Corbera, a partir de Barbault, 1974
Un primer exemple simple és donat per la influència de l’aigua i de la humitat més o menys gran del sòl. Especialment importants a les sabanes, que sovint són medis força secs, aquests factors varien molt amb la situació topogràfica. A la sabana de Lamto, per exemple, les fondalades sovint són molt humides mentre que els pendissos, més ben drenats, i els planells enlairats són clarament secs. Animals com els amfibis, per exemple, són molt sensibles a aquestes condicions. La resposta és molt diferent segons les espècies; algunes són més abundants a la zona d’altiplà, com el gripau Bufo regularis, o l’espècie petita de desenvolupament directe Arthroleptis poecilonotus, mentre els rànids Ptychadaena pumilio i Phrynobatrachus accraensis només es troben a les fondalades humides i, fins i tot, inundades.
Una distribució igual de contrastada es dona entre les serps de la mateixa sabana, condicionada en bona part pel règim alimentari, compost en gran mesura per espècies com Gonionotophis grantii i Natriciteres olivaceus, de batracis, mentre que altres, com Echis carinatusi Psammophis sibilans, s’alimenten preferentment de llangardaixos i de petits rosegadors. La densitat dels petits rosegadors també depèn molt de la humitat del sòl i, en conseqüència, del micro-relleu superficial. A una altra escala, la dependència respecte als punts d’aigua influeix molt en la distribució dels grans mamífers, durant l’estació seca, que ocasiona concentracions importants que desapareixen quan tornen les pluges.
La vegetació és, amb l’aigua, el factor més gran de la localització de les espècies. En efecte, és alhora font d’aliment pels fitòfags —per tant, secundàriament, pels zoòfags—, i refugi on trobar recer. Les sabanes que només tenen un estrat herbaci, sense arbres ni arbustos, són força escasses, però existeixen. Una de les seves característiques és la presència de grans ramats d’ungulats. Aleshores és la vida gregària la que garanteix a les espècies que hi viuen una certa seguretat. Facilita la vigilància i permet donar l’alerta més fàcilment quan s’acosta un depredador. Afavoreix eventualment la supervivència de la majoria pel sacrifici d’un individu, sovint ja desafavorit per l’edat o per una malaltia. El problema més gran que es dona en aquests medis és la recerca durant tot l’any d’herbes suficientment riques, és a dir joves. Pel que fa als ocells, l’obligació de niar a terra limita el nombre d’espècies i orienta les seves estratègies en el sentit d’una fecunditat més gran i d’un desenvolupament més ràpid dels pollets, que són nidífugs.
Les sabanes arbòries, en canvi, proveeixen als animals de llocs de repòs protegits de les mirades, molt útils quan l’herba és curta. L’augment de la densitat de les plantes llenyoses incrementa molt d’aquesta manera la seguretat dels grans mamífers. Si en una sabana herbàcia la silueta d’una girafa o d’un antílop es perfila a l’horitzó, en canvi resulta poc visible enmig d’un bosquet. A més de la seguretat, la presència d’arbres també aporta un cert confort arrecerant els animals durant les hores més caloroses del dia. Al costat d’aquesta seguretat aportada als herbívors, els arbres també són un recer per als carnívors. En algunes sabanes, on els arbres són escassos i dispersos, cadascun és sovint el lloc de repòs d’un lleó. Les feres, principalment els lleopards, de vegades també en fan el seu rebost penjant d’una branca les restes d’una presa incompletament devorada i conservada per als propers dies. Les sabanes arbòries i boscoses són així força més densament i diversament poblades de mamífers, com també d’animals dels grups més diversos, que les sabanes herbàcies.
Els ocells, evidentment, encara són més afavorits per l’abundància de plantes llenyoses. A la regió del Nimba, per exemple, la sabana herbosa d’altitud sense arbres és veïna d’una altra formació de dominància herbàcia, però amb nombrosos arbres petits escampats que donen al paisatge l’aspecte d’un verger. Paral·lelament la densitat d’ocells, molt baixa a la sabana herbàcia, augmenta considerablement, sobretot durant l’estació seca —l’hivern de l’hemisferi nord— durant la qual arriben xifres importants de migradors paleàrtics, molt especialment passeriformes. Moltes formes de la fauna tropical també hi són instal·lades permanentment, com els bulbuls Pycnonotus barbatusi Clororychla simplex, els cistícoles Cisticola brachypterusi C. lateralis, els suimangues, Anthreptes collarisi A. coccinigastero el teixidor Ploceus nigricollis.
En el cas dels grans mamífers, no és sols l’heterogeneïtat intrínseca d’una sabana arbòria allò que té un paper preponderant, sinó també la coexistència, a poca distància, de medis diferents com ara sabana herbosa, sabana més o menys llenyosa o bosc en galeria. Sota climes d’estacions tan contrastades com els de les zones de sabana, aquesta diversitat espacial compensa la variació del medi al llarg del temps. La multiplicitat dels tipus d’hàbitats afavoreix la supervivència de les poblacions. Ja se n’han remarcat els efectes en el cas de les sabanes del Serengueti, on sobre una extensió a l’escala de la capacitat de desplaçament dels grans mamífers coexisteixen diferents tipus de sabanes.
L’heterogeneïtat dels paisatges dona també un ventall més ampli de possibilitats de diversificació de maneres de viure, una més gran diversitat de nínxols ecològics, i augmenta així la riquesa d’espècies en el territori. En el parc del riu Comoé, on hi ha barrejades sabanes herboses i llenyoses, boscos clars i boscos densos al llarg dels cursos d’aigua, la localització habitual de les principals espècies de grans mamífers fitòfags es pot definir amb precisió.
La tria de l’aliment pels fitòfags
Per als fitòfags, la tria de l’hàbitat depèn d’una manera estreta de l’aliment que busquen. Segons els tipus de vegetació, els diversos grups animals tenen importàncies diferents. Les zones d’herba curta són preferides pels tèrmits per una banda, i pels grans ungulats per l’altra. En canvi, aquests darrers estan poc representats a les sabanes amb vegetació rupícola, més accessibles, en canvi, als petits rosegadors, als acrídids... i als depredadors.
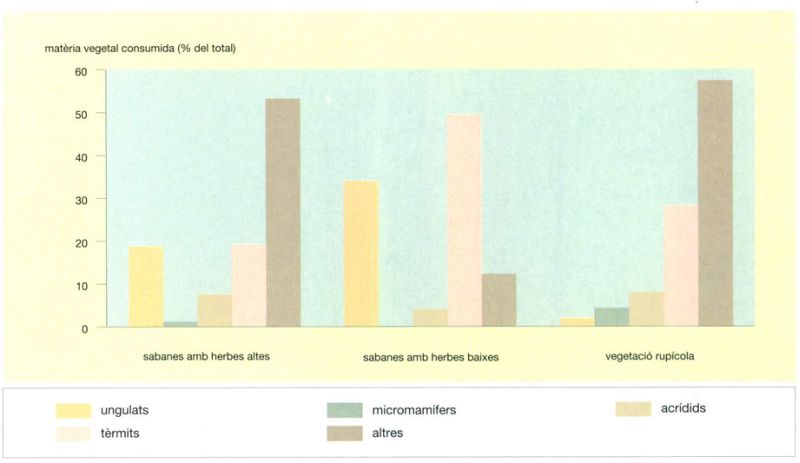
Consum de matèria vegetal dels principals grups d’herbívors i detritívors als tres tipus de sabana del Serengueti, a Tanzània. Cada hàbitat ofereix un tipus o altre d’aliment vegetal, i així la importància de les diferents espècies herbívores és variable. Els ungulats, per exemple, són responsables del consum del 34% de la matèria vegetal de les sabanes d’herbes baixes, i del 18,8% en les sabanes d’herbes altes. En canvi, pràcticament no troben aliment entre la vegetació rupícola, que fa de més bon aprofitar per herbívors més petits.
Jordi Corbera, a partir de Sinclair, 1975
S’han fet molts estudis, en diverses sabanes, sobre les preferències alimentàries dels grans mamífers fitòfags. Així com en els medis herbosos la tria és limitada, no passa el mateix a les sabanes llenyoses. En aquestes, en efecte, mentre que algunes espècies, les pasturadores, consumeixen només gramínies, d’altres són brostejadores que utilitzen fulles d’arbres, més riques en proteïnes però també més indigestes pels tanins que contenen. Altres, encara, tenen un règim que comprèn els dos tipus d’aliment, sigui simultàniament, sigui successivament segons l’estació. Al Serengueti és palesa la diferència entre la gasela de Thomson, més d’un terç del règim alimentari de la qual consisteix en fulles de dicotiledònies (arbustos i petits arbres), i les tres espècies (el nyu blau, el sassabi i la zebra de Burchell) exclusivament pasturadores. D’aquestes darreres, la zebra manifestament usa de les gramínies una quantitat més gran de tiges i força menor de fulles que el nyu i el sassabi.
Un estudi similar de la dieta dels grans mamífers fitòfags ha estat realitzat al parc nacional del riu Comoé. Distingeix tres fonts d’aliment: les fulles d’espècies llenyoses, les plantes herbàcies no gramínies (mimosàcies, per exemple) i les gramínies. En funció de la importància relativa d’aquests tres tipus d’aliment, les espècies es poden repartir en tres categories que són els brostejadors, els omnívors de tendència vegetariana i els pasturadors. L’elefant, per exemple, és quasi exclusivament un brostejador de fulles igual que els antílops de selva; els babuïns són uns vegetarians omnívors, mentre que el búfal, els antílops de sabana, l’hipopòtam i el facoquer són principalment pasturadors. En el cas de les espècies vegetals llenyoses també ha estat possible precisar la part consumida de la planta: escorça, arrel, fulla, flor o fruit.
Al costat d’aquests resultats estadístics, l’observació directa dels animals mentre s’alimenten fa veure amb més precisió les modalitats del consum de l’herba per part d’algunes espècies dominants, i la contribució aportada, d’aquesta manera, al repartiment dels recursos. Mentre que elefants, búfals i hipopòtams poden utilitzar herba alta —i també, en el cas dels elefants, fulles d’arbustos—, l’antílop equí i els búbals busquen herba més aviat mitjana. Els cobs comuns mengen, en canvi, herba curta i els facoquers herba encara més curta.
La distribució dels mamífers
A causa de l’extensió dels territoris que ocupen i com que les condicions del medi són relativament semblants, les sabanes africanes ressalten el paper de l’aïllament físic sobre la diferenciació de les espècies. Entre les diverses regions, a vegades, hi ha obstacles que són franquejables per les espècies de talla gran i capaces de fer grans desplaçaments. Aquest fet és particularment evident quan s’observa l’elefant o l’hipopòtam, perfectament ubics. Altres espècies de grans mamífers, però, tenen diferenciacions locals causades per un aïllament relatiu que porta a subespeciacions i a especiacions dins d’un mateix gènere. Sovint també són reforçades per la desaparició d’espècies que s’havien produït en un o altre territori. Dins els grups menys mòbils, com els petits rosegadors i la majoria dels invertebrats, es desenvolupen, sovint, divergències més notòries que poden afectar els gèneres, o fins i tot ser més específiques. Aquestes diferenciacions, vinculades a la distància, augmenten també per la intervenció d’altres condicions ecològiques, principalment climàtiques.
Els mamífers d’àrea restringida
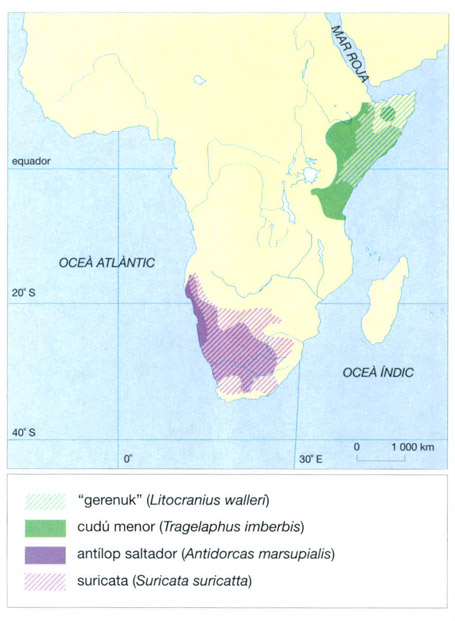
Distribució de quatre espècies de mamífers de les sabanes africanes, d’àrea restringida. El “gerenuk” o antílop girafa (Litocranius walleri) habita només les zones àrides del corn africà. Se’l troba freqüentment a les sabanes arbustives amb una coberta esclarissada de matolls espinosos, dels quals brostegen les fulles i els brots tendres. El cudú menor (Tragelaphus imberbis) habita els paratges arbrats o arbustius més o menys densos de l’extrem de l’Àfrica oriental, i no se’l troba mai en les planures herbàcies, ja que s’alimenta de fulles, tiges joves i branquetes d’arbres i arbustos. L’antílop saltador ( Antidorcas marsupialis) habita al SW del continent africà. Els seus ramats nombrosos sovint cobreixen les>planures obertes seques fins a perdre’s de vista. Finalment, la suricata (Suricata suricatta) és una mangosta petita exclusiva de les planes seques de l’Àfrica meridional, on excava llodrigueres subterrànies i caça insectes i petits vertebrats.
IDEM, a partir de fonts diverses
Tanmateix, fins i tot entre les espècies de grans mamífers, n’hi ha que només es troben en un territori restringit, cas que, a l’Àfrica, és més freqüent entre les espècies de la selva que entre les formes de sabana. Un cert nombre d’ungulats, per exemple, es troben localitzats exclusivament en algunes sabanes i subdeserts de la part oriental d’Àfrica, és el cas d’algunes gaseles com la de Speke (Gazella spekei) i la de Clarke o “dibatag” (Ammodorcas clarkei), de l’òrix beisa o beisa d’Abissínia (Oryx gazella beisa), del niala de muntanya (Tragelaphus buxtoni), de l’antílop nan (Dorcatragus megalotis), dels dic-dics (Madoqua guentherii M. swaynei) i del “gerenuk” o antílop girafa (Litocranius walleri); també les d’altres mamífers com la llebre d’Abissínia (Lepus habessinicus), i nombroses espècies de petits rosegadors.
Els mamífers de les sabanes meridionals
Altres espècies estan confinades a les zones meridionals i algunes, més particularment, al SW del continent. Entre els ungulats hi hauria el cèlebre “springbok” o antílop saltador (Antidorcas marsupialis), que en el passat comptava els seus efectius per milions però en una àrea relativament restringida, l’antílop cabirol (Pelea capreolus), el niala (Tragelaphus angasi), de la costa del SE africà, l’òrix (Oryx gazella) i el “blesbok” de front blanc (Damaliscus dorcas). Entre els rosegadors hi ha esquirols terrestres com Xerus inaurisi la rata “dassie” (Petromus typicus) i entre els lagomorfs, la llebre de bosquina (Lepus saxatilis) i la llebre dels boiximans (Bunolagus monticularis). També s’hi troba el babuí sud-africà (Papio ursinusi diversos carnívors com el gat de peus negres (Felis nigripes), la guineu del Cap (Vulpes chama), la hiena bruna (Hyaena brunnea), la suricata (Suricata suricatta), l’“aardwolf” o pròteles (Proteles cristatus), que s’alimenta de tèrmits, principalment dels del gènere Trinervitermes, i la mangosta groga (Cynictis penicillata). Entre els insectívors de l’Àfrica meridional destaquen l’eriçó del Cap (Erinaceus frontalis) i els estranys macroscèlids, de musell allargat en forma de trompa mòbil: la musaranya elefant d’orelles curtes (Macroscelides proboscideus) i les d’orelles llargues (Elephantulus myurus, E. intufii E. rupestris).
Els mamífers de les sabanes septentrionals
Moltes espècies ocupen una llarga franja de sabana que s’estén d’E a W, a l’hemisferi nord, entre el Senegal i el Sudan nilòtic, però no s’estenen cap a l’Àfrica central. Aquest és el cas, entre els ungulats, de la gasela de front roig (Gazella rufifrons), i de la gasela dama ( G. dama) —aquesta amb una àrea de distribució força fragmentada—, del duiker de flanc ros (Cephalophus rufilatus), de les sabanes més humides, com també del redunca comú o “nagor” (Redunca redunca), d’aspecte de cabirol, que es troba igualment a les muntanyes fins a Tanzània. Entre els carnívors, la guineu pàl·lida (Vulpes pallida) té també una àrea de distribució d’aquest tipus. Entre els rosegadors l’esquirol terrestre africà (Xerus erythropus), que desborda un xic l’àrea citada per l’orient africà, i, fins i tot, el porc espí (Hystrix cristata), que, cap a l’W, puja fins al Magreb.
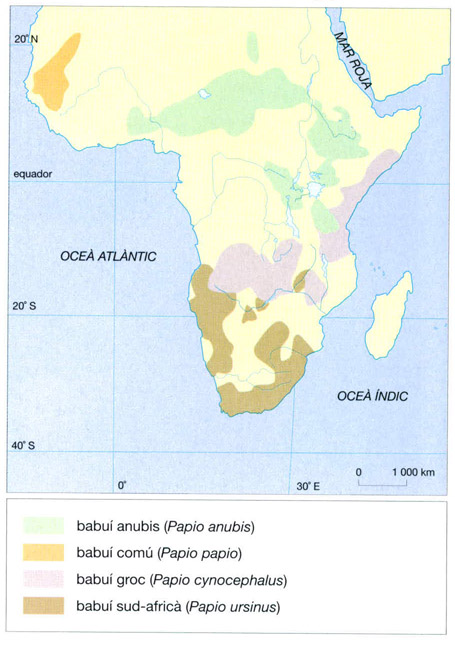
Àrea de distribució de quatre de les cinc espècies de babuïns (Papio) de les sabanes africanes. Els primats d’aquest gènere són els únics que viuen exclusivament als hàbitats de sabana, però alguns ocupen, també, zones més o menys boscoses i hi ha espècies que fins i tot habiten en àrees semidesèrtiques i en roquissars. El babuí comú (P. papio), per exemple, es mou sobretot pels paratges amb roques soltes, i el babuí sud-africà (P. ursinus) es troba freqüentment a prop dels “kopjes”, uns turons rocallosos dispersos típics de les planures sud-africanes amb matollars. El babuí groc (P. cynocephalus), per contra, es pot trobar tant a les sabanes com a les zones boscoses obertes.
IDEM, a partir de fonts diverses
No és rar entre els animals de selva de l’Àfrica occidental que es localitzin en una àrea limitada; entre les espècies de mamífers de sabana, en canvi, només es poden assenyalar els casos de la mangosta de Gàmbia (Mungos gambianus) i del babuí comú (Papio papio), que és diferent del babuí anubis (P. anubis) de l’Àfrica oriental. Les formes de talla més petita tenen una capacitat menor de desplaçament que sovint ha fet possible una especiació i moltes espècies endèmiques de petits rosegadors i de musaranyes només viuen a l’extrem W africà, al Senegal, a Guinea i a la Costa d’Ivori.
Els mamífers de les sabanes orientals
Es tracta de les espècies l’àrea de distribució de les quals s’estén des del NE africà pel corn d’Àfrica, i baixa fins al S del continent, passant per la regió dels grans llacs. Aquesta àrea, per tant, exclou els territoris de l’Àfrica occidental i la part més austral del continent. Entre els antílops distribuïts d’aquesta manera hi ha el “grysbok” o raficer del Cap (Raphicerus melanotis), una forma de talla petita, molt propera a l’“steenbok” o raficer comú (R. campestris) de l’Àfrica meridional, el saltaroques (Oreotragus oreotragus), que és una forma adaptada a les roques, l’impala (Aepyceros melampus), el nyu blau (Connochaetes taurinus), la zebra de Burchell (Equus burchelli), el damà gris de Bruce (Heterohyrax brucei) i el pangolí del Cap (Manis temmincki).
El cas dels rinoceronts requereix una atenció especial. El rinoceront negre (Diceros bicornis) era molt estès, encara no fa un segle, a tot el conjunt de la zona de les sabanes d’Àfrica. L’àrea on viu actualment comprèn alguns territoris molt reduïts del centre, l’E i el S del continent i és el resultat de l’extermini de què ha estat víctima. El rinoceront blanc (Ceratotherium simum) ha estat víctima d’un extermini encara més eficaç, i per això només es troba en alguns sectors aïllats de l’Àfrica oriental però també en algunes àrees cap al Txad i cap a Namíbia.
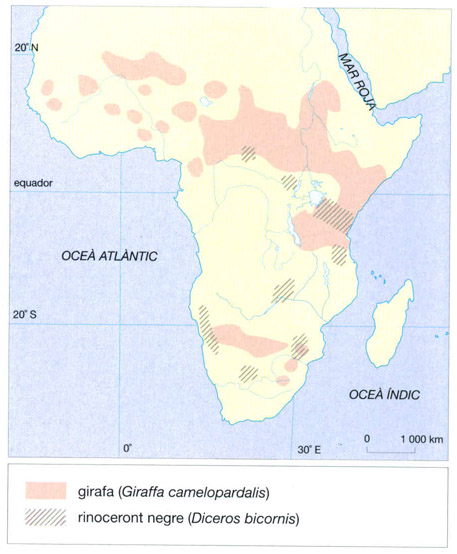
Àrea de distribució de la girafa (Giraffa camelopardalis) i del rinoceront negre (Diceros bicornis). Ambdues espècies antigament s’estenien per una àrea molt més àmplia. La girafa es trobava gairebé per tot el continent africà, fins i tot a la regió del Sàhara central, bé que la creixent aridesa del clima l’obligà a marxar. Posteriorment, la caça descontrolada l’eliminà de la major part de l’Àfrica occidental, tal com succeí amb el rinoceront negre, avui dia greument amenaçat.
IDEM, a partir de fonts diverses
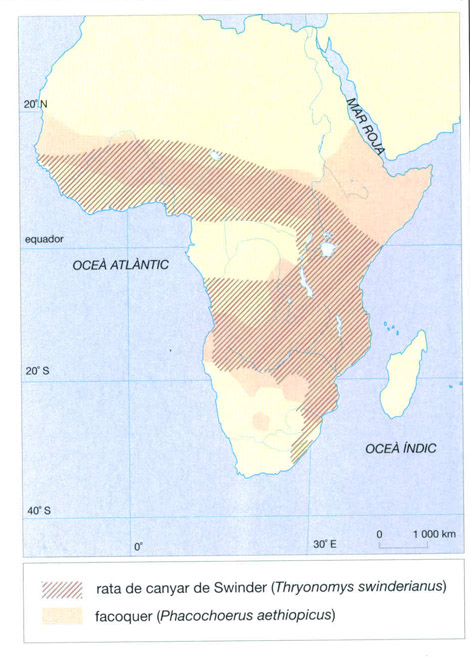
Àrea de distribució del facoquer (Phacochoerus aethiopicus) i de la rata de canyar de Swinder (Thryonomys swinderianus), dos mamífers àmpliament difosos per la majoria de sabanes africanes. El facoquer és un porc salvatge que habita les sabanes obertes. Se’l pot trobar, sobretot, en àrees amb aigua disponible per beure i rebolcar-se, però també viu en zones molt seques, on l’aigua escasseja durant la major part de l’any. El cas de la rata de canyar de Swinder és remarcable perquè es tracta d’una espècie ubiqua que habita des de les sabanes del Senegal fins a les de Sud-àfrica, i també s’estén per algunes àrees de selva plujosa. Aquest rosegador, d’uns 50 o 60 cm de llargada, sense comptar la cua, i fins a 7 kg de pes, es troba, principalment, entre els canyars i els matollars de les ribes dels rius i els llacs, bé que també freqüenta hàbitats més secs.
IDEM, a partir de fonts diverses
Els mamífers d’àrea ubiqua
A diferència de totes aquestes espècies, poc o molt localitzades, algunes altres són molt ubiqües i es troben a tota l’Àfrica tropical. A més de l’elefant i de l’hipopòtam es pot citar, entre els herbívors, el búfal africà (Syncerus caffer), el duiker comú (Sylvicapra grimmia), el búbal (Alcelaphus buselaphus), el sassabi (Damaliscus lunatus), l’antílop equí (Hippotragus equinus), el facoquer (Phacochoerus aethiopicus), el gàlag del Senegal (Galago senegalensis), el cob comú (Kobus kob) i el cob aquàtic (K. ellipsiprymnus), la llebre Lepus whyteii la llebre del Cap (L. capensis), i la rata de canyar de Swinder o “aulacodi” (Thryonomys swinderianus). Entre els carnívors, les espècies ubiqües són el lleopard (Panthera pardus), el lleó (P. leo), el licaó (Lycaon pictus), el caracal (Felis caracal), el gat de Líbia (F. silvestris libyca), que es troba també a Aràbia i al Pròxim Orient, el serval (F. serval) i les mangostes Ichneumia albicaudai Atilax paludinosus. Moltes espècies abasten àrees que depassen Àfrica, entre d’altres el lleopard, el guepard i el lleó. També és el cas de la llebre del Cap (Lepus capensis) o de la hiena ratllada (Hyaena hyaena), absent al centre i el S d’Àfrica, però present a Aràbia i a l’Índia.
Aquestes distribucions, bé que globalment ubiqües, no són idèntiques entre elles ja que depenen de les exigències ecològiques de l’espècie. Algunes formes, més eurítopes, és a dir més tolerants quant al tipus d’hàbitat, poden, per exemple, entrar a la selva o bé a les zones més seques, i la seva distribució aleshores és molt contínua. D’aquest tipus són les àrees del lleopard o de l’elefant. Altres espècies, més exigents, estan limitades a un biòtop particular i la seva àrea de distribució aleshores comprèn territoris més o menys separats. La majoria dels ungulats que tenen un tipus d’alimentació específic tenen àrees de distribució d’aquestes característiques. L’extensió de les poblacions humanes provoca sempre una fragmentació més gran de les àrees de distribució, tal com s’observa amb les de la girafa, el lleó i encara més amb les dels rinoceronts.
La distribució dels ocells
En els ocells es retroben àrees de distribució que recorden les dels mamífers. Alguns, com la gallina de Guinea comuna (Numida meleagris), algun teixidor com la quelea de bec vermell (Quelea quelea) i l’esplugabúfals Buphagus africanus, com també l’àguila bel·licosa (Polemaetus bellicosus), l’àguila saltimbanqui (Terathopius ecaudatus) i el serpentari o secretari (Sagittarius serpentarius) es troben en una àrea que cobreix totes les sabanes d’Àfrica. L’estruç (Struthio camelus) també està estès de l’W a l’E i al S d’Àfrica però no es troba a les sabanes humides d’herba alta i té l’àrea de distribució limitada a les regions més seques. L’àguila de Verreaux (Aquila verreauxii), en canvi, està limitada a l’Àfrica oriental i meridional.
Entre els calaus, el calau de terra cafre (Bucorvus cafer) ocupa tot Àfrica al S de l’equador, mentre que el calau de galtes argentades (Ceratogymna brevis), únicament s’estén per una franja ampla dels territoris orientals, des de la mar Roja fins a Moçambic; el calau de bec groc (Tockus flavirostris) es limita al corn d’Àfrica, igual que la gallina de Guinea vulturina (Acryllium vulturinum) i el francolí de gorja groga (Francolinus leucoscepus). Un altre teixidor, el Bubalornis albirostris, i la perdiu roquera (Ptilopachus petrosus) tenen la clàssica distribució en una franja que va d’E a W. La majoria de les espècies de mida petita es troben en àrees més limitades.
La distribució dels vertebrats de sang freda i dels invertebrats
Els mamífers poc o molt coneguts —sobretot els de talla gran i mitjana— i els ocells, sigui quina sigui la seva mida, han estat objecte de molts estudis arreu d’Àfrica. En canvi, amb algunes excepcions, amb els coneixements actuals és difícil donar amb precisió les àrees de distribució de la major part dels invertebrats i dels vertebrats inferiors, com els amfibis i els rèptils. D’una banda hi ha molts punts foscos de taxonomia per aclarir, i de l’altra les investigacions són encara força insuficients per a la majoria de les espècies que no siguin les més típiques. Només es pot dir que les formes amb una capacitat baixa de dispersió —per exemple la majoria dels cucs de terra i també dels mol·luscs, dels miriàpodes i dels rosegadors— tenen àrees de distribució força més reduïdes que els animals grossos.
El poblament animal d’algunes sabanes africanes extremes
Per a comprendre la distribució de les espècies al si dels grans conjunts continentals cal completar l’aspecte geogràfic de la distribució dels diversos grups taxonòmics animals amb una anàlisi fina dels factors ecològics. Fins i tot en un territori limitat, els medis aplegats sota el nom de sabana són de fet molt variats. Encara amb més raó poden variar entre espais geogràfics allunyats entre ells. Segons la seva naturalesa, aquests factors s’han de considerar a escales diferents. Les diferències climàtiques i dels grans tipus de vegetació es perceben tan sols a escala de desenes, fins i tot centenes de quilòmetres. En els grans paisatges definits així, es manifesta una heterogeneïtat a totes les escales. L’estudi de l’efecte d’aquesta heterogeneïtat sobre el poblament animal s’ha de fer per a cada un dels diferents grups taxonòmics perquè no pot ser el mateix per als petits invertebrats que per als grans mamífers.
L’estudi de la composició de les poblacions animals en funció de les condicions climàtiques i del tipus de vegetació és una primera aproximació de l’acció dels factors del medi. Això afecta totes les subdivisions del medi i totes les espècies i s’expressa, per tant, a escales molt diverses, que poden anar del centímetre cúbic per als organismes del sòl de mida molt petita fins a extensions d’uns quants quilòmetres quadrats per als animals grans. Aquí es consideraran més especialment els grups taxonòmics amb representants fàcilment visibles i els factors del medi immediatament accessibles a l’observació.
El poblament animal de la sabana humida d’altitud de Nimba
Les carenes dels monts Nimba, situades a la frontera entre Guinea i la Costa d’Ivori, reben precipitacions importants, de 1 500 a 3 500 mm anuals segons l’orientació. Fora de l’estació de pluges el clima és, en canvi, molt sec, de manera que les estacions estan molt contrastades. Les carenes, per sobre dels 1 400 m d’altitud, són cobertes d’una sabana densa, a vegades anomenada praderia, formada per un gramenet de poca alçada. Els sòls, a causa de l’erosió vinculada al relleu accidentat, són esquelètics, cosa que explica l’absència quasi total d’arbres i arbustos. Aquesta sabana d’altitud és un illot aïllat enmig d’una zona forestal. També acull una fauna particular que conté desenes d’espècies animals endèmiques, bé que amb una riquesa específica global inferior que a les grans sabanes africanes de la plana.
L’escassa superfície no permet la diferenciació d’espècies originals de mamífers. Els que hi ha passen gran part de la seva vida —sobretot durant el dia— a la selva veïna. Aquest és el cas, entre els mamífers, del búfal roig de selva (Syncerus caffer nana), de l’antílop jeroglífic o “guib” (Tragelaphus scriptus), i dels duikers, principalment el roig (Cephalophus dorsalis), tots ells cada cop més rars. En canvi, hi ha dues espècies de mida mitjana que són abundants: el damà roquer (Procavia capensis) i la rata de canyar de Swinder o “aulacodi” (Thryonomys swinderianus). Són les víctimes principals dels lleopards (Panthera pardus), els excrements dels quals són nombrosos arran de carena. Entre els carnívors també hi ha la civeta africana (Civettictis civeta) i alguna geneta com Genetta tigrina. Els petits rosegadors són representats per quatre espècies i les musaranyes per dues, però la densitat és sempre baixa.
La humitat extrema que hi ha durant la meitat de l’any a la sabana d’altitud és la responsable de l’escassetat de rèptils, especialment llangardaixos, per als quals l’estació seca seria la favorable. Entre les serps, només són freqüents les colobres Dipsadoboa unicolorque s’alimenten principalment dels petits gripaus endèmics Nectophrynoides occidentalis, un element molt notable de la fauna d’aquesta sabana.
Les característiques biològiques d’aquest petit amfibi, els adults del qual fan entre 18 i 27 cm de longitud, són estrictament adaptades al medi. La manca de cursos d’aigua que determina el relleu, es tradueix en una viviparitat completa, és a dir, les seves cries, quan neixen, estan totalment metamorfitzades i mesuren de 7 a 8 mm de longitud. La fecundació té lloc entre setembre i octubre, just abans de l’estació seca, i els naixements al juny, un cop ja ha començat l’estació de pluges. Les femelles, tan bon punt són fecundades, s’enterren a les fissures del sòl i del subsòl on romanen, sense menjar, fins a l’arribada de les primeres pluges de l’any següent, entre març i abril. El creixement dels embrions, molt alentit mentre les mares són colgades, s’accelera aleshores fins al moment del naixement de les cries. En aquesta sabana d’altitud, N. occidentalis, molt abundant, pot superar densitats d’un individu per metre quadrat i una biomassa de 15 g/m2.

Composició numèrica i ponderal del poblament d’invertebrats de la sabana dels monts Nimba, a Guinea. Si bé la metodologia de què es disposa no permet calcular amb exactitud el nombre i la biomassa dels petits invertebrats, sí que ha permès observar que aquests organismes són molt més abundants a les sabanes durant l’estació humida que no pas a l’estació seca, quan les condicions es tornen molt més dures. La major part dels invertebrats de les sabanes, però, han desenvolupat adaptacions específiques per afrontar la sequera estacional, especialment esteses entre les espècies que habiten les sabanes més seques, com per exemple la dels monts Nimba. Hi ha espècies que es traslladen temporalment cap a hàbitats amb unes condicions de vida més suportables, especialment a l’ecotò entre el bosc i la sabana. Altres invertebrats resten a la sabana, i sobreviuen al període sec enterrats al sòl en formes de resistència. D’aquesta manera no tan sols queden protegits de la dessecació sinó també dels depredadors i de la intensa escalfor dels incendis que assolen la sabana.
Dades proporcionades per l’autor
Pel que fa als insectes, són nombrosos, sobretot durant l’estació seca, les llagostes, els pregadéus i els dermàpters (insectes bastó). A les espècies endèmiques, la majoria sense ales o amb ales reduïdes, com si es tractés d’una població insular, s’afegeixen formes diverses, normalment alades, provinents de les sabanes de la plana. Durant l’estació de les pluges, en canvi, pul·lulen espècies higròfiles d’opilions, de miriàpodes, de dermàpters i de paneroles, tot plegat moltes desenes d’espècies sovint pròpies d’aquesta sabana.
L’abundància de cargols és una altra característica d’aquest medi, en el qual aprofiten el llarg període de plugim i d’humitat persistent. L’espècie més abundant, Gymnarion grandis, té una closca reduïda en la qual l’adult no es pot retreure, ni per tant sobreviure, durant l’estació eixuta. Els joves nascuts algunes setmanes abans que comenci aquella estació són els que la poden superar, a recer d’una closca en la qual encara caben, fixats contra pedres i roques. L’estratègia dels Gonaxis, dels quals hi ha dues espècies endèmiques, és molt diferent. La seva closca ben desenvolupada permet l’estivació a l’interior del sòl; el primer any en estat juvenil; ja que el seu desenvolupament dura, efectivament, dos anys.
Els cucs de terra, de talla mitjana o petita, són molt nombrosos als gramenets d’altitud de Nimba. La densitat s’acosta als 200 individus/m2, similar a la de les sabanes de la plana. Durant l’estació de les pluges es beneficien de les condicions especialment favorables d’humitat i d’abundància d’humus del sòl.
Els tèrmits tenen un paper anàleg. Nombrosos al peu de cada mota d’herba, són, quasi tots, tèrmits que pertanyen a l’antic gènere Anoplotermes, caracteritzat per l’absència de soldats. El règim alimentari humívor es fa aparent immediatament en observar l’abdomen de les obreres que s’ha tornat negre a causa de la terra absorbida. Una diferència fonamental amb les sabanes de la plana del voltant és el fet curiós que no hi hagi tèrmits fitòfags, cultivadors o no de fongs. Tampoc no hi ha espècies que construeixin nius epigeus, ja que els Anoplotermesnomés formen colònies difoses dins el sòl.
En conjunt, l’originalitat del poblament >animal de la sabana d’altitud dels monts Nimba és causada per un aïllament molt marcat, però també per les condicions especials d’un clima sensiblement menys càlid que el de la plana i on alternen una estació molt seca i una estació molt humida. Aquestes condicions són les que han permès la individualització d’una fauna higròfila que ha implicat moltes espècies endèmiques i un funcionament dominat per la preponderància total de sapròfags i geòfags —tèrmits i cucs de terra principalment— respecte als herbívors.
El poblament animal de la sabana seca de Nylsvley
La sabana de Nylsvley, on va funcionar una estació permanent de recerca en ocasió del Programa Biològic Internacional en la dècada del 1960, ja aleshores havia perdut quasi tota la fauna de grans mamífers, desplaçada per la cria extensiva de bovins. La transformació en reserva permeté reconstituir en part aquesta gran fauna al mateix temps que estudiar la resta del poblament animal, poc modificat.
El clima es caracteritza per una pluviositat anual baixa, entre 600 i 700 mm. A diferència de la majoria de les sabanes tropicals, les temperatures mitjanes mensuals són de 19°C (i màximes de 24°C), però el més important és que té 20 dies anuals de gelades nocturnes. El recobriment de les plantes llenyoses és de prop del 25%; la dominància respectiva permet distingir dues fàcies ben diferenciades, una amb Burkeai l’altra amb Acacia. Totes les espècies animals que hi viuen es troben a les grans sabanes veïnes d’Àfrica oriental però aquí la fauna està molt empobrida.
Els mamífers són representats per 42 espècies: 4 insectívors, 2 lagomorfs, 18 rosegadors, 11 carnívors, 1 tubulidentat i 6 ungulats. Entre els fitòfags falten l’elefant africà (Loxodonta africana), l’hipopòtam (Hippopotamus amphibius) i el búfal africà (Syncerus caffer). Les zebres (Equus zebra, E. burchelli) i les girafes (Giraffa camelopardalis) són poc nombroses. Les altres espècies són pasturadors com el nyu blau (Connochaetes taurinus), el cob dels canyissars (Redunca arundinum), el cob defassa (Kobus defassa) i l’antílop equí (Hippotragus equinus) i brostejadors com el gran cudú (Tragelaphus strepsiceros), el dic-dic de Serengueti (Madoqua kirki) i el raficer comú (Raphicerus campestris). El règim de l’impala (Aepyceros melampus) és mixt però amb un clar predomini de gramínies, mentre que el facoquer (Phacochoerus aethiopicus) i el senglar de riu (Potamochoerus porcus) són omnívors. Els rosegadors petits són representats principalment per Dendromus melanotis, un ratolí arborícola de cua semiprènsil; Tatera leucogaster, el gran jerbu descalç de ventre blanc; Saccostomus campestris, una rata galtuda africana, i Mus minutoides.
La població d’ocells sembla relativament pobra, amb 78 espècies i una densitat de 5,86 individus per ha a la sabana amb Burkea, i 63 espècies i 11,85 individus per ha a la sabana amb Acacia.
Els insectes, en canvi, tenen un paper important entre els consumidors d’herba. Els acrídids són els principals, i consumeixen al voltant del 6% de la producció epigea. Les fulles dels arbres són menjades principalment per erugues, i la defoliació pot arribar alguns anys al 30% de la producció. Les larves de coleòpters intervenen principalment sobre les llavors de lleguminoses, i els hemípters, xucladors de saba, tenen un paper difícil d’avaluar bé que significatiu.
Amb una biomassa abundant, més de 10 vegades superior a la de la fauna epigea, la població hipogea és responsable del consum d’una quantitat important de matèria vegetal. Constitueix, per tant, i de molt, la part principal del món dels fitòfags de la sabana. Els tèrmits són representats per una vintena d’espècies, diversitat sensiblement inferior a la de les sabanes humides de l’Àfrica occidental. Depenent de les fàcies de sabana, vinculades al tipus de sòl, la seva abundància és molt variable ja que passa de 0,16 g/m2 a la sabana amb Acaciaa 3,17 a la sabana amb Burkea. La biomassa mitjana de les poblacions és clarament molt més baixa que la dels tèrmits de Lamto i el consum és també menor: 10 g/m2 i any en comptes dels 27 de Lamto.
Els tèrmits humívors, fet curiós per a una sabana seca, són abundants, en indrets arriben a constituir fins al 68% de la biomassa del grup. Aquesta anomalia s’explica, sens dubte, per la manera molt especial d’alimentar-se d’aquests tèrmits, que aquí consumeixen les radicel·les de les plantes a mesura que es descomponen, sense ingerir el sòl, massa sorrenc, que les envolta.
El poblament animal de la sabana àrida de Fété-Olé
Les sabanes àrides de la regió de Fété-Olé, al Senegal, sempre a Àfrica però a l’hemisferi nord, que han estat objecte d’estudis aprofundits, són un bon exemple d’aquest tipus de sabana que s’acosta ja al subdesert. El clima, caracteritzat per una pluviositat de només 400 mm de mitjana, és molt variable segons els anys. La vegetació té un estrat arbori relativament desenvolupat, de tipus decidu, i un estrat herbaci amb espècies anuals que desapareixen totalment a l’estació seca. A causa de la manca de punts permanents d’aigua la fauna gran mai no hi ha estat gaire abundant. Amb tot, hi vivien girafes, lleons i, fins i tot, esporàdicament, elefants, com també diferents antílops sahelians com l’òrix comú (Oryx dammah), l’addax (Addax nasomaculatus), el sassabi (Damaliscus lunatus) i el búbal (Alcelaphus buselaphus).
Aquesta gran fauna actualment ha desaparegut quasi del tot, al mateix temps que el territori ha estat envaït pel bestiar domèstic gràcies a l’excavació de nombrosos pous. D’ungulats queden el facoquer (Phacochoerus aethiopicus) i tres gaseles (Gazella rufifrons, G. dorcasi G. dama); un lagomorf, la llebre Lepus crawshi; un tubulidentat, el porc formiguer (Orycteropus afer); un primat, la mona vermella o “patas” (Erythrocebus patas), i quatre carnívors: el xacal comú (Canis aureus) i el xacal ratllat (C. adustus), la guineu pàl·lida (Vulpes pallida) i la mofeta africana (Ictonyx striatus).
També s’han comptat 8 rosegadors: l’esquirol de Gàmbia (Heliosciurus gambianus), l’esquirol terrestre africà (Xerus erythropus), les rates Praomys natalensisi Arvicanthis niloticus, els jerbus Taterillus pygargus, T. gracilisi Desmodilliscus brauerii el porc espí (Hystrix cristata); també quatre insectívors, les musaranyes Crocidura sericea, C. lusitanica, C. lamotteii l’eriçó africà de ventre blanc (Erinaceus albiventris), i cinc espècies de quiròpters: Rousettus aegyptiacus, Taphozous perforatus, Nycteris thebaica, Asellia tridensi Tadarida major.
El nombre considerable d’espècies de petits mamífers no ha de disfressar el fet que l’efectiu de cadascuna d’elles és molt baix, fins i tot dels rosegadors. Aquesta pobresa és, d’altra banda, molt variable segons els anys. Augmenta amb el grau d’aridesa, mentre que, en relació estreta amb la producció vegetal, una fecunditat potencial alta permet als efectius remuntar ràpidament quan arriben anys plujosos.
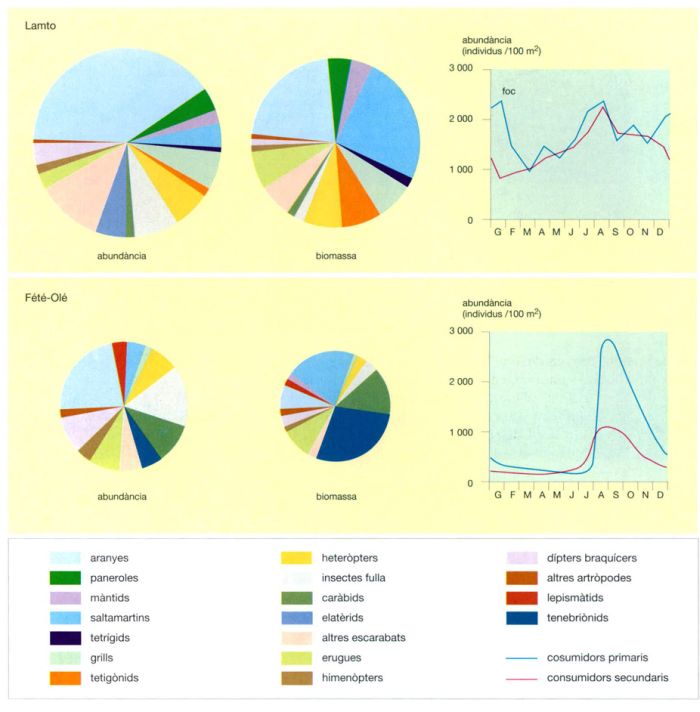
Biomassa i abundància relatives dels principals grups d’artròpodes de l’estrat herbaci en dues sabanes africanes força diferents entre elles, la de Lamto, a la Costa d’Ivori, i la de FétéOlé, al Senegal. La sabana de Fété Olé, molt més àrida, presenta un estrat herbaci format per espècies anuals que a l’estació seca desapareixen completament. Aquest fet explica que tant el nombre d’individus com la biomassa dels artròpodes que hi habiten prenguin valors més baixos que a les sabanes de Lamto, més humides i amb una producció herbàcia més elevada. L’abundància considerable d’alguns grups d’artròpodes, com per exemple les aranyes, no es contradiu amb el fet que la seva biomassa sigui relativament molt menys important, ja que es tracta d’espècies de mida petita. En canvi, grups amb pocs efectius, com per exemple els escarabats tenebriònids, representen una fracció gran de la biomassa total, en tractar-se d’animals també més grans. D’altra banda, l’abundància dels diferents grups en conjunt presenta variacions estacionals molt marcades a les sabanes de Fété Olé. El màxim s’assoleix el mes d’agost, al principi de l’època de pluges. Aleshores el nombre total d’artròpodes (consumidors primaris i secundaris junts) és unes quinze vegades més alt que el mínim del mes d’abril, en plena època de sequera. A Lamto, en canvi, l’abundància no presenta variacions tan marcades al llarg de l’any.
Jordi Corbera, a partir de Gillon, 1974
Entre els insectes, els tèrmits representen una fracció essencial de la biomassa i dels efectius, amb 1 500 a 3 000 g de matèria seca per ha. N’hi ha una vintena d’espècies, però quatre d’elles són molt àmpliament dominants ja que representen el 95% de la biomassa. Aquestes són Psammotermes hybostoma, un tipus de tèrmit amb niu hipogeu característic dels medis sorrencs desèrtics; Odontotermes smeathmanii Macrotermes subhyalinus, totes dues espècies cultivadores de fongs amb niu epigeu en cúpula (de talla gran en el cas Macrotermes) i, finalment, Trinervitermes trinervius, amb niu epigeu en cúpula de talla petita (alguns decímetres), consumidor de gramínies. La densitat de termiters pot superar el nombre de 5 per hectàrea en el cas de Trinervitermes, 2 de Psammotermesi Odontotermesi 0,5 de Macrotermes, però sempre són colònies molt populoses. Si es compara aquesta població de tèrmits amb la de Lamto s’observa que els cultivadors de fongs tenen un lloc clarament més important a Lamto que a Fété-Olé, mentre que els humívors, pràcticament absents a Fété-Olé, són també força més abundants a Lamto.
Els efectius i les biomasses dels insectes no socials, força menys altes que en una sabana menys àrida com la de Lamto, són, a més, extremament variables amb les estacions. Arriben el mes d’octubre (final de l’estació de les pluges) a un màxim quatre vegades superior al mínim de juliol, al final de l’estació seca. Aquestes variacions del conjunt dels artròpodes emmascaren desviacions força més grans en alguns grups com les erugues, els acrídids, els corídids, els ligèids i els elatèrids. També són caracteritzades per la curta durada del període en què la població és abundant. Aquest pic és força diferent de la constància relativa de la població de Lamto, on el mínim és a penes inferior a la meitat del màxim.
El poblament animal del medi hipogeu
Un gradient de S a N a les sabanes de l’Àfrica occidental, des de la Costa d’Ivori fins a Burkina Faso, destaca les conseqüències d’una aridesa creixent sobre la població del sòl. A Lamto, la sabana es pot considerar no sols poc àrida, sinó fins i tot relativament humida, amb 1 300 mm de pluges anuals i una estació seca tan sols de quatre mesos. Més al N, a Ouango-Fitini, dins de la sabana del parc nacional del riu Comoé, les pluges estan més agrupades en el temps, i per això l’estació seca és sensiblement més llarga i rigorosa. A les sabanes del centre de la Costa d’Ivori, al Foró, hi ha una situació intermèdia.
Les poblacions de cucs de terra responen de formes diferents a aquest grau creixent d’aridesa. Aquestes respostes consisteixen, en primer lloc, en fluctuacions estacionals més o menys accentuades de la densitat global: d’oscil·lacions que van de l’ordre del senzill al quàdruple com a màxim a Lamto (fins i tot sovint només del senzill al doble), passen a oscil·lacions que poden multiplicar per vint la població en els moments favorables en comparació amb els menys favorables. Les poblacions també responen per una disminució sensible de la biomassa total màxima, que pot caure dels 30 g/m2 a la sabana humida de Lamto a menys de 5 g a la sabana seca de Comoé. Encara és més notable el canvi de la composició del poblament, on els tipus detritívors i geòfags del sòl superficial, de vida curta i reproducció ràpida, esdevenen majoritaris mentre desapareixen les formes de profunditat, més fràgils i de reproducció lenta. Aquesta adaptació ja existeix a les sabanes intermèdies del Foró on el nombre d’espècies en conjunt disminueix a la meitat, passant de deu a cinc. En una sabana encara més seca com la de Fété-Olé, al N del Senegal, els cucs han desaparegut del tot. Es pot dir, per tant, que aquests animals són característics de les sabanes més humides —és a dir, d’estació seca curta i poc rigorosa— i que les poblacions es modifiquen i després desapareixen tan bon punt el clima es torna àrid, per poc que sigui.
La situació dels tèrmits és molt diferent. D’una manera general, les variacions dels seus efectius són petites al llarg de les successives estacions de l’any: a Ouango-Fitini encara són poc marcades i fins i tot a Fété-Olé, encara que es tracti d’un medi més sec. Hi ha sempre un escurçament notable del període en què es produeixen les eixamenades, molt vinculades a la humitat de l’aire. També es manifesta una diferència en la davallada en profunditat, més gran a les sabanes més àrides. Així, les moles de fongs, a Ouango-Fitini, passen de l’estrat superficial del sòl, a l’estació de les pluges, a una profunditat de 30 i fins i tot 40 cm a l’estació seca. La biomassa total dels tèrmits, com la producció vegetal, disminueix quan augmenta la sequera, però lentament; es passa de 12,6 g/m2 a Lamto, a encara 10 g/m2 al Foró i a 5,2 g/m2 a Ouango-Fitini. A Fété-Olé, tot i l’aridesa d’aquesta sabana, encara hi ha una biomassa relativament important. La composició del poblament, en canvi, es modifica més. Hi ha una forta disminució dels consumidors d’humus (herbívors) mentre que els macrotermitins, recol·lectors de gramínies mortes i cultivadors de fongs, esdevenen molt preponderants. Aquesta tendència també augmenta a les fàcies més seques com a Fété-Olé. Dues característiques de la vida dels tèrmits són un avantatge considerable a les sabanes seques. La primera és l’aptitud d’acumular reserves per als períodes desfavorables en què la vegetació herbàcia desapareix. Per a alguns, com els Trinervitermes, aquestes reserves són brins d’herba tallats, per a altres, com Macrotermes, són les nombroses moles de fongs. El segon avantatge dels tèrmits és el manteniment en els termiters voluminosos d’una temperatura i una humitat pràcticament constants. Associada a la presència permanent d’aliment abundant, aquesta climatització del termiter els permet mantenir una vida activa durant tot l’any amb efectius relativament poc variables, al contrari de quasi tots els altres organismes.
La vida en els sòls de la sabana, dominada pels cucs de terra a les sabanes pre-forestals humides, és completament diferent a les sabanes seques i àrides, on els tèrmits esdevenen, i de lluny, els actors principals del funcionament gràcies a la vida social i a les construccions que saben edificar.
Fluctuacions estacionals en les colònies de tèrmits
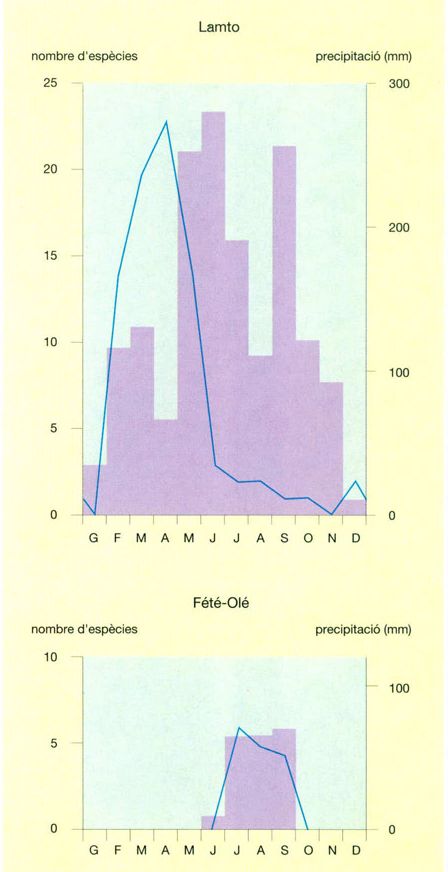
Jordi Corbera, a partir de Goodall, 1983
El nombre d’espècies de tèrmits que s’eixamenen cada mes a les sabanes de Lamto, la Costa d’Ivori, i de Fété-Olé, al Senegal, mostra una relació molt clara amb la pluviositat mitjana mensual (representada per la línia blava). La producció d’eixams d’individus alats és l’expressió més evident de les fluctuacions estacionals de les colònies de tèrmits. La majoria d’espècies produeixen els alats a l’estació seca, i l’eixamenament es dona a principi de l’estació humida, tal com s’observa a Lamto. Normalment té lloc després d’un ruixat fort, o bé un o dos dies després, a la posta del sol o a la nit. L’alliberament dels alats de les diferents espècies al mateix temps, quan la humitat de l’aire i del sòl és l’apropiada, segurament té un valor adaptatiu, ja que els depredadors no poden caçar-los tots. En climes més estacionals, l’eixamenament dels alats encara depèn més de les precipitacions, com passa per exemple a Fété-Olé. Al llarg de tota l’època seca, ni un sol alat emprèn el vol. D’altra banda, el nombre d’espècies de tèrmits, com també les seves densitats i biomasses mitjanes, presenten diferències entre els diversos grups alimentaris (xilòfags, cultivadors de fongs, farratjadors o humívors), i no és pas igual a Fété-Olé i a Lamto. En aquesta darrera sabana, més humida, s’han comptat més de 9 milions d’individus per hectàrea, la majoria cultivadors de fongs (gairebé el 50%) o de règim humívor (el 30%). En canvi, a Fété-Olé no s’arriba als 2 milions i mig per hectàrea, més de la meitat farratjadors i una bon part (40%) cultivadors de fongs, mentre els humívors són molt poc comuns. Els tèrmits xilòfags no són especialment abundants en cap de les dues sabanes, ja que tant a Lamto com a Fété-Olé amb prou feines arriben al 4% del nombre total d’individus.
La fauna sabanícola sud-americana
L’Amèrica del Sud, igual que Àfrica, és situada en una franja ampla dins la zona intertropical però, en canvi, és en gran part recoberta per selves, mentre que les sabanes només ocupen una superfície relativament limitada. Tanmateix, el fet capital de la història biogeogràfica de l’Amèrica del Sud és que ha estat aïllada de l’Amèrica del Nord (i de totes les altres terres emergides) durant molt de temps, des del principi del Cenozoic fins a la meitat del Pliocè, és a dir durant el llarg període en què es van diferenciar els mamífers moderns. La formació, relativament recent, de l’istme de Panamà, estret i muntanyós, no ha permès el pas d’un gran nombre d’espècies; la seva vegetació de denses selves plujoses ha estat particularment desfavorable al pas d’espècies pròpies de medis predominantment herbosos. A l’Amèrica del Nord, per altra banda, els medis amb predomini de la vegetació herbàcia són situats fora de la zona intertropical, i per tant no aixopluguen una veritable fauna de sabana que recordi l’africana. La fauna de l’Amèrica del Sud és, per tant, particularment pobra en grans mamífers de sabana i, a més, els que hi ha, tenen clares afinitats amb els de la selva plujosa. La fauna de les sabanes de l’Amèrica del Sud, en definitiva, és totalment diferent de la de les sabanes africanes. Unes i altres han estat aïllades des de fa molt de temps entre elles i només tenen en comú molt poques espècies; l’esplugabous (Bubulcus ibis), que té una capacitat de desplaçament suficient per a travessar l’Atlàntic, seria un d’aquests escassos exemples.
Les vastes superfícies, la diversitat del medi i l’aïllament han permès una diferenciació, en el mateix lloc, que porta a un endemisme considerable per a tots els grups taxonòmics de talla petita, des dels ocells, rèptils, amfibis i rosegadors fins a diversos invertebrats. De fet, les poblacions d’ocells i de rats-penats, per exemple, són d’una riquesa excepcional, com les dels rèptils, els amfibis i molts altres grups. A més a més, són essencialment formes forestals a causa de la predominància de les selves a totes les zones tropicals del continent. Per a moltes espècies, les sabanes no són res més que un hàbitat colonitzat secundàriament.
Els vertebrats sabanícoles i les seves àrees de distribució
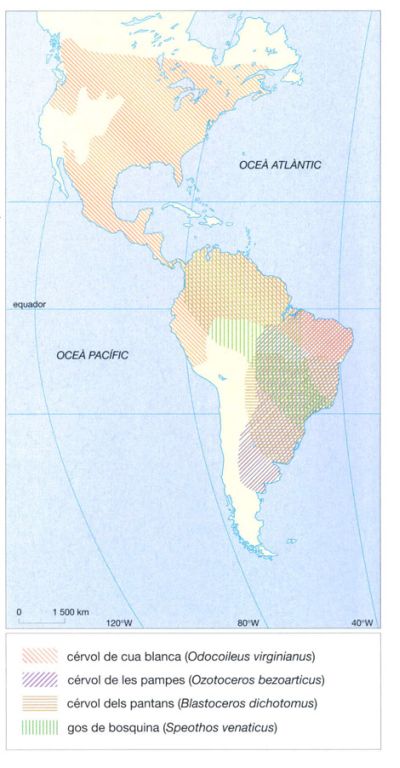
Àrea de distribució de quatre espècies de mamífers de les sabanes sud-americanes. El cérvol de cua blanca (Odocoileus virginianus) s’estén pràcticament per tota l’Amèrica del Nord, i per la part més septentrional de l’Amèrica del Sud. Si bé habita preferentment els voltants de les zones forestades, ocupa, també, les sabanes que es troben dins la seva àrea de distribució. El cérvol de les pampes (Ozotoceros bezoarticus) viu a les praderies obertes de poca altitud del SE de l’Amèrica del Sud, incloent-hi la zona de les pampes, d’aquí el seu nom. L’àrea de distribució del cérvol dels pantans (Blastoceros dichotomus) se superposa en gran part amb la del cérvol de les pampes, però, a diferència d’aquest darrer, prefereix els aiguamolls, les sabanes humides d’herbes altes i els marges dels boscos humits. El gos de bosquina (Speothos venaticus) habita els boscos i les sabanes humides de la zona central i septentrional de l’Amèrica del Sud, i sovint se’l pot veure vora els cursos d’aigua.
IDEM, a partir de fonts diverses
Una primera característica remarcable de la fauna de les sabanes sud-americanes és l’absència de bòvids, com els antílops tan característics de les sabanes africanes. Els únics ungulats grans del continent sud-americà són cèrvids, la majoria de les espècies dels quals són forestals. Només el cérvol de cua blanca (Odocoileus virginianus) es pot considerar una forma típicament sabanícola. La seva àrea de distribució cobreix tota l’àrea septentrional del continent sud-americà, principalment les sabanes de Veneçuela, Colòmbia i Guyana, però també les praderies d’Amèrica del Nord d’on és originària (és absent, en canvi, de les zones no forestals del Brasil, com per exemple els “cerrados”). Molt tolerant pel que fa al biòtop, sembla que hauria arribat a l’Amèrica del Sud des de la seva àrea d’origen seguint els medis oberts de la zona costanera de l’istme centramericà. La discontinuïtat relativa de la seva àrea, vinculada a la seva afinitat pels medis herbacis, ha determinat la formació de moltes races geogràfiques. En general, els individus d’O. virginianusde l’Amèrica del Sud són més petits que els de l’Amèrica del Nord, amb un pes inferior als 50 kg en comptes dels 200 kg de les formes canadenques. La seva alimentació consisteix sobretot en fulles i branques de diverses dicotiledònies, en particular de Mimosa, i també fruits de les sabanes, entre altres els de les palmeres Copernicia. En la seva dieta, en canvi, les gramínies tenen un paper poc important.
Hi ha dos cèrvids més que freqüenten els medis herbacis sud-americans. Com el seu nom indica, el cérvol de les pampes (Ozotoceros bezoarticus) és una espècie arribada a les sabanes des de la part meridional del continent, on els medis herbacis ocupen extenses superfícies fora de les regions tropicals. L’àrea de distribució d’aquest animal s’estén des de l’Argentina fins a diverses comunitats vegetals no boscoses de la zona central del Brasil, però no arriba a les sabanes de Veneçuela. És un animal elegant, de talla petita, amb només 70 cm a l’alçada de la creu. El cérvol dels pantans (Blastocerus dichotomus) és de talla més gran, ja que la seva alçada arriba als 130 cm a la creu i el seu pes és de 100 kg. Està adaptat a les extenses zones inundables del centre del continent, però la seva àrea s’estén també a la zona dels “cerrados”. L’adaptació als terrenys pantanosos es tradueix en peülles amples i relligades per una membrana. Té les banyes molt desenvolupades. La caça activa ha fet disminuir molt les poblacions, tant de cérvol de les pampes com de cérvol dels pantans, i per això actualment es pot considerar que són uns animals en perill d’extinció, tot i que antigament eren molt abundants.
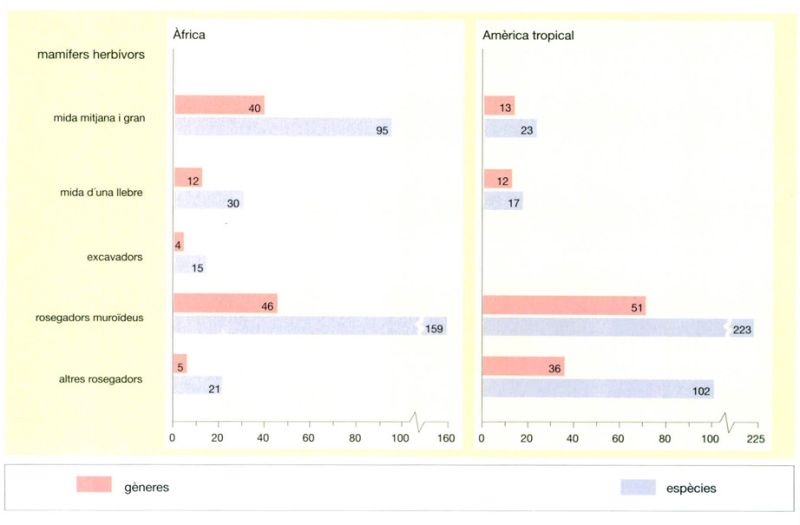
Comparació entre el nombre de gèneres i espècies de mamífers herbívors terrestres a les zones tropicals d’Àfrica i d’Amèrica. Els mamífers de mida mitjana i gran són molt abundants al continent africà, però en canvi escassegen a l’Amèrica tropical. Aquest fet s’explica perquè bona part de l’Amèrica del Sud és —o ha estat fins fa molt poc— recoberta per selves o vegetació forestal, de manera que els hàbitats de sabana amb predomini d’una vegetació herbàcia, on més abundants són els grans mamífers, ocupen extensions molt més petites que al continent africà. El fet que l’Amèrica del Sud quedés aïllada de la del Nord i de la resta de terres emergides fins a temps geològics molt recents (mitjan Pliocè) dificultà el pas de les espècies animals cap al S del continent. Els que més difícil ho tingueren foren els animals de sabana, ja que, a més a més d’haver de creuar per l’estret istme de Panamà, havien de travessar grans extensions de selves tropicals abans d’arribar a les sabanes. Una altra característica remar-cable de les zones tropicals americanes és l’abundància d’espècies de rosegadors i la manca de mamífers excavadors.
IDEM, a partir de Keast, 1972
Els pècaris semblen petits senglars i són igualment herbívors, o més aviat omnívors. Són animals de la gran selva i també freqüenten les comunitats obertes i les sabanes, en particular, el pècari de collar (Tayassu tajacu), que és molt abundant. Els pècaris viuen en ramats que poden ser molt nombrosos, i que es desplacen molt. Tenen una considerable longevitat (25 anys) i una fecunditat gran. Els seus principals depredadors són els jaguars i, dels joves, els ocelots.
Els carnívors, sobretot si són de mida gran, ocupen a l’Amèrica del Sud àrees geogràfiques dilatades i medis més diversificats que els herbívors que estan estretament lligats al tipus de vegetació. El nombre dels que entren a les sabanes és també considerable, encara que el seu hàbitat més freqüent sigui la selva plujosa. És el cas del jaguar (Panthera onca), el fèlid més gran d’Amèrica. La seva àrea de distribució arriba a la Patagònia, pel S, i a Arizona, pel N; exclou només la zona dels Andes. És un animal de cap ample i cos massís, que pot pesar més de 150 kg. El seu pèl recorda el del lleopard, amb taques clares perfilades de negre. Caçador nocturn, s’alimenta principalment de pècaris, cérvols, capibares i altres rosegadors grossos. També captura micos i tapirs a la selva, a més de tortugues aquàtiques, caimans i fins i tot peixos, ja que és un bon nedador.
El jaguarundí (Felis yagouaroundi) té la mateixa àrea de distribució que el jaguar. Més petit, més esvelt, amb el cap més estret, de cua molt llarga i amb el pelatge uniforme —però variable segons els individus—, s’alimenta de preses força més petites, tant de mamífers com d’ocells. La seva dependència de la selva és molt elevada, igual com passa amb molts altres gats salvatges, en particular l’ocelot (Felis pardalis), animal d’un metre de llargada (cua inclosa) i amb un pes d’uns 10 kg. El seu pelatge, tacat com el del jaguar, que li ha valgut un gran èxit entre els pelleters, ha estat a punt de ser la causa de la seva perdició. El felí més freqüent a les sabanes americanes és, però, el puma (Felis concolor) encara que no n’és ni de bon tros exclusiu, ja que la seva àrea de distribució abasta pràcticament tot el continent americà, des del territori de Yukon i Nova Escòcia, al N, fins a la Patagònia, i des del nivell de la mar a l’alta muntanya.
Al costat dels fèlids, a l’Amèrica del Sud tropical hi ha altres carnívors que es localitzen també preferentment a la selva. Alguns, a més, freqüenten les comunitats vegetals seques i embardissades del centre del continent i fins i tot la pampa argentina. Són principalment la guineu menjadora de crancs (Cerdocyon thous), la guineu argentada (Urocyon cinereoargenteus) i el gos de bosquina (Speothos venaticus).
Els opòssums o sarigues (Didelphis marsupialis, D. albiventris, etc.) són representants del grup dels marsupials, a la vegada abundants i àmpliament distribuïts per tots els ambients de l’Amèrica del Sud i fins i tot de Mèxic i del S dels Estats Units en el cas d’algunes espècies. Fan uns 40 cm de llargada, sense comptar la cua, mancada de pèl, que té aproximadament la mateixa longitud. Són omnívors; consumeixen fruits, insectes i petits vertebrats. Els opòssums són força coneguts per l’original forma que tenen de transportar les seves cries quan ja han deixat la bossa marsupial: penjades per les cues de la cua de la seva mare, que aquesta recorba sobre l’esquena.
A més dels animals de règim carnívor veritable, a l’Amèrica del Sud hi ha zoòfags especialitzats en el consum d’insectes. L’ós formiguer (Myrmecophaga tridactyla) viu en els medis de sabana llenyosa i de bosc clar. És un animal de coll allargat i musell llarg i punxegut, amb una llengua llarga i vermiforme, de més de 50 cm, que amb la saliva pot aglutinar centenars de formigues i tèrmits, tal com fan a l’Àfrica els pangolins (Manis) i el porc formiguer (Orycteropus afer). Detecta la presència de preses gràcies a un olfacte molt desenvolupat i esventra els nius de formigues i tèrmits gràcies a unes potents urpes. Amb les urpes també es defensen dels seus depredadors tot dreçant-se sobre les potes de darrere com els óssos. El seu pelatge molt dens és una protecció eficaç contra les mossegades i les picades de les formigues.
També hi ha altres mamífers que mengen insectes: són els tatús o armadillos, que es caracteritzen per la forta cuirassa articulada que els defensa. El seu règim és, de fet, força omnívor, ja que s’alimenten de vegetals i de diverses preses petites. La seva originalitat és poder enrotllar-se en forma de bola protegint així les parts toves del cos; el tatú de bola o “bolita” (Tolypeutes tricinctus) forma una esfera quasi perfecta. Els tatús són excavadors que poden cavar-se un cau ràpidament. El tatú pròpiament dit (Dasypus novemcinctus) té una àrea de distribució molt vasta que arriba fins i tot als Estats Units, pel nord, mentre que les altres espècies estan més localitzades i són presents només a l’Amèrica del Sud, com l’armadillo gegant (Priodontes maximus), l’àrea del qual s’estén per la major part de l’Amèrica del Sud a l’E dels Andes. Es tracta d’una espècie de gran talla (pot arribar al metre i mig, inclosa la cua) que viu en caus relativament profunds excavats en nius de tèrmits, dels quals s’alimenta.
Els mamífers de talla petita com els rosegadors, al contrari del que passa amb els grans, han pogut diferenciar-se en formes nombroses gràcies a la diversitat de nínxols ecològics. Encara són sobretot les zones de selva les que han permès especiacions; el cas dels rats-penats és el més típic, amb una riquesa taxonòmica força superior a la de tots els altres continents. La mateixa observació es pot fer a propòsit de les poblacions d’ocells, de les quals a l’Amèrica del Sud n’hi ha més de 3 000 espècies enfront de les 1500 de l’Àfrica, però només 521 enfront de 708 es troben a la sabana.
Els invertebrats sabanícoles i les seves àrees de distribució
La situació dels invertebrats de la sabana sud-americana és semblant a la dels petits vertebrats. Una diversitat específica molt alta i un endemisme molt elevat caracteritzen el conjunt de les poblacions del continent, però les de les sabanes són molt pobres. Tot i així, contenen formes molt originals, sovint compartides amb les selves. És el cas particular d’algunes formigues. Entre les nombroses espècies hi ha dos grups particularment remarcables.
Els cultivadors de fongs, semblantment a alguns tèrmits africans, tallen fulles que porten en llargues fileres cap al seu niu. Amb aquestes fulles, barrejades amb saliva, fan una pasta sobre la qual creix un fong que els serveix d’aliment. Així es resol la dificultat de digestió que planteja als animals el consum d’aliments massa rics en cel·lulosa i massa pobres en proteïnes. Moltes espècies d’aquest tipus consumeixen fulles de dicotiledònies i en malmeten les plantacions, mentre que altres, en canvi, exerceixen la seva activitat sobre l’herba graminoide de la sabana. En concentrar en els nius els elements minerals continguts en aquestes fulles, afavoreixen la formació de bosquetons al mig de la sabana, tal com també fan alguns tèrmits africans.
L’altra categoria és representada per les anomenades formigues legionàries, com Neivamyrmex nigrescens. Aquestes formigues, cegues però amb un olfacte molt desenvolupat, són, pel seu elevat nombre, temibles depredadors. Repartides en tots els medis des del S del continent fins als Estats Units, recorren el terreny en fileres immenses de centenars de milers d’individus. Canvien contínuament de nius, deixant cada dia fileres de caça. Les úniques parades llargues que fan, de dues a tres setmanes, estan vinculades a la nimfosi de les larves. És sorprenent la convergència entre aquestes formigues i les d’Àfrica, els dorilínids africans del gènere Anomma.
Els tèrmits, menys nombrosos que a l’Àfrica, també són presents a l’Amèrica del Sud i algunes espècies construeixen termiters que poden assolir grans dimensions. Contràriament a les formes que dominen a les sabanes africanes, aquests tèrmits no són cultivadors de fongs sinó humívors, alguns constructors de termiters i altres no. Les construccions que fan han donat lloc en alguns sectors de la sabana a una topografia molt particular, els “murundus”, paisatges caracteritzats per la presència de nombrosos túmuls de terra, d’un a tres metres d’alçada, construïdes sobre una superfície perfectament plana, fenomen semblant al que s’ha observat a l’Àfrica.
La fauna sabanícola australiana
Austràlia pot acollir poblacions animals molt importants i molt variades gràcies a la seva extensa superfície i a la diversitat del seu relleu, del seu clima i de la seva flora. De tots els continents, és el més aïllat de la resta i el que ho ha estat de fa més temps. La seva fauna presenta així mateix una originalitat excepcional que es manifesta en tots els grups taxonòmics i de manera encara més palesa en els mamífers. Més complicat resulta referir-se més específicament a la fauna sabanícola, ja que la major part d’Austràlia no està compresa al si de la zona intertropical, sinó que el seu centre se situa a la latitud del Sàhara i queda sotmesa a climes desèrtics o subdesèrtics que afavoreixen més aviat les plantes llenyoses que les herbàcies. Aquests climes, amb nits força fredes, són molt diferents dels de les zones tropicals. Només la franja de territori compresa entre les selves i els boscos monsònics del N i NE del país, i els espais subdesèrtics del centre es poden considerar sabanes tropicals del mateix tipus que les d’altres continents, en les quals els animals depenen en bona part de les gramínies de l’estrat herbaci.
A les zones més temperades, i especialment a la zona del continent, nombroses artigades i aclarides dels boscos han permès que es desenvolupessin formacions herbàcies secundàries —no pròpiament sabanes—, destinades a la cria del bestiar, a les quals s’han adaptat tot seguit moltes espècies autòctones d’animals herbívors. A l’inrevés, la introducció continuada d’herbívors, primer bovins però després, principalment, ovins i conills, ha degradat considerablement les pastures i ha reduït les possibilitats de supervivència de la fauna autòctona. La caça i la multiplicació de gossos i de gats ha completat aquesta acció. En conjunt, doncs, les poblacions animals d’Austràlia tenen una originalitat excepcional. Dins el món de grans mamífers, en concret, res no pot recordar el que s’observa a l’Àfrica ni amb prou feines de la fauna de l’Amèrica del Sud, que tanmateix comparteix els marsupials amb Austràlia. Menys aparent en els altres grups taxonòmics, l’especificitat de les poblacions no és menor i correspon a un grau molt alt d’endemisme. És difícil, en canvi, al contrari del que passa a l’Àfrica, definir una veritable fauna sabanícola.
Els marsupials sabanícoles i les seves àrees de distribució
Originàriament, a les sabanes australianes i formacions herboses afins, naturals o secundàries, avui explotades per enormes ramats d’animals domèstics, no hi havia altres mamífers que marsupials i, més en concret, els únics herbívors presents eren cangurs (els grans) i ualabis (els petits).
Bé que els cangurs i altres marsupials colonitzen a Austràlia tots els medis, moltes espècies es distribueixen preferentment a les sabanes, herboses o llenyoses, del N del país. Es tracta de l’euro o “wallaroo” (Macropus robustus), el cangur antilopí (M. antilopinus), el ualabi àgil (M. agilis), i els ualabis de roca, més localitzats, Petrogale penicillata, P. brachyotis, P. burbidgeii Peradorcas concinna. Aquests darrers, com també Macropus robustus, són espècies més aviat dependents de les zones muntanyoses i rocalloses; a les zones més seques i espinoses també es troben ualabis de cua unglada, com Onychogalea unguifera, ualabis llebre, com Lagorchestes conspicillatus, i el cangur roig (Macropus rufus). Algunes altres espècies, com el ualabi de cua de fuet (Macropus parryi) i el cangur gris (Macropus giganteus), no són formes de la regió tropical, però hi arriben de forma local estenent la seva àrea cap al N al llarg de la costa oriental.
A aquestes espècies autòctones cal afegir el búfal aquàtic (Bubalus bubalis), una espècie domèstica que ha estat introduïda i que prospera en llibertat a les sabanes de la plana costanera septentrional; per la seva banda, a les regions més eixutes són els dromedaris (Camelus dromedarius), també introduïts, els que s’han assilvestrat i es multipliquen.
A Austràlia no hi havia animals zoòfags de l’ordre dels carnívors o dels insectívors. Els nínxols ecològics corresponents eren ocupats per marsupials. Entre aquests es poden citar els de la família dels peramèlids o bandicuts, per exemple Isoodon auratus, un dels bandicuts de morro curt. És un animal de musell allargat (bé que no tant com els altres bandicuts), d’uns 30 cm de llarg que s’alimenta de rosegadors i d’insectes. Els ratolins marsupials de cap aplanat, Planigale, de la família dels dasiúrids, són, al contrari, molt petits (menys de 10 cm) i pesen només alguns grams. Tenen un crani aplanat que els permet esmunyir-se en les escletxes de les roques. El règim alimentari és insectívor.
A més d’aquests carnívors autòctons es troba un cànid, el dingo (Canis familiaris dingo), que envaí el continent portat pels primers colons humans, fa diversos mil·lennis. Intel·ligent, s’ha naturalitzat ràpidament i ha esdevingut el principal depredador de cangurs i ualabis. També és un enemic temut dels ramats, sobretot dels d’ovins, i ha estat sotmès a una caça intensa per mantenir els seus efectius per sota d’una xifra que pugui considerar-se perillosa.
Els ocells sabanícoles i les seves àrees de distribució
Tret dels corredors o ratites, els ocells, per la seva aptitud de volar, han pogut arribar a Austràlia després que aquest continent se separés de les altres terres emergides. Per tant, constitueixen poblacions amb un aïllament taxonòmic menor que el dels mamífers, però tot i així dotades d’una originalitat indiscutible. El més gros de tots, l’emú (Dromaius novaehollandiae), no és pas un ocell volador sinó corredor, un casuariforme d’aproximadament 1,80 m d’alt i un pes que pot superar els 50 kg. Es troba arreu del continent australià (també en els territoris subdesèrtics i també en la part no tropical) excepte als boscos i a les zones ara massa antropitzades. L’emú viu en ramats petits que no paren de moure’s, poden córrer ràpid i fins i tot nedar. Com l’estruç, té un règim alimentari molt variat, sobretot herbívor, però que també inclou insectes. L’emú és molt aficionat al gra i pot causar greus perjudicis a les collites de cereals, cosa que ha ocasionat que se’l combatés acarnissadament. La femella pon els ous en un niu rudimentari fet de fullaraca apilada a terra. Cada posta és sovint de més d’una dotzena d’ous, cadascun dels quals pesa més de 500 g. El mascle els cova durant unes vuit setmanes i els pollets, amb un plomatge ratllat característic, es desenvolupen ràpidament, bé que es queden durant més d’un any sota la protecció dels adults.
El nombre elevat de vegetals llenyosos, fins i tot a les zones subdesèrtiques, ha permès el desenvolupament d’una fauna ornítica rica. Les formes més notables, com els ocells del paradís (família paradisèids), els ocells lira (Menura novaehollandiae, M. alberti) o els casuaris (Casuarius), són animals més aviat de selva. En canvi els curiosos ocells d’enramada (família ptilonorrínquids), els mascles dels quals estableixen closos nupcials, de vegades molt complicats, on acumulen objectes diversos susceptibles d’atraure l’atenció i l’interès d’una companya, es troben en medis tropicals més descoberts. Lloros, cotorres, cacatues, periquitos, en una paraula, tota la varietat dels psitàcids, són els ocells més importants d’Austràlia per la seva abundància. Encara que la seva diversitat és gran, la seva morfologia és molt uniforme, amb un bec molt robust que els permet obrir llavors dures i una llengua carnosa que els fa capaços d’imitar la veu humana. Tot i que la majoria de les espècies són forestals, n’hi ha unes quantes que viuen fora de la selva. Totes aquelles espècies que, com els periquitos (Melopsittacus undulatus), coneguts a Austràlia com a “budgerigars”, i d’altres, viuen en les zones semidesèrtiques es reuneixen en estols nombrosos i sorollosos per arribar als punts d’aigua durant l’estació seca. Llur àrea de distribució s’ha estès gràcies a l’activa multiplicació d’aquestes espècies i a la seva colonització de noves terres.
Els rapinyaires són igualment nombrosos i tenen un paper important al si d’uns ecosistemes on els mamífers carnívors són escassos i en perill d’extinció. El més conegut és sens dubte el baza emplomallat (Aviceda subcristata), consumidor de petits vertebrats i d’insectes, però també de fruits, com també molts altres falcons, principalment el falcó comú australià (Falco cenchroides), el falcó bru australià (F. berigora), el falcó gris australià (F. hypoleucos) i les formes ornitòfagues, com el falcó negre australià (F. subniger), i una raça particular de falcó pelegrí (F. peregrinus). També hi ha diversos esparvers, tots de règim molt eclèctic. Les àguiles són representades per la petita àguila calçada australiana (Hieraaetus morphnoides) i l’àguila de cua en tascó (Aquila audax), capaç d’atacar des de cangurs petits o ocells grans fins a llangardaixos, però que també consumeix carronyes en el lloc dels voltors, que són absents a Austràlia. Entre els rapinyaires nocturns hi ha diferents òlibes dels gèneres Tyto(T. alba, T. novaehollandiae), extensament difós per tot el continent australià, i Ninox(N. rufa, N. boobook, N. connivens), de mides molt diverses.
Els rèptils i els insectes sabanícoles i les seves àrees de distribució
Pel que fa a la resta de la fauna, llangardaixos i serps ofereixen una gran diversitat, en particular a la zona subdesèrtica. Molts amfibis depenen de les aigües temporals i estan adaptats a una rigorosa estació seca. Pràcticament tots plegats pertanyen a tàxons endèmics.
També la fauna d’insectes és molt característica. Els acrídids són abundants, fins i tot en els sectors àrids. Pertanyen a espècies i també a gèneres endèmics però, per a un profà, el seu aspecte és el d’una llagosta vulgar. Els coleòpters copròfags, en canvi, es troben en una situació molt especial. La fauna d’aquests insectes era molt reduïda a Austràlia a causa de la manca de grans ungulats i estava especialitzada a atacar i reciclar els petits cagallons dels cangurs. La introducció en massa de bestiar domèstic, bovins i ovins, ocasionà una acumulació considerable d’excrements que necessitaven setmanes per a descompondre’s i provocaven la formació d’eixams de mosques i d’altres dípters. Per això s’introduïren moltes espècies al·lòctones d’escarabats copròfags, provinents de Sud-àfrica i de la conca mediterrània, per combatre aquestes acumulacions. Moltes d’aquestes espècies s’han aclimatat i han proliferat.
La fauna sabanícola asiàtica i malgaixa
Una gran part de les sabanes de l’Àsia meridional i de Madagascar són secundàries. Això fa, per una banda, que la fauna pròpiament sabanícola sigui més pobra que en altres continents com Amèrica, Austràlia i, sobretot, Àfrica i, per una altra, que hi siguin presents espècies que més aviat són de selva, o si més no de clariana o de vorada de selva. La situació, tanmateix, és una mica diferent a l’Índia, on l’extensió de les sabanes (i també la de boscos monsònics, més esclarissats que la selva plujosa), ja abans de la intervenció humana, era més gran que al sud-est asiàtic o a Madagascar.
La fauna de les formacions sabanoides índies
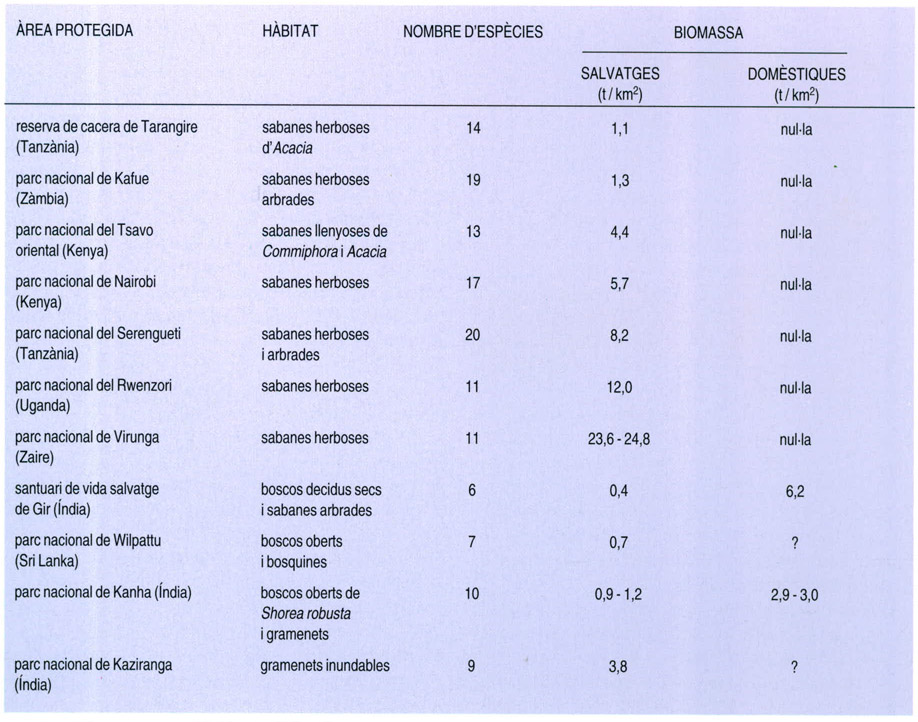
Comparació de les biomasses d’ungulats salvatges i domèstics d’alguns parcs nacionals de les sabanes d’Àfrica i d’Àsia meridional. Les sabanes tropicals ofereixen un hàbitat excel·lent als herbívors pasturadors i brostejadors, tant de mida gran com petita, i ja siguin salvatges o domèstics. Els més comuns són els ungulats, especialment abundants, tant en nombre d’espècies com d’individus, a les sabanes seques del continent africà. Tanmateix, la biomassa corresponent als grans ungulats només pren valors elevats en algunes zones protegides de les sabanes africanes, especialment les de la zona oriental i meridional del continent, ja que, tal com ha succeït a l’Àsia tropical, la pressió antròpica ha reduït considerablement les poblacions salvatges que en altres temps vagarejaven lliurement per les extenses pastures verges. Un cop eliminats o reduïts els ramats salvatges, els domèstics ocuparen el seu lloc i acabaren fent-se dominants, tal com mostren, per exemple, els valors elevats de biomassa corresponents al bestiar domèstic de les zones protegides de Gir (6,2 t/km2) i de Kanha (2,9-3 t/km2), comparats amb els de les poblacions salvatges (0,4 i 0,9-1,2 t/km2, respectivament).
Bourlière, 1983
La fauna, especialment rèptils (sobretot serps), ocells, rosegadors i grans herbívors de les formacions sabanoides índies, és força rica. Si es té en compte els medis de bosc clar que contenen alguns parcs nacionals, el nombre d’espècies d’ungulats sembla prou alt, però es troba molt lluny de la diversitat observada a les sabanes d’Àfrica. La biomassa que hi representen aquests animals és igualment força més baixa que la de les poblacions d’ungulats africanes, tot i que que als parcs nacionals indis hi viu també una quantitat considerable de caps de bestiar domèstic.
Entre els grans herbívors, el bestiar domèstic (ovelles, cabres i camells) destaca per la seva gran densitat, i té un paper molt significatiu en el manteniment, i també en la degradació, d’aquestes sabanes llenyoses, ja que s’alimenten d’arbres i arbustos espinosos. Entre els ungulats salvatges es poden trobar el sambar (Cervus unicolor), el nilgai (Boselaphus tragocamelus), l’antílop negre (Antilope cervicapra), el txusinga (Tetracerus quadricornis), l’axis (Axis axis), la gasela de l’Índia (Gazella benetti) i el barasinga (Cervus duvauceli), la densitat dels quals, a les àrees on no entren en competència amb els animals domèstics, sovint supera els 50 caps per km 2.
A les zones que corresponen més ajustadament a la noció de sabana, únicament hi ha tres espècies d’ungulats: el nilgai, el txusinga i l’antílop negre. El nilgai ( Boselaphus tragocamelus), com suggereix la seva denominació científica genèrica, té alhora trets de brau (‘bos’) i de cérvol (‘elaphus’), pesa prop de 200 kg, i el seu pelatge és de color bru agrisat, amb una anella blanca a les potes, per damunt de les peülles. La femella no té banyes i les del mascle són relativament curtes. El nilgai és un menjador de fulles, fruits i herba, i per tant pastura i brosteja. Les poblacions del nilgai no es pot dir que estiguin en perill d’extinció, per tal com es beneficien de la rarificació del tigre, el seu principal enemic, i del respecte de les poblacions índies. El txusinga o antílop de quatre banyes (Tetracerus quadricornis) té la curiosa característica d’estar dotat al front de quatre banyes punxegudes, les dues més petites en posició anterior, just damunt de les òrbites. A l’Índia està protegit i s’ha mantingut en abundància tot i la superpoblació humana, encara que les diferents poblacions es troben cada cop més isolades entre elles. L’antílop negre (Antilope cervicapra) viu en harems que poden ser de moltes desenes d’individus. És de mida mitjana, amb una alçada de 80 cm a la creu i un pes de 30 a 35 kg. S’alimenta de gramínies ja que viu sobretot en medis descoberts. És un animal extremament ràpid, capaç de fer salts enormes. La bellesa de les seves banyes ha perjudicat molt l’espècie, que ha esdevingut escassa fora de les reserves.
És del tot natural que l’elevada densitat d’herbívors als boscos espinosos comporti la presència d’un gran nombre de depredadors, com guineus (Vulpes bengalensis, V. vulpes), xacals comuns (Canis aureus), hienes ratllades (Hyaena hyaena), tigres ( Panthera tigris) i lleons asiàtics (P. leo persica). La creixent pressió de la urbanització, mitjançant l’esclarissament de la vegetació natural, ha donat com a resultat una ràpida disminució de les poblacions, tant d’herbívors com de tigres. Tanmateix, de les 14 reserves de tigres del país, dues es troben dintre del la regió de bosc espinós de Sariska (prop de Jaipur) i Ranthabore (prop de Sawai Madhopur). Un altre santuari dins de l’àrea de bosc espinós de Gujarat constitueix un refugi per al famós lleó del Gir, la subspècie endèmica esmentada més amunt. En el Rann de Kutch, a l’extrem occidental dels boscos sabanoides espinosos, viu un altre animal endèmic i amenaçat, l’ase salvatge (Equus hemionus).
Cap gran felí de l’Índia, tret potser del localitzadíssim lleó de Gujarat, no pot ser considerat com un animal pròpiament de sabana, ja que tant el tigre com el lleopard depenen de la selva. Aquesta pobresa de la gran fauna pròpia dels medis oberts ha estat esgrimida com una prova de l’absència de veritable sabana primitiva. A les zones subdesèrtiques, en canvi, potser hi viuen encara alguns guepards (Acinonyx jubatus). També hi ha cànids, com el cuon o gos salvatge asiàtic (Cuon alpinus), amb un pes proper als 20 kg, que viu en ramats organitzats amb una manera de caçar semblant a la dels licaons d’Àfrica; ha esdevingut escàs i les gossades estan reduïdes a alguns individus. El llop (Canis lupus pallipes), representat per una subspècie més petita que les de regions més septentrionals, com també la hiena ratllada (Hyaena hyaena), amb una àrea que s’estén fins a l’Àfrica, també són presents a l’Índia. La situació dels mamífers de mida més petita és més favorable. A les regions meridionals, també molt antropitzades, subsisteixen nombroses espècies de rosegadors, d’insectívors i de mangostes que pertanyen a la fauna original.
L’Índia, com Austràlia, és situada parcialment fora de la zona intertropical. Fins i tot, encara que les formacions herbàcies representin una part important de la vegetació, no sempre es tracta de veritables sabanes, excepte a la regió del Dècan. Al Dècan mateix, per altra banda, la intervenció humana recent ha modificat totalment l’estat, ja prou alterat per mil·lennis de civilització agrícola, de la vegetació. La pressió demogràfica ha portat a construir moltes petites rescloses que fan possible la irrigació i els conreus intensius que exigeix la pressió demogràfica creixent i que han fet desaparèixer les comunitats vegetals precedents i amb elles la fauna que en depenia. Els piocs, les gangues i molts d’altres ocells de sabana han cedit el lloc a espècies limícoles.
Això no obstant, l’avifauna sabanícola índia mereix una menció especial per diverses raons. El paó (Pavo cristatus), present en gran nombre a tota la regió, és l’ocell nacional de l’Índia. Un altre ocell endèmic, el gran pioc indi (Ardeotis nigriceps), que ha esdevingut espècie amenaçada a causa d’una caça excessiva durant dècades, actualment està protegit. A l’àrea dels boscos sabanoides espinosos hi ha molts aiguamolls estacionals, els quals allotgen grans poblacions de nombroses espècies d’ocells aquàtics, tant sedentaris com migradors, a més de molts ocells terrestres. Un d’aquests aiguamolls és el parc nacional de Keoladeo, a Bharatpur (prop d’Agra), famós per les seves grues siberianes (Grus leucogeranus), que ha estat designat com a zona d’importància internacional per la convenció de Ramsar. Només en aquest parc conviuen més de 350 espècies d’ocells, de les quals més de 120 són migradores. Un altre indret inscrit a la convenció de Ramsar, el llac Sambhar —el llac salat més gran de l’Índia—, també es troba a la zona de bosc espinós i és visitat anualment per milers de flamencs Phoenicopterus rubera més de molts altres ocells. Una altra reserva important d’ocells, Nal Sarovar (Gujarat), també es troba en aquesta zona.
La fauna de les sabanes del sud-est asiàtic
Encara que les sabanes de la part continental del sud-est asiàtic, càlides i seques, no tenen una fauna específica especialment rica, hi destaquen una colla de grans herbívors. Entre d’altres cal esmentar, a Cambodja, l’espècie més grossa del món de bovins salvatges, el “kouprey” (Bos sauveli), avui dia malauradament a punt d’extingir-se, el banteng (Bos javanicus), el muntjac (Muntiacus muntjak), el sambar (Cervus unicolor) i fins i tot, en alguns llocs, l’elefant asiàtic (Elephas maximus). Durant el dia, si el cel és cobert, també es poden trobar espècies més rares, com el gos salvatge asiàtic (Cuon alpinus). Es tracta d’animals que acostumen a viure als boscos tropicals semiperennifolis més ombrívols i als boscos monsònics mentre dura la calor del dia, però que, quan arriba la fresca del vespre, se’n van als boscos oberts i a les sabanes, on s’alimenten d’herbes i fruites caigudes i on llepen les eflorescències salines que són tan comunes als sòls secs i sovint salins de les sabanes d’aquesta regió.
Una cadena tròfica característica de les sabanes llenyoses dels tròpics asiàtics la conformarien el tigre d’Indo-xina (Panthera tigris corbettii) caçant els sambars, els muntjacs i les llebres siameses (Lepus peguensis siamensis), els quals al seu torn s’alimenten de les plantes de l’estrat herbaci i aprofiten les petites eflorescències salines. Alguns llangardaixos són també especialment característics dels boscos oberts, com ara l’agàmid, sovint bípede en els seus desplaçaments, Leiolepis belliana rubritaeniata, que és una menja molt apreciada per molts habitants de la regió a l’estació seca.
Pel que fa a les sabanes herboses, són pocs els animals que se’n puguin considerar característics. Un cas especial, però molt localitzat, seria el del dragó de Komodo (Varanus komodoensis) al parc nacional de Komodo. Un altre cas ben característic seria el de l’ualabi àgil (Macropus agilis) a les sabanes del SE d’Irian Jaya.
La fauna sabanícola malgaixa
Encara que una part notable de la superfície de Madagascar és ocupada per formacions herbàcies, no totes poden qualificar-se de veritables sabanes. La majoria, en efecte, són situades fora de la zona intertropical i a més es tracta de comunitats secundàries d’origen recent atribuïbles a l’acció humana. Madagascar, altrament, per mor de la seva superfície limitada, comparada amb els grans continents d’Àfrica i d’Amèrica, i aïllada a més a més des de fa molt de temps, acull una fauna que, per més que la proporció de formes endèmiques sigui alta, és en conjunt relativament pobra. Aquesta fauna és principalment forestal i hi dominen els prosimis i els insectívors. L’origen molt recent de la majoria de les formacions herbàcies explica la pobresa particular de la seva població animal, sobretot pel que fa a mamífers, fins al punt que no hi ha a Madagascar cap espècie herbívora autòctona que en depengui, ni que estigui particularment adaptada a aquests medis.
Els amfibis i els rèptils de Madagascar, com tots els altres grups animals, presenten un grau elevat d’endemisme, però la fauna de les sabanes és en conjunt prou pobra. A remarcar l’absència de cap mena de serp verinosa. Els ocells, en canvi, per als quals un braç de mar no representa un gran obstacle, estan representats per nombroses espècies. Tot i així es tracta també majoritàriament d’espècies forestals. Cal assenyalar de tota manera l’existència d’un remarcable ocell corredor de mida enorme, l’ Aepyornis, extingit durant el període històric i versemblantment sota l’acció humana. Se’n troben ous sovint. Un ocell d’aquest tipus sens dubte només podia viure en medis oberts, cosa que planteja de forma diferent el problema de l’origen de les sabanes actuals.