Al ritme de les pluges
Les sabanes estan àmpliament distribuïdes en un context geogràfic molt extens, caracteritzat per variacions climàtiques considerables. Tal com s’ha assenyalat, de tots els factors que dominen l’ecologia de les sabanes, el primordial és probablement la sequera estacional, que limita el creixement de les plantes i no permet el desenvolupament d’un cobricel dens de capçades.
Els models climàtics
Tanmateix, es donen models climàtics associats a aquests ecosistemes, com ara temperatures elevades tot l’any i una precipitació anual concentrada principalment en una estació, coincident amb l’estiu, durant la qual cau entre el 86% i el 98% de la precipitació anual total, i sequeres periòdiques amb una durada mitjana que pot oscil·lar entre quatre mesos a la zona septentrional de l’Amèrica del Sud i vuit mesos al NE d’Austràlia. L’extrem inferior d’aquesta gamma climàtica correspon a les regions àrides i semiàrides del Sahel, amb una precipitació inferior a 500 mm, i l’extrem superior a les sabanes dels “llanos” orientals de Colòmbia, amb una precipitació que supera els 2 000 mm.
Allà on els sòls ho permeten, les sabanes llenyoses ocupen l’extrem més plujós del continuum de la sabana. Si l’estació seca és inferior a tres mesos, es produeix una gradació de les sabanes llenyoses cap als boscos tropicals secs decidus o semidecidus. Si el període eixut supera els sis mesos, la gradació condueix de les sabanes llenyoses cap a les herboses. De vegades, es pot apreciar aquest continuum al si d’un determinat paisatge. Al capdamunt d’un vessant, els sòls tendeixen a ser poc profunds i sorrencs, amb poca capacitat de retenir l’aigua; la sabana té, en aquest cas, una coberta arbòria relativament esclarissada. Més avall del pendent, els sòls són més profunds i reben aigua per drenatge de la part superior del pendent, a més a més de les precipitacions; aquests sòls poden suportar una sabana més tancada. Al peu del pendent hi pot haver una sabana periòdicament inundable o un bosc de ribera o, fins i tot, ambdues formacions. El període eixut coincideix amb la part més fresca de l’any, i dura entre cinc i set mesos. La temperatura de l’aire oscil·la de temperada a alta. Les gelades són poc freqüents, però quan es produeixen (per exemple als límits latitudinals o altitudinals de la distribució de les sabanes llenyoses), poden tenir un efecte permanent en l’estructura i la composició de la formació.
Normalment hi ha una estació plujosa anual, però en algunes àrees es poden donar condicions multimodulars que siguin causa d’un clima amb més d’una temporada de pluja, com passa en algunes regions de clima monsònic de l’Àsia meridional properes a l’equador. Anàlogament a la temporada humida es poden presentar períodes de sequera de duració curta que poden incidir en el funcionament de les sabanes. Aquests “veranitos”, com són coneguts a l’Amèrica Llatina, poden afectar fins al 68% de l’àrea dels “cerrados” del Brasil i representen un risc seriós per a l’ús del sòl, a causa de la baixa capacitat de retenció d’humitat d’aquests.
Cal destacar que en regions extenses de sabanes, la dinàmica de les pluges és condicionada per l’oscil·lació meridional d’El Niño. Aquest fenomen de variació atmosfèrica i oceanogràfica, entre el Pacífic meridional i l’oceà Índic, presenta una quasi-periodicitat de 2 a 10 anys, que influeix en la precipitació a Indonèsia, a diferents regions de l’Índia, a l’E i el S de l’Àfrica, el N de la Xina, el centre i el SE d’Amèrica i l’E d’Austràlia. Així, l’alta variabilitat característica de la precipitació anual a les sabanes d’Austràlia contrasta amb la d’altres regions de sabana, on la importància extrema d’aquest fenomen és palesa en les variacions de la composició florística i en l’optimització d’estratègies per al maneig de sabanes.
En tot cas, la supervivència, ni que sigui només a nivells de subsistència, de les plantes llenyoses i fins i tot de les herbàcies de les sabanes, suscita alguns interrogants atès que han d’enfrontar-se anualment a unes difícils condicions de sequera durant un període més o menys prolongat. L’explicació rau en el balanç hídric anual d’aquests ecosistemes i a l’estructura de la comunitat vegetal, que ha evolucionat en resposta a la successió de les estacions al llarg del cicle anual i és capaç d’equilibrar el balanç entre la reserva hídrica acumulada durant l’estació humida i els limitats requeriments que té durant l’estació seca.
Els factors que intervenen en l’existència i l’estructura de les sabanes
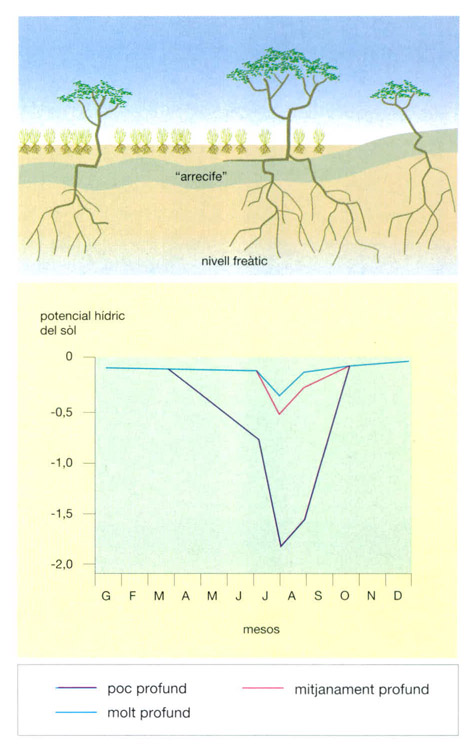
Jordi Corbera, a partir de fonts diverses
La representació esquemàtica d’una secció dels “llanos” veneçolans a prop de Calabozo pot servir per a explicar el fet que l’existència i l’estructura de les sabanes és resultat de la interacció de causes molt diverses. En aquesta zona, sota la superfície del sòl, a poca profunditat, hi ha una capa molt dura, cimentada per hidròxid de ferro, anomenada localment “arrecife”, que en algunes zones emergeix a la superfície. Aquesta capa resulta un obstacle insalvable per a les arrels de les plantes. Com que és impermeable, tota l’aigua de pluja queda emmagatzemada a la capa de sòl superficial, de manera que al començament de l’època seca s’evapora ràpidament. En aquesta situació no podrien existir arbres, ja que no hi ha prou aigua perquè puguin sobreviure, com ho demostra el potencial hídric baix que durant bona part de l’any mantenen les capes més superficials del sòl (a baix). Tanmateix, apareixen aïllades i més o menys regularment espaiades en la matriu de pastures espècies arbòries dels gèneres Byrsonima i Curatella, que, a més a més, són perennifòlies i durant la prolongada sequera no perden la fulla, encara que la sabana s’assequi completament. Unes observacions de camp mostren que un exemplar adult de Curatella transpira, durant la sequera, uns 10 litres d’aigua diaris. La presència d’aquestes espècies s’explica per l’existència d’esquerdes en l’“arrecife”, que permeten a les arrels llargues dels arbres traspassar-lo i tenir accés a la capa freàtica situada a nivells inferiors. D’aquesta manera els arbres disposen d’aigua també a l’època seca, ja que el potencial hídric de les capes més profundes del sòl pràcticament no varia durant tot l’any.
Els índexs d’humitat i l’esclerofil·lisme
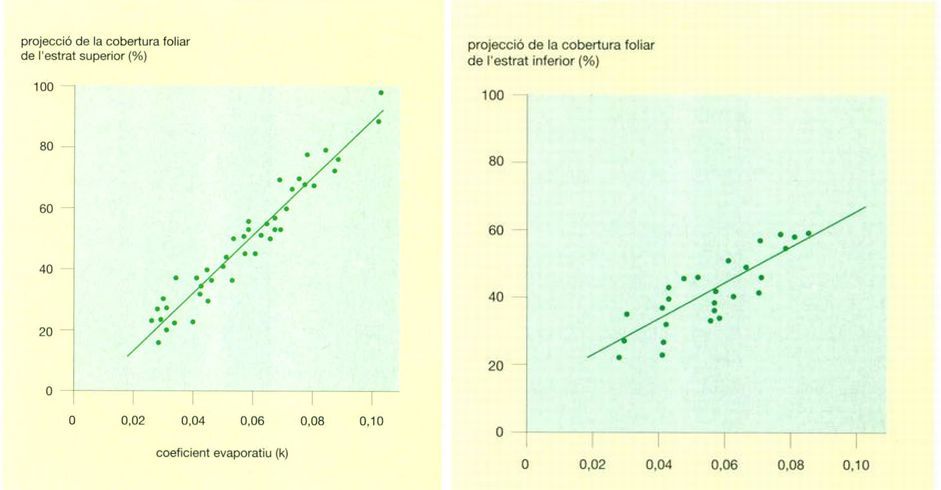
La relació entre la cobertura foliar i la disponibilitat hídrica en les comunitats vegetals perennifòlies de la sabana australiana es fa palesa en aquest gràfic, en el qual es relacionen els percentatges de projecció de la cobertura foliar amb diferents valors del coeficient evaporatiu. Com més elevat és el valor d’aquest coeficient, menys actua l’aigua com a factor limitador del creixement vegetal, i més gran és la projecció de la cobertura foliar. A l’estrat superior (a l’esquerra) la cobertura foliar augmenta molt més ràpidament amb el coeficient evaporatiu que a l’estrat inferior (a la dreta).
Jordi Corbera, a partir de dades proporcionades per l’autor
En efecte, l’índex mensual d’humitat (Ih), que és el quocient entre l’evapo-transpiració real (Ea) i l’evapo-transpiració potencial (Eo) per a un mes donat, és inferior a 1 durant l’estació eixuta (de fet pot acostar-se a un valor nul al pic de la sequera) però aquest quocient no és independent de l’aigua realment disponible al sòl (W) que és la suma de les precipitacions (P) més la reserva extraïble del perfil del sòl al començament del mes (Rext) menys l’aigua perduda per escorrentia (E) o per drenatge cap a nivells massa profunds del perfil del sòl inaccessibles a les arrels (D); així, per a una regió climàtica determinada, es pot escriure:
Ih = Ea / Eo = k · W = k (P + Rext - E - D)
El coeficient k, que expressa la relació entre l’evaporació relativa i l’aigua disponible, s’anomena coeficient evaporatiu; per al cicle anual d’un macroclima donat aquest coeficient es manté més o menys constant, però cada mes el valor de l’índex d’humitat varia en funció del balanç hídric de cada microhàbitat. El seu valor s’acosta a 1 durant els mesos més humits de l’any i s’acosta a 0 en els mesos més eixuts.
A les sabanes llenyoses, a mesura que el clima esdevé més àrid, els arbres i els arbustos es fan més dispersos. L’índex foliar de l’estrat arbori decreix també quan el poder evaporatiu de l’atmosfera creix, és a dir, quan hom es desplaça d’una zona climàtica més humida cap a una de més àrida. Al llarg del continuum sòl-planta-atmosfera, és precisament en la frontera entre la capçada de fullam i l’atmosfera on el potencial hídric pot experimentar la màxima reducció. La conductància més elevada (o el que és el mateix, la mínima resistència) per al flux d’aigua des del sòl cap a l’atmosfera, que és tant com dir la màxima evapo-transpiració, la proporciona un cobricel ben dens (un índex foliar elevat) mentre que unes capçades esclarissades i disperses disminueixen la superfície evaporadora en contacte amb l’atmosfera i l’evapo-transpiració es redueix (el coeficient evaporatiu, k, és menor). Un segon factor que redueix la conductància de l’aigua és el gradual increment de l’esclerofíl·lia (presència de fulles gruixudes i dures, no carnoses, amb estomes escassos, petits i enfonsats) al llarg de transsectes segons un gradient de zones climàtiques (de més humida a més àrida). L’esclerofil·lisme implica una disminució de l’àrea de fulla per unitat de massa foliar, oscil·lant de 6 a 14 m2/kg de massa seca ja que la fulla té un contingut mineral extraordinàriament alt. Unida a una llarga vida de les fulles l’esclerofíl·lia s’interpreta, normalment, com un mecanisme conservador dels nutrients, comú en ambients poc fèrtils; però molts arbres de sabanes llenyoses són decidus i, per tant, les seves fulles tenen una vida curta; per això, sembla més probable que, en aquestes formacions, l’esclerofil-lisme estigui relacionat més aviat amb la conservació d’aigua.
El ritme de creixement i mort de les fulles a l’estrat herbaci de les sabanes segueix de manera palesa un ritme estacional en consonància amb l’alternança d’estacions plujoses i seques. Aquest model estacional és el resultat d’un aprofitament oportunista de l’aigua disponible: el ràpid desenvolupament de les fulles de l’estrat herbaci explota al màxim l’aigua de l’horitzó superficial del sòl, fruit d’un estiu plujós com per exemple el que es dóna sota el clima monsònic de l’Austràlia septentrional; això deixa en aquesta capa superficial tot just l’aigua indispensable per permetre als rizomes i els òrgans subterranis (que també poden extreure una certa quantitat d’aigua de capes una mica més profundes) sobreviure a la llarga sequera hivernal. En el clima subhumit de Darwin, on les pluges són molt abundants (1 491 mm/any) hi ha només petites diferències entre l’evapo-transpiració de l’aigua de les comunitats perennifòlies i estacionals; el nivell d’aigua del sòl és també similar. En canvi, a les zones semiàrides (com Daly Waters) o decididament àrides (com Tennant Creek), la utilització de l’aigua durant l’estació humida per a un creixement vigorós de les fulles és molt més alt als estrats inferiors de la sabana (índexs foliars del 40 al 56%) que als de les comunitats perennifòlies (índexs foliars del 27 al 29%). El dipòsit d’aigua del sòl s’esgota molt més ràpidament en el cas dels primers però les fulles entren en senescència i moren al final de l’estació humida, cosa que permet conservar prou aigua al perfil del sòl per a permetre sobreviure els rizomes i altres òrgans subterranis.
La producció i els fluxos al llarg de l’any
Com en qualsevol ecosistema, la transferència i l’intercanvi de massa i d’energia en el continuum sòl-vegetació-atmosfera, modulades per les variacions en el temps i en l’espai de les interaccions ecològiques, defineixen les característiques bioenergètiques i biogeoquímiques de la sabana. El funcionament bioenergètic es pot mesurar en termes de producció primària total (PT), és a dir, de fixació d’energia radiant per unitat de superfície vegetal i per unitat de temps. En concret, la producció primària total es pot expressar com:
PT = ∆B + M + D + R
on ∆B és la variació de la biomassa en un temps donat, que s’obté per diferència entre la massa se- ca recollida per unitat de superfície en diferents temps; M és l’índex de mortalitat del teixit funcional; D és l’índex de pèrdua de massa vegetal com a conseqüència de processos de descomposició i consum i R és la taxa de respiració de la comunitat vegetal. Altrament, la quantitat de massa tangible, fixada com a producció primària total, correspon a la producció primària neta (PN) que és la diferència entre la producció primària total i la respiració:
PN = PT - R
Tant PN com PT es mesuren en unitats de massa o d’energia per unitat de superfície i de temps (per exemple, kg/m2 de biomassa seca per dia o MJ/m2 per dia).
La producció biològica
La mitjana anual de producció primària neta i de biomassa en les sabanes està positivament correlacionada amb la mitjana anual de precipitacions. El límit inferior de l’eficàcia fotosintètica correspon a les sabanes, condicionades climàticament, de les regions àrides i semiàrides; així, a les sabanes d’Àfrica sud-oriental hi ha una producció de 100 g de matèria seca/m2. A l’extrem superior de l’eficàcia es troben els valors de producció primària neta estimats en sabanes de Loudetia a Lamto (Costa d’Ivori) i a Nigèria (3 000 g/m2). Tanmateix, la producció primària no és controlada per les precipitacions, sinó per la durada del període en el qual és possible el creixement, que està en funció de la pluja, de les exigències d’evaporació i de les reserves d’aigua del sòl. Això explica la paradoxa aparent de les sabanes llenyoses, que és que tenen tant una limitació d’aigua com de nutrients: l’aigua limita la durada de creixement, i els nutrients en limiten la taxa.
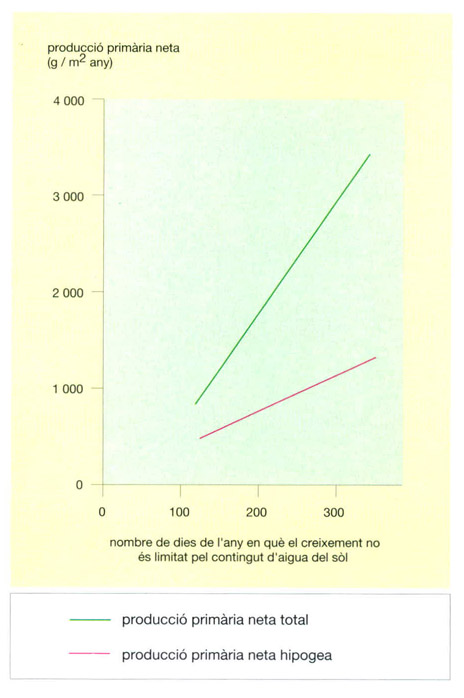
Relació entre la disponibilitat d’aigua del sòl i la producció primària neta de les sabanes llenyoses tropicals. La disponibilitat d’aigua s’ha expressat en funció del nombre de dies de l’any en què el contingut d’aigua del sòl és suficient per a compensar l’evapo-transpiració potencial, i, per tant, no limita el creixement vegetal. La producció de les sabanes llenyoses amb prou feines arriba a la meitat de la dels boscos tropicals humits, però s’assoleix amb una desena part de la biomassa, de manera que les sabanes es poden considerar ecosistemes més eficients que les selves. A les sabanes, fins al 50% de la producció primària té lloc a sota terra, un percentatge elevat comparat amb el d’altres biomes. La producció hipogea, però, es va fent menys important com més humides són les sabanes, tal com mostren les rectes divergents del gràfic. Quan el contingut d’aigua del sòl es manté elevat durant la major part de l’any, el percentatge de producció hipogea disminueix fins al 30% o menys.
Jordi Corbera, a partir de dades proporcionades per l’autor
La majoria de les estimacions de producció de les sabanes es basen en els valors màxims de fitomassa acumulada per la vegetació, que podrien situar-se entre un 30% i un 70% per sota dels valors reals. Així i tot, bo i tenint presents aquestes consideracions, les dades disponibles fins ara indicades posen de manifest que la capacitat bioproductiva total de les sabanes pot oscil·lar entre uns límits molt amplis. El límit inferior (100 g/m2 anuals) equivaldria als valors màxims de producció neta coneguts de les comunitats arbustives de climes àrids i semiàrids; el límit superior (3 000 g/m2 anuals) és similar als valors màxims enregistrats als boscos tropicals.
Les sabanes llenyoses fan la impressió de ser ecosistemes amb una productivitat primària baixa. El baix índex foliar, la durada restringida de les fulles, la baixa conductància als estomes i el baix contingut de nutrients de la fulla fan difícil assolir taxes gaire elevades de producció primària, però això queda compensat, en part, per un nivell d’ener- gia solar sempre alt i per l’elevada temperatura de l’aire. L’índex de creixement anual de les plantes perennes és inferior a l’assolit pels ecosistemes contínuament humits, com seria el cas dels boscos tropicals, però les taxes de producció assolides a les sabanes llenyoses durant l’estació humida són força similars a les enregistrades en boscos tropicals pobres en nutrients. L’índex del total de la producció primària neta anual de les formacions llenyoses tropicals és d’entre 950 i 2 750 g/m2, i se- gurament és d’una mitjana de prop de 1 300 g/m2. A l’extrem inferior de la taxa de producció, el 50% o més d’aquesta producció primària té lloc a sota terra. En formacions llenyoses més humides, la frac- ció de producció subterrània baixa fins a menys d’un 40%, que malgrat tot és alta amb relació a d’altres biomes. Aquesta alta fracció de producció subterrània és probablement la raó principal que els índexs enregistrats pel que fa a l’increment de diàmetre en les formacions llenyoses tropicals siguin més baixos que els dels boscos de plantació cultivats en els mateixos emplaçaments. La taxa anual d’increment de diàmetre en les formacions llenyoses tropicals és aproximadament d’un 3-4% de l’àrea basal de l’arbre.
La fracció de la producció primària de l’ecosistema atribuïble a l’estrat herbaci disminueix de manera no lineal amb l’augment del recobriment de les capçades dels arbres. Al límit inferior del que es consideren sabanes llenyoses, la producció herbàcia pot ser aproximadament igual que la producció de les plantes llenyoses. A mesura que el cobricel de les capçades dels arbres es va tancant, la producció de l’estrat herbaci disminueix, primer ràpidament, per quedar després estabilitzada a un nivell baix.
Les tècniques de mesura
La producció primària es pot mesurar de manera directa o indirecta. En el cas de les sabanes, les mesures directes de producció primària es basen en balanços de massa que consisteixen a recollir el material vegetal a l’inici i a la fi d’un espai de temps. La massa epigea (és a dir, la que sobresurt de la superfície del sòl) d’una superfície representativa de la comunitat se sega i se’n separa la massa funcional (verda) de la no funcional (no verda). L’índex de mortalitat es mesura pels canvis de la massa morta en un espai de temps definit i les pèrdues per descomposició, mitjançant tècniques diverses que poden consistir a marcar el material mort, en l’ús de ‘bossetes de fullaraca’ o en observacions directes dels òrgans morts. Les determinacions de la massa hipogea, la que és subterrània, es fan mitjançant excavacions de blocs de sòl, a l’inici i a la fi de l’interval de temps, i després se separen les arrels del bloc mitjançant procediments de flotació i rentat. Els índexs de descomposició i de mortalitat de les arrels es determinen mitjançant càlculs similars als emprats per a la massa epigea, però la dificultat rau a separar el material funcional del no funcional. El recurs d’utilitzar sals reductores de tetrazole dóna bons resultats en aquesta separació.
Pel que fa a les mesures indirectes, es pot acudir a la mesura de l’intercanvi de gasos, emprant cambres d’assimilació o tècniques microclimàtiques. En el cas de les cambres d’assimilació, una part de la comunitat es tanca en una cambra climatitzada i es mesuren la taxa d’intercanvi de diòxid de carboni i de vapor d’aigua, amb analitzadors infrarojos de gasos disposats en un sistema controlat. Els mètodes microclimàtics consisteixen a mesurar les transferències de massa i d’energia entre el sòl de les sabanes i l’atmosfera, emprant tècniques de balanç d’energia i intercanvi de moment. Generalment es mesuren, sobre la vegetació herbàcia i sobre el cobricel, la radiació neta, el flux de calor al sòl, la temperatura de l’aire, la velocitat de l’aire, les concentracions de diòxid de carboni i de vapor d’aigua a l’aire i l’índex foliar de la comunitat.
Els resultats obtinguts pel mètode directe de recollida resulten similars als que s’obtenen pels mètodes no destructius. Així, els valors de les taxes netes de creixement a les sabanes de l’Orinoco, mesurats mitjançant la tècnica de balanç de massa (entre 2,8 i 6,9 g de matèria seca/m2 dia), resulten similars als mesurats mitjançant tècniques de balanç energètic (entre 0,102 i 0,127 MJ/m2 dia o de 6,6 a 7,9 g de matèria seca/m2 dia).
Els marges de variació
Aquests mètodes han estat emprats per a caracteritzar la producció primària de les sabanes arreu del món. Uns estudis a llarg termini fets a les sabanes de l’Orinoco, emprant mètodes de balanç de massa, indiquen que la fitomassa epigea varia, segons els punts, entre 122 i 688 g/m2 i la hipogea mostra una acumulació que pot variar entre 142 i 427 g/m2. La producció primària neta de la regió mostra variacions d’un punt a un altre de més del doble, amb valors extrems de 388 g/m2 any a les sabanes estudiades de San Diego de la Cabrutica (Veneçuela), i 968 g/m2 any a les sabanes protegides del foc i del pasturatge. A l’estat d’Amazones, a Veneçuela, on s’enregistren precipitacions anuals superiors als 2 400 mm, la producció epigea neta en sabanes cremades és de 276 g/m2 any i la producció hipogea és de prop de 200 g/m2 any. Als altiplans dels “llanos” orientals de Colòmbia, els valors de producció neta varien entre 250 i 560 g/m2 any.
A les sabanes de l’Orinoco, un nombre limitat d’espècies aporta una elevada proporció de la biomassa. Així, per exemple, resulta comú que tres espècies constitueixin del 60% al 87% de la fitomassa epigea total. La gramínia Trachypogon vestitus, per exemple, produeix el 79% de la fitomassa epigea en sabanes cremades. Per altra banda, en sabanes protegides del foc i la pastura, hi ha un model més homogeni de distribució de fitomassa entre espècies, en el qual tres espècies de poàcies, la ja esmentada T. vestitus, Axonopus canescens i A. purpusii, es reparteixen el 84% del total. Aquests resultats indiquen que els factors determinants de les variacions espacials de la producció de les sabanes de l’Orinoco s’associen a canvis ambientals.
Als “cerrados” brasilers, la producció ha estat avaluada en poques localitats, si es compara amb la seva extensió. Les dades de producció epigea reportades per als “campos cerrados” de Pirassununga, a l’estat de São Paulo, varien entre 550 i 780 g/m2 any per als “campos sujos” del districte federal de Brasília entre 297 i 550 g/m2 any, i per als “cerrados” pròpiament dits entre 90 i 327 g /m 2 any.
A les sabanes australianes, la producció epigea varia de 70 a 420 g/m2 any, amb els valors més baixos mesurats a les sabanes arbustives d’Acacia dominades per gramínies dels gèneres Enneapogon i Thyridolepis i els més alts en sabanes en què les poàcies dominants eren Heteropogon contortus, Themeda australis i diferents espècies del gènere Bothriochloa. La producció anual de la fitomassa epigea varia entre 181 i 570 g/m2 anuals, amb els valors més baixos mesurats a les sabanes arbustives d’Acacia i els més alts a les sabanes monsòniques de T. australis, Sehima nervosum i Chrysopogon fallax.
A les sabanes africanes, la capacitat de producció de les pastures és condicionada per la variació espacial de la precipitació. Sota règims de pluviositats compresos entre 500 i 1 200 mm, la producció neta epigea en les sabanes de la regió sudano-zambeziana varia entre 100 i 800 g/m2 any. A les sabanes de clima similar a les de l’Orinoco, els “cerrados” o Austràlia, es produeix una quantitat més gran de matèria orgànica. Així, les sabanes de Loudetia i Andropogon de Lamto, a la Costa d’Ivo- ri, en produeixen de 810 a 2 150 g/m2 any i algu- nes sabanes septentrionals de Nigèria n’acumulen 1 734 g/m2 any.
Al N de l’Índia, amb una precipitació anual inferior a 400 mm, la producció epigea en sabanes on les gramínies dominants són l’“anjan” (Cenchrus ciliaris) i Aristida depressa és de 1 600 g/m2 any i la hipogea de 600 g/m2 any. En les sabanes més humides (900 mm anuals), amb “zarga” (Dichanthium annulatum) i “gandhejghas” (Cymbopogon martinii) com a gramínies dominants, la producció epigea és de 460 g/m2 any. A la regió de Varanasi, amb una pluviositat de 1 200 mm, la producció epigea de sabanes amb gram (Cynodon dactylon), anomenat en hindi “doob”, és de 3 400 g/m2 any i la hipogea de 1 200 g/m2 any.
A les regions de sabana inundables com, per exemple, als “llanos” de l’Orinoco, hi ha sabanes de marge, d’aiguamoll i d’“estero”, amb una formació paisatgística que comprèn des de bancals o cavallons no inundables (marges) fins a depressions (aiguamolls i “esteros”) sotmeses a inundacions estacionals. Als marges permanentment emergits, les produccions epigees netes de la vegetació herbàcia dominada per Paspalum chaffanjohnii, Sporobolus indicus i Axonopus purpusii s’han mesurat entre 425 i 890 g/m2 any i de les hipogees en 460 g/m2 any; als aiguamolls, on les plantes dominants són Leersia hexandra, Panicum laxum i P. versicolor, s’assoleixen valors de 550 g/m2 any de matèria epigea, i als “esteros”, on l’espècie dominant és Hymenachne amplexicaulis, la producció epigea pot variar entre 500 i 2 207 g/m2 any i s’hi han mesurat 1 195 g/m2 any de producció hipogea. En sòls més fèrtils, amb una làmina d’aigua que arriba fins a 3 m de profunditat entre juny i novembre, en sabanes de Paspalum fasciculatum s’han mesurat produccions de matèria epigea d’entre 1 400 i 2 540 g/m2 any.
Els factors modificadors
Tot i que les sabanes neotropicals són a les zones de precipitació més alta, el seu comportament productiu contrasta amb la capacitat bioproductiva de sabanes de condicions climàtiques similars. Així, la precipitació sembla tenir una incidència menor en els seus processos productius. En condicions d’alta humitat relativa com les que hi ha a Puerto Ayacucho (Veneçuela), on es dóna una precipitació superior als 2 400 mm, la producció total és de 579 g/m2 any. Aquest valor és similar als enregistrats en altres sabanes neotropicals amb precipitacions de 1 300 mm. A més a més, els valors de producció estimats en aquestes sabanes neotropicals a partir dels models usuals basats en les dades de precipitacions i temperatures, foren entre 1,7 i 2,1 vegades superiors als valors màxims mesurats.

La comparació entre les dades climàtiques anuals i les estratègies d’utilització de l’aigua de les comunitats perennifòlies i de sabana estacional de tres localitats del Territori Septentrional d’Austràlia mostren una relació clara entre el recobriment foliar i la pluviositat. La localitat de Darwin, amb un clima subhumit, presenta una cobertura foliar, tant de l’estrat superior com de l’estrat inferior, molt més gran que no pas el de Daly Waters, de condicions semiàrides, i encara més que el de Tennant Creek, més àrid. Cal destacar que les diferències entre el recobriment de la formació perennifòlia i el de la sabana són només a nivell de l’estrat inferior.
Dades proporcionades per l’autor
Els resultats obtinguts indiquen, doncs, que a les sabanes neotropicals operen altres limitacions diferents de les climàtiques; aquesta capacitat bioproductiva menor que la determinada per les condicions climàtiques indica que els factors edàfics poden actuar directament sobre la vegetació o modular l’acció del clima sobre els processos productius. Així, certament, a les sabanes són comunes l’estacionalitat i la variació anual de les precipitacions, però durant l’estació de pluges poden haver-hi períodes d’estrès hídric condicionats per una baixa capacitat d’emmagatzemament i retenció d’aigua dels sòls. A més, la baixa disponibilitat d’humitat edàfica determina que al començament de l’estació eixuta, la quantitat d’aigua emmagatzemada al sòl minva ràpidament i, així, es redueix l’índex de creixement de la vegetació. L’eliminació de l’estrès hídric a l’estació seca gràcies a la irrigació augmenta la producció de matèria seca a les sabanes de l’Orinoco i als “cerrados” brasilers. S’han evidenciat altres restriccions edàfiques a les sabanes de l’Orinoco, on la productivitat neta estacional depèn de la massa efectiva del sòl, expressada per la fracció textural inferior a 2 mm, de tal manera que la matèria seca acumulada a les sabanes de sòls profunds és dos cops més gran que la mesurada en sabanes sobre sòls superficials. La disponibilitat d’aigua es redueix, per tant, considerablement a les sabanes en què la massa efectiva del sòl varia entre el 6% i el 14% de la massa total del sòl. Encara més, en sòls superficials, el creixement de les espècies també es veu afectat per la resistència mecànica a la penetració de les arrels. Els resultats exposats sobre les limitacions edàfiques en el funcionament de les sabanes de l’Orinoco foren corroborats mitjançant l’anàlisi multivariable de l’expressió regional de les sabanes de l’Orinoco.
La producció de matèria seca en sabanes neotropicals també és condicionada per la baixa disponibilitat de nutrients als sòls. La fertilització experimental dels sòls dels “llanos” de l’Orinoco amb nitrogen, fòsfor i potassi determina un augment significatiu de la biomassa vegetal i de la concentració de nutrients a la massa herbàcia. En el cas de les sabanes de l’Orinoco, s’establí experimentalment que l’augment de l’índex de producció el provoquen simultàniament un desenvolupament més gran de l’àrea foliar i un augment de l’eficàcia fotosintetitzadora de les gramínies dominants. L’estrès nutritiu també és comú a les sabanes australianes, on el dèficit de nutrients és el factor principal que limita la productivitat de la coberta herbàcia en sòls amb sesquiòxids. Les sabanes monsòniques i les semiàrides d’Austràlia es caracteritzen per limitacions de nutrients que interactuen amb l’estrès hídric que hi ha a l’hivern. Quan es corregeixen les deficiències de nitrogen i fòsfor a les sabanes de Katherine, la producció d’herba augmenta fins a arribar al valor potencial típic de l’estació.
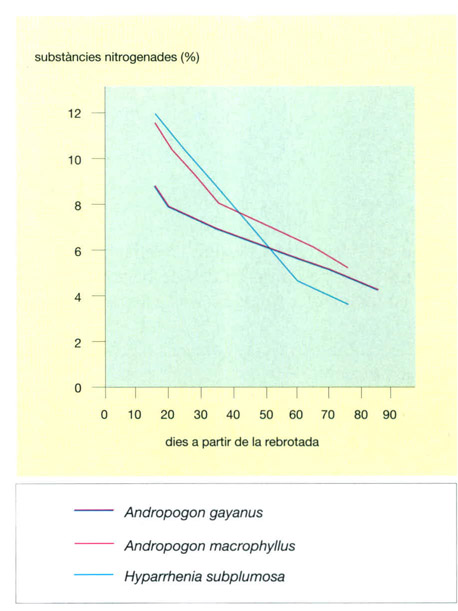
Variacions en el contingut en substàncies nitrogenades durant el creixement de tres espècies de gramínies de les sabanes africanes. A mesura que les plantes creixen, els teixits es carreguen de cel·lulosa, i el seu contingut en proteïnes disminuex. En totes tres espècies, apareix un punt d’inflexió, que correspon a un canvi en la velocitat de creixement. Les espècies del gènere Andropogon el presenten pocs dies després d’haver rebrotat, mentre que Hyparrhenia subplumosa continua creixent a gran velocitat durant un període més llarg.
Jordi Corbera, a partir de dades proporcionades per l’autor
En el cas de les sabanes inundables, la productivitat neta és més gran que a les sabanes ben drenades; tot i això, la presència d’una làmina d’aigua sobre la coberta vegetal determina que la durada total del període de creixement sigui menor, de manera que la productivitat es redueix en funció de la permanència de la inundació. Així, l’alt potencial de producció total, mesurat en sabanes inundables (2 300-5 200 g /m2 any), s’expressa només parcialment quan la làmina d’inundació persisteix durant un temps considerable. En casos extrems, la producció anual epigea de les sabanes inundades (860-912 g/m2 any) és similar a la mesurada en les sabanes ben drenades.
El flux d’aigua
El gradient horitzontal d’humitat a través del paisatge de les sabanes és un dels factors operatius que determinen les variacions en la vegetació. El gradient fisionòmic de les sabanes, que comprèn des de vegetació herbàcia fins a vegetació arbòria, s’ha explicat basant-se en la interacció entre les precipitacions, les formes vitals que es desenvolupen a les sabanes i les característiques físiques del perfil del sòl.
En una revisió sobre el flux hídric de les sabanes d’arreu del món s’ha trobat que el valor mitjà de precipitació arriba al 3% de la precipitació global i l’evaporació a l’atmosfera és aproximadament del 2%; l’equilibri es manté pel drenatge superficial i intern del sistema encara que al bioma es donen situacions amb índex d’evaporació potencial que poden duplicar les precipitacions totals.
Estimacions del flux hídric a les “matas”, els bosquetons semicaducifolis que es troben escampats al si de la matriu herbàcia de les sabanes de l’Orinoco, indiquen que el component principal d’equilibri hídric és l’evapo-transpiració, amb valors que varien entre el 63% i el 72% de la precipitació anual. Al gramenet, l’evapo-transpiració és similar a la dels bosquetons (72% de la transpiració total). S’han assenyalat valors de 900 a 1 200 mm anuals al “miombo” del Zaire i als boscos semicaducifolis de Yapo, a la Costa d’Ivori. Pel que fa a les sabanes australianes, varia des dels 233 mm observats en gramenets amb Acacia fins als 573 mm indicats per a “tussocks” de “Mitchell grass” (Astrebla) sobre sòls argilosos.
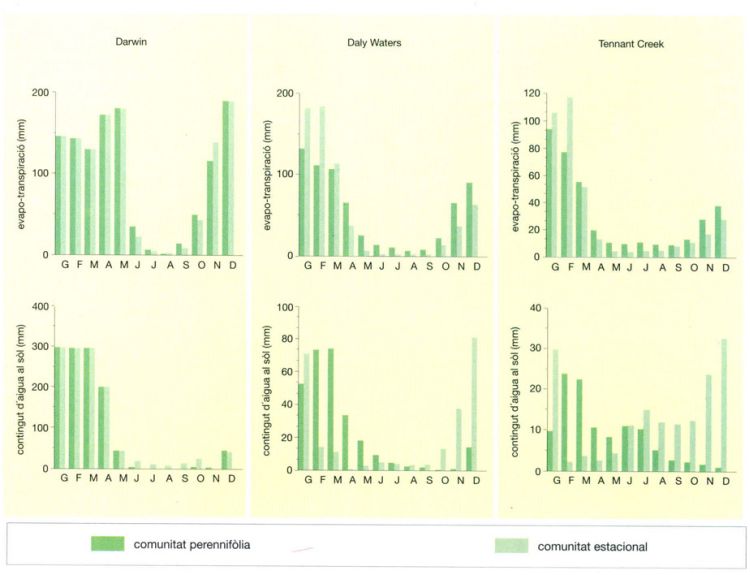
Els valors d’evapo-transpiració i contingut d’aigua al sòl en tres localitats del Territori Septentrional d’Austràlia il·lustren diferents patrons d’explotació de l’aigua per a dues comunitats diferents. Al clima subhumit de Darwin hi ha poca diferència entre l’evapo-transpiració de la comunitat perennifòlia i la de la sabana estacional. La quantitat d’aigua del sòl és també molt semblant i elevada en ambdós ecosistemes. En canvi, a la localitat semiàrida de Daly Waters, durant l’estació humida, les plantes de la sabana utilitzen més aigua que les de la comunitat perennifòlia. Tanmateix, al final de l’estació humida, el fullatge de la comunitat estacional es marceix, i durant tota l’estació seca són les plantes perennifòlies les que presenten uns valors d’evapo-transpiració més elevats. Aquest fenomen s’accentua a Tennant Creek, on el clima és encara més sec.
IDEM, a partir de dades proporcionades per l’autor
Els càlculs d’evapo-transpiració potencial anual (ETP) de les regions cobertes de sabanes indiquen que a les sabanes neotropicals són típics els valors propers als 1 300 mm. A les sabanes africanes, els valors reportats varien entre 1 600 i 1 700 mm mentre l’evapo-transpiració actual (ETA) ho fa entre 490 i 1 050 mm. Així, el quocient ETA/ETP presenta una gamma de valors que va de 0,28 a 0,65. Aquests valors contrasten amb els quocients propers a la unitat típics de cultius ben irrigats.
La relació entre l’increment de matèria seca per unitat de superfície i la pluviositat s’anomena eficiència en l’ús de l’aigua. El seu valor permet relacionar les pèrdues d’aigua de la vegetació a través del procés de transpiració amb la quantitat de matèria orgànica produïda per les sabanes. L’eficiència en l’ús de l’aigua mitjana de les sabanes d’arreu del món ha estat estimada en 1,2 g de matèria seca/kg de precipitació. En el cas de les sabanes del SW d’Àfrica, amb pluviositats inferiors als 700 mm, aquesta eficiència és d’1,0 g/kg, mentre que a les sabanes australianes, amb pluviositats d’entre 500 i 1 800 mm, varia de 2,6 a 3,7 g/kg. S’han calculat valors menors a les sabanes de l’Orinoco, on hi ha les precipitacions més elevades enregistrades en àrees cobertes de sabana (1 000-2 800 mm).
La vegetació afecta el flux d’aigua, no sols per l’efecte de la transpiració, sinó també pel de la intercepció. En el cas dels bosquetons de les sabanes dels “llanos” de l’Orinoco, s’ha mesurat una intercepció que supera el 20% de la precipitació anual (1 100-1 300 mm). Així, la vegetació llenyosa afecta l’energia cinètica de les pluges i, per tant, la quantitat d’aigua infiltrada i l’erosió del sistema. A més, la vegetació redistribueix la precipitació a través del fullam i dels troncs dels arbres, amb l’arrossegament de partícules i de materials solubles, que modifica la quantitat de nutrients a la superfície del sòl.
El flux de nutrients
Les anàlisis de les relacions tròfiques a les sabanes indiquen que la disponibilitat de nutrients al sòl és baixa. En casos extrems, encara que el sòl tingui un grau de fertilitat més gran, les condicions climàtiques, en particular les precipitacions, poden limitar el desenvolupament de la vegetació, com passa amb els sòls amb un horitzó d’acumulació d’argila i una alta saturació de bases (alfisòls) d’algunes sabanes australianes.
Tal com s’ha posat de relleu al capítol dedicat als sòls de les sabanes, molts d’aquests (oxisòls, entisòls i ultisòls) es caracteritzen per un baix contingut de nutrients, que es reflecteix en les característiques xeromòrfiques de la vegetació. La hipòtesi d’un escleromorfisme lligat a l’oligotròfia del sòl fou formulada per explicar l’aparença coriàcia de la vegetació dels “cerrados” al Brasil, basant-se en el baix contingut de nutrients al sòl. El gradient oligotròfic dels sòls dels “cerrados” es considera com a determinant del con- tinuum de variants fisiognòmiques de la vegetació. Aquesta hipòtesi de l’escleromorfisme lligat a l’oli- gotròfia es complementà amb la inclusió de l’efecte de l’acidesa del sòl com a conseqüència de l’alta concentració de l’alumini al complex de saturació.
L’efecte de l’estrès per mancances tròfiques sobre el creixement de la vegetació de les sabanes ha estat analitzat experimentalment. Així, l’acumulació de matèria seca i el contingut nutritiu de la capa herbàcia de les sabanes de l’Orinoco i dels “cerrados” brasilers es poden incrementar com a resultat de l’addició de nutrients com ara nitrogen, fòsfor i potassi. La dinàmica dels nutrients en la sabana s’analitza mitjançant l’estudi de les entrades i sortides del sistema. Així, per exemple, han estat avaluades les pèrdues de nutrients de l’estrat herbaci de les sabanes resultat d’incendi, emissió, lixiviació i volatilització. S’han fet mesures de volatilització d’amoníac en sabanes americanes protegides que posen de manifest que les pèrdues de nitrogen durant l’estació de pluges van de 0 a 0,4 mg/m2 hora. La font d’amoníac es localitza en el material vegetal que es descompon a la part inferior del cobricel i a les espigues de les herbes graminoides. També es donen pèrdues de nitrogen en forma d’òxids de nitrogen quan el sòl s’humiteja al començament de l’època de pluges; s’han estimat valors de 2,6 kg/ha per al monòxid de nitrogen i de 0,65 kg/ha per al diòxid de nitrogen. Els incendis freqüents comporten una pèrdua petita però repetida de nitrogen i òxids de nitrogen en el fum. Una part d’aquest nitrogen torna en forma de pols o naturalment de pluja àcida, però una altra part es perd damunt d’altres tipus de vegetació o dels oceans.
Les pèrdues de nutrients del sistema poden recuperar-se per la deposició atmosfèrica, fins i tot amb escreix, com en el cas del magnesi que enfront d’una pèrdua d’1,72 kg/ha any, pot incorporar, a partir de la deposició atmosfèrica 11,30 kg/ha any, amb un superàvit de 9,58 kg/ha any. En canvi, el balanç és quasi nul per al calci (3,50 kg/ha any d’entrades enfront de 2,55 de pèrdues); el potassi (3,61 i 2,34 respectivament) i el fòsfor (0,43 i 0,32). En el cas del nitrogen, les pèrdues (12,5 kg/ha any) es compensen mitjançant la incorporació anual per precipitació (2,2 kg/ha any) i la fixació biològica (entre 0,2 i 222 kg/ha any). La fixació de nitrogen per les lleguminoses també contribueix a la dota- ció de nitrogen al sistema. Tanmateix, l’aportació de les lleguminoses a la fitomassa total de les sabanes és relativament baixa (exceptuant les sabanes que es cremen sovint). Ben paradoxalment, tot i que moltes sabanes, sobretot llenyoses, de sòls infèrtils són dominades per espècies arbòries lleguminoses, molt poques d’aquestes espècies tenen la capacitat de formar nòduls d’arrels que continguin bactèries fixadores de nitrogen simbiòtiques. La major part de la fixació de nitrogen en les formacions llenyoses es porta a terme per bacteris independents, per algues verdes que creixen en la superfície del sòl o bé per simbionts de l’arrel de petites lleguminoses que creixen en l’estrat herbaci. En canvi, en una sabana herbosa com els “llanos” de l’Orinoco, s’ha observat nodulació en el 86% de les 127 espècies de lleguminoses recol·lectades.
La baixa disponibilitat de fòsfor pot ser l’origen del baix índex de fixació biològica del nitrogen. L’escassesa de fòsfor és conseqüència del baix contingut inherent a la geologia d’aquestes zones, a la intensa erosió, i a la fusió del fòsfor amb els òxids de ferro en el sòl. La major part de les arrels d’arbres i herbes de les sabanes llenyoses tenen associacions fungals ectomicoríziques, que sembla que incrementen la capacitat de les plantes hostes d’obtenir fòsfor del sòl. Una gran part dels nutrients són reciclats dins de les plantes, més que no pas entre la planta i el sòl.
A l’estrat herbaci de les sabanes, hi ha mecanismes de conservació de nutrients que representen adaptacions als ambients oligotròfics. A les sabanes de l’Orinoco, per exemple, el 72% del nitrogen es redistribueix (3,8 g/m2) mentre que la resta, incorporat a la vegetació, prové de la mineralització de matèria orgànica del sòl i de l’atmosfera. A les sabanes australianes d’herbes altes, la retranslocació cap a la fitomassa subterrània comporta del 0,2 al 0,4% de nitrogen romanent a la fitomassa aèria. Aquest mecanisme de recuperació de nutrients determina que les pèrdues de nitrogen per mineralització ràpida en sabanes cremades es redueixin substancialment. En el cas de les sabanes monsòniques i orientals d’Austràlia, les pèrdues són respectivament d’entre 8 i 10 kgN/ha i d’uns 16 kgN/ha. En els components llenyosos, la redistribució és ineficient i el contingut de N a les fulles dels arbres abans dels incendis és significatiu i arriba a valors de 2,9 g/m2 en les sabanes tropicals altes. A més dels processos meteorològics relacionats amb el flux de nutrients, existeixen efectes locals. Així, el rentat foliar i el caulinar constitueixen vies alternatives de redistribució de nutrients. En sabanes subtropicals altes, la quantitat de nutrients mobilitzats en aquests processos de rentat pot ser igual a la que s’afegeix al sistema per la fullaraca.
Les estratègies radiculars i foliars davant de l’economia hídrica
En general, la fisiognomia de les sabanes es caracteritza per dos grups contrastats de formes biològiques: les plantes herbàcies, principalment les graminoides, i les llenyoses. Unes i altres integren des de les sabanes exclusivament herbàcies fins a les sabanes llenyoses que poden presentar expressions fisiognòmiques de densitat arbòria elevada, com és el cas dels “cerradões”. Només les sabanes condicionades per un clima tropical estacional, amb pluges a l’estiu de menys de 600 mm, tal com succeeix al S de l’Àfrica, poden considerar-se pròpiament climàtiques. A Namíbia, on hi ha des de sabanes herbàcies, restringides a regions amb precipitació entre 100 i 250 mm, fins a sabanes humides, en què la precipitació és suficient per al funcionament d’herbes i arbres, ha estat descrit un gradient fisiognòmic lligat a la pluviositat. En climes més humits, hi pot haver sabanes o vegetació arbòria, depenent de la interacció de la precipitació amb altres factors ecològics com el sòl i el foc. En sabanes ben drenades, per exemple, el gradient observat des de sabanes herboses fins a sabanes llenyoses densament boscades ha estat explicat basant-se en la interacció entre la capacitat potencial de les formes de vida per a explorar el perfil del sòl i les característiques físiques i químiques d’aquest perfil.
Estructura i sistema radicular de la vegetació de sabana al llarg d’un gradient de precipitació creixent

IDEM, a partir de Walter, 1986
L’exemple correspon a un conjunt de sabanes molt seques del SW d’Àfrica. El sòl d’aquesta regió és sorrenc i fi, i pot emmagatzemar la major part de l’aigua de pluja. Tal com s’observa a l’esquema, el volum de precipitacions afecta directament la proporció entre gramínies i plantes llenyoses. Quan les precipitacions anuals no superen els 100 mm (a), l’aigua no pot arribar a les capes profundes del sòl, ja que la poca que cau és ràpidament absorbida pel sistema radical de les plantes herbàcies. Durant la llarga estació seca, que es prolonga 8 mesos, no hi ha disponibilitat d’aigua, de manera que les plantes llenyoses no poden subsistir. Quan les precipitacions són una mica més elevades (uns 200 mm anuals) (b), la situació és bàsicament la mateixa, de manera que la coberta vegetal es limita també a gramínies, bé que en aquest cas poden assolir major alçada. Una precipitació de 300 mm anuals (c) és prou abundant per a permetre la subsistència de petits arbustos. Finalment, amb uns 400 mm de precipitació anual (d) ja poden créixer arbres aïllats, si bé les gramínies continuen sent dominants. En una darrera etapa, no representada, el dosser arbori arribaria a fer-se continu i la seva ombra eliminaria les gramínies. Aquest equilibri és extremament delicat, tal com ho demostren els efectes de la ramaderia. El bestiar consumeix la part aèria de les herbes, de manera que la transpiració disminueix i queda un romanent d’aigua al sòl que pot ser utilitzat per les plantes llenyoses. En poc temps, les sabanes de pastures es cobriran d’arbustos, que desplaçaran les gramínies i acabaran cobrint la major part de l’àrea. Aquest fenomen ha afectat extenses zones, no sols a l’Àfrica sinó també a l’Índia, a la part septentrional de Veneçuela i a altres zones seques.
La competència pels recursos hídrics
A les sabanes hiperestacionals o estacionalment inundables que caracteritzen les posicions topogràfiques deprimides a les regions de sabana amb clima humit, el sòl, generalment molt més argilós que en el cas suara considerat i amb un drenatge intern lent o quasi nul, roman durant una part de l’estació de pluges saturat d’aigua o fins i tot inundat amb una làmina d’aigua de 10 a 20 cm. Sota aquestes condicions, en les quals l’aigua desplaça l’aire dels porus del sòl, només poden persistir espècies adaptades a condicions de deficiència d’oxigen, cosa que moltes espècies herbàcies assoleixen desenvolupant teixits radicals que permeten un flux intern d’aire que s’anomena aerènquima. En contrast amb aquesta situació, durant l’estació seca el sòl s’asseca completament, no sols superfícialment sinó en tot el seu perfil, ja que generalment es tracta de sòls amb horitzons argilosos molt poc permeables a pocs centímetres de profunditat. Així doncs, no queda aigua disponible per a espècies llenyoses que arrelen en profunditat. Fins i tot en els casos que existeixi un nivell freàtic permanent a poca fondària, les arrels, incapaces de travessar l’horitzó argilós, no poden aprofitar aquest recurs hídric. Per aquesta raó les sabanes estacionalment inundades només tenen unes poques espècies llenyoses o manquen completament d’arbres. Més sovint, com a única forma no pròpiament herbàcia, hi apareixen algunes espècies de palmes que, pel que fa a la seva economia hídrica, es comporten igual que les gramínies perennes.
A les sabanes de climes molt més secs, on la precipitació anual és menor de 700 mm, la situació i els factors que regulen la competència entre herbes i plantes llenyoses són totalment diferents. Les pluges que s’infiltren mai no són suficients per a acumular als horitzons inferiors del sòl una reserva d’aigua utilitzable per les espècies d’arrels profundes, de manera que l’únic recurs disponible és el de l’aigua superficial, la qual, per altra banda, es troba en quantitats molt més limitades que a les sabanes humides. Sota aquestes condicions, la com- petència interspecífica s’aguditza i afavoreix les gramínies, que troben avantatges competitius en el seu sistema radical intensiu. Quan les precipitacions són molt escasses, entre 200 i 400 mm anuals, només arriben per mantenir un estrat herbaci. Si les precipitacions són més abundoses, comença a aparèixer un estrat d’arbustos o d’arbres baixos, generalment decidus o perennifolis microfil·les, molt obert. L’estrat llenyós pot augmen- tar d’importància quan les pluges ultrapassen els 600 mm anuals. D’aquesta manera, la quantitat disponible d’aigua en el sòl derivada directament de les precipitacions, regula totalment la relació entre plantes herbàcies i llenyoses. Però si algun factor extern introdueix una pressió selectiva en contra de les gramínies, com podria ser, per exemple, la sobrepastura, aquestes disminuiran el seu recobriment i deixaran aigua disponible per als arbustos. El resultat, si es mantenen aquestes condicions de sobrepastura durant uns quants anys consecutius, serà la contracció gradual de la cobertura herbàcia fins arribar a desaparèixer del tot. Es transforma així l’ecosistema de sabana seca en un matoll espinós tancat i d’escàs valor farratger. En moltes zones tropicals i subtropicals (N de Tanzània, N i E de Kenya, etc.) aquesta arbustificació massiva d’antigues sabanes com a conseqüència directa de la sobrepastura ha tingut conseqüències econòmiques i socials desastroses, i ha obligat a emigrar en recerca de noves pastures considerables contingents de població.
Tot sovint els incendis periòdics tenen també un paper important en l’equilibri entre les plantes herbàcies i les llenyoses en aquestes zones més seques, ja que els arbustos i els arbrissons baixos són més sensibles al foc que no pas les gramínies, de manera que els incendis freqüents tendeixen a mantenir una coberta herbàcia contínua i una baixa densitat d’arbustos. Si la sobrepastura introdueix discontinuïtats a la coberta herbàcia originàriament contínua, el foc es propagarà cada cop amb més dificultat, fins que arribarà un punt en què es farà impossible la seva propagació a través de la vegetació. A partir d’aquell moment els arbustos començaran a resultar afavorits i l’arbustificació es farà pràcticament irreversible.
La captura i la utilització de l’aigua
Les gramínies tenen un sistema radicular molt fi i superficial. Les seves arrels ocupen un volum comparativament petit de sòl, però d’una manera compacta i completa, cosa que possibilita una explotació intensa i una extracció de la major part de l’aigua que conté. En canvi, les plantes llenyoses tenen arrels més gruixudes, que s’estenen sobre un volum molt més gran de sòl però de manera més laxa i l’exploten extensivament. Normalment arriben a capes més profundes i poden explotar aigua d’uns nivells inaccessibles per a les gramínies.
La segona diferència entre les gramínies i les plantes llenyoses és l’economia de l’aigua. Les primeres, quan hi ha aigua suficient, transpiren i fotosintetitzen molt activament; per això la seva producció primària pot ser molt elevada en períodes molt curts de temps. Alhora, però, els seus mecanismes per a estalviar aigua quan aquesta és escassa són molt ineficients. Quan comença la sequera la seva transpiració no disminueix i esgoten ràpidament les reserves d’aigua del sòl. És per això que tota la part aèria de la planta mor i només les arrels i els meristems terminals de la tija persisteixen, protegits de la dessecació per una coberta de fulles seques. És així com les gramínies aconsegueixen sobreviure a l’estació seca. L’estratègia dels arbres i dels arbustos és totalment diferent. Tenen una forta capacitat per a regular la pèrdua d’aigua quan aquesta és escassa, tancant els estomes de les fulles i perdent-les eventualment quan això no és suficient. Llavors només romanen la tija i les branques. Això no obstant, és fàcil demostrar que aquestes parts continuen perdent una petita quantitat d’aigua durant la sequera, de manera que esdevé impossible la subsistència de la planta si no manté, almenys, un petit subministrament d’humitat. Quan el sòl és sec, si no hi ha aigua disponible, la planta no pot sobreviure.
Per això, climàticament parlant, el que determina el triomf de les gramínies per sobre de les plantes llenyoses és l’existència d’un període anual durant el qual no hi ha aigua disponible per a ésser utilitzada pels arbres. Això no és una conseqüència inevitable de l’existència d’una època de sequera. En començar la sequera, l’evaporació asseca ràpidament les capes superficials del sòl, però molt més lentament les capes profundes. Les plantes llenyoses tenen arrels molt més profundes que les gramínies. Per tant, el sòl pot estar completament sec per a les plantes herbàcies però, si hi ha aigua (per exemple per la presència d’una capa freàtica) a un nivell a l’abast de les arrels dels arbres, el bosc substituirà i eliminarà la sabana encara que hi hagi una intensa sequera. Per als arbres, existirà aleshores una sequera climàtica, però no edàfica, i no se’n veuran afectats. L’important per als arbres no és que plogui o no, sinó simplement la disponibilitat d’aigua al sòl a una profunditat assolible.
Es donen múltiples situacions en què pot haver-hi aigua disponible per als arbres durant la sequera. En molts casos permeten explicar perquè en àrees predominantment de sabana poden aparèixer arbres aïllats o clapes de bosc. Però també hi ha altres factors que poden impedir la colonització d’una àrea pels arbres i afavorir el desenvolupament d’una sabana. A més a més del clima, els més importants són les característiques del sòl (nutrients i granulometria), la posició topogràfica, el drenatge, la presència o no de capes impermeables i cuirasses, la localització de la capa freàtica, la freqüència més o menys gran del foc i l’explotació pels herbívors.
Per tant, es pot dir que el desenvolupament i el manteniment d’un ecosistema de sabana és, en la majoria dels casos, un fenomen complex on actuen simultàniament molts factors. La influència d’aquests és diferent en les diverses zones i produeixen, per tant, diferents tipus de sabana. Pràcticament cada àrea de sabana és un cas particular on es combinen diferents variables ambientals (incloent-hi el clima, el sòl, l’acció antròpica, etc.) per a produir un cert tipus de sabana.
Els atots de les plantes herbàcies
Les herbes dominants de les sabanes són majoritàriament espècies perennes i amb unes fulles relativament amples i fines. Algunes poden arribar a ser molt altes (fins a 3 m), tot i que la seva alçada normal és d’un metre. El desavantatge d’aquesta forma de vida és la necessitat de tornar a desenvolupar anualment les parts aèries. Un altre grup de plantes herbàcies és format per aquelles que sobreviuen durant l’estació seca en forma de llavors. Aquest grup inclou algunes herbes graminoides anuals, però principalment consisteix en petites plantes dicotiledònies. Rarament constitueixen una gran fracció de la biomassa, excepte després d’una alteració física del sòl.
El total de biomassa herbàcia en peu arriba al seu màxim aproximadament a la meitat de l’estació plujosa i persisteix fins que comença l’estació seca. Una part significativa de matèria morta roman a lloc i serveix de combustible dels freqüents incendis que es produeixen. En àrees que no han patit incendis durant alguns anys, s’acumula la matèria morta. El foc té un important paper com a accelerador del cicle de nutrients perquè ho transforma tot en cendra. En les àrees cremades la brotada comença pocs dies després del foc, i cap a la meitat de l’estació plujosa següent la biomassa viva en peu assoleix els mateixos nivells que en parcel·les no cremades. La matèria morta, però, constitueix tan sols una part molt petita en comparació amb la matèria viva, a diferència d’àrees que no han estat afectades pel foc durant uns quants anys on la biomassa viva pot ser només una petita part respecte a la biomassa morta. Generalment els ramaders recor-ren als incendis per incrementar la quantitat d’herba comestible per al bestiar en les pastures de sabana i formacions associades.
Moltes espècies de gramínies posseeixen rizomes subterranis que els permeten brotar abans que comenci l’estació plujosa, sovint després dels focs de cap al final de l’estació seca. En el cas dels sòls de baixa disponibilitat d’aigua, l’efecte estacional de la precipitació és reforçat per un estrès hídric de tipus edàfic. A més, les gramínies es caracteritzen per un ús poc eficient de l’aigua, a causa de l’escàs control que tenen de les pèrdues d’aigua per transpiració; així, al començament de la temporada de sequera, la coberta de la vegetació es deshidrata.
Els recursos de les plantes llenyoses
En contrast amb les herbàcies, les plantes llenyoses presenten un sistema d’arrels força extens que té la capacitat d’explotar un volum de sòl considerable. Tanmateix presenten estratègies diferents en funció de com aconsegueixen sobreviure durant l’estació seca.
Les plantes llenyoses sobreviuen gràcies a una superfície foliar reduïda, com també a l’esclerofil·lisme de les fulles, que també poden ser caduques. Aquestes adaptacions restringeixen la taxa de pèrdua d’aigua, però també limiten la taxa màxima de fotosíntesi. Generalment, les plantes amb aquesta forma de creixement (bàsicament arbres) floreixen i produeixen fulles noves unes poques setmanes abans del principi de les pluges. Aquest naixement primerenc els possibilita un accés preferent als pol·linitzadors i a l’increment de nutrients que es produeix al principi de l’estació humida. Aquesta desclosa és possible gràcies a l’emmagatzemament d’aigua i hidrats de carboni de l’estació humida anterior. L’accés a l’aigua emmagatzemada a un nivell profund del perfil del sòl també pot col·laborar en aquesta plenitud primerenca, però no és l’úni- ca raó, si fos així, els arbres no haurien perdut les fulles anteriorment. Una segona estratègia de supervivència consisteix que les parts aèries de les plantes morin durant l’estació eixuta. La planta sobreviu en forma de tubercle, rizoma o de bulb subterrani. Moltes plantes llenyoses de poca alçada, tal com succeeix amb les herbàcies, pertanyen a aquesta categoria. Aquesta forma de creixement no solament permet evitar la sequera, sinó que també aporta resistència enfront el foc i el pasturatge.
En les formacions llenyoses tropicals es troben molts exemples de veritables arbres subterranis. Són plantes amb un òrgan subterrani gran, llenyós i perenne, que tenen només unes poques fulles i branques baixes que sobresurten a la superfície. En emplaçaments de baixa fertilitat, sorrencs i amb incendis freqüents, poden originar una formació llenyosa o fins i tot un bosc d’un metre d’alçada amb prou feines. La seva fenologia és sovint antiestacional, i creixen durant l’estació seca, després dels incendis, de tal manera que eviten la competència amb les herbes. Com a conseqüència, la majoria d’aquestes plantes són molt poc palatables o fins i tot verinoses per als mamífers.
En espècies caducifòlies, la pèrdua del fullam s’associa a la reducció de la disponibilitat hídrica del sòl. Per altra banda, en les formes perennifòlies, l’activitat metabòlica es manté tot l’any, encara que disminueixi en la temporada de sequera. Així, en les sabanes arbustives d’Àfrica sud-occidental, amb un estrat herbaci dominat per Aristida uniplumis i un de llenyós amb la caparidàcia Boscia foetida i les bignoniàcies Catophractes alexandrii i Rhigozum trichotomum, B. foetida transpira diàriament el 7,8% del seu pes sec, i C. alexandrii i R. trichotomum en perden aproximadament el 5%. A les sabanes dels “llanos” de l’Orinoco, les pèrdues diàries d’aigua de la dil·leniàcia arbòria Curatella americana sobrepassen els 16 litres durant la temporada de sequera.
S’ha relacionat la contínua activitat metabòlica de les espècies perennifòlies amb les fonts permanents d’aigua que es trobarien al nivell freàtic o a les capes de sòl permanentment humides. Així, les plantes llenyoses i les herbàcies no competirien per l’aigua emmagatzemada al sòl, ja que les herbàcies tenen sistemes d’arrels superficials i les llenyo- ses tenen accés a l’aigua emmagatzemada en capes més profundes, per sota de les arrels de les gramínies. La hipòtesi ja proposada indica que hi ha una separació espacial de les fonts de subministrament hídric per als dos grups de formes biològiques i, així, no tindrà lloc una competència per l’aigua. Tanmateix, aquesta hipòtesi no explica l’omnipresència d’espècies perennes en sabanes amb sòls divergents per les seves característiques físiques (amb restriccions per a l’emmagatzemament de l’aigua i la penetració d’arrels o amb perfils que comporten un nivell freàtic profund). Tal paradoxa ha estat dilucidada per investigacions sobre la capacitat funcional de les plantes llenyoses perennes que han permès constatar tres tipus d’adaptacions. En primer lloc un quocient alt entre la superfície d’absorció d’aigua i la superfície d’evaporació, fins i tot quan el creixement del sistema radicular dels arbres es veu limitat per una crosta superficial; en segon lloc, un transport eficient de l’aigua per a l’activitat metabòlica; i, finalment, una separació temporal i espacial del nínxol ecològic que permet la coexistència de les activitats fenològiques dels arbres i de les herbes.
Fitòfags, sapròfags i zoòfags: els consumidors
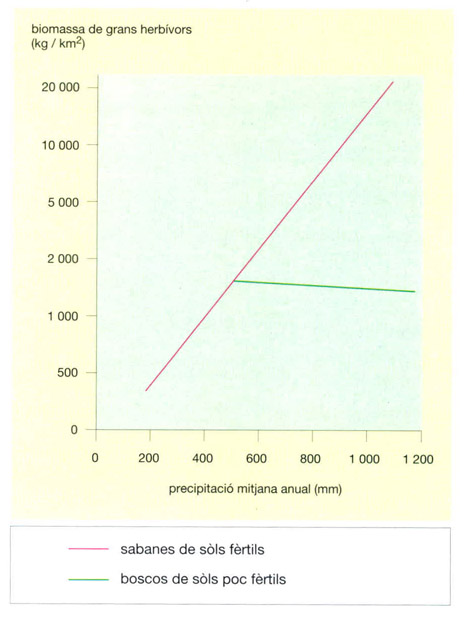
La biomassa de grans herbívors de les àrees protegides de les sabanes africanes varia segons el tipus de comunitat vegetal que es desenvolupi, cosa que a la vegada és determinada per les condicions climàtiques i edàfiques. En terrenys fèrtils, sobre els quals creixen sabanes obertes i gramenets, la biomassa per unitat de superfície augmenta en incrementar-se les pluges. En canvi, a les sabanes arbrades que creixen sobre sòls pobres, la biomassa de grans herbívors disminueix, bé que molt lleugerament, en augmentar la precipitació mitjana anual (la biomassa s’ha representat sobre una escala logarítmica), ja que les pluges permeten el desenvolupament de plantes llenyoses menys palatables i més difícils de digerir.
Jordi Corbera, a partir de dades proporcionades per l’autor
Més enllà de la diversitat del poblament vegetal o animal, les sabanes presenten característiques generals comunes en el seu funcionament tròfic i energètic. Així, per exemple, l’ús de la matèria vegetal sintetitzada es porta a terme seguint l’esquema habitual de tots els ecosistemes, però tot i així, les característiques pròpies de les sabanes, i concretament el baix contingut de proteïnes de les fulles de gramínies i la seva mediocre digestibilitat, influeixen molt en l’abundància relativa dels diferents components de la xarxa tròfica i, per tant, en les modalitats del funcionament ener-gètic.
La producció primària de les sabanes és consumida per una gran varietat d’herbívors, encara que la fracció de producció vegetal consumida per tots ells, és probablement inferior a un 10%. Els més importants en termes de biomassa, fins i tot per damunt dels grans mamífers, són segurament els insectes, en particular llagostes i erugues. La biomassa de grans mamífers, també considerable, sobretot a l’Àfrica, varia molt d’un continent a un altre i d’un tipus de sabana a un altre. És més gran a les sabanes herboses, més seques i més fèrtils, que a les llenyoses, que disminueix amb l’augment de les pluges. En efecte, bé que la producció total de les plantes augmenta amb l’increment de pluviositat, la palatabilitat i digestibilitat del conjunt de la biomassa vegetal disminueix. Per altra banda, l’elevat contingut tànic i fenòlic de les fulles dels arbres de les sabanes, sobretot de les llenyoses, té com a conseqüència que les fulles mortes i caigudes siguin resistents a la descomposició. Abans de la caiguda de la fulla, gairebé tots els nutrients que no estan dins de les parets cel·lulars són retirats de la fulla i reciclats per la planta. Això, juntament amb els productes químics secundaris, fa que els residus de fulles de les formacions llenyoses tropicals siguin un substrat de ben poc profit per als organismes descomponedors.
Els balanços i fluxos energètics i les relacions tròfiques
Per tal de conèixer ben bé l’aspecte dinàmic del funcionament de l’ecosistema, cal que siguin quantificades les transferències tròfiques d’un grup de consumidors a un altre. Aquestes transferències es poden expressar en biomasses, però és més racional traduir-les en equivalents energètics.
A escala d’un individu o d’una població, el balanç energètic pot expressar-se per dues relacions que reflecteixen simplement el principi de conservació de l’energia. La primera,
I = NA + A
expressa les dues fraccions en què es pot dividir l’equivalent energètic de l’aliment ingerit (I>): l’energia no assimilada (NA), i per tant excretada, i l’assimilada (A>). Només aquesta segona es tradueix en producció de biomassa (P) si bé una part és dissipada en la respiració (R); de manera que es pot escriure:
A = P + R
L’equivalent energètic de la producció de matèria (P) reflecteix l’energia invertida en el creixement i la reproducció, és a dir, en la síntesi de matèria orgànica nova, mentre que R representa l’energia que és dissipada per la respiració. El valor de R equival al conjunt de les despeses de funcionament de l’organisme (per exemple, en el treball mecànic i químic, el manteniment de la temperatura, etc.) i correspon a l’energia química que deixa per mineralització la fracció vivent de l’ecosistema. La respiració, en alguns casos, es pot mesurar directament pel conjunt de les poblacions que viuen en un mateix biòtop, com és el cas del conjunt dels organismes del sòl. S’ha aconseguit també alguna vegada seguir a distància l’energia respiratòria d’un animal, ocell o mamífer, retransmesa per biotelemetria. En general, però, la determinació de R d’una manera directa al camp no és possible gaire sovint, i aleshores és necessari fer estudis de captivitat, en condicions tan similars a les de la natura com sigui possible.
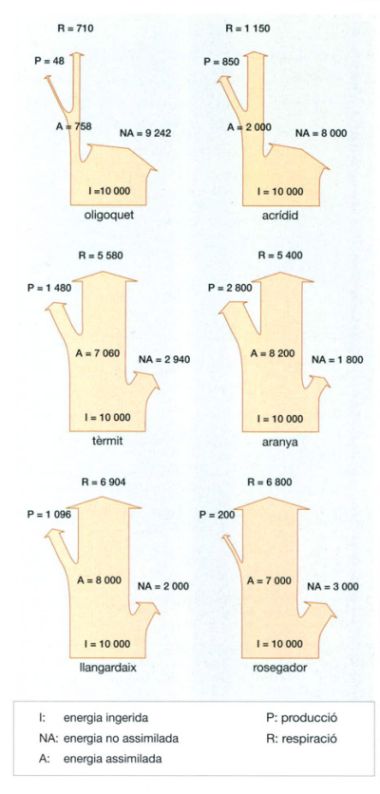
Els balanços energètics d’algunes espècies abundants a la sabana de Lamto, a la Costa d’Ivori, permeten comparar l’eficiència amb què espècies de diferent categoria taxonòmica i grup tròfic utilitzen l’energia que consumeixen. Del total d’energia que ingereix un animal (I), una part no és assimilada (NA), i de la que sí que ho és (A), una part es perd per respiració (R) i l’altra es destina a la producció (P), és a dir, al creixement de l’individu i a la seva reproducció. L’eficiència en l’assimilació de l’energia consumida és determinada tant per la posició taxonòmica com per la categoria tròfica i les característiques fisiològiques de cada espècie. Fins i tot en un mateix individu, l’eficiència varia segons l’estadi de desenvolupament i l’estació de l’any, que determina la quantitat i la qualitat de l’aliment disponible i, per tant, el valor nutritiu de la dieta bàsica. En línies generals, però, es pot afirmar que els animals detritívors, com per exemple els oligoquets, són els menys eficients, ja que tan sols poden utilitzar una part molt petita del total que ingereixen. En canvi, els carnívors, com per exemple les aranyes, són capaços d’assimilar la major part de l’energia que han consumit.
IDEM, a partir de Godall, 1986
Les relacions A/I (coeficient d’assimilació), P/A (coeficient de creixement tissular) i P/I (coeficient ecològic de creixement) expressen les característiques principals del balanç energètic. Tanmateix, per a una població és interessant definir, a més, les relacions P/B i I/B de la producció i del consum de la biomassa mitjana, tant si el marc de vida és d’una generació, com si és d’un any. La relació P/B caracteritza la taxa de renovació de la biomassa. Aquests balanços energètics s’han d’extrapolar al conjunt de l’ecosistema, extrapolació sortosament facilitada per les analogies en el funcionament tròfic que es troben força sovint entre espècies que pertanyen a una mateixa unitat taxonòmica. Es basa també en l’existència d’algunes relacions generals de les quals la més útil és la que vincula la producció d’una generació a la seva biomassa. Aquesta relació P/B pot ser, en efecte, deduïda de les corbes de supervivència i de creixement de l’es- pècie, característiques biològiques relativament ac- cessibles: dóna la producció anual a partir de la biomassa mitjana i del nombre de generacions per any. El coneixement de la xarxa tròfica i dels balanços energètics dels diversos grups taxonòmics permet aleshores definir els fluxos d’energia d’una manera més precisa com més detallada és la xarxa tròfica.
Aquest esquema de funcionament seria aplicable teòricament a qualsevol ecosistema, eventualment, amb algunes modificacions relatives als grups taxonòmics implicats. Tanmateix, els ecosistemes amb predomini de la vegetació herbàcia, i més especialment les sabanes, imposen sempre determinats constrenyiments que en concreten l’originalitat funcional. Es poden analitzar seguint el destí de la matèria vegetal que és a la base de tota xarxa tròfica i que, amb el control dels factors climàtics, determina les modalitats de les transformacions ulteriors successives d’aquesta matèria. Després de la primera fase, i fonamental, del consum d’una part de la matèria vegetal encara viva —la fitofàgia—, hi ha dues destinacions diferents per la part no consumida. La primera és una mineralització brutal per l’acció del foc; la segona és una descomposició progressiva pel complex joc de les innombrables interaccions entre els animals, els fongs i els bacteris. A les fases de fitofàgia i descomposició s’afegeixen, a tots els nivells, fenòmens de depredació i de parasitisme, que també diversifiquen les transferències de matèria que impliquen els fluxos d’energia: tots ells es poden reagrupar amb el nom de zoofàgia.
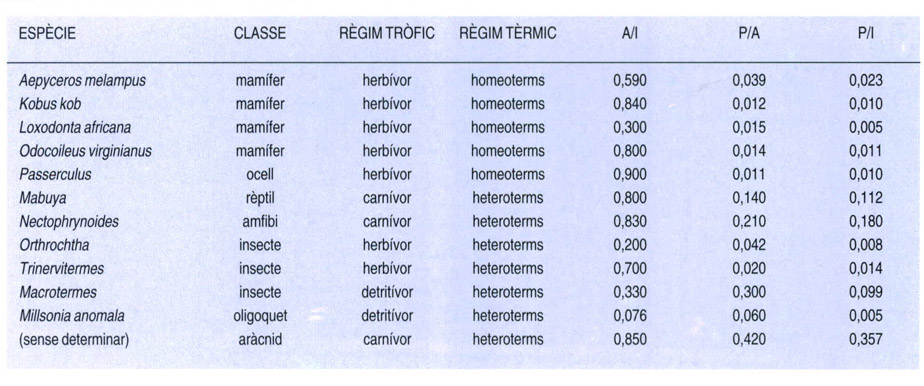
Fluxos energètics de diferents espècies animals i els seus règims tèrmic i tròfic (I=energia ingerida, A=energia assimilada, P=producció). La producció neta es refereix tant al propi creixement com a la reproducció. A partir dels valors d’I, A i P, és possible calcular diferents coeficients que donen una idea de l’eficiència amb què cada organisme aprofita els aliments a la seva disposició. El coeficient d’assimilació (A/I), per exemple, representa la proporció d’aliment ingerit que és realment assimilat per l’organisme, molt variable segons l’espècie. Però el més utilitzat és el coeficient ecològic de creixement, que resulta del quocient P/I. Aquest coeficient varia considerablement amb l’edat de l’animal i amb el tipus de règim alimentari. Bé que hi ha moltes excepcions, en general l’eficiència és més elevada en els animals carnívors, especialment en els invertebrats, que en els herbívors. Així, el valor més elevat del quadre és el que correspon a l’aràcnid (P/I=0,357), mentre que l’elefant africà (Loxodonta africana) presenta un dels valors més baixos (P/I=0,005).
Dades proporcionades per l’autor
A partir dels règims alimentaris de les diverses espècies, es poden reconstruir les relacions tròfiques que les uneixen i establir la xarxa tròfica de l’ecosistema en què s’integren. Tanmateix, atesos el nombre sempre elevat de les espècies que formen una població de sabana (de prop d’uns quants centenars, o fins i tot milers) i l’eclecticisme alimentari de la majoria d’elles, una xarxa tròfica completa d’una sabana és, evidentment, impossible de conèixer amb detall. Per a tenir-ne una idea prou aproximada es poden aplegar primer en un mateix grup tròfic les espècies que tenen comportaments alimentaris propers, per exemple els diversos antílops o el conjunt dels petits rosegadors o fins i tot de les aranyes. També és desitjable no tenir en compte espècies rares, en general molt nombroses a causa de l’estructura específica habitual de les biocenosis. Per altra banda, moltes de les relacions tròfiques són purament fortuïtes i es pot simplificar la xarxa no prenent-les. En un medi que té alhora un tapís herbaci i un estrat llenyós ni que sigui discontinu, també es podrà, en una primera aproximació, presentar per una banda la xarxa tròfica de l’estrat herbaci, de l’altra la de l’estrat arbori, i considerar només, a partir d’això, les relacions tròfiques entre els dos conjunts.
La fitofàgia: pasturadors, brostejadors i altres herbívors
El consum de la matèria orgànica vegetal, fonament de totes les xarxes tròfiques, té modalitats molt diverses, fins i tot pot variar al si d’un tipus determinat d’ecosistema, i encara més en totes les comunitats vegetals considerades aquí com sabanes. L’originalitat principal de les sabanes és donada per l’abundància d’herbes, principalment gramínies, que co- existeixen amb arbres i arbustos les fulles dels quals són també utilitzades. Així el règim dels brostejadors o folívors (“browsers”) s’ha contraposat al dels pasturadors o pròpiament herbívors (“grazers”), bé que moltes espècies tenen un règim mixt.
Entre els herbívors pasturadors hi ha el búfal afri- cà (Syncerus caffer) o l’hipopòtam (Hippopotamus amphibius), però també les zebres (Equus burchelli i altres congèneres), els nyus (Connochaetes) i altres antílops. Alguns tèrmits dels anomenats farratjadors, com els del gènere Trinervitermes, consumeixen també herba que tallen, en petits trossets, durant la nit i porten en filera cap al niu. Molts ortòpters, especialment acrídids, s’alimenten també de gramínies. Diversos petits rosegadors són també, almenys en bona part, afeccionats a l’herba tendra.
Entre els brostejadors folívors cal recordar evidentment la girafa (Giraffa camelopardalis), però també el gran cudú (Tragelaphus strepsiceros), el “gerenuk” (Litocranius walleri), i altres formes més petites, com els dic-dics (Madoqua) o algun duiker comú (Sylvicapra grimmia), que sovint consumeixen també fruits caiguts a terra i petites plantes herbàcies dicotiledònies a més de les fulles de les llenyoses. A la sabana australiana, on les fulles d’Eucalyptus contenen olis aromàtics, una gran diversitat de brostejadors marsupials s’han adaptat a menjar aquestes fulles, però no consumeixen una part gaire gran del total de les fulles disponibles; de fet, al continent australià, els únics grans herbívors que hi havia abans de la recent introducció del bestiar oví i boví, del cavall i del búfal aquàtic, eren marsupials, com els cangurs, que mai no es presentaven en gran nombre.
Altres espècies tenen un règim mixt de fulles d’arbre i d’herba, en proporcions variables, com les gaseles pròpiament dites (Gazella), l’antílop equí (Hippotragus equinus), els èlands (Taurotragus) i, per descomptat, els elefants. Caldria afegir, a aquesta categoria, les formigues talladores de fulles de l’Amèrica del Sud, encara que, segons les espècies, tant puguin ser consumidores de gramínies com de fulles de dicotiledònies. A l’Amèrica del Sud, abans de la introducció del bestiar domèstic d’origen europeu fa pocs centenars d’anys, les formacions llenyoses tropicals havien tingut un nombre extraordinàriament petit de grans mamífers brostejadors durant tota la seva història evolutiva. El principal brostejador indígena és la capibara (Hydrochaeris hydrochaeris).
Les fulles dels arbres —per tant, de dicotiledònia— són en general relativament riques en proteïnes, i molt especialment les fulles d’acàcia, que es troben entre les més consumides. Són un bon aliment que els ungulats, amb l’ajuda de la flora simbiòtica del rumen, o del cec en el cas de les zebres, poden assimilar en gran part. El primer problema és abastar les fulles i, en aquest sentit, tenen un clar avantatge les girafes, que no tenen cap competidor per sobre dels tres metres, i els antílops de mida gran. Un segon problema està vinculat al fet que les fulles dels arbres, sovint riques en tanins, són de difícil digestió, i fins i tot en algunes espècies hi ha substàncies secundàries que les fan verinoses. Per això la biomassa de mamífers brostejadors és relativament petita a les sabanes llenyoses. En els ambients àrids, la defensa de l’arbre està basada sobretot en les espines, però els consumidors folívors ja hi estan adaptats, especialment les girafes.
L’herbivoria basada en el consum de gramínies planteja problemes encara més complexos, vinculats alhora a l’estació i al clima. Al principi del creixement, durant algunes setmanes, els teixits de l’herba, encara tendra, són rics en proteïnes, fins i tot en els ambients humits. Mancades de substàncies secundàries, encara tendres i pobres en silici, les gramínies són un aliment selecte del qual s’aprofiten tant els acrídids i les llebres com els ungulats. A mesura que la planta creix, els teixits es carreguen de cel·lulosa, la proporció de proteïnes disminueix i alhora perden també, a poc a poc, el valor alimentari per als animals. Però, com que progressivament es formen nous rebrots, al consumidor sempre li queda la possibilitat d’escollir, en la mesura que li ho permeti la mida de la seva boca: aquest és el cas dels rosegadors i dels acrídids. La tasca ja és més delicada per als petits ungulats de boca estreta. Per als mamífers de talla gran i boca ampla és difícil, fins i tot si saben seleccionar amb la llengua una part de la mota. Aquesta selecció de l’herba més tendra es posa en evidència per la diferència que sistemàticament hi ha entre el contingut en proteïnes de la sabana i el de l’aliment ingerit.
Per a defugir la dificultat que representa el consum d’un aliment exageradament ric en cel·lulosa, l’herbivoria dels ungulats s’acompanya d’un aparell digestiu molt notable, el funcionament del qual ha estat estudiat amb molta precisió en el bestiar domèstic. L’estómac dels remugadors és enorme i complex. Permet el desenvolupament d’un nombre considerable de microorganismes, bacteris i protoctists ciliats, que garanteixen la digestió de bona part de la cel·lulosa. Més endavant del tub digestiu, l’intestí pot aleshores absorbir els productes d’aquesta descomposició, com també una part dels cossos bacterians que l’han provocada. Una simbiosi com aquesta permet als remugadors utilitzar herba relativament pobra, cosa que en canvi és impossible per a les llagostes o per als rosegadors. Un mecanisme del mateix tipus és present als equins, la zebra per exemple, gràcies a un cec molt desenvolupat. Així i tot, quan el contingut en proteïnes de les gramínies està per sota d’un cert llindar, l’herba esdevé impròpia per al creixement de l’animal, de manera que, fins i tot menjant molt, l’animal perd pes. Aquest fet s’observa en algunes regions, on el bestiar domèstic s’aprima al final de l’estació de les pluges quan l’herba és, en canvi, sobreabundant.
Això fa referència sobretot a les sabanes de les zones humides, com les guineanes en el cas d’Àfrica. Allà, en efecte, les altes temperatures i la gran quantitat de precipitacions asseguren a les plantes un creixement molt ràpid però determinen per això mateix una limitació de les sals minerals subministrades a la planta, la qual cosa provoca una insuficiència en la síntesi de les proteïnes respecte a la de la cel·lulosa. A les sabanes llenyoses humides, el contingut de nitrogen de l’estrat herbaci graminoide només supera el llindar de l’1% (el mínim necessari per als remugants per a complementar els bacteris intestinals que digereixen la cel·lulosa) durant la primera part de l’estació de les pluges o just després d’un incendi. En canvi, així que el clima esdevé més sec el factor limitador ja no són les sals minerals, sinó la manca d’aigua. El contingut en proteïnes és aleshores més alt, encara que disminueix al llarg del creixement, i l’herba es manté comestible força més temps, de vegades fins que mor. Aleshores, fins i tot l’herba seca esdevé un aliment de valor, una mena de palla o farratge natural. Això passa al Sahel, on el consum de la vegetació herbàcia epigea pot ser aleshores total.
Hi ha una diferència important en l’ús de les gramínies segons que es produeixin en clima més o menys humit. A les zones on la qualitat de l’herba esdevé ràpidament insuficient, els animals herbívors han de desplaçar-se per cercar herba comestible o, eventualment, completar la seva dieta amb les fulles més riques dels arbres. Quan el foc destrueix les grans herbes seques, la rebrotada aporta un aliment especialment valuós, que explica la concentració d’animals en aquests sectors. Els caçadors, que cremen parcel·les limitades, i es posen a l’aguait, coneixen bé aquest fenomen i se n’aprofiten.
Tot i que els grans mamífers no consumeixen una part gaire important de la biomassa vegetal, també poden tenir un impacte important en l’ecologia de les sabanes, no sols amb relació a espècies concretes i amb la composició florística d’algunes comunitats, sinó fins i tot pel que fa a l’estructura de l’hàbitat. Els grans herbívors, tant salvatges com domèstics, poden afectar el cicle de nutrients mitjançant l’addició de fems però també pot tenir com a conseqüència la compactació del sòl, com passa a les pastures naturals explotades per ramats nombrosos de grans herbívors. A molts indrets, la reducció de biomassa s’ha atribuït al trepig pels herbívors, tant si són bous com capibares, elefants com nyus, i no directament al consum que fan. L’excés de pasturatge és causa de la reducció de les taxes d’infiltració i de la intensificació de l’erosió del sòl.
Entre els diferents grups animals que formen el conjunt dels fitòfags, cal fer esment encara dels consumidors de llavors o de fruits (ocells, rosegadors, algunes formigues i alguns hemípters heteròpters), que són nombrosos sobretot a les sabanes seques, on les llavors són més abundants; el seu ús no provoca problemes especials ja que es tracta de midó i de proteïnes de fàcil digestió. Passa el mateix amb els succionadors de saba (o del contingut d’algunes granes), com diversos homòpters (pugons, cotxinilles, aleurodes i cicadinis), que d’altra banda només fan servir una part dels glúcids, i molts heteròpters (amb exclusió dels redúvids). Un quart grup el formarien aquells que s’alimenten d’altres matèries vegetals (nèctar, pol-len, espores), com nombrosos dípters, en particular drosòfiles, o himenòpters mel·lífers, o collèmbols, o diverses formigues. I, finalment, els que s’alimenten d’arrels, com els nematodes fitoparàsits i diverses larves de coleòpters.
La saprofàgia: detritívors i humívors
El material vegetal, un cop mort i degradat pels microorganismes del sòl (els quals són, de fet, per a molts animals, el veritable aliment), és ingerit per espècies sovint qualificades de detritívores o sapròfagues, especialment cucs de terra dels horitzons superficials, tèrmits cultivadors de fongs, paneroles, diverses larves de coleòpters i miriàpodes diplòpodes a la virosta i símfils en el sòl. Aquests organismes no descomponen necessàriament una gran part de la brossa per si mateixos, però la trenquen i la mouen cap a llocs on els microorganismes la poden descompondre més ràpidament. En un estadi més avançat de descomposició, la matèria vegetal íntimament barrejada amb el sòl és utilitzada per altres animals entre els quals destaquen els cucs de terra geòfags i els tèrmits humívors, com els bacteris i els fongs.
Hi ha, en efecte, tres tipus diferents de cucs de terra: els que viuen a la capa de virosta i mengen fulles mortes; els que viuen únicament al sòl i que digereixen la matèria orgànica d’aquest, i els que viuen al sòl, però a la nit s’alimenten a la capa de virosta. Aquests darrers són importants pel fet que traslladen matèria orgànica i nutrients des de la capa de virosta cap a dins del sòl on serveixen d’aliment precisament als geòfags. Allà on són comuns aquests darrers, el sòl té una estructura engrunada, constituïda quasi totalment per boletes fecals de cucs. Els cucs poden processar moltes tones de sòl per hectàrea anualment i són inexistents a les sabanes llenyoses amb pluviositats per sota de 800 mm.
Tal com s’ha fet amb diverses espècies herbívores i carnívores també han estat estudiats els balanços energètics d’un cert nombre d’espècies detritívo- res i humívores (geòfagues). Del tèrmit cultivador de fongs Macrotermes subhyalinus, per exemple, l’espècie més abundant a moltes sabanes africanes, els resultats obtinguts corresponen a un coeficient d’assimilació (A/I) de 0,30 aproximadament i un coeficient ecològic de creixement (P/I) de 0,09 amb una taxa de renovació de la biomassa (P/B) de 6,5. En el cuc de terra geòfag Millsonia anomala, molt abundant a la sabana de Lamto, el coeficient d’assimilació (A/I) és només de 0,08. Aquest valor tan baix és determinat probablement per la dificultat que té l’animal per recuperar les partícules alimentàries enmig de l’enorme massa de terra ingerida. El rendiment ecològic de creixement (P/I), de 0,005, és també molt baix, per més que l’animal sigui un heteroterm. La causa és el considerable treball que representa el trànsit intestinal, com també el desplaçament en el sòl. Aquest valor és sempre menystingut del fet que la producció de mucus no s’ha tingut en compte mentre que representa certament una part considerable de la producció. Un tèrmit humívor, Cubitermes exiguus, geòfag com el cuc M. anomala té un coeficient d’assimilació (A/I) molt baix, 0,15, i un coeficient ecològic de creixement (P/I) també molt baix, de 0,01, més acostats als del cuc de terra que als del tèrmit cultivador de fongs. El coneixement de la dinàmica de les poblacions de les principals espècies sapròfagues —i geòfagues— permet així una extrapolació a escala del conjunt del poblament de la sabana fent el total de les energies que corresponen a les ingestions, a les produccions i a les excrecions, aquí considerables ja que les taxes d’assimilació de tots aquests organismes són mínimes.

Balanços energètics de les sabanes de Lamto, a la Costa d’Ivori, i de Serengueti, a Tanzània. De la producció primària total, una part és consumida pel foc (F), una altra és ingerida pels herbívors (Ih) i pels geòfags i detritívors sapròfags (Isg), de manera que s’aprofita per al creixement i trigarà a reciclar-se, i una altra retorna al sòl mineralitzada, bé pels herbívors (Mh), bé pels sapròfags i geòfags (Msg). La resta de la producció no és utilitzada. Els valors de producció total per a cada sabana figuren al peu de cada quadrat. Les quantitats consumides i reciclades per cada fracció es donen també en milions de kcal per hectàrea i any. Noteu com la sabana de Lamto, molt més humida, és també molt més productiva. Els sapròfags d’aquesta sabana tenen un paper molt més important que el dels herbívors. Mentre que els primers consumeixen aproximadament un 60% de la producció primària total, els segons tan sols arriben a l’1%. En canvi, a la sabana més seca de Serengueti, aquestes diferències no són tan marcades; els sapròfags consumeixen aproximadament el 26% de la producció primària, i els herbívors un 12 o 13%.
IDEM, a partir de fonts diverses
A la sabana de Lamto, per exemple, el paper dels sapròfags (detritívors i geòfags) és molt superior al dels herbívors que utilitzen la matèria vegetal verda, ja que representa un consum de Isg = 61 x 106 kcal per hectàrea i any en lloc de Ih = 1 x 106 kcal per hectàrea i any. La superioritat dels sapròfags sobre els herbívors és menor en una sabana més seca com la del Serengueti, on el consum representa tanmateix Isg = 13,6 x 106 kcal per hectàrea i any enfront Ih = 6,4 x 106 kcal per hectàrea i any.
En la piràmide tròfica, sapròfags i geòfags es col·loquen sempre en el mateix nivell que els herbívors ja que, com ells, consumeixen matèria vegetal. Els resultats dels balanços energètics, per tant, s’han d’afegir als dels herbívors, el conjunt que constitueix el nivell dels consumidors primaris, el que ve immediatament després del nivell dels vegetals clorofíl·lics productors.
La zoofàgia: depredadors i paràsits
Tots els organismes que s’han citat representen una part essencial del grup dels consumidors primaris. Serveixen d’aliment per als depredadors —i paràsits— que formen el grup dels consumidors secundaris. En tots els poblaments animals hi ha carnívors i paràsits que fan servir la matèria orgànica de qualitat que es troba en els cossos dels altres animals, siguin herbívors, detritívors o també carnívors. Gairebé la totalitat de la presa consumida té proteïnes directament utilitzables amb un bon rendiment, a diferència del que passa en el consum dels vegetals, que tenen les molècules de cel·lulosa i de lignina que són difícils d’hidrolitzar, i només són assimilables indirectament, amb l’ajut de microorganismes simbionts.
Els balanços energètics establerts per a diferents espècies zoòfagues han destacat la gran diferència entre els rendiments que corresponen als homeoterms, és a dir els animals anomenats correntment de sang calenta (mamífers i ocells) i als heteroterms, és a dir tots els altres (amfibis, rèptils, artròpodes, etc.). Els coeficients d’assimilació (A/I) són sempre alts, de 0,80 a 0,90, mentre que el coeficient de creixement tissular (P/A) i per tant P/I (= P/A2A/I) són molt diferents segons es tracti d’animals homeoterms o heteroterms. Com als altres medis terrestres, el coeficient ecològic de creixement (P/I) dels carnívors heteroterms és de prop de 0,35, mentre que el dels homeoterms és, al contrari molt baix, inferior a 0,02. El consum de carn animal ofereix per tant un interès evident respecte a la fitofàgia i encara més respecte a la saprofàgia, però té l’inconvenient que la biomassa dels animals-preses disponibles és força més baix que en els vegetals.
Entre els animals zoòfags de la sabana hi ha vertebrats, com els mamífers carnívors i insectívors, llangardaixos, amfibis i molts ocells, i també diversos invertebrats, alguns dels quals viuen a l’estrat herbaci, com els pregadéus i els hemípters redúvids, i altres a la superfície del sòl o dins d’aquest, com els coleòpters caràbids, la majoria de les formigues, alguns àcars depredadors i, menys nombrosos, els miriàpodes quilòpodes. Les aranyes són, després de les formigues, el grup dominant, present arreu, sobre l’herba i sobre el sòl.
Hi ha també organismes que consumeixen a la vegada matèries d’origen vegetal i altres d’origen animal. Per tant pertanyen almenys a dos nivells tròfics. És el cas dels mamífers que s’anomenen omnívors, entre els quals figuren, a la sabana, alguns rosegadors, i el de nombrosos ocells, que són alhora granívors i insectívors. Dintre dels invertebrats, també es troben exemples d’omnívors, com moltes formigues i nombroses espècies de paneroles, grills i saltamartins. Finalment també pertanyen a aquesta categoria alguns microorganismes descomponedors —bacteris i fongs— que fan servir matèries orgàniques molt diverses, tant d’origen vegetal com animal.
Molts depredadors devoren altres animals que també són carnívors, de manera que se situen en un lloc més avançat de la xarxa tròfica. Per això alguns autors els han definit com a consumidors terciaris. Alguns animals, com les serps que s’alimenten d’amfibis i de llangardaixos, es classifiquen entre aquests superpredadors, però és també el cas dels mateixos amfibis i llangardaixos quan consumeixen aranyes o pregadéus.
La zoofàgia no és fonamentalment diferent a les sabanes que en altres medis com les selves. En els invertebrats, per exemple, la composició de les poblacions no ofereix característiques especials. En els mamífers, en canvi, es destaca la importància que sovint tenen les espècies grans, com el lleó (Panthera leo), el lleopard (P. pardus), el licaó (Lycaon pictus), les hienes (Hyaena, Crocuta), etc., congruents amb la gran talla de molts dels herbívors que els serveixen de preses, com els ungulats.
El bestiar i el foc: la sabana antròpica
Els ramats de bestiar domèstic han compartit des de fa milers d’anys amb els grans ramats d’ungulats salvatges l’ús dels recursos de moltes sabanes africanes, encara que la prevalença de la tripanosomiasi o malaltia de la son i del seu vector, la mos- ca tse-tse (Glossina), n’impedeix encara l’extensió, sobretot a les regions de sabana llenyosa. Més recentment s’ha introduït el bestiar domèstic a les sabanes neotropicals (principalment bovins) i a les australianes (principalment ovins). L’assimilació de la matèria vegetal pels animals fitòfags és especialment difícil a les sabanes humides d’herba alta amb un contingut proteic baix. El consum és més fàcil a les sabanes més seques, d’herba més rica en proteïnes, però la part no utilitzada és igualment important. Quan finalitzen les pluges, aquesta part no utilitzada esdevé sempre presa de les flames; aquests focs de brolla són una de les característiques més importants de les regions de sabana. Cap reglament administratiu no ha pogut fins avui dia fer-los desaparèixer i per tant representen un factor que s’ha de considerar com a natural. Remunten, sens dubte, a la nit dels temps i són certament força anteriors a l’acció dels humans, que no han fet res més que incrementar-ne la freqüència.
Altrament, el foc ha estat un factor ambiental natural en l’evolució de les sabanes i totes les seves espècies estan d’una manera o altra adaptades a sobreviure amb una certa freqüència d’incendis. A més, si més no algunes espècies, depenen totalment del foc per a la seva supervivència en un exemple sorprenent d’adaptació selectiva a un factor extern que podria semblar negatiu per al manteniment de qualsevol organisme viu. Conjuntament amb la disponibilitat d’aigua i de nutrients i amb el consum de la biomassa pels herbívors, el foc forma part essencial de la constel·lació de factors determinants dels ecosistemes de sabana i dels làbils equilibris entre les espècies llenyoses i les herbàcies a les regions tropicals i subtropicals del planeta.
Els efectes dels incendis i del pasturatge
El feble contingut de nutrients de l’herba de les sabanes ha estat històricament un dels factors que n’ha limitat l’ús. Les poblacions humanes de les sabanes s’han adequat a les deficiències en la qualitat de l’herba a través d’un ús extensiu del sistema i del foc, que augmenta transitòriament el nivell de nutrients de les pastures, ja que en renova el creixement. En pastures cremades de les sabanes neotropicals, el contingut de proteïna és d’un 15% (25 mg N/g de matèria seca), mentre que en pastures tallades, el contingut n’és del 8% (13 mg N/g de matèria seca).
L’establiment de pastures i l’ús del foc per part dels humans ha tingut un impacte decisiu en la fisiognomia de les sabanes tal com es coneixen actualment. En canvi, hi ha molt poques experiències a llarg termini que posin de manifest els efectes del foc i del pasturatge a tota la gamma de sabanes. Certament, són palesos els canvis en la composi- ció florística de les sabanes per efecte del foc i de l’abús del pasturatge, i en les sabanes massa pasturades, la diversitat pot augmentar amb l’increment de les espècies de poca palatabilitat. Així, per exemple, s’ha remarcat que si s’elimina el foc, el sobrepasturatge estimula la invasió de diferents espècies d’Acacia, que formen mates espesses. En reduir-se l’àrea foliar de les gramínies a causa de la sobreexplotació pels herbívors s’eliminà superfície de transpiració, cosa que ocasiona una disponibilitat més gran d’aigua al sòl, que facilita la invasió d’espècies llenyoses, disseminades pels herbívors.
En el cas de les sabanes llenyoses, quan s’evita que es cremin, la biomassa, la densitat i el recobriment dels arbres s’incrementa i les proporcions de les espècies poden canviar, però les sabanes llenyoses no es converteixen necessàriament en boscos secs i tancats. Generalment, la competència entre els arbres per l’aigua durant l’estació seca limita el seu creixement abans que el cobricel es tanqui i l’estratificació de les espècies llenyoses es complexifiqui.
S’han fet algunes experiències amb parcel·les permanents de sabana en les quals s’han evitat durant un període més o menys dilatat els efectes del foc i de la pastura per analitzar els canvis en la vegetació. A les sabanes dels “llanos” de l’Orinoco, una parcel·la establerta el 1961 ha estat analitzada al llarg de 25 anys de protecció contra el foc i el pasturatge. L’anàlisi indica que, a partir de la protecció, la vegetació canvià de composició específica i de dominància. En la vegetació herbàcia, Trachypogon plumosus, que era la poàcia dominant abans de la protecció, reculà a partir d’aquesta mentre ocupava el seu lloc Axonopus canescens; Andropogon hirtiflorus, una gramínia de port alt que es trobava a la localitat de les sabanes estudiades, augmenta considerablement la seva presència, igual que la invasora Hyparrhenia rufa, una gramínia d’origen africà. Pel que fa a la vegetació llenyosa, la sabana protegida caracteritzada de bon començament per una fisiognomia oberta amb petits bosquetons, ha anat canviant progressivament cap a una sabana arbrada. L’augment de la vegetació llenyosa ha reduït l’estrat herbaci i ha afectat l’estructura vertical de la comunitat.
Com a resultat del baix grau d’herbivoria i de la lenta descomposició de les fulles, les sabanes llenyoses acumulen herba seca que roman a lloc i una capa de fullaraca seca i branquillons. Això, unit al període sec anual, fa que els incendis siguin tan freqüents: una brossa que s’encén fàcilment coincideix amb un clima ideal per als incendis i amb l’abundància de fonts d’ignició.
A les sabanes llenyoses rarament passen més de tres anys sense que es cremi l’estrat herbaci al- menys una vegada, normalment durant l’estació se- ca. Generalment, els incendis són provocats per caçadors i ramaders per eliminar l’herba vella no palatable per als grans herbívors i promoure un nou creixement d’herba més tendra i agradosa o agricultors que preparen la terra per als cultius. En aquesta mena de vegetació, cremar és necessari per a una ramaderia sostenible amb una baixa aportació de fertilitzants i es fa des de fa milers d’anys. Els incendis provocats pels humans no es poden considerar aliens a la natura mateixa d’aquestes formacions, ja que una gran part del seu caràcter paisatgístic natural n’és una conseqüència. En temps pre-homínids, però, ja hi havia focs; els llamps eren, probablement, la principal font d’ignició.
Els típics incendis de sabana, fins i tot els de les sabanes llenyoses, no són gaire intensos, ja que el foc rarament encén les capçades dels arbres. Generalment, crema per l’estrat herbaci, arran de terra. La quantitat de llenya que es consumeix és d’1 a 3 t per hectàrea. Els arbres madurs de les formacions llenyoses tenen una variable susceptibilitat al foc. La majoria estan protegits per l’escorça: tot i que les fulles poden socarrar-se i caure, el tronc de l’arbre no s’arriba a morir per causa del foc llevat que l’escorça estigui danyada, per exemple, per una petita cicatriu causada per un porc espí. Els arbrissons que tinguin la capçada totalment dintre de la zona de les flames poden quedar cremats fins arran de terra, però rarament moren; els arbres de les sabanes tenen una extraordinària capacitat de rebrotar de soca o d’arrels.
El foc i el funcionalisme de la sabana
Hi ha prou evidències que diferents freqüències o una distribució diferent del foc al llarg del cicle anual tenen conseqüències diferents per a la composició de les sabanes, com per al seu funcionament i la seva dinàmica temporal, inclòs l’equilibri entre les diferents formes vitals. D’aquests estudis es pot concloure que el foc és un factor important de mortalitat tant entre les formes llenyoses com entre les herbàcies. Quasi totes les poblacions de la sabana, fins i tot les anuals, queden malmeses pels incendis. També s’ha demostrat que encara que que- din afectades, les espècies també resisteixen una certa freqüència de focs, particularment quan es donen durant l’època seca.
De fet, cada cop sembla més clar que nombroses espècies de la sabana, no solament resisteixen el foc sinó que se n’han tornat dependents. Moltes espècies no dispersen els seus fruits o les seves llavors fins que arriben a ser cremades, mentre que d’altres només floreixen després del foc, el qual pot actuar també indirectament sobre la germinació de les llavors mercès a la modificació de les condicions ambientals que introdueix. Alguns autors suggereixen que els incendis sovintejats de les sabanes són necessaris per al manteniment de les poblacions d’arbustos per mitjà de l’eliminació de la coberta herbàcia i la incidència de la radiació en els llocs on es troben les llavors. Un altre efecte indirecte del foc sobre la germinació pot ser determinat per l’acció de la composició química dels residus de la crema sobre la germinació de les llavors. Els propàguls de la labiada Hyptis suaveolens, per exemple, un mendastre característic de les sabanes americanes pertorbades, semblen germinar amb particular facilitat en sabanes que han estat cremades i pasturades. L’acció del foc, en tot cas, no pot ser reemplaçada per un altre factor amb un efecte directe aparentment similar, com podria ser la pastura intensiva o la defoliació experimental. Fins i tot, mitjançant l’ús de models de simulació, s’ha suggerit que algunes espècies no sobreviurien a la sabana si la freqüència d’incendis no assolís un cert valor mínim, que en el cas d’algunes gramínies és tan baix com un foc cada tres anys, per sota d’aquesta freqüència l’espècie s’extingiria localment.
Les elevades temperatures de les flames que abrusen la part aèria de l’estrat herbaci, que poden assolir els 600°C, no són prou per a produir danys aparents als meristemes apicals de les gramínies que dominen, per exemple, a les sabanes dels “llanos” de l’Orinoco, concretament Axonopus canescens i Trachypogon, que es troben arran i per sota de la superfície del sòl, on les temperatures, només a 5 cm de fondària sota el terra, no ultrapassen els 50°C. En canvi, pel que fa a les llavors, les dades indiquen que la seva supervivència depèn de l’exposició al foc i la resistència a les temperatures altes de les de cada espècie. S’ha comprovat experimentalment que les llavors d’espècies com la curatel·la (Curatella americana) presenten germinabilitat (un 12%) quan les unitats hidratades se sotmeten a temperatures de 55°C durant 60 segons. En llavors d’espècies dels “llanos” sotmeses a l’efecte de temperatures altes i cremes en condicions de camp, s’ha trobat que el tant per cent de germinació en totes les espècies estudiades era molt baix excepte en el cas del “corobore” o garrofer de l’Orinoco (Hymenaea courbaril, família de les mimosàcies), que té llavors de testa dura i impermeable que s’escarifiquen per l’efecte de les temperatures altes. En les espècies restants, el percentatge de germinació minva en funció de la temperatura. Aquests resultats indiquen que la capacitat de germinació de llavors sotmeses a l’efecte de temperatures elevades podria representar una pre-adaptació a l’efecte del foc.
El foc també interactua amb els processos de reproducció asexuada. Així, les espècies llenyoses poden presentar borrons gemnífers al sistema d’arrels que permeten la propagació d’individus després dels incendis o d’altres accions traumàtiques. Aquest tipus de reproducció ha estat observat en espècies molt diverses: flacourtiàcies com el “macapiritú” (Casearia sylvestris), mirtàcies com el “guisaro” o guaiaber de sabana (Psidium guineense), boraginàcies com Cordia curassavica, cesalpiniàcies com Cassia moschata o papilionàcies com la “sucupira” (Bowdichia virgilioides) o Lonchocarpus arnestii.
Una altra estructura que s’ha relacionat amb la resistència al foc és la presència d’òrgans suberosos subterranis, els xilopodis, que presenten teixits gemmífers que podrien originar nous mòduls de vegetació. Els xilopodis són òrgans subterranis típics de les espècies arbòries i arbustives de les sabanes llenyoses, que s’originen a partir de l’hipocòtil o del coll de l’arrel; s’han trobat aquesta mena d’òrgans, per exemple, en rabasses de “camará” (Lantana camara, família de les verbenàcies) sotmeses a incendis freqüents als “cerrados” del Brasil i també en les d’espècies de les sabanes de l’Orinoco, com la dil·lenàcea Curatella ame- ricana, les papilionàcies Bowdichia virgilioides, Galactia jussiaeana o les del gènere Eriosema, o la boraginàcia Cordia curassavica. En totes, del teixit meristemàtic dels xilopodis poden rebrotar nous exemplars.
L’efecte del foc és crític per a la supervivència dels nous plançons de les plantes que intenten establir- se a les sabanes. Si els individus no es cremen, les arrels poden arribar a un desenvolupament adequat per a la supervivència. En cas de focs recurrents, les poblacions es redueixen a causa de la minva de brots i la minva de bancs de llavors per l’eliminació de les estructures reproductives. Així, els efectes espacials i estacionals dels focs determinen les ‘oportunitats’ perquè els plançons nous es puguin establir a les sabanes i arribar a l’estadi reproductiu.
Les estratègies relacionades amb els tipus de reproducció de les espècies de les sabanes cremades són complexes. En les sabanes neotropicals, la reproducció per llavors i estructures vegetatives es dóna en un nombre important d’espècies, però la seva expressió relativa ha de ser quantificada amb relació a l’establiment de nous individus fèrtils a les sabanes. Les anàlisis fetes en sabanes cremades indiquen que les espècies de sabana presenten una sèrie de característiques morfològiques que els donen resistència al foc i els permeten la supervivència en condicions de focs sovintejats. Així, s’han reportat espècies amb tiges que presenten un desenvolupament exuberant de teixit suberós que probablement redueix el dany al càmbium vascular i el teixit meristemàtic. Aquest tipus de desenvolupament es pot observar en espècies com la curatel·la (Curatella americana), el “nancé” o “manteco sabanero” (Byrsonima crassifolia, família de les malpighiàcies) o la rubiàcia Palicourea rigida. Aquestes espècies generalment presenten també fulles coriàcies lignificades que poden resistir el foc.
Els arbres i els arbustos de la regió del “cerrado” presenten adaptacions importants per a resistir el foc com ara escorces gruixudes suberoses, gemmes adventícies que broten després de l’incendi, xilopodis i els tubercles d’alguns arbres que els asseguren la supervivència en l’estadi de plançó. Una altra adaptació important de les espècies arbòries de “cerrado” per a resistir els incendis i la sequera és el seu sistema radicular difús, del qual surten diferents rebrots que creixen fins a arribar a l’estadi adult. D’aquesta manera, individus diferents que a nivell aeri no tenen relació aparent poden estar units a través de les arrels. Encara que les llavors de moltes espècies germinen bé in vitro, es creu que la regeneració per mitjà de llavors és molt rara en condicions naturals. La tortuositat dels arbres del “cerrado” s’atribueix sovint al fet que quan les gemmes apicals són destruïdes pels incendis broten les gemmes laterals. Aquesta particularitat no es troba en els arbres de boscos caducifolis o de boscos plujosos. Malgrat la coincidència general dels investigadors en el fet que el foc té un paper important en l’ecologia d’aquests ecosistemes, hi ha pocs estudis sistemàtics sobre els efectes del foc en les plantes de sabana.
L’acció mineralitzadora del foc
El foc accelera el procés de mineralització al sòl i augmenta momentàniament el contingut de nutrients disponibles per a les plantes. Després de la crema, hi ha un augment de la concentració de nutrients (calci, magnesi, potassi, sodi, fòsfor) en la solució del sòl. Tot i això, aquests cations no es retenen gaire a les partícules del sòl i es lixivien fàcilment o els absorbeix la vegetació. Els ions nitrogenats no augmenten immediatament, ja que el nitrogen es volatilitza en una proporció més gran. Tot i això, la concentració de nitrogen als plançons de la vegetació herbàcia cremada augmenta ràpidament després de l’incendi, la qual cosa indica que existeix una font d’origen diferent. En aquest cas, el foc pot estimular la fixació simbiòtica i no simbiòtica de nitrogen pels bacteris del sòl associat a un increment del pH, la temperatura i la disponibilitat dels nutrients. Entre aquests bacteris s’han reportat espècies dels gèneres Beijerinckia i Clostridium.
La part endògena de la vegetació escapa de la combustió. Sota del sòl, la fracció destruïda és variable. És màxima a les sabanes humides de tipus guineà, ja que la quantitat de combustible produïda és considerable i la part extreta pels herbívors molt baixa. A Lamto, on la producció primària és de prop de 26 t per hectàrea, anualment es cremen aproximadament una mitjana de 8 t per hectàrea, principalment herba, però també una part variable de vegetals llenyosos, depenent dels anys i de la data de l’incendi. La part incendiada és força més reduïda a les sabanes més seques. La producció primària proveeix una quantitat menor de combustible, de manera que el foc és menys violent i la combus- tió esdevé menys completa. A més, en tractar-se d’herba de millor qualitat la proporció consumida pels herbívors és força més important. En el cas que una part de la vegetació epigea no és consumeixi al moment de l’incendi, aquesta representarà, per tant, una fracció força menys gran que a les sabanes humides: així ho mostra la comparació de les sabanes de Lamto i del Serengueti. En els medis encara més secs, i quan l’explotació pels herbívors és més completa, el foc es pot evitar, almenys durant alguns anys; només es produirà quan s’hagi acumulat una quantitat suficient de combustible. Les sabanes de l’Àfrica meridional, per exemple, cremen aproximadament cada tres o quatre anys. Al límit, en els medis en què l’estructura és una estepa de mates d’herba molt discontínues, el foc ja no pot passar. El consum més intens d’aquesta herba de contingut ric en proteïnes també accentua aquesta eliminació del foc.
La part llenyosa de la vegetació de les sabanes també pateix l’acció del foc, més com més intens és el foc, és a dir, com més abundant és l’estrat herbaci. En una sabana en equilibri, la fracció cremada —fulles, branques i troncs morts— correspon sensiblement a la producció. Quan l’acció dels humans multiplica els focs, l’estrat llenyós té tendència a reduir-se i la sabana esdevé més oberta. Quan és possible una protecció davant del foc, en canvi, es produeix un repoblament progressiu que pot portar a un bosc clar.
La vehiculació atmosfèrica dels gasos i les cendres
Des del punt de vista energètic, els incendis són un agent mineralitzador. Eliminen de la biocenosi una part de l’energia química introduïda per la fotosíntesi, en la mateixa dosi que el consum per la població animal i la descomposició pels microorganismes. Durant els indencis, els nutrients de la fullaraca i del sòl poden ser, bé volatilitzats, bé suspesos a les cendres i dispersats amb elles per convecció, bé mineralitzats en formes disponibles per a la vegetació. En els casos de volatilització i dispersió a través de les cendres, les pèrdues de nutrients depenen de la temperatura a què arriba el material. Així, la volatilització del nitrogen i del sofre es produeixen quan s’assoleixen temperatures pròximes als 200°C, mentre que el fòsfor, el potassi, el calci, el magnesi i altres cations necessiten temperatures d’aproximadament 500°C.
Les pèrdues de nutrients de les sabanes neotropicals en un incendi s’han quantificat, pel que fa a nitrogen i sofre, en valors de prop d’11,5 i 3,2 kg/ha. Quant a les pèrdues de cations, poden ser significatives quan es desplacen les cendres a través de la convecció. Les estimacions de les pèrdues totals de nutrients cap a l’atmosfera, per volatilització i convecció de cendres, indiquen una reducció apreciable del contingut de nutrients als sòls de les sabanes. Tot i això, cal assenyalar que existeixen mecanismes meteorològics que permeten la incorporació de nutrients que provenen d’altres sistemes i/o la redistribució en el mateix sistema. Així, una part de les pèrdues es compensen per l’aportació a través de la precipitació. En el cas de les sabanes de l’Orinoco, la precipitació subministra el 19% del nitrogen, el 17% del fòsfor i el 30% del potassi necessaris cal perquè la vegetació herbàcia pugui arribar a una fitomassa de 463 g/m2 durant l’estació de creixement.
Una altra font de nutrients per a les sabanes és l’aportació a través d’aerosols, partícules de pols, sals, grans de pol·len i, sobretot, cendres, que són interceptats per la vegetació i dipositats sobre les fulles i les branques. Aquests residus dipositats són arrossegats per la precipitació, des de les capçades cap al sòl i, en el cas particular del rentat de les tiges, els nutrients són dirigits de dret cap a les arrels de les plantes. D’aquesta manera, els nutrients escampats amb els aerosols s’acumulen a terra al voltant dels individus que els han retingut. La cendra, en concret, és redistribuïda pel vent i els residus que queden al sistema poden ser incorporats al sòl a través de la precipitació. L’acumulació depèn de les característiques físiques de la capa superficial del sòl i de les característiques iòniques dels residus, que reflecteixen la composició del material combustible.
A causa de la seva immensa extensió i de la freqüència amb què cremen, les sabanes representen aproximadament la meitat de la biomassa vegetal que es crema al món. Com a resultat dels focs, són expulsats a l’atmosfera molts gasos en quantitats suficients perquè hi hagi conseqüències d’escala global. Alguns dels gasos residuals, com ara el diòxid de carboni, el metà i l’òxid nitrós, són gasos amb efecte hivernacle, però els incendis de sabanes no són els responsables de l’augment de l’efecte hivernacle que tant preocupa actualment a nivell internacional: sempre s’han cremat, i no hi ha cap evidència que suggereixi que la freqüència global amb què es cremen hagi augmentat. El diòxid de carboni que el foc allibera a l’atmosfera torna a ser capturat al llarg dels anys següents per la vegetació que creix. Alguns d’aquests gasos que resulten dels incendis participen en les reaccions que condueixen a la formació d’ozó a les capes baixes de l’atmosfera. Els incendis de les sabanes estan, segurament, entre les fonts naturals més importants d’ozó. En tot cas, és un fet que l’atmosfera de les àrees de sabana, durant la temporada d’incendis, està permanentment emboirada, a causa de la presència de minúscules partícules de sutge.