El plàncton marí
El plàncton és el conjunt d’organismes que viuen en suspensió a l’aigua. Encara que alguns tinguin moviment propi, el moviment passiu provocat pel desplaçament de l’aigua és sempre més important. El component algal del plàncton, anomenat fitoplàncton, és constituït per algues unicel·lulars, que poden ser lliures o poden formar colònies. Els organismes més abundants del fitoplàncton marí són les diatomees (o bacil·lariofícies), les dinofícies, les coccolitoforals (haptòfits), alguns altres grups de flagel·lats i, ocasionalment, cianofícies. Tots són autotròfics, és a dir, utilitzen l’energia solar i les molècules inorgàniques per a sintetitzar els components orgànics que necessiten per a viure. Ara bé, en alguns casos, i sobretot entre les dinofícies, poden utilitzar components orgànics dissolts en aigua de mar, com a font de matèria i d’energia. Aquesta heterotròfïa pot ser parcial o total.
Les algues del fitoplancton tenen mides molt variables, però les més freqüents estan entre 10 i 200 µm. Aquestes algues són els productors primaris de la mar, i la major part de l’energia que hi va a parar es fixa gràcies a ells. Si els comparem amb els productors primaris terrestres (herbes d’un prat, arbres d’un bosc, plantes d’un conreu), trobem grans diferències, causades per la inestabilitat del medi aquàtic, inestabilitat que també comporta una gran facilitat per al transport de substàncies. En efecte, els vegetals terrestres inverteixen en estructura (arrels, troncs, branques, teixit conductor, etc.) una gran part de l’energia procedent de la fotosíntesi, mentre que, als organismes del fitoplàncton, aquesta energia es canalitza cap a la reproducció, raó per la qual presenten uns temps de duplicació de cada alga de solament un a alguns dies.
Molts dels organismes del fitoplancton són cosmopolites, i la majoria de les espècies tenen àrees de distribució molt grans. Si comparem el fitoplàncton d’una regió amb el d’una altra, les principals diferències no són florístiques, sinó de relació quantitativa entre els diversos organismes. La composició del fitoplàncton de les costes catalanes presenta poques diferències amb la de la resta de la Mediterrània. Les principals diferències de composició que podem observar van lligades a l’estació de l’any i als diferents tipus d’aigua, com ara aigua de ports, de badies, propera a la costa, de mar oberta, etcètera.
Cada espècie fitoplanctònica està adaptada a unes condicions ecològiques concretes, sovint difícils de precisar, ja que ens falta informació sobre una part important de les variables del medi ambient, que poden repercutir de manera diversa sobre el desenvolupament dels diferents organismes. Tot i així, els diferents grans grups d’organismes presenten algunes característiques generals d’abundància i de distribució. Així, les diatomees creixen ràpidament en condicions de turbulència de l’aigua, quan estan ben proveïdes de sals nutritives. Són elles les que dominen en la comunitat del fitoplàncton, quan aquesta presenta el seu màxim desenvolupament. Però quan la concentració de sals nutritives baixa, les diatomees es van fent més rares. En aquestes noves condicions ambientals la població algal és més baixa, i es presenta dominada per les dinofícies, que tenen moviment propi i estan més ben adaptades a la supervivència en condicions d’estratificació i de pobresa en sals nutritives.
Les coccolitoforals (haptòfits) tenen una coberta formada per plaques calcàries. Tendeixen a ocupar una posició intermèdia entre les diatomees i les dinofícies. Els silicoflagel·lats (crisofícies) presenten un esquelet intern de sílice i assoleixen les concentracions màximes durant l’hivern i la primavera. A l’estiu, es refugien en nivells per sota del superficial. Alguns grups de petits flagel·lats, com ara determinades haptofícies i criptomonadals, poden tenir un paper molt important en àrees particulars. Al fitoplàncton hi poden haver fins i tot una sèrie de minúsculs organismes fotosintetitzadors, que no són retinguts en filtres de 2 µm de malla, i que pertanyen a diferents grups de bacteris i d’algues. Poden tenir un significat important pel que fa a la biomassa i a la producció.
Factors condicionadors de la distribució del fitoplàncton
La sedimentació dels organismes

Una pesca de fitoplancton acabada d’agafar amb xarxa de malles molt fines, vora les illes Medes, a començaments del mes de març. L’aspecte floculant és degut a l’abundància de diatomees filamentoses, amb sedes, del gènere Chaetoceros.
X. Llimona
Els organismes del fitoplàncton es troben en suspensió en una aigua sotmesa a un moviment gairebé permanent. Si poguéssim seguir la trajectòria d’una cèl·lula, veuríem que és molt irregular, però que presenta una resultant dirigida cap avall, car la seva densitat, encara que molt propera a la de l’aigua, és sempre una mica superior. La nostra cèl·lula necessita llum suficient per a la fotosíntesi, però la penetració de la llum a la mar es limita a la capa d’aigua superficial. Per sota d’uns 100 m de profunditat, la llum no permet a l’alga fotosintetitzar tot el que necessita. La velocitat de sedimenta ció dels organismes planctònics és variable, però la majoria de les cèl·lules es sedimenten a velocitats compreses entre 1 i 5 m/dia. A causa d’això, la permanència de les algues a la zona il·luminada és condicionada per la turbulència de l’aigua, l’única força capaç de retornar més amunt una fracció del total de les cèl·lules. Molts organismes del fitoplancton presenten expansions en forma de seda, de banya, de collar o d’ala, que frenen com un paracaiguda el moviment de sedimentació. Però totes aquestes adaptacions serien insuficients si no hi hagués la turbulència.
Els elements nutritius limitants
En principi, a l’aigua marina hi ha en dissolució tots els elements i compostos químics necessaris per a la vida dels organismes. Però el fitoplancton n’absorbeix alguns de manera preferent i arriba a deixar l’aigua tan empobrida que les condicions es fan limitants. El fòsfor i el nitrogen, que normalment es presenten en forma de fosfat i de nitrat, són les substàncies que més sovint es tornen limitants a l’aigua de la mar. El fitoplàncton es sedimenta en part, i en part és consumit pel zooplàncton, que se’l menja. D’aquesta manera, els preciosos fòsfor, nitrogen i altres elements limitants van a parar al fons de la mar, mentre les capes superiors se’n veuen gradualment privades. A la Mediterrània, i sobretot durant l’estiu, es pot produir un reciclatge de nutrients limitat a les capes il·luminades i degut principalment a l’activitat dels bacteris.
Una part dels nutrients pot regenerar-se in situ, gràcies, principalment a l’activitat dels bacteris i del zooplàncton, però pot ser molt més important quantitativament el retorn de substàncies nutritives, des de capes profundes a la superfície, a causa dels moviments de l’aigua. A la costa, el desplaçament de l’aigua superficial mar enfora pot fer pujar aigua profunda, més rica en nutrients. Aquests corrents són activats pels vents costaners. Un altre agent de fertilització important vora la costa és la descàrrega dels rius (per exemple, l’Ebre o el Llobregat), que aboquen a la mar les sals nutritives que transporten i, alhora, fan que el corrent d’aigua dolça mar enfora afavoreixi la barreja de les aigües. Hi ha també mecanismes de fertilització general, no tan localitzats com en el cas de les costes, i que tenen influència sobre la producció de la mar catalana. El més important és el que acompanya l’anomenada formació hivernal d’aigua profunda del golf de Lleó, causada per l’enfonsament d’aigua densa, freda i relativament salada, originada en la superfície, que provoca una uniformització de tota la columna d’aigua i l’accés de nutrients profunds a les capes il·luminades.
El plàncton i el cicle físico-químic anual de l’aigua
En efecte, durant l’any, el fitoplàncton passa per una sèrie de canvis estacionals que van estretament lligats a factors físico-químics de l’aigua. Aquests factors ecològics regulen el desenvolupament del fitoplàncton, tant pel que fa a les espècies com pel que fa a la seva abundor relativa. A la tardor, l’aigua superficial es va refredant i la temperatura i la densitat s’uniformitzen en tota la columna d’aigua, que perd estabilitat. Aquestes condicions, unides a l’acció del vent, afavoreixen la barreja de les aigües i, per tant, l’arribada de sals nutritives a les aigües superficials. Això fa que en aquesta època es produeixi a la nostra mar un important desenvolupament del fitoplancton, protagonitzat per unes poques espècies de diatomees pennals, com Asterionella japonica, Thalassionema nitzchioides, Thalassiothrix mediterranea, algunes espècies de Nitzschia, i de centrals, com Chaetoceros (per exemple Ch. compressus) i Rhizosolenia stolterfothii, etc. A mida que avança l’hivern, es va incrementant l’homogeneïtzació de les aigües superficials amb les profundes. En aquestes condicions, el fitoplàncton, dispersat per la turbulència de l’aigua, passa una part important del temps en zones profundes, de manera que, en conjunt, les poblacions no tenen suficient llum per a créixer.
A les nostres aigües, la màxima abundor de fitoplàncton marí es produeix al final de l’hivern o al principi de la primavera, quan en augmentar la temperatura ambient, les capes d’aigua superficials s’escalfen i es tornen menys denses. Es produeix així una classificació de l’aigua per densitats. Aquesta estratificació de la columna d’aigua limita la barreja del fitoplàncton cap a zones poc il·luminades i afavoreix el creixement de les algues. El color de la mar es torna més verdós i l’aigua perd transparència. Però aquesta gran població d’algues microscòpiques captura compostos solubles, principalment de nitrogen, de fòsfor i de silici, fins a arribar a reduir molt llur concentració. Això és especial ment important, perquè, al mateix temps, en anar-se intensificant l’estratificació de l’aigua, acaba formant-se, entre les capes d’aigua superiors, calentes, i les inferiors, més fredes, una zona en la qual, en pocs metres, la temperatura i la densitat de l’aigua canvien fortament, l’anomenada termoclina.
La presència d’aquesta discontinuïtat representa una barrera que disminueix l’intercanvi entre les capes d’aigua superficials i les profundes. A causa d’això, a la zona il·luminada s’esgoten les últimes reserves de sals nutritives, àvidament recercades per les algues, i s’estableix un gradient de nutrients oposat al de la llum. Aquest factor, juntament amb la tendència de les cèl·lules a sedimentar, determina la distribució vertical del fitoplàncton durant l’estiu. La concentració d’algues a les capes superficials, ben il·luminades, però empobrides en sals nutritives, disminueix, i això fa que intensitats de llum apreciables puguin penetrar més profundament. En aquestes condicions, el fitoplàncton acaba formant un màxim entre 40 i 90 m de profunditat, en una zona on, d’una banda, pot tenir accés als nutrients de les capes fondes i rep encara llum suficient per a fotosintetitzar, i d’una altra, l’existència del gradient de densitat redueix la dispersió de les cèl·lules.
La distància a la costa

"Marea roja", provocada per una gran abundància d’individus de Noctiluca scintillans, regolfada al port de Vallcarca (Garraf, maig de 1984); en aquesta ocasió el fenomen s’inicià a les costes de Castelló i acabà afectant tot el litoral català.
Xavier Tomàs / Francisco A. Comín
En general, la concentració de fitoplàncton és més elevada a les costes que a mar endins. Això és causat per factors com la limitació de profunditat i la barreja d’aigües per acció dels vents, afavorida per la topografia de la costa, o per la influència dels rius. En la nostra costa, es produeix un màxim de productivitat costanera vora les desembocadures de l’Ebre i del Llobregat. De vegades, es produeixen acumulacions anormals de dinofícies, que donen colors brunencs o vermellosos a la mar (marea roja). Quan la dinofícia Noctiluca scintillans es fa abundant, comunica una tonalitat rosada a les aigües, que de nit presenten punts lluminosos. Però, si bé en molts llocs les marees roges han provocat grans pèrdues econòmiques, no han estat detectats efectes tòxics significatius a les marees roges mediterrànies.
L’aigua dels ports sol ésser molt rica en matèria orgànica i en sals nutritives. A la primavera i a l’estiu, aquestes aigües es converteixen en una sopa de fitoplàncton, i presenten una intensa coloració. Sovint, el color és vermellós, indici molt probable d’elevada concentració de dinofícies (sobretot diverses espècies dels gèneres Gymnodinium, Prorocentrum i Gonyaulax), algunes de les quals tenen capacitat per a la utilització de matèria orgànica dissolta en l’aigua. Si la tonalitat de les aigües és verdosa, probablement hi dominen les diatomees. La poca renovació de l’aigua dels ports i l’abocament d’aigües residuals condueix a una eutrofització d’aquests sistemes. L’oxidació de l’excés de matèria orgànica, sobretot en condicions d’estratificació tèrmica, provoca sovint l’esgotament de l’oxigen dissolt, que pot tenir conseqüències negatives a la fauna marina.
A la mar catalana, el sistema costaner del delta de l’Ebre és especialment important. A les seves badies marginals es pot trobar una concentració de fitoplàncton diverses vegades superior en quantitat a la que trobem a mar oberta. L’arribada d’aigua dolça té un paper important en la fertilitat d’aquestes badies. El cicle anual del fitoplancton mostra també un màxim de primavera i un altre de tardor, com a la costa prope ra, però hi destaquen la presència d’una abundància relativa molt més gran de fitoplàncton durant l’estiu i una participació important de les diatomees durant tot l’any.
Composició del fitoplàncton
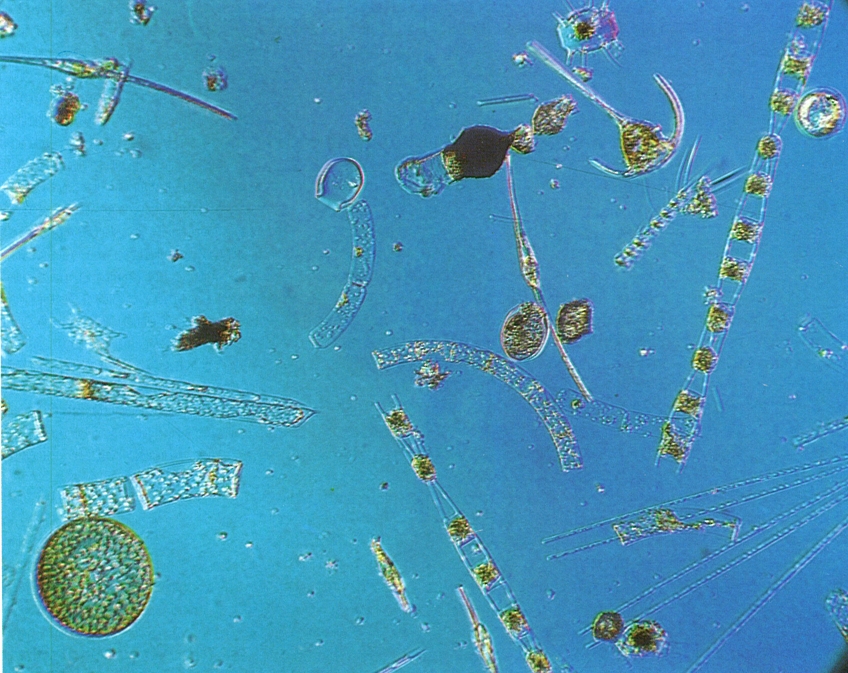
Fitoplàncton de la mediterrània pescat amb malles molt fines i observat al microscopi interferencial. S’hi observen diverses diatomees, com Hemiaulus (cadena amb obertures, a la dreta), Asterionella mediterranea (feix a l’angle inferior dret), Rhizosolenia stolterfothii (cadenes corbades de la part central) i una altra Rhizosolenia (cilindrica, acabada en un esperó, al centre esquerra), un Chaetoceros (cadeneta amb sedes al voltant, vers l’angle superior dret), i una Biddulphia ¡a dalt i a la dreta, amb apèndixs radials a les valves); també s’observen dinòfits, com Ceratium tripos (en forma d’àncora, vers l’angle superior dret) i C. fusus (cèl·lules fusiformes, llargues, al centre i a l’angle superior esquerre).
Claude Carré / Jacana
L’observació del fitoplàncton viu, directament o amb microscopi, és un dels plaers més grans que podem oferir a la nostra vista. Es tracta d’un món d’organismes diminuts, on predomina el color verd de la clorofil·la, quan la comunitat és jove, i els tons groguencs i vermellosos a mida que la successió avança. A causa de la poca profunditat de camp del microscopi, i de la gran distància que hi ha entre els organismes de l’aigua de la mar comparada amb la seva mida, es fa necessari concentrar el fitoplàncton. Això es pot fer filtrant l’aigua de mar amb una xarxa de malles molt fines, arrossegadas des d’una embarcació, o bé deixant sedimentar un determinat volum d’aigua en una cubeta de fons de vidre prim i observant-la per sota amb l’ajut d’un microscopi invertit. Si utilitzem la xarxa, obtenim la fracció del fitoplàncton més grossa, la que forma colònies o tè cèl·lules d’una bona mida. És la fracció més vistosa, i els organismes que en formen part presenten estructures de gran complexitat, que no es veuen tan sovint en el plàncton sedimentat, que comprèn la totalitat dels organismes. En realitat, ambdues tècniques es complementen.
A les nostres aigües podem observar algunes espècies notables, sovint rares, bellament ornamentades, com Ornithocercus magnificus, Amphisolenia bidentata, Ceratium platycorne, C. ranipes, C. candelabrum, i els gèneres Histioneis, Ceratocorys i Dinophysis, entre les dinofícies típiques del període estival; Planktoniella sol, Stephanopyxis turris, Asteromphalus flabellatus, Biddulphia mobiliensis, Corethron hystrix, Grammatophora marina, Asterolampra, Coscinodiscus, Actinopthychus, entre les diatomees; i Coronosphera mediterranea, Discosphaera tubifer, Rhabdosphaera clavigera, Helicosphaera carteri, Ponthosphaera discopora, P. syracusana i Sciphosphaera asteinii, entre les coccolitoforals (haptòfits).
Entre les espècies característiques del màxim del final de l’hivern i del principi de la primavera, hi ha molts nanoflagel·lats (haptòfits, criptòfits, etc.), dinoflagel·lats dels gèneres Gyrodinium, Gymnodinium, Amphidinium, Peridinium i Prorocentrum, i diatomees com Leptocylindrus danicus, L. minimus, Nitzschia pungens, Rhizosolenia calcar-avis, R. delicatula, R. fragilissima, R. imbricata, R. stolterfothii, Skeletonema costatum, Hemiaulus sinensis, Bacteriastrum, Eucampia zodiacus, Chaetocerós decipiens, Ch. lorenzianus, Ch. curvisetus, Ch. pseudocurvisetus, Ch. peruvianus i diverses espècies de Thalassiosira. Totes aquestes diatomees formen cadenes o associacions en les quals les cèl·lules es disposen en rengles. L’única espècie de coccolitoforal que es troba de manera abundant és Emiliania (= Coccolithus) huxleyi.
Al màxim de tardor, les espècies més abundants són diatomees pennades, com Asterionella japonica, Thalassionema nitzschioides, Thalassiothrix frauenfeldii i T. mediterranea.
Hi ha algunes espècies del fitoplàncton que podem trobar durant tot l’any. Citem com a exemple Ceratium fusus, C. furca, Dinophysis caudata i Dictyocha fíbula. Altres espècies, com Noctiluca scintillans i Ceratium fusus, escintillen en la foscor, i comuniquen lluminositat a l’aigua de la mar. Algunes espècies que podem trobar en el plàncton, com les dels gèneres Diploneis, Pleurosigma, Navicula, Amphora i Amphiprora, són d’origen bentònic, però poden passar molt temps en suspensió a l’aigua.
Importància del fitoplàncton
El fitoplàncton és la part més important de la primera baula de la cadena tròfica de la mar. Els organismes que el componen utilitzen energia lluminosa, sals minerals i carboni dissolts en l’aigua per a sintetitzar matèria orgànica rica en energia. Aquesta fotosíntesi que té lloc al fitoplàncton representa la porta bàsica d’entrada d’energia als ecosistemes marins. La resta dels organismes que viuen a la mar depenen, directament o indirectament, de la matèria orgànica fabricada per la fotosíntesi del fitoplancton.
L’abundància de fitoplàncton va lligada als moviments advectius (horitzontals) i turbulents de les aigües, i aquests depenen en bona part de l’energia externa aportada per l’atmosfera i la hidrosfera. La manera més senzilla de determinar l’abundància relativa de fitoplàncton, a més de l’apreciació visual, és la mesura de la transparència de l’aigua que s’obté enfonsant lentament un disc blanc de 30 cm de diàmetre (disc de Secchi), penjat horitzontalment d’una corda i anotant la profunditat en què hom deixa de distingir-lo. Hi ha altres procediments, més laboriosos, com l’anàlisi de pigments fotosintetitzadors continguts en un volum d’aigua i el recompte, amb l’ajut d’un microscopi invertit, del nombre de cèl·lules sedimentades sobre la base d’una cubeta, a partir d’un volum conegut d’aigua de mar. A la Mediterrània, el fenomen que més contribueix a la producció del fitoplàncton durant tot l’any és el desenvolupament massiu que té lloc al final de l’hivern i a la primavera. L’etapa més pobra és l’estival.
Així com la vegetació dels ecosistemes terrestres ens mostra una composició en espècies i una abundància d’aquestes que varia de manera estacional i també espacial, en relació amb les condicions del medi, el fitoplàncton presenta unes certes pautes semblants. Hi podem apreciar una relació entre les propietats del medi i la seva composició en espècies. En aquest sentit cal recordar la successió, ja explicada, que va des dels medis sotmesos a barreja d’aigües per turbulència i molt fèrtils, dominats per les diatomees, fins a medis estratificats i pobres, on destaquen, encara que més rarament, els dinoflagel·lats. A partir d’un esdeveniment pertorbador del medi, com la introducció de quantitats importants de nutrients a la capa il·luminada de la mar, s’inicia una successió d’organismes. Aquesta successió no arriba als nivells de maduresa assolits pels ecosistemes terrestres, a causa de la inestabilitat del medi aquàtic.
El bentos marí

Zonació de comunitats sobre roques batudes del cap de Creus (Alt Empordà). De dalt a baix s’hi observen les comunitats de Chthamalus stellatus (glans de mar), Rissoella verruculosa, Lithophyllum tortuosum, Cystoseira mediterranea i Corallina elongata.
Enric Ballesteros / Javier Romero
El bentos marí és el conjunt dels organismes que viuen fixos al fons de la mar, o en íntima relació amb ell. Si tenim en compte que les tres quartes parts de la superfície del món estan cobertes per la mar, ens adonarem de l’extensió del bentos a nivell mundial. Però hem de considerar també, que l’aigua és un medi menys transparent que l’aire, i absorbeix la llum que travessa les seves capes superiors, de forma que per sota de 100-150 metres de profunditat, hi ha foscor total. Com que la profunditat mitjana dels oceans és d’uns 4000 m, és fàcil veure fins a quin punt són limitades les zones on la llum fa possible el desenvolupament de vegetals bentònics. En efecte, aquests resten limitats a una estreta franja que envolta les terres emergides, i que representa un percentatge molt migrat del total dels fons marins.
Com que les algues no tenen òrgans excavadors com les arrels, necessiten gairebé totes un substrat rocós sobre el qual fixar-se, i això redueix encara més la superfície apta per a la vida de les algues bentòniques. Així doncs, les algues bentòniques queden restringides a uns hàbitats relativament poc extensos, però llur accessibilitat fa que els donem una importància en els oceans que, a escala global, no tenen. Ara bé, en aquests hàbitats, la vegetació algal sovint pot ser qualificada d’exuberant, amb comunitats algals d’una complexitat estructural i d’una riquesa específica realment notables, així com d’una curiosa vistositat, en una escala relativament petita, podríem dir que "miniaturitzada".
Als apartats que seguiran, oferirem una visió succinta de les comunitats del bentos marí amb presència vegetal des d’una perspectiva triple: autoecològica (amb l’estudi dels factors que influeixen en la vida i la distribució de les algues), paisatgístico-bionòmica (amb la descripció de les principals comunitats del nostre litoral) i dinàmica (amb el paper de les algues en el sistema bentònic).
Factors condicionadors de la distribució de les algues bentòniques
Si volem comprendre la vegetació del bentos marí, és millor que oblidem els coneixements que hem après sobre la vegetació terrestre. En efecte, les "regles del joc" són totalment diferents: hi ha aigua abundant, no hi ha sòl d’on extreure nutrients, la llum és filtrada per l’aigua, la gravetat és compensada per la flotació, etc. Els factors més importants que tenen influència sobre la distribució de les algues són el substrat, l’hidrodinamisme, la llum i la temperatura; tenen també una certa importància la pressió, la concentració de nutrients, la salinitat, etc. En general, aquests factors actuen en conjunt, interaccionant entre ells. A més, cal afegir-hi el que en diem factors biòtics (interacció entre les espècies). De fet, no hem d’oblidar que l’acció de l’ambient sobre les espècies actua modificant les relacions de competència.
El substrat
En contrast amb el que succeeix amb la vegetació terrestre, la interacció entre l’alga i el substrat és dèbil, ja que l’alga no n’extreu res, i solament l’utilitza com a suport. Per això, la interacció és més de tipus físic que no pas químic. La importància de la natura física dels fons es reflecteix en les grans diferències que hi ha entre la vegetació dels fons rocosos i la dels fons de sediment. Als primers, la duresa, d’una banda, i la textura i el microrelleu de la roca, d’una altra, semblen influir decisivament en la fixació de les espores. En fons de material fi, la mobilitat d’aquest material és el factor negatiu més important.
L’hidrodinamisme
L’hidrodinamisme, o moviment de l’aigua, pot consistir en moviments de vaivé (ones) o en fluxos unidireccionals (corrents). A les zones situades una mica més amunt del nivell de la mar, l’hidrodinamisme és sinònim d’humectació. Les ones i llurs exquitxos són l’única aportació d’aigua de mar que reben les espècies que viuen emergides. Al nivell de la mar, o una mica per sota, l’hidrodinamisme significa energia mecànica que, quan ultrapassa un cert límit, pot arribar a arrencar les algues, actuant a favor de les espècies més robustes o dotades de millors òrgans de fixació. L’energia mecànica més moderada és favorable, ja que facilita la circulació de l’aigua i la renovació dels nutrients i del CO2 dissolt. Les aigües massa encalmades afavoreixen la sedimentació dels materials en suspensió, fet que pot interferir en l’activitat demoltes espècies, fins i tot eliminar-les. En general, hom parla de costes batudes (o mode batut) i de costes calmes o no batudes (o mode encalmat).
La il·luminació
La llum és un recurs vital per a les algues. En penetrar a través de l’aigua, la llum va essent progressivament absorbida, i s’atenua ràpidament en augmentar la profunditat, fins que deixa d’ésser efectiva per a la fotosíntesi de les plantes. El límit de distribució en profunditat de les algues coincideix amb aquella fondària on l’energia lumínica que arriba ja no els permet fotosintetitzar tot el que necessitarien per a viure (profunditat de compensació). On se situï aquesta profunditat, dependrà de la transparència de l’aigua, que varia segons la meteorologia, la mar de què es tracti, i l’època de l’any. Per exemple, mentre a la Costa Brava trobem algues fins als 70-80 m (excepcionalment 100 m) de profunditat, a les costes de Mallorca no és rar que pescadors que arrosseguen les arts per fons de més de 150 m en trobin restes. Pel damunt de la profunditat de compensació, l’adaptació de les espècies a les diferents intensitats de llum influeix clarament en l’establiment de les successives comunitats que trobaríem al llarg d’un hipotètic recorregut perpendicular a la costa, i cap al fons.
La importància d’aquest factor ressalta quan observem, a poca profunditat, els llocs poc illuminats, per estar protegits per algun sostre o trobar-se a l’entrada d’una cova. En aquests casos, hi trobem espècies pròpies d’una major profunditat. Per altra part, l’absorció de la llum per l’aigua no és igual per a les diferents longituds d’ona. Així, els colors freds (blau, verd) penetren a major profunditat que els càlids (groc, taronja, vermell), de forma que, a mida que la llum penetra, no solament es va atenuant, sinó que la seva composició espectral, és a dir, el seu color, va canviant. Per a alguns, les diverses algues no solament deuen estar adaptades a la intensitat de la llum que els arriba, sinó també a la seva qualitat, de forma que el conjunt dels pigments que conté cada espècie, i per tant, el color que presenten, hauria d’ésser complementari del color de la llum que reben (teoria de l’adaptació cromàtica). Fins a quin punt això és exacte ha estat motiu d’una polèmica que encara no sembla haver-se tancat, i en la qual no entrarem. Ara bé, en la pràctica, la intensitat de la llum és un factor més important que la qualitat.
La temperatura
La temperatura té, segons sembla, una importància decissiva com a factor de distribució geogràfica a gran escala. A les nostres costes, les variacions estacionals de la temperatura de l’aigua superficial i les diferències entre la temperatura de la superfície i la de les capes profundes (per sota de 30-40 m) durant l’estiu, que poden superar els 10°C, són fenòmens que sens dubte tenen influència sobre els cicles vitals i l’ecologia de les diferents espècies.
L’espai disponible
Al bentos marí, les espècies competeixen també pel recurs que podríem anomenar espai, que comprèn tant el substrat on s’han de fixar, com el volum on han de créixer i captar la llum. Una gran part de les morfologies i els tipus de creixement de les algues marines es poden interpretar com a diferents estratègies ecològiques de cara a la competència per a aquest recurs. Així, les algues de gran mida i creixement vertical creen microhàbitats ombrívols, on poden créixer espècies reptants o aplicades al substrat. També és molt freqüent que les espècies petites utilitzin l’alga gran com a suport, tot obtenint unes condicions de llum avantatjoses. Aquest epifitisme es pot produir novament sobre una alga ja epifítica, de forma que l’encadenament d’espècies pot portar a una gran complexitat i a un exhaustiu aprofitament de la llum i de l’espai.
Paper ecològic de les algues al bentos
Les algues bentòniques tenen, en primer lloc, un paper energètic, és a dir, capten energia i sintetitzen matèria orgànica, que representa una font potencial d’aliment per als animals marins. Ara bé, encara que alguns sistemes algals siguin dels més productius del planeta, els animals només consumeixen, de forma directa, una part relativament petita de les algues produïdes. Al bentos marí, els herbívors (garotes, cargols, crustacis, alguns peixos), són més aviat escassos, de forma que podem dir que les algues es mengen poc directament, potser per la dificultat de digerir molts dels seus components (polisacàrids complexos). Això no vol dir que la matèria sintetitzada per elles es perdi, ja que és aprofitada de forma indirecta per la via dels detritívors. En efecte, quan la fronda es desprèn, sofreix una sèrie de modificacions, primer físiques (disgregació), després químiques. Hi intervenen processos, tant abiòtics com biològics (acció dels bacteris i d’altres organismes), fins que, al final, el material algal entra a la reserva ("pool") de matèria orgànica, dissolta o en forma de partícules, i pot ser així aprofitat per molts animals bentònics.
Les algues exerceixen també un paper que en podríem dir estructural, modificant les característiques físiques de l’espai (ombra, suport per a altres algues, protecció davant l’hidrodinamisme). Les algues calcificades modifiquen el substrat, tot creant cavitats on poden refugiar-se diversos animals. En general, l’existència de coberta vegetal representa una diversificació d’hàbitats i de condicions ecològiques que, aprofitada tant per vegetals com per animals, permet constituir una comunitat molt complexa.
Zonació de les comunitats bentòniques
La zona favorable per a la colonització algal és compresa entre uns decímetres per sobre del nivell de la mar fins a uns 100 m (o més) de profunditat. Si comparem les condicions (temperatura, il·luminació, agitació de l’aigua, etc.) de cadascun d’aquests límits, veurem que són radicalment distintes. Els factors que acabem d’esmentar, tenen una variació vertical (en profunditat) molt acusada. Per això, la substitució d’unes espècies per unes altres (i, també, d’unes comunitats per unes altres) quan seguim un tall perpendicular a la costa, és molt neta, i determina un paisatge en bandes horitzontals, cada una d’elles corresponent a una comunitat. L’amplada de cada una d’elles depèn de la rapidesa amb què canvien els factors ambientals sobre l’eix vertical. La variació d’aquests factors és, precisament, molt més brusca als primers metres superiors, de forma que és aquí on les bandes són més estretes i més ben definides, mentre que, en augmentar la profunditat, llur amplitud augmenta i les fronteres es fan més laxes. Aquesta successió en zones horitzontals rep el nom de zonació.

Zonació en un indret molt batut i inclinat de la Costa Brava. De dalt a baix, s’observen, en primer lloc (estatge supralitoral), les taques negres del liquen Verrucaria symbalana i immediatament després les closques blanques del cirripede Chthamalus stellatus i altres congèneres (glans de mar), els quals formen un poblament dens que marca l’origen de l’estatge mediolitoral, constituït per l’horitzó fosc de l’alga Rissoella verruculosa i la cornisa organogènica, blanquinosa, de Lithophyllum tortuosum. Aquesta cornisa permet l’establiment, ja a l’estatge infralitoral, d’una comunitat esciòfila de petits rodòfits (Lomentaria, Schottera, etc.), molt característica: fins a 5 m de profunditat es fa una comunitat d’algues fotòfiles dominada per Asparagopsis armata, fins que una mica més acall Dictyota dichotoma, de color bruno-olivaci, inicia la transició vers la comunitat del cloròfit Codium vermilara, àmpliament estesa entre els 10 i els 15 m, mentre que a les parets verticals i poc il·luminades que segueixen s’instaura una altra comunitat de cloròfits (Halimeda tuna / Udotea petiolata/ Ja a l’estatge circalitoral, el feòfit Cystoseira zosteroides forma poblaments esclarissats sobre substrats horitzontals o quasi, fins entorn dels 20 m; a la base de la roca litoral, entre els 20 i els 40 m, s’estableix el complex coral·ligen, caracteritzat per la presència d’algues coral·ligenes incrustants, en el qual abunden diverses espècies d’animals bentònics (esponges, briozous, antozous, etc.), com la vistentgorgónia Paramuricea clavata. En llocs sorrencs, tanmateix, s’instal·la la comunitat del grapissar ("maërl"), constituïda per rodòfits calcificats de vida lliure.
Mikel Zabala / Jordi Corbera
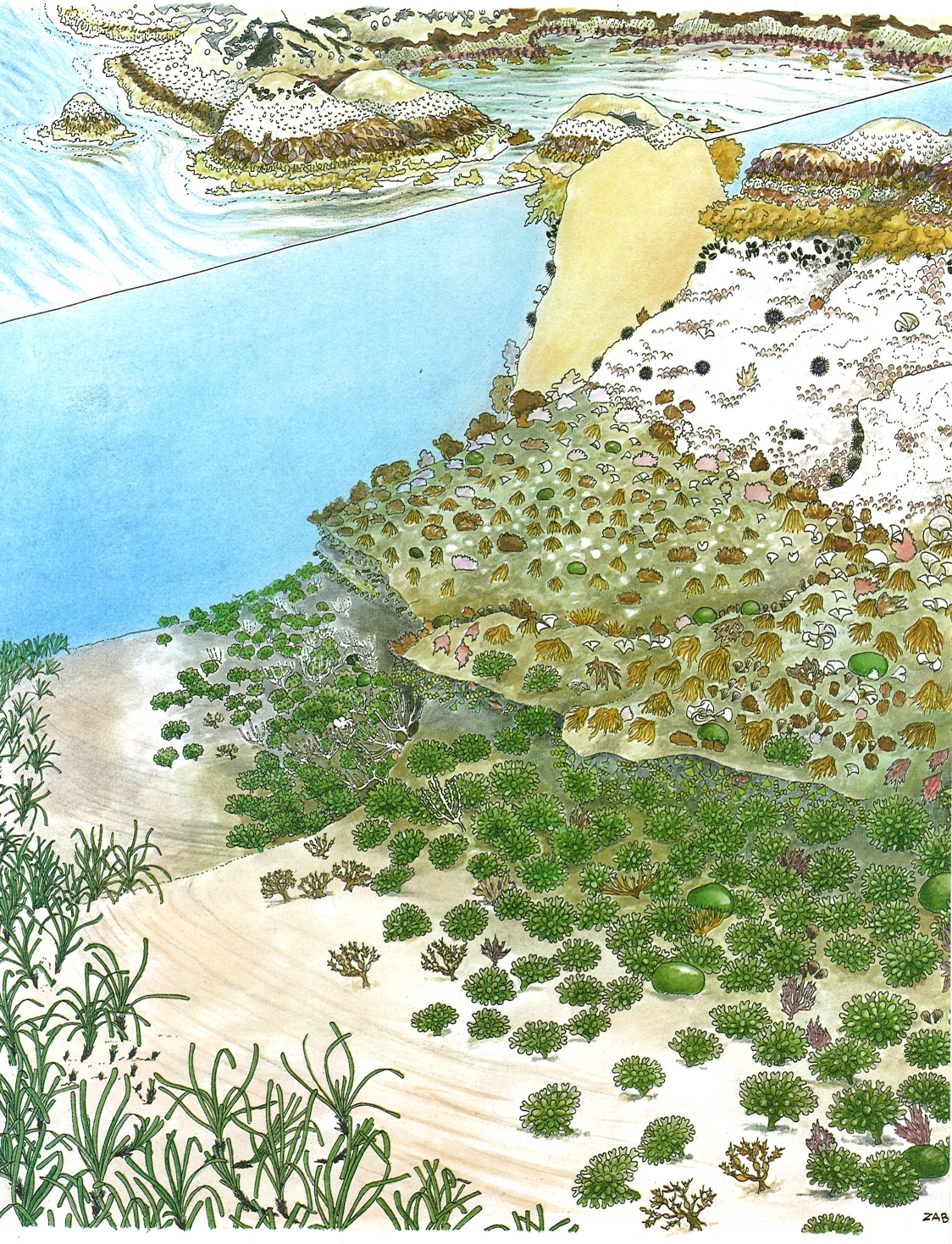
Zonació en un indret mitjanament batut i poc inclinat a la Costa Brava. De dalt a baix, i a la banda dreta, s’observa, en primer lloc, la comunitat liquènica de Verrucaria symbalana (taques negres) que constitueix l’estatge supralitoral, immediatament per sota de la qual es disposen, en diverses bandes estretes, les comunitats de l’estatge mediolitoral (fins a penes 0,5 m de profunditat): les closques blanques del cirrípede Chthamalus stellatus i altres congèneres (glans de mar), el feófit Mesospora macrocarpa, l’horitzó de Porphyra i del feòfit incrustant Ralfsia verrucosa, l’horitzó bru i estret de Rissoella verruculosa, l’horitzó de Lithophyllum tortuosum, que no arriba a formar tenassa ("trottoir"), intercalat amb els tal·lus de color verd oliva de Nemoderma tingitanum, i la banda vermellosa de Corallina elongata; al centre de l’esquema és representada una cubeta d’aigües encalmades, on la zonació és prou diferent. L’estatge infralitoral, que arriba fins a uns 5 m de profunditat, s’inicia amb una banda bruna de Cystoseira mediterranea, continua amb una faixa ampla de Lithophyllum incrustans i garotes, la qual és seguida per una comunitat molt diversificada d’algues fotòfiles, i acaba amb una ampla zona dominada pel cloròfit Codium vermilara. Després, el substrat sedimentan, pla, acull les praderies submarines de la fanerògama Posidonia oceanica.
Mikel Zabala / Jordi Corbera
El concepte implicat en aquest nom ha estat, però, molt discutit a nivell teòric, ja que sembla presuposar l’existència d’unitats discontínues, quan seria potser preferible parlar d’un continu on la dominància d’algunes espècies en caracteritzaria l’aspecte, en funció de la profunditat. En tot cas, les transicions són relativament sobtades, perquè resulten de la interacció dels efectes de dos gradients oposats.
En moltes mars altres que la Mediterrània, l’existència de marees sensibles provoca l’aparició d’una zona àmplia, rica en algues, alternativament emergida i submergida, que ha estat molt estudiada pels autors anglosaxons i francesos. La terminologia proposada pels primers no és, òbviament, aplicable a les costes mediterrànies. Per això, seguim la classificació proposada pels autors francesos, principalment de l’Escola de Marsella, encara que no ens proposem oferir aquí una exposició fitosociològica sistemàtica, sinó esbossar un breu esquema del nostre paisatge litoral i submarí. Per raons pràctiques, delimitarem i definirem les zones en base a criteris biològics, és a dir, atenent als límits de distribució vertical de les espècies característiques. No obstant això, el concepte de zona es basa més en les característiques ambientals, de les quals també farem esment, fins on podrem.
La primera zona que podem considerar com a pertanyent al bentos marí és la zona supralitoral, o zona dels esquitxos, on l’aigua arriba solament en forma d’esquitxos projectats per les ones, quan la mar està més o menys agitada. La seva amplitud vertical varia d’un lloc a un altre, i depèn de la freqüència de dies de mar prou agitada, de la força mitjana de les ones, i de la inclinació de la superficie rocosa. Pot anar des d’alguns centímetres fins a pocs metres.
Immediatament per sota, apareix la zona mediolitoral o de rompent, banyada pel vaivé de les ones. Es tracta d’una zona no submergida però que, excepte de forma esporàdica (calmes duradores o minves) té l’aportació hídrica assegurada. L’amplitud d’aquesta zona també varia, per les mateixes raons que a la zona anterior. A les nostres costes, és de l’ordre d’uns decímetres.
L’amplitud vertical dels estatges següents és ja considerablement més gran, i la seva característica principal és la d’estar permanentment submergits. La zona infralitoral, ben illuminada, va des de la superfície de la mar, fins a la profunditat on solament arriba el 5-10% de la llum de la superfície. Això equival, segons el lloc i l’època de l’any, entre els 15 i els 30 m de profunditat.
Finalment, la zona circalitoral, o zona poc illuminada, baixa fins a les màximes profunditats compatibles amb la vida vegetal que, en els llocs d’aigües més transparents, pot atènyer els 200 m (a les nostres costes, com hem vist, se situa entre els 70 i el 150 m).
Per sota d’aquesta darrera zona, només es troben comunitats exclusivament formades per animals i organismes descomponedors que, en una bona part, es nodreixen dels excessos de producció de les zones superiors.
La zona supralitoral

Comunitat d’una cubeta supralitoral, amb Enteromorpha intestinalis.
Enric Ballesteros
La vida és molt difícil en aquesta zona, sobretot a causa de la manca d’aigua en absència de condicions de mar agitada (que pot durar dies, i fins mesos en determinades èpoques de l’any), de les grans oscil·lacions termomètriques, de les fortes variacions de salinitat, quan plou o quan l’aigua queda embassada, etc. És per això que només uns quants organismes han pogut desenvolupar les adaptacions necessàries per a sobreviure en condicions com aquestes. Possiblement, el problema més gran de la supervivència en aquest nivell és la pèrdua d’aigua per evaporació. Això fa que els organismes fotosintetitzadors més abundants siguin els líquens (Verrucaria symbalana) i determinats cianòfits que sols creixen dins la roca (endolítics) o a les esquerdes humides. Les algues superiors no troben, a la zona supralitoral, condicions aptes per a llur supervivència, si exceptuem les cubetes (clots a la roca on s’acumula aigua durant un temps) i les coves.
Encara que les condicions físico-químiques de les cubetes supralitorals siguin molt canviants, la presència, més o menys duradora, d’una certa massa d’aigua, fa possible que hi visquin una sèrie d’algues, eurioiques, però molt típiques d’aquests hàbitats. Hi abunden diverses espècies de cloròfits unicel·lulars i flagel·lats que, si la quantitat de nutrients és elevada (per efecte d’ocells marins, residus duts per la mar i altres aportacions de matèria orgànica), proliferen fins a tenyir l’aigua de color verd. Brachiomonas submarina, Tetraselmis fontiana i diverses espècies dels gèneres Chlamydomonas i Pyramimonas, en són els més corrents. Els cianòfits, les diatomees i algunes dinofícies i crisofícies completen aquesta llista. En algunes cubetes amb aigua amb poca sal, apareixen algunes algues pluricel·lulars dels gèneres Enteromorpha i Cladophora.
Les coves i esquerdes una mica grans ofereixen també una certa constància de condicions d’humitat i de temperatura, les quals permeten la vida de diverses algues. En aquest cas, el factor limitant és la llum. Per aixó només hi trobem espècies esciòfiles, més pròpies de l’estatge mediolitoral o, àdhuc, de l’infralitoral. Hildenbrandia rubra, una alga incrustant de color púrpura i tacte relliscós, sol ser molt abundant. L’acompanyen algunes coral·linàcies, també incrustants (sobretot Phymatolithon lenormandi), que comuniquen a les parets i al sostre un suau matís rosat.
Pel que fa a la producció primària, cal dir que el creixement dels líquens és molt lent, i que els cianòfits contribueixen poc a la producció total, que és molt baixa. En canvi, quan la concentració del fitoplàncton és elevada, hi podem trobar valors de producció considerables, equiparables als de les llacunes eutròfiques i als conreus d’algues.
La zona mediolitoral

Cornisa, al nivell mediolitoral, de Lithophyllum tortuosum, a les illes Medes (Baix Empordà).
Enric Ballesteros / Javier Romero
La disponibilitat hídrica d’aquesta zona és més constant, i això permet el creixement d’una sèrie d’espècies adaptades a viure fora de l’aigua, però regularment mullades pels esquitxos de les onades. És en aquest tros on la disposició en bandes de la vegetació es fa més aparent. Els diferents horitzons, definits per l’alga o les algues dominants, es succeeixen en un espai molt reduït, sovint inferior a 1 m. A la Costa Brava és on es presenten la majoria d’horitzons de què parlarem, principalment a les zones silíciques. A la resta dels Països Catalans, la zonació no és gens aparent (cas del País Valencià i les Balears) i, d’ésser-ho, sol limitar-se a les comunitats portuàries, on diverses espècies d’afinitats tionitròfiles (d’aigües riques en nutrients) formen horitzons clars durant la primavera i l’hivern. Un bon exemple de la zonació mediolitoral que podem trobar a qualsevol punt de la costa gironina, sempre en llocs batuts, sobre roques silíciques, principalment granit (sector Blanes-cap de Begur) permet d’il·lustrar les característiques d’aquest estatge bentònic.

Comunitats nitrófiles d’espècies oportunistes, amb Enteromorpha compressa (verd clar) i Bangia atropurpurea (roig fosc), sobre roques de platges sovintejades. La zonació és nítida en la foto inferior.
Enric Ballesteros
Just per sota de la zona supralitoral, que vindria definida per les taques negres del liquen Verrucaria symbalana, en llocs mitjanament inclinats, apareix un horitzó molt clar, dominat per dues espècies de cirrípedes (Chthamalus), envoltades per un tegument format per escuts calcificats. Creixen densament serrats, i entre ells proliferen nombroses colònies de cianòfits (Bachytrichia quojii, Rivularia atra), que presenten l’aspecte de petits hemisferis negres, durs i relliscosos. Als indrets orientats al N, el feòfít incrustant Mesospora macrocarpa dona al conjunt un color brunenc ferruginós molt característic. Sobre les closques dels Chthamalus i altres superfícies calcàries, podem veure petits grànuls o puntets negres, que corresponen als peritecis del liquen Arthopyrenia littoralis.
En un nivell inferior, i durant l’hivern, apareixen uns tels rosats o daurats, laminars i amb plecs, que corresponen a diverses espècies de Porphyra (P. leucosticta, P. umbilicalis, P. linearis), generalment acompanyades per Polysiphonia sertularioides i els feòfits Feldmannia irregularis i Scytosiphon lomentaria. Més avall, els tal·lus arrissats, brunencs o rossos del rodòfit Rissoella verruculosa, formen un horitzó vistós i molt ben delimitat, d’alguns centímetres d’amplada. L’acompanyen una mena de fideus, més o menys gruixuts, de color vermell fosc i de consistència gelatinosa tenaç, que corresponen al rodòfit Nemalion helminthoides, i les crostes negres i lluents del feòfít Ralfsia verrucosa.

Detall de la banda mediolitoral, de trànsit en l’estatge mediolitoral típic i l’infralitoral, dominada per Ceramium ciliatum.
Enric Ballesteros
La part inferior de la zona mediolitoral és ja molt rica en algues, que hi apareixen amb una diversitat considerable. Sota la banda de Rissoella, hi podem trobar dues comunitats: la de Nemoderma tingitanum, un feòfít incrustant, de color verd oliva o verd ocraci, que es presenta als llocs menys batuts, o bé la de Lithophyllum tortuosum, pròpia dels llocs més batuts. Si l’agitació de l’aigua és suficient i el substrat és inclinat, aquesta coral·linàcia, d’aspecte cerebriforme, va creixent i acumulant carbonat càlcic, fins a arribar a establir autèntiques cornises, de fins a més d’1 m de gruix. Aquesta cornisa o "trottoir" és a la zona de les illes Medes i el Montgrí, a determinats punts del cap de Begur i del cap Norfeu i a la costa septentrional menorquina (on és anomenada tenassa), on assoleix un desenvolupament més gran; en aquests casos, observem un empobriment dels horitzons superiors, perquè, a causa de llur forma, les tenasses de Lithophyllum tenen tendència a apartar l’aigua que incidiria sobre les zones més altes del litoral. Finalment, i ja en contacte amb les comunitats infralitorals dominades per Cystoseira mediterranea, apareix una comunitat de transició, d’aspecte variable, on determinades espècies, com Ceramium ciliatum, Gelidium pusillum s.l., Corallina elongata, Hypnea musciformis i diverses Enteromorpha i Cladophora poden formar poblacions més o menys efímeres o més o menys constants.
Com passa amb tantes altres comunitats naturals, aquest exemple de zonació, que hauria d’ésser normal, es va tornant una excepció, en una bona part per l’acció de l’home. En efecte, els abocaments d’aigües residuals, la construcció de ports, l’alteració, en general, de les condicions físiques de la línia de costa, provoquen la desaparició dels horitzons esmentats i la seva substitució per altres comunitats, menys riques i vistoses. Unes poques espècies eurioiques en resulten afavorides. Lliures de competència, creixen durant l’hivern, afavorides per una irradiació solar més baixa i per una disponibilitat d’aigua més constant. A la part superior apareix llavors un horitzó fosc de Bangia atropurpurea o de Lyngbya confervoides i, a sota, una comunitat de Scytosiphon lomentaria, Porphyra leucosticta, diverses Cladophora, Blidingia minima i altres espècies. Pel quefa a l’aspecte, són revestiments de poc gruix, on dominen els vermells foscos i, sobretot, els verds més o menys llampants. El pas cap a la zona infralitoral, que aquí no és rica en Cystoseira, sinó en espècies d’ambients inestables, com Corallina elongata, Ulva rigida, Gelidium pusillum, és marcat per la presència de poblacions denses d’Enteromorpha compressa.
Si bé, en aquesta zona, la vegetació de les coves i esquerdes fosques és semblant a la vista a la zona supralitoral, les cubetes mediolitorals són ben diferents. Les condicions ambientals, més constants, fan que les poblacions fitoplanctòniques no hi estiguin tan ben representades i, en canvi, hi creixin espècies bentòniques, de caràcter oportunista. Entre altres, hi són freqüents les clorofícies Enteromorpha compressa, Chaetomorpha aerea, Cladophora vagabunda, C. albida i força més espècies d’aquest gènere, els feòfits Ectocarpus siliculosus varietat confervoides i Scytosiphon lomentaria, i alguns rodòfits dels gèneres Ceramium i Polysiphonia.
La dinàmica de les comunitats mediolitorals es caracteritza per un creixement ràpid de la majoria de les espècies en els mesos d’hivern i del principi de la primavera, quan les condicions de llum, nutrients i agitadó de la mar són òptimes. Al final de la primavera, la majoria de les espècies entren en un període d’envelliment i van desapareixent a mida que entra l’estiu, quan la lluminositat i la temperatura esdevenen excessives, alhora que l’encalmament progressiu de la mar els limita l’aportació d’aigua. Hi ha, però, especies d’algues, com Mesospora macrocarpa, Lithophyllum tortuosum i Ralfsia verrucosa, que es presenten durant tot l’any, encara que durant l’estiu es mantenen en condicions precàries i es mostren seques i poc acolorides.
La zona infralitoral

Poblaments característics, emergits, de Cystoseira: C. mediterranea (a dalt), en un lloc batut del cap de Tossa (Selva), i C. compressa (a baix), en un lloc arrecerat del cap de Creus (Alt Empordà).
Enric Ballesteros i Javier Romero
Les condicions ecològiques de la zona infralitoral son lés més favorables per a la vida dels vegetals marins. No existeix el perill d’emersions contínues o periòdiques, que era propi dels nivells superiors, i la immersió permanent assegura una relativa constància de les condicions del medi. La llum, encara poc atenuada pel gruix de la massa d’aigua, és suficient per a una fotosíntesi activa, i el moviment de les ones assegura una adequada renovació dels nutrients.

Visió detallada de la comunitat de Cystoseria mediterranea. L’espècie principal modifica les condicions ambientals i crea diversos microhàbitats que són colonitzats per diferents organismes animals i vegetals: els epífits dels ràmuls, algues menudes i marcadament fotòfiles, entre les guals la incrustant Dermatolithon pustulatum, els filaments brunencs de Feldmamia caespitula i els filaments vermells de Ceramium; els epífits dels troncs, una mica més grans, que viuen a l’ombra, com la coral·linàcia ramificada Jania rubens, els rodòfits filamentosos Callithamnion granulatum i Polysiphonia mottei, o els animals sèssils, com l’hidrari Aglaophenia kirchenpaueri, o mòbils, com el cranc Pisa corallina; les algues de l’estat basal, erectes les unes (com Corallina elongata, de tal·lus articulats, Laurencia pinnatifida, de frondes purpuràcies, o Valonia utricularis, en forma de petites vesícules verdes i lluents), incrustants les altres (com Lithophyllum incrustans, de color rosa clar, o Hildebrandia canariensis, de color vermell fosc); i, finalment diversos animals fixos al substrat, com el musclo (Mytilus galloprovinciale), o les glans de mar (Balanus perforatus).
Mikel Zabala / Jordi Corbera
Pel que fa a la producció (síntesi de matèria orgànica per unitat de temps), tots aquests factors es combinen de manera òptima en la banda compresa entre 0 i 0,5 m de profunditat, especialment en les zones ben il·luminades i exposades a les ones. És aquí on s’estableixen les comunitats de Cystoseria de llocs batuts (Cystoseira mediterranea a les costes catalanes, C. stricta a altres indrets de la Mediterrània), de desenvolupament i complexitat estructural notables. Els tal·lus de Cystoseira, bruns i esponerosos, sovint amb aspecte de cua de guineu, poden atènyer fins a 25-30 cm d’alçada, i constitueixen un estrat "arbori" sobre el qual es fixen nombrosos epífits. Al seu recer, protegits per ells de la llum i de la força de l’aigua, hi pot haver un estrat "arbustiu" d’algues erectes i ramificades, com Corallina, Jania i altres.
Algunes espècies cespitoses, de mida petita, com Valonia utricularis i Gastroclonium clavatum, formen part de l’estrat "herbaci", per sota del qual trobem encara un estrat incrustant (sense cap d’anàleg als boscos), constituït per algues calcàries que creixen aplicades al substrat, com Lithophyllum incrustans, Hildenbrandia canariensis, etc. Aquesta estratificació i diversificació d’hàbitats té com a conseqüència una gran diversitat en espècies, que repercuteix també en la fauna acompanyant (poliquets, petits crustacis, mol·luscs, etc.).

Poblament tipic, submergit, a 1 m de profunditat, de Cystoseria caespitosa, prop de Tossa de Mar (Selva).
Enric Ballesteros

Comunitat d’algues esciòfiles, en un lloc batut, amb predominança de Valonia utricularis.
Javier Romero
Quan l’agitació de l’aigua és menor, a la mateixa profunditat apareixen unes altres espècies de Cystoseira, com C. caespitosa, C. crinita, C. baleárica, C. compressa i C. ercegovicii. Les comunitats dominades per aquestes espècies, encara que mantinguin una complexitat important, presenten una producció inferior a la de les anteriors, a causa del moviment menor de l’aigua, que dificulta la renovació dels nutrients. Totes les comunitats de Cystoseira tenen un ritme estacional ben marcat, definit per l’espècie dominant que, al final de l’estiu, i fins al principi de la primavera, resta reduïda als troncs i les branques principals. Durant aquest període, en quedar desprotegits, els estrats inferiors poden resultar seriosament afectats i reduïts, i en el lloc de les espècies que els formaven, en creixen d’altres, en general de caràcter oportunista (Ulva, etc.), lligades a l’augment de nutrients en superfície.
Més avall de les poblacions de Cystoseira, trobem unes comunitats d’algues fotòfiles, que continuen estant caracteritzades per l’exuberància del component vegetal. En llocs batuts apareix Asparagopsis armata (més sovint en la seva fase tetrasporofítica, dita Falkenbergia rufolanosa, en forma de floc de cotó rosat), Laurencia obtusa, Corallina elongata, etc. El conjunt presenta un aspecte frondós i de colors diversos, gronxat o, fins i tot, a vegades agitat violentament per les ones.
En llocs encalmats, es fa una comunitat de colors menys vistosos, on dominen els tons bruns, produïts per la presència de nombrosos feòfits (Halopteris scoparia, que forma petites escombres fosques, amb les puntetes daurades, Cladostephus hirsutus, Dilophus fasciola), juntament amb moltes espècies pantropicals, que podríem dir que hi afegeixen la nota exòtica: les "margaridetes" d’ Acetabularia acetabulum, les paperines o embuts de Padina pavonica, les petites porres verdoses de Dasycladus vermicularis (sobretot abundant a les Balears), etc. En aquests llocs més tranquils, el dipòsit de materials transportats per l’aigua és important, de forma que el sediment juga també un cert paper en l’establiment i la persistència de la comunitat, ja que moltes espècies no poden viure en condicions com aquestes.
L’existència de pertorbacions locals altera aquestes comunitats, que són substituïdes per unes altres, en general de complexitat i diversitat inferiors. Si les pertorbacions són d’origen humà (contaminació), s’arriba a l’establiment de fàcies nitròfiles, amb abundor de cloròfits (Ulva, Enteromorpha) i de certs rodòfits (Gelidium, Corallina). Altres pertorbacions són d’origen natural i potser la més espectacular de totes és l’atribuïble a la pressió excessiva dels herbívors, principalment garotes, pagellides i mol·luscs diversos; la presència en gran nombre d’aquests herbívors pot arribar a eliminar la co berta d’algues erectes. Queda llavors solament una capa de Lithophyllum incrustans adherida a la roca, que li dona una tonalitat rosada, sobre la qual destaquen les taques negres dels eriçons o garotes. Es un "paisatge" en certa manera "desolat", si hom el compara amb les comunitats algals descrites més amunt.

Comunitats d’espècies fotòfiles en un indret arrecerat (a baix), amb Padina pavonia, Halopteris scoparia, Cladostephus hirsutus, etc., i sobre una paret (a dalt), amb Dictyota dichotoma i Falkenbergia rufolanosa, a les illes Medes (Baix Empordà).
Javier Romero i Mikel Zabala / Josep M. Gili
Factors com l’orientació vers el N, la inclinació (vertical o bé balmada) o el microrelleu (cornises i ressalts) poden fer que, al mateix estatge infralitoral, les condicions d’il·luminació es trobin, en alguns llocs, molt atenuades. Això fa possible l’ascens de comunitats circalitorals, si bé en certes condicions hi poden haver comunitats infralitorals específiques. Quan la illuminació és baixa, però l’hidrodinamisme és accentuat, condicions típiques de la part inferior de la cornisa de Lithophyllum tortuosum, apareix una comunitat molt ben caracteritzada. A les costes del Principat, dominen rodòfits de mida petita, com Schottera nicaensis, Lomentaria articulata, Plocamium cartilagineum, etc., que constitueixen un conjunt amb aspecte de fina gespa vermellosa, bastant curiós. Més cap al S, i també a Mallorca, hi entren espècies d’afinitats termòfiles, com Botryocladia botryoides, Polyphysa parvula, etc. L’impacte de l’aigua a què aquestes comunitats es veuen sotmeses comporta la destrucció de petites parcel·les de vegetació, que queden distribuïdes aquí i allà, una mica a l’atzar. Per això, el conjunt queda format per un mosaic de peces, cada una de les quals correspon a una fase diferent de la regeneració de la comunitat.
En zones menys agitades, i generalment més profundes, apareixen altres comunitats esciòfiles (pròpies de llocs poc il·luminats), que hom denomina genèricament el pre-coral·ligen. De fet, corresponen a la transició entre els estatges infralitoral i circalitoral, i en parlarem al proper apartat.
Si volem completar aquesta breu panoràmica de l’estatge infralitoral, no podem pas deixar de fer referència a les comunitats que colonitzen el substrat sedimentari, és a dir, els fons sorrencs o fangosos. Gairebé sempre, l’espècie dominant en aquestes condicions, poc adequades per a la fixació de les algues, és una fanerògama, els rizomes i les arrels de la qual (la mata), li permeten fixar-se sobre el substrat tou, formant prades submarines més o menys extenses, dites alguers , que sovint destaquen com a clapes fosques, sobre els fons arenosos clars. El cas més freqüent i espectacular és el de Posidonia oceanica, dita "alga", encara que es tracti d’una fanerògama, una espècie endèmica de la Mediterrània.
L’aparell vegetatiu de Posidonia oceanica crea unes condicions favorables per a l’establiment de nombroses espècies d’algues, tant oferint-los un substrat on fixar-se, com creant zones protegides de la llum o de la força de l’aigua. Com a resultat, tenim un poblament molt complex que, pel que fa a les algues, es pot dividir en dos estrats molt diferents. Sobre els rizomes, poden créixer des d’espècies fotòfiles (Acetabularia, Jania, Sphacelaria) fins a fortament esciòfiles (Rhodymenia, Peyssonnelia), segons sigui la densitat i, per tant, el recobriment degut a la fanerògama; les comunitats que hi trobem s’assemblen a d’altres d’infra o circalitorals, més o menys empobrides. En canvi, sobre les fulles cintiformes apareixen multitud de petites algues, molt més especialitzades, ja que han d’estar adaptades al particular ritme de creixement i de caiguda del substrat, a la seva natura orgànica i al constant moviment de balanceig al qual estan sotmeses. Han d’ésser, doncs, espècies de creixement ràpid, de cicle vital curt (que s’ha de completar abans de la caiguda de la fulla) i de mida petita. A la primavera, la comunitat de les fulles és dominada per petits feòfits (Castagnea cylindrica, C. irregularis, Myrionema magnusii, Giraudia sphacelarioides, Myriactula gracilis, etc.), mentre que, al final de l’estiu, el verd intens de les fulles va essent en bona part substituït per un color blanquinós, produït per les petites rodofícies incrustants que han anat cobrint, de manera gairebé contínua, la part més vella de les fulles o les fulles senceres: Fosliella lejolisii i altres espècies del gènere, Dermatolithon, etc. Si examinem a la lupa una fulla adulta de Posidonia oceanica, ens trobarem amb un fascinant microcosmos d’espècies animals i vegetals. A més, com que les fulles van creixent per la base i envellint i erosionant-se per la punta, hi tenim escrita, d’un extrem a l’altre, tota la història de la successió d’aquesta curiosa comunitat.
Les altres fanerògames marines presents a les nostres costes (Cymodocea, Zostera), viuen sobre sediments més fins, en badies i llocs que reben aportacions d’aigua dolça i donen suport a comunitats d’algues semblants, però més pobres. Les úniques algues veritables que aconse gueixen colonitzar alguns fons de sediment són les coletes, el cloròfit Caulerpa prolifera, que unes vegades conviu amb les dues fanerògames anteriors, i d’altres forma densos prats monospecífics. Les frondes de Caulerpa estan gairebé del tot desproveïdes d’epífits, probablement a causa d’una substància tòxica segregada per aquesta alga termòfila, d’afinitats pantropicals.
La zona circalitoral

Comunitat d’espècies esciòfiles de poca fondària, a la península del cap de Creus (Alt Empordà). S’hi observen — ultra uns ràmuls d’Asparagopsis armata —, Jania corniculata, Valonia utricularis, Myriogramme minuta, Cladophora pellucida, etc.
Javier Romero
El límit de la zona infralitoral ve marcat per l’atenuació progressiva de la llum i l’absorció selectiva de determinades longituds d’ona en augmentar la fondària, fenòmens que provoquen un canvi en la vegetació bentònica. Les espècies esciòfiles (que resisteixen la baixa il·luminació), substitueixen progressivament les fotòfiles. Per establir exactament on se situa el pas de la zona infralitoral a la circalitoral, hom ha pretès de donar una sèrie d’espècies indicadores, pròpies d’una zona o altra o que rarament surtin d’ella. Amb freqüència hom ha triat el límit inferior de Palguer de Posidonia oceanica com a frontera entre ambdós estatges. D’altres opinen que la substitució del feòfit Halopteris scoparia pel seu congènere H. filicina és útil per a fer aquesta diferenciació. En realitat, es tracta més aviat d’un pas gradual, marcat per una sèrie de comunitats de trànsit (les pre-coral·lígenes), molt riques en espècies, i que se situen en fondàries variables, segons la transparència de les aigües. Com a exemple, diguem que el pre-coral·ligen se situa entre 10 i 20 m a la Costa Brava, mentre que, a les Balears, ho fa a una fondària molt més gran, de 30 a 40 m. Cal remarcar que aquests límits batimètrics no són gens rígids. Recordem que comunitats com aquesta pugen a menys profunditat, a l’empara de les coves i de les superfícies balmades.

Comunitat d’espècies esciòfiles de fondària, situada a uns 15 m de profunditat, a les illes Medes (Baix Empordà), amb dominància c/’Udotea petiolata.
Mikel Zabala / Josep M. Gili
Una sèrie d’espècies són abundants quantitativament en aquestes comunitats de precoral·ligen. A la Costa Brava, és freqüent la comunitat de Codium vermilara, que comença a aparèixer en els fons lleugerament inclinats, vers 6-8 m de fondària, per a baixar fins als 25 m. Una altra comunitat típica, la de Cystoseira spinosa, també present a la costa catalana, és potser més abundant a les Illes i al País Valencià; ambdues comunitats tenen un nucli d’espècies comunes. Val a dir que la majoria de fideus (nom que els pescadors dónen a Codium vermilara) que s’enganxen a les xarxes i que tant molesten a la gent de mar provenen d’individus que han crescut en aquesta zona i que, arrencats pels corrents i els temporals, es dipositen sobre els fons de pesca.

Comunitat coral·lígena, amb dominància d’espècies animals filtradores i presència d’algunes algues (Mesophyllum lichenoides, Lithophyllum expansum, Udotea petiolata), a 25 m de profunditat, a les illes Medes (Baix Empordà).
Mikel Zabala / Josep M. Gili
Per sota de les comunitats de pre-coral·ligen apareixen, sobre substrat dur, les típiques comunitats circalitorals, caracteritzades per una sèrie d’espècies que tenen llur màxim desenvolupament en aquesta zona. Generalment, hi abunden les coral·linàcies incrustants (Mesophyllum lichenoides, Lithophyllum expansum), que van acumulant carbonat càlcic fins a formar un sòcol coral·ligen, que és una roca biogènica constituïda pels tal·lus morts de les algues incrustants i pels nombrosos organismes sèssils (esponges, briozous, madreporaris, mol·luscs, etc.), que hi creixen, omplint els forats deixats per les algues. Tenen, doncs, una estructura molt complicada, ja que el creixement del conjunt no és uniforme i hi abunden espais buits. Se’ls ha comparat amb trossos de formatge de "gruyère", les cavitats dels quals estarien ocupades per un conjunt d’éssers vius, de gran diversitat i bellesa, que crida poderosament l’atenció dels escafandristes. Vista amb llum natural, l’aspecte és un xic tenebrós, ja que, a més de la baixa intensitat lumínica, cal afegir-hi que els colors són blavissos i freds. Però n’hi ha prou amb encendre una llanterna o un focus perquè apareguin els colors càlids i el coral·ligen esdevingui un fantàstic i barroc concert de colors.

Comunitat de fondària, sobre grapissar ("maërl"), a 45 m de profunditat, prop de Tossa de Mar (Baix Empordà), amb Lithothamnium fruticulosum, Phymatolithon calcareum, Lithophyllum expansum i altres coral·linàcies.
Javier Romero
Les comunitats coral·lígenes són variades, i és probable que se’n puguin tipificar diverses. La comunitat de Cystoseira zosteroides, estructuralment semblant a les pre-coral·lígenes, és dominada encara per espècies amb formes arborescents. És pròpia de zones amb corrents intensos. Entre 15 i 35 m de fondària, és molt freqüent a la Costa Brava una comunitat dominada per Mesophyllum lichenoides i Halimeda tuna. Els grans blocs de coral·ligen originats per acumulació de Lithophyllum expansum es localitzen per sota dels 35 m. Les dades que tenim sobre altres punts de les costes dels Països Catalans són escasses i, actualment (1985), no permeten oferir una descripció detallada. No obstant això, és possible un paral·lelisme amb les comunitats estudiades a la Costa Brava, que es trobaran a una fondària més alta o més baixa, segons la terbolesa de l’aigua. Així, a les clares aigües de les Balears, no trobarem un coral·ligen fins per sota dels 40-50 m de fondària.
Sobre substrats tous, les comunitats d’algues de la zona circalitoral tenen una certa importància. L’estabilitat del medi és superior a la dels substrats tous infralitorals, i això permet el creixement d’una sèrie d’espècies. Una comunitat dominada per feòfits, els més aparents dels quals són Arthrocladia villosa i Sporochnus pedunculatus, de desenvolupament primaveral, tipifica els fons de sorra gruixuda i pedruscall de la major part de les nostres costes. A la Costa Brava, apareix per sota dels 15 m de fondària, i deixa d’ésser dominant cap els 35 m. A les Balears, mostra un desplaçament batimètric idèntic al de les altres comunitats circalitorals, i assoleix fondàries molt més importants, superiors fins i tot als 100 m.
Les comunitats dominades per coral·linàcies arbusculars lliures (dites grapissar, a Catalunya, i avellanor, a les Balears) amb Phymatolithon calcareum, Lithothamnium corallioides, L. fruticulosum, L. valens i altres, marquen el límit de la vegetació circalitoral (60-90 m a les costes peninsulars, 150 m a les Balears). Podem descriure l’aspecte general d’aquestes comunitats dient que sembla lunar, ja que la poca llum existent, l’elevada uniformitat del poblament, i el substrat pla, però irregular, a causa de l’encavallament dels tal·lus arbusculars de les coral·linàcies, donen al conjunt un aspecte irreal. L’estrat elevat pot ser escàs, i quedar reduït a petites espècies que creixen entre el grapissar, a la Costa Brava. Però a determinades zones, com ara a les Balears, trobem, formant extenses prades, diverses algues laminars, ben conegudes dels pescadors, com ara l’herba torta (Vidalia volubilis), l’herba cuquera (Phyllophora crispa) o l’herba col (Laminaria rodriguezii), l’única laminària que hi ha a les nostres mars.
La producció de les comunitats circalitorals és molt variable i, òbviament, depèn de la llum disponible. Hi tenen especial importància les espècies dominants a la comunitat, ja que és probable que es comportin de formes diferents. Així, les Cystoseira de profunditat renoven la totalitat de llurs branques un cop l’any. Les coral·linàcies creixen molt lentament, i els seus tal·lus es mantenen fotosintèticament actius durant un temps desconegut, però que segurament és de diversos anys. Algunes espècies de Sporochnus, Arthrocladia i Dictyota, produeixen un nou tal·lus cada any. Halimeda tuna, en canvi, va renovant els articles que componen el seu tal·lus durant tot l’any, si bé hom hi observa un màxim creixement al principi de l’estiu. La quantificació de la producció és, doncs, complicada i difícilment generalitzable a totes les comunitats circalitorals. Sens dubte, però, és inferior a la de les comunitats infralitorals i mostra un descens en sentit vertical. De forma global, cal també dir que la renovació de la biomassa és força més lenta que a nivells superiors.