Característiques del grup
Els rodòfits, o algues vermelles, comprenen més de 4000 espècies, agrupades en uns 675 gèneres. Les característiques principals que les defineixen com a grup fan referència, principalment, als pigments fotosintetitzadors, a les substàncies de reserva, a alguns aspectes citològics i als tipus de reproducció.
Pel que fa als pigments, hem de dir que, al costat de la clorofil·la a, no en tenen de b ni de c. En algunes espècies hi ha una discutida clorofil·la d, exclusiva d’aquest grup. Com a complement, tenen α i β-carotè, diverses xantofil·lès i dues ficobilines, ja conegudes de les cianofícies. la ficocianina (blava) i la ficoeritrina (vermella), que formen part (com a grup prostètic de ficobiliprotèids) d’uns grànuls discoïdals o hemisfèrics units a la superfície dels tilacoides, els ficobilisomes (recordem que les cianofícies també en presenten). Ben ordenats a l’interior dels ficobilisomes (la ficocianina a l’interior, la ficoeritrina a la perifèria) aquests pigments capten energia a partir de llum de longituds d’ona curta, i la traspassen a la clorofil·la a, que no seria capaç de captar-la. Normalment la ficoeritrina emmascara els altres pigments i, per aquesta causa, els rodòfits tenen el color vermell que justifica el seu nom. Amb tot això, la gamma de colors que presenten els rodòfits és molt més vasta, i va des de colors blavosos, daurats i brunencs, fins a gairebé el negre, tot passant, evidentment, pels diferents matisos del vermell.
Les substàncies de reserva són diverses: gotetes de lípids, floridòsid (un derivat de galactosa i glicerina), etc., però la més típica és el midó de florídies, un polisacàrid de molècula arrodonida, semblant a la del midó de les plantes superiors, però més ramificada, com el glucogen. Forma granets, que amb iode només es tornen groguencs o vermellosos, no blaus com els de midó, situats a la superfície dels plastidis o al citoplasma.
La paret cel·lular està diferenciada en una capa interna, de cel·lulosa, i una capa externa, amorfa, de natura pèctica, sovint rica en col·loides gelificats com l’agar (galactana) i la carrageenina (galactan-sulfat). A la família de les coral·linàcies (i en alguns representants d’altres famílies) la paret cel·lular està fortament incrustada de carbonats, sobretot carbonat càlcic, però també poden tenir carbonat magnèsic. Els plastidis (anomenats també cromatòfors o rodoplasts) poden tenir formes diverses: petits i discoïdals, estrellats, lobulats, cintiformes, etc. Els seus tilacoides no es disposen mai en grups. Estan més o menys separats i equidistants, igual que a les cianofícies. Una de les característiques més notables des del punt de vista citològic és l’existència, en moltes espècies, de sinapsis, o connexions entre cèl·lules contigües, que poden procedir d’una septació incompleta en el moment de la divisió cel·lular, o es poden formar posteriorment. Però encara no es veu pròu clar si es tracta de veritables connexions intercel·lulars, ni quin és el mecanisme de llur formació.
Però la característica més original dels rodòfits és la seva reproducció, amb cicles vitals notablement complexos, sovint amb tres fases, dues de vida lliure i una altra més o menys paràsita, on no intervé absolutament cap cèl·lula flagel·lada. Més avall, ho expliquem amb més detall.
Hi ha dos grans grups a l’interior dels rodòfits, el de les bangiofícies (Bangiophyceae) i el de les florideofícies (Florideophyceae). Hi ha encara una certa polèmica sobre la categoria taxonòmica (de classe o de subclasse) que cal donar a aquests grups. Els arguments, en un o en un altre sentit, són sovint subtils. Els tractarem aquí com a classe. Les bangiofícies acostumen a tenir una morfologia i un cicle vital més senzills que els de les florideofícies.
Morfologia i tipus de creixement
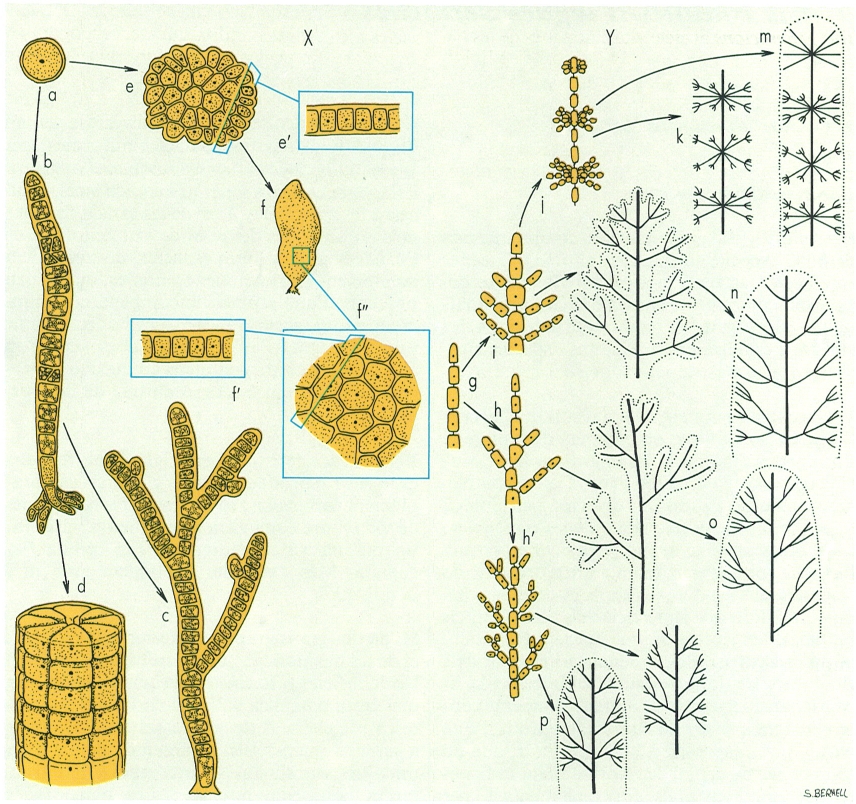
Tipus d’organització dels rodòfits. A l’esquerra, tipus organitzatius de les bangiofícies, ordenats per complexitat creixent, que posen de manifest l’evolució des del tipus unicel·lular (a Porphyridium) fins al pseudo-parenquimàtic (f’(f’/f") Porphyra), passant pel filamentós (c Goniotrichum, d Bangia) i altres etapes intermèdies (b Erythrotrichia, e (e’) Erythrocladia). A la dreta, ídem per a les florideofícies, amb filaments uniseriats (g), amb ramificacions esparsa (h/h’), oposada (i) o verticil·lada (j), o formant tal·lus filamentosos diversament ramificats (k, l) o pseudo-parenquimàtics (m, n, o, p).
Maber, a partir de Scagel, modificat pels autors
Els rodòfits no atenyen la complexitat morfològica que caracteritza els feòfits. En canvi, la diversitat de formes i aspectes que hi trobem és realment notable: espècies d’estructura coccal, en filaments senzills o ramificats, en cintes, làmines, formes folioses, etc.
Les bangiofícies són les que presenten morfologies més senzilles, amb representants unicel·lulars, filamentosos (ramificats o no, uni o pluriseriats) i laminars, aquests darrers, amb una o dues capes de cèl·lules. A les florideofícies no trobem cap representant unicel·lular. Les espècies més senzilles són formades per filaments ramificats. Sovint hi és ben palesa una organització heteròtrica, és a dir, un sistema de filaments reptants, a partir dels quals creixen filaments erectes. A les espècies més complexes, formades per agregació o reunió lateral de filaments, fins arribar a un aspecte cel·lulós (pseudo-parènquima), podem distingir dos grans grups de morfologies, segons si els filaments que s’han reunit procedeixen majoritàriament de la part reptant o de la part erecta d’un sistema heteròtric com el que hem descrit abans.
D’aquesta manera, en el primer cas, tenim un tal·lus de tipus prostrat, aplicat al substrat, com els que són típics de les criptonemials i de les gigartinals, més o menys incrustants. Entre les formes erectes, en les quals la part reptant resta reduïda al sistema de fixació, la diversitat d’aspectes possibles és molt més gran, i hi podem distingir dos tipus de creixement: l’uniaxial, quan tot el tal·lus s’ha originat a partir d’una cèl·lula apical única (és a dir, consta de totes les ramificacions que surten d’un sol eix, reunides), i el multiaxial, amb molts filaments, és a dir eixos, de creixement independent, paral·lels i amb més o menys branques, reunits entre ells. El creixement intercalar és infreqüent.
L’especialització de les cèl·lules vegetatives no és gaire important. Amb freqüència podem distingir-hi un còrtex compacte, format per cèl·lules ben pigmentades, i una medul·la a dins, més o menys laxa, on és possible distingir els filaments (medul·la filamentosa), o bé formada per cèl·lules incolores més o menys grosses (medul·la cel·lulosa).
Encara que tot l’aparell vegetatiu dels rodòfits es basa solament en la combinació d’aquests elements de base, relativament senzills (pel que fa a diferències entre cèl·lules i a tipus d’organització), la diversitat de formes que trobem a les florideofícies és molt gran, com a resultat de la combinació de tipus de ramificació, de simetries, de formacions foliàcies, etc.
Reproducció i cicles vitals

Cicle vital d’una bangiofícia, concretament d’una espècie del gènere Porphyra (to clar, fase haploide (n); to fosc, fase diploide (2n); M reducció meiòtica): a1/a2 gametòfits masculí/femení (a’1/a’2 detall de talls transversals de la làmina, augmentats, amb cèl·lules vegetatives); b1/b2 formació, a la làmina, de gametangis masculins/femenins (espermatangis/carpogonis); c1 espermacis; d fusió de gàmetes (singàmia); e carpòspores; f germinació de la carpòspora; g formació i desenvolupament de l’esporòfit (estadi Conchocelis); h monòspora; i concòspores; j germinació de la concòspora; k inici de desenvolupament del gametòfit; I aplanòspora.
Maber, a partir de dades facilitades pels autors
Si l’aparell vegetatiu dels rodòfits és relativament senzill, la màxima complexitat d’aquestes algues la trobem, com deiem més amunt, a l’aparell reproductor i al cicle vital. Molts dels cicles dels rodòfits solament han pogut ésser compresos després d’un seguiment molt precís de cultius mantinguts en aquari. Tot i així, encara persisteixen algunes incògnites, per exemple, referents a la meiosi i al moment del cicle en què es situa en algunes espècies, el significat de determinades estructures, etc. Per a descriure tota aquesta complexitat d’estructures i de processos ha calgut crear un ric lèxic científic que, malgrat que part dels termes han caigut ja en desús, és encara una font de dificultat per als estudiants, i de disgust, potser justificat, per als no especialitzats.
D’una manera molt general, cal dir que hi ha tres generacions al cicle vital, una més de les habituals a les algues, que són l’esporòfit (diploide) i el gametòfit (haploide). Als rodòfits, hi ha una tercera generació, també diploide, que forma una petita planta que viu fixa sobre el gametòfit femení, tot traient-ne els aliments que necessita per a viure. És el gonimoblast o carposporòfit.
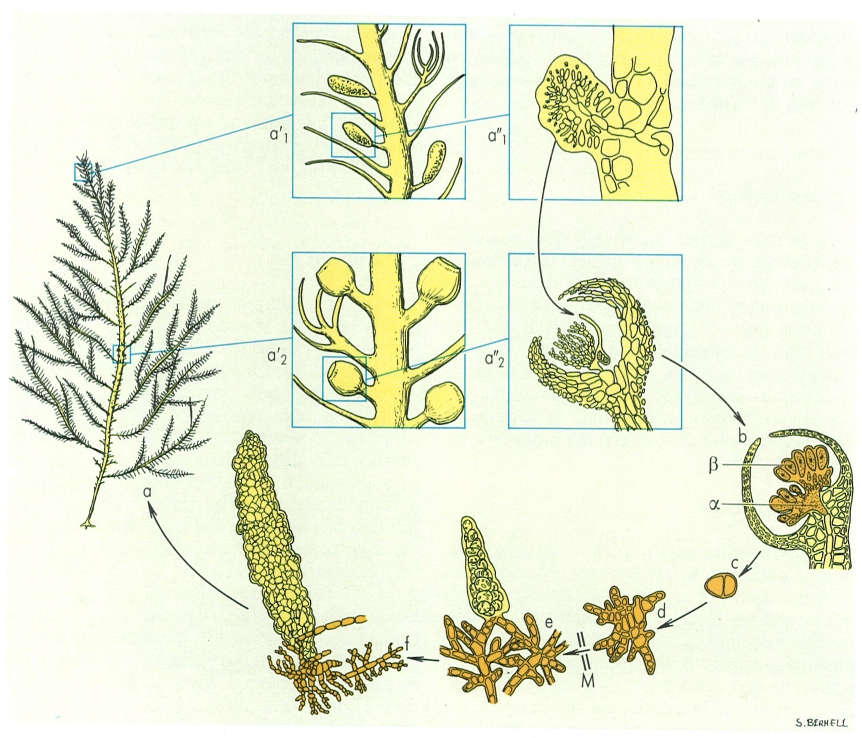
Cicle vital d’una florideofícia (II), concretament de l’espècie Bonnemaisonia asparagoides (to clar, fase haploide (n); to fosc, fase diploide (2n); M reducció meiòtica): a gametòfit (a’1/a’2 espermatangiófors/cistocarps, augmentats; a1"/a2" espermatangiòfor amb esperma tangís/cistocarp amb el carpogoni, més augmentats); b formació del carposporòfit (α) dins del cistocarp, després de la fecundació, i inici de la formació de carpòspores (β); c carpóspora en germinació; d inici de desenvolupament de l’esporòfit; e esporòfit (estadi Hymenoclonium serpens), amb probable meiosi en algunes cèl·lules; f desenvolupament gradual del gametòfit.
Maber, a partir de fonts diverses facilitades pels autors
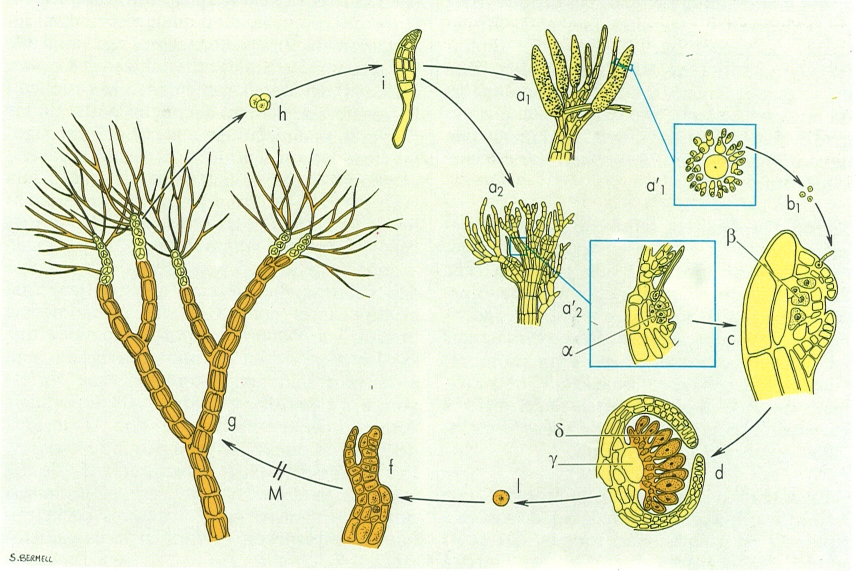
Cicle vital d’una florídeoficia (I), concretament d’una espècie del gènere Polysiphonia (to clar, fase haploide (n); to fosc, fase diploide (2n); M reducció meiòtica): a1/a2 gametòfits masculí/femení, el primer amb tricoblasts modificats on s’allotgen els espermatangis (espermatangiòfors), el segon amb una branca carpogonial (a’1 secció transversal d’un espermatangiòfor amb espermatangis; a’2 detall de la branca carpogonial, constituida per quatre cèl·lules, on pot observar-se la cèl·lula sustentadora basal (α)); b1 espermacis; c fase de fecundació amb la subsegüent ulterior migració del nucli diploide de la cèl·lula auxiliar (β); d cistocarp (γ) i carposporòfit (δ) amb carpòspores en formació; e carpòspora; f detall d’un ràmul del tetrasporòfit; g tetrasporòfït amb tetràspores en formació a la part terminal dels ràmuls; h tetràspora; i germinació de la tetràspora.
Maber, a partir de fonts diverses facilitades pels autors
Els rodòfits, i gairebé sense cap excepció, presenten reproducció sexual per oogàmia. La generació gametofítica, haploide, porta anteridis (anomenats espermatangis) i oogonis (anomenats carpogonis), tots sobre un mateix individu, o en individus separats. Els espermatangis produeixen espermacis en gran nombre. Aquests són cèl·lules uninucleades, de moviments passius, ja que no tenen flagel, però el seu paper és el mateix que el d’un espermatozoide: aportar el nucli masculí.
El carpogoni té una papil·la o, més sovint, una prolongació filiforme, la tricògina, destinada a captar els espermacis quan aquests, que acostumen a madurar tots alhora, queden lliures a l’aigua en gran nombre. Amb tot, la probabilitat que un espermaci trobi una tricògina i hi quedi enganxat, no és pas gaire gran. Per això molts rodòfits presenten adaptacions tendents a multiplicar l’efectivitat de la fecundació, basades en la multiplicació del nucli i del zigot per mitosi i fent que, allí on hi ha hagut una sola fecundació, acabin tenint molts nuclis, equivalents a molts zigots.
Així, és relativament freqüent entre les florideofícies més evolucionades, que del carpogoni fecundat surtin unes prolongacions que van a trobar una o més cèl·lules especialitzades (cèl·lules auxiliars) del mateix gametòfit, s’hi fusionen i hi transfereixen còpies del nucli zigòtic. En tot cas, ja sigui directament a partir dels carpogonis fecundats, ja a partir de les cèl·lules auxiliars, comencen a créixer uns petits arbúsculs, formats per cèl·lules diploides, que viuen fixes sobre el gametòfit, a les seves expenses: són els carposporòfits (o gonimoblasts), i corresponen a aquesta generació, exclusiva de la majoria dels rodòfits, que té com a paper principal multiplicar el nombre de nuclis diploides i produir carpòspores, espores diploides que es dispersen i, en germinar, originen una altra generació també diploide, que correspon a l’esporòfit veritable. Als esporangis formats per l’esporòfit té lloc la meiosi, de manera que s’hi formen quatre espores. Per això se’n diuen tetrasporangis, de les espores, tetràspores, i de l’esporòfit, tetrasporòfit. En dispersar-se i després de germinar les tetràspores, es formen nous gametòfits.
No és rar que, en formar-se el carposporòfit, hi creixi al voltant una coberta protectora, originada pel gametòfit. L’estructura resultant, que sovint té l’aspecte d’una boleta, rep el nom de cistocarp.
Sobre la base d’aquest cicle vital típic, hi ha no pas poques variacions, de les quals esmentarem solament algun exemple. Així, a les bangiofícies, ens trobem amb molts casos en què la sexualitat és encara desconeguda. Això no passa amb Bangia i Porphyra, dos dels gèneres més característics d’aquest grup. Les cèl·lules del tal·lus de Porphyra, que és foliad, es converteixen en espermatangis i en carpogonis. Un cop realitzada la fecundació, el zigot es converteix en un cert nombre de carpòspores diploides. Falta, doncs, la fase carposporofítica (o és extraordinàriament reduïda). Aquesta és una diferència fonamental, respecte a les florideofícies. Les carpòspores, en germinar, produeixen un esporòfït filamentós, poc visible (que havia estat descrit com un gènere independent, Conchocelis), el qual, prèvia meiosi, produeix les concòspores, que donaran lloc, novament, a la fase gametofítica. Les dues generacions, la laminar i la filamentosa, es poden reproduir elles mateixes, asexualment. Hom ha trobat que aquests processos van regulats per la longitud del dia. D’aquesta manera, durant l’hivern i el començament de la primavera trobem la fase foliosa, mentre que la resta de l’any, són els petits filaments de l’esporòfit els que persisteixen, normalment vivint sobre conquilles de mol·lusc buides.
El cicle de Polysiphonia és un dels típics de les florideofícies. El gametòfit porta espermatangis o carpogonis, sobre individus separats. Els carpogonis es formen a l’interior de cistocarps en forma de copa. El tetrasporòfït és idèntic, morfològicament, al gametòfit.
A Bonnemaisonia trobem un segon tipus de cicle vital, caracteritzat per un gametòfit i un esporòfit d’aspecte molt diferent. Les carpòspores, en germinar, originen un individu de mida petita, filamentós, que havia estat descrit com un gènere independent, anomenat Trailiella o Hymenoclonium. Gairebé no es coneix el camí que porta a la formació del gametòfit, la fase més grossa i aparent, ja que no ha estat demostrada la presència de tetràspores a la fase filamentosa. Podria ésser que, en algunes espècies, la fase haploide es produís després de la meiosi d’alguna cèl·lula somàtica de la fase filamentosa.
A part els exemples suara comentats, han estat descrites moltes altres variacions del cicle vital dels rodòfits: presència de carpotetràspores, diversos tipus de mitòspores i de meiòspores, presència, en un mateix individu, de carpòspores i tetràspores, etc. Recordem que el cicle vital d’una espècie no és una cosa totalment immutable. En efecte, a partir d’un patró bàsic actuen, tot modulant-lo, els factors ambientals els quals, de retruc, són reflex de les fluctuacions climàtiques, localització geogràfica, etc.
Ecologia, distribució i interès econòmic
D’una forma esquemàtica, podríem dir que les florideofícies viuen en condicions ambientals que es mantenen relativament estables. Molt sovint, es tracta d’espècies de creixement lent i d’estratègia ecològica de tipus "conservador" (k-estrategues). Amb tot, les excepcions són freqüents. Així, el component algal de moltes zones molt il·luminades i mitjanament pol·lucionades, el constitueixen florideofícies com Corallina i Gelidium. En canvi, les bangiofícies es caracteritzen per un creixement molt ràpid i per una certa tolerància a les condicions fluctuants. Això les caracteritza com a espècies oportunistes (r-estrategues), que poden proliferar en llocs i en èpoques en què l’ambient es mostri canviant de forma imprevisible, unes condicions que resulten desfavorables per a altres tipus d’algues.
Els rodòfits apareixen des de la zona mediolitoral fins a les profunditats més grans que siguin compatibles amb la vida vegetal (a les nostres costes, 100 m o més). A alguns autors els ha semblat que l’explicació de la seva supervivència a unes profunditats tan considerables es troba en la natura dels seus pigments, el color dels quals és complementari del de la llum que hi ha a partir de 20-30 m, d’una tonalitat clarament blavosa, a causa de l’absorció selectiva de l’aigua sobre els colors més càlids.
A diferència dels feòfits, que tenen el seu centre de gravetat a les aigües boreals, els rodòfits presenten molts representants de distribució centrada a les aigües càlides o tropicals. Però no seria prudent que generalitzéssim massa aquesta tendència. Per altra banda, la major part dels rodòfits són marins, però hi ha no pas poques espècies (tant de bangiofícies com de florideofícies), que viuen a les aigües dolces o salabroses.
A nivell mundial, els rodòfits tenen una certa importància econòmica. Alguns (per exemple, Porphyra) són molt usats com a aliment i condiment (i no tan sols pels xinesos i els japonesos, sinó també en moltes costes atlàntiques europees). Però des del punt de vista industrial, és molt més important la seva recol·lecció (de les platges, o mitjançant escafandrers) com a font de polisacàrids, procedents de les substàncies gelificades de la seva paret cel·lular. Les més importants són l’agar (emprat per a gelificar els medis de cultiu en microbiologia, entre moltes altres aplicacions) i la carrageenina (utilitzada en aprestos, farmàcia, alimentació, etc.). No hi ha tradició en la producció d’aquests productes a les nostres costes.
En algunes regions, les espècies de la família de les coral·linàcies, incrustades de carbonat càlcic (grapissar), s’empren per a corregir l’aciditat excessiva de certes terres de conreu. En altre temps, s’havien usat també (per exemple, Corallina officinalis i espècies afins), com a antiàcid digestiu. La utilització d’aquest grapissar ("maërl") en agricultura és important en regions com Bretanya, Galícia o Irlanda. Però, en part per manca d’espècies adequades, de tradició i, encara més, perquè no acostumen a haver-hi problemes d’aciditat als nostres camps, no té gaire importància la utilització de les algues coral·lines als Països Catalans.
Sistemàtica
Dividim el fílum dels rodòfits en dues classes, la de les bangiofícies (Bangiophyceae), d’organització i cicle vital més simples, i la de les florideofícies (Florideophyceae), molt més gran i diversificada, d’organització i cicle vital més complexos. Es també freqüent que hom atorgui a aquests dos grans grups categoria de subclasse; tenim, llavors, les bangiofícides i les florideofícides (hom es refereix sovint a aquestes darreres amb el nom de florídies).
La classe de les bangiofícies consta de tres ordres: el de les porfiridials (Porphyridiales), el de les bangials (Bangiales) i el de les compsopogonals (Compsopogonales); la de les florideofícies en consta de cinc: el de les nemalionals (Nemalionales), el de les gigartinals (Gigartinales), el de les criptonemials (Cryptonemiales), el de les rodimenials (Rhodymeniales) i el de les ceramials (Ceramiales).