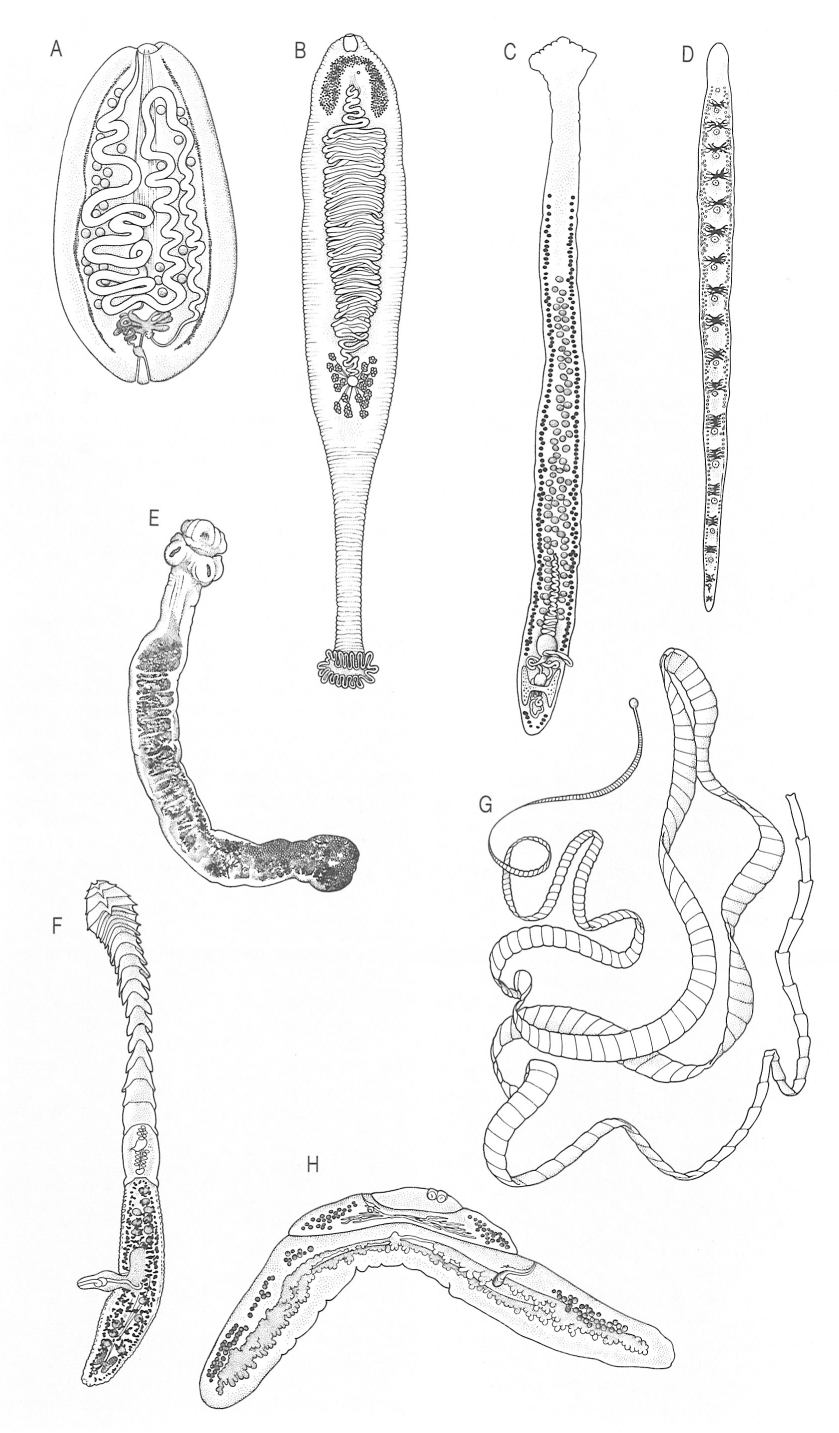
Aspecte general dels cestodes monozoics i dizoics. A, B Cestodes monozoics cestodaris: A adult d’Amphilina foliacea, B adult de Gyrocotyle. C, D Cestodes monoics eucestodes: C adult de Caryophyllaeus laticeps, D adult de Spathebothrium simplex. E, F, G, H Cestodes polizoics: E adult de Nematoparataenia southwelli, sense segments, ni porus sexuals, ni bossa del cirrus, ni vagina; els seus òrgans sexuals ocupen zones indistintes dins el curt estróbil; F adult de Litobothrium alopias, amb segments anteriors modificats i proglotis terminal madur. G adult de Taenia saginata (diversos metres de llargada), H adult de Meggittina baerí, amb tendència a patir divisió estrobilar posterior.
Biopunt, a partir de fonts diverses
La classe dels cestodes comprèn platihelmints que en l’estat adult parasiten l’intestí o els seus diverticles (rarament el celoma) de vertebrats i, només excepcionalment, d’invertebrats: actualment hom només en coneix Archigetes, un cariofil·lideu, que parasita, en estat adult, oligoquets dulciaqüícoles, i Cyathocephalus truncatus, un espatebotrideu, que viu a l’hemocel d’un amfípode. En conjunt, presenten caràcters típics dels platihelmints, com la superfície proveïda d’un veritable tegument, però no tenen boca ni tracte digestiu. Hom en distingeix dos grans grups: els eucestodes i els cestodaris. Els eucestodes tenen forma de cinta segmentada (excepte alguns cariofíl·lides i els espatebòtrides), amb una estructura de fixació anomenada cap o escòlex a l’extrem anterior. Formen el grup de cestodes més nombrós i transcendent, pel seu interès aplicat en medicina i veterinària.
Els cestodaris
Els cestodaris formen un grup petit i poc conegut. Tots són endoparàsits de l’intestí i el celoma de peixos, rarament de rèptils, i tenen característiques intermèdies entre els veritables cestodes i els trematodes. Els cestodaris adults no tenen aparell digestiu i tenen músculs parenquimàtics. La larva és del mateix tipus que la dels eucestodes, i essencialment allò que la’n diferencia és que la forma inicial, anomenada licòfora, presenta 10 ganxos, i no 6 com la larva dels eucestodes. S’assemblen als trematodes pel fet de tenir ventoses semblants a les dels digenis, no tenir escòlex i presentar un sol conjunt d’òrgans reproductors hermafrodites, a més de la manca d’estrobilització o segmentació. La consideració d’aquestes característiques anatòmiques, i de la posició filogenètica dels seus peixos hostes, suggereix que es tracta d’un grup de paràsits arcaic, originat durant el Mesozoic, o probablement abans. Hom en distingeix dos ordres: els amfilínides (Amphilinida), paràsits del celoma d’esturions, altres peixos primitius i tortugues, i els girocotílides (Gyrocotylida), paràsits intestinals de quimèrids marins, d’anatomia molt diferent. No es coneix cap cicle complet de cestodari, però hom ha suggerit una certa diheteroxènia, amb almenys la intervenció d’un crustaci amfípode com a hoste intermediari, i de l’esturió com a hoste definitiu (amfilínides) i fins i tot potser un cicle directe (girocotílides).
Els eucestodes
Morfologia

L’escòlex, part terminal anterior del cos dels cestodes, s’ha especialitzat a fixar el paràsit en l’hoste. Pot tenir formes i elements de fixació molt diversos. El dibuix de la mostra, a dalt, diferents tipus de ganxos rostel·lars de cestodes ciclofíl·lids, corresponents als gèneres següents: 1 Dipylidium, 2 Biuterina, 3 Davainea, 4 Hymenolepis, 5 ganxo gros de Taenia, 6 ganxo petit de Taenia. A baix, hom ha representat l’escòlex de: A Orygmatobothrium musteli, B Duthiersia fimbriata, C Calliobothrium verticillatum, D Eubothrium crassum, E Echeneibothrium fallax, F Polypocephalus medusia i G Anthobothrium cornucopia.
Biopunt, a partir de fonts diverses
En el cos dels eucestodes típics hom distingeix tres regions: l’escòlex o cap, amb els òrgans de fixació; el coll, regió indivisa, i l’estròbil, zona segmentada que forma la cadena d’anells típica de les tènies. Les dimensions que poden atènyer són d’allò més variables: des d’espècies microscòpiques, que no arriben al mil·límetre, fins a espècies gegantines, d’uns quants metres de longitud, com Diphyllobothrium latum, paràsit de carnívors i de l’home, que arriba a fer fins 15 m.

Anells sexuals madurs d’una tènia (Taenia).
C. Gautier / Jacana.
L’escòlex fa la funció de fixar el verm al microhàbitat que ocupa en l’hoste. Pot presentar formes molt diverses i és equipat amb tota mena de ventoses, solcs, ganxos, espines o glàndules, que li faciliten la fixació; en algunes espècies és molt simple o no existeix (Spathebothrium simplex), i en alguns casos (Fimbriaria fasciolaris) hi ha al seu lloc un altre òrgan de fixació, el pseudoscòlex, format a partir de l’extrem anterior de l’estròbil.
Els òrgans de fixació principals són musculars o quitinosos i en forma de ganxo. Entre els musculars destaquen: les ventoses o acetàbuls, en forma de calze i amb musculatura pròpia (típiques dels ciclofil·lideus i presents generalment en nombre de 4); els botridis, ventoses rudimentàries allargades i foliàcies, molt mòbils, amb musculatura pròpia només als marges i als envans transversals (típiques dels tetrafil·lideus i generalment en nombre de 4); i els botris, fenedures o solcs longitudinals, sense musculatura pròpia (típics dels pseudofil·lideus i generalment en nombre de 2). A més, pot haver-hi un òrgan musculós addicional, el rostel, apical, protràctil i retràctil generalment, que s’allotja dins una bossa o beina, i que pot portar ganxos o no. Entre els òrgans de fixació quitinosos i en forma de ganxo, que generalment es localitzen en el rostel, n’hi ha de diversos tipus, i són especialment típics els que tenen forma d’ungla de gat (típics dels tènids), en forma de martell (davainèids), de forca (himenolepídids) o d’espina de roser (dilepídids); sempre s’hi poden diferenciar tres regions, que reben el nom de fulla, mànec i guarda. A més dels òrgans de fixació esmentats, l’escòlex pot presentar cèl·lules glandulars, que en alguns casos (cariofíl·lides i pseudofí·lides) sembla que produeixen secrecions especials per a ajudar a adherir-se a la mucosa intestinal de l’hoste.
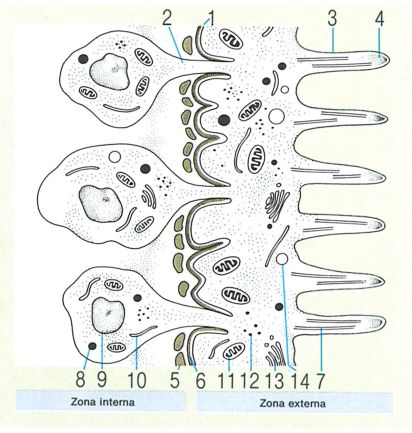
Estructura del tegument dels cestodes: 1 làmina basal, 2 pont citoplasmàtic, 3 microvellositats, 4 àpex electrodens de les microvellositats. 5 músculs longitudinals, 6 músculs circulars, 7 microtúbul, 8 acumulacions de lípids, 9 nucli, 10 reticle endoplasmàtic, 11 mitocondri, 12 grànuls de glicogen, 13 aparell de Golgi, 14 vesícula.
Biopunt, a partir de fonts diverses
El coll, la regió compresa entre l’escòlex i el cos segmentat o estròbil, és indivís i generalment més estret que l’escòlex. Té a la seva part posterior cèl·lules germinals que originen els anells de l’estròbil; en les espècies que no tenen coll, aquesta zona germinal es troba a la part posterior de l’estròbil.
L’estròbil constitueix la major part del cos del cestode adult. En general és integrat per una sèrie repetitiva d’unitats funcionals, separades exteriorment per constriccions, que internament contenen un conjunt complet (o dos en alguns casos) d’òrgans reproductors. Cadascuna d’aquestes unitats funcionals és un anell o proglotis. L’estròbil, per tant, no és una cadena d’unitats independents i separades sinó d’unitats funcionals que presenten plecs tegumentaris seriats o constriccions concèntriques que originen una repetició d’estructures externes, la qual pot correspondre o no a una repetició interna de sistemes d’òrgans. La formació de nous anells (proglotització) a la part posterior del coll fa que cadascun d’ells es vagi allunyant progressivament de l’escòlex i, en aquesta aproximació a la part posterior del cos, els anells van madurant sexualment; la longitud de l’estròbil va augmentant a mesura que creix el nombre d’anells, que, però, creix només fins a un cert nombre (diferent per a cada espècie), a partir del qual els anells que arriben a l’extrem final del cos, que són madurs sexualment, es desprenen.
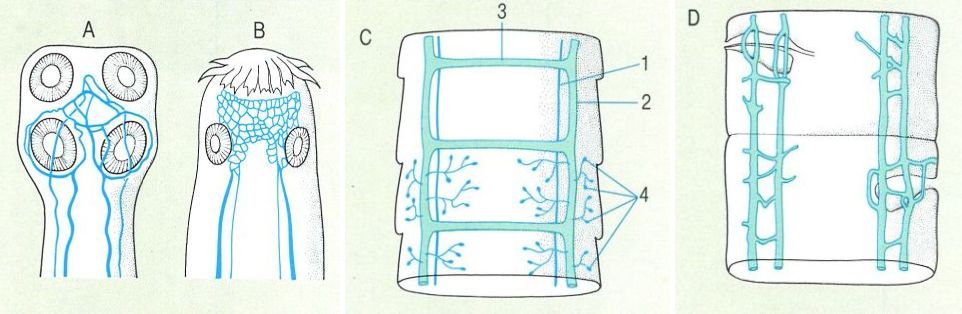
Sistema excretor dels cestodes. A Sistema excretor en l’escòlex de Proteocephalus torulosus. que mostra la connexió en anell únic; B sistema excretor de tipus reticular en l’escòlex de Taenia; C sistema excretor a l’altura de l’estròbil; D canals excretors longitudinals dorsals de Proteocephalus ambloplitis. Hom ha representat en el dibuix: 1 canal lateral longitudinal dorsal, 2 canal lateral longitudinal ventral, 3 canal transversal ventral, 4 cèl·lules flamígeres.
Biopunt, a partir de fonts diverses
Al llarg d’aquest desplaçament cada anell passa per diferents estadis de maduresa. Així, en un estròbil es poden distingir tres regions: la primera, pròxima a l’escòlex, és la dels anells immaturs, que són curts i petits, i tenen només vestigis d’òrgans genitals; la segona és la dels anells sexuals o anells madurs, més llargs i amples, que presenten els òrgans genitals ben formats i funcionals; i la tercera és la dels anells gràvids, més grossos i portadors d’un úter totalment desenvolupat i farcit d’ous, amb els òrgans genitals en fase de degeneració o totalment desapareguts. En alguns casos hom pot distingir encara altres estadis intermedis de maduració: per exemple, en els ciclofíl·lides, hom ha descrit les regions d’anells immadurs, prematurs, madurs, postmadurs, pregràvids, gràvids, i fins i tot postgràvids. Els anells no sempre arriben al seu estat final de gravidesa adherits a l’estròbil. Segons això, hom parla de cestodes (o anells) apolítics, en les espècies en què l’anell es desprén de l’estròbil quan l’úter ja és totalment gràvid (tènids); de cestodes (o anells) pseudoapolítics, o simplement anapolítics, quan no es desprenen fins que els ous no hagin estat expulsats, sigui per dehiscència (tetrafíl·lides) o per un orifici de posta (difil·lobòtrides); de cestodes (o anells) euapolítics quan el despreniment té lloc abans que l’úter sigui totalment gràvid (tetrarínquides, alguns tetrafíl·lides i ciclofíl·lides); i de cestodes hiperapolítics, quan es desprenen abans que els òrgans genitals no hagin adquirit la maduresa (alguns casos, com en el gènere Pseudhymenolepis). D’altra banda, es parla d’anells craspedotes quan els marges posteriors de l’anell anterior cobreixen parcialment els marges anteriors de l’anell que ve al darrere (alguns anoplocefàlids, per exemple), i d’anells acraspedotes quan no és així i apareixen les constriccions circulars típiques de l’estròbil (propi dels dipilídides).

Anells hiperapolítics aïllats del cestode Pseudhymenolepis redonica, paràsit intestinal de la musaranya comuna (Crocidura russula) al nostre país. Aquest tipus d’anells es desprenen de l’organisme mare abans que els òrgans genitals siguin madurs.
Santiago Mas-Coma.

Els anells gràvids dels ciclofíl·lides (a l’esquerra, de Pseudocatenotaenia matovi, paràsit del ratolí de bosc) contenen els ous que s’acumulen a l’úter cec; l’aparell reproductor hermafrodita ha desaparegut. En els pseudofílides els anells gràvids mantenen l’aparell reproductor hermafrodita mentre l’úter és obert, de manera que els ous són expulsats per un porus especial. Hom ha representat en el dibuix diferents tipus d’úters: A úter ramificat, amb porus uterí, d’un tetrafíl·lide; B úter en anell, de Cyclustera capito; C úter resolt en càpsules parenquimàtiques pluriovulades, de Raillietina anaplocephaloides; D úter amb òrgan parauterí, de Biuterína distincta; E úter en doble ferradura, de Paronia ambigua. Hom ha indicat en el dibuix: 1 porus uterí, 2 òrgan parauterí.
Santiago Mas-Coma; Biopunt, a partir de fonts diverses
Els cestodes no tenen aparell digestiu, ja que l’absorció de nutrients presents a l’intestí de l’hoste es fa a través de la paret externa del cos; aquesta és un tegument sincític: la zona externa és citoplasmàtica, i la interna és formada per cossos cel·lulars connectats amb la zona externa per mitjà de ponts citoplasmàtics. El citoplasma distal és limitat per membranes plasmàtiques, una d’externa i una d’interna; la superfície del tegument és revestida de glicocàlix (capa de polisacàrids), la interna descansa sobre una làmina basal de fibretes connectives. El tegument presenta unes formacions característiques, els microtriquis, projeccions molt fines de la zona externa que fan que augmenti la superfície tegumentària lliure i intervenen en l’adhesió de l’animal a la mucosa de l’hoste. En alguns estadis larvals, el parènquima pot acumular substàncies inorgàniques en forma d’anells concèntrics anomenats corpuscles calcaris, separats del citoplasma cel·lular per una membrana molt plegada. La musculatura subtegumentària és constituïda per una capa externa de fibres circulars i una capa interna de fibres longitudinals; la musculatura parenquimàtica, formada per fibres longitudinals (externes), transverses i circulars (internes), divideix el parènquima en dues zones: una de medul·lar, per l’interior de la musculatura parenquimàtica, i una de cortical, per l’exterior d’aquesta. Les ventoses, el rostel i el sistema reproductor disposen de fibres musculars més especialitzades. No hi ha aparell respiratori ni sistema circulatori, com tampoc aparell digestiu. El sistema excretor és constituït per tres elements principals: en primer lloc, els canals col·lectors, dos de ventrals, amples, i dos de dorsals, més prims, que recorren el cos longitudinalment, connectats generalment per altres canals transversals; en segon lloc, les cèl·lules protonefridials excretores o flamígeres, que es troben disperses pel parènquima; i en tercer lloc, el túbuls que connecten aquestescèl·lules amb els canals col·lectors. El sistema nerviós consisteix en dos ganglis laterals situats a la regió de Fescòlex i connectats per una comissura transversal; d’aquest anell parteixen fibres nervioses cap endavant, que innerven l’escòlex, i els troncs laterals, dorsals i ventrals que es dirigeixen cap a la part posterior del cos, també connectats entre ells per comissures transversals, en nombre variable segons les espècies.
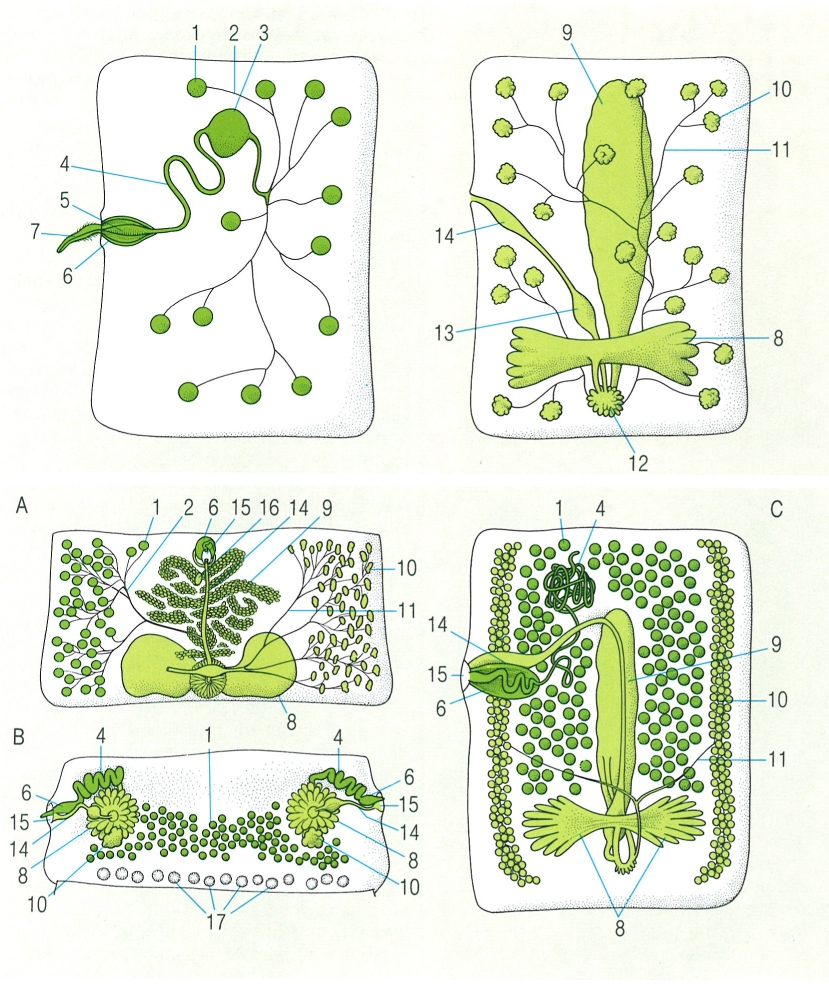
Aparell reproductor dels cestodes. A dalt, organització de l’aparell reproductor (masculí, a l’esquerra; femení a la dreta) en els proglotis. A baix, distribució dels òrgans reproductors en proglotis madures en tres casos: Diphyllobothrium latum (A) Moniezia expansa (B) i en un tetrafíl·lid (C). Hom ha indicat en el dibuix: 1 testicles, 2 conducte eferent, 3 vesícula seminal externa, 4 espermiducte, 5 vesícula seminal interna, 6 bossa del cirrus, 7 cirrus, 8 ovaris, 9 úter, 10 fol·licles vitel·lins, 11 conducte vitel·lí, 12 glàndula de Mehlis al voltant de l’ootip, 13 receptacle seminal, 14 vagina, 15 porus genital, 16 porus uterí, 17 glàndules interproglotidianes.
Biopunt, a partir de fonts diverses
En general, són hermafrodites, i en els casos excepcionals d’espècies dioiques sembla que cada estròbil pot madurar potencialment com a mascle i com a femella. L’estructura dels òrgans reproductors és semblant a la dels altres platihelmints, i la seva disposició i distribució en el cos varien molt segons els grups. Cada anell presenta en general un aparell reproductor masculí i un de femení (excepcionalment dos de cada sexe o altres combinacions), però en alguns grups els aparells masculí i femení se situen en diferents anells del mateix estròbil. L’aparell reproductor masculí consta d’un nombre variable de testicles immersos en el parènquima medular, de cadascun dels quals surt un conducte eferent, que va a confluir al conducte deferent comú o espermiducte. Aquest pot presentar nanses o una vesícula seminal externa per a emmagatzemar els espermatozoides, i condueix a una bossa del cirrus, dins la qual es converteix en l’aparell copulador. Aquest consta d’una part inicial anomenada conducte ejaculador, que es pot dilatar formant una vesícula seminal interna, i una regió muscular distal o cirrus, espinosa o llisa, protráctil, que s’evagina a l’exterior a través del porus genital masculí. Aquest pot obrir-se al marge lateral de la proglotis (en els tetrafíl·lides i els ciclofíl·lides) o a la seva superfície plana (en els pseudofil·lideus). L’aparell reproductor femení consta d’un ovari únic amb estructures associades, com ara l’oviducte, que surt de l’ovari a través d’un esfínter (oocapte) i va a desembocar al conducte vitel·lí comú, resultant de la confluència dels conductes procedents de les glàndules vitel·lògenes. Aquestes poden disposar-se formant un vitel·lari compacte (ciclofíl·lides) o formant fol·licles (pseudofíl·lides, tetrafíl·lides). Els conductes vitel·lins poden formar un magatzem vitel·lí, del qual parteix el conducte vitel·lí comú. Més endavant de la confluència d’aquests conductes, l’oviducte s’eixampla i forma l’ootip, que, com en tots els platihelmints, és envoltat de cèl·lules glandulars, el conjunt de les quals rep el nom de glàndula de Mehlis. De l’ootip emergeix la vagina, que connecta amb l’exterior pel porus genital femení; aquest generalment s’obre, amb el porus masculí, en un atri genital comú. La vagina presenta, en la seva trajectòria, una dilatació anomenada receptacle seminal, que emmagatzema els espermatozoides després de la inseminació. L’úter també emergeix de l’ootip; es tracta d’un conducte que es dilata molt en els anells gràvids, quan al seu interior s’acumulen els ous madurs. En algunes espècies, l’úter comunica amb l’exterior per un porus de posta o tocostoma, a través del qual s’expulsen els ous; en d’altres, el conducte uterí forma un sac cec i els ous surten a l’exterior perquè es trenca la paret de l’anell gràvid, ja després de l’estròbil. La forma de l’úter varia molt segons les espècies. A vegades els ous resten dins l’úter recoberts d’unes càpsules uterines que poden ésser recobertes al seu torn d’un teixit parenquimatós dens, formant en conjunt les anomenades càpsules parenquimatoses; també es poden formar estructures anomenades parauterines, que es desenvolupen per a fer la funció de l’úter quan aquest es desintegra.
Biologia
Reproducció i desenvolupament
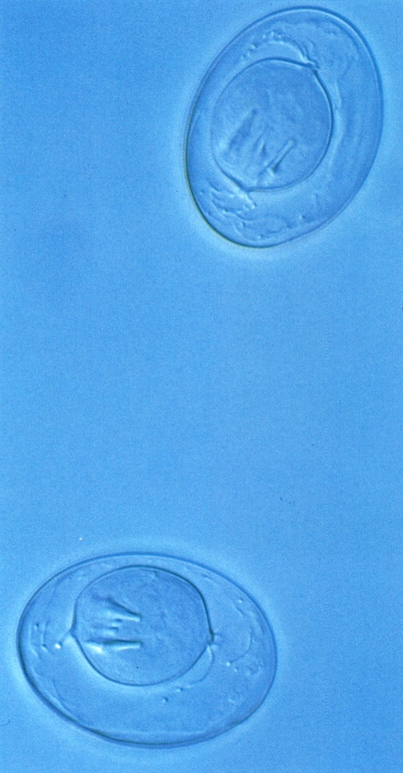
Ous de la tènia Hymenolepis fraterna, paràsit de la rata (Rattus norvegicus) vistos al microscopi interferencial. Els ous embrionats de les tènies contenen el primer estadi larval, anomenat oncosfera o embrió hexacant, que ja disposa de ganxos per a la fixació de la futura larva hexacanta que emergirà de l’ou, dins l’hoste intermediari.
Santiago Mas-Coma.
En general, els cestodes són proteràndrics, és a dir, maduren primer els òrgans masculins, que emmagatzemen l’esperma produït fins a la maduració dels òrgans femenins; alguns, però, són proterogínics, és a dir, la maduració es fa en l’ordre invers. Els espermatozoides són emmagatzemats en les nanses de l’espermiducte o en la vesícula seminal externa, des d’on van a parar al receptacle seminal, per mitjà d’inseminació a través de la vagina. En els hermafrodites (la majoria), la inseminació pot ésser creuada —entre individus diferents— o pot recórrer a l’autofecundació d’un mateix individu, sia dins una mateixa proglotis o entre proglotis diferents d’un mateix estròbil. Hi ha casos extraordinaris en què, no havent-hi obertura vaginal (Dioecotaenia cancellatum), els espermatozoides s’introdueixen dins el parènquima per mitjà del cirrus, trencant la paret de l’anell, alliberant l’esperma (fenòmen anomenat impregnació hipodèrmica) i arribant d’alguna manera fins al receptacle seminal. El contacte entre l’espermatozoide i l’oòcit es produeix generalment dins l’oviducte (rarament en l’ootip o l’úter). En l’ootip es prepara la formació de la càpsula de l’ou, on intervenen les secrecions de les cèl·lules vitel·lògenes i la glàndula de Mehlis.
Cal distingir dos tipus principals d’ous. El primer tipus, propi dels pseudofíl·lides, els tetrafíl·lides i els tripanorinquis, és un ou proveït d’una càpsula gruixuda, constituïda per esclerotina, i d’un opercle; aquests ous s’eliminen abans d’embrionar i alliberen a l’aigua una larva ciliada nedadora, el coracidi, que serà ingerit per l’hoste intermediari. El segon tipus, típic dels ciclofil·lideus, són ous sense càpsula o amb una càpsula molt fina, sense opercle, que quan s’alliberen ja són embrionats i són ingerits per l’hoste abans d’eclosionar. En general, l’ou dels cestodes té, de fora a dins, una closca externa formada per dues membranes, una coberta vitel·lina externa (que és la substància de reserva per a l’embrió), un embriòfor o coberta interna (molt variable i molt característica de cada espècie) i, finalment, l’embrió hexacant o oncosfera (primer estadi larval dels cestodes, caracteritzat pel fet de presentar 6 ganxos distribuïts en tres grups de dos). La larva hexacanta, un cop alliberada de les cobertes externes de l’ou, dins l’hoste intermediari, envaeix un hàbitat paraentèric i pateix una metamorfosi, per la qual canvia gairebé totes les seves estructures i es converteix en el metacestode. Generalment, en els cestodes es reserva el nom de larva per a referir-se a l’oncosfera, i el de metacestode per a totes les fases compreses entre l’oncosfera i l’adult.
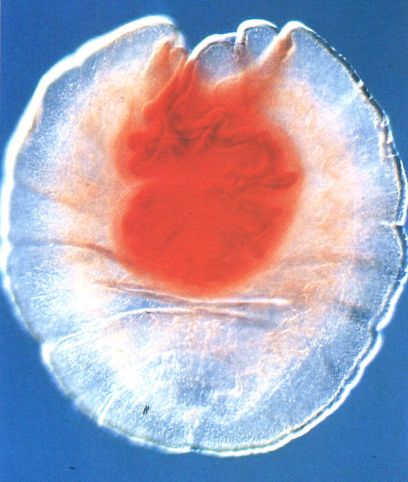
Els tipus d’estadis larvals o metacestodes són molt diversos. En la fotografia hom pot veure un cas molt especial, anomenat tetratiridi (amb l’escòlex invaginat), propi del gènere Mesocertoides.
Santiago Mas-Coma.
El desenvolupament de l’oncosfera en metacestode pot ésser de tipus primitiu o de tipus neotèric. Per l’evolució de tipus primitiu s’arriba a I’estat de metacestode sensé que es formi una cavitat primària interna, i es passa pels estats anomenats procercoide i plerocercoide. En I’estat procercoide, originat en un microhàbitat paraentèric del primer hoste, la larva no té escòlex però sf que té un apèndix posterior (cercòmer), encara dotat dels sis ganxos de l’oncosfera. S’arriba aleshores, generalment en un segon microhàbitat (en els pseudofíl·lides, en un segon hoste), al plerocercoide, estat també acístic, que ja ha perdut els ganxos i el cercòmer, i comença a adquirir una forma de cinta, amb un escòlex ben desenvolupat. En alguns casos (tripanorincs), hi ha una fase peculiar anomenada plerocerc, proveïda d’una estructura posterior vesicular (blastocist), que dona pas a un altre estadi anomenat neoplerocercoide. Hom ha pensat que els cestodes dels grups cariofíl·lides, espatebòtrids i nippotènids es podrien considerar procercoides o plerocercoides que inicien i completen el seu desenvolupament en un hoste únic.
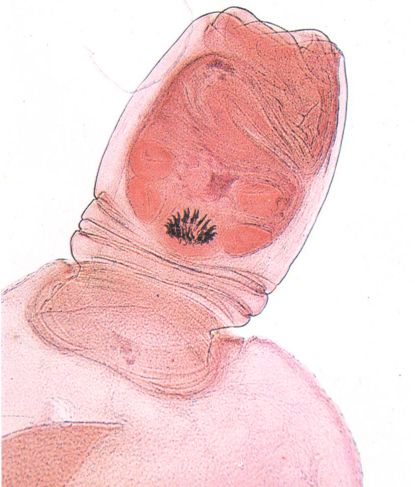

Diferents aspectes de les formes de desenvolupament de les tènies. A dalt, escòlex invaginat en una part desembeinada del cisticerc de Taenia pisiformis. A baix, corones de ganxos rostel·lars d’un escàlex de larva o d’adult.
AGE / Fotostock, Hervé Chaumeton / Jacana, C. Gautier / Jacana.
Pel tipus de desenvolupament neotèric es forma una cavitat primària a I’interior de l’oncosfera. L’escòlex es pot formar externament i després retreure’s per originar una fase anomenada cisticercoide, o bé pot tenir un origen endogen, i llavors es formen fases anomenades cisticercs o cenurs (si s’origina dins el canal d’invaginació) o hidàtide (si s’origina dins la cavitat primària). Aquest tipus de desenvolupament és típic dels ciclofíl·lides. Els metacestodes d’aquest grup es poden reunir, doncs, en dues categories: els monocèfals, proveïts d’un sol escòlex, que inclouen els tipus cisticerc, estrobilocerc, cisticercoide, estrobilocercoide i tetratiridi; i els policèfals, proveïts de diversos escòlexs (protoscòlex), que inclouen el cenur, I’equinococ o hidàtide unilocuar i I’equinococ o hidàtide multilocular (alveolar o alveococ).
El cicle vital
L’estratègia reproductiva dels cestodes es basa fonamentalment en la reproducció sexual dels adults, generalment hermafredites, en els quals té una gran incidèneia el fenomen de I’autofecundació; la reproducció asexual es limita als metacestodes dels ciclofíl·lides de cicle terrestre. Les estratègies de transmissió estan orientades a aconseguir la dispersió de I’espècie en Tespai i en el temps, i la infestació dels hostes que intervenen en el cicle. La inclusió en el cicle d’un estadi de resistèneia (metacestode) hi contribueix decisivament. En generai hi ha una especificitat de tipus fisiològic força gran entre I’adult d’una espècie de cestode i I’hoste definitiu, que es tradueix en l’elecció d’hostes de parentiu estret (de la mateixa família o del mateix gènere), encara que hi ha algunes excepcions ( Diphyllobothrium latum i Hymenolepis diminuta, per exemple, són molt poc específics). L’anàlisi de l’evolució dels grups de cestodes i dels grups d’hostes permet de pensar en una certa especificitat filogenètica. En canvi, pel que fa a l’estadi de metacestode, l’especificitat amb l’hoste intermediari és molt menys acusada. Cada classe de vertebrats té una fauna de cestodes endèmica, i dins una mateixa classe, cada ordre té espècies infestants característiques. Per exemple, els taurons i les rajades tenen una cestodofauna diferent. Entre els rèptils hi ha els paràsits Duthiersia (paràsit de varans) i Bothridium (paràsit de pitons), de distribució discontínua sobre tres continents, cosa que permet pensar en casos de parasitisme molt antics. Cada ordre d’ocells té també els seus cestodes paràsits, i sorprèn el fet que ordres que inclouen espècies amb idèntica alimentació (per exemple, rapinyaires diürns, falconiformes, i rapinyaires nocturns, estrigiformes) tenen paràsits diferents. Diverses espècies del gènere Hymenolepis parasiten insectívors, com eriçons, talps i musaranyes; i, entre les musaranyes, les de dents blanques (crocidurins), tenen espècies paràsites diferents de les de dents vermelles (soricins).
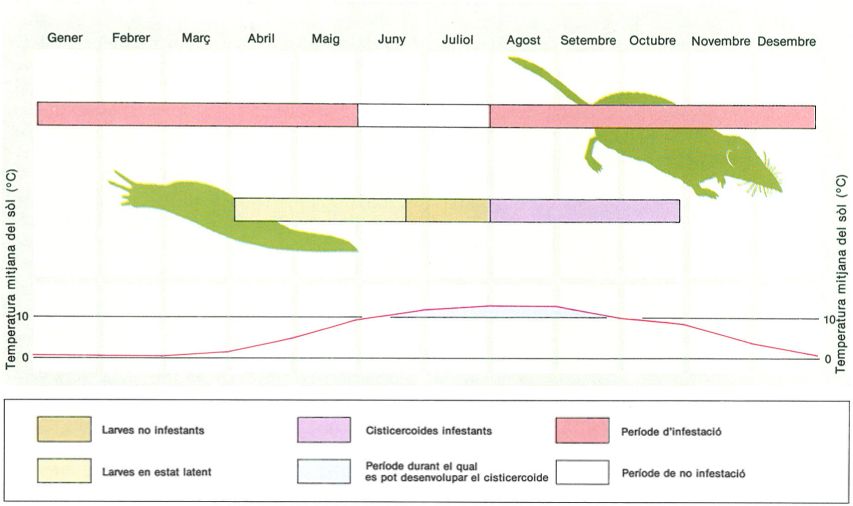
Etapes de la infestació de la musaranya cua-quadrada (Sorex araneus) i del llimac Arion lusitanicus (hoste intermediari) pel cestode Choanotaenia crassiscolex en els Pirineus orientals: correlació entre la dinàmica anual de la població del cestode en els seus hostes i variacions estacionals de la temperatura mitjana del sòl. Al llarg del seu cicle biològic, els llimacs es troben parasitats successivament per larves en estat de latència, larves no infestants i per cisticercoides infestants. En l’hoste definitiu hi ha un període de no infestació, molt breu, i un període llarg d’infestació.
Biopunt, original de Jourdane 1977.
Els cestodes poden utilitzar crustacis, insectes, mol·luscs, àcars, anèl·lids i vertebrats com a hostes intermediaris, mentre que els hostes definitius són gairebé sempre vertebrats. En general, quan l’hoste definitiu és aquàtic, les formes joves també tenen hostes intermediaris aquàtics; el mateix es pot dir dels que tenen un hoste definitiu terrestre. No obstant això, encara hi ha molts grups el cicle biològic dels quals no es coneix. En general s’accepta que els cestodes segueixen cicles indirectes, que necessiten la intervenció d’un o dos hostes intermediaris (cicles diheteroxens o triheteroxens).
En el cicle vital triheteroxè, l’adult deixa anar ous operculats no embrionats al medi aquàtic extern, a través del tocostoma, barrejats amb les femtes de l’hoste definitiu. Un cop a l’exterior, els ous s’embrionen i es desenvolupa al seu interior un coracidi ciliat. Després l’ou es desclou i el coracidi neda fins que és ingerit pel primer hoste intermediari, generalment un artròpode (habitualment, un crustaci). Llavors perd el seu embolcall ciliat i l’oncosfera, ja lliure dins la cavitat digestiva de l’hoste, travessa la paret intestinal i arriba a l’hemocel, on pateix la metamorfosi que el convertirà en un procercoide. El segon hoste intermediari, generalment un peix, s’infesta en menjar-se el primer hoste intermediari, portador del procercoide; aquest travessa la paret de l’intestí de l’hoste i va a la cavitat peritoneal i als músculs esquelètics, on es converteix en un plerocercoide. Quan el que actua d’hoste definitiu, generalment un mamífer terrestre depredador de peixos, es menja el segon hoste intermediari, els plerocercoides infestants queden lliures al seu intestí i es fixen a la mucosa, on atenyen l’estat adult. Si el segon hoste intermediari és ingerit per un vertebrat diferent de l’hoste definitiu habitual ( hoste d’espera o paratènic), els plerocercoides no avancen en el seu desenvolupament, sinó que s’esperen, encapsulats, a ésser ingerits per l’hoste adequat. Aquest cicle és típic dels pseudofil·lideus, i més concretament de Diphyllobothrium latum, l’adult del qual arriba a parasitar també l’home. Però dins el mateix ordre dels pseudofíl·lides hi ha cicles diheteroxens, com el de Bothriocephalus rarus, paràsit de tritons, que té un únic hoste intermediari (copèpode); o el de B. claviceps, paràsit de l’anguila (Anguilla anguilla), també amb un únic hoste intermediari (copèpode), però amb la possible intervenció d’un hoste paratènic (peix), la paret del tub digestiu del qual no travessa. Els proteocefalats tenen un cicle també modificat, ja que passen per dos estadis de plerocercoide l’un en el primer hoste intermediari (artròpode), i l’altre en el segon hoste intermediari.
Els cicles típicament diheteroxens, amb la intervenció d’un únic hoste intermediari, són ben coneguts en l’ordre dels ciclofíl·lides. Els ous, que contenen una larva hexacanta o oncosfera, s’alliberen trencant l’anell gràvid (dins o fora de l’hoste definitiu) o bé després que l’hoste intermediari hagi ingerit l’anell. L’oncosfera queda lliure al tracte digestiu, on travessa la paret de l’intestí i va a instal·lar-se (activament o passivament, a través de la sang) al seu microhàbitat, on pateix la metamorfosi i es converteix en metacestode. En els hostes intermediaris invertebrats formen metacestodes de tipus cisticercoide, i en els vertebrats es formen metacestodes monocèfals (com els cisticercs), o policèfals (com els cenurs o hidàtides), però, en qualsevol cas, l’hoste definitiu s’infesta per ingestió de l’hoste intermediari, i l’escòlex o els protoescòlexs es fixen a la seva mucosa intestinal i inicien el desenvolupament de l’adult. Un cas especial és el del gènere Mesocestoides, l’adult del qual és paràsit de mamífers carnívors o d’ocells rapinyaires, i que segueix un cicle triheteroxè obligat, amb un primer hoste intermediari (àcar copròfag, oribàtid), en el qual es forma un tetratiridi que no s’arriba a desenvolupar totalment fins que un segon hoste intermediari (rèptil, rosegador, insectívor) se’l menja; llavors es desenvolupa el tetratiridi i s’arriba a l’estat metacíclic infestant per a l’hoste definitiu; es tracta, doncs, d’un cicle triheteroxè terrestre que s’assembla força al cicle aquàtic dels pseudofíl·lides (el tetratiridi és l’equivalent del plerocercoide). Una altra excepció es la d’Hymenolepis fraterna, una espècie amb dues races o subespècies (H. fraterna fraterna, paràsit de rosegadors, i H. fraterna nana, de l’home), que té un cicle diheteroxè facultatiu: és l’únic cestode conegut fins ara que pot completar el seu cicle sense passar forçosament per un hoste intermediari (en condicions de laboratori, pot passar també amb H. straminea [=H. microstoma ]); en el seu cicle normal, té com a hoste intermediari un coleòpter —que ingereix el ous eliminats amb les femtes de l’hoste definitiu—, i torna a infestar un altre hoste definitiu quan aquest es menja l’insecte infestat. L’home pot infestar-se si ingereix accidentalment un cuc de la farina (Tenebrio molitor) infestat, per exemple menjant pa poc cuit. Aquest cestode té la capacitat excepcional de poder seguir un cicle monoxè. Els ous disseminats pel sòl i ingerits per l’hoste definitiu alliberen l’oncosfera dins l’intestí d’aquest; aleshores l’oncosfera en penetra les vellositats intestinals i es desenvolupa fins a l’estat de cisticercoide infestant. Més tard, trenca la vellositat intestinal i es deixa caure a l’interior de l’intestí, on fixa el seu escòlex a la mucosa i es desenvolupa com a adult. Els ous que produeix aquest adult surten a l’exterior amb les femtes de l’hoste i així es tanca el cicle. Aquest pot donar-se en els rosegadors o en l’home, i permet que l’hoste s’autoinfesti amb el mateix paràsit, autoinfestació que pot ésser exògena (per contacte amb excrements) o endògena (quan la larva surt de l’ou abans que aquest surti a l’exterior amb els excrements).
Els factors que fan viable el cicle biològic dels eucestodes intervenen, d’una banda, sobre els organismes que intervenen en el cicle (diferents estadis del paràsit i diferents hostes), en el qual cas es tracta de factors biòtics i abiòtics del medi, i, de l’altra, sobre la transmissió, més condicionada per factors de comportament. La transmissió dels cestodes troba moltes dificultats. S’ha calculat que del milió d’ous que produeix diàriament Diphyllobothrium latum, únicament quatre arriben a produir un adult. La fase lliure d’ou, i també la de larva coracidi, tenen contacte directe amb el medi i, per tant, estan sotmesos a canvis sobtats dels factors físics i químics externs, en l’eclosió de l’ou sembla que intervé la llum (activació enzimàtica). Per afavorir la infestació de l’hoste intermediari, els ous solen recórrer a adaptacions morfològiques (semblança amb les preses de l’hoste, per exemple); quan el cicle inclou un segon hoste intermediari, que aquest s’infecti o no depèn, a part el seu comportament, dels factors ambientals (pressió parcial d’oxigen, pH i temperatura) que permeten que la larva es transformi en plerocercoide. La infestació de l’hoste definitiu és afavorida pel canvi de comportament que sol mostrar l’hoste intermediari, provocat per la infestació per part dels metacestodes; un cop dins l’hoste, el desencistament i l’evaginació de l’escòlex depenen de certes reaccions enzimàtiques i de sals biliars i, més tard, de factors del medi intern de l’hoste, com el pH, la pressió parcial d’oxigen, l’augment de la temperatura, les reaccions immunitàries, etc., que permeten que s’arribi a convertir en adult.
Filogènia i sistemàtica
Sobre l’origen i l’evolució dels cestodes s’han desenvolupat diverses teories. Algunes sostenen que els avantpassats s’establiren inicialment en artròpodes i posteriorment passaren als vertebrats. Hom ha pensat en un model ancestral de tipus rabdocel proveït d’apèndix caudal nedador i de ventosa terminal com a adult de vida lliure; en aquest cas, els ous, no operculats, haurien d’ésser ingerits per un artròpode, i l’embrió arribaria al tracte digestiu i posteriorment a l’hemocel, on es convertiria en metacestode. Amb el temps, en ésser ingerits els artròpodes pels vertebrats, els metacestodes es desenvoluparien dins el seu tracte digestiu fins a donar adults paràsits. D’altres teories, en canvi, sostenen que primer van adquirir l’hoste vertebrat i posteriorment l’intermediari; en aquest cas, la larva que sortís de l’ou quedaria lliure dins el tracte digestiu d’un micròfag, i faria servir els ganxos, no per fixar-se, sinó per penetrar a través de la paret intestinal i arribar a l’hemocel. La diferència entre ambdues teories és que la primera postula la procedència dels cestodes a partir de formes de vida lliure, mentre que la segona els fa procedir de monogenis paràsits. Totes dues, però, coincideixen a fer-los derivar de formes amb tub digestiu, cosa que implica una evolució regressiva posterior. Altres teories pensen, contràriament, que els cestodes mai no han tingut aparell digestiu, i que els digenis i els cercomeromorfs (monogenis i cestodes) deriven d’un avantpassat comú primitiu mancat de tub digestiu.
El seguiment de l’evolució dels cestodes es basa en les característiques de l’escòlex i dels seus cicles vitals. Generalment s’accepta que l’avantpassat comú dels cestodes era un protocestode monofosat (amb un sol òrgan de fixació o ventosa apical única), el qual hauria originat tota una branca monozoica, que finalment hauria donat lloc als actuals cestodes monozoics (cariofíl·lides) i a una branca polizoica, la qual, al seu torn, hauria donat lloc als actuals monofosats (estapebòtrids marins i d’aigua dolça, i nippotènids d’aigua dolça), difosats (pseudofíl·lides marins i d’aigua dolça), ditetrafosats (tripanorincs exclusivament marins) i tetrafosats (tetrafíl·lides marins, i proteocefàlides d’aigua dolça i ciclofíl·lides d’aigua dolça i terrestres).
Els hostes definitius més primitius dels cestodes, els elasmobranquis, allotgen paràsits molt diferenciats, i molt específics. Els cestodes dels teleostis marins són molt diferents dels d’aigua dolça. Tots els teleostis, marins o d’aigua dolça, allotgen pseudofíl·lides, però en els d’aigua dolça dominen els ictiotenídids, amb l’escòlex proveït de quatre ventoses però amb anatomia semblant als tetrafíl·lides; és probable que en aquest darrer ordre de cestodes s’hagi originat la línia evolutiva dels paràsits de vertebrats terrestres. Tots els grups de vertebrats terrestres tenen paràsits excepte els cocodrils, cosa que potser té relació amb l’origen marí d’aquest grup: els hostes intermediaris eren absents a les aigües dolces, i es trencà la cadena de transmissió.
Emmarcats dins el fílum dels platihelmints, la seva classificació interna ha sofert molts canvis. Antigament foren considerats com un grup allunyat dels monogenis, mentre que avui hom ha tendit a considerar la classe cestodaris (Cestodaria), al costat dels monogenis, dins la superclasse dels cercomeromorfs (Cercomeromorphae), per separar-los de la superclasse dels trematodes, que inclou els digenis. Alguns autors han proposat la creació del fílum dels cercomeris (Cercomeria), que inclouria trematodes i cercomeromorfs, mentre que d’altres han proposat el fílum cestodes (Cestoda). Hom seguirà aquí la classificació que considera els cestodaris (Cestodaria) com una classe, que inclou dues subclasses, la dels cestodaris (Cestodaria) i la dels eucestodes (Eucestoda). La primera ha inclòs tradicionalment els ordres dels amfilínides (Amphilinida) i els girocotílides (Gyrocotylida). Per a la subdivisió dels eucestodes hom considerarà la revisió feta recentment (1985) per G.D. Schmidt, que estableix 12 ordres: els cariofíl·lides (Caryophyllida), paràsits de teleostis i anèl·lids aquàtics; els espatebòtrides (Spathebothriida), de teleostis; els tripanorinquis (Trypanorhyncha), d’elasmobranquis; els pseudofíl·lides (Pseudophyllida), de peixos, amfibis, rèptils, ocells i mamífers; els lecanicefàlides (Lecanicephalida), d’elasmobranquis; els aporideus (Aporidea), d’anseriformes, els tetrafíl·lides (Tetraphyllida), els difíl·lides (Diphyllida) i els litobòtrides (Litobothrida), tots tres d’elasmobranquis; els proteocefalats (Proteocephalata), de peixos, amfibis i rèptils; els ciclofíl·lides (Cyclophyllida), d’amfibis, rèptils, ocells i mamífers; i els nippotènides (Nippotaeniida), de teleostis.
Principals representants
Moltes de les espècies d’eucestodes tenen importància en el camp de la veterinària, ja que infesten animals domèstics, i d’altres en tenen en el camp mèdic, ja que infesten l’home. L’home només té dues espècies de cestodes pròpies i específiques: l’anomenada tènia de l’home (Taenia solium), que se serveix del porc com a hoste intermediari, i la tènia anomenada tènia del bou (T. saginata), perquè aquest és el seu hoste intermediari. Tota la resta de tèniasis o cestodiasis humanes s’han de considerar zoonosis: Diphyllobothrium latum i espècies pròximes (que infesten carnívors depredadors de peixos), Hymenolepis fraterna nana i H. diminuta (que parasiten rosegadors), Dipylidium caninum (paràsit del gos i el gat), diverses espècies de Raillietina (paràsites de rosegadors), Bertiella studeri (de primats), Inermicapsifer madagascariensis (de rosegadors), poden parasitar l’home quan són en fase d’adult. D’altres el poden parasitar en fase de metacestode: diverses espècies de Spirometra, les larves de la qual li produeixen la denominada esparganosi; la tènia de l’home (Taenia solium), els cisticercs de la qual produeixen la cisticercosi, l’anomenada tènia hidatídica (Echinococcus granulosus), els hidàtides o quists de la qual produeixen l’equinococcosi, o E. multilocularis, els hidàtides de la qual causen l’alveococcosi o equinococcosi multilocular.