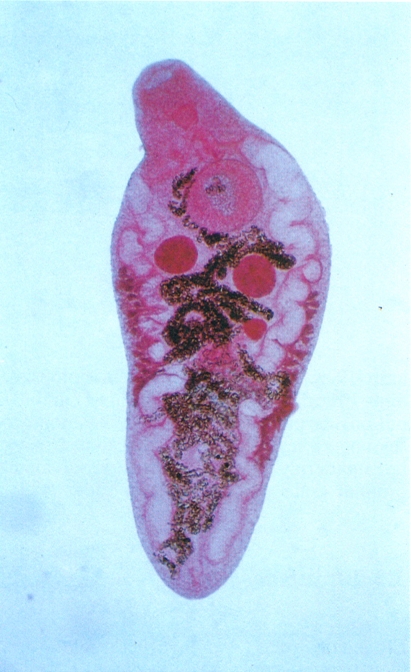
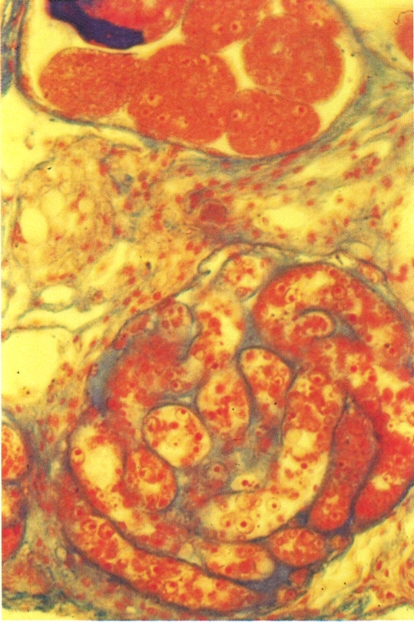
Els trematodes digenis són tots paràsits. Hi ha espècies que són conegudes des de l’antigor, pel fet de parasitar l’home i produir-li greus malalties; és el cas per exemple de l’esquistosomiasi, per exemple. D’altres tenen una gran incidència en els animals, ja que parasiten animals domèstics i produeixen grans pèrdues econòmiques; no així l’espècie de la fotografia, Platynosomum soricis, un cuc pla que parasita les musaranyes del nostre país. Els digenis es troben gairebé a tot el món (continents, mars i oceans). La distribució d’una espècie concreta depèn de l’especificitat per al primer hoste intermediari i de l’existència de les condicions ecològiques necessàries perquè sigui possible el cicle biològic.
Santiago Mas-Coma.
La classe dels trematodes comprèn platihelmints que viuen paràsits de tota mena de vertebrats. Hom la divideix en dos grans grups, de categoria taxonòmica discutida: els digenis i els aspidogastris. Els digenis constitueixen el grup més nombrós i són els cucs paràsits més comuns i abundants després dels nematodes, com també els trematodes més estudiats, pel seu gran interès mèdic, veterinari i econòmic.
La transcendència econòmica, sanitària i mèdica dels trematodes rau en el fet que diverses espècies de digenis arriben a parasitar l’home i els animals domèstics i silvestres explotats per l’home, i són agents causants de malalties. Algunes són les productores de les distomatosis humanes. Segons dades de l’Organització Mundial de la Salut, entre 1975 i 1981, el nombre de persones afectades d’esquistosomiasi era de 200 milions, i entre mig milió i un milió les morts anuals; la incidència en l’home es reparteix entre les espècies següents: uns 100 milions de casos d’esquistosomiasi vesicular (Schistosoma haematobium); uns 60 milions d’esquistosomiasi intestinal (S. mansoni); i uns 100 milions d’esquistosomiasi japonesa (S. japonicum). Els segueix en importància la clonorquiasi i l’opistorquiasi, amb 19 milions d’afectats; la fasciolopsiasi, amb 10 milions; i la paragonimiasi, amb 3,2 milions de parasitats. Són molt més nombroses encara les espècies que incideixen sobre la ramaderia, algunes de les quals arriben a originar grans pèrdues econòmiques.
Els aspidogastris
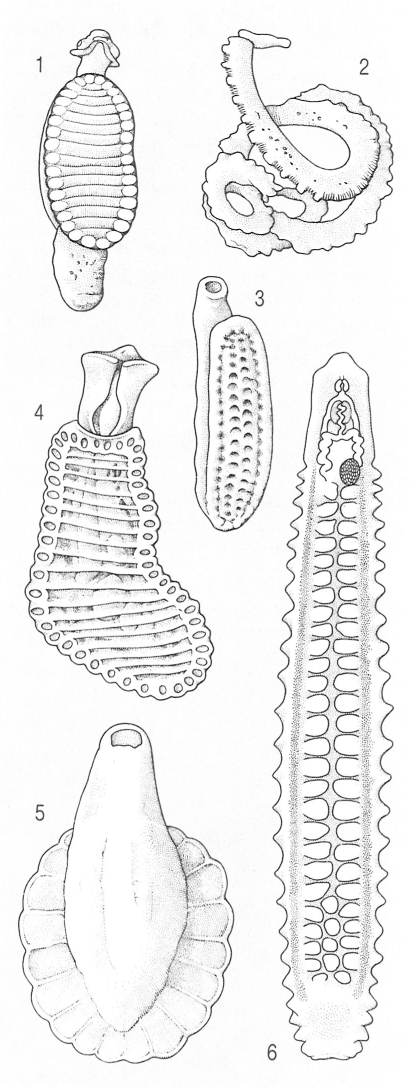
Tipus morfològics generals dels aspidogastris, segons les característiques de la ventosa ventral, que pot ésser grossa i dividida en alvèols, dividida en una filera de ventoses longitudinals o feta de plecs transversals. Hom ha representat en el dibuix: 1 Lobatostomum ringens, 2 Stichocotyle cristata, 3 Lophotaspis vallei, 4 Cotylogaster michaelis, 5 Cotylaspis insignis, 6 Rugogaster hydrolagi.
Biopunt, a partir de fonts diverses
Els aspidogastris (Aspidogastrea), també anomenats aspidocotilis (Aspidocotylea) o aspidobotris (Aspidobothria), comprenen un nombre reduït d’espècies, menys estudiades a causa de la seva escassa importància econòmica i sanitària. Es tracta de trematodes que han establert, en la majoria dels casos, relacions parasitàries vagues amb mol·luscs, si bé alguns són paràsits facultatius o obligats de peixos o tortugues. Presenten tres tipus anatòmies bàsics. El primer és el dels aspidogàstrides (Aspidogastrida), que tenen una ventosa ventral immensa, dividida en alvèols per parets septals musculoses longitudinals i transversals, ventosa que usualment porta cossos marginals o tentacles curts entre els alvèols. En els esticocotílides (Stichocotylida), en canvi, s’observa una sèrie longitudinal de ventoses individuals. En els rugogàstrides (Rugogastrida), l’òrgan ventral de fixació és constituït per plecs transversals. En tots ells, el tegument i els diferents sistemes anatòmies són similars als dels digenis. La majoria d’espècies són de cicle directe. Sembla que només les espècies paràsites de vertebrats requereixen un hoste intermediari, ja que no es coneix cap cas en què la larva de vida lliure que emergeix de l’ou, anomenada cotilocidi (cotylocidium), sigui infestant per als vertebrats. Els adults són molt poc específics, de manera que és habitual, per exemple, de trobar, a l’intestí d’alguns peixos, espècies que normalment són paràsites dels mol·luscs. D’altres, en canvi, sembla que no poden madurar en mol·luscs i que necessiten un peix hoste definitiu.
Els digenis
Morfologia
Els trematodes digenis (Digenea) adults presenten una gran varietat de formes. La majoria són plans i d’aspecte foliaci, si bé també n’hi ha de piriformes, cònics, cilíndrics, etc. Les seves dimensions oscil·len entre uns quants microns (200 µm) i diversos centímetres (7 cm, en Fasciolopsis buski); els didimozoides constitueixen una excepció, ja que poden arribar a fer 15 m. Com a òrgans de fixació presenten habitualment dues ventoses: l’oral, anomenada també bucal o apical, situada a la part anterior i en el fons de la qual s’obre gairebé sempre la boca (rarament absent); i la ventral, o acetàbul, a la zona ventral mitjana i més o menys allunyada de la primera (absent en determinats grups). Segons això, hom distingeix diferents tipus morfològics, amb denominacions que actualment no tenen validesa taxonòmica: d’una banda, els prosostomes (boca a la ventosa apical), que inclouen els distomes (dues ventoses pròximes), amfistomes (acetàbul terminal), monostomes (sense acetàbul), equinostomes (dues ventoses, l’anterior amb collar d’espines peribucal), holostomes (dues ventoses acompanyades d’un òrgan ventral o òrgan tribocític, de funció adhesiva i glandular) i esquistosomes (dioics) i, d’altra banda, els gasterostomes (boca a l’acetàbul).
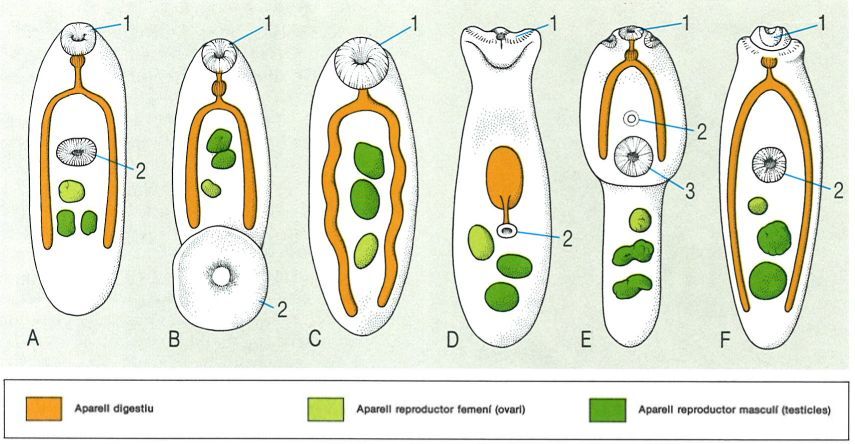
Hom ha descrit uns tipus morfològics generals de digenis monoics, segons la posició de la boca i de la ventosa apical, si bé actualment aquests tipus no tenen validesa taxonòmica. El dibuix en mostra els principals, com també la disposició general dels aparells digestiu i reproductor: A distoma, amb les dues ventoses, bucal o apical (1) i ventral o acetàbul (2) pròximes; B amfistoma, amb la ventosa ventral al final del cos; C monostoma, sense ventosa ventral; D gasterostoma, amb la boca situada en la ventosa ventral; E holostoma, amb un òrgan tribocític (3) acompanyant les dues ventoses; F equinostoma, amb la ventosa bucal proveïda d’espines i la ventral normal. Hi ha encara el cas de l’esquistosoma, que és dioic.
Biopunt, a partir de fonts diverses
La paret del cos és un tegument metabòlicament actiu, semblant al dels monogenis i els cestodes, constituït per un síncit amb els nuclis situats sota d’una capa muscular superficial i connectats, mitjançant expansions, al citoplasma distal extern, anucleat i continu; superficialment, la paret pot ésser llisa o lluir una ornamentació de diferent mena, a base d’espines, protuberàncies, rugositats, forats, etc. Sembla que les funcions d’aquest tegument són l’absorció de nutrients i la síntesi i la secreció de materials protectors. Cal destacar l’existència d’abundants papil·les sensorials en el tegument, el nombre i la distribució de les quals (quetotàxia) són d’un gran interès taxonòmic, sobretot pel que fa a les fases larvals. Les capes musculars superficials són de disposició circular, longitudinal i diagonal, i permeten notables canvis de la forma del cos. El cos és reblert de teixit connectiu parenquimàtic, que fa una funció mecànica, esquelètica, protectora i metabòlica; aquest teixit envolta els òrgans interns.
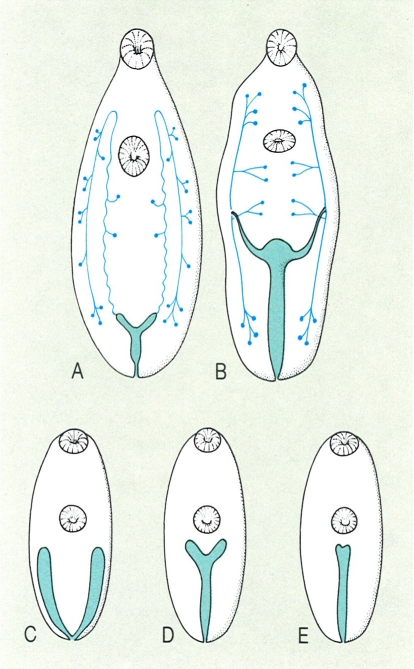
Aparell excretor dels digenis. A dalt, diferents models de distribució dels conductes excretors i dels solenòcits (en blau ple) en el cas dels gèneres Opisthorchis (A) i Dicrocoelium (B). A baix, diferents tipus de vesícules excretores (en blau tramat): en forma de lletra V (C). de lletra Y (D) i de I (E).
Biopunt, a partir de fonts diverses
Els trematodes digenis no tenen aparell respiratori ni circulatori. L’aparell digestiu comprèn una boca (generalment al fons de la ventosa oral, i rarament de l’acetàbul), una prefaringe (de vegades absent), una faringe (bulb esofàgic musculós apte per a la succió), un esòfag (de vegades absent) i dos cecs intestinals (de llargada variable i senzills en el cas de Dicrocoelium, ramificats en Fasciola o fusionats posteriorment en Schistosoma), i, solament en algunes famílies (opecoèlids), un orifici anal. El sistema nerviós és d’estructura semblant al dels turbel·laris, amb una massa ganglionar periesofàgica, de la qual surten els plexes nerviosos longitudinals que innerven tot el cos (dues fileres ventrals, dues de dorsals i dues de laterals), units per comissures transversals. L’aparell excretor és un sistema protonefridial ben desenvolupat, igual que el de tots els platihelmints. Els protonefridis, també anomenats cèl·lules flamígeres o solenòcits, són cèl·lules en forma de matràs, amb les parets filtrants i amb cilis vibràtils a l’interior, que amb el seu moviment («flama vibràtil») donen la força motora perquè flueixi el sistema. Aquests solenòcits (el nombre i disposició dels quals té un gran interès sistemàtic sobretot pel que fa a les larves) es troben repartits pel mesènquima, i són connectats per petits túbuls col·lectors que desemboquen en dos col·lectors principals laterals que, al seu torn, desemboquen en una vesícula excretora (en forma de Y, V i I, i d’una gran importància taxonòmica); aquesta s’obre a l’exterior mitjançant un únic porus excretor (de vegades doble, en estadis larvals), en general terminal. Segons el trajecte dels dos canals col·lectors principals es poden distingir dos grans tipus d’aparells excretors: el tipus mesostoma, en el qual els dos canals, un cop rebuts els secundaris (prop de l’acetàbul), es dirigeixen directament a la vesícula; i el tipus estenostoma, en el qual, en canvi, els dos principals, després de rebre els secundaris, pugen primer fins a l’altura de la faringe i, fent un bucle, baixen fins a la vesícula. L’embriogènesi de la vesícula excretora té un gran interès sistemàtic perquè constitueix la base de la distinció entre els dos grans i únics grups (superordres) de digenis: durant la formació de l’estadi larval de cercària (cercariogènesi), en el cas dels anepiteliocistidis (Anepitheliocystidia) es conserva la vesícula excretora primitiva, que resulta de la fusió dels dos col·lectors principals, sense que entrin en la seva constitució cèl·lules epitelials, mentre que en els epiteliocistidis (Epitheliocystidia) la vesícula primària és envoltada, i després substituïda, per cèl·lules d’origen mesodèrmic i la vesícula definitiva té una paret epitelial espessa.
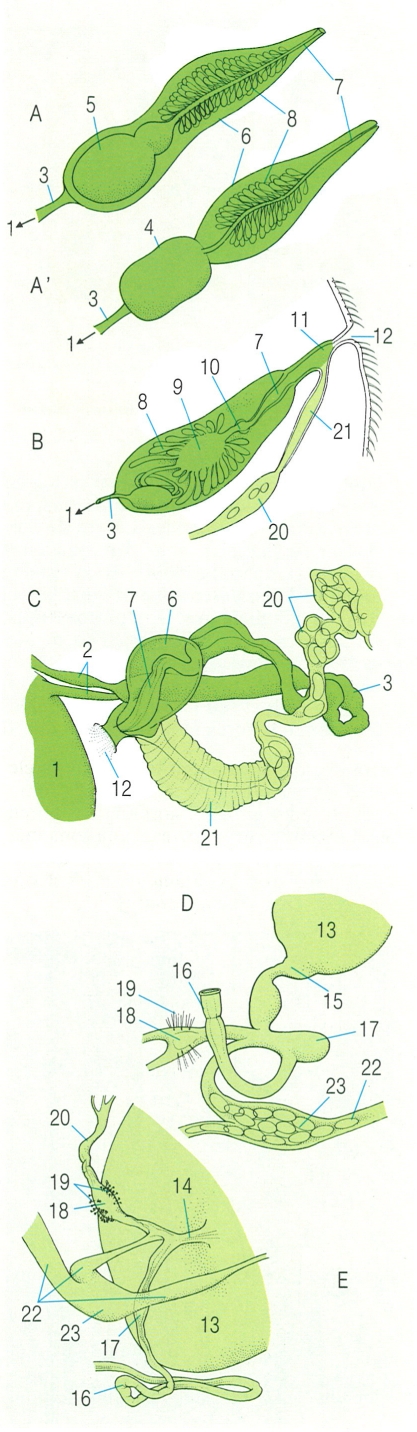
Aparell reproductor terminal dels digenis. A dalt, diferents òrgans i conductes genitals terminals: A, A’ estructures masculines en els casos de vesícula seminal interna o externa, respectivament; B esquema de l’estructura interior en un tall que passa per l’atri genital i la bossa del cirrus en Pleurogenes medians; C visió ventral de les estructures genitals terminals i el gonòpor en Scaphiostomum palaearcticum. A baix, aparell productor dels ous (oogenòtop) en Opisthioglyphe ranae (D) i en Scaphiostomum palaearcticum (E). 1 Testicle, 2 conductes eferents, 3 conducte deferent, 4 vesícula seminal externa, 5 vesícula seminal interna, 6 bossa del cirrus, 7 cirrus, 8 glàndules prostàtiques, 9 pars prostàtica, 10 canal ejaculador, 11 atri genital, 12 porus genital, 13 ovari, 14 oviducte, 15 oviscapte, 16 canal de Laurer, 17 receptacle seminal, 18 ootip, 19 glàndula de Mehlis, 20 úter, 21 metraterma, 22 vitel·loducte, 23 magatzem de vitel.
Biopunt, a partir de T.C. Cheng, J.G. Baer, Ch. Joyeux i S. Mas-Coma.
La majoria dels digenis adults són hermafrodites, si bé alguns són dioics, és a dir, amb els sexes separats, com en els esquistosomàtides. L’aparell genital masculí comprèn generalment dos testicles, més rarament un (monòrquides), i a vegades més de dos o fins i tot diverses dotzenes (esquistosomàtides), la posició dels quals respecte de l’ovari és d’un gran interès sistemàtic. La seva forma pot ésser des d’arrodonida fins a ramificada, segons les espècies. De cada testicle surt un conducte deferent, i la seva unió dona lloc al conducte deferent o espermiducte, que va a parar generalment a la bossa del cirrus, molt musculosa; hi ha una vesícula seminal interna i un conducte ejaculador que s’estén fins al final de la bossa i dona lloc a l’òrgan copulador masculí o cirrus, evaginable, llis o armat d’espines. El conducte ejaculador es troba generalment envoltat de nombroses cèl·lules glandulars prostàtiques, i pot mostrar una dilatació anomenada pars prostàtica. La bossa del cirrus desemboca al porus genital masculí, pròxim al femení, en un atri genital comú. Hi ha moltes variacions en aquestes estructures segons els grups taxonòmics. L’aparell genital femení consta d’un sol ovari, de forma (generalment arrodonida, rarament lobulada o ramificada) i localització diferents segons les espècies. De l’ovari surt un curt oviducte, que comunica lateralment amb el receptacle seminal i amb l’anomenat canal de Laurer; aquest, que acaba cec en el parènquima o s’obre dorsalment a través del tegument, sembla que és un vestigi de vagina no funcional; posteriorment l’oviducte rep el conducte vitel·lí comú, que porta els productes recollits pels vitel·loductes, procedents dels dos (en general) camps de fol·licles o glàndules vitel·lines. L’oviducte s’eixampla posteriorment per formar l’ootip, estructura al voltant de la qual hi ha nombroses glàndules unicel·lulars anomenades en conjunt glàndula de Mehlis; les cèl·lules vitel·lines són les precursores de la coberta de l’ou, mentre que la funció de la glàndula de Mehlis està relacionada amb la formació d’una membrana al voltant de la massa cel·lular ovífero-vitel·lina. El complex estructural que hem descrit, incloent-hi el principi de l’úter, rep el nom de oogenòtop o aparell formador d’ous. A continuació de l’ootip pot haver-hi encara, en algunes espècies, un curt receptacle seminal uterí, emmagatzemador d’espermatozoides, a més de l’úter pròpiament dit, que s’allarga més o menys segons les espècies i desemboca en el porus genital femení. L’última part de l’úter és sovint musculosa i exerceix la funció d’ovojector i de vagina, i rep el nom de metraterma.
Biologia i ecologia
Reproducció i desenvolupament

Hom anomena esquistosoma el tipus de digeni dioc, és a dir, aquell del qual hi ha individus mascles i individus femelles. L’espècie Schistosoma mansoni n’és un exemple ben conegut a causa de la seva incidència en les comunitats humanes, com a agent de l’esquistosomiasi intestinal. En la fotografia podem veure’n la femella, prima i cilíndrica, allotjada en l’interior del canal ginecòfor del mascle, més gros, on es realitza la còpula (tots dins el cos de l’hoste definitiu, en aquest cas, l’home).
Santiago Mas-Coma.
En els trematodes digenis és general que es doni el fenomen de la proteràndria. En el cas de les espècies hermafrodites sembla que la reproducció creuada de dos individus és la norma general; tanmateix, en absència de cap altre individu de la mateixa espècie dins l’hoste, recorren a l’autofecundació. Tant en un cas com en l’altre, l’esperma es vessa a l’úter després de la introducció del cirrus; en espècies sense cirrus, l’atri genital es tanca per contracció muscular i, en els casos excepcionals d’espècies amb individus unisexuals i desproveïts de cirrus, com els esquistosomàtides, la còpula té lloc quan la femella s’allotja en una canal ventral longitudinal del mascle (canal ginecòfor). De tota manera, els espermatozoides pugen per l’úter i s’emmagatzemen en el receptacle seminal.
La reproducció sexual té lloc dins l’hoste definitiu. Tanmateix, hi ha espècies progenètiques els òrgans genitals de les quals són capaços de madurar precoçment i produir ous en estadis anteriors a l’adult (metacercària o estadi fisiològicament equivalent) i, per tant, encara en l’hoste intermediari. També s’han citat processos partenogenètics en espècies hermafrodites, tant en adults com en metacercàries progenètiques, si bé els coneixements en aquest sentit encara són insuficients.
Els estadis larvals
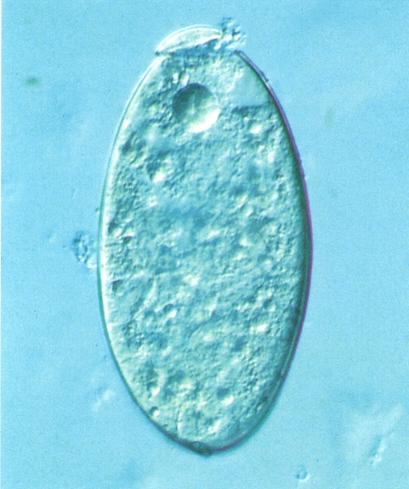
Ou operculat i sense embrionar del distoma hepàtic (Fasciola hepatica), vist al microscopi. En els esquistosomes, els ous manquen d’opercle.
Santiago Mas-Coma.
Els ous són generalment el·lipsoïdals, de color marronós clar o fosc, amb una gran variabilitat de forma, dimensions, grossària de la coberta i color. La majoria tenen un opercle en un dels seus pols, per on sortirà l’embrió o miracidi; però els esquistosomàtides no en tenen, i l’embrió surt a través d’una zona més prima de la coberta de l’ou. De vegades els ous presenten apèndixs filamentosos (notocotílides) o esperons (Schistosoma).
El miracidi és la primera forma larval del cicle d’un digeni, i per tant apareix directament del desenvolupament de l’ou. Per metamorfosi es converteix en el següent estadi larval, un esporocist dins l’hoste intermediari, el qual al seu torn es transformarà en alguns casos en rèdia, i aquesta, en cercària. Aquestes fases larvals desenvolupen diverses generacions per reproducció asexual dins l’hoste intermediari, però, finalment, la cercària surt a l’exterior i cerca el seguent l’hoste.
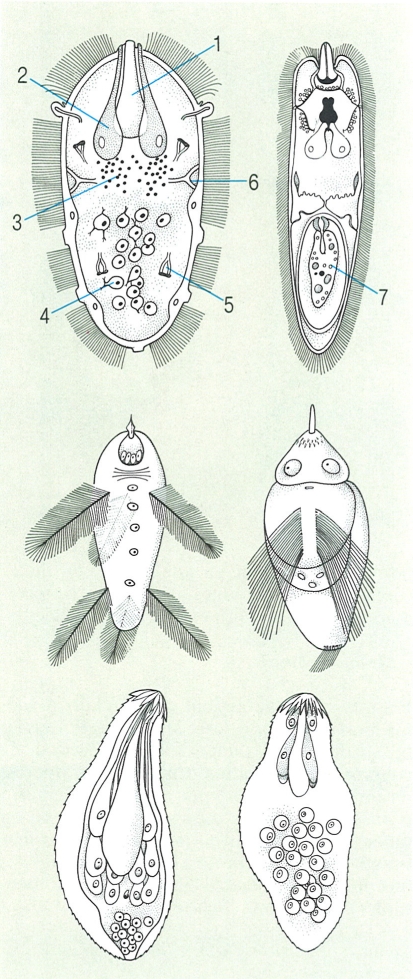
Tipus de miracidis en els trematodes digenis. A dalt, miracidis amb cilis nombrosos distribuïts per tota la superfície externa; al mig, miracidis amb arestes ciliades i amb cilis agrupats en plomalls; a baix, miracidis desproveïts de cilis, recoberts d’espines i amb un aparell glandular voluminós. Hom hi ha indicat: 1 glàndula apical, 2 glàndula de penetració, 3 cervell, 4 cèl·lula germinal, 5 cèl·lula flamígera, 6 nervi lateral, 1 rèdia jove (cas especial de Parorchis acanthus.
Biopunt, a partir de fonts diverses
El miracidi acabat de sortir de l’ou generalment és piriforme i recobert de 4 o 5 séries transversals de plaques epidèrmiques planes, ciliades o no, de disposició, nombre i forma propis de cada espècie. Aquestes plaques són interrompudes per expansions de cèl·lules subepitelials; per sota de l’epiteli hi ha fibres musculars circulars i longitudinals. A l’extrem anterior hi ha una papil·la apical o terebratori, no ciliada, sovint retràctil, que presenta moites terminacions sensorials; hi desemboquen els conductes de les glàndules de penetraci ó i de la glàndula apical; en algunes espècies hi ha també un estilet apical o espines. El sistema excretor és bilateral, amb un o dos pareils de cèl·lules flamígeres i dos porus excretors laterals. El sistema nerviós consta d’una massa ganglionar cérébral anterior, de la qual surten plexes nerviosos que innerven la musculatura i les terminacions sensorials; generalment hi ha un parell de taques oculars o fotoceptors pigmentats, papil·les sensorials de diferents tipus (de nombre i distribució molt constants en cada espècie) i dues papil·les laterals localitzades en els soles transversals. La meitat superior del miracidi conté cèl·lules germinals que seran transportades a l’estadi larval següent, l’esporocist. En tots els casos el miracidi fa una metamorfosi dins el primer hoste intermediari, i origina un esporocist. Un cas particular és el de les espècies de Philophthalmus, els miracidis de les quals inclouen ja un esporocist larval diminut. També són excepcionals les espècies Parorchis acanthus i Typhlocoelium cymbium, el miracidi de les quals conté ja al seu interior un estadi larval distint, anomenat rèdia, que és alliberat quan entra a l’hoste. Hi ha també d’altres casos en què l’estadi subsegüent del miracidi és el de rèdia i, per tant, no passen per la fase d’esporocist (Stichorchis, Caecincola).
L’esporocist no és més que un sac germinal sense boca ni sistema digestiu. La paret és constituïda per un tegument sineític extern actiu (la nutrició és transtegumentària), una capa muscular de fibres longitudinals i circulars (tenen una certa mobilitat i capacitat de variar la forma) i un epiteli germinal intern. El sistema excretor és ben desenvolupat, amb un nombre de cèl·lules flamígeres que s’incrementa amb l’edat de l’esporocist. Les cèl·lules germinals de l’epiteli intern es van desprenent successivament de la paret interior del sac esporocístic i es diferencien fins a donar lloc al següent estadi larval, que és una altra generació d’esporocists (esporocists fills, per a diferenciar-los de l’esporocist de primera generació o esporocist mare) o fins a donar una rèdia.
Els esporocists fills són també sacs germinals, simples (ovoïdals o allargats) o ramificats. Les cèl·lules germinals de la paret donen lloc al següent estadi larval de cercària en un procès anomenat cercariogènesi. Així, es parla en aquests casos d’esporocists cercariógens, per diferenciar-los de l’esporocist mare o esporocistogen. En alguns casos, però, com ara en Schistosoma, en comptes de cercàries, s’originen generacions subsegüents d’esporocists (esporocists replicants bipotencials, és a dir, capaços de produir tant cercàries com nous esporocists). En molts casos, els esporocists presenten porus de naixement o orificis de posta per a deixar sortir els estadis interns, que de vegades són precedits d’una cambra d’ejecció per a l’expulsió de les cercàries (braquilàimids); en altres casos no hi ha porus, i els estadis interns surten trencant la paret de l’esporocist; i en d’altres, les cercàries es transformen directament, dins l’esporocist, en el següent estadi larval de metacercària enquistada (leucoclorídids). Hom pot afegir, finalment, que també s’ha descrit algun cas en què els esporocists (mare i fills) són productors de miracidis.
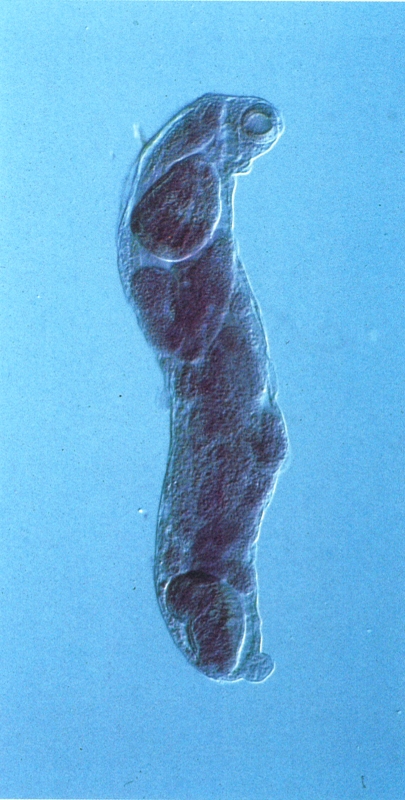
Rèdia cercariògena, és a dir, formadora de cercàries, de trematode digeni. Noteu la voluminosa faringe, a l’extrem anterior, i la presència a l’interior del cos de petites cercàries en diferents estadis de desenvolupament.
Santiago Mas-Coma.
La rèdia és també un sac germinal més o menys allargat, el·lipsoïdal o cilíndric, d’extrem posterior generalment rom, de vegades amb una o dues prolongacions o apèndixs (lòbuls) a la meitat posterior i un collar musculós a prop de l’extrem anterior, que els facilita la locomoció a través dels teixits de l’hoste. Presenta un aparell digestiu característic, rudimentari però funcional, amb una boca (sense ventosa), una faringe musculosa i un cec intestinal curt o llarg, no ramificat. La paret del sac redial és igual que la de l’esporocist (la nutrició també és transtegumentària), però en algunes espècies s’hi ha detectat la presència de papil·les sensorials. En el sistema excretor, les cèl·lules flamígeres desemboquen en dos troncs principals que s’obren a l’exterior mitjançant els respectius porus laterals. Les cèl·lules germinals de la paret cauen dins el sac redial i donen rèdies de segona generació (idèntiques a la rèdia mare) o cercàries, segons els casos, que surten a través d’un porus de naixement generalment situat prop de la faringe. Sembla que la majoria d’espècies que presenten rèdies tenen una única generació de rèdia unipotencial, ja cercariogènica, o bé dues generacions de rèdies, amb una primera d’unipotencial (rèdia mare rediogènica) i una segona, també unipotencial (rèdies filles cercariogèniques). No obstant això, en el cas de Fasciola hepatica, hi poden haver fins un màxim de quatre generacions de rèdies, i les tres primeres generacions són totes bipotencials (productores tant de rèdies com de cercàries). En el cas dels equinostomàtides, el nombre de generacions redials pot ésser il·limitat.
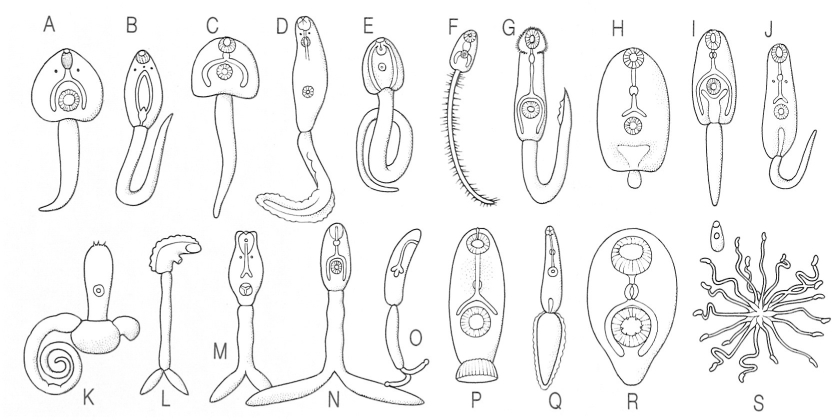
Diferents tipus de cercàries de trematodes digenis. A amfistoma, B monostoma, C cercària glmnocèfala, D cercària gimnocèfala de tipus pleurolofocerca, E cistòfora, F tricocerca, G equinostoma, H microcerca, I xifidiocerca, J oftalmoxifidiocerca, K furcocercària del tipus gasterostoma, L furcocercària del tipus lofocerca, M furcocercària afaríngia, N furcocercària amb faringe, O furcocercària monostoma sense faringe ni ventosa oral, P cotilocerca, Q ropalocerca, R cercària, S cercària del tipus «Rettenkönig».
Biopunt, original d’O.W. Olsen.
La cercària representa, en la majoria dels trematodes digenis, el final de la seqüència de formes larvals lligades per metamorfosi i multiplicació asexual regenerativa que es fa dins el primer hoste intermediari. La cercariogènesi té lloc a l’interior dels esporocists o les rèdies i és un procés complex que parteix d’una única cèl·lula germinal i culmina amb la formació de la cercària completament desenvolupada, si bé en alguns casos les cercàries acaben de madurar, un cop fora de l’esporcist o la rèdia, en els teixits del primer hoste intermediari. La cercària madura constitueix ja un estadi juvenil de l’adult. Presenta en general un cos globular, cilíndric o oval, i una cua, de llargada i forma diferents, que de vegades pot no ésser-hi. En les primeres fases de la cercariogènesi, la paret és un epiteli primitiu que degenera quan, per sota, es forma el que serà ja el tegument definitiu de la cercària. Aquest tegument definitiu és un síncit citoplasmàtic, envoltat externament d’una fina membrana plasmàtica suau i llisa i, per sobre, d’una capa muscular (fibres circulars externes i longitudinals internes), que recorda el tegument de l’adult que, de vegades és espinós. El cos presenta una ventosa oral proveïda d’un estilet protràctil amb base musculada (xifidiocercàries) o armada d’espines corbes (furcocercàries), i una ventosa ventral de diferent localització, que pot faltar en algunes espècies. L’aparell digestiu consta d’una boca, generalment situada a la ventosa oral, rarament a la ventral (gasterostomes), que és seguida d’una prefaringe, una faringe, l’esòfag i finalment l’intestí, generalment bifurcat en dues branques cegues simples (fins i tot en aquelles espècies en què aquestes són ramificades en l’adult, com en Fasciola hepatica). Una peculiaritat de l’estadi de cercària és la presència general de cèl·lules glandulars en el cos, la naturalesa de les quals es relaciona amb els diferents tipus de desenvolupament i comportament (enquistament, migració intraorgànica, penetració activa en el següent hoste, etc.); les cercàries dels azígids i els bivesicúlids són les úniques que no presenten cap mena de glàndules (són ingerides directament per l’hoste). Les anomenades glàndules de penetració, de funció encara no del tot aclarida, són constituïdes per entre 2 i 8 parells de cèl·lules proveïdes de conductes que s’obren a l’exterior a través de porus anteriors; comprenen les anomenades glàndules d’escapada, el contingut de les quals es vessa durant la migració de sortida de la cercària en el primer hoste intermediari, les glàndules apicals, de funció posterior a la penetració, les glàndules postacetabulars, importants per a l’adherència a superfícies i amb d’altres funcions, i les glàndules preacetabulars, útils en la penetració. Les glàndules cistògenes són presents al subtegument de totes les cercàries que s’enquisten (en el medi extern o en el segon hoste intermediari) i secreten les múltiples capes del quist que embolcalla el següent estadi larval de metacercària. Un tercer tipus són les conegudes com a glàndules mucoidees, localitzades a la regió mitjana ventral d’algunes cercàries i de funció encara desconeguda, bé que hom els ha atribuït les de protecció, lubrificació, etc. El sistema excretor de la cercària madura és idèntic que el de l’adult, però molt més simple, amb menys cèl·lules flamígeres, però amb un nombre i distribució constants per a cada espècie; en algunes, la vesícula excretora s’obre a l’exterior a través d’un o dos porus a prop de l’apèndix caudal. El sistema nerviós també és del mateix tipus que el de l’adult. Cal destacar la presència de generalment dos, rarament tres, taques oculars (notocotílids, opistòrquids) i també de papil·les sensorials tegumentàries, el nombre i distribució de les quals (quetotàxia cercàriana) són característics per a cada espècie i permeten fins i tot de distingir les cercàries d’espècies que tenen els adults morfològicament idèntics. Generalment, a la regió postacetabular es distingeix una agrupació de cèl·lules que representa l’esbós dels futurs òrgans sexuals (primordi genital).
La mesocercària constitueix el següent estadi larval en alguns grups com els estrigeoïdeus (com per exemple Alaria) i els hemiuroïdeus (com per exemple Halipegus), en els cicles dels quals intervenen tres hostes intermediaris. S’origina en el segon hoste intermediari i és morfològicament intermèdia entre la cercària (primer hoste intermediari) i la metacercària (tercer hoste intermediari). Estructuralment és com una cercària més gran, amb tegument, espinulació i ventoses similars, amb cecs intestinals una mica més desenvolupats i un aparell excretor més complex, però encara amb les glàndules de penetració i el primordi genital sense desenvolupar.
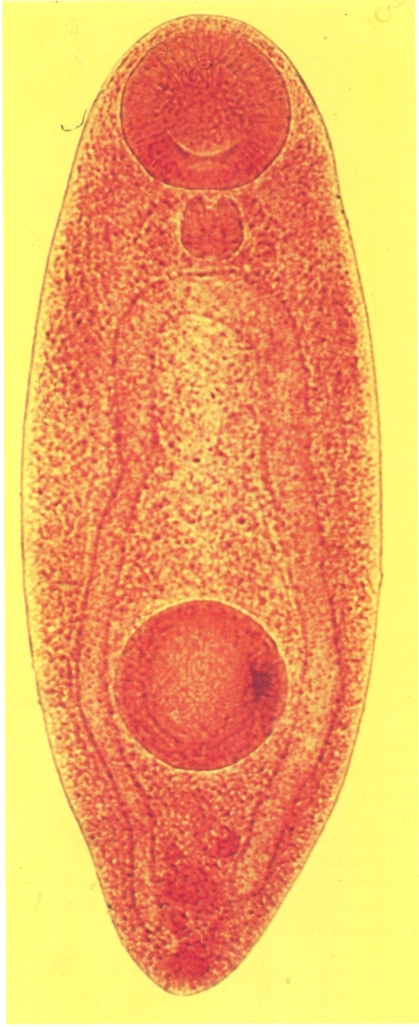
Metacercària de Dollfusinus frontalis, una espècie que passa un cicle vital complex, pel qual parasita successivament tres hostes diferents, els dos primers cargols terrestres i el definitiu un micromamífer. Hom ha estudiat el seu cicle a Formentera, on parasita com a hoste definitiu l’eriçó (els sins frontals) o la rata cellarda (les fosses nasals); de l’ou, en surt el miracidi, un cop dins el cargol terrestre Trochoidea caroli; el miracidi dona lloc a dues generacions d’esporocists; posteriorment s’esdevé l’emissió de cercàries, les quals abandonen el cos del primer cargol i penetren al segon hoste intermediari, Cernuella virgata, per infestació activa, a través de l’orifici excretor del cargol, i es traslladen fins a la cavitat pericàrdica, on es transformen en metacercàries com la de la fotografia. Les metacercàries del cargol s’hi mantenen fins que el cargol és ingerit per un eriçó o una rata cellarda.
Santiago Mas-Coma.
La metacercària és l’estadi larval que segueix al de la cercària en la majoria dels trematodes digenis, i les excepcions són els casos rars amb mesocercàries, ja esmentats, i aquells en què les cercàries infesten directament l’hoste definitiu i donen lloc a l’adult (esquistosomatoïdeus). En unes espècies, la metacercària és lliure i viu enquistada en el medi extern, mentre que en d’altres és paràsita (enquistada o no), en general del segon hoste intermediari (del tercer en els estrigeoïdeus i els hemiuroïdeus, amb presència obligada o facultativa de mesocercàries en un segon hoste intermediari). Constitueix una forma de transició entre cercària i adult. Es distingeix de la cercària per la pèrdua de certs òrgans larvals, que deixen d’ésser funcionals o útils (la cua que servia de mitjà de locomoció, l’estilet, les glàndules de penetració, etc.), i també pel desenvolupament d’altres òrgans, necessaris per al posterior estadi d’adult (sobretot l’aparell genital). El tegument cercària persisteix, almenys en aquelles espècies que no s’enquisten. Les modificacions de l’aparell digestiu afecten només l’extensió dels cecs intestinals, que tenen generalment ja les característiques de l’adult. El sistema excretor en uns casos no es modifica gens respecte del de la cercària (fasciòlids, equinostomàtides, microfàl·lids, al·locreàdids), i en d’altres, mostra un lleuger augment del nombre de cèl·lules flamígeres (Opisthorchis); en d’altres, encara, té un considerable desenvolupament, i apareixen solenòcits addicionals i nous conductes excretors (estrigeoïdeus). Pel que fa al sistema nerviós, cal destacar la reducció o total desaparició de les papil·les sensorials en les metacercàries que s’enquisten i, en canvi, l’increment notable en les que no s’enquisten, com en el cas dels braquilàimids. L’aparell genital adquireix una importància preponderant, però en grau variable segons les espècies, des d’un simple esbós de les futures estructures sexuals de l’adult (variació escassa respecte de la cercària), fins a estructures ja completament conformades, encara que no funcionals (per tant, sense producció d’ous, excepte en metacercàries progenètiques), passant per casos de desenvolupament intermedi. Els estrigèids són els que presenten metacercàries més evolucionades, amb una complexitat molt pròxima a la de l’adult. Pel que fa a les metacercàries enquistades, es pot distingir entre les de vida lliure i les paràsites. L’estructura de la paret quística en les de vida lliure és bastant complexa i resistent, amb un nombre de capes que varia segons les espècies i que, d’acord amb els estudis duts a terme amb Fasciola hepatica, depèn del nombre de tipus de cèl·lules cistògenes de la cercària. En F. hepatica hi ha dues capes concèntriques, una d’externa i una altra d’interna, els precursors de les quals són produïts per quatre tipus de cèl·lules cistògenes. Els quists de les metacercàries enquistades paràsites també són de forma arrodonida, amb una paret de gruix variable però sempre més fina i menys complexa que la dels quists lliures. En els casos de metacercàries paràsites es pot diferenciar entre la porció quística elaborada per l’hoste o càpsula (resultant d’un procés d’encapsulament) i la porció elaborada pel paràsit o quist pròpiament dit.
El cicle vital
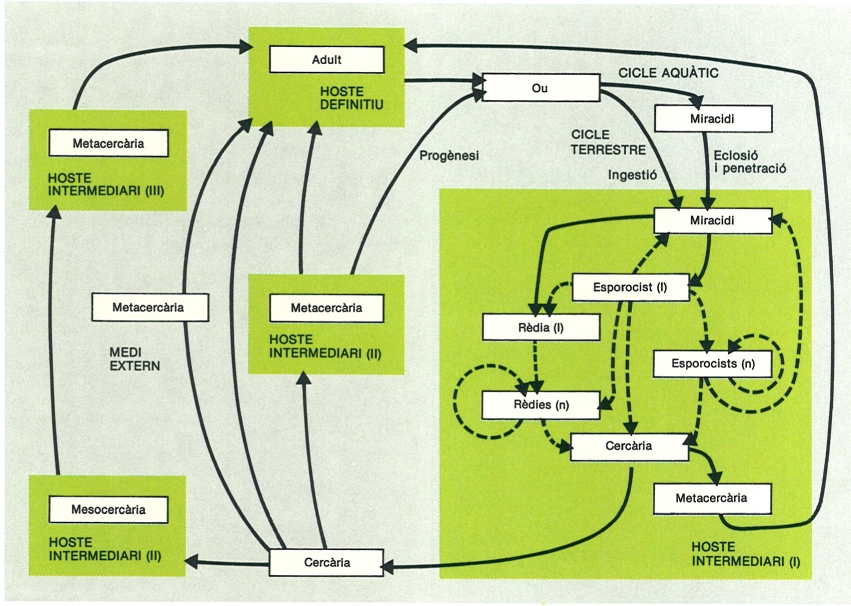
Esquema general dels diferents tipus de cicles vitals que es poden trobar en els digenis. Les línies discontínues indiquen processos de multiplicació asexual.
Biopunt, original de l’autor.
El cicle vital dels trematodes digenis requereix sempre la intervenció de com a mínim dos hostes diferents (heteroxè): l’hoste final o definitiu on s’allotgen en l’estadi adult, i l’hoste o hostes intermediaris, on viuen durant els estadis larvals. Hi ha digenis que utilitzen un, dos o tres hostes intermediaris (cicles di, tri, i tetraheteroxens, respectivament), però la majoria són triheteroxens.
L’hoste definitiu és quasi sempre un vertebrat, excepcionalment altres animals: Alloglossidium macrobdellensis paràsita una sangonera, i Asymphylodara demeli, l’hoste definitiu normal del qual és un peix, es desenvolupa també en un anèl·lid (nerèid) com a hoste vicàriant; potser caldria afegir el cas de les metacercàries progenètiques, que utilitzen invertebrats com a hostes intermediaris, perquè, ja que arriben a la maduresa sexual i donen ous viables, cal considerar-les en certa manera també adults.
El primer hoste intermediari constitueix un denominador comú en el cicle dels trematodes digenis, tant per la seva naturalesa com pels processos que s’hi esdevenen. Es tracta sempre d’un mol·lusc (excepcionalment és un anèl·lid), en general un gasteròpode, menys sovint un bivalve i excepcionalment un escafòpode. En aquest primer hoste intermediari el cicle té com a finalitat produir un gran nombre de larves mitjançant la metamorfosi i la multiplicació regenerativa, de manera que a partir d’un únic individu (miracidi), i passant per diferents estadis larvals segons els casos (esporocists, rèdies), s’origina un gran nombre d’individus finals (cercàries). El segon i tercer hostes intermediaris, quan n’hi ha, poden ésser invertebrats o vertebrats, i la seva funció essencial és de transportar el digeni fins a l’hoste definitiu. En absència d’intermediaris, el paràsit ateny l’hoste definitiu per mitjà d’una forma de vida lliure (cercària, metacercària enquistada) o bé mitjançant una forma paràsita del primer hoste intermediari (metacercària enquistada).
En els cicles amb un sol hoste intermediari, les cercàries poden restar-hi o bé ésser expulsades abans de la infecció de l’hoste definitiu. En el primer cas, pot ésser que la cercària s’encisti a dins mateix de l’esporocist (és el cas de Leucochloridium, l’esporocist del qual, ple de metacercàries, s’instal·la als tentacles del mol·lusc i els tenyeix de bandes de colors que atreuen l’atenció dels ocells, que fan d’hostes definitius), o bé que s’encisti directament en el mol·lusc (els ous de Typhlocoelium cymbium comencen a eclosionar dins l’úter i en surt un miracidi que ja és portador d’una rèdia, que, després de penetrar en un mol·lusc planòrbid, és alliberada i produeix dues cercàries sense cua que s’enquisten en els teixits del mol·lusc). Un cas especial és el Dollfusinus frontalis, que fa servir un mateix individu de mol·lusc simultàniament de primer i de segon hostes intermediaris. La segona modalitat és que la cercària abandoni el primer hoste intermediari. Pot ésser que la cercària que ha sortit del mol·lusc, de vida nedadora, sigui ingerida per l’hoste definitiu (per exemple, els azígids, que quan són adults viuen a l’esòfag i a l’estómac de peixos d’aigua dolça), o bé que hi penetri activament, com és el cas dels coneguts esquistosomes, causants de l’esquistomiasi o bilharziasi, que penetren a través de la pell i s’instal·len en el corrent sanguini. Un cas semblant però amb producció de metacercàries és el d’Haplometra cylindracea, paràsit d’amfibis, en el qual les cercàries que han sortit d’un mol·lusc limnèid penetren a través del tegument de la cavitat bucal dels amfibis i s’hi encisten, però en menys de 48 hores les metacercàries formades es desencisten i migren fins als pulmons. I el cas més freqüent és que les cercàries, després de sortir de la rèdia que parasita el mol·lusc, nedin fins als vegetals de la vora de l’aigua o vers altres objectes i hi restin encistades en forma de metacercària fins que l’hoste definitiu se les menja, com passa en Fasciola hepatica o Fasciolopsis buski, o també en Notocotylus neyrai, un paràsit de rates d’aigua freqüent al nostre país.
Quan el cicle inclou dos hostes intermediaris, generalment el primer és un mol·lusc aquàtic, rarament terrestre (en els dicrocèlides i els braquilàimides) i el segon és un invertebrat o vertebrat de sang freda. En la majoria dels cicles, les cercàries es transformen en metacercàries sense passar una metamorfosi, en dues modalitats, segons que les cercàries siguin produïdes per rèdies (hi ha cicles lleugerament diferents dins aquesta modalitat, com els de Lissorchis mutabile o de Skrjabinophyetus neomydis i de . soricis , o Nephrotrema truncatum) o bé per esporocists (també amb cicles diferents dins d’aquesta segona modalitat, com els de Ptychogonimus megastoma, Dicrocoelium dentriticum, Gorgodera euzeti, Haematoloechus pyrenaicus, Plagiorchis neomydis, Pseudocephalotrema pyrenaica o Brachylaima ruminae i Pseudoleucochloridium pericardicum). Només en els estrigèids, les cercàries passen per una metamorfosi per atènyer l’estadi de metacercària, i ho fan generalment dins un vertebrat (peix, amfibi o rèptil), rarament un mol·lusc o una sangonera, que actua de segon hoste intermediari.
El cicle amb tres hostes intermediaris és molt rar i constitueix una adaptació d’un digeni de cicle aquàtic a hostes terrestres. Pot tractar-se d’una tetraheteroxènia obligada, com passa en el gènere Strigea (el qual empra com a hostes intermediaris un mol·lusc, un amfibi i un tercer hoste amfibi, serp, ocell o mamífer) i també en Halipegus ovocaudatus (que utilitza un mol·lusc, un crustaci i un insecte odonat com a subsegüents hostes intermediaris); o d’una tetraheteroxènia facultativa, que hom troba en els gèneres Alaria i Pharyngostomum (paràsits intestinals de mamífers carnívors).
Els factors ecològics que intervenen en les etapes successives d’un cicle afecten tant el paràsit com els hostes; alhora, cal distingir entre els factors que influeixen els trematodes en els estadis lliures i en els estadis paràsits d’una banda, els que influeixen els hostes, i, encara, els que determinen la transmissió.
Els estadis lliures en els digenis són les fases externes d’ou, de miracidi, de cercària i de metacercària. L’ou és una fase passiva que, gràcies a la closca, demostra una resistència als factors ambientals. En el cas d’espècies els ous de les quals arriben al medi extern encara sense embrionar, la seva viabilitat i la seva eclosió (en els casos de miracidi amb vida lliure), són sensibles a factors abiòtics. Els terrestres són sensibles a la temperatura, la llum (radiació ultraviolada), la humitat i la dessecació; els aquàtics ho són al pH, la salinitat, la concentració d’oxigen i la pressió osmótica. I fins i tot són sensibles a factors biòtics, com els efectes ovicides d’algunes espècies de fongs o la depredació per hostes inadequats. El miracidi (de vida lliure únicament quan els ous fan eclosió a l’aigua) és una fase nedadora, en la qual incideixen factors essencialment físico-químics, com la temperatura, la llum solar i la gravetat, i etològics, d’atracció cap al mol·lusc (primer hoste intermediari). La fase lliure de cercària ha estat la més estudiada des del punt de vista ecològic. Diferents factors afecten la producció de cercaries (el nombre de miracidis que infesten el mol·lusc, la capacitat de replicació dels esporocists o rèdies cercariògenes, temperatura) i altres afecten la seva emissió en ritmes d’emergència; sembla que en les cercàries aquàtiques els sincronitzadors principals d’aquests ritmes són el fotoperíode i el termoperíode, mentre que en les terrestres és la presència d’aigua, i també l’activitat i el comportament del mol·lusc emissor. La dispersió de cercàries aquàtiques depèn poc del seu comportament i molt de les característiques hidrodinàmiques del medi. La capacitat d’infecció de les cercàries depèn del període en què l’esporocist les ha produït (de producció alta o baixa), del moment del ritme circadià en què han estat emeses, del temps transcorregut després de la sortida a l’exterior i de la temperatura. La metacercària, quan és de vida lliure (només en algunes espècies), sempre és una fase passiva i aquàtica, i la seva viabilitat depèn tant de factors físico-químics com de factors biòtics (existència de suports vegetals o animals, depredació per animals que són inadequats com a hostes).
Els estadis paràsits són els de miracidi (dins el mol·lusc), esporocist, rèdia, cercària (dins el mol·lusc), mesocercària, metacercària i adult. El miracidi, en l’etapa que és dins el mol·lusc, depèn essencialment de l’especificitat i dels mecanismes de defensa de l’hoste. L’especificitat és molt marcada, de tipus fisiològic, fins al punt que una espècie de digeni només es pot desenvolupar en una única espècie (oioxènia), o en espècies molt pròximes i emparentades, de mol·luscs (estenoxènia), en un lloc determinat, si bé en punts geogràfics distants hi pot haver races fisiològiques diferents del mateix digeni que utilitzen espècies de mol·luscs filogenèticament una mica més allunyades (oligoxènia estreta); per exemple, Fasciola hepatica (cosmopolita) només utilitza espècies de la família dels limnèids; Dollfusinus frontalis utilitza, a Formentera, dues espècies del cargol terrestre Trochoidea (subgènere Xerocrassa), mentre que al migjorn francès utilitza una espècie d’un gènere proper de la mateixa subfamília. El mol·lusc està dotat de mecanismes interns de defensa innats i adquirits, semblants als immunitaris dels vertebrats; fins i tot s’ha demostrat que races geogràfiques diferents d’una espècie de mol·lusc resisteixen diferentment una espècie determinada de digeni paràsit (per exemple, Schistosoma mansoni i Biomphalaria glabrata). El desenvolupament d’esporocists i rèdies també es veu afectat per l’especificitat, la «immunitat», la temperatura, el nombre de miracidis originadors de la infestació, la coexistència amb estadis larvals d’altres espècies de digenis, l’alimentació i l’edat del mol·lusc.
Les cercàries, en la curta fase que passen dins el mol·lusc, són sotmeses a les seves reaccions de defensa, sobretot aquelles que s’han equivocat de via de migració i han anat a parar a un carreró sense sortida. Especificitat i reaccions de defensa són també els factors més importants en la penetració del següent hoste. Mesocercàries i metacercàries, quan són paràsites, en són d’invertebrats o de vertebrats, i de vegades s’enquisten i d’altres no. Sobre elles actuen factors com els físico-químics del microhàbitat de parasitació, l’especificitat i les reaccions de defensa. Les formes enquistades són sempre més resistents que les que no ho són. Pel que fa a l’especificitat, els digenis solen ser-ne poc per al segon (i tercer) hoste intermediari, i en tot cas és més aviat de tipus ecològic i etològic (requeriments ambientals, règim alimentari) que no fisiològic, de manera que els que evolucionen en insectes no ho fan ni en crustacis, ni en mol·luscs, ni en vertebrats, i al revés, guiats per les característiques ecoetològiques d’aquests grans grups.
En el cas dels adults també són importants els factors del microhàbitat de parasitació, l’especificitat i les reaccions immunitàries de l’hoste (vertebrat); aquestes són evadides mitjançant diverses estratègies; en el cas d’Schistosoma, els adults, que viuen a l’interior dels vasos sanguinis, engeguen un curiós mecanisme pel qual s’envolten superficialment d’antígens del mateix hoste. L’especificitat dels digenis adults sol ésser fonamentalment ecoetològica (vertebrats que consumeixen els hostes intermediaris o vegetals que porten les metacercàries). Tanmateix, hi ha casos en què l’especificitat de l’hoste és molt estricta. Hi ha espècies d’Schistosoma pròpies de l’home, que únicament utilitzen com a «reservoris» alguns simis antropomorfs i rosegadors; Schistosoma haematobium, per exemple, és molt difícil d’introduir en altres possibles hostes. Aquest últim fenomen té relació amb la manera d’infestació activa transcutània que segueixen les cercàries d’Schistosoma. De tota manera, entre els digenis que infesten l’hoste definitiu per via oral també hi ha casos amb una certa especificitat que va més enllà de la simplement ecoetològica, com succeeix en molts representants antics de la família dels braquilàimids, paràsits de mamífers i ocells. Dins els paràsits de peixos són majoria les espècies oioxenes (paràsites d’una sola espècie de peix), nombroses les oligoxenes (paràsites de peixos pròxims taxonòmicament, per exemple pertanyents a una mateixa família) i molt menys nombroses les eurixenes (paràsites de peixos allunyats taxonòmicament).
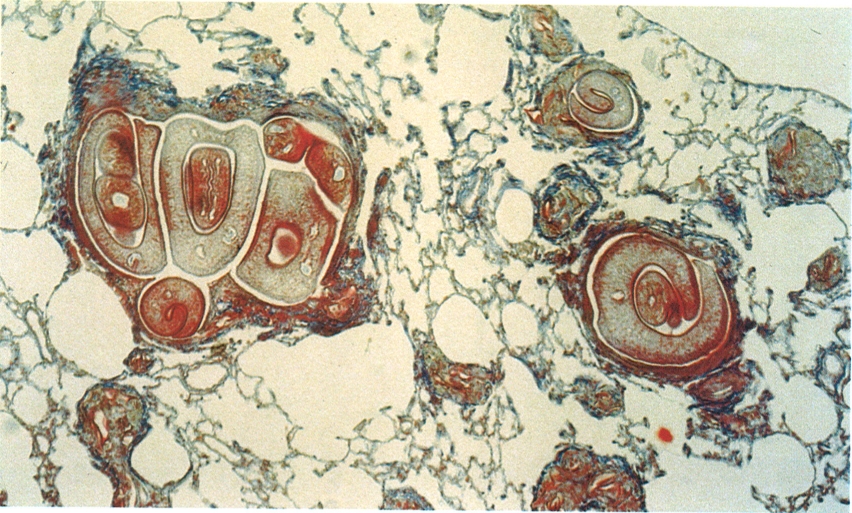
La bilharziosi es manifesta a nivell histològic per la presència de Schistosoma mansoni en els teixits de l’hoste afectat. La fotografia correspon al teixit pulmonar d’una rata afectada per aquesta malaltia, i hom hi pot distingir parelles (mascles i femelles) d’aquest digeni.
Santiago Mas-Coma.
Des d’un punt de vista ecològic hi ha dos tipus de cicles en els digenis. Els uns es completen en un sol hàbitat, aquàtic o terrestre, i llavors la continuïtat del cicle només és assegurada quan els estadis lliures del digeni i de l’hoste mostren, almenys en un moment donat, les mateixes exigències ecològiques (ocupació d’un mateix biòtop). Els altres inclouen dos o més ambients, i és usual que els digenis en requereixin un d’aquàtic i un altre de terrestre. El pas de l’aigua cap a terra ferma es fa amb hostes intermediaris que surten de l’aigua, com els insectes que emergeixen de larves aquàtiques o com els amfibis, o bé mitjançant hostes definitius terrestres que mostren tendències aquàtiques en el seu comportament o alimentació. El pas invers, del medi terrestre a l’aquàtic, va lligat al contacte entre l’hoste definitiu i l’aigua, on arriben els ous del digeni. La intervenció d’hostes capaços de fer desplaçaments notables és el motor essencial de la dispersió del digeni i permet d’explicar, per l’aïllament, les variacions observades en les àrees de repartició. El cicle biològic d’un paràsit no es fonamenta, tanmateix, sobre una trobada purament fortuïta dels medis successius. En la majoria dels digenis, la infestació del primer hoste intermediari freqüentment va lligada al comportament del paràsit (quimiotàxia) i, més rarament, al comportament de l’hoste (depredació d’ous en alguns cicles terrestres). La infestació del segon hoste intermediari va lligada o bé al comportament del paràsit (tropisme), o bé al de l’hoste (per ingestió de la cercària). Finalment, la infestació de l’hoste definitiu generalment va lligada al comportament de l’hoste (depredació del segon hoste intermediari) o, més rarament, al del paràsit (cas dels cicles reduïts o simplificats en què la cercària penetra directament en l’hoste definitiu, el qual se situa com a segon hoste). Pel que fa al pas al medi exterior, pot tractar-se d’un fenomen passiu (evacuació d’ous) o actiu, lligat a un tropisme (sortida de cercàries). En definitiva, la viabilitat d’un cicle en un lloc determinat és influïda per l’abundància d’hostes (sobretot quan el paràsit manifesta una especificitat estricta), pels factors biòtics i abiòtics que poden reduir la població parasitària (principalment en els estadis paràsits i secundàriament en els lliures), i pel comportament dels hostes, especialment el de l’hoste definitiu.
Infestació del primer hoste intermediari
Replicació dels esporocists de Schistosoma mansoni en l’hepatopàncrees d’un cargol de l’espècie Biomphalaria glabrata. En la part superior de la fotografia apareix un esporocist cercariogen, és a dir, amb cercàries en formació al seu interior, i a la part inferior, un esporocist replicador o esporocistogen.
Santiago Mas-Coma.
La infestació del mol·lusc (primer hoste intermediari) té lloc o bé per penetració activa del miracidi a través del tegument (cicles aquàtics), o bé passivament, quan el mol·lusc ingereix l’ou i aquest allibera el miracidi en el seu tub digestiu (cicles terrestres i alguns d’aquàtics). En el primer cas, el miracidi, de durada breu (usualment de no més d’un dia) neda fins aproximar-se al mol·lusc adequat i, quan hi és a prop, sembla que responent a estímuls quimiotàctics procedents essencialment del mucus del mol·lusc, hi entra en contacte i s’hi adhereix mitjançant secrecions especials o bé per l’acció succionant de la papil·la apical; seguidament penetra el tegument mitjançant els moviments d’aquesta papil·la i de tot el miracidi i gràcies a una acció enzimàtica citolítica. Ja dins el mol·lusc, en general el miracidi fa una migració curta: es transforma en el següent estadi larval en el mateix punt de penetració o molt a prop, generalment en el teixit del mantell, del peu o dels tentacles. En els casos en què el mol·lusc ingereix l’ou i aquest fa eclosió en el seu aparell digestiu, el miracidi travessa la paret i fa la metamorfosi ben a prop, habitualment en l’hepatopàncrees, on continuarà el desenvolupament larval. L’esporocist mare originat en el punt de penetració produeix rèdies o esporocists fills, segons les espècies, que l’abandonen en general a través d’un porus de sortida i migren cap a l’hepatopàncrees o a prop, on passen a produir cercàries. En aquelles rares espècies en les quals el miracidi inclou ja una rèdia, aquesta és alliberada després de penetrar el tegument i fa una migració ràpida cap al cor; si el que inclou és un esporocist, el miracidi penetra la pell del mol·lusc només en part i l’esporocist surt per consumar el procés d’infestació i migra cap al cor, on produeix unes quantes rèdies mare, que a la vegada són productores de nombroses rèdies filles, que migren cap a l’hepatopàncrees i donen lloc a rèdies nétes ja cercariògenes. L’hepatopàncrees constitueix el microhàbitat de parasitació d’esporocists i rèdies cercariògens per excel·lència. Els esporocists no tenen boca i han d’absorbir tots els nutrients a través de la seva paret, estimulant la històlisi de les cèl·lules hepatopancreàtiques per a obtenir materials lliures absorbibles; en canvi, les rèdies tenen un aparell digestiu simple i són capaces tant d’ingerir cèl·lules com d’absorbir materials a través de la paret. Tant en un cas com en l’altre, les cercàries acabades de formar surten del sac germinal en general a través d’uns porus especials, si bé hi ha casos en què han de trencar-ne la paret i d’altres en què no arriben a sortir-ne (casos en què els sacs poden romandre dins el mol·lusc o bé ésser expulsats a l’exterior). Un cop fora, les cercàries inicien una migració cap a l’exterior (menys en els casos rars en què s’enquisten). Aquesta té lloc en general a través del sistema circulatori del mol·lusc, i finalment en surten per orificis naturals com l’anus o el pneumostoma, bé travessant activament teixits (esquistosomàtids) o bé seguint conductes naturals (braquilàimids).
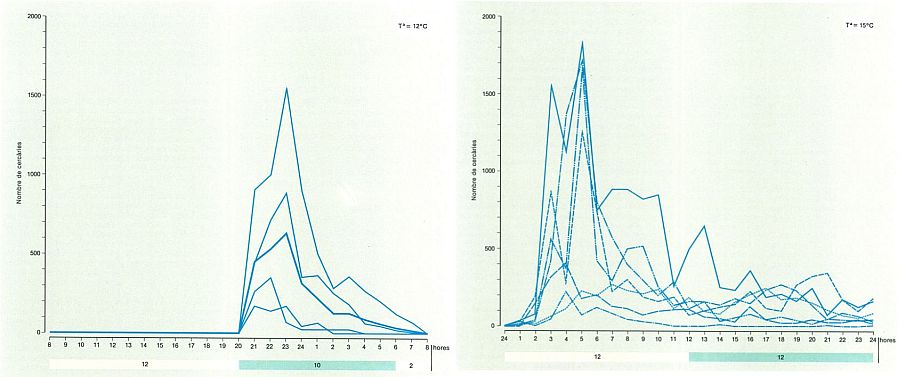
L’emissió de les cercàries a partir de l’hoste parasitat ha estat motiu de diversos estudis, en els quals hom compara les corbes d’emissió cercariana que s’obtenen alterant les fases de llum i fosca i la temperatura en diferents espècies. La figura en mostra alguns resultats. A dalt hom ha dibuixat les corbes d’emissió cercariana del plagiòrquid Plagiorchis muris; representen el nombre de cercàries d’aquesta espècie emeses durant 32 hores per diferents cargols parasitats (Radíx limosa), en condicions de ritme de fosca i llum i temperatura constants (la mitjana és indicada per la línia gruixuda). A baix, corbes d’emissió cercàriana de Brachylaima ruminae en un dia, corresponents a diferents cargols parasitats (Rumina decollata), en condicions igualment constants.
Carto-Tec, amb originals d’A. Theron, S. Mas-Coma i M. Gracenea.
L’emergència de cercàries a l’exterior o emissió cercariana pel mol·lusc segueix, en la majoria dels digenis, un ritme concret. El més general és que emetin les cercàries amb un ritme circadià, amb un sol pic de màxima o acrofase cada 24 hores; es distingeixen espècies amb cercàries de dia (en Schistosoma mansoni, S. haematobium, etc., l’acrofase es dona en hores de llum solar), espècies amb cercàries de nit (Plagiorchis vespertilionis, Riberoia marini, etc.) i casos intermedis (acrofase a l’alba o al capvespre, com en Petasiger caribbensis). Altres casos menys nombrosos (Clinostomum golvani) són de ritme ultradià, amb dos o més punts àlgids cada 24 hores. D’altres estan sotmesos aparentment a un ritme infradià, amb punts d’emissió màxima irregulars i amb una periodicitat superior a 24 hores, que poden estar superposats als anteriors i en certa manera relacionats amb les estacions de l’any (en espècies de cicle terrestre com Brachylaima ruminae o Pseudoleucochloridium pericardicum). També s’ha detectat alguna espècie en què l’emissió de cercàries no segueix cap ritme concret, com Fasciola hepatica. L’existència de ritmes d’emissió és un mecanisme que afavoreix la infestació de l’hoste següent, perquè procura una alta concentració de cercàries en el moment més oportú per a la infestació; es comprèn, doncs, que no hi sigui en Fasciola hepatica, ja que la seva cercària, en lloc d’infestar un hoste, entra en un période de latència (metacercària enquistada). Estudis recents han demostrat fins i tot l’existència d’una variabilitat intraspecífica en l’emissió, amb races cronobiològiques diferents en zones geogràfiques diverses, que són adaptacions a les condicions locals de transmissió. Un exemple excel lent és el de Schistosoma mansoni a l’illa caribenya de Guadalupe, on s’observen uns màxims diaris d’emissió diferents en zones diverses de l’illa segons que el principal hoste definitiu sigui l’home o la rata: en els llocs en què l’home i la rata presenten una incidència de parasitació alta i baixa respectivament, l’emissió màxima és poc abans del migdia (cosa que afavoreix la infestació en els homes, ja que és l’hora en què és més probable el bany), mentre que en llocs despoblats, on la rata sofreix una alta taxa d’infestació, l’emissió màxima és tardana, com una adaptació a l’horari de màxima activitat d’aquest animal, que és preferentment crepuscular.
Infestació del segon hoste intermediari
En els cicles triheteroxens o tetraheteroxens, l’activitat i el comportament de la cercària un cop en el medi extern van sempre destinats a afavorir la infestació del segon hoste intermediari. En el cas de cicles diheteroxens, la cercària adopta un comportament destinat a infestar l’hoste definitiu, en general per via transcutània o, accidentalment, oral (Schistosoma), bé es dirigeix cap a vegetals o altres suports (conquilles de mol lusc, etc.), on s’enquista ràpidament i forma la metacercària per a facilitar que l’hoste definitiu la ingereixi (Fasciola hepatica). Les cercàries aquàtiques neden utilitzant la cua com a órgan locomotor, excepte aquelles que, pel fet d’ésser microcerques o acerques, repten sobre el substrat del fons (Paragonimus westermani). Les cercàries terrestres, només amb un rudiment caudal, repten sobre el soi utilitzant les dues ventoses, com fan els braquilàimids. L’activitat i el comportament de les cercàries lliures depenen d’estímuls fototàctics (positiu o negatiu segons les espècies, essent més sensibles les que presenten taques oculars), mecànics (agitació i corrent d’aigua), geotàctics (positiu o negatiu segons les espècies) i químics (la proximitat de l’hoste provoca un marcat increment de l’activitat). La viabilitat de les cercàries lliures és reduïda, i, en general, és més petita en les aquàtiques (de 4 a 11 hores) que en les terrestres (entre unes hores i dos dies, en els braquilàimids). Hi ha casos excepcionals de cercàries que presenten cobertes de resistència que els permeten de mantenir-se més temps en el medi extern, com les cercàries aquàtiques d’Halipegus ovocaudatus, que són emeses a l’aigua enquistades i romanen viables fins 41 dies, a 14°C, o les cercàries terrestres de Dicrocoelium dendriticum, que són expulsades enquistades i en paquets coberts d’un moc que els permet de suportar un cert grau de sequedat.
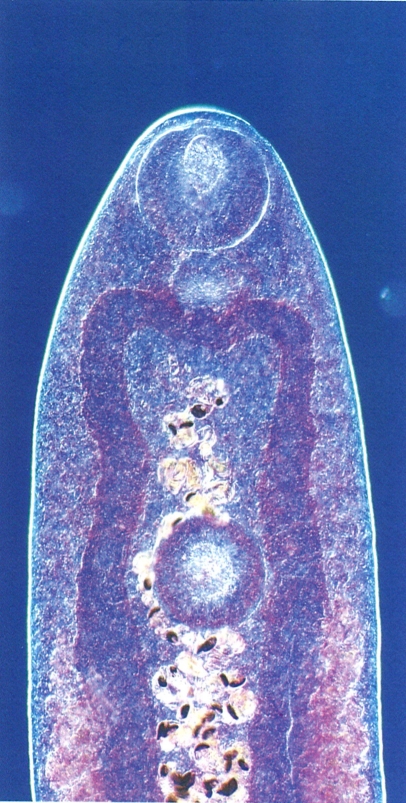
Detall de l’extrem anterior del cos d’un exemplar de 15 dies de vida del digeni monoic Brachylaima ruminae, paràsit de rosegadors.
Santiago Mas-Coma.
La infestació del segon hoste intermediari es pot fer, un cop establert el contacte, de tres maneres. La primera és la de les cercàries que penetren activament, a través de la pell, en invertebrats o vertebrats, que perden la cua i migren cap al microhàbitat (cavitat corporal, ronyó, sistema nerviós, etc., en invertebrats; musculatura o altres teixits, en vertebrats); alla originaran l’estadi de metacercària, habitualment enquistada, o l’estadi de mesocercària, no enquistada. En el cas de metacercàries enquistades, la seva presència provoca habitualment una resposta dels teixits de l’hoste, que consisteix a formar una càpsula al seu voltant; així, cal distingir entre el material quístic d’origen parasitari i el procedent de l’hoste (Clonorchis sinensis, en peixos ciprínids). El procès d’enquistament en general és molt ràpid, i en les metacercàries que s’enquisten dins un hoste intermediari s’originen menys capes quístiques que en les que ho fan en el medi extern. La segona manera és la de les cercàries que accedeixen al segon hoste intermediari passivament, per ingestió. De vegades, la cercària, aquàtica, per atreure’l, fa moviments que recorden les preses habituais de l’hoste, mentre que altres vegades és únicament el comportament alimentari de l’hoste el que determina la infestació, com passa amb les cercàries aquàtiques, actives peró sensé cua, de Lissorchis mutabile, que són ingerides per l’anèl·lid Chaetogaster, o amb les cercàries terrestres i immòbils (estan incloses en paquets) de Dicrocoelium dendriticum, que són ingerides per formigues. En tots aquests casos, la cercària travessa la paret de l’intestí de l’invertebrat, perd la cua (si en tenia) i arriba fins a l’hemocel, on origina l’estadi de metacercària, enquistada o no. En el cas de Dicrocoelium dendriticum es dóna, a més, el fet curiós que la primera cercària que penetra a l’hemocel de la formiga es dirigeix al gangli subesofàgic i s’enquista, cosa que fa que la formiga canviï el seu comportament: durant el dia puja per les tiges de les herbes i s’hi fixa ben amunt amb les mandíbules (així afavoreix la possibilitat d’ingestió per herbívors), mentre que al vespre recupera els seus moviments i torna a terra. Una tercera manera és la de les cercàries que penetren activament en el segon hoste intermediari a través d’orificis naturals, com succeeix amb les cercàries terrestres i microcerques d’espècies de braquilàimids (Brachylaima ruminae, Pseudoleucochloridium pericardicum o Dollfusinus frontalis), que penetren en cargols terrestres a través del seu orifici excretor, i migren seguint els conductes naturals fins al ronyó o la cavitat pericàrdica, on originen les metacercàries, que no són mai enquistades.
A diferència del que succeeix en metacercàries que s’enquisten en el medi extern i atenyen l’estadi infestant poc temps després (hores o dies), la maduració de les metacercàries en el segon hoste intermediari necessita generalment un temps més llarg (entre unes quantes setmanes i uns mesos), que varia segons les espècies però que s’escurça en augmentar la temperatura.
Infestació de l’hoste definitiu
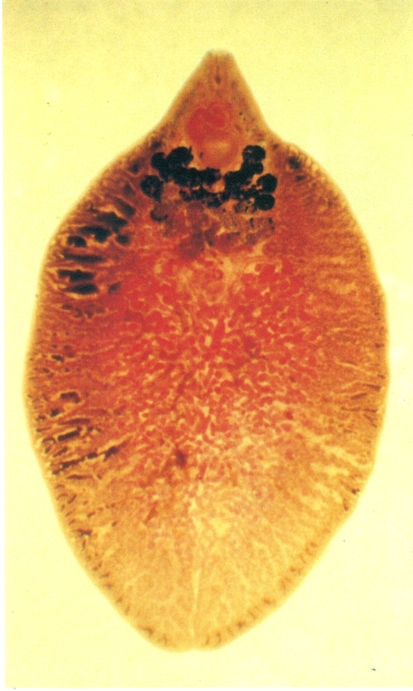
Distoma hepàtic (Fasciola hepatica), paràsit, en estat adult, dels conductes biliars de les ovelles, que ocasionalment pot afectar l’home. El distoma hepàtic té un cicle biològic complex, a través d’un cargol d’aigua dolça (limnèid) com a hoste intermediari. L’individu fa aproximadament 3-cm de llargada.
Santiago Mas-Coma.
En cicles dihetereroxens, la infestació de l’hoste definitiu pot ésser passiva, per via oral; en aquest cas, l’hoste ingereix el mol·lusc, que té metacercàries enquistades dins l’esporocist (Leucochloridium) o en els seus mateixos teixits (Typhlocoelium cymbium), o bé sense enquistar, en els seus òrgans (Dollfusinus frontalis). Altres vegades es fa per ingestió directa de les cercàries lliures (cas dels azígids), que neden fent moviments que recorden els de les larves de mosquits i atreuen els peixos d’aigua dolça. La infestació també es pot produir si l’hoste ingereix metacercàries enquistades que estan adherides en vegetals o altres elements del medi (Fasciola hepatica). En cicles diheteroxens també pot donar-se una infestació activa de l’hoste definitiu, en què la cercària el penetra a través del tegument i, un cop dins, es poden formar metacercàries enquistades (Haplometra cylindracea) o no (Schistosoma) abans de donar lloc als adults. En els cicles tri i tetraheteroxens, la infestació és sempre passiva, per via oral: l’hoste definitiu ingereix el segon o, més rarament, el tercer hoste intermediari, que porta les metacercàries.
Mentre que en el cas de la ingestió de metacercàries no enquistades es pot iniciar de seguida la migració o el desenvolupament, en el cas de les metacercàries enquistades ha de tenir lloc abans el desenquistament. Aquest procés és complex i diferent segons les espècies, ja que hi incideixen factors tant extrínsecs (enzims digestius de l’hoste, temperatura, condicions reductores, pH, concentració d’anhídrid carbònic, i presència de sals biliars) com intrínsecs (secrecions del paràsit); la majoria de quists metacercarians necessiten almenys pepsina (secreció gàstrica) i normalment tripsina (enzim intestinal) per a desenquistar-se i alliberar la metacercària. En el cas de la ingestió de cercàries d’azígids, aquestes originen directament els adults a l’estómac o a l’esòfag de peixos d’aigua dolça. Pel que fa als casos en què la cercària penetra activament a través del tegument, en espècies com Haplometra cylindracea, penetren en la cavitat bucal de capgrossos i amfibis adults, on s’enquisten i, ràpidament (en menys de 48 hores), es desenquisten i migren directament als pulmons, on originen els adults; en els esquistosomàtides, la cercària penetra ràpidament (en la pell humana, en mitja hora o menys), aprofitant una discontinuïtat de la pell (fol·licles pilosos), perd la cua, es transforma morfològicament i canvia fisiològicament fins a formar l’anomenat esquistosòmul, que accedeix ràpidament als vasos de la zona de penetració i migra, per via limfàtica o sanguínia, fins als vasos de vísceres internes, on dona lloc a l’adult.
Exemplar del digeni Prosthodendrium parvouterus, paràsit de quiròpters del nostre país.
Santiago Mas-Coma.
Les metacercàries d’espècies els adults de les quals tenen el microhàbitat de parasitació fora de l’aparell digestiu, han de fer una migració mes o menys llarga per arribarhi. Els mecanismes fisiològics que acompanyen aquest tropisme encara no han estat del tot aclarits: cal recordar que els digenis han arribat a colonitzar microhàbitats tan diversos com ara el tracte digestiu, el fetge, el pàncrees, els pulmons, l’aparell circulatori, els ronyons, la bufeta urinària, els sinus nasals i frontals, els ulls, etc., i que de vegades espècies diferents atenyen un mateix microhàbitat seguint camins migratoris diferents. Per exemple, Fasciola hepatica i Dicrocoelium dendriticum són tots dos paràsits de l’obertura dels canals hepàtics i de la vesícula biliar, però mentre que . hepatica hi arriba travessant la paret intestinal (penetra des de la cavitat peritoneal en el parènquima hepàtic), D. dendriticum ho fa seguint l’obertura del colèdoc.
Els digenis adults de cada espècie volen en general un òrgan de residència molt concret, si bé de vegades es troben excepcions (com els adults de Fasciola hepatica, anormalment localitzats als pulmons). Aquesta especificitat està relacionada amb la naturalesa de la seva alimentació: s’alimenten per succió, mitjançant la ventosa oral i la faringe, i fan digestió intestinal, si bé també poden absorbir petites molècules a través del tegument extern. Així, els que fan vida intestinal s’alimenten de mucus, mucosa epitelial, de vegades sang i també aliments de l’intestí de l’hoste a mig digerir; els paràsits de la bufeta urinària s’alimenten predominantment de teixit epitelial; els que viuen als pulmons i al sistema circulatori s’alimenten de sang; i els que viuen al fetge mengen bilis i teixit hepàtic. Quant al metabolisme, és predominantment anaerobi, fins i tot en espècies que viuen en medis aerobis, com els esquistosomes, i la font primària d’energia són els carbohidrats. En el microhàbitat final, la taxa de maduració dels adults varia segons l’espècie, la seva localització final en l’hoste, el grau de desenvolupament i diferenciació que tenia la metacercària en el moment de la infestació, l’espècie d’hoste, i fins i tot de vegades també la raça. Així, dins un mateix microhàbitat, les espècies de digenis de dimensions més grans necessiten més temps per a madurar; d’altra banda, creixen i maduren més ràpidament les que viuen en microhàbitats amb aliment abundant, i maduren més de pressa les que han partit d’una metacercària en un estat avançat de desenvolupament (per exemple, amb els òrgans sexuals ja diferenciats), com també les que tenen un hoste definitiu habitual per a l’espècie, etc. De vegades, aquestes diferències poden arribar a ésser molt grans entre espècies d’una mateixa família, com en el cas dels braquilàimids: els adults de Pseudoleucochloridium pericardicum, paràsit intestinal de musaranyes, maduren molt ràpidament, i són evacuats, sense tenir temps de créixer, al cap de 24 o 48 hores de produir-se la infestació; en canvi, els adults de Brachylaima ruminae, paràsit intestinal de rosegadors, són ovífers ja a partir dels 3 o 6 dies (segons l’edat de les metacercàries ingerides), i el creixement i la vida es prolonguen fins no gaire més enllà dels 150 dies de postinfestació; no és així per a Dollfusinus frontalis, paràsit dels sins nasals i frontals d’eriçons i rosegadors, els adults del qual allarguen la seva vida més d’un any.
Els adults s’atreuen mútuament, mitjançant estímuls químics, per a fer la fecundació. Els ous són arrossegats a través dels conductes naturals fins que surten a l’exterior per un orifici natural de l’hoste: en els paràsits del sistema digestiu o de fetge i pàncrees surten amb els excrements; els ous dels que habiten al sistema excretor (ronyó, bufeta urinària) surten amb l’orina; i els dels paràsits dels sins nasals o frontals i de les vies respiratòries, surten amb els esputs o els estornuts, o bé són deglutits i travessen el tracte digestiu. Els digenis que viuen a l’interior del sistema circulatori (esquistosomàtides) són un cas especial. En aquest darrer cas, el ous deixats anar primer han de travessar la paret dels vasos (cosa que fan aprofitant l’aprimament que presenta aquesta paret quan els ous s’envolten de petits coàguls de sang i té lloc la infiltració de fibroblasts i l’acció de cèl·lules endotelials), i després els teixits circumdants (gràcies a l’acció d’enzims histolítics secretats pel miracidi intern) fins que cauen en l’obertura de l’òrgan més pròxim (intestí o bufeta urinària, segons les espècies) i finalment surten pels excrements o bé per l’orina (respectivament). Segons l’embrionament i el comportament del miracidi, es poden distingir quatre tipus principals d’ous: els que encara no estan desenvolupats en el moment de la posta i han de fer l’embrionament complet fora de l’hoste (Fasciola hepatica, Paragonimus westermani), els que ja es troben parcialment desenvolupats i demanen un breu període a l’exterior per a completar l’embrionament (Haematoloechus), els que estan completament embrionats quan abandonen l’hoste i fan eclosió immediatament quan arriben a l’aigua (Schistosoma) o després d’ésser ingerits pel mol·lusc apropiat (Dicrocoelium dendriticum, Haplometra cylindracea) i, finalment, aquells en què l’eclosió del miracidi pot tenir lloc fins i tot dins l’úter del digeni (Typhlocoelium cymbium, Parorchis acanthus).
Filogènia i sistemàtica
La reconstrucció de l’origen i evolució dels trematodes és difícil per l’absència de dades paleontològiques; només tenim indicis indirectes, com per exemple les perles trobades en bivalves triàsics, possiblement causades per aquests paràsits. En una escala temporal més petita, són útils les troballes d’ous en materials fecals en les excavacions arqueològiques, com els de Schistosoma, trobats en mòmies d’Egipte.
Els digenis adults són presents en tots els grans grups de vertebrats, des dels elasmobranquis fins als ocells i els mamífers, la qual cosa fa pensar en un origen antic del grup. La majoria dels pocs digenis paràsits d’elasmobranquis deriven d’espècies paràsites de teleostis, la qual cosa indica que els elasmobranquis els van adquirir en una època relativament més recent. D’altra banda, cal recordar que el primer hoste intermediari gairebé sempre és un mol·lusc gasteròpode, d’alt grau d’especificitat, per la qual cosa es creu que l’origen, dels digenis almenys, és posterior al dels cargols pulmonats d’aigua dolça. D’una manera general, hom pensa que els trematodes en els seus orígens eren formes paràsites només de mol·luscs, derivades de turbel·laris semblants als rabdocèlids, i que posteriorment van desenvolupar una cua per a la natació, convertint-se en paràsits d’un vertebrat després d’haver estat ingerits per aquest en un estadi semblant al de cercària, lliure o encistada, o amb el mol·lusc sencer. Per això, es pot pensar que aquest precursor dels trematodes actuals devia ésser semblant als aspidogastris, poc adaptats a la vida parasitària, i de cicle senzill, amb un sol hoste.
Hom pot suposar, doncs, que el cicle primitiu dels trematodes incloïa dos hostes (un mol·lusc aquàtic i un peix, segurament teleosti): la complicació del cicle i l’aparició de la larva metacercària devien ésser posteriors. L’estadi de metacercària, capaç de transformar-se en adult sense passar una metamorfosi complicada, hauria fet possible la invasió dels vertebrats terrestres. Posteriorment s’hauria pogut adoptar un segon hoste intermediari, o tornar a un cicle amb dos hostes per pèrdua de l’hoste definitiu o del segon intermediari. La colonització del medi terrestre, amb un cicle que inclou cargols terrestres com a primers hostes intermediaris i cercàries també terrestres, es considera molt més moderna, i només hi han arribat algunes famílies (dicrocèlids, braquilàimids, leucoclorídids). Per a seguir les línies evolutives de cada grup, cal considerar la condició primitiva o moderna de diferents caràcters a cada estadi, els seus microhàbitats i les seves especificitats. L’anàlisi filogenètica dels digenis feta per mètodes cladístics considerant només les larves està d’acord amb una altra de feta prenent només els adults, la qual cosa demostra que els estadis larvals no són recapitulacions de les formes adultes ancestrals.
La classificació dels trematodes, fins poc després de la primera meitat d’aquest segle, es basava essencialment en la morfologia dels adults, i distingia els dos grans grups dels prosostòmats (Prosostomata) i els gasterostòmats (Gasterostomata), segons si la posició de la boca era en la ventosa apical o en la ventral, respectivament; els aspidogastris (Aspidogastrea) eren considerats un subgrup dels digenis (Digenea) o un grup independent però pròxim, i de vegades com un grup intermedi entre aquells i els monogenis (Monogenea). A la segona meitat d’aquest segle, els aspidogastris ja són considerats unànimement com un grup independent i de la mateixa categoria taxonòmica que els digenis, per als quals ja es comencen a tenir en compte els estadis larvals a l’hora de fer-ne la classificació. Hom proposa per als digenis la distinció entre esporocistoineïns (Sporocystoinei) i redioineïns (Redioinei), amb cercàries produïdes, respectivament, per esporocists o per rèdies. La Rue va proposar (1957) una classificació basada en les característiques de la cercària i el sistema excretor, i distingia els anepiteliocistidis (Anepitheliocystidia) i els epiteliocistidis (Epitheliocystidia). Els didimozoïdeus (Didymozoidea) ja eren considerats un grup a part dels digenis i dels aspidogastris, bé que recentment alguns autors els situen dins els digenis.
Principals parasitosis produïdes per trematodes digenis sobre diversos animals, que poden afectar l'espècie humana
Dades elaborades per l’autor.
| TREMATODES AGENTS DE PARASITOSIS | ANIMAL AFECTATS | ABAST GEOGRÁFIC | |
| PARASITOSIS INTESTINALS | |||
| Fasciolopsis buski | Porc, i accidentalment, l’home | Extrem Orient | |
| Echinostoma iliocanum i E. revolutum | Vertebrats semiaquàtics | Extrem Orient | |
| Heterophyes heterophyes | Carnivors depredadors de peixos | Africa, Asia Menor, Extrem Orient | |
| Metagonimus yokogawai | Carnivors depredadors de peixos | Proxim Orient | |
| Gastrodiscoides hominis | Primats i rosegadors | Extrem Orient | |
| Nanophyetus salmincola | Diversos animals, esporàdicament, l’home | ||
| PARASITOSIS HEPÀTIQUES | |||
| Clonorchis sinensis | Porc, gos i gat | Extrem Orient | |
| Opisthorchis felineus | Diversos carnivors | Asia, Europa oriental | |
| Dicrocoelium dendriticum | Herbivors (bens, vaques) | cosmopolita | |
| Fasciola hepatica | Herbivors domestics | cosmopolita | |
| Fasciola gigantea | Herbivors | Africa, Orient | |
| PARASITOSIS SANGUÍNIES | |||
| Schistosoma japonicvm | Diversos mamifers | Extrem Orient | |
| S. haematobium | Home | Africa, Prôxim Orient | |
| S. intercalatum | Primats i herbivors | Africa | |
| S. mekongi | Asia | ||
| S. mansoni | Primats (home inclos) i rosegadors | Africa, America Central, America del Sud | |
| PARASITOSIS PULMONARS | |||
| Paragonimus westermani | Diversos animals domestics i silvestres | Extrem Orient, Africa | |
| P. mexicanus | Diversos animals | America Central, America del Sud | |
| PARASITOSIS DERMATÍTIQUES | |||
| Cercàries d’esquistosomatoïdeus | Diversos animals, esporàdicament, l’home |