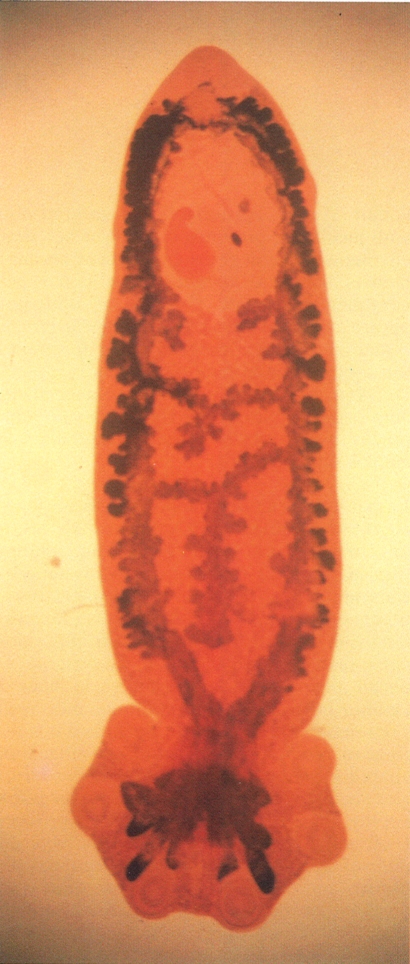
Monogeni paràsit del gripau d’esperons (Pelobates cultripes), pertanyent a l’espècie Polystoma pelobatis. La fotografia ha estat feta a partir d’exemplars trobats als Pirineus catalans.
Santiago Mas-Coma.
Els monogenis són platihelmints aquàtics de cicle biològic simple. D’hoste únic, la majoria són paràsits externs de peixos, marins i d’aigua dolça, però també n’hi ha que parasiten crustacis i cefalòpodes; d’altres són paràsits interns d’amfibis, quelonis i rarament (una sola espècie) de mamífers. Externament, el seu cos és allargat i pla, amb un òrgan característic a la regió posterior del cos, especialitzat en fixar-se a l’hoste, l’opistohaptor. La majoria són molt petits, d’escala mil·limètrica (les dimensions mínimes són de 30 µm i les màximes, de 2 cm), i d’un to grisós o translúcid, encara que el color dels ous, dels òrgans interns o de l’aliment ingerit poden fer que es vegin vermells, rosats, brunencs, groguencs o totalment negres. Tenen un gran interès zoològic i han estat considerats com un grup situat entre els trematodes i els cestodes.
Els monogenis no són paràsits de l’home, però l’afecten indirectament, ja que originen grans pèrdues en espècies de peixos d’interès comercial, tant en la pesca marina com en l’aqüicultura. Un gran nombre d’espècies (Dactylogyrus) són patògenes per als alevins de peixos de les piscifactories, i la seva infestació origina pèrdues de sang, erosió dels epitelis i dona accés a infeccions fúngiques i bacterianes que els arriben a matar; d’altres (Gyrodactylus) desencadenen hiperinfestacions en el mateix hoste, i són la causa de grans pèrdues, també en les piscifactories. Hi ha exemples semblants que afecten la pesca marina. Per exemple, el monogeni Nitzschia sturionis fou introduït, juntament amb el seu hoste, l’esturió Acipenser stellatus, a la mar d’Aral, on va produir un brot infecciós a l’espècie local d’esturió, A. nudiventris, que no n’havia estat afectat mai abans per aquest paràsit i que va patir una reducció alarmant del nombre d’exemplars de les seves poblacions.
Morfologia
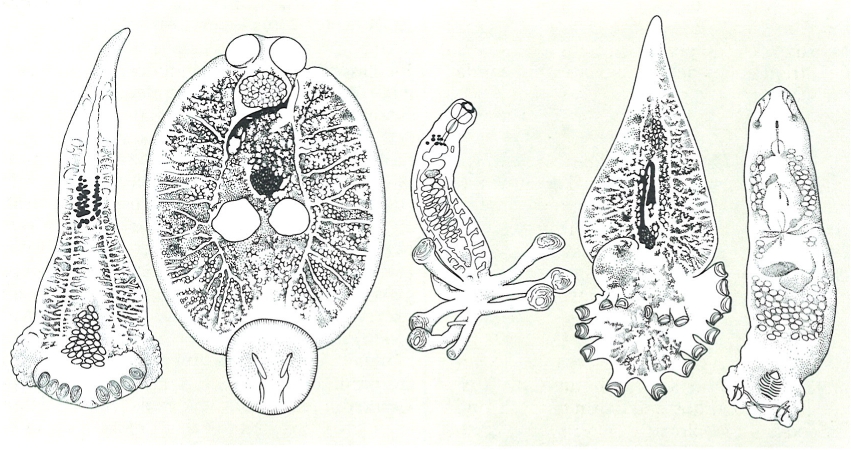
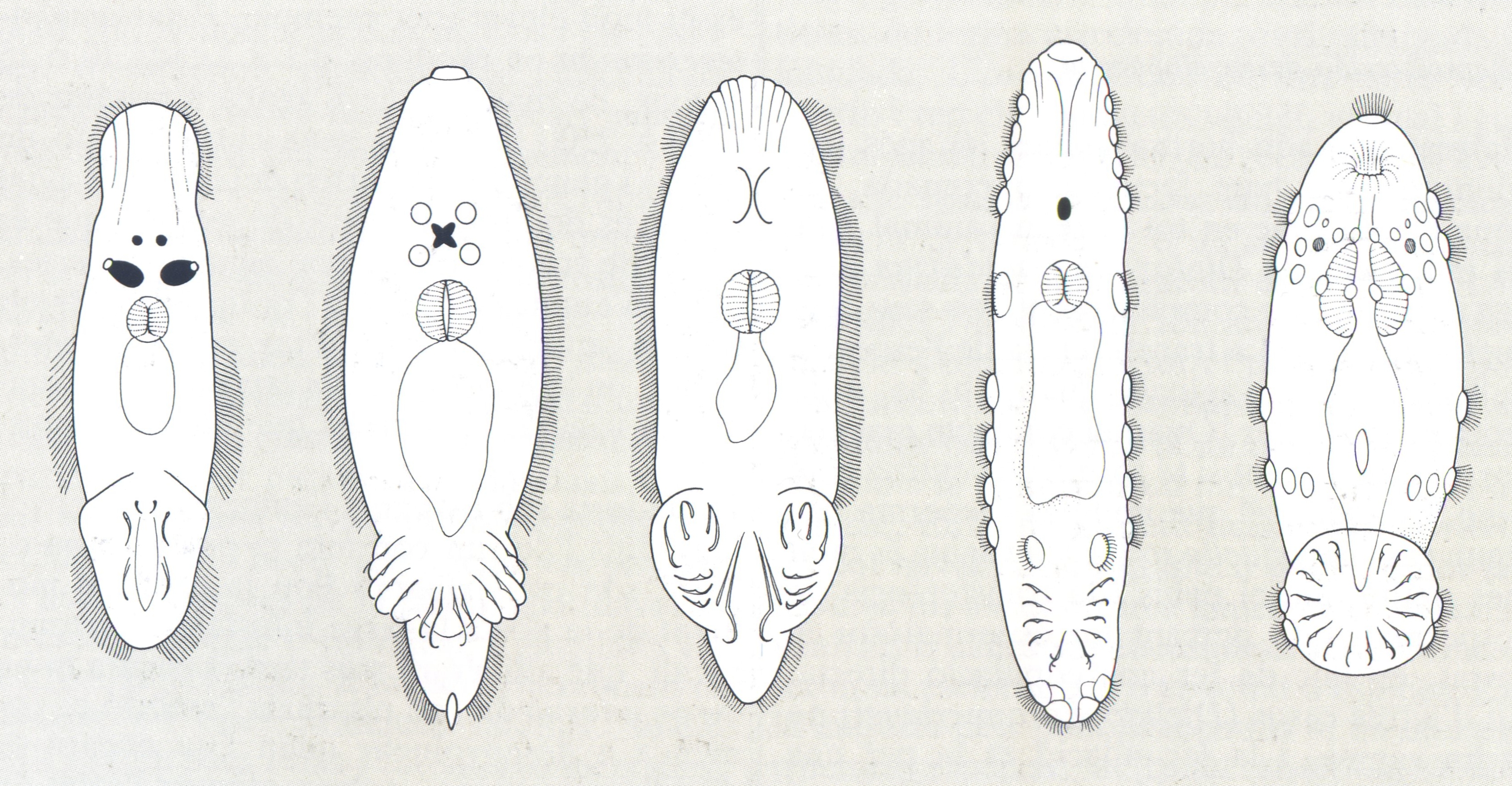
Diversos tipus morfològics dels monogenis, que mostren els diferents models d’opistohaptor.
Biopunt, a partir de fonts diverses
El cos és de simetria bilateral, encara que en algunes espècies pot ésser lleugerament alterada. Té una regió cefàlica, portadora del prohaptor i anterior a la faringe, el tronc o cos pròpiament dit, el peduncle i, al final del cos, l’opistohaptor.
El prohaptor és el conjunt d’òrgans de fixació i d’alimentació, constituït bàsicament per formacions glandulars. Hi ha dos tipus de prohaptor: el que no és connectat amb l’embut bucal i el que sí ho és. El primer es troba en les formes més primitives, l’extrem cefàlic de les quals acostuma a ésser truncat, lobulat o ample i arrodonit; generalment presenta glàndules cefàliques, que secreten una substància enganxosa per a fixar-se a l’hoste, llargues i abundants microvellositats, ben diferents de les de la resta del cos, i de vegades botris musculosos profunds, que serveixen de ventoses. El segon tipus de prohaptor comporta l’especialització de la boca i l’embut bucal; en el cas més simple hi ha una ventosa oral que envolta la boca; altres vegades poden ésser dos òrgans o ventoses bucals dins les parets de l’embut, proveïdes de components musculars, glandulars i sensorials, els quals, sembla, tenen algun paper en l’alimentació de les espècies hematòfagues.
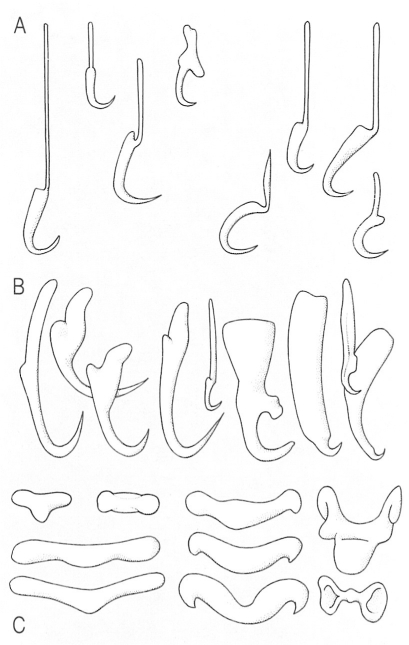
Diferents models d’armadura de l’opistohaptor dels monogenis, situat al final del cos: A ganxos marginals, B ganxos centrals, C esclerites accessòries o barres connectives. La disposició dels ganxos (àncores o hamuli), la presència de pinces i òrgans adhesius configuren en conjunt les característiques d’aquest òrgan de fixació adequades a cada cas.
Biopunt, a partir de fonts diverses
A l’extrem posterior del cos, l’opistohaptor pot ésser ben separat pel peduncle o bé no ésser més que un eixamplament, que de vegades s’estén cap endavant per la part ventral. Durant l’ontogènia es diferencien els dos tipus principals d’opistohaptors. La larva que surt de l’ou sempre té un opistohaptor minúscul armat de ganxos o espines esclerotitzats. En el primer tipus, aquest opistohaptor larval pot mantenir-se o evolucionar vers l’òrgan adult, o mantenir-se jove i que aparegui al costat un altre òrgan nou que donarà lloc a l’opistohaptor definitiu de l’adult. Aquest opistohaptor és un gran disc amb lòculs poc profunds o ventoses ben desenvolupades, amb grans ganxos centrals anomenats àncores o hamuli i petits ganxos generalment marginals. En el segon tipus pot perdre’s completament l’opistohaptor larval, o mantenir-se petit sobre o dins l’opistohaptor adult o fixat al final d’un apèndix muscular. Aquest opistohaptor és més especialitzat, amb òrgans adhesius individualitzats i/o amb pinces esclerotitzades, valves musculars o ganxos suplementaris.
La paret externa dels monogenis és un tegument viu, semblant al dels digenis i els cestodes, d’estructura sincítica. La musculatura principal del cos és feta de músculs superficials situats just per sota del tegument i disposats en capes circulars, diagonals i longitudinals; la de l’opistohaptor és molt important per a l’adhesió de l’animal al seu hoste. L’aparell digestiu és molt simple: la boca i l’embut bucal solen tenir ventoses; més avall s’obre una prefaringe curta, seguida d’una faringe muscular i glandular, que constitueix un aparell xuclador potent. Les glàndules faríngies secreten una forta proteasa que degrada l’epidermis de l’hoste (en xuclen els productes lisats). Després de la faringe pot haver-hi un esòfag, simple o ramificat, i glàndules digestives unicel·lulars. L’intestí generalment es divideix en dues branques laterals, sovint molt ramificades i fins i tot interconnectades. No hi ha anus. El sistema nerviós correspon al model general, amb ganglis cerebrals dels quals surten troncs nerviosos que van cap a la regió posterior, connectats transversalment per comissures. Els òrgans adhesius són ben innervats i en general hi ha una gran varietat d’òrgans sensorials. La majoria de les espècies tenen ulls de tipus rabdomèric (com els turbel·laris i algunes larves de digenis) en l’estat larval aquàtic lliure: quatre taques oculars en els monopistocotilis, que poden persistir en l’adult una mica reduïdes, i dues en els poliopistocotilis, que es perden durant el procés de maduració. S’han descrit diversos òrgans sensorials ciliars. Pel que fa al sistema excretor, la unitat de funcionament és el protonefridi cel·lular flamíger, típic dels platihelmints; externament, s’observen dos porus excretors laterals, prop de l’extrem anterior de l’animal.
L’aparell genital masculí inclou generalment un sol testicle (excepcionalment, n’hi ha diversos, fins al cas extrem, d’una espècie que en té 200). De cada testicle surt un conducte eferent que s’eixampla i es converteix en conducte ejaculador . No sembla que hi hagi cap formació semblant a la bossa del cirrus o al cirrus evaginable dels trematodes i els cestodes; però mentre que en alguns casos el conducte ejaculador és simple i acaba en un atri genital poc profund, sovint amb una ventosa, en d’altres té els marges esclerotitzats i, finalment, en d’altres els teixits del final del conducte ejaculador, musculosos i engruixits, formen una mena de penis, moltes vegades armat de ganxos de mida i forma variable. Alguns tenen una vesícula seminal sacciforme simple i la majoria tenen glàndules prostàtiques unicel·lulars. En famílies determinades hi ha un altre tipus d’òrgan copulador, que reuneix el conducte ejaculador amb un aparell copulador esclerotitzat d’estructura complexa. El sistema genital femení comprèn sempre un sol ovari, generalment situat per davant dels testicles, de forma arrodonida, oval, allargada o lobulada. L’oviducte que en surt corre vers l’ootip i rep els conductes vitel·lí, vaginal i genitointestinal al llarg del seu recorregut. El receptacle seminal pot ésser un simple eixamplament de l’oviducte o un sac especial amb un conducte separat que hi desemboca. Les glàndules vitel·lògenes són abundants. Generalment s’estenen per tot el parènquima i sovint fins i tot per l’opistohaptor; cada grup de glàndules vitel·lògenes té un conducte eferent i es fon a la regió ventral mitjana prop de l’oviducte, formant un petit magatzem de vitel. Hi pot haver vagina simple o doble, o pot no haver-n’hi; les obertures poden ésser dorsals, ventrals o laterals, generalment amb el tracte terminal esclerotitzat o envoltat d’espines. En la majoria dels poliopistocotilis hi ha una estructura especial, el canal genito-intestinal, que connecta l’oviducte i una branca de l’intestí, la funció del qual es desconeix, bé que hi ha diverses hipòtesis. L’ootip no és sinó una expansió muscular del conducte femení. La glàndula de Mehlis comprèn cèl·lules mucoses i cèl·lules seroses i se suposa que té alguna funció en l’origen i la producció dels precursors de l’ou, semblant a la que tenen els digenis. A continuació de l’ootip hi ha l’úter, que es dirigeix cap endavant fins a obrir-se a l’atri genital, juntament amb el conducte ejaculador.
Biologia
Reproducció i desenvolupament
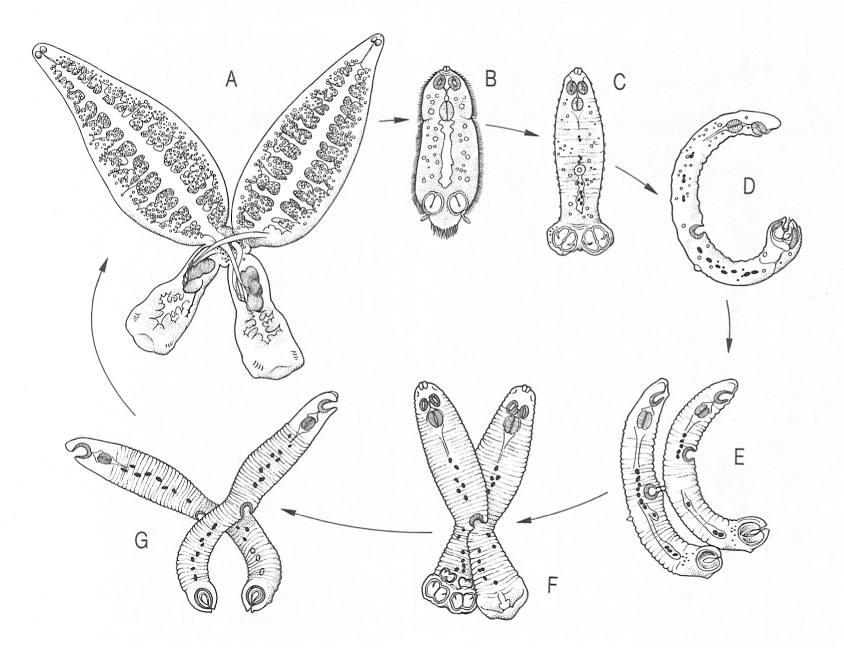
Fases del desenvolupament de Diplozoon paradoxum, un monogeni paràsit de peixos d’aigua dolça que té característiques biològiques excepcionals dins el seu grup, pel fet que els adults arriben a fusionar totalment els seus conductes genitals per transferir-se l’esperma mútuament. Hom ha representat en el dibuix: A adults fusionantse en còpula permanent, B larva nedadora acabada de sortir de l’ou, C larva diporpa, D-G larves diporpes adherint-se les unes a les altres.
Biopunt, a partir de fonts diverses
Malgrat que són hermafrodites, és més freqüent que hi hagi fecundació creuada. Es dona el cas, únic en tot el regne animal, que els individius de Diplozoon paradoxum arriben a fusionar-se completament pels seus conductes genitals. La transferència de l’esperma té lloc als porus vaginals, quan n’hi ha, o directament a l’úter. Quan el conducte ejaculador és simple, tant si té forma de ventosa com si no, és aquest el que impulsa l’esperma. En algun cas (Entobdella soleae) la vagina és molt més petita que el penis i la transferència de l’esperma es fa deixant un espermatòfor al costat, en lloc de fer una copulació directa. En d’altres casos (Diclidophora merlangï) no hi ha vagina, i la fecundació es fa per una mena d’impregnació hipodèrmica. Quan el penis té forma de ventosa, s’adhereix a l’altre animal per darrere dels porus genitals i amb les espines arrenca una petita part del tegument; l’esperma penetra a través de la ferida oberta i s’obre pas (1 o 2 mm) cap al receptacle seminal. Després de la fecundació, el zigot i les cèl·lules vitel·lines arriben a l’ootip, on es comencen a formar els ous. Els ous poden tenir formes molt variades (ovalats, triangulars, allargats, tetraèdrics, etc.), però sempre tenen un opercle en un pol i sovint un filament adhesiu o dos (un a cada pol) per a enganxar-se entre ells o per a fixar-se a l’hoste o a qualsevol substrat. El nombre d’ous pot ésser molt elevat (per exemple, Polystoma integerrimum en produeix d’1 a 3 cada 10 o 15 segons, de manera que n’arriba a produir de 2000 a 2500 durant la seva curta estació de posta) i generalment surten del cos molt aviat (a través de l’úter i l’atri), de manera que és difícil de trobar-n’hi gaires; algunes espècies, però, n’emmagatzemen a l’ootip i després els deixen anar a través d’un porus. L’úter, doncs, en la majoria de les espècies no té funció de vagina, al contrari que en els digenis.

Tipus d’ous en els monogenis. Noteu la forma general ovalada dels ous, i els apèndixs filamentosos i els esperons que duen per a facilitar la seva fixació. Dels que no tenen opercle, l’embrió en surt trencant la paret per una zona més fina.
Biopunt, a partir de fonts diverses
La majoria són ovípars i l’embrió es desenvolupa dins l’ou, un cop feta la posta. Alguns, però, són ovovivípars i l’ou ja conté al moment de la posta la larva infestant o oncomiracidi, que alliberarà més tard. I encara hi ha casos intermedis, en els quals l’embrió es comença a desenvolupar quan l’ou encara és dins l’úter però no arriba a atènyer l’estat de larva infestant fins després de la posta. També n’hi ha que, essent típicament ovípars (Polystoma integerrimum, P. pelobatis), recorren a l’ovoviviparisme en l’últim ou de cada posta. Finalment, n’hi ha de vivípars (tots els girodàctils excepte Oogyrodactylus farlowellae), en els quals l’ou s’obre quan encara és dins l’úter, i en surt una larva lliure, que es desenvolupa fins a l’estat jove i no surt a l’exterior fins que no és un subadult funcional, però amb la particularitat que, dins aquest jove sovint es pot veure un segon, un tercer (dins el segon) i fins un quart estadi jove (dins el tercer) per un tipus d’embriogènesi encara no prou conegut, potser poliembrionia seqüencial, capaç de donar lloc fins a quatre individus a partir d’un mateix zigot; en aquests casos de viviparisme no es passa per l’estat de larva oncomiracidi de vida aquàtica lliure i el jove nascut s’instal·la directament al mateix hoste, al costat del seu predecessor, i dona lloc al naixement de següent estadi jove que tenia al seu interior. És llavors que un ou del seu ovari pot tornar a ésser fecundat i repetir-se la seqüència.
Diferents oncomiracidis de monogenis.
Biopunt, a partir de fonts diverses
El més corrent és que es passi per una fase larval única, l’oncomiracidi (antigament giractiloide), que fa vida aquàtica lliure. Externament, aquesta larva recorda un protozou, amb cèl·lules ciliades distribuïdes de vegades en tres zones diferenciades; excepcionalment, hi ha oncomiracidis no ciliats (els girodactílids vivípars i els ovípars Acanthocotyle lobianchi, Sphyranura oligorchis, Epicotyle torpedinis, Microcotyle salpae, entre altres). L’oncomiracidi també té glàndules cefàliques i taques oculars, i un nombre variable de papil·les sensorials o sensors argentòfils (papil·les sensorials tegumentàries que s’impregnen de nitrat d’argent), els models de distribució de les quals són d’interès taxonòmic (quetotàxia). Internament, l’oncomiracidi té l’aparell digestiu ben diferenciat, els porus excretors ja formats i un òrgan opistohaptor larval, amb ganxos i esclerits, amb el qual s’adherirà a l’hoste abans de començar a perdre les cèl·lules ciliades per completar el desenvolupament.
Generalment s’anomena postlarva la fase que va del moment en què l’oncomiracidi es fixa per primera vegada a la paret externa de l’hoste al moment en què es converteix en preadult i es fixa al seu microhàbitat definitiu. Aquesta fase es caracteritza, en els monoopistocotilis, per una migració des del punt inicial de contacte fins al microhàbitat final (en els poliopistocotilis no hi ha cap migració, ja que l’oncomiracidi es fixa directament al microbiòtop branquial definitiu). Morfològicament, la fase de postlarva es caracteritza per la transformació de l’opistohaptor simple de l’oncomiracidi en l’opistohaptor complex de l’adult; en els poliopistocotilis aquesta transformació és uniforme (procés anomenat "metamorfosi"), amb desaparició de certs ganxos larvals i formació d’òrgans nous (les pinces); en els monoopistocotilis no hi ha transformacions profundes i sembla que hi ha dos tipus de desenvolupament: en un cas (en els dactilogírids) hi ha modificacions induïdes per l’aparició d’estructures com els hàmuls en una adaptació dependent del mode d’adherència i de la naturalesa del substrat, i en l’altre (en els monocotílids, capsàlids, acantocotílids i girodactílids) no hi ha modificacions importants, ja que l’adherència continua essent de tipus cutani (sense excloure el parasitisme branquial).
En el cas d’algunes espècies (polistomàtids) paràsites d’amfibis, hi ha unes formes intermèdies entre l’estadi d’oncomiracidi de vida lliure i el d’adult paràsit de la bufeta urinària, que reben el nom de larves de creixement lent i formes neotèniques. Les larves de creixement lent són aquelles que, quan passen a les brànquies del cap-gros, canvien poc i més tard emigren cap a la bufeta urinària de l’amfibi jove, quan el cap-gros fa la metamorfosi, per a convertir-se en paràsit adult; llavors, les transformacions inclouen la desaparició de les cèl·lules ciliades, el creixement lent dels hàmuls, l’aparició molt lenta de ventoses al voltant dels ganxos i el canvi d’aspecte de les glàndules cefàliques, que esdevenen refringents. Les formes neotèniques són paràsits deles brànquies, de creixement ràpid, que s’originen en el cap-gros a partir de l’oncomiracidi, hi arriben a madurar, a fecundar-s’hi i a produir-hi ous viables, i moren just al moment que el cap-gros comença la metamorfosi; sempre són més petites que els adults, no tan planes, i tenen els hàmuls molt reduïts. En realitat són formes realment diferents, tant morfològicament com anatòmicament. En els polistomàtids també hi ha unes formes, intermèdies entre els individus neotènics perfectes (sense hàmuls) i els verms adults (amb hàmuls), que reben el nom de formes metaneotèniques, que s’originen a les brànquies quan el cap-gros és infestat per oncomiracidis en el moment en què ja inicia la metamorfosi. Les formes intermèdies entre l’estadi d’oncomiracidi i l’estadi neotènic reben el nom de metagirodactiloides o metaoncomiracidi, i no són altra cosa que larves que han començat a absorbir sang de les brànquies de l’hoste però el desenvolupament neotènic de les quals ha quedat inhibit.
El cicle vital
Els coneixements sobre els cicles vitals dels monogenis es limiten a algunes espècies. Però en general es pot afirmar que segueixen un cicle simple, directe i monoxè, amb un sol hoste, i que s’hi distingeixen les tres fases essencials, d’ou, oncomiracidi i adult, paràsit d’un únic hoste i, per tant, hoste definitiu. No obstant això, sembla que hi ha un gran nombre de variacions i modalitats de cicles, i fins i tot hi ha la prova que dues espècies de gastrocotílids paràsits de peixos depredadors passen per un període de desenvolupament en un peix que és presa habitual del peix que esdevindrà el seu hoste definitiu. Entre els cicles més ben coneguts, podem distingir-ne dos tipus: els dels paràsits de peixos i els dels paràsits de tetràpodes.
Cicle biològic dels monogenis paràsits de peixos
Cicle biològic del monogeni Benedenia melleni, paràsit de peixos. A Ous, B oncomiracidi acabat de sortir de l’ou, C oncomiracidi que ja ha perdut la coberta ciliada i ha desenvolupat l’opistohaptor, D adult jove, paràsit d’un peix, E adult. A baix a la dreta hom ha indicat les vies d’infestació del peix hoste pels dactilogírids (paràsits branquials).
Biopunt, a partir de fonts diverses
En el model més general, els adults paràsits del peix són ovípars i deixen anar els ous, sense embrionar, al medi aquàtic extern; allà, els ous s’embrionen i quan eclosionen en surt l’oncomiracidi, que neda activament fins a trobar el peix hoste definitiu. Però aquest model general té moltes variacions, que representen estratègies ecològiques i etològiques molt diverses.
En primer lloc, hi ha espècies (Dactylogyrus vastator) que fan ous sense filaments i d’altres que en fan amb filaments (Acanthocotyle lobianchi, A. greeni i diverses espècies de Dionchus i Lamellodiscus). Sembla que en alguns casos aquests filaments serveixen per a adherir-se als altres ous de la posta i formar petits raïms molt lleugers, que poden ésser arrossegats amb l’aigua que entra per les brànquies del peix, però en d’altres casos (Dionchus agassizi i D. remorae, que parasiten Echeneis naucrates) sembla que serveixen perquè quedin adherits al mateix hoste que ocupa el progenitor.
En la gradació que va de l’oviparisme al viviparisme, passant pel pas intermedi de l’ovoviviparisme, s’observa una certa tendència a reduir el cicle biològie en l’espai. El més comú és que el monogeni sigui ovípar i que tingui un úter curt, que li impedeix de retenir els ous, que, per tant, ha de deixar anar contínuament. Amb tot, alguns monogenis, també ovípars, poden retenir els ous a l’úter (microcotílids i hexabòtrids) i deixar anar de cop postes nombroses; aquest és un cas intermedi entre l’oviparisme i l’ovoviviparisme autèntic (Callorhynchicola multitesticulatus, Lithidiocotyle seconda, Plectanocotyle gurnardi, Carangoides ovovivipara o Oogyrodactylus farlowellae). El cas extrem és el del viviparisme dels girodactílids, en els quals s’esdevé la formació de l’individu jove perfectament conformat quan encara és dins l’úter del progenitor, de manera que es pot desenvolupar un segon jove dins aquest i fins i tot un tercer dins el segon i un quart dins el tercer, per un mecanisme de poliembrionia seqüencial.
Entre les espècies que passen per una etapa larval, podem diferenciar dos casos: amb l’oncomiracidi ciliat i, per tant, amb capacitat natatòria, i amb l’oncomiracidi no ciliat. En el primer cas és possible la infestació activa del peix hoste i, en aquest sentit, és important l’adaptació del cicle del monogeni, que fa coincidir el seu moment de producció d’ous amb el moment de la reproducció del peix, perquè això facilita l’abundància d’hostes disponibles. En el segon cas, quan l’oncomiracidi no és ciliat, com per exemple en Oogyrodactylus farlowellae, paràsit d’un peix gat sudamericà que viu en corrents molt ràpids, l’ou, embrionat al moment de la posta queda adherit al substrat gràcies a unes gotes adhesives i infesta el peix quan aquest va a fixar-se per la boca al mateix substrat.
Pel que fa a les vies d’infestació, també hi ha dues modalitats. En la primera, sembla que els oncomiracidis penetren d’una manera passiva en la cambra branquial de l’hoste, amb el corrent d’aigua respiratòria, com passa en els poliopistocotilis. En la segona, típica de diverses espècies de monopistocotilis, hi ha un contacte inicial de l’oncomiracidi amb la pell del peix i a continuació una migració secundària de la postlarva cap al microhàbitat branquial definitiu; encara no se sap, però, si aquest procès és general.
Hi ha, encara, el cas particular de les espècies del génere Diplozoon, paràsits de les brànquies de ciprínids, on es dona la fusió de dos individus. Els ous d’aquesta espècie tenen un filament polar molt llarg i fi, gràcies al qual s’entortolliguen formant petits raïms, que acaben caient al fons; les larves ciliades, capaces de sobreviure fins a 6 hores lliurement, són aspirades amb el corrent inhalant i van a fixar-se a les brànquies, on perden els cilis i passen la metamorfosi, durant la qual els creix una petita ventosa circular a la cara ventral i un petit sortint de la cutícula a la cara dorsal. Quan dues d’aquestes larves, que reben el nom de diporpa, es troben, cadascuna s’agafa al sortint dorsal de l’altra per mitjà de la seva ventosa, es fusionen pel seu punt d’inserció comú i inicien la metamorfosi conjuntament, per la qualo es fusionen els conductes sexuals masculins i femenins i queden acoblades definitivament; llavors, la fecundació sempre és creuada.
Cicle biològic dels monogenis paràsits de tetràpodes
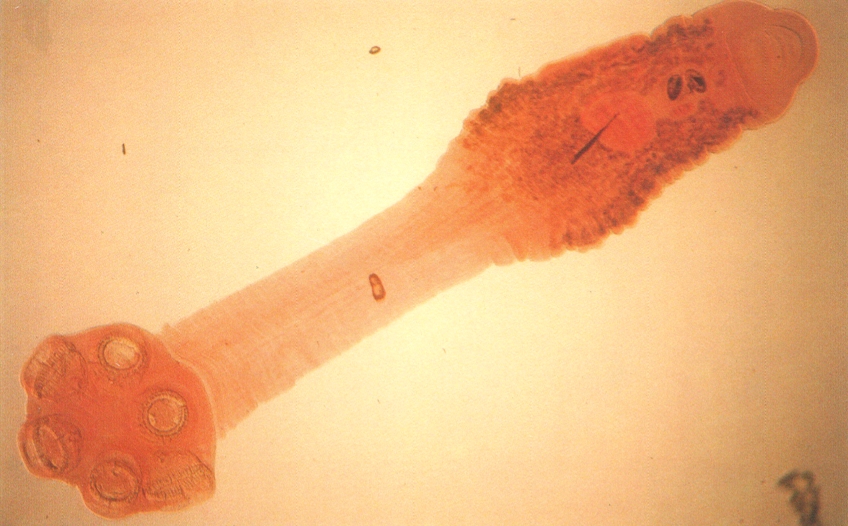
El monogeni Oculotrema hippopotami de la fotografia, paràsit de l’hipopòtam a l’Àfrica, permet d’observar la forma del cos i de l’opistohaptor.
Santiago Mas-Coma.
Els monogenis que han esdevingut paràsits de tetràpodes d’hàbits amfibis pertanyen principalment als poliopistocotilis i, dins aquests, a les famílies dels polistomàtids (Polystomatidae), principalment paràsits interns d’anurs, i esfiranúrids (Sphyranuridae), paràsits externs d’urodels. Entre els monopistocotilis hi ha únicament alguns girodactílids (Gyrodactylidae) paràsits de brànquies de cap-grossos d’anurs i la cavitat bucal d’anurs adults, i alguns de la família dels iagotremàtids (Iagotrematidae), de la bufeta urinària de quelonis i urodels.
Dels monopistocotilis hom coneix força bé el cicle d’un iagotremàtid, Euzetrema knoepffleri, que viu a la bufeta urinària de l’urodel Euproctus montanus, de Córsega. És ben simple: els ous no embrionats surten a l’aigua i s’incuben en unes 6 setmanes, després de les quals en surt un oncomiracidi ciliat capaç de nedar, que infesta un nou hoste entrant-hi directament per la cloaca; a continuació, puja pels urèters fins als ronyons, on la postlarva, després de nodrir-se durant uns 2 mesos, torna a baixar pels urèters fins a la bufeta urinària, i hi madura fins que esdevé el paràsit adult.
En els polistomàtids la complicació dels cicles és, en canvi, extraordinària. Hi ha una gran varietat de possibilitats de cicles, adaptats a tota mena de condicions ecològiques. El cicle més simple i més generalitzat és el que no inclou ni estadis neotènics ni postlarves migradores, ni cicle intern, ni periple renal, com tampoc dualitat larval. Així, per exemple, en el cicle de Protopolystoma xenopi, una espècie paràsita del gripau Xenopus laevis, de l’Àfrica tropical, i el d’Eupolystoma alluaudi, paràsit d’un altre gripau africà, Bufo regularis, els adults ponen ous ininterrompudament, i els oncomiracidis penetren directament a la cloaca dels gripaus joves.
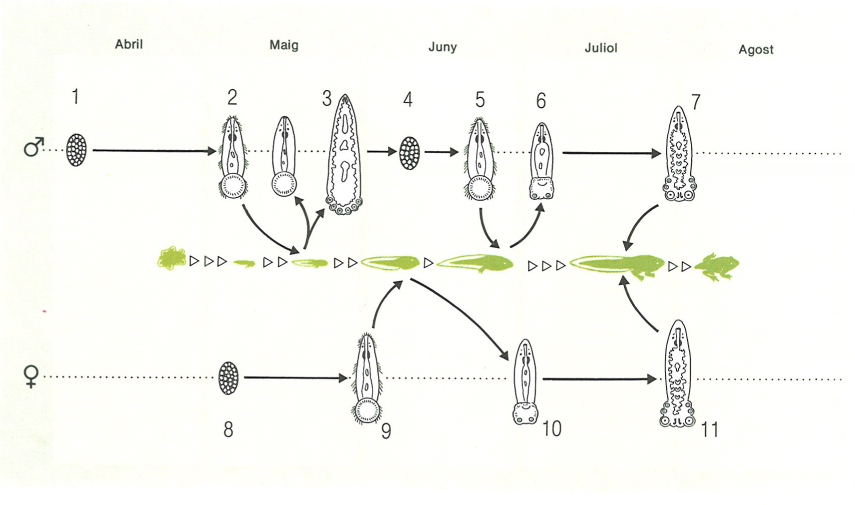
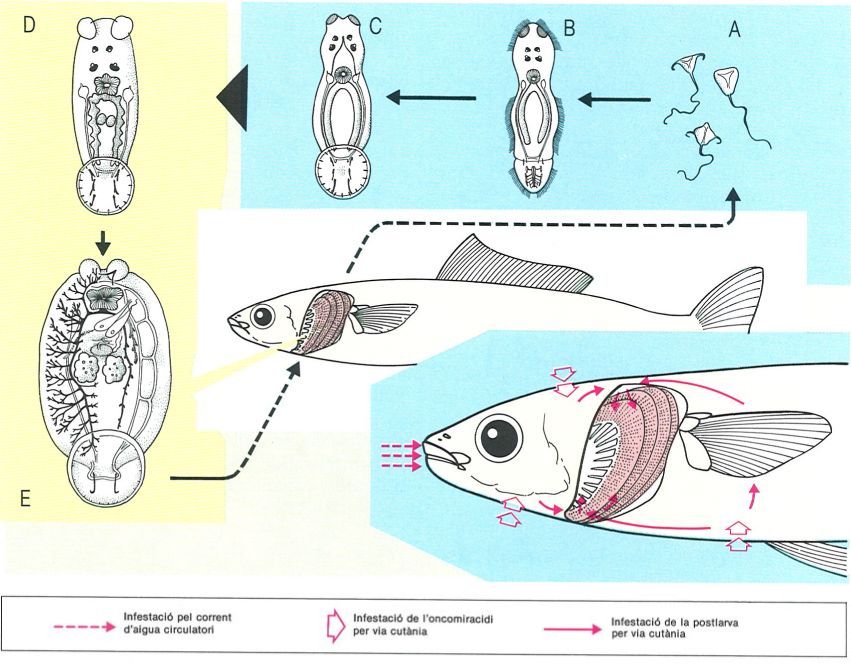
Cicle biològic de Polystoma integerrimum, paràsit de la bufeta urinària de la granota roja (Rana temporaria). 1 Ous del paràsit alliberats a l’aigua pels paràsits dels mascles de la granota, al principi de l’abril; 2 larves que, provinents de la línia «masculina» (granota mascle), podran adherir-se a les brànquies dels cap-grossos joves i transformar-se en neotènics (3), productors d’ous (4) que es desenvolupen més de pressa gràcies a la temperatura superior de l’aigua, i donen larves (5) que es fixen a capgrossos més desenvolupats, que fan la metamorfosi aviat (6), amb la qual cosa el paràsit passa de la cavitat branquial a la bufeta urinària (7); 8 ous del paràsit alliberats a l’aigua pels paràsits de les femelles de la granota, les quals s’uneixen als mascles al final de l’abril; 9 larves que surten de la línia «femenina», que es troben directament amb cap-grossos més desenvolupats i no donen generació neotènica, sinó que es transformen directament en formes capaces d’arribar a la bufeta en el moment que el cap-gros fa la metamorfosi (10, 11).
Biopunt, a partir de fonts diverses
Altres polistomàtids són de cicle directe però passen per formes neotèniques, com per exemple Polystoma integerrimum, paràsit de la bufeta urinària de la nostra granota roja (Rana temporaria) dels Pirineus catalans. Els ous dels polistomes surten a l’exterior en el moment de la posta de la granota. Com que els mascles de les granotes inicien la fresa abans que les femelles, els ous de polistoma que infestaven els mascles de la granota inicien el seu desenvolupament a l’aigua abans que els ous procedents dels paràsits de les femelles. Els oncomiracidis es fixen a les brànquies externes dels cap-grossos i s’alimenten de sang i, quan es forma la cambra branquial, hi queden tancats i es converteixen en individus neotènics, és a dir, amb caràcters larvals però ja capacitats per a reproduir-se. L’individu neotènic només produeix un ou, i l’oncomiracidi ciliat que en surt es fixa a les brànquies d’altres cap-grossos, els quals mai no donaran formes neotèniques. Quan el cap-gros inicia la metamorfosi, les postlarves dels polistomes surten a l’exterior pel forat que formen les potes del davant de la granota en emergir de la cavitat branquial, i a continuació recorren els flancs de la granota i hi penetren pel forat cloacal, fins a la bufeta urinària, on es converteixen en polistomes adults. Hi ha, doncs, d’una banda, un cicle directe amb neotènics o cicle neotènic i, de l’altra, un cicle amb intervenció d’una postlarva migratòria i d’adults a la bufeta, o cicle vesical.
En alguns polistomàtids hi ha un cicle intern directe, com per exemple en Polystoma integerrimum, paràsit de la granota roja (Rana temporaria), en P. pelobatis, paràsit del gripau d’esperons (Pelobates cultripes), ambdues dels Pirineus catalans, i en les espècies africanes Eupolystoma alluaudi, paràsit de Bufo regularis, i Metapolystoma cachani, paràsit de Ptychadena longirostris. En aquest cicle, el polistoma adult paràsit de la bufeta es converteix de cop d’ovípar en ovovivípar, gairebé en vivípar, perquè després de deixar anar una multitud d’ous sense embrionar en guarda un de sol, l’últim, dins l’úter, on desenvolupa completament l’embrió i on finalment eclosiona, de manera que en aquest cas surt del progenitor una larva conformada, que directament va a fixar-se a la bufeta del mateix hoste, al costat del progenitor. Però en espècies com Eupolystoma alluaudi i Metapolystoma cachani, aquesta larva que surt del progenitor, abans d’instal·lar-se a la bufeta urinària, fa tot un viatge o cicle renal pujant pels urèters fins al ronyó i després baixa altra vegada fins a la bufeta. Tant en aquest cas com en els anteriors, es tracta d’un cicle d’autoinfestació endògena (no surt el paràsit a l’exterior) que porta a la hiperinfestació del mateix hoste. En el cas de M. cachani coexisteixen, a més, diverses d’aquestes modalitats: cicle neotènic, cicle vesical, cicle intern i periple renal. La plasticitat d’aquests cicles biològics permet moltes possibilitats d’infestació a hostes diferents. En generai, la facilitat de passar d’ovoviviparisme a viviparisme els permet d’adaptar-se a condicions d’espai i temps extremes i, per tant, ajuda a assegurar la perpetuació de l’espècie.
Val la pena de citar també el cas extrem de Pseudodiplorchis americanus, paràsit de la bufeta urinària del gripau Scaphiopus couchii dels deserts nord-americans. Aquest gripau viu enterrat 10 mesos l’any i només surt després de pluges intenses. Fresa la mateixa nit i abandona l’aigua dos o tres dies després; durant els aproximadament dos mesos que fa vida terrestre no entra a l’aigua per res. El monogeni només pot infestar-lo durant els dos o tres dies que dura la fresa, i només durant el dia (perquè de nit el gripau surt de l’aigua), és a dir, aproximadament durant unes 12 o 18 hores l’any. El monogeni arriba a formar fins 250 ous, que es desenvolupen al llarg de tota l’estació de letargia de l’hoste, és a dir, durant la tardor i l’hivern; aquests ous surten del cos del progenitor tots de cop en el moment de la posta del gripau, eclosionen immediatament i infesten els gripaus reunits en el lloc.
Finalment, cal esmentar la dualitat morfològica de l’oncomiracidi d’espècies d’Eupolystoma: després de la posta eclosionen larves ciliades que asseguren la transmissió d’un hoste a l’altre, i dins l’úter eclosionen larves no ciliades que surten i es fixen al mateix hoste, sense que intervinguin cap-grossos en el cicle.
Filogènia i sistemàtica
Malgrat que durant molt de temps s’ha considerat els monogenis (amb els digenis i els aspidogastres) com un ordre o una subclasse dels trematodes, actualment es parla més de la seva proximitat als cestodes, sobretot per la semblança química i estructural dels ganxos dels oncomiracidis i dels estadis larvals dels cestodaris i eucestodes. Hom considera que els digenis i els aspidogastres van sorgir separadament dels monogenis. Es parla d’una evolució paral·lela de la capacitat natatòria dels vertebrats i del sistema d’ancoratge de les larves dels monogenis; se suposa que la colonització del medi terrestre fou posterior a l’aparició dels tetràpodes amfibis.
No hi ha una classificació unànimement acceptada dels monogenis. En general però, hi ha consens a distingir, dins la classe dels monogenis (Monogenea), dos grans grups o subclasses: els monopistocotilis (Monopisthocotylea) i els poliopistocotilis (Polyopisthocotylea).
Dins els monopistocotilis s’inclouen els grups (considerats també ordres per alguns autors) dels dactilogírids (Dactylogyridae), els ancirocefàlids (Ancyrocephalidae), calceostomàtids (Calceostomatidae), els diplectànids (Diplectanidae) i els tetraònquids (Tetraonchidae), paràsits de teleostis; els monocotílids (Monocotylidae) i capsàlids (Capsalidae), paràsits de celacis; i els acantocotílids (Acanthocotylidae), de selacis i teleostis. El grup dels iagotremàtids (Iagotrematidae), paràsits d’amfibis, representa una família relicta intermèdia entre els monopistocotilis i els poliopistocotilis, bé que hi ha qui tendeix a incloure-la dins els primers. Els girodactílids (Gyrodactylidae), paràsits de peixos, amfibis, cefalòpodes i crustacis, són també de posició intermèdia i conflictiva. Els udonèl·lids (Udonellidae), paràsits de peixos marins i de copèpodes paràsits de peixos marins, també de posició discutida, solen incloure’s en els monopistocotilis.
Els poliopistocotilis inclouen com a grups principals els polistomàtids (Polystomatidae), paràsits de tetràpodes; els hexabòtrids (Hexabothriidae), paràsits de selacis; els diclidofòrids (Diclidophoridae), els discocotílids (Discocotylidae), els gastrocotílids (Gastrocotylidae), els mazocrèids (Mazocraeidae) i els microcotílids (Microcotylidae), tots paràsits de teleostis; els quimericòlids (Chimaericolidae), paràsits d’holocèfals, i finalment els diclibòtrids (Diclybothriidae), paràsits de condrostis. Els diplozoids (Diplozoidae), paràsits de les brànquies de peixos d’aigua dolça, constitueixen un grup d’identitat pròpia dins els poliopistocotilis, però alguns autors els situen dins dels mazocrèids.