
Els elements florals poden prendre aspectes molt variables. El cas més simple, en què els sèpals, exteriors i verds, envolten els pètals, acolorits i de consistència diferent, a l’entorn de les parts fèrtils (gineceu i androceu), no és pas l’únic entre les plantes superiors, ja que les diverses peces poden modificar-se de forma i de color. Les flors de l’aranya (Nigella damascena), que veiem a la fotografia, presenten un involucre de bràctees laciniades disposades a tocar del periant, format per cinc peces petaloides. Segueix un verticil amb cinc petits nectaris de color fosc, un androceu integrat per nombrosos estams i un gineceu pentacarpel·lar amb els estils i els estigmes lliures. Les peces acolorides del periant corresponen en realitat als sèpals, i els nectaris no són altra cosa que pètals transformats.
Enric Curto
Les flors són els òrgans on s’encadenen els esdeveniments més decisius del cicle biològic dels espermatòfits, des de la formació d’espores, passant pel desenvolupament dels gametòfits i la reproducció sexual, fins a la constitució dels embrions i les llavors (i els fruits, si és el cas). Caldrà, per totes aquestes raons, que hi dediquem una atenció particular, començant per la morfologia i continuant pels processos i les transformacions que donen lloc, finalment, a la formació de la llavor. Convé recordar, abans de continuar, que el reconeixement de les particularitats del cicle vital de les plantes superiors, que es relaciona amb el dels pteridòfits heterosporis, no tingué lloc fins fa poc més d’un segle i quart. Aquest fet explica que encara avui utilitzem un vocabulari doble per referir-nos als elements florals, amb un nom històric, que podríem qualificar de morfològic, i un altre que indica la seva situació i la funció dins del cicle biològic. Insistirem sobre aquest punt en parlar del cicle, però podem avançar, per posar un parell d’exemples, que els estams corresponen a microsporofil·les i els grans de pol·len a micròspores.

Els diversos graus de simetria floral es posen de manifest a les fotografies que il·lustren aquest comentari. Les flors (a dalt) de la canya d’Índia (Canna indica), una cannàcia ornamental representada als nostres jardins per nombroses varietats, no tenen cap element de simetria i són, doncs, asimètriques. Al mig, flors zigomorfes, amb un sol pla de simetria, de l’orquídia Himantoglossum longibracteatum. Les espècies d’Iris (a baix) tenen les flors actinomorfes, amb tres plans de simetria i un eix ternari.
Jordi Vidal i Enric Curto
Podem imaginar una flor com una rama de creixement limitat que porta al seu àpex un nombre variable de fulles modificades anomenades antofil·les. Els elements essencials de la flor són les fulles fèrtils, portadores d’esporangis, que poden ser de dos tipus: estams i carpels. El conjunt dels primers s’anomena androceu, mentre que parlem de gineceu per referir-nos als segons. Ben sovint, però quasi exclusivament en el grup de les angiospermes, acompanyen les fulles fèrtils un conjunt de peces estèrils, el periant, que les envolta i les protegeix. Tant les unes com les altres arrenquen de l’eix floral, que de vegades s’insereix directament a la tija o bé, cosa més freqüent, s’hi uneix mitjançant un mànec que rep el nom de peduncle (o bé de pedicel, si la flor fa part d’una inflorescència composta). En el primer cas diem que la flor és sèssil i en el segon, que és pedunculada.
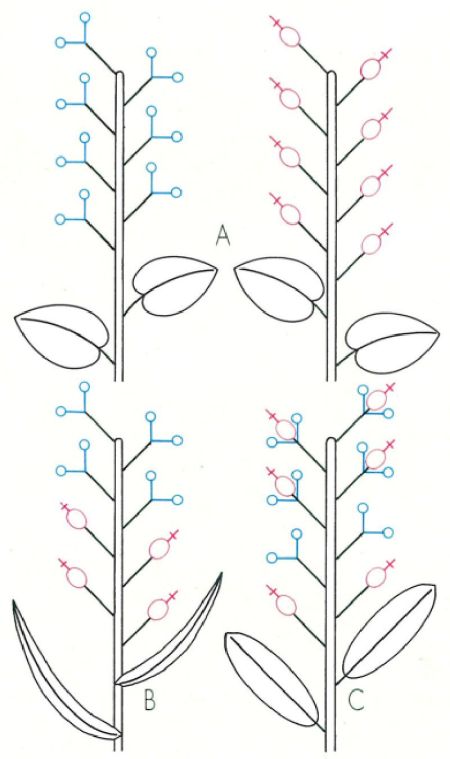
Esquema de la disposició de les flors a les plantes dioiques (A), monoiques (B) i polígames (C). L’androceu és representat de color blau i el gineceu de color vermell.
Mercè Cartañá, a partir de fonts diverses
Habitualment, les flors són hermafrodites, amb androceu i gineceu alhora. Però també sovintegen les flors unisexuals, anomenades masculines si tenen estams però no carpels, i femenines si no presenten androceu. Els espermatòfits més primitius, entre els quals trobem la majoria de les actuals gimnospermes, tenen les flors unisexuals i generalment amb un periant nul o bé rudimentari. Les flors de les fanerògames més evolucionades, en canvi, són molt sovint hermafrodites, com a adaptació al transport del pol·len per animals; només alguns grups que secundàriament han retornat a la primitiva pol·linització mitjançant el vent presenten, entre els espermatòfits més avançats, flors unisexuals. Hi ha vegetals com les alzines, els pins, etc. que tenen flors masculines i flors femenines en un mateix individu; parlem en aquest cas de plantes monoiques. Les plantes dioiques, en canvi, tenen uns peus que només porten flors masculines i uns altres que només en fan de femenines; aquest és el cas dels salzes, del teix i del malcoratge (Mercurialis annua), entre d’altres. Més rarament podem veure plantes polígames, que fan flors unisexuals i hermafrodites en un mateix individu o bé en individus diferents; en són exemple el garrofer (Ceratonia siliqua), el veladre (Veratrum album) i moltes de les compostes que fan part de la nostra flora.

A les plantes monoiques la disposició de les flors masculines i les flors femenines pot ser molt variable. En el càrex (Carex) de la fotografia, les flors s’agrupen en espigues que també són unisexuals; la superior és mas-culina, com posen de manifest els estams sortints; les altres espigues, femenines, mostren els estigmes que ultrapassen les bràctees florals.
Ramon Dolç / Sebastià Hernandis
Si deixem de banda les flors acícliques, amb llurs peces disposades helicoïdalment, la immensa majoria de les flors cícliques de les angiospermes presenten els elements florals, en particular els del periant, ordenats i simètrics. El grau de simetria floral pot ser, tanmateix, divers. Les flors radiades, amb diversos plans de simetria, reben també el nom de regulars o, més pròpiament, d’actinomorfes; en trobem a l’herba fetgera (Hepatica nobilis), a les gencianes (Gentiana) i a moltíssimes altres de les plantes de la nostra flora. Les flors dissimètriques o de simetria bilateral, amb dos plans perpendiculars de simetria, són poc freqüents; en posseeixen la ballarida (Hypecoum imberbe) i, si preníem en consideració l’androceu, la majoria de les crucíferes. La simetria dorsiventral, caracteritzada per l’existència d’un sol pla de simetria, és representada a moltes de les famílies més conegudes, com ara les papilionàcies (ginesta, alfals, etc.) i les labiades (espígol, timó, etc.); aquestes flors reben el nom de zigomorfes. Les flors asimètriques, finalment, no tenen elements de simetria; són pròpies d’algunes plantes tropicals, com ara les Canna, plantades a molts dels nostres jardins.
L’eix floral
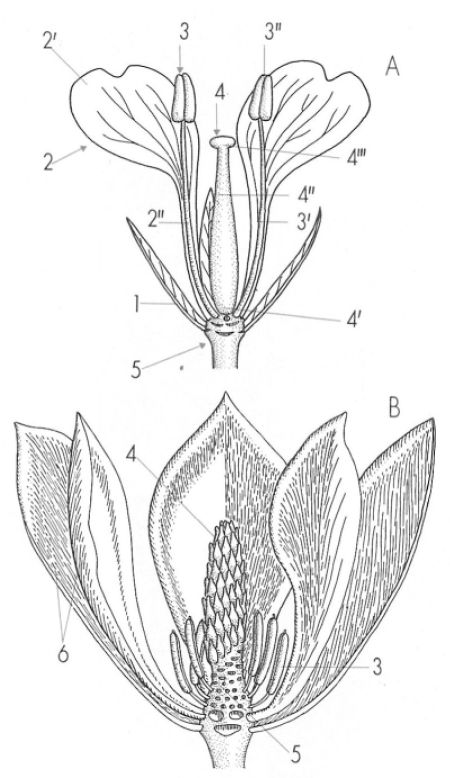
Disposició dels elements florals en una flor cíclica (A) i en una d’acíclica (B). En el primer cas les peces se situen en verticils i en el segon ho fan de manera helicoidal. Hom pot reconèixer: 1 sèpal, 2 pètal (2’limbe, 2" ungla), 3 estam (3’ filament, 3" antera), 4 pistil (4’ ovari, 4" estil, 4’" estigma), 5 receptacle floral, 6 tèpal.
Mercè Cartañà, a partir de fonts diverses
L’eix floral és el suport físic de totes les peces de la flor. Allargat i més o menys cònic a les gimnospermes i a les angiospermes més primitives, es va anar escurçant progressivament en el decurs de l’evolució fins a esdevenir pla o còncau. L’eix floral que té forma de disc, de copa o de tub, propi de les angiospermes més evolucionades, rep el nom de receptacle o també de tàlem. Els receptacles còncaus poden soldar-se amb la base del gineceu, de la qual cosa resulta una estructura poc o molt gruixuda destinada a protegir dels animals els primordis seminals que se situen a l’interior de l’ovari.
Coincidint amb aquest procés de contracció de l’eix, les peces florals passaren d’una disposició inicialment helicoidal a una altra de verticil·lada que mirarem d’explicar breument. En el primer cas, les peces florals es disposen com si fossin fulles esparses separades per entrenusos curtíssims. Si, en canvi, la disposició és verticil·lada, totes les peces d’un mateix tipus surten d’un sol nus (o bé de dos nusos) fent un verticil, o com a màxim dos. Parlem de flors acícliques sempre que les peces es disposen en hèlix. Si la disposició és verticil·lada, cal parlar de flor cíclica. Les flors acícliques tenen un nombre de peces generalment molt elevat i variable, fins i tot dins d’una mateixa espècie. En el cas de les flors cícliques, cada planta sol presentar un nombre definit de verticils i de peces per verticil. El nombre més freqüent de peces és de cinc, cas que correspon als verticils anomenats pentàmers, però també n’hi ha de quatre (tetràmers); a les monocotiledònies sovintegen els verticils trímers, és a dir, de tres peces. Si prenem en consideració el nombre de verticils, les flors pentacícliques i les tetracícliques es compten segurament entre les més abundants. En el primer cas hi ha cinc verticils, dos dels quals solen correspondre a l’androceu, i, en el segon, n’hi ha quatre.
El periant


Diversitat morfològica del periant. El de les tulipes (Tulipa), homoclamidi, es disposa en dos verticils i rep el nom de perigoni perquè les peces (anomenades tèpals) són aproximadament equivalents; la tulipa de la fotografia (a dalt) és una de les nombrosíssimes varietats cultivades. Les flors dels rosers (Rosa) tenen perfectament diferenciats el calze i la corol·la que són, respectivament, dialisèpal i dialipètala; la corol·la dels rosers silvestres, com el de la fotografia central, té només cinc pètals, però les formes cultivades en tenen molts més, en general com a resultat de la metamorfosi d’altres peces florals.
Jordi Vidal
Les peces estèrils de la flor, situades a la seva perifèria, s’expliquen com una adaptació funcional al reclam i a la recepció d’animals transportadors de pol·len. Per aquesta raó són pràcticament inexistents a les gimnospermes, que han encarregat el transport de llur pol·len al vent, i en canvi són freqüents a les angiospermes.
El periant consta típicament de dos tipus de peces que són, de fora cap a dins, els sèpals i eh pètals, que constitueixen, respectivament, el calze i la corol·la. Habitualment, el calze té el color i la consistència de les fulles, mentre que la corol·la, més tendra, és de colors vius; recordem, per exemple, les estepes (Cistus), els ametllers (Prunus dulcis) i tantes altres plantes. També podem trobar flors en què el calze i la corol·la no es diferenciïn morfològicament, cosa freqüent en moltes angiospermes primitives i entre les lílides (cas del cebollí, Asphodelus fistulosus), o bé que tinguin un sol verticil periàntic, com en el cànem (Cannabis sativa). El periant sol rebre en aquests casos el nom de perigoni i les peces corresponents, el de tèpals. Cal afegir, encara, que certes angiospermes, entre les quals podem esmentar els pollancres (Populus), no tenen penant perquè no el necessiten, car els animals no s’encarreguen del transport del pol·len. Esmentem, finalment, que les flors de certes plantes, com ara les malvàcies o els clavells, són embolcallades per un conjunt de peces estèrils, generalment verdes i situades per fora del calze, que rep el nom de calicle.
Segons els elements que integren el periant, hom parla de flors heteroclamídies, si tenen calze i corol·la ben diferenciats; homoclamídies, si, tot i que hi ha dos verticils, llurs peces són tan semblants que no es poden distingir morfològicament; haploclamídies o monoclamídies, quan tenen un sol tipus d’embolcall, i, finalment, aclamídies, si no hi ha periant.

Les flors acícliques de les nimfees (Nymphaea) tenen el periant format de quatre sèpals caducs i de molts pètals lliures; l’androceu consta de nombrosos estams, els externs amb el filament gradualment petaloide; el gineceu, parcialment enfonsat al receptacle, és pluricarpel·lar.
Jordi Vidal
L’especialització de les peces estèrils de la flor ha tingut lloc segurament per camins diferents. Tant el calze com el calicle semblen provenir evolutivament de fulles que s’han atançat a les peces fèrtils. Entre les proves que ho demostren hi ha la mateixa anatomia dels sèpals, perfectament comparable a la de les fulles normals. La corol·la, en canvi, prové molt sovint d’estams estèrils, incapaços de produir pol·len, que han modificat la seva forma. Les relacions entre corol·la i androceu són evidents a les flors de la nimfea (Nymphaea alba), en la qual es poden observar tots els estadis que van des dels estams encara funcionals fins als pètals més exemplars.
Les peces del periant poden ser lliures o bé soldar-se pels marges, i en aquest darrer cas donar lloc a tubs més o menys llargs. Si els sèpals són independents es diu que el calze és dialisèpal (o també corisèpal); si ho són els pètals, la corol·la és dialipètala, i, en el cas del perigoni, parlem de dialitèpal. Si les peces s’han soldat, els prefixos que s’utilitzen són els de gamo-, de manera general, o bé sin-, per a sèpals i tèpals, i sim-, per als pètals. Amb alguns exemples ho entendrem millor: la corol·la d’un codonyer (Cydonia oblonga) és dialipètala, la de la didalera (Digitalis purpurea), simpètala o gamopètala, el calze dels colitxos (Silene vulgaris) és sinsèpal i el perigoni del marcòlic (Lilium martagon), dialitèpal. En els pètals lliures hom distingeix una zona basal, estreta, anomenada ungla, i una zona superior, més ampla, que rep el nom de limbe. Les corol·les gamopètales solen constar d’un tub, que correspon a la part soldada, i dels lòbuls que representen el fragment lliure dels pètals.

El color de la corol·la d’algunes angiospermes pot canviar amb el temps. La del millgruà (Buglossoides purpurocaerulea), com la d’altres plantes (sobretot de la seva mateixa família, les boraginàcies), té inicialment coloracions vermelloses que passen més tard a blavenques. Aquests canvis es deuen a la virada del color dels antocíans vacuolars, sovint com a resposta a canvis del pH cel·lular.
Ramon M. Masalles
A causa de la seva coloració, els pètals són les peces més vistents de la flor; això és tan cert, que parlem del color de la flor referint-nos normalment al color dels pètals. Aquesta coloració té el seu origen en la presència de pigments dissolts en els vacúols (flavones i antocíans, per exemple) o bé situats en els cromoplasts (com alguns carotenoides). De consistència més feble que no pas el calze, la corol·la té una vida generalment molt curta que comença a extingir-se en el mateix moment de la fecundació.
L’androceu
Els estams són fulles modificades portadores d’esporangis, ací anomenats sacs pol·línics, a l’interior dels quals es produeixen les espores o grans de pol·len. El conjunt dels estams d’una flor rep el nom d’androceu.
Els sacs pol·línics són esporangis del tipus eusporangi. Consten d’un teixit fèrtil, intern, anomenat arquespori, envoltat d’una paret formada per quatre o més capes de cèl·lules. Entre l’arquespori i la paret hi ha les cèl·lules del tapet, que nodreixen els grans de pol·len i col·laboren activament en l’engruiximent de la seva coberta.
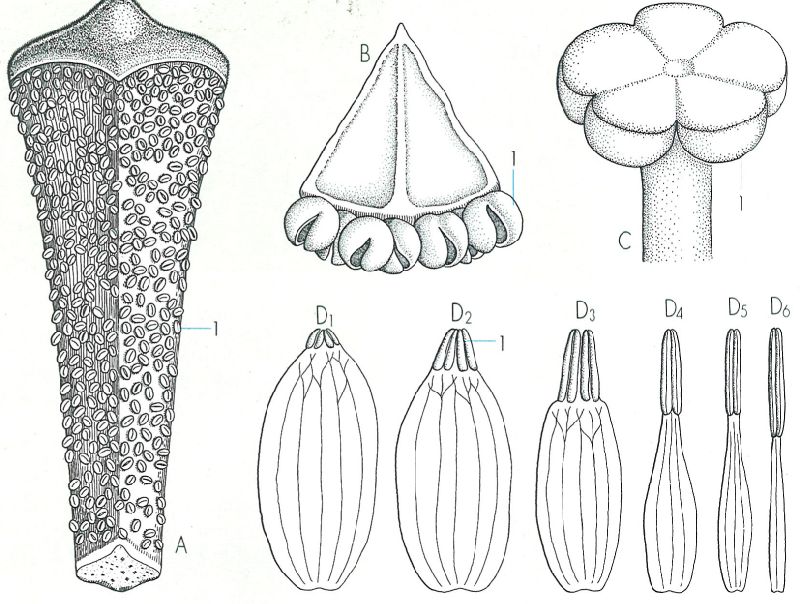
Diversitat morfològica dels estams de diverses plantes superiors: Cycas (A, x 3, 5). Juniperus (B, x 21). Taxus (C, x 45) i Nymphaea (D, x 1,5). Les flors de les nimfees mostren el pas d’estams a pètals (procés que, d’altra banda, explica les flors dobles de moltes de les nostres plantes de jardí). 1 sac pol·línic.
Mercè Cartañá, a partir de fonts diverses
Els estams de les gimnospermes presenten una morfologia molt diversa: són esquamosos a les coníferes, en forma de fulla allargada a Cycas, de llumí a Ephedra, peltats a Taxus, etc. També és variable el nombre de sacs pol·línics per estam, com també el d’estams per flor; en tots els casos, però, els estams formen part de flors unisexuals que tenen sovint forma de pinyes en miniatura. Els estams de les angiospermes, en canvi, tenen tots aproximadament la mateixa forma. Consten d’un peu llarg i prim anomenat filament que porta al capdamunt quatre sacs pol·línics agrupats de dos en dos. Cada grup de dos sacs forma una teca; i les dues teques de l’estam, separades per una mena de solc però soldades per una continuació del filament que rep el nom de connectiu, constitueixen l’antera.
Les flors acícliques, com les de Magnolia, tenen molts estams disposats helicoïdalment entorn d’un eix floral més o menys llarg. Per contra, els estams de les flors cícliques solen ser poc nombrosos i es disposen en un sol verticil o bé en dos. En el cas que hi hagi un sol verticil, en general hi ha tants estams com sepals i com pètals; així ocorre a les umbel·líferes i a les solanàcies, per exemple. Si hi ha dos verticils, com passa a les ericàcies, rutàcies, etc., el nombre d’estams duplica sovint el de pètals. Poden produir-se, tanmateix, variacions notables, que van des de la reducció del nombre d’estams fins a la seva multiplicació. No és pas rar l’avortament d’un o més estams, que es transformen en estaminodis estèrils, com a les escrofulariàcies. També és corrent la multiplicació del nombre d’estams, anomenada poliàndria, que hom pot observar a les rosàcies, a les gutíferes, etc. La diversitat de l’androceu pel que fa al nombre, la disposició, el grau de soldadura, etc. de les peces, serví de base a Linné per a establir el seu sistema sexual de classificació, una mena de clau de determinació, segurament la primera que han tingut els botànics a l’abast.
Els estams de les flors cícliques es disposen, com la resta dels elements florals, alternant amb les peces dels verticils contigus. En cas de desaparició d’un dels verticils, però, es pot arribar a una «superposició» de les peces de dos verticils successius. En realitat, aquests verticils són successius físicament, però no pas evolutivament, ja que entremig en manca un altre (que de vegades es posa de manifest mitjançant estaminodis). Aquesta situació es presenta, per exemple, a les primulàcies, algunes de les quals (Soldanella alpina o Samolus valerandi) conserven encara un verticil d’estaminodis que alternen amb els pètals.
Tot i el fet de néixer independents en els primordis florals, els estams poden soldar-se entre ells o bé amb altres peces en el decurs del seu desenvolupament, moltes vegades com a derivació de certs processos de poliàndria. A les papilionàcies, per exemple, es produeix la concrescència dels filaments estaminals, ja sigui en un sol grup (androceu monadelf) o bé en dos grups (diadelf). L’androceu del pericó (Hypericum perforatum), format per diversos grups d’estams, rep el nom de poliadelf. A les compostes també es fusionen els estams, però no pas pels filaments, sinó per les anteres. Altres vegades es produeix la soldadura de l’androceu amb les peces del periant, sobretot en el cas de les corol·les simpètales, com ocorre a les labiades, a les boraginàcies, etc. A les orquidàcies, hom pot observar, encara, la soldadura de l’androceu, format generalment per un sol estam, amb el gineceu.
L’alliberament dels grans de pol·len es produeix en esquinçar-se la paret de l’antera madura, en general com a conseqüència del dessecament i la contracció de les seves cobertes. Sovint s’estripa longitudinalment el tabic que comparteixen els dos sacs pol·línics de cada teca. Altres vegades l’obertura és més localitzada i consisteix en un porus més o menys gros, com a les ericàcies, o fins i tot en finestretes que conserven, com en les lauràcies, els corresponents finestrons. Habitualment els grans de pol·len són alliberats independents els uns dels altres, però en determinades plantes surten de les anteres agrupats. A les ericàcies, per exemple, els grans de pol·len es dispersen en grups de quatre (tètrades) i a les mimosàcies, en grups de vuit o de pocs més (políades). Les orquidàcies i les asclepiadàcies dispersen conjuntament tot el pol·len d’una teca, cimentat formant un pol·lini.
L’arquespori dels sacs pol·línics és un teixit que es divideix mitòticament fins a produir les cèl·lules mare del pol·len, cadascuna de les quals dona lloc a quatre cèl·lules pol·líniques o grans de pol·len mitjançant un procés de meiosi. Així doncs, els grans de pol·len són espores haploides.
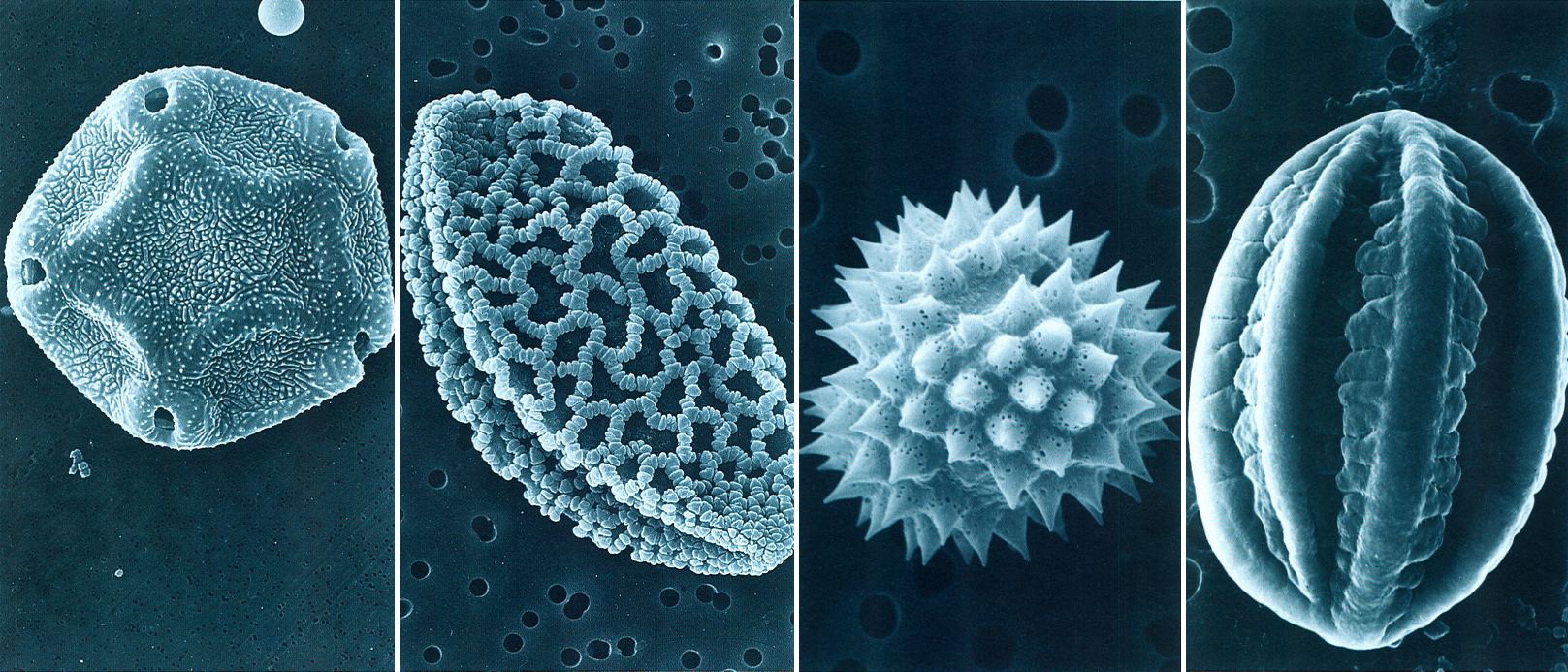
Morfologia de diversos grans de pol·len al microscopi electrònic d’escandallatge. Els de l’èfedra (Ephedra major), una gimnosperma, no tenen apertura i són el·lipsoïdals (fotografia superior a l’esquerra, en visió meridiana) i mostren de 5 a 25 solcs longitudinals que no són pròpiament apertures veritables: en el moment de la germinació, el gra es trenca per un dels solcs i permet la sortida del tub pol·línic (x 2 000). Els de l’àrnica (Arnica montana), una composta, són aproximadament esferoïdals (fotografia superior a la dreta, en visió polar) i equinats (és a dir, l’exina té nombroses espines perforades, curtes i amples com a adaptació a la pol·linització entomòfila), amb tres apertures compostes o colporus (x 2 400). A baix, a l’esquerra, hi ha un gra de pol·len aproximadament esferoidal (oblat) de vern (Alnus glutinosa) en visió polar; de contorn pentagonal i especialment adaptat a la pol·linització anemòfila, presenta quatre o cinc porus arrodonits i envoltats d’un anell prominent (és, doncs, aspidoporat) i l’exina espinuloso-verrucosa (x 2 400). El gra de forma navicular de la fotografia inferior a la dreta correspon al marcólic (Lilium martagon), una liliàcia; és monocolpat (el colp coincideix amb el solc llarg que recorre tot el pol distal) i d’exina reticulada, amb lúmens amples a la part central i compacto-verrucosa als extrems equatorials (x 1 200).
María Suárez
Com que el pol·len ha de suportar durant un cert temps les inclemències atmosfèriques, convé que sigui resistent. Els grans de pol·len tenen, per aquesta raó, una coberta gruixuda anomenada esporodermis, en la constitució de la qual intervenen dos estrats ben diferenciats, l’exina i la intina. Vexina, l’estrat exterior, és particularment resistent a la descomposició a causa de la impregnació amb esporopol·lenina, un polímer complex d’elevat pes molecular que també es troba a les espores de molts fongs. És formada per diverses capes, les més exteriors de les quals presenten ornamentacions especials relacionades amb els mecanismes de pol·linització. El seu gruix, variable, és particularment minso en determinades zones anomenades punts germinatius, sovint més o menys circulars (porus) o allargats (colps). A través d’aquestes zones poc resistents sortirà el tub pol·línic en el moment de la germinació del gra de pol·len. La intina consta també de diverses capes, però és molt més delicada i, sobretot, elàstica, ja que envolta el tub pol·línic durant tot el seu creixement.
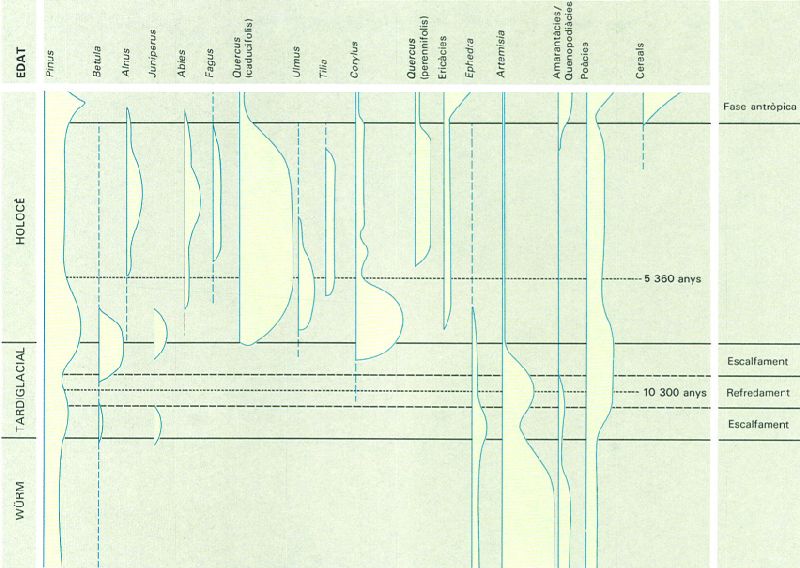
Diagrama pol·línic de síntesi corresponent al Pleistocè superior i a l’Holocè, elaborat a partir de diversos sondatges efectuats a la plana d’Olot (Garrotxa). El predomini del pol·len d’Artemisia i, secundàriament de poàcies i d’amarantàcies o quenopodiàcies, fa pensar en l’existència durant el Würm d’un paisatge vegetal de caràcter estèpic que, amb lleugeres oscil·lacions, es manté fins a l’acabament del Tardiglacial, progressivament substituït per bedollars (de vegades amb pins). Des de l’inici de l’Holocè, les rouredes de roure pènol esdevenen dominants i més tard apareixen avetoses i fagedes als obacs, vernedes a la vora dels rius i encara alzinars als solells. La presència de l’home condiciona el paisatge vegetal dels darrers dos mil anys, caracteritzat per la desaparició de molts boscos i l’increment dels conreus i de la vegetació ruderal.
Carto-Tec, a partir d’originals de Ramon Pérez-Obiol, 1987
La ciència que estudia els grans de pol·len i, de manera més general, les espores rep el nom de palinologia. És una disciplina antiga que ha pres, però, molta volada durant els darrers decennis, d’una banda perquè les tècniques actuals permeten la realització de treballs de gran precisió i, de l’altra, pel fet que la morfologia de cada gra de pol·len (forma, dimensions, ornamentació, etc.) permet de reconèixer la planta de la qual prové. En relació amb això, són nombroses les aplicacions de la palinologia. Es pot saber, per exemple, quines espècies han estat visitades per les abelles en el procés d’elaboració de la mel simplement estudiant el pol·len que conté. I, atesa la resistència de l’exina, que fa possible la conservació del pol·len a l’interior de les roques sedimentàries, dipòsits lacustres, etc., es pot arribar a esbrinar la història de la flora i de la vegetació d’un indret determinat, si més no pel que fa als períodes geològics més recents. Mitjançant l’anàlisi pol·línica quantitativa i qualitativa dels materials extrets per sondatge, es pot deduir la composició aproximada de les comunitats vegetals situades a la zona mentre va durar la formació del jaciment. Encara podem afegir que el pol·len de certes plantes, sobretot de les gramínies però també de les urticàcies, de certes compostes, etc., provoca malalties de tipus al·lèrgic conegudes des de fa poc més d’un segle amb el nom genèric de pol·linosis, el tractament i la prevenció de les quals no serien possibles sense les aportacions de la palinologia.
El gineceu
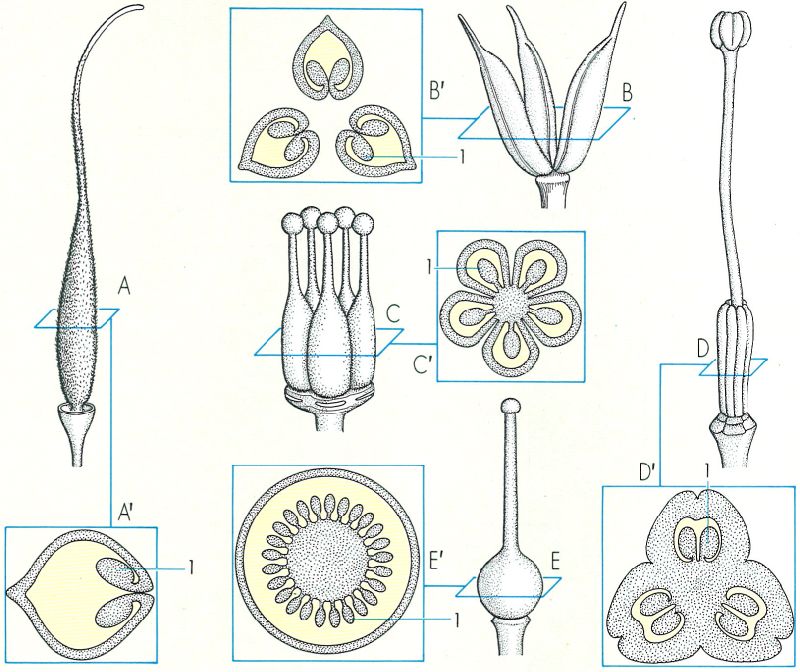
Gineceus de diverses angiospermes, amb les seccions transversals dels ovaris corresponents. El pistil de les papilionàcies (A) és monocarpel·lar; la secció de l’ovari (A’) mostra els primordis seminals disposats als marges de la fulla carpel·lar. En el cas dels esperons (Delphinium), el gineceu consta de diversos pistils monocarpel·lars (B); el tall transversal (B’) mostra una placentació marginal, com en el cas precedent. Els cinc carpels d’Oxalis (C) s’han soldat en un ovari únic amb cinc cavitats o lòculs (C), però conserven lliures els cinc estils amb els estigmes corresponents. A Lilium (D) hi ha un pistil tricarpel·lar amb un ovari trilocular (D’) i un estil, però amb tres estigmes. Els cinc carpels que formen el gineceu de Primula (E) són soldats formant un ovari, un estil i un estigma únics; E’ secció transversal de l’ovari, unilocular i amb placentació central. 1 primordi seminal.
Mercè Cartañá, a partir de fonts diverses
La part femenina de la flor consta d’un nombre variable de carpels portadors de primordis seminals. Els carpels són fulles modificades que, ben sovint, conserven encara l’aspecte foliar. Els primordis seminals estan formats d’un esporangi i una coberta protectora que hom suposa derivada, en el decurs de l’evolució, d’antics esporangis avortats. El conjunt dels carpels, amb els primordis seminals, rep el nom de gineceu.
Els carpels de les gimnospermes són poc homogenis. A la majoria de les espècies conegudes consisteixen, simplement, en un suport per als primordis seminals. Només en uns pocs grups (cas de Cycas) trobem carpels morfològicament comparables a fulles, portadors d’un nombre generalment reduït de primordis seminals. En tots els casos, tanmateix, són carpels oberts que deixen els primordis seminals directament accessibles als grans de pol·len.
Els carpels de les angiospermes, malgrat que estan sempre tancats i soldats pels marges, deixen reconèixer encara el seu origen foliar. Els recipients que constitueixen, típicament en forma d’ampolla, reben el nom de pistils (del llatí «pistillum», mà de morter). El pistil consta d’una part basal més o menys inflada, l’ovari, dins del qual es disposen els primordis seminals; d’una zona apical o estigma, preparat per a la recepció i la germinació del pol·len; i d’una columna, anomenada estil, que uneix aquestes dues parts. El pistil es justifica evolutivament per la necessitat de protegir els primordis seminals, ja sigui de la dessecació o bé de les mandíbules dels animals encarregats de transportar el pol·len. En contrapartida, els grans de pol·len ja no tenen accés directe als primordis seminals, sinó que troben en el seu camí un filtre especialitzat, l’estigma.

El gineceu de l’hibisc de la Xina (Hibiscus rosa-sinensis) consta de cinc carpels fusionats formant un ovari súper i un estil que es ramifica a la meitat superior i porta cinc estigmes amples i capitats. La fotografia permet observar, encara, el tub que formen els filaments soldats dels nombrosos estams (androceu monodelf) i la presència d’un cal·licle o epicalze.
Jordi Vidal
Les flors de les angiospermes primitives solen presentar molts carpels disposats helicoïdalment sobre l’eix floral. Cadascun d’aquests carpels forma habitualment un pistil independent, de manera que el nombre de pistils coincideix amb el de carpels; és el cas dels gineceus anomenats coricàrpics (o, també apocàrpics) dels botons d’or (Ranunculus), de les magnòlies (Magnolia grandiflora), etc. Més sovint, però, els carpels, com totes les peces florals, tenen disposició verticil·lada i es troben en nombre poc elevat. En aquests casos és molt freqüent la concrescència entre ells, que pot afectar només la regió de l’ovari (el resultat és un pistil amb un sol ovari però amb estils nombrosos) o bé incloure també els estils i, de vegades, fins i tot els estigmes. Els pistils formats de diversos carpels soldats corresponen als gineceus anomenats sincàrpics (o, també, cenocàrpics) que trobem en moltíssimes famílies de la nostra flora: crucíferes, escrofulariàcies, etc.
La posició de l’ovari en relació amb les altres peces florals és un caràcter important. Parlem d’ovari súper quan el gineceu se situa sobre un receptacle més o menys pla, de tal manera que la resta de les peces florals semblen arrencar de sota l’ovari. L’ovari és ínfer quan es troba soldat a un receptacle molt còncau en forma de copa; en aquest cas, el periant i l’androceu semblen sortir per damunt l’ovari. Si la soldadura afecta només la part inferior de l’ovari, es diu que aquest és semiínfer. Els receptacles en forma de copa, habitualment soldats a l’ovari (però no sempre, com en els rosers), reben el nom d’hipant.
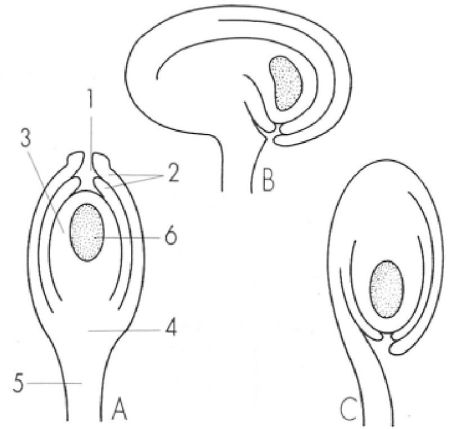
Els primordis seminals, de vegades anomenats òvuls, són òrgans propis de les plantes superiors que contenen els megasporangis i posteriorment les megàspores i el megaprotal·lus. Els del dibuix consten de: 1 micròpil, 2 teguments, 3 nucel·la, 4 càlaza, 5 funicle, 6 megaprotal·lus. Segons l’orientació del micròpil, es parla de primordis àtrops o ortòtrops (A), anàtrops (C) i campilòtrops (B).
Mercè Cartañá, a partir de fonts diverses
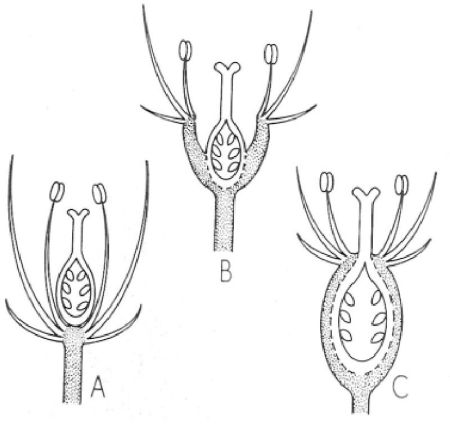
La situació de l’ovari respecte del receptacle floral es pot referir a tres tipus bàsics: ovari súper (A), semiínfer (B) i ínfer (C). A la figura, el receptacle i el peduncle florals són representats en puntejat.
Mercè Cartañá, a partir de fonts diverses
Cada un dels primordis seminals consta d’un nucli central, la nucel·la, envoltat d’una o dues cobertes que s’anomenen teguments. Aquests s’originen en una zona basal que rep el nom de càlaza i deixen a l’extrem oposat una petita obertura, el micròpil. De la càlaza surt, també, el funicle, el mànec que uneix el primordi amb el carpel. Segons llur posició respecte del funicle, parlem de primordis àtrops (rectes, amb el micròpil oposat a la càlaza i al funicle), anàtrops (girats, amb el micròpil al costat del funicle) o campilòtrops (disposats transversalment al funicle). La nucel·la és un esporangi amb un teixit fèrtil central anomenat arquespori. Algunes de les cèl·lules de l’arquespori actuen com a cèl·lules mare del sac embrional, una de les quals es dividirà meiòticament i originarà quatre espores haploides, les cèl·lules del sac embrional. Normalment, tres de les meiòspores així originades s’atrofien, de manera que només una esdevé adulta. Contràriament a les espores originades en els sacs pol·línics (és a dir, els grans de pol·len), que són disseminades tan bon punt han madurat, la cèl·lula del sac embrional resta dins de l’esporangi on ha estat formada.
Els primordis seminals poden aparèixer en llocs diversos de l’ovari, però s’insereixen sempre en unes zones carpel·lars concretes que reben el nom de placentes. Als pistils monocarpel·lars, les placentes se situen sovint als marges del carpel, però també ho poden fer a la zona medial. En alguns pistils sincàrpics, la concrescència dels carpels ha comportat reordenacions internes que han desplaçat secundàriament els primordis cap al centre de l’ovari, cap a la seva base, etc. Per aquestes raons la disposició dels primordis seminals dins l’ovari, anomenada placentació, presenta una certa diversitat que podem referir, això no obstant, a dos tipus bàsics: en primer lloc, el tipus parietal, en el qual els primordis s’insereixen a les parets de l’ovari, com passa a la família de les violàcies; i, segonament, el tipus central, en el qual els primordis seminals neixen de l’eix central del pistil. Trobem bons exemples de placentació central a les flors de les primulàcies i de les cariofil·làcies.
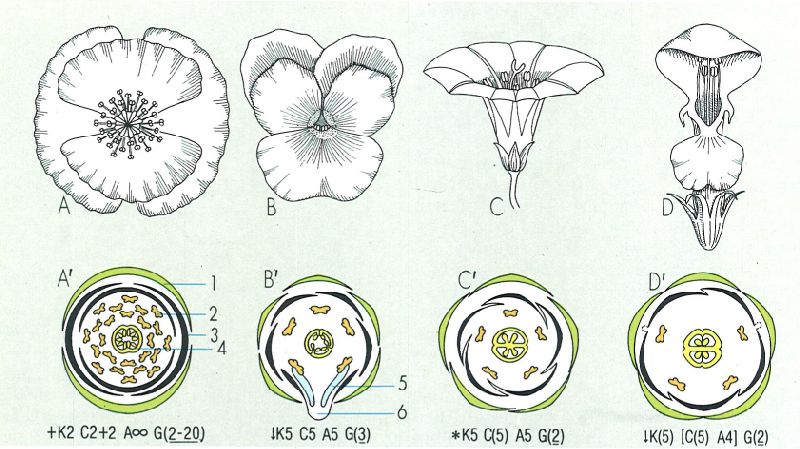
Flors, diagrames florals i fórmules florals de plantes corresponents a quatre gèneres de la nostra flora: A i A’ rosella (Papaver), B i B’ pensament (Viola), C i C corretjola (Convolvulus), D i D’ Lamium. 1 calze, 2 corol·la, 3 androceu, 4 gineceu, 5 nectaris, 6 esperó.
Mercè Cartañá, a partir de fonts diverses
Els diagrames i les fórmules florals són expressions convencionals de les característiques fonamentals d’una flor. Els primers són projeccions de les flors en què les peces més externes corresponen als verticils inferiors; equivalen a talls transversals, idealitzats, de la flor i hi són representats el nombre de peces de cada mena, el grau de soldadura de les peces i la disposició relativa dels elements florals. Les fórmules florals són un conjunt de símbols, lletres i números que expliquen, de manera ordenada, la simetria de la flor, el nombre de peces per verticil, la soldadura de les peces (si n’hi ha), la posició de l’ovari, etc. Pel que fa a la simetria, els codis utilitzats són aquests: @, flor acíclica; *, flor actinomorfa; ↓, flor zigomorfa i +, flor dissimètrica. El nombre de peces florals s’indica a continuació dels símbols corresponents al calze (K), la corol·la (C), el perigoni (P), l’androceu (A) i el gineceu (G). Si són dos els verticils que hi intervenen, cal explicitar el nombre de peces per verticil (un androceu format per dos verticils de tres peces cadascun, per exemple, es representa així: A3+3). Les peces soldades s’indiquen posant entre parèntesis el número corresponent, o bé entre claudàtors, si la soldadura afecta dues menes de peces; la combinació [C(5)A4] vol dir que els cinc pètals de la corol·la estan soldats entre ells i amb els quatre estams de l’androceu. El caràcter súper o ínfer de l’ovari es representa mitjançant una línia situada, respectivament, a sobre o a sota de la xifra que indica el nombre de carpels. Així doncs, G(2) significa que el gineceu, format de dos carpels soldats, té l’ovari súper. Un aclariment, encara: si el nombre de peces és molt elevat (més de 15 o 20) no cal especificar-lo exactament, sinó que s’indica amb el símbol que representa el concepte d’infinit ∞ (no cal dir que es tracta d’un infinit que no té gran cosa a veure amb el dels matemàtics); la fórmula floral de la rosella (Papaver rhoeas), per exemple, és: + K2 C2+2 A∞ G(5-18).
Les inflorescències
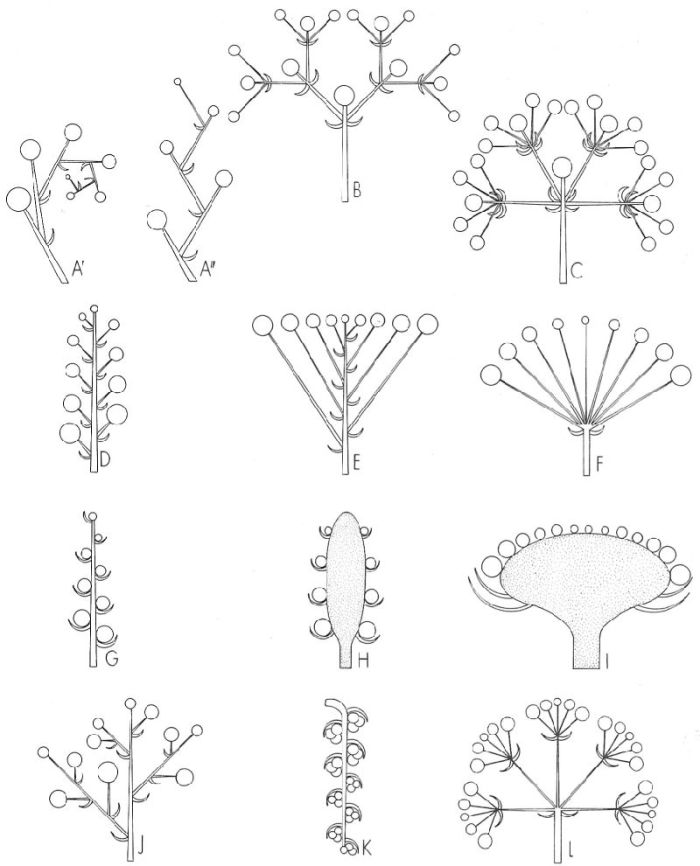
Esquema dels principals tipus d’inflorescències cimoses (A-C), racemoses (D-l) i compostes (J-L). A cima unípara o monocasi (A’ cima escorpioide, A" ripidi), B cima bípara o dicasi, C cima multípara o pleocasi, D raïm, E corimbe, F umbel·la, G espiga, H espàdix, I capítol, J panícula, K ament, L umbel·la composta.
Mercè Cartañá, a partir de fonts diverses
Una inflorescència és un sistema de rames especialitzat, generalment amb fulles simplificades o bé sense fulles, que porta un conjunt de flors. Segons el tipus de ramificació, les inflorescències es poden qualificar de cimoses, si la ramificació és simpòdica, o bé de racemoses, si és monopòdica. Un altre caràcter important és la presència o no de flor terminal a l’àpex de l’eix central; en el primer cas es tracta d’una inflorescència oberta i, en el segon, d’una de tancada. De manera general, les inflorescències cimoses són tancades mentre que les racemoses solen ésser obertes.
Els tipus més freqüents d’inflorescències racemoses són: el raïm, l’espiga, l’espàdix, el corimbe, la umbel·la i el capítol. Les flors del raïm es disposen poc o molt separades les unes de les altres al llarg d’un eix, amb els peduncles florals tots d’una llargada aproximadament equivalent; és la inflorescència típica de moltes de les nostres crucíferes. L’espiga es caracteritza perquè les flors s’insereixen directament (sense peduncle) a l’eix principal, talment com un raïm que tingués les flors sèssils; trobem espigues en els plantatges (Plantago) i també en les gramínies, però en aquest cas les espigues solen formar inflorescències compostes (una espiga d’espigues, per exemple, a l’ordi, Hordeum vulgare). Vespàdix és una mena d’espiga que té l’eix principal engruixit i les flors unisexuals; el de la sarriassa (Arum italicum), amb les flors femenines situades a la part basal de l’eix i les masculines situades més amunt, és envoltat per una gran bràctea en forma de paperina que rep el nom à’espata. En el corimbe, hi ha un conjunt de flors que arrenquen de l’eix principal com en el raïm, però que arriben totes aproximadament al mateix nivell; això comporta que la longitud dels pedicels florals es redueixi progressivament de les flors inferiors a les superiors. Fan corimbes moltes crucíferes, entre les quals els carraspics (Iberis). Parlem à’umbel·les quan les flors provenen de nusos tan atançats que sembla que surtin d’un únic punt i arriben totes al mateix nivell. La majoria de les umbel·líferes fan umbel·les compostes, això és, umbel·les que, en comptes de flors, porten al seu torn altres umbel·les, més petites, anomenades umbèl·lules. A la base de la umbel·la hi pot haver un grup de bràctees, l’involucre i, anàlogament, pot existir un involucel a sota de cada umbèl·lula. El capítol consta d’un receptacle en forma de disc o de con damunt del qual es disposen, densament juntes, les flors; el conjunt d’aquestes flors és envoltat per un collar de bràctees, anomenat involucre. Els capítols són característics de les famílies de les compostes, les globulariàcies, les dipsacàcies, etc.

Els ciatis són inflorescències pròpies de les lletreres o lletereses (Euphorbia), que solen agrupar-se, encara, en inflorescències cimoses del tipus dicasi o pleocasi. Cada ciati és format per diverses flors masculines i una de femenina (un ovari inflat, amb tres estils i sis estigmes), envoltat tot el conjunt per un involucre de cinc bràctees portador de quatre glàndules que poden prendre a moltes espècies l’aspecte d’un «croissant», com en el cas de la fotografia.
Enric Curto
A les inflorescències cimoses l’eix principal atura el seu creixement quan dona lloc a una flor, però a partir d’aquest eix es formen una o més rames laterals, les quals acaben també en una flor i poden ser substituïdes per rames terciàries que es comporten igual, i així successivament. Hi ha tres tipus bàsics de cimes: el monocasi, el dicasi i el pleocasi. En el monocasi o cima unípara, una sola branca reprèn el creixement quan l’eix precedent produeix una flor i atura el seu; hi ha diversos tipus de monocasis segons quina sigui l’orientació de les rames que es van formant. Si totes les rames provenen del mateix costat de l’eix respectiu, el conjunt es cargola com la cua d’un escorpí i, llavors, es tracta d’una cima escorpioide, fàcilment identificable a moltes boraginàcies. En el dicasi o cima bípara, l’eix principal, acabat en una flor, produeix dues rames, cadascuna de les quals pot seguir el mateix procés; presenten dicasi moltes cariofil·làcies, entre les quals hi ha els colitxos (Silene vulgaris). Si són tres, o més de tres, les rames que provenen de l’eix principal en formar, aquest, una flor terminal, parlem de pleocasi o cima multípara; els pleocasis són freqüents a les lletereses (Euphorbia). Les inflorescències cimoses, també anomenades cimes, poden presentar les flors sense peduncles. En aquests casos solen prendre un aspecte si fa no fa esfèric i reben el nom de glomèruls; són molt freqüents a la família de les labiades (l’espígol, la menta, etc.).

Els capítols són inflorescències característiques, entre d’altres, de la família de les compostes. Els del gira-sol (Helianthus annuus) són de grans dimensions i permeten observar les flors ligulades, estèrils, a la perifèria i les tubuloses o flòsculs a la zona central; alguns dels flòsculs mostren una columna central, buida i fosca, formada per les anteres soldades, que alliberen el pol·len, de color groc, a l’interior del tub.
Ramon M. Masalles
Les inflorescències compostes són formades per la reunió de dues o més inflorescències simples, i es caracteritzen perquè porten inflorescències allí on les simples duen flors. Hem parlat ja de la umbel·la i de l’espiga compostes; conjuntament amb la panícula i l’ament, que comentarem seguidament, són les més ben representades a la nostra flora. La panícula és un raïm compost o raïm de raïms. L’eix principal, de creixement indefinit, produeix branques portadores de flors pedicel·lades. Les flors dels ceps (Vitis vinifera) s’agrupen en panícules. D’altres panícules porten espigues en comptes de flors: l’anomenada «espiga» de moltes de les gramínies, com és ara la civada (Avena sativa), la canyota (Sorghum halepense), etc., és en realitat una panícula d’espigues.
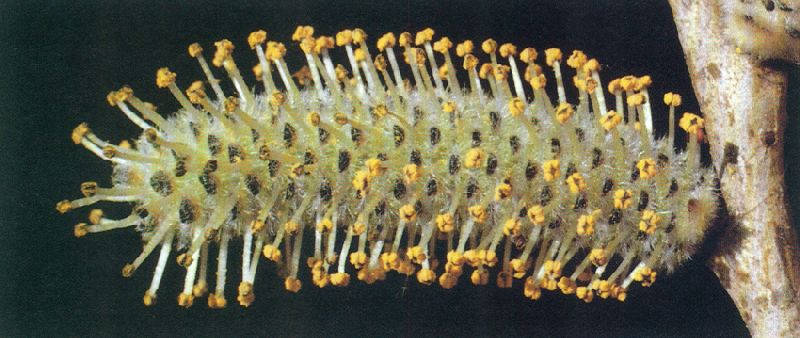
Ament masculí de salze (Salix). Els aments són inflorescències generalment pèndules i unisexuals, en forma d’espiga densa, presents a molts dels arbres planifolis de la nostra flora.
Antoni Agelet
L’ament és un raïm contret, amb aspecte d’espiga i generalment pèndul, que porta glomèruls de flors unisexuals. El nombre de flors per glomèrul és variable. Presenten aments molts dels nostres arbres planifolis: bedoll (Betula pendula), castanyer (Castanea sativa), etc.