Consideracions generals sobre els nematodes
Els nematodes constitueixen un grup de verms de cos cilíndric, no segmentat, i recobert d’una cutícula resistent que els dóna un aspecte molt característic. Essent alguns, especialment els paràsits de l’home, coneguts des de l’antiguitat, el conjunt del grup ha anat prenent cada vegada més importància al llarg de la història de la ciència. Els nematodes han tingut un gran èxit en la colonització de la Terra. Apareguts durant el Paleozoic, han penetrat, a partir de l’ambient marí, en les aigües dolces i el medi terrestre, on un gran nombre d’espècies viuen com a paràsits d’animals o plantes, fins al punt de constituir un dels grups d’animals més abundants de la Terra (hom els situa en segon lloc després dels insectes): actualment es parla d’unes 20 000 espècies, però és probable que n’hi hagi moltes més, si es té en compte que cada espècie d’insecte allotja, com a mínim, una espècie de nematode paràsit, i que n’hi ha molts que parasiten altres grups d’animals, i, encara, molts més de fitoparàsits i de vida lliure en el medi aquàtic i terrestre. D’altra banda, a més del gran nombre d’espècies, destaca també llur gran abundància: hom n’ha comptat fins a 90 000 exemplars en una poma podrida, aproximadament un miler (pertanyents a 36 espècies diferents) en una mostra de 6 o 7 ml de llot, etc. Atesa la gran diversitat de formes de vida, hom els ha estudiat clàssicament en branques diferents, ja sigui des del punt de vista de la parasitologia o helmintologia, ja sigui des de la fitoparasitologia i la zoologia aquàtica i terrestre (nematologia).
Morfologia

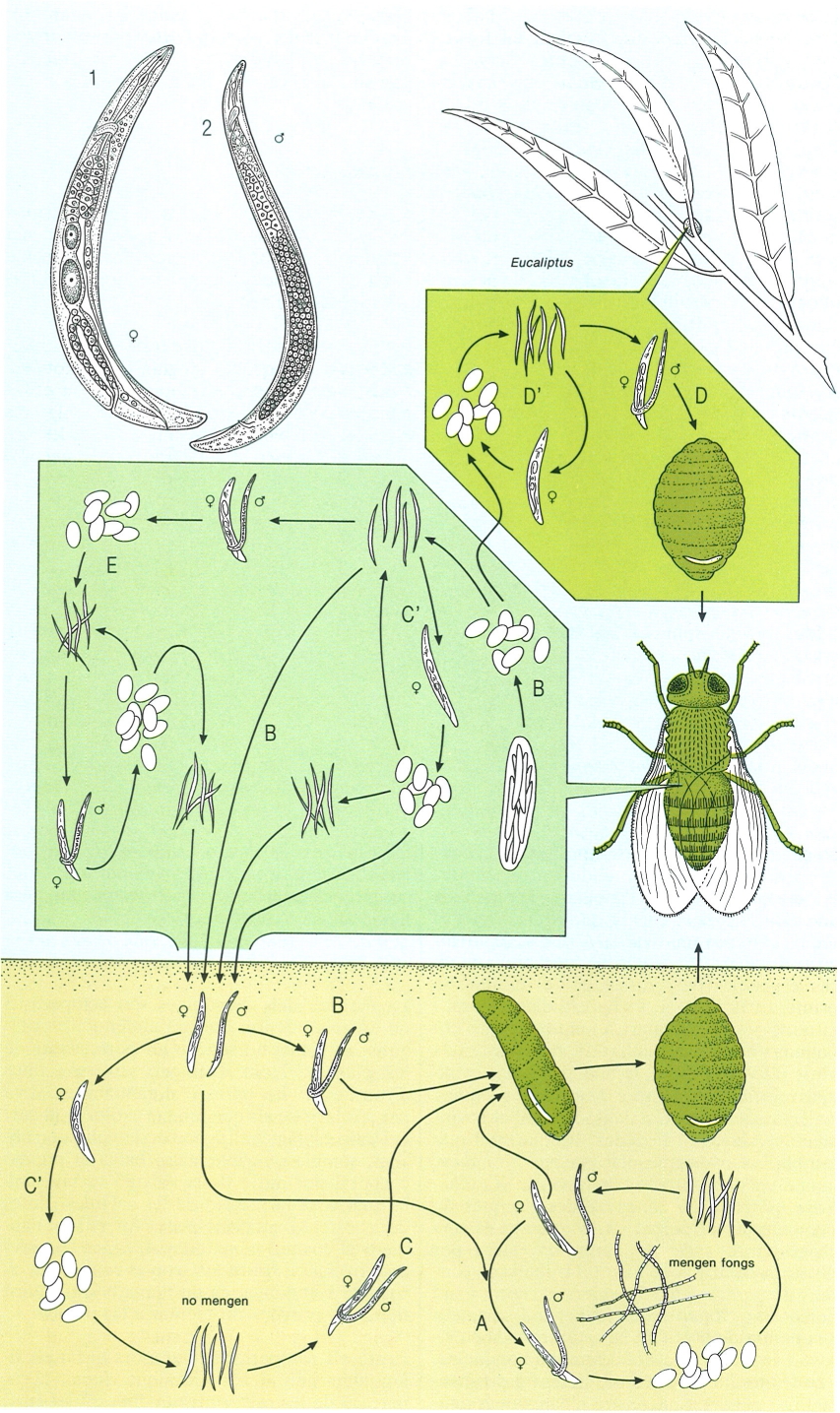
La gran diversitat morfològica dels nematodes és en relació amb la gran diversificació ecològica d’aquest extens fílum animal. Efectivament, els nematodes ocupen nínxols ecològics molt diversos i són d’una gran importància quantitativa i qualitativa en pràcticament tots els ecosistemes: fan poblacions de nombrosos individus, i reuneixen un gran nombre d’espècies, de vida lliure, paràsites d’animals (l’home inclòs), paràsites de vegetals, paràsites d’altres paràsits, etc. La figura vol mostrar alguns exemples d’aquesta riquesa morfològica. Hom hi ha representat: 1 Halalaimus (mar Antàrtica), 2 Dracograllus (oceà Atlàntic), 3 Echinotherisus (mar del Nord), 4 Greffiella, 5 Pselionema (mar del Nord), 6 Desmoscolex, 7 Richtersia (oceà Atlàntic), 8 Trichurls (noteu l’espícula i la beina evaginada del mascle), 9 Criconema, 10 Tetrameres (femella), 11 Necator americanus (mascle), 12 Soboliphyme (femella), 13 Bunonema, 14 Chaetosoma, 15 Kahmannia eliomydis (paràsit de la rata cellarda, Balears).
Biopunt, a partir de fonts diverses
La morfologia general dels nematodes és molt homogènia, malgrat la gran diversitat d’hàbitats i nínxols ecològics que ocupen. La llargada del cos és altament variable, des de menys d’un mil·límetre ( Hapalomus [Greeffiella] minutus mesura 82 µm), fins a uns quants metres (8 m en el cas de Placentonema gigantissima, paràsit de catxalots). Entre els paràsits de l’home hi ha igualment una gran varietat de mides: Strongyloides stercoralis no arriba als 2 mm de llargada, mentre que Dracunculus medinensis pot arribar a fer 1 m. En general són transparents, blanquinosos o groguencs i si presenten algun altre color, aquest correspon al del contingut intestinal. La forma del cos, d’una simetria bilateral primària, també és molt variable (filiforme, fusiforme, globosa, esfèrica, etc.), però en general és cilíndrica i allargada, amb els extrems que s’aprimen gradualment. Els extrems anterior o cefàlic i posterior o caudal solen tenir formacions particulars.

Detall de la regió cefàlica d’una femella del nematode Choanolaimus psammophilus, on s’aprecien esplèndidament els amfidis, en forma de petites fossetes laterals al cap. La fotografia és feta al microscopi òptic. Aquesta espècie, depredadora, ha estat trobada en un salicornar del delta de l’Ebre.
Alejandro Palomo.
Hom pot considerar el cos dividit en quatre regions diferents: la regió cefàlica, l’esofàgica, la intestinal i la caudal. La regió cefàlica presenta, a l’extrem anterior, l’obertura oral, amb llavis proveïts de papil·les i setes tàctils, i, a cada costat del cap, òrgans quimioceptors (amfidis); les estructures fotoceptores són taques pigmentàries, i són especialment abundants en les formes aquàtiques. La regió esofágica presenta externament el porus excretor (ventral), i internament correspon a l’estomodeu. La regió intestinal, la més llarga, correspon interiorment a l’intestí i els òrgans genitals. La cua és la regió posterior i sol ésser una mica o molt corbada en els mascles.
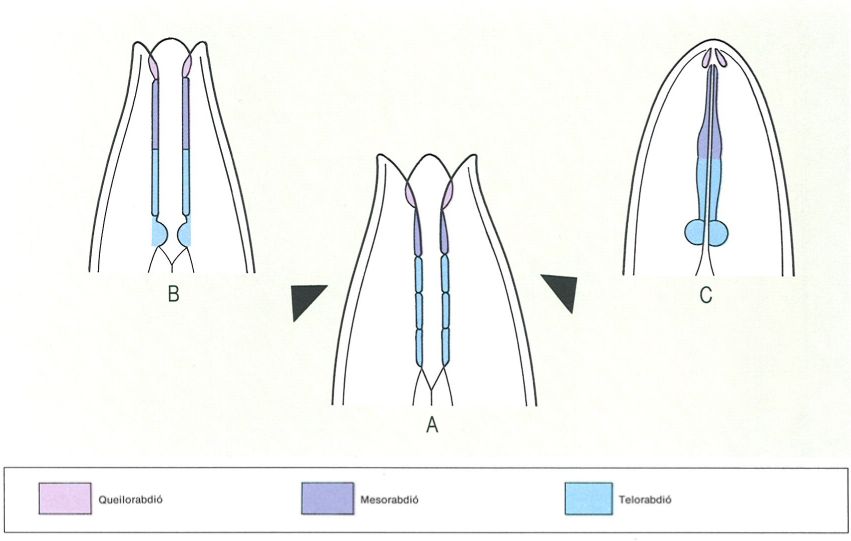
Origen i formació de l’estoma o cavitat bucal dels nematodes micròtrofs rabditoïdeus (B) i de l’estilet dels nematodes paràsits de plantes i d’invertebrats (C), a partir d’una forma edàfica primitiva (A).
Biopunt, a partir de fonts diverses
La paret del cos és constituïda per la cutícula, l’hipoderma i la musculatura de la paret. La cutícula és formada per tres capes (cortical, mitjana o mesocutícula, i interna o endocutícula), fetes al seu torn de diverses capes, en la composició de les quals hi ha lípids, una proteïna relacionada amb la ceratina, i fibres de colagen. La cutícula gairebé mai no és llisa, sinó que acostuma a presentar una estriació transversal, a més d’ornamentacions i formacions diverses (reforços anul·lars, berrugues, espines, arestes, ales cefàliques, ales caudals, bossa copuladora, papil·les cefàliques i papil·les caudals). L’hipoderma és un síncit prim situat immediatament per sota de la capa basal de la cutícula, la funció del qual és secretar cutícula nova. Presenta quatre engruiximents o cordons hipodèrmics, que recorren longitudinalment el cos formant uns sortints interns (dorsal, ventral i dos laterals) en la cavitat general, i que delimiten quatre zones o camps hipodèrmics longitudinals. La musculatura de la paret és constituïda per fibres longitudinals de cèl·lules que se situen entre els cordons hipodèrmics. Poden adoptar diferents posicions. Hom parla de musculatura de tipus polimiari —típica dels nematodes grossos, com Ascaris— quan té les fibres musculars molt nombroses i molt sortints; la del tipus meromiari—típica de nematodes petits (oxiúrides) i d’estadis larvals— presenta només dues o tres fibres de cèl·lules aplanades; el tipus holomiari comprèn fibres musculars abundants, iguals i disposades molt juntes, de manera que només deixen lliure el cordó ventral (triquiúrides). A més de la musculatura de la paret del cos, hi ha molts altres músculs, més especialitzats, com ara els esofàgics, els intestinals o els associats a la reproducció.
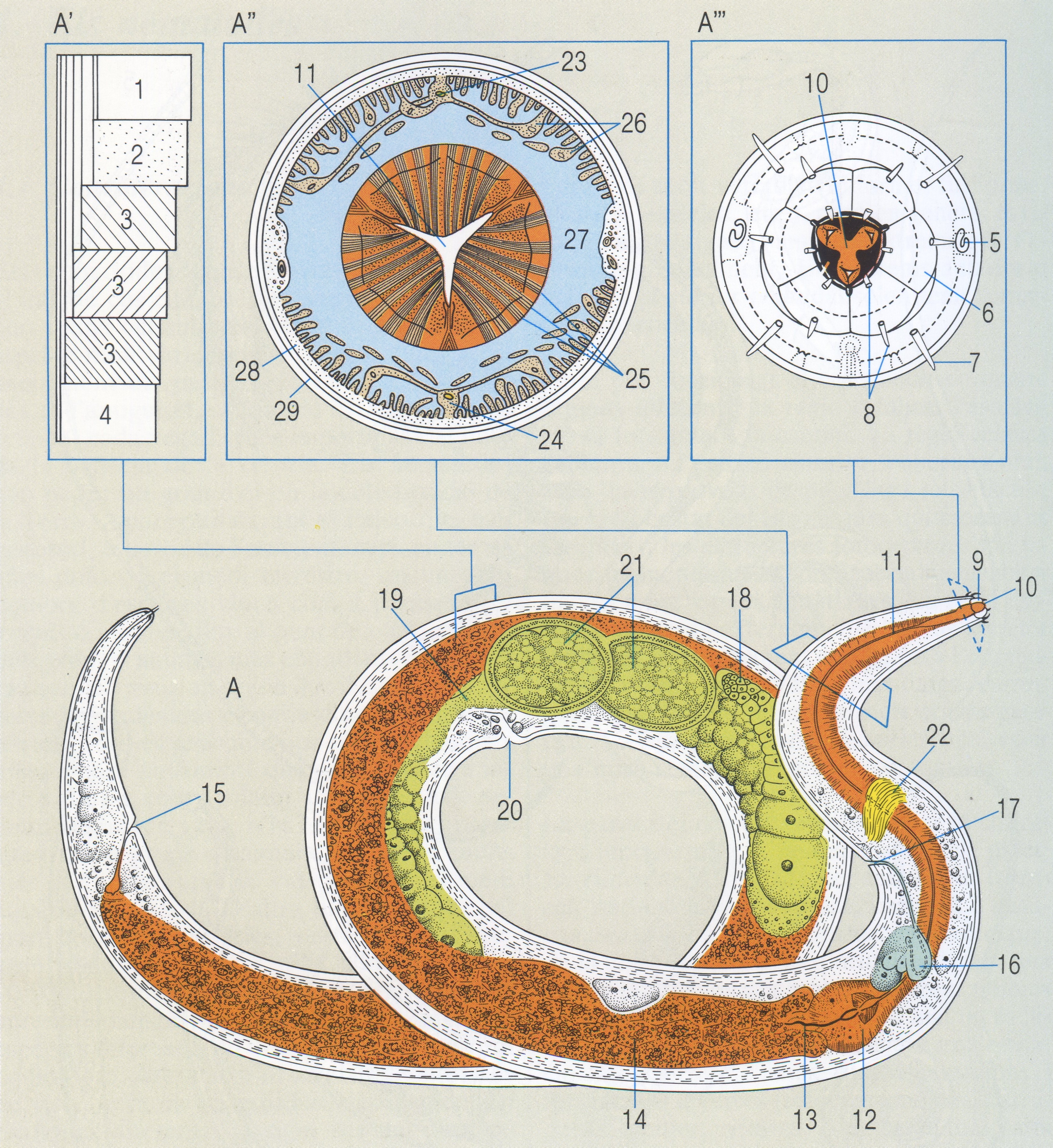
Morfologia general interna dels nematodes: A tall longitudinal d’un nematode que es pot considerar típic (concretament, una femella adulta d’adenofori. de l’espècie Plectus parietinus); A′ detail de les capes cuticulars. A″ tall transversal del cos a l’altura de l’esòfag, i A″′ visió frontal de la regió cefàlica. Hom n’ha indicat: 1 còrtex, 2 capa homogènia, 3 capa fibril·lar, 4 membrana basal, 5 amfidis, 6 llavi, 7 seda cefàlica, 8 papil·la cefàlica, 9 càpsula bucal, 10 boca, 11 esòfag, 12 bulb cardíac, 13 vàlvula esofàgico-intestinal, 14 intestí 15 anus, 16 cèl·lula excretora, 17 porus excretor, 18 ovari, 19 ùter, 20 vulva, 21 ous, 22 anell nerviós, 23 cordó nerviós dorsal, 24 cordó nerviós ventral, 25 musculatura radial, 26 musculatura longitudinal, 27 cavitat del cos (pseudoceloma), 28 epidermis, 29 cuticula.
Biopunt, a partir de fonts diverses
La cavitat del cos, el pseudoceloma, conté un líquid pseudocelomàtic que envolta els órgans. Aquests són fixats a la paret del cos per fines trabècules de teixit conjuntiu. El pseudoceloma té com a funció fer d’esquelet hidrostàtic i intervé en l’intercanvi de substàncies entre teixits i en la difusió dels gasos. Els nematodes no disposen d’aparells respiratori ni circulatori, sinó que prenen l’oxigen per difusió a través de la paret del cos.
L’aparell digestiu és un tub senzill, i complet en la majoria de les espècies, amb boca i anus (excepcionalment, en els mermítids i alguns filàrids, sembla que l’anus és atrofiat). L’estomodeu (cavitat bucal i esófag) i el proctodeu (recte) són recoberts de cutícula. La boca, que s’obre en l’extrem terminal (o subterminal) anterior, és una obertura circular envoltada per, com a màxim, sis llavis (de vegades, reduïts o absents), que duen dues corones de papil·les labials lliures o soldades; a més, hi ha quatre papil·les cefàliques darrere els llavis, conjuntament amb els amfidis, un pareil d’òrgans sensorials situats a banda i banda del cap, que són molt més desenvolupats en els nematodes de vida lliure que no en els paràsits. La cavitat bucal, o estoma, situada entre la boca i l’esòfag, pot tenir una importància molt variable. Moltes vegades constitueix una prominència ben distinta, i mostra regions diferents: el queilostoma (l’anterior), el protostoma, el mesostoma, el metastoma i el telostoma (la posterior); totes aquestes regions són revestides de cutícula, i cada tram o rabdió rep també noms específics (queilorabdió, prorabdió, etc.). En moltes espècies paràsites hi ha un estilet bucal, rígid i esmolat, buit per dins, que serveix per a perforar els teixits de l’hoste i obtenir l’aliment; aquest estilet consta d’una zona anterior (el con) i una de posterior (el vàstag o la base), que presenta tres nòduls basals. En d’altres hi ha una estructura que forma una mena de dent esmolada, massissa, que surt de la paret de l’esòfag (en Dorylaimus, per exemple). En d’altres, la paret de la cavitat bucal és gruixuda i forma una estructura rígida anomenada càpsula bucal, sovint armada de làmines foliàcies, llancetes, dents, etc., que actuen d’òrgans de fixació i perforació. L’esòfag és un òrgan musculós de secció triradiada, que actua d’òrgan de succió d’aliment; per això, disposa d’una musculatura especial i de tres glàndules esofàgiques (la dorsal més gran), que produeixen secrecions digestives i anticoagulants. En diversos grups, l’esòfag s’eixampla en determinats punts, formant els bulbs, ja sigui a la part distal de l’esòfag (bulb basal, terminal o esofàgic) o a la part mitjana (bulb mitjà); també pot aparèixer un pseudobulb, que no presenta aparell valvular. L’esòfag comunica amb l’intestí per mitjà d’una vàlvula esofàgico-intestinal. L’esòfag pot tenir formes molt diferents, i hom en distingeix diferents tipus, d’una gran importància per a la classificació de les espècies.
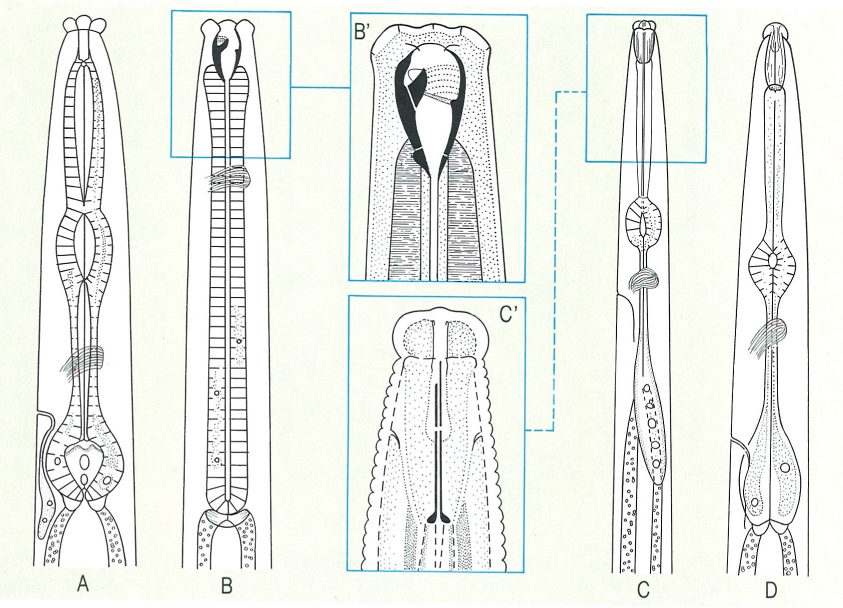
La morfologia de l’esòfag és un caràcter important en els nematodes, ja que pot presentar molts tipus diferents, que hom utilitza per a la determinació de les espècies. Hom n’ha representat quatre casos diferents: A esòfag de tipus rabditoide (Rhabditis); B esòfag de tipus cilíndric (Mylonchulus), i B′ detall de la regió cefàlica, amb l’estoma ample, proveït de parets fortament cuticularitzades i d’una dent dorsal; C esòfag de tipus afelencoide (Aphelenchus) i C regió cefàlica de l’estilet (Aphelenchoides), amb nòduls basals poc desenvolupats i regió labial llisa i poc separada del cos; D esòfag de tipus tilencoide (Tylenchorhynchus).
Biopunt, a partir de fonts diverses
L’esòfag moniliforme (característic dels triquiniel·loïdeus) té una primera part muscular cilíndrica i curta, i una part posterior llarga de naturalesa glandular, anomenada esticosoma, que és constituïda per cèl·lules rectangulars (esticòcits), de funció secretora. L’esòfag cilíndric o tubular (típic dels espirúrids i els filarioïdeus) té l’aspecte d’un tub de diàmetre uniforme, encara que té una primera part muscular i una segona part, més llarga i ampla, glandular. L’esòfag estrongiloide (típic dels estrongílides) té forma de maça de beisbol, i es va eixamplant cap enrere. L’esòfag oxiuriforme (típic dels oxiúrides, però també present en els cosmocèrides, els heteràkides i els subulúrides), té la part anterior estrongiloide i un bulb terminal esfèric. L’esòfag rabditoide (típic dels rabdítids) té la part anterior (procorpus) cilíndrica, seguida d’un pseudobulb (metacorpus) i finalment d’una zona cilíndrica (istme) i un bulb posterior. L’esòfag seuratiforme o claviforme (típic dels seuràtides) té forma de porra i només s’hi diferencia, pel diàmetre, una part anterior, més curta i prima. L’esòfag ascaridoide (típic dels ascarídides) és llarg, cilíndric i muscular, uniforme; però en diversos gèneres hi ha, en el punt d’unió entre l’esòfag i l’intestí, una zona molt especialitzada, que pot consistir simplement en un tram curt glandular anomenat ventricle (en Toxocara), o bé pot ésser més complex, amb entre 1 i 5 cecs esofàgics originats al ventricle i dirigits cap enrere (en Raphidascaris i Contracaecum) o cap endavant (en Multicaecum), o bé encara pot complicar-se més amb l’aparició d’un (en Angusticaecum, Porocaecum, Paradujardinia, Multicaecum, Contracaecum) o dos (en Crossophorus) cecs intestinals que parteixen de la part anterior de l’intestí i es dirigeixen cap a l’esòfag. L’esòfag dorilaimoide (típic dels dorilàmides de vida lliure) és semblant al tipus espiruroide, cilíndric o tubular, però amb la part anterior prima més llarga que la posterior. L’esòfag diplogasteroide (típic dels diplogastèrides paràsits d’insectes) té la regió anterior muscular que acaba fent un bulb mitjà, seguit d’una regió glandular posterior que acaba en un pseudobulb. L’esòfag tilencoide (típic dels tilènquides) és semblant al diplogasteroide però té la regió anterior més prima (típic de Tylenchus); no obstant això, també es pot complicar, per exemple perquè la part glandular es projecti cap enrere i se superposi a la regió intestinal anterior (en Helicotylenchus i en Aphelenchus), o bé pot ésser que no hi hagi bulb mitjà (Neotylenchus). L’esòfag mermitoide (típic dels mermitoïdeus paràsits d’invertebrats quan són joves, i de vida lliure quan són adults) es compon d’una part anterior tubular, llarga i molt prima, i una segona part ampla i encara més llarga (esticosoma); els esticòcits són inclosos dins el teixit esofàgic i no fent una fila.
L’intestí és una estructura simple, en forma de tub, no muscular, feta per una capa de cèl·lules epitelials intestinals amb microvellositats. Hom hi ha definit tres regions diferents, segons el gruix del lumen i les funcions, tot i que són difícils de distingir morfològicament: la regió anterior o ventricular és essencialment secretora, mentre que la regió mitjana i la regió posterior o prerectal són de naturalesa absorbent. El recte és un tub curt i aplanat. En les femelles, el recte va a parar a l’anus (terminal o subterminal), mentre que en els mascles desemboca a la cloaca, on dona també l’aparell reproductor, a través del conducte deferent.
El sistema nerviós és relativament simple. Hi ha dos anells nerviosos, l’un a la regió esofágica i l’altre a la regió anal. El centre nerviós principal és constituït per un collar de ganglis al voltant de l’esòfag (anell nerviós anterior), dels quals surten cap enrere els troncs nerviosos longitudinals que recorren longitudinalment el cos per l’interior dels cordons hipodèrmics, i cap endavant els destinats a innervar els òrgans sensorials cefàlies. Els troncs nerviosos posteriors s’uneixen entre ells per fines anastomosis transversals i prop del recte formen la comissura rectal o anell nerviós posterior. Hi ha, a més dels òrgans sensorials relacionats amb la boca, altres òrgans sensorials. Les papil·les cervicals o deiridis, es troben en la majoria dels nematodes paràsits en nombre de dos, situades a l’altura de l’anell nerviós anterior. Les papil·les caudals són més desenvolupades en els mascles i tenen algun paper en la còpula. Els anomenats fasmidis (la presència o absència dels quals permet distingir els dos grans grups de nematodes) són un parell d’òrgans cuticulars, bilaterals, situats a l’extrem posterior del cos, que tenen una estructura semblant als amfidis.
El sistema excretor, excepte en alguns grups, en què manca (tricurats i dioctofimats), no és comparable al sistema protonefridial dels platihelmints. En general hi ha un porus excretor a la cara ventral, situat a diferents punts de la regió esofàgica). Es distingeixen dos tipus bàsics de sistema excretor. El tipus glandular (propi dels afasmidis o adenoforis, de vida lliure) és constituït per cèl·lules glandulars (renetes o glàndules renoides), que poden desembocar independentment en el porus excretor o bé a través d’un peduncle comú. El tipus tubular (més típic de les formes paràsites) consisteix bàsicament en dos conductes llargs que corren per dins dels cordons hipodèrmics laterals i es connecten entre ells per un conducte transversal (per la qual cosa adopta la forma de la lletra H), prop de la regió anterior, del qual neix un conducte que va a desembocar al porus excretor; aquest sistema pot presentar moltes variacions.
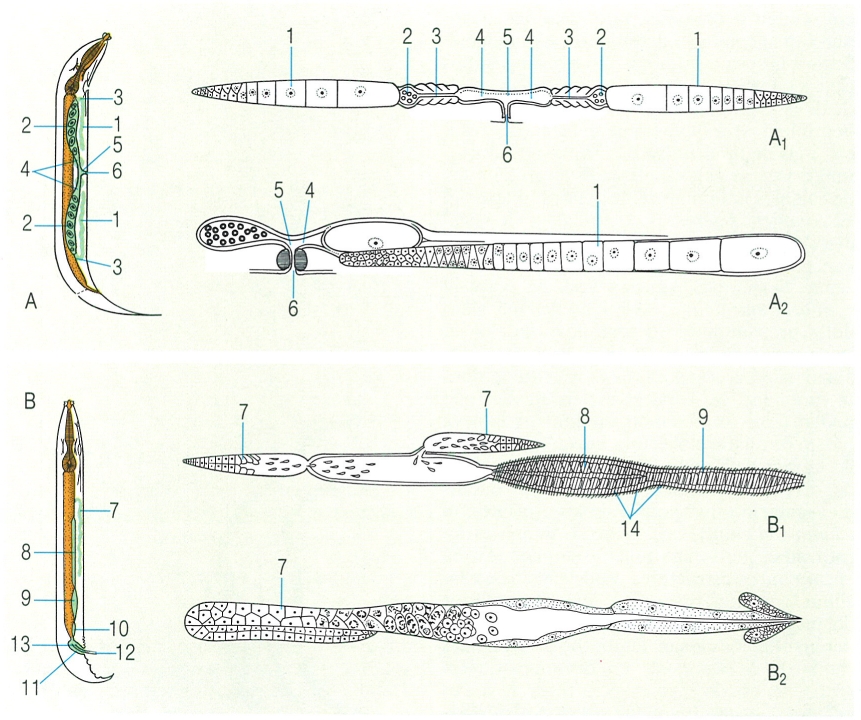
Localització de l’aparell reproductor femení (A) i masculí (B) en els nematodes mascles i femelles, respectivament, i tipus d’aparell reproductor. A1 aparell reproductor femení amfidelf recte, A2 aparell reproductor femení opistodelf retroflex, B1 aparell reproductor masculí d’un enòplide, amb la vesícula seminal i el vas deferent molt musculós, B2 aparell reproductor masculí d’un secernenti, sense musculatura però amb glàndula present. Hom hi ha indicat: 1 ovari i glàndula vitel·lògena, 2 úter, 3 oviducte, 4 ovojector, 5 vagina, 6 vulva, 7 testicle, 8 conducte deferent o espermiducte, 9 vesícula seminal, 10 conducte ejaculador, 11 cloaca, 12 espícules, 13 governacle, 14 bandes musculars.
Biopunt, a partir de fonts diverses
La majoria dels nematodes són dioics. Mascles i femelles són molt semblants externament, excepte en les estructures reproductores i en les dimensions (les femelles són més grosses que els mascles); en algunes espècies, però, hi ha un dimorfisme sexual molt marcat, especialment en tetramèrids zooparàsits i heterodèrids fitoparàsits, els mascles dels quals presenten l’anatomia típica dels nematodes, mentre que les femelles són simples bosses d’úter inflades. En algunes espècies només es coneixen les femelles, i hom les considera partenogenètiques. L’hermafroditisme s’ha comprovat en pocs casos (Heterogonema ovomasculis).
Les gònades són cordons sòlids de cèl·lules que per un extrem es transformen en conductes i desemboquen a l’exterior. En els mascles hi ha generalment un sol testicle (monòrquides), però n’hi pot haver dos (diòrquides). El testicle pot ésser relativament curt i sense enrotllar, però en els nematodes grossos sol ésser molt llarg i s’embolica al voltant de l’intestí. En el testicle es distingeixen dues zones: la zona germinal o distal, on tenen lloc les divisions espermatogèniques, i la zona de creixement o proximal, que és la que continua cap al conducte deferent o espermiducte. Segons la seva morfologia, hom distingeix dos tipus de testicles: els de tipus telogònic, en els quals la proliferació de cèl·lules germinals té lloc solament a l’extrem cec, distal, del testicle, i els de tipus hologònic (típic dels dioctofimats i tricurats), en els quals la proliferació de cèl·lules germinals té lloc en tot el testicle. El conducte deferent o espermiducte de vegades s’eixampla per la part terminal i forma la vesícula seminal o òrgan destinat a emmagatzemar l’esperma. Aquesta vesícula porta al conducte ejaculador, sovint proveït de glàndules prostàtiques, i desemboca a la cloaca.
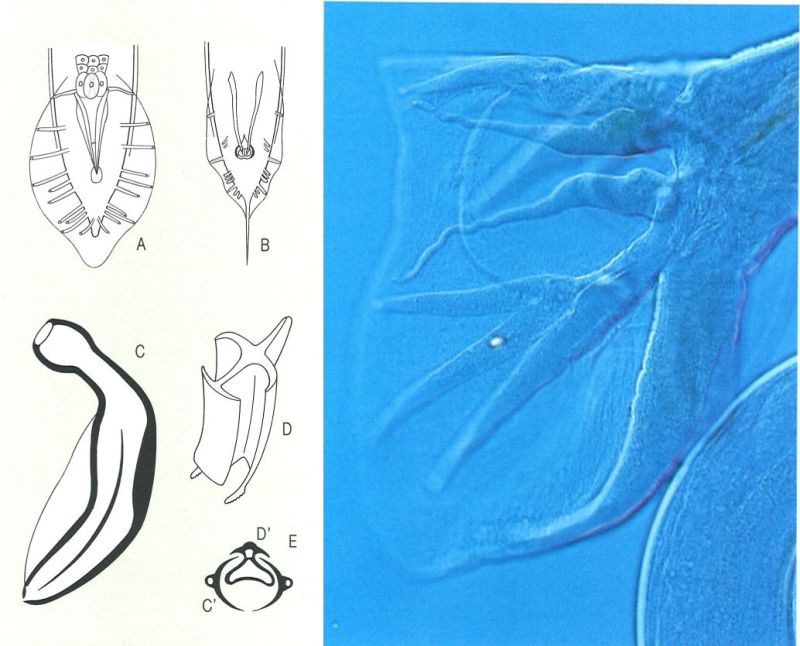
Les estructures auxiliars de la còpula adquireixen diferents nivells de complicació en els mascles dels nematodes. En el dibuix, hom ha representat: A bossa copuladora o bursa pelodera, típica dels rabdítides, B bursa leptodera, C espícula copuladora, D governacle, i E tall transversal de les peces copuladores d’un mascle d’una altra espècie, on s’aprecia la disposició de les espícules (C’) i el governacle (D’). La fotografia mostra un detall de l’extrem posterior d’un mascle d’Heligmosomoides polygyrus, que mostra la bursa o bossa copuladora, amb els lòbuls ben desenvolupats.
Biopunt, a partir de fonts diverses; Santiago Mas-Coma.
A l’interior de la cloaca hi ha les estructures auxiliars de la còpula: les espícules (una, dues o, rarament, cap), peces esclerotitzades que poden tenir formes molt diferents, i la seva funció és dilatar l’obertura vulvar de la femella durant la còpula; el governacle, una peça dorsal esclerotitzada que guia els moviments d’evaginació i invaginació de les espícules; el telamó (telamon), peça ventral, també esclerotitzada, que té una funció semblant. Hi ha encara altres estructures especialitzades a facilitar la còpula, presents a l’extrem posterior del mascle, com les papil·les genitals (molt ben desenvolupades quan també ho són les ales caudals), de les quals en distingim quatre grups, segons la seva posició respecte a l’obertura cloacal (papil·les precloacals, cloacals, postcloacals i caudals). Quan les ales caudals són dues simples expansions laterals de l’extrem caudal del mascle (en espirúrides), s’anomenen bossa caudal, i quan són molt desenvolupades, es parla de bossa copuladora o bursa; aquesta bossa, circular o oval, sovint és dividida en dos lòbuls laterals separats per un lòbul dorsal i sostinguda per unes expansions digitiformes anomenades radis o costelles (en els ancilostomatoides, els estrongiloides, els tricostrongiloides i els metastrongiloides); la morfologia i la disposició d’aquestes costelles, com la de totes les altres peces anomenades de l’aparell copulador, tenen una gran importància taxonòmica.

La regió vulvar de les femelles dels nematodes pot presentar diferents estructures situades a la cara ventral de l’animal. Les fotografies mostren, a dalt, la regió uterina terminal d’una femella de Trichuris muris, amb ous madurs a l’interior, i, a baix, un detall de la vulva de l’espècie Aspiculuris tetraptera, en el moment d’expulsar un ou. Ambdues espècies són paràsites del ratolí de bosc (Apodemus sylvaticus).
Santiago Mas-Coma.
L’aparell reproductor femení en la majoria dels nematodes consta de dos ovaris (en algunes espècies, n’hi ha fins més de sis), i el model general és molt semblant al dels mascles, excepte pel que fa a l’orifici genital, que desemboca, independentment del sistema digestiu, directament a l’exterior. Els ovaris són cordons sòlids, tubs llargs embolicats al voltant del tub digestiu. L’extrem proximal de l’ovari és la zona germinal, que produeix els oogonis, els quals després passen a la zona de creixement de l’ovari, en direcció a l’oviducte. A l’oviducte hi ha el crustaformeri, propi dels tilènquides. Més endavant, l’oviducte s’eixampla lleument per a formar el receptacle seminal o espermatoteca, que l’uneix a l’úter. L’úter és tubular i acaba formant la vagina, la qual dona a la vulva o orifici femení, situat sempre a la cara ventral de l’animal. La vulva és formada per dos llavis, de vegades sortints; en alguns grups hi ha una estructura cuticular anomenada epistigma, d’interès taxonòmic. En algunes espècies, entre el tram final de l’úter i la vulva hi ha una regió muscular, l’ovojector, que expulsa els ous a través de la vulva. Poden haver-hi un, dos o més tubs uterins, i segons això hom parla de femelles monohistèrides o monodelfes, femelles dihistèrides o didelfes(el cas més freqüent), i femelles polihistèrides o polidelfes. Entre les femelles didelfes, hom distingeix encara entre les amfidelfes ( Strongyloides stercoralis i els tricostrongílides), que tenen un úter dirigit cap endavant i l’altre cap enrere, que al final conflueixen en una vagina, i aquesta amb una vulva situada a la part mitjana del cos, les prodelfes, quan tots dos úters corren paral·lels i convergeixen en una vagina i una vulva que s’obren a la regió posterior del cos (en els metastrongiloïdeus), i les opistodelfes, quan els úters corren paral·lels però procedeixen de la regió posterior i conflueixen en una vagina i una vulva situades en posició anterior (en els filàrids i el gènere Enterobius).
Biologia i ecologia

Aparellament de dos nematodes.
Fernando García del Pino.
En els nematodes la fecundació és obligatòriament creuada, ja que les espècies presenten els sexes separats. De tota manera, hi ha algunes excepcions, tant d’espècies hermafrodites com d’espècies que tenen femelles partenogenètiques. En general, el mascle copula amb la femella ajudat dels seus òrgans coadjuvants de la còpula (ales caudals, papil·les caudals, espícules guiades pel governacle, bossa copuladora). Els espermatozoides, allargats, cònics o esfèrics, desproveïts de flagel, amb moviments ameboides, són dipositats a la vagina de la femella i ascendeixen pel conducte uterí fins a fecundar els òvuls. En les espècies en què la femella és proveïda de receptacle seminal, un únic intercurs sexual pot ésser suficient. En algunes espècies s’ha pogut observar, però, que la femella copula seriadament amb diferents mascles. I se’n coneixen algunes espècies en les quals, un cop el mascle ha trobat una femella, l’envolta amb la seva potent bossa copuladora i es manté en còpula durant la resta de la seva existència adulta ( Pseudophysaloptera kahmanni, paràsit estomacal de la rata cellarda, a les Balears).
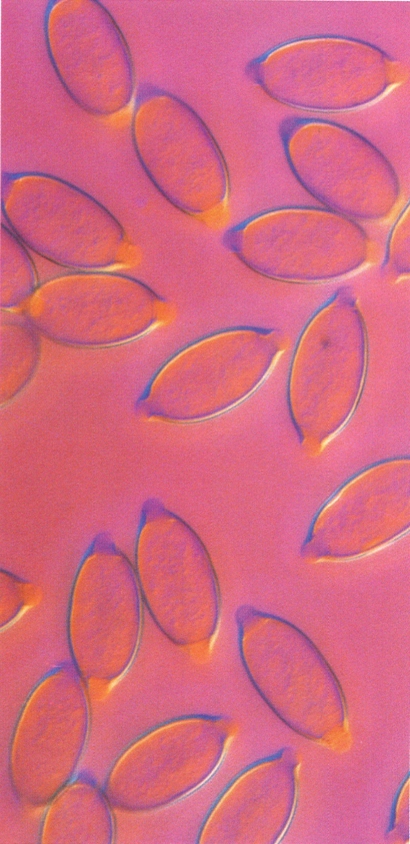
Ous del nematode zooparàsit Trichuris muris, als quals es veuen perfectament els dos taps polars.
Santiago Mas-Coma.
Els ous tenen una coberta que, en la majoria dels nematodes, és constituïda per tres capes: una membrana vitel·lina, de naturalesa lipoïdea, al voltant del zigot; una capa quitinosa denominada còrion, que constitueix la closca de l’ou; i una membrana albuminosa, la més externa, de naturalesa proteica, que pot ésser llisa o ornamentada. La forma i l’estructura dels ous dels nematodes no varien gaire, i en general se’n poden distingir fonamentalment tres tipus bàsics: de closca fina, normalment el·líptics i allargats (ancilostòmides, filàrides i rabdítides); de closca gruixuda, més grossos i arrodonits (ascàrides); amb taps polars, en forma de llimona, amb la closca gruixuda i amb dos taps mucosos albuminoides als extrems (triquiúrides i capil·làrides). Els ous, un cop formats a dins la femella, són expulsats a l’exterior. En el moment de sortir per la vulva poden contenir una sola cèl·lula ou, o bé un blastòmer, o bé un embrió (sempre desproveït de cilis o ganxos). Pel que fa a la posta, cal distingir entre les femelles ovípares (alliberen els ous quan aquests contenen una sola cèl·lula ou o un blastòmer) i les femelles ovovivípares (el desenvolupament de l’embrió dins de l’ou es fa quan aquest és encara dins l’úter matern, de manera que quan la femella allibera els ous, aquests ja duen un embrió), i, finalment, les femelles vivípares (alliberen embrions o larves lliures, ja que l’ou desclou dins l’úter).
El desenvolupament postembrionari dels nematodes comprèn, en general, un total de cinc estadis evolutius: primer, segon, tercer i quart estadis larvals, i un cinquè estadi, que ja és el d’adult (des de jove immadur fins a adult madur). Cadascun dels estadis passa al següent després de passar una muda; hi ha, per tant, un total de 4 mudes al llarg del desenvolupament. Qualsevol procés de muda, també anomenat d’ecdisi o desembeinament, comprèn la formació d’una nova cutícula, la pèrdua de la cutícula anterior i la ruptura d’aquesta cutícula antiga i sortida de la larva. Sembla que les larves dels nematodes secreten un fluid que facilita la muda justament abans de perdre la cutícula antiga, i que aquest fluid ataca la cutícula solament des de l’interior i en una regió concreta. Cal fer notar la diferència d’aquest tipus de muda amb el que és característic dels artròpodes: en els nematodes les dimensions del cos continuen creixent entre una muda i una altra (bé que, excepcionalment, pugui haver-hi un període de letargia immediatament abans i després de la muda). No obstant, en alguns casos, un estadi larval pot mantenir la coberta cuticular de l’estadi anterior; es tracta de les anomenades larves embeinades. Això sol passar entre el segon i el tercer estadis larvals, coincidint amb la segona muda. Aquestes larves embeinades són de vida lliure i solen aparèixer com una adaptació a les condicions desfavorables del medi extern; moltes vegades són resistents a la dessecació. En altres casos, la larva infestant pot ésser ingerida per un hoste que no és l’apropiat com a hoste definitiu, i llavors aquestes larves travessen la paret del tub digestiu de l’hoste i hi romanen en estat de latència. És el cas de l’anomenada paratènesi i dels hostes paratènics. Hom utilitza altres termes per a designar estadis larvals concrets de certs grups de nematodes, que, per exemple, poden fer referència al tipus d’esòfag de la larva ( larva rabditoide, corresponent al primer estadi larval; larva estrongiloide, corresponent al segon i tercer estadis larvals) o no ( larva microfilària, un estadi de desenvolupament embrionari molt precoç, anterior al primer estadi larval, en els filarioïdeus).

El tipus d’alimentació dels nematodes és molt variat i hom pot veure-hi una relació amb les estructures bucals, especialitzades en cada cas a xuclar, mastegar, etc. Les fotografies mostren, a dalt, la regió cefàlica d’una femella de Prionchulus, de règim alimentari carnívor, que s’alimenta de petits organismes, incloent-hi altres nematodes. A baix, l’estilet de Macroposthonia xenoplax (criconemàtid), ectoparàsit molt freqüent al Penedès, causant de la clorosi de la vinya; la femella de la fotografia (feta al microscopi electrònic d’escandallatge) fa 0,3 mm de longitud.
Alejandro Palomo.
Els nematodes es nodreixen de microorganismes o macroorganismes vius o bé de teixits de plantes i animals. Hom reserva el qualificatiu de depredador o macròtrof per als nematodes que mengen animals pluricel·lulars, i anomena micròtrofs els que s’alimenten de microorganismes; tota la resta dels nematodes poden ésser agrupats en fitòfags o fitoparàsits, i zoòfags o zooparàsits. Els nematodes macròtrofs solen tenir l’obertura bucal àmplia, cilíndrica o en forma de tassa, i les parets poden tenir dents o altres estructures per a la trituració de l’aliment. Molts nematodes micròtrofs terrestres tenen un esòfag muscular, amb una valva situada sota el bulb terminal (rabditoïdeus) o a la regió mitjana de l’esòfag (diplogasteroïdeus), o valves a totes dues regions (pseudodiplogasteroïdeus); la seva funció és fer avançar l’aliment cap a l’intestí.
El paper dels nematodes micròtrofs aquàtics i terrestres en el reciclatge de la matèria orgànica és molt important, ja que ocupen el segon graó de la cadena tròfica, just per sobre dels bacteris i fongs, com a descomponedors. Els terrestres són capturats per altres invertebrats, i els aquàtics, per peixos.
Els nematodes edàfics viuen just per sota de la superfície del sól, de manera que estan molt condicionats pels canvis ambientals. Durant els períodes secs, quan les condicions físiques són adverses, molts nematodes bacteriòtrofs, sia en forma d’adults, d’ou o en l’etapa juvenil, responen amb la criptobiosi (pèrdua de l’activitat metabòlica, sense producció de CO2 ni de productes de desfet) o la quiescència (període d’inactivitat, amb disminució de l’activitat metabòlica); de vegades la quiescència inclou el manteniment de formes juvenils anomenades larves dauer, considerades com una forma de dispersió, ja que es poden fixar a l’exosquelet dels artròpodes. Una altra estratègia davant la manca d’aliment (bacteris) en l’època desfavorable consisteix, en el cas de les femelles, en no expulsar els ous a l’exterior sinó deixar que es desenvolupin al seu interior, a costa dels seus propis teixits.
Filogènia i sistemàtica
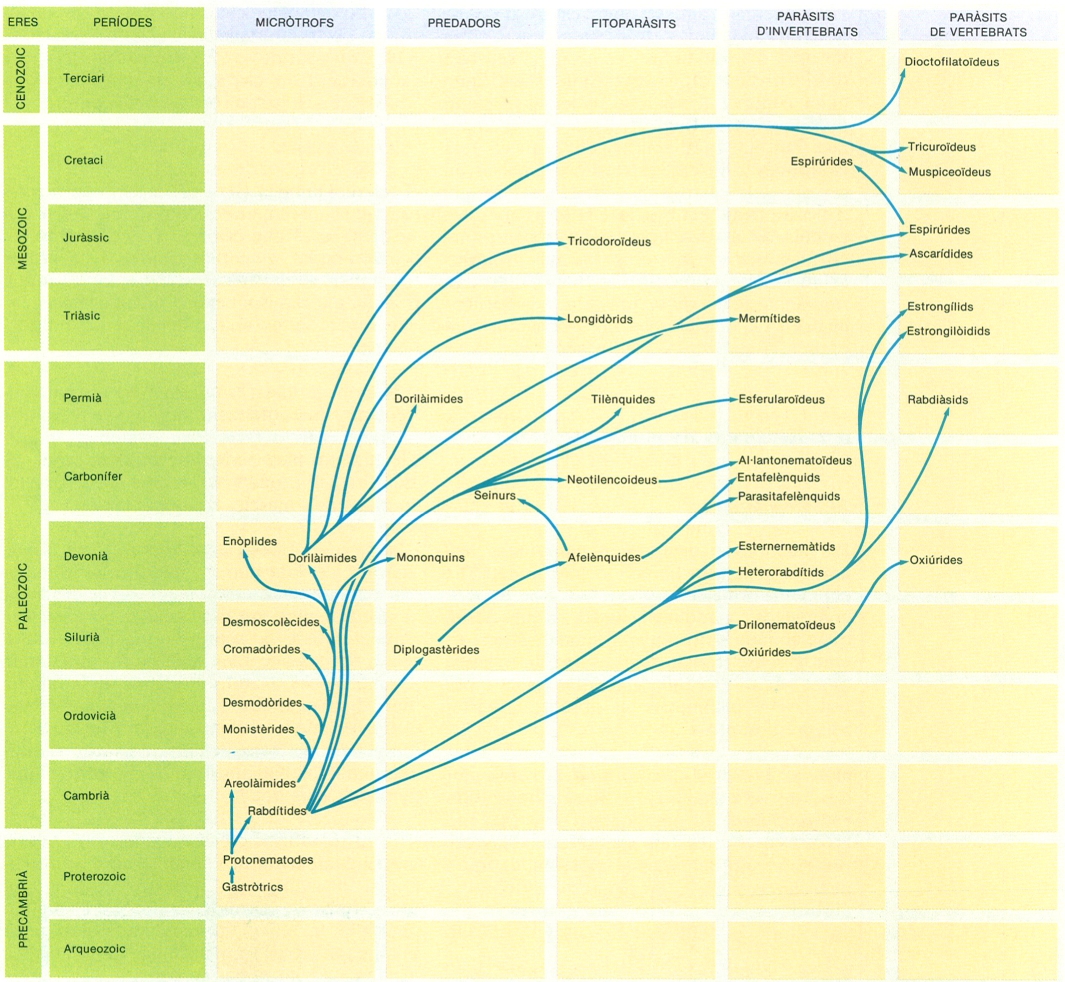
Esquema de les relacions filètiques més probables entre els principals grups de nematodes secernentis.
Carto-Tec, a partir de GO. Pomar, 1983
El fílum dels nematodes (Nematoda) és subdividit en dues classes: la dels adenoforis (Adenophorea) i la dels secernentis (Secernentea). La classe dels adenoforis comprèn la subclasse dels cromadoris (Chromadoria) i la dels enoplis (Enoplia).
La filogènia d’aquest fílum és encara força desconeguda, tant pel que fa a la seva afinitat amb d’altres fílums, és a dir, el seu punt d’origen d’entre els organismes pseudocelomats, com pel que fa a les relacions filètiques que uneixen els mateixos grups de nematodes. Pel que fa al seu origen, hom els relaciona més amb els gastròtrics o amb els ancessors dels gastròtrics actuals que no amb d’altres grups. Pel que fa a la filogènia interna dels nematodes, hom tendeix a pensar que els adenoforis deuen ésser els més primitius, ja que inclouen les formes lliures, que haurien donat lloc a dues grans línies, la dels cromadoris i la dels enoplis; els secernentis comprenen la majoria de les formes paràsites.
Els grups de nematodes
Els adenoforis
Els adenoforis comprenen nematodes amb amfidis generalment ben desenvolupats (excepte en formes paràsites), sovint amb porus complexos, glàndules caudals i hipodèrmiques comunes, fasmidis absents, sistema excretor sense canals laterals, format bàsicament per una cèl·lula glandular única i ventral, o bé completament absent, i deiridis sempre absents.
Dels nou ordres que formen la classe dels adenoforis, solament un, el dels mermítides (Mermithida) és enterament constituït per espècies paràsites (en aquest cas, d’invertebrats); els dorilàimides (Dorylaimida) contenen alguns paràsits de plantes i vertebrats, i els altres set són essencialment micròtrofs de vida lliure o depredadors: els monistèrides (Monhysterida), els desmodòrides (Desmodorida), els desmoscolècides (Desmoscolecida), els cromadòrides (Chromadorida), els monònquides (Mononchida), els enòplides (Enoplida) i els areolàimides (Araeolaimida).
Els cromadoris
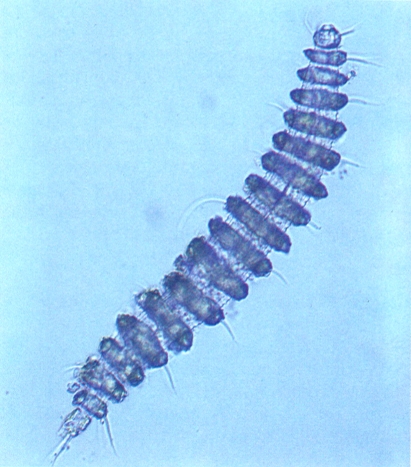
Desmoscolècide del gènere Desmoscolex. Noteu l’aspecte especial de la cutícula, amb els anells ben marcats, i les setes locomotores externes, que li donen una aparença similar a la dels poliquets.
David Masalles
Els areolàimides (Araeolaimida) són formes de vida lliure, marines, dulciaqüícoles o terrestres, totes micròtrofes. Tenen amfidis espirals amplis, glàndules caudals i un esòfag muscular. Presenten tres llavis dobles i quatre setes al cap; les papil·les genitals dels mascle són tubulars i la cutícula sense puntuacions.
Els monistèrides (Monhysterida) són tots micròtrofs, principalment marins, però també d’aigua dolça i terrestres; contenen amfidis circulars, o més o menys espirals; l’esòfag és cilíndric i el cap té 4, 6 o 8 setes, i les papil·les genitals dels mascles són papil·loides.
Els desmodòrides (Desmodorida) són també marins, de vida lliure, i micròtrofs: la majoria s’alimenten de diatomees, algues i protozous. Tenen els amfidis circulars o espirals, la cutícula anellada, sense puntuacions i l’estoma o cavitat bucal variable de forma (simple o amb dents). Alguns membres de la família dels draconemàtids (Draconematidae) tenen setes locomotores ventrals, amb les quals es desplacen pel fons del mar, cas que, juntament amb el d’alguns desmoscolècides és únic entre els nematodes.
Els cromadòrides (Chromadorida), també principalment marins, amb alguns representants dulciaqüícoles i terrestres, tots micròtrofs, presenten amfidis amb espiral simple o múltiple, la cutícula ornamentada amb punts o altres dibuixos formant línies transversals, i l’estoma sovint armat de dents.
Els desmoscolècides (Desmoscolecida) són tots marins (excepte algunes Desmoscolex, que són d’aigua dolça i terrestre), micròtrofs, i es caracteritzen perquè tenen una cutícula amb anells prominents i setes, el cap més o menys triangular, amb llavis i estoma reduïts, i perquè tenen tantes setes locomotores a la seva superfície (dorsal i ventral) que és fàcil de confondre’ls amb anèl·lids.
Els enoplis
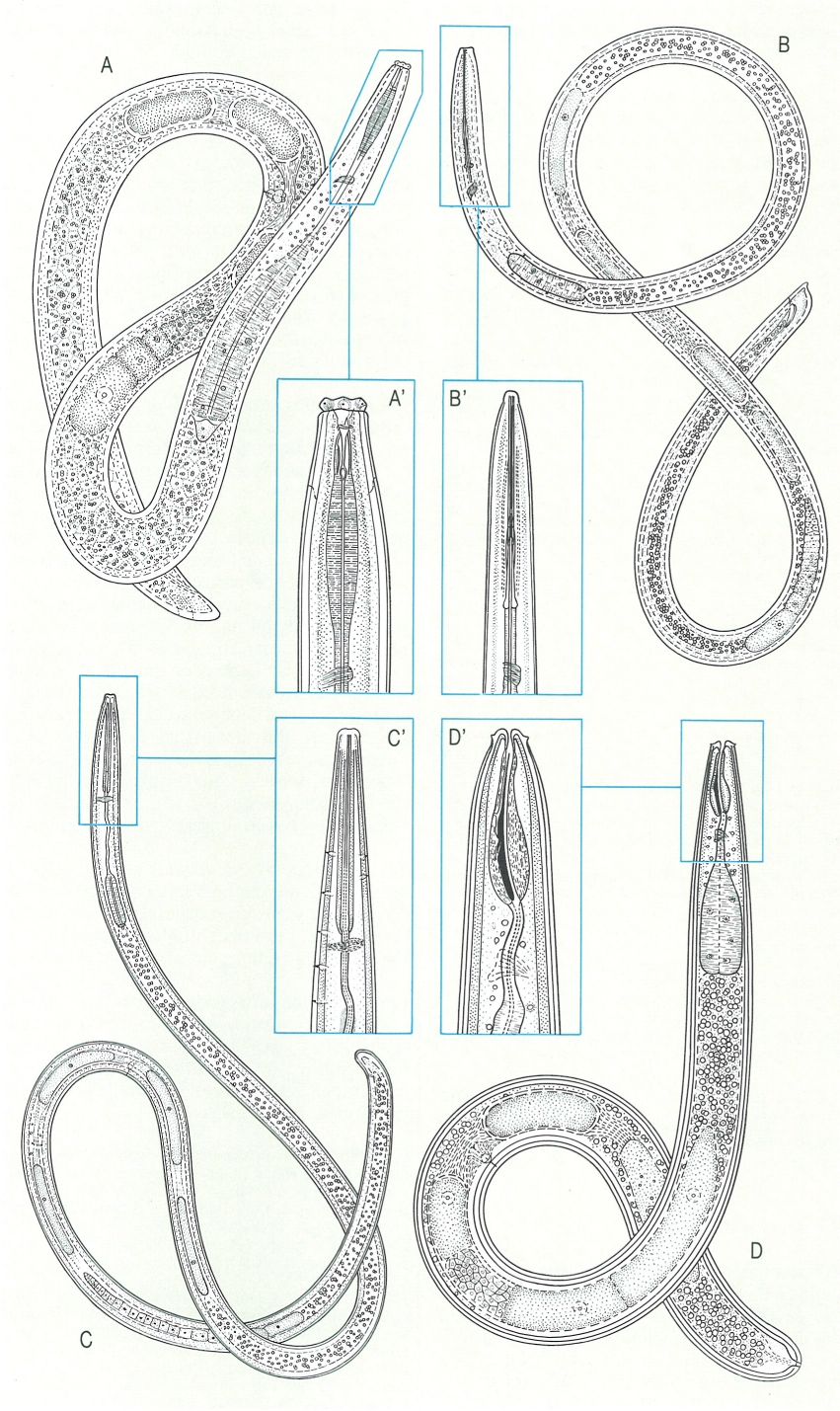
Morfologia general dels dorilàimides i detall de la regió anterior del cos, en alguns exemples trets de la nematofauna catalana. Els dorílàimides tenen una gran importància en l’agronomia per la seva qualitat de transmissors de virus infecciosos, causants de malalties per a les plantes de conreu. A aspecte general de Dorylaimus. amb l’estilet curt, l’esòfag cilíndric i l’ovari amfidelf reflex, i detall (A’) de la part anterior, amb sis llavis angulars, separats del cos per una constricció, i un amfidi lateral en forma de copa. B aspecte general de Xiphinema, amb l’esòfag estret en la zona anterior i cilíndric en la posterior, i els ovaris amfidelfs o opistodelfs, i detall (B’) de la part anterior del cos, amb l’odontstilet llarg, amb l’anell guia en el terç basa I, l’odontòfor acabat en un nòdul trilobulat, i l’amfidi lateral en forma de copa. C Aspecte general de Longidorus, amb l’esòfag cilíndric estret al davant i més ample al darrere, i els ovaris amfidelfs, i detall de la regió anterior (C), amb l’odontòstil amb l’anell guia situat prop de la regió labial, l’odontòfor sense engruiximent basal i els amfibis laterals i en forma de bosses. D Aspecte general de Trichodorus, amb l’esòfag cilíndric i acabat en un bulb basal piriforme, l’ovari amfidelf i la cua curta, de rodona a hemisférica, i detall de la regió cefàlica (D’). amb llavis no diferenciats, amfidis en forma de porus i estilet corbat dorsalment.
Biopunt, a partir de fonts diverses
Els enòplides (Enoplida) són majoritàriament marins, tret d’alguns dulciaqüícoles i terrestres, i presenten amfidis en forma de butxaca o de tassa, el cap amb un cercle de sis papil·les labials internes o setes, sis papil·les labials externes o setes i quatre papil·les postlabials o setes; les dues darreres corones generalment són fusionades en un cercle de deu papil·les o setes; l’esòfag generalment és cilíndric o ben eixamplat a la base.
Els dorilàimides (Dorylaimida) constitueixen l’ordre que té més espècies micròtrofes o depredadores, tant d’aigua dolça com terrestres, però destaquen per la seva transcendència en l’agronomia com a transmissors de microorganismes infecciosos, que duen al tracte digestiu, a més de la seva acció directa sobre els conreus, ja que destrueixen les rels de les plantes.
Hom coneix quatre gèneres de dorilàimides transmissors de virus, que pertanyen a les famílies dels longidòrids (Longidoridae) i tricodòrids (Trichodoridae). Els longidòrids es diferencien de la resta de dorilàimides perquè tenen un estilet molt llarg, que introdueixen a les cèl·lules radiculars de les plantes, i per la gran llargada del seu cos (1,5 a 13 mm) en relació amb el diàmetre; el cos es corba per la part ventral i pren forma de lletra C, amb una curvatura més accentuada a la regió caudal, especialment en els mascles. L’estilet o odontòstil, és axial, buit per dins, i té una prolongació (odontòfor) que es continua amb l’esòfag i acaba en un bulb esofàgic posterior, musculós i força prominent. El bulb esofàgic té un conducte glandular dorsal i una glàndula a la seva part anterior, a més d’un parell de glàndules subventrals a la part central, amb una vàlvula entre l’esòfag i l’intestí (càrdies). El gènere Longidorus es caracteritza perquè té un anell guia, que dirigeix la sortida de l’estilet, senzill, a la part anterior de l’odontòstil; la unió de l’odontòstil amb l’odontòfor és simple, mai dentada, i l’odontòfor no presenta rebabes basals; la vulva, en les femelles, es troba a la part mitjana del cos. Les branques genitals són dobles, oposades, amb els ovaris retroflexos. Els amfidis, grossos i sacciformes, tenen una obertura petita i puntiforme. La cua és cònica i arrodonida. El gènere Xiphinema es caracteritza, en canvi, per tenir les obertures amfidials amples, que ocupen gairebé tota la base de la regió labial, els amfidis en forma d’embut o d’estrep, la base de l’odontòstil amb dues o tres dents en el punt d’unió amb l’odontòfor, i aquest amb rebabes prominents; a la base de l’odontòstil hi ha dos anells guia, el posterior dels quals és el més visible. La posició de la vulva és variable en les espècies amb gònades dobles, els ovaris són oposats i retroflexos, la vulva se situa a la part mitjana del cos (no quan la femella és monodèlfica), i la cua pot ésser arrodonida, amb un mugró terminal, cònica, allargada o filiforme. Des del descobriment de Xiphinema index com a espècie vectora del «fan leaf» de la vinya, el 1959, s’ha estudiat a fons la transmissió de virus per nematodes i s’ha comprovat que tant poden transmetre’ls els adults com les formes juvenils. Els nematodes els adquireixen i els transmeten en alimentar-se de l’extrem apical de les rels de la planta hoste. Les partícules del virus es retenen d’una manera selectiva.
Els mermítides (Mermithida) són nematodes filiformes que en l’estat juvenil parasiten insectes, mentre que en l’estat adult viuen lliures en el sòl i en l’aigua.
Els secernentis
Els secernentis comprenen nematodes amb amfidis generalment poc desenvolupats, amb porus petits i simples pròxims als llavis, glàndules caudals i hipodèrmiques absents, fasmidis presents, sistema excretor amb un o dos canals laterals, amb cèl·lules glandulars associades o sense, deltíridis usualment presents.

Extrem anterior del cos d’un tilènquide, Crossonema menzeli, en visió lateral. La fotografia ha estat feta al microscopi electrònic d’escandallatge.
Miquel Escuer / Alejandro Palomo.
En la classe dels secernentis, predominantment terrestre, hi ha quatre ordres exclusivament paràsits d’animals: els estrongílides (Strongylida), els ascarídides (Ascaridida), els oxiúrides (Oxyurida) i els espirúrides (Spirurida); dos més són compostos d’espècies paràsites de plantes: els tilènquides (Tylenchida) i els afelènquides (Aphelenchida), i són els nematodes pròpiament fitoparàsits; el darrer ordre, el dels rabdítides (Rhabditida) conté majoritàriament formes terrestres micròtrofs, que són paràsits obligats d’invertebrats i vertebrats. Són pocs els nematodes marins micròtrofs de la classe dels secernentis, però Rhabditis marina (de l’ordre dels rabdítides, intertidal i cosmopolita) ho és excepcionalment.
Els rabdítides (Rhabditida) de vida lliure i micròtrofs són essencialment terrestres, i sembla que la colonització de l’hàbitat marí ha estat relativament recent. Des del punt de vista del comportament, hom els considera entre els més primitius de la seva classe i de tots els nematodes, i alhora es creu que d’ells s’han originat la resta de línies filètiques dels nematodes secernentis paràsits (fitoparàsits i zooparàsits). És l’ordre present en un nombre més gran d’hàbitats. Les formes que ingereixen microorganismes presenten una valva en el bulb basal, mentre que les que ingereixen aliment semifluid extret de l’hoste presenten la valva degenerada o absent; en casos rars, el bulb basal també és petit. Dins l’ordre dels rabdítides destaca una espècie, de la família dels estrongiloídids (Strongyloididae), Strongyloides stercoralis, causant de l’estrongiloïdiasi o anguilulosi de l’home.
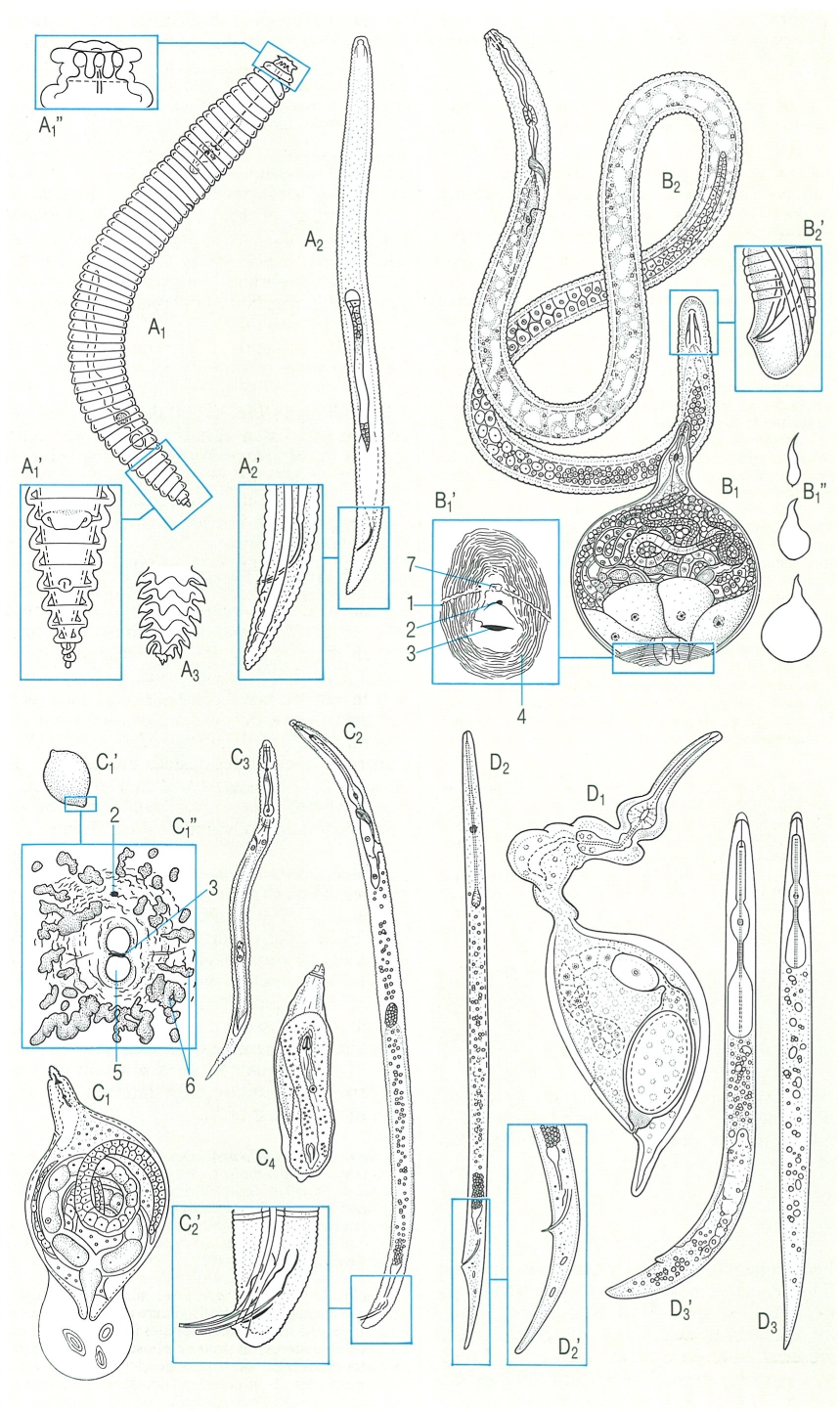
Morfologia general de les principals famílies de tilènquides. Hom hi ha representat l’aspecte general del cos i detalls de la regió cefàlica i el tipus d’esòfag. A Criconemàtids (Criconema annulifer): A1 femella adulta, amb l’esòfag del tipus criconematoïdeu, l’estilet llarg, l’ovari prodèlfic recte, i la vulva posterior; A1 regió caudal ventral, amb la vulva situada en l’anell desè, tancada i amb el llavi vulvar superior solapat a l’anterior, i l’orifici anal situat en el setè anell; A1" regió cefàlica, amb dos anells clarament diferenciats; A2 mascle de la mateixa espècie, sense esòfag ni etilet; A2’ regió caudal, amb l’espieula prima i corbada, i amb la bursa poc desenvolupada; A3 regió caudal d’un jove, amb les projeccions disposades en bandes longitudinals. B Meloidogínids (Meloidogyne): B1 femella adulta, amb un gran desenvolupament de l’ovari i de les glàndules rectals; B’1 tall transversal de la zona perianovulvar (1 banda lateral, 2 orifici anal, 3 orifici vulvar, 4 rugositats cuticulars, 7 final de la cua); B"1 aspecte extern d’algunes fases del desenvolupament; B2 aspecte d’un mascle; B2’ regió caudal, amb una espícula corbada i una banda lateral amb quatre línies, sense bursa. C1 Heterodèrids (He terodera): C1 femella; C’1 cist; C1" tall transversal de la zona perianovulvar (2 orifici anal, 5 semifenestra, 3 orifici vulvar, 6 bules); C2 aspecte general del mascle; C2 regió caudal, amb espícula corba, governacle, banda lateral amb quatre línies i sense bursa; C3 jove del segon estadi, a dins de l’ou, abans d’eclosionar; C4 jove del segon estadi. D Tilencúlids (Tylenchulus semipenetrans).’ D1 femella, amb l’ovari prodèlfic, la vulva posterior i el porus excretor situat prop de l’orifici vulvar; D2 mascle; D2’ regió caudal del mascle, amb una espícula prima i lleugerament corba, sense bursa; D3 jove; D3’ femella preadulta.
Biopunt, a partir de fonts diverses
Els tilènquides (Tylenchida) són paràsits de plantes, bé que també n’hi ha que en són d’insectes. Són fàcils de reconèixer per la presència d’estilet, per l’obertura de la glàndula esofágica dorsal (que se situa prop de la base de l’estilet), perquè tenen el bulb esofàgic mitjà ben desenvolupat i perquè el mascle no té mai costelles a la bossa copuladora. Les formes sedentàries i les paràsites d’insecte tenen el cos inflat, encara que els joves són sempre vermiformes. Esmentarem les famílies més importants d’aquest ordre.
Els tilènquids (Tylenchidae), cosmopolites i abundants en els sòls rics en algues i molses, tenen un estilet moderadament llarg (12-15 µm), el bulb esofàgic mitjà i basal ben desenvolupat, l’ovari prodèlfic, recte, i la bossa copuladora petita; la cua en general és filiforme, recta o corbada; Tylenchus, Filenchus, Aglenchus i Coslenchus en són els gèneres principals.
Els dolicodòrids (Dolichodoridae) són paràsits migradors, que solen produir atròfia i necrosi a les rels que parasiten; gairebé tots són cosmopolites i polífags i molt comuns en els conreus, els prats i els boscos; les plantes parasitades manifesten símptomes de clorosi, retard en el creixement i fins i tot poden morir, quan la infecció és acusada. Tylenchorhynchus goffarti té la cutícula amb anells d’1 µm d’ample, la regió cefàlica rodona, sortint del contorn del cos per una constricció; T. huesingi, més robust i de gairebé 1 mm de llargada, té la regió cefàlica rodona, però no sortint del contorn del cos; Merlinius brevidens té sis línies a la banda lateral, la regió cefàlica anulada i dividida en sis porcions; i Paratrofurus loofi té la regió cefàlica atenuada, llisa, i la cutícula molt ampla a la regió caudal.
Els hoplolàimids (Hoplolaimidae) són ectoparàsits migradors, amb un estilet robust i la carcassa cefàlica massissa; poden penetrar els teixits més profunds de la rel. Els del gènere Helicotylenchus són coneguts amb el nom de nematodes espirals, i tenen la glàndula esofàgica dorsal solapant la regió ventral de l’intestí; en el gènere Rotylenchus aquesta superposició és a la regió dorsal de l’intestí. Scutelonema té el fasmidi molt gran, situat a la cua o a qualsevol lloc del cos i les glàndules esofàgiques lliures, que se superposen a l’intestí dorsalment, lateralment o ventralment.
Els rotilencúlids (Rotylenchulidae) representen una transició entre les formes ectoparàsites i les endoparàsites sedentàries; les larves, mascles i femelles (preadults) viuen al sòl i són vermiformes. La femella preadulta és la que penetra la rel, indueix els teixits a formar cèl·lules gegants per a nodrir-se’n i s’engruixeix en la part posterior del cos.
Els pratilènquids (Pratylenchidae), allargats i prims, tenen una carcassa cefàlica ben desenvolupada, amb dos o tres anells, l’estilet moderadament desenvolupat (generalment fa menys de tres vegades l’amplada de la regió cefàlica), i les glàndules esofàgiques superposades a l’intestí. Són endoparàsits, i la seva acció patògena no solament es deu a l’acció directa sobre la rel sinó també al transport d’altres microorganismes (bacteris i fongs). Les espècies de Pratylenchus són monodèlfiques, amb sac postvulvar; en Pratylenchoides són didèlfiques, i amb glàndules esofàgiques lleugerament per sobre de l’intestí; en Zygotylenchus són didèlfiques, amb les glàndules esofàgiques molt allargades.
Els meloidogínids (Meloidogynidae) són endoparàsits sedentaris formadors de gal·les o nusos a les rels. Meloidogyne, el gènere més representatiu, pot arribar a constituir el principal factor limitador de la producció vegetal dels països tropicals i subtropicals; a les regions temperades hi ha menys espècies i solament són actius a l’estiu, però també són motiu de forts danys en fruiterars, horta, cereals i vinyes. Les espècies del gènere es diferencien per l’ornamentació de la zona perianovulvar i per la forma dels estadis juvenils. En l’actualitat s’empren tècniques serològiques i d’hibridació de DNA per a determinar les espècies.
Els heterodèrids (Heteroderidae) són nematodes que formen cists. Heterodera, el gènere principal, inclou endoparàsits sedentaris d’una gran importància econòmica per a l’agricultura. La femella adulta, obesa, manté la part anterior del cos clavada a la rel, on s’alimenta del síncit format per la ruptura de les parets cel·lulars. La paret del cos de la femella, un cop fecundada, s’engruixeix i es forma el cist, el qual resta al sòl, ple d’ous i de larves del segon estadi, capaços de resistir llargs períodes d’inactivitat, fins que les condicions de temperatura i humitat, i la presència de l’hoste, indueixen la seva activació. Heterodera forma complexos amb fongs patògens d’efectes molt més nocius que el sol nematode o el fong; per exemple, H. schachtii, associat a Rhizoctonia solani, parasita la remolatxa; H. rostochiensis, amb Verticilium dahliae, la patata.
Els criconemàtids (Criconematidae) tenen femelles i joves desproveïts de banda lateral, vermiformes i anellats externament; els mascles no tenen estilet, i són prims, amb banda lateral i anells molt petits. L’ovari és prodèlfic, recte, i els mascles tenen la bursa poc desenvolupada. Són ectoparàsits migradors àmpliament distribuïts per tota mena de sòls, conreats o naturals. Hi ha espècies que tenen projeccions en el marge dels anells, més freqüents en els medis naturals, mentre que els anellats i sense projeccions són més abundants en camps de conreu. En el gènere Macroposthonia la femella i els joves tenen el marge dels anells llis o carenat, mai amb projeccions, la regió cefàlica no diferenciada de la resta del cos i lòbuls sublaterals; la vulva és oberta, la vagina recta o sigmoide. Xenocriconemella macrodora té l’estilet molt llarg (un 50% de la llargada del cos) i és molt freqüent en boscos d’alzina. En Criconema, les formes juvenils tenen el marge dels anells amb projeccions disposades en bandes longitudinals, les femelles tenen el marge dels anells llis, la regió cefàlica amb un o dos anells clarament diferenciats, la vulva tancada i moltes vegades amb el llavi superior superposat a l’inferior. Ogma té el marge dels anells amb projeccions disposades en línies longitudinals. Crossonema presenta una banda contínua de projeccions en el marge dels anells. En Hemicriconemoides la femella té els anells rodons, la membrana cuticular és doble i els nòduls basals de l’estilet tenen forma d’ancla; els joves tenen projeccions.
Els hemicicliofòrids (Hemicycliophoridae) són àmpliament distribuïts pels sòls naturals i humits. Són anellats externament i amb la membrana cuticular doble, els anells rodons, sense lòbuls sublaterals, amb un estilet llarg i amb els nòduls basals ovals i amb el marge dirigit cap a la cua; els mascles tenen l’esòfag degenerat, la bursa molt desenvolupada i l’espícula llarga i prima, de corbada a gairebé semicircular.
Els tilencúlids (Tylenchulidae) tenen un marcat dimorfisme sexual: la femella té esòfag i un estilet ben desenvolupat, la meitat anterior del cos cilíndrica i la posterior corbada pel ventre i obesa, i el porus excretor situat prop de la vulva; el mascle, en canvi, no té ni esòfag ni estilet. Tylenchulus semipenetrans és el nematode dels cítrics.
Els paratilènquids (Paratylenchidae) són vermiformes, petits (menys de 0,5 mm), corbats pel ventre, amb la cutícula fina, anellada, i amb l’estilet dotat d’un con més llarg que la base. Són ectoparàsits obligats de moltes plantes, i formen poblacions molt nombroses. Paratylenchus neoamblycephalus pot restar com a forma de resistència en el quart estadi juvenil, i es reconeix perquè té l’estilet reduït o absent i l’esòfag degenerat.
Els anguínids (Anguinidae) s’alimenten de fongs i parasiten les parts aèries de les plantes. En el gènere Anguina, la femella madura es disposa formant una espiral; són paràsits obligats que formen gal·les sobre les fulles, tiges i inflorescències de diverses monocotiledònies. Ditylenchus destructor és el nematode que podreix la patata.
Els afelènquides (Aphelenchida) són nematodes proveïts d’un estilet, amb nòduls poc o gens desenvolupats, amb el bulb mitjà molt gros i esfèric i el terminal rarament present; les glàndules esofàgiques són lliures (a la cavitat del cos, generalment superposades a l’intestí); la glàndula esofàgica dorsal s’obre al bulb mitjà; l’ovari és prodèlfic, recte, i la bursa és present solament en el gènere Aphelenchus. Viuen lliures en el sòl, o bé com a fitoparàsits (de fulles, tiges i bulbs) o associats a insectes. Destaquen les famílies dels afelènquids i afelencòlids.
Els afelènquids (Aphelenchidae) tenen l’estilet sense nòduls i una banda lateral que conté entre 10 i 12 línies; els mascles tenen bursa amb costelles"., i les femelles tenen la cua curta i cilíndrica. Aphelenchus avenae és freqüent en sòls conreats.
Els afelencòidids (Aphelenchoididae), reuneixen gèneres paràsits de fulles i tiges, i d’insectes. Tenen un estilet amb nòduls reduïts o sense nòduls, l’esòfag sense bulb terminal, i les glàndules lliures formant grans lòbuls; la cua de les femelles és aguda i la bursa, quan n’hi ha, és reduïda a un lòbul. Aphelenchoides, Bursaphelenchus i Rhadinaphelenchus són tres dels gèneres principals.
Els estrongílides (Strongylida), paràsits de vertebrats, entre els quals l’home, es caracteritzen per tenir la faringe cilíndrica, una càpsula bucal quitinosa ben formada; els mascles tenen bursa. Destaca especialment la família dels ancilostomàtids (Ancylostomatidae), que inclou els ancilòstoms (Ancylostoma duodenale), nematodes de quasi 2 cm de llargada que parasiten l’home, en el qual provoquen l’anèmia dels minaires, d’una gran incidència als països tropicals; una altra espècie pròxima, Necator americanus, té importància als continents americà i africà.

Els ascarídides reuneixen nematodes paràsits d’animals, molts dels quals són ben coneguts com a productors de malalties a l’home i als animals domèstics. Pertany en aquest grup l’ascàride del gos i el gat (Toxocara canís), que veiem a la fotografia, i d’altres com l’ascàride de l’home, el del porc i el del cavall.
ACE / Fotostock.
Els ascarídides (Ascaridida) es caracteritzen pel fet de tenir l’obertura bucal proveïda de tres grans llavis. Els mascles tenen dues espícules. Són nematodes grossos (entre 10 i 35 cm, generalment) i reuneixen alguns dels principals paràsits de l’home, com el conegut ascáride de l’home (Ascaris lumbricoides), i nombroses espècies d’interès veterinari, com l’ascàride del cavall (Parascaris equorum), els ascàrides del gos i del gat ( Toxocara canis i T. cati) i l’ascàride del porc (Ascaris suum).
Els oxiúrides (Oxyurida) tenen el cos fusiforme, acabat en una cua punxeguda en el cas de les femelles, i tres llavis a l’obertura bucal. Inclouen els coneguts oxiürs ( Enterobius vermicularis i E. gregorii), paràsits intestinals de l’home, especialment en edat infantil.
Els espirúrides (Spirurida) tenen dos llavis a la boca i els mascles presenten dues espícules desiguals a la bossa caudal. Són paràsits de vertebrats, i inclouen espècies que afecten l’home. És especialment important la família dels filàrids (Filariidae), que inclou els causants de malalties com els tumors de Calabar (Loa loa), l’elefantiasi (Wuchereria bancrofti) i l’oncocercosi o ceguesa dels nius (Onchocerca volvulus), i també la superfamília dels dracunculoïdeus (Dracunculoidea), que inclou l’espècie causant de la filariasi de Medina (Dracunculus medinensis).
Els nematodes marins

Dos exemples de nematodes marins: A dalt, espècie del gènere Prochaetopoma A baix, exemplar del gènere Draconema.
David Masalles
Els nematodes marins són els metazous més abundants en els sediments oceànics, i són representats per un gran nombre d’espècies diferents. Morfològicament tenen alguns trets en comú; per exemple, es distingeixen dels altres grups de nematodes pel fet que presenten unes gònades relativament curtes i simples, una cutícula gruixuda amb setes ben visibles, i una sèrie de glàndules situades bàsicament en posició caudal, que els permeten d’adherir-se a les partícules de sediment. La llargada dels nematodes marins oscil·la normalment entre 1 i 3 mm, però també n’hi ha alguns de mida superior a 1 cm i d’altres de menys de 0,2 mm.
Els nematodes marins són dioics. De desenvolupament directe i bentònic, es reprodueixen per mitjà d’ous. Presenten quatre estadis juvenils diferents abans d’arribar a adults. En algunes espècies vivípares, els ous es desclouen dins l’úter. L’absència de fases planctòniques en el desenvolupament afecta la dispersió de les espècies; no obstant això, per causa de les seves petites dimensions, poden ésser transportades pels corrents marins. Els cicles vitals són curts (normalment duren entre 20 i 30 dies) i la reproducció és contínua. Algunes espècies de mida grossa presenten un sol cicle anual. El temps de generació i la fecunditat depenen fonamentalment de la temperatura.
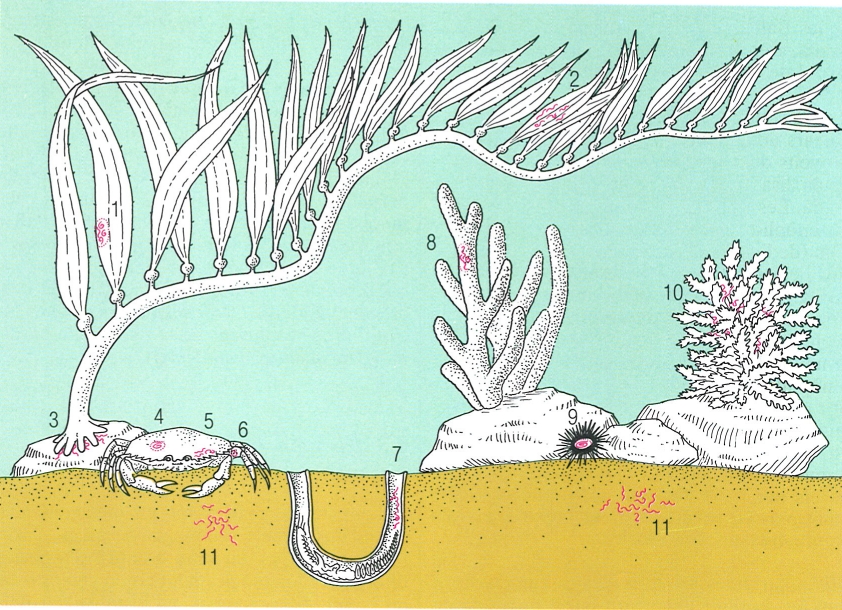
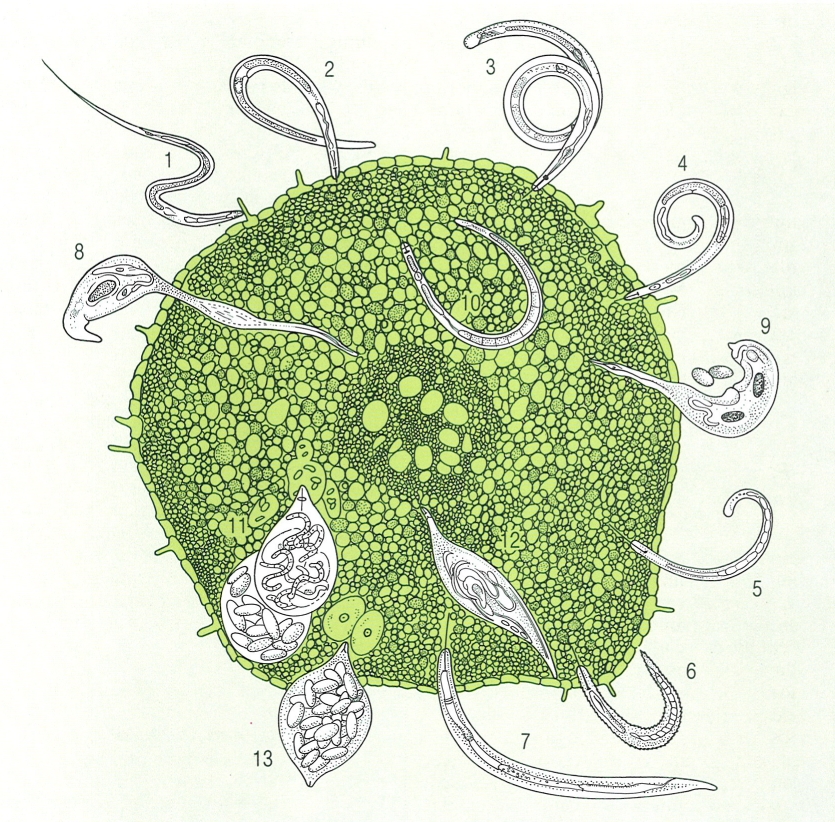
Nematodes micròtrofs presents en diferents hàbitats marins: 1 Halenchus fucicola, que s’alimenta d’algues marines; 2 cromadòrides que viuen en les algues submergides; 3 espècies de Thoracostoma i d’altres gèneres s’instal·len a la base de les algues marines; 4 nematodes paràsits com els espirúrids (Ascarophis, Proleptus, Rhabdochona) utilitzen els crancs marins com a hostes intermediaris; 5 en les cambres respiratòries dels crustacis bentònics s’hi allotgen membres dels gèneres Rhabditis, Diplolaimella, Theristus, Tripylium i Monhystrium; 6 als apèndixs dels mateixos crustacis hi viuen alguns cefalòbids; 7 algunes espècies (com Enoplus communis) viuen dins els tubs dels poliquets; 8 alguns enòplides viuen dins les esponges; 9 hi ha equinomermèl·lids (Echinomermella grayi) paràsits de les garotes; 10 d’altres viuen entre els coralls; 11 el sediment és ocupat per representants dels ordres dels areolàimides, monistèrides, desmodòrides, cromodòrides, enòplides i desmoscolècides.
Biopunt, original de George O. Poinar.
Dins el medi marí, gràcies a la seva capacitat d’adaptació ecofisiològica, han arribat a colonitzar tots els hàbitats bentònics. Són particularment abundants en els fons sedimentaris, i atenyen les densitats més elevades en zones estuarianes (fins 20 × 106 individus per metre quadrat), i les més baixes en platges sorrenques exposades a un fort onatge (0,1 × 106 individus per metre quadrat). La taxa metabòlica és alta, i poden assolir valors elevats de producció. Els que viuen en sediments sorrencs, especialment de gra gros, s’han adaptat al medi intersticial i són capaços de desplaçar-se horitzontalment i verticalment; en aquest ambient, la quantitat d’oxigen que hi ha a l’aigua és el factor limitador de la seva distribució. Els nematodes intersticials generalment són petits o mitjans, allargats, tenen la cutícula gruixuda i llargues i nombroses setes per afavorir el desplaçament. En sediments fins i fangosos, molt més compactes, els nematodes colonitzen únicament la capa superficial; en aquest cas tenen mides corporals més grans i en general menys setes. Els de les zones fitals solen trobar-se en els dipòsits de sediment o detrits acumulats damunt les plantes. En ambients fotòfils poden desenvolupar ocels o pigments fotosensibles, que els permeten de diferenciar la intensitat de la llum i n’afavoreixen el desplaçament. La diversitat específica varia segons l’hàbitat, però en general és alta: a les platges tranquil·les se’n poden trobar fins 100 espècies diferents, mentre que en fons fangosos i comunitats d’algues el seu nombre oscil·la entre 30 i 70; algunes espècies (espècies oportunistes) són capaces d’adaptar-se a zones sotmeses a pressió ambiental (contaminació, entrades d’aigua dolça o de salinitat elevada), i dominar totalment la comunitat de nematodes.
L’aliment potencial dels nematodes marins inclou detrits orgànics, organismes en descomposició, bacteris, diatomees i altres organismes vius. Els que viuen en el mateix medi competeixen mútuament pels recursos alimentaris, si bé la gradació de mides de l’aliment i de la boca permet una bona distribució de l’aliment entre les espècies. Generalment, l’estructura de la cavitat bucal és un bon indicador de l’estratègia alimentària de l’espècie: una cavitat bucal petita o absent indica que digereix només petites partícules o fluids (alimentació molt selectiva), mentre que una cavitat bucal desenvolupada i sense dents indica la capacitat de digerir partícules més grans, i fins i tot diatomees (alimentació menys selectiva); quan la cavitat bucal té petits denticles, és que poden foradar cèl·lules o partícules i absorbir líquids interns o també brostejar superfícies més grans; finalment, la presència de grans mandíbules i dents indica que l’espècie és depredadora i omnívora. La proporció relativa de les diferents estratègies en la comunitat de nematodes depèn de la naturalesa i percentatge de l’aliment disponible, cosa que generalment es relaciona amb el tipus de sediment.
Els nematodes fan part de l’alimentació d’un gran nombre d’animals bentònics marins. Els seus principals consumidors són els sedimentívors, que ingereixen sense seleccionar grans quantitats de sediment, com per exemple els poliquets, els holoturoïdeus i d’altres; alguns crustacis i peixos (especialment els estats larvals de burrets —gòbids— i llisseres —mugílids—) s’alimenten activament de nematodes.
Els estudis fets als Països Catalans se centren en les poblacions de les comunitats fotòfiles de les Balears i de zones concretes del litoral català, i en les comunitats sedimentàries del litoral de Barcelona i del delta de l’Ebre.
En les comunitats d’algues fotòfiles catalanobalears dominen els nematodes de talla gran, proveïts de setes cefàliques curtes, ocels, cutícula llisa i cavitat bucal ben diferenciada i armada. Les famílies més ben representades són els enòplids, els fanodermàtids i els enquelídids (de l’ordre dels enòplides), els cromadòrids i els monopòstids (de l’ordre dels cromadòrides) i els desmodòrids (de l’ordre dels desmodòrides). Aquests nematodes s’adapten perfectament a les condicions que els ofereix el medi algal. Generalment, els més grossos (enòplids) tenen preferència per la part superior de les plantes, que és la menys densa, ja que allà resisteixen més bé l’hidrodinamisme; en les parts més denses de les algues (zona intermèdia, entre la base i les ramificacions) hi ha els nematodes més petits (cromadòrids), mentre que a la base s’hi tornen a situar enòplids grossos.
Els sediments sorrencs deltaics (delta de l’Ebre) presenten espècies de cutícula fortament estriada (anellada, en general), amb setes llargues i abundants, amfidis circulars o espirals, de mida mitjana i proveïts de cavitat bucal ben desenvolupada i armada. Les famílies més abundants són els ciatolàimids i els selaquinemàtids (ordre dels cromadòrides), i els desmodòrids (ordre dels desmodòrides). Les zones de sediments fangosos i llimosos són habitades per espècies de mida més grossa, cutícula llisa o amb fileres de punts i cavitat bucal menys desenvolupada i moltes vegades no armada; hi dominen els comesomàtids (cromadòrides), els desmodòrids (desmodòrides), els xiàlids (monistèrides) i els linomèids (areolàimides). Les zones que reben influència de les aigües dolces tenen unes poblacions menys estructurades, en les quals dominen les famílies ara citades, en una proporció que depèn de les característiques del sediment de cada punt.
El litoral barceloní presenta zones fortament contaminades, on hi ha espècies de nematodes adaptades o molt resistents a la pol·lució; hi dominen els comesomàtids (cromadòrides) i els xiàlids (monistèrides). A les zones menys afectades per la contaminació s’hi troben espècies de ciatolàimids i comesomàtids (cromadòrides) i de linomèids (areolàimides).
Els nematodes dulciaqüícoles i terrestres
És impossible de fer una divisió clara entre els nematodes lliures terrestres i els d’aigua dolça, ja que un gran nombre d’espècies es troben, indistintament, en un o altre medi, si bé cal dir que cada espècie té una freqüència d’aparició més gran en un d’ells, que es considera el seu hàbitat específic. No hi ha espècies exclusives de determinats medis o biòtops, sinó que solament es pot parlar d’espècies que dominen en determinats medis; per tant, més que d’espècies característiques, hom parla de comunitats caracteritzades per la dominància de certes espècies.
A les torberes de l’alta muntanya, constituïdes per esfagnes, es troba una comunitat de nematodes caracteritzada per la dominància constant d’Ironus ignavus, Actinolaimus macrolaimus i Eumonhystera vulgaris, sovint acompanyades de Teratocephalus crassidens, Alaimus filiformis i Rhabditis filiformis, que són muscícoles i humícoles. En estanys i llacs pirinencs d’origen glacial, a aproximadament 2000 m d’altura, es troben dues espècies dominants, Dorylaimus stagnalis i Tobrilus gracilis, sovint acompanyades de diverses espècies de Monhystera, Plectus i Mononchus.
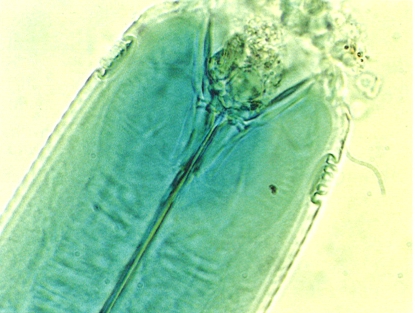
Detall de la regió cefàlica d’una femella del nematode Choanolaimus psammophilus, on s’aprecien esplèndidament els amfidis, en forma de petites fossetes laterals al cap. La fotografia és feta al microscopi òptic. Aquesta espècie, depredadora, ha estat trobada en un salicornar del delta de l’Ebre.
Alejandro Palomo.
En rius i canals, la fauna de nematodes és més pobra; hi dominen Tobrilus gracilis, Dorylaimus stagnalis o Diplogaster rivalis. Un biòtop molt característic és el que es troba en sèquies de rec de la Plana de Castelló, on el corrent és lent, amb abundants detrits vegetals i algues; hi domina absolutament Diplogaster rivalis.
Les comunitats de nematodes lliures terrestres presenten una certa complexitat ecològica. Els biòtops terrestres són caracteritzats pel tipus de sòl, pel recobriment vegetal i per tot un conjunt de factors físics i biòtics. Una comunitat completa és integrada per representants depredadors (principalment monònquids, tripílids i altres grups menors), fitòfags (gairebé exclusivament dorilàimids i monistèrids) i micròtrofs (plèctids i fonamentalment rabdítids). Pel que fa a la distribució general de la nematofauna edàfica, en el primer horitzó del sòl, a la zona superficial, hi ha una presència majoritària d’espècies micròtrofes i depredadores, mentre que en el segon horitzó, més profund, la fracció fitófaga és més ben representada. En relació amb les estacions de l’any, sembla que a l’hivern hi ha una fauna de nematodes més rica que a l’estiu, la qual cosa sembla demostrar la importància del gradient d’humitat en el sòl per al desenvolupament d’aquestes comunitats. Pel que fa a la textura dels sòls, els que tenen un component de llim i argila més elevat, tenen una proporció més alta d’espècies micròtrofes i depredadores, mentre que els sòls de textura sorrenca tenen la fracció fitófaga més ben representada que les altres. Analitzant les dades obtingudes fins a l’actualitat, sembla que hi ha una relació entre les espècies micròtrofes i els medis àcids, de pH baix, i que el medi alcalí és preferit per les fraccions depredadora i fitófaga.
Els nematodes fitoparàsits
Els nematodes fitoparàsits pertanyen fonamentalment als ordres dels tilènquides, els afelènquides i els dorilàimides. Els dorilàimides fitoparàsits són tots ectoparàsits obligats. Pel que fa als tilènquides i els afelènquides, es poden reconèixer dos grans grups: els ectoparàsits, tot el cos o la major part del cos dels quals és el que es troba exposat a les condicions ambientals mentre s’alimenten, i els endoparàsits, tot el cos dels quals és recobert pel teixit de la planta.
A Cicle anguinoide. B Cicle al·lantonematoide. C Cicle iotoncoide
Biopunt, a partir de fonts diverses
Els nematodes fitoparàsits i entomoparàsits poden tenir diferents tipus de cicles biològics. A Cicle anguinoide: típic dels tilènquides paràsits de plantes, els estadis de desenvolupament dels quals són lliures i s’alimenten de cèl·lules de rels o d’algues, o bé d’hifes fúngiques. B Cicle al·lantonematoide: el mascle i la femella, lliures, s’acaben de desenvolupar en el sòl; el mascle mor després de la impregnació de la femella; la femella penetra en l’hemocel de l’insecte, i hi deixa ous i joves; els preadults surten a l’exterior a través de l’oviducte i de l’anus. C Cicle iotoncoide: té una generació parásita i partenogenètica, que viu en l’hemocel d’un insecte, i una generació heterosexual, lliure; la femella acaba de desenvolupar-se al sòl, i per partenogènesi dona lloc a femelles i mascles (C’); les noves femelles, després de la impregnació, penetren en un nou insecte. D Cicle neotilencoide: té una forma heterosexual, parásita d’un insecte, però amb un estadi lliure al sòl (B), i una altra forma heterosexual lliure al sòl, que menja fongs. En el gènere Fergusobia (1 femella, 2 mascle), el cicle té una generació heterosexual dins un insecte productor de gal·les en els eucaliptus; mascles i femelles són lliures dins de la gal·la; la femella impregnada penetra en la cavitat del cos de la larva de l’insecte abans que aquesta surti de l’ou (D); l’altra part del cicle fa vida lliure en l’interior de les gal·les, s’alimenta de les cèl·lules de la paret de la gal·la i es reprodueix per partenogènesi (D’). En el gènere Parasitylenchus hi ha dos cicles heterosexuals, l’un amb un estadi lliure al sòl (B i l’altre que es completa dins l’insecte, de manera que dins d’aquest, trobem tres tipus d’adults (E): la femella heterosexual primària amb estadi lliure al sòl, la femella heterosexual secundària amb el cicle complet dins la cavitat de l’insecte, i els mascles del cicle heterosexual secundari.
Els ectoparàsits
Els dorilàimides ectoparàsits
Els dorilàimides ectoparàsits pertanyen únicament a dues famílies, els longidòrids i els tricodòrids. Els gèneres Trichodorus, Paratrichodorus, Longidorus i Xiphinema tenen una gran importància com a transmissors de virus de plantes. Pel que fa als dorilàimides transmissors de virus en diferents conreus, cal dir que el seu estudi és recent. Fins a l’actualitat hom ha trobat a la península Ibèrica diverses espècies del gènere Xiphinema. X. brevicolle s’ha trobat en conreus de cítrics de la costa castellonenca. X mediterraneum és la més freqüent i cosmopolita de totes les de la conca mediterrània, i hom tendeix a relacionarla amb la malaltia de l’avellaner anomenada «de sant Nicolai», encara que no transmet el GFV («Grapewine Fanleaf Virus»). X. diversicaudatum és vectora de diferents tipus de virus específics del presseguer, el cirerer, la prunera i la figuera, i s’ha estudiat a l’oest i el nord-oest peninsular. X.index, la més coneguda per la seva facultat de transmissora de virus GFV a la vinya, té una distribució mundial meridional, sempre associada a aquest conreu; als Països Catalans s’ha trobat sovint afectant vinyes al Penedès, bé que, pel que sembla, va essent desplaçada per X. italiae, també vectora del virus de la vinya. X. sahelense es troba en zones càlides i seques de l’Andalusia occidental. X. vuittenezi és una espècie més aviat de l’interior, estesa per l’Europa central, que ha estat trobada en conreus de pereres i albercoquers al centre i sud peninsular. X. turcicum (coneguda solament a la península Ibèrica), ha estat trobada en vinyes i cítrics de la costa llevantina i andalusa, i en fruiterars del nord-oest i centre peninsular. Del gènere Longidorus s’han trobat quatre espècies als Països Catalans: L. attenuatus, L. caespiticola, i L. profundorum, a les comarques valencianes, i L. macrosoma, a les lleidatanes i barcelonines.
Els tilènquides i els afelènquides ectoparàsits
Esquema idealitzat dels tipus de fltoparasitisme que podem trobar entre els nematodes tilènquides que s’alimenten de rels de plantes. Ectoparàsits: 1 Cephalenchus, 2 Tylenchorhynchus, 3 Rotylenchus, 4 Helicotylenchus, 5 Rotylenchulus, 6 Macroposthonia, 7 Hemicycliophora. Semiendoparàsits: 8 Tylenchulus, 9 Rotylenchulus. Endoparàsits sedentaris: 10 Pratylenchus. Endoparàsits sèssils: 11 Meloidogyne, 12 Nacobus, 13 Heterodera.
Biopunt, a partir de fonts diverses
La majoria dels tilènquides són ectoparàsits de les rels, mentre que els afelènquides i alguns anguínids ho són de les parts aèries de les plantes. Hom distingeix diversos tipus d’estratègia. Es parla d’alimentació passiva per a referir-se als que tenen l’estilet molt curt i el bulb esofàgic mitjà poc desenvolupat o absent; sembla que mantenen baixa la pressió corporal per mitjà de la vàlvula rectal, amb la qual cosa la pressió de les cèl·lules i hifes de l’hoste és suficient per a fer entrar l’aliment al cos del nematode; Aphelenchus avenae, trobat en fruiterars, vinyes i camps de cítrics de Catalunya, és un nematode d’aquest tipus; no provoca problemes a les plantes.
Entre els ectoparàsits migradors hi ha els tilènquides i els afelènquides que viuen a la superfície de les rels, proveïts d’un estilet curt, i que es mouen ràpidament d’un lloc a l’altre, com Aglenchus agricola, freqüent en fruiterars, hortes (Segrià, Baix Llobregat) i boscos (Osona, Selva), i A. bryophilus i Ecphyadophoroides tenuis, tots dos associats a cítrics (País Valencià). D’altres tenen un estilet llarg, que claven profundament als teixits de la rel, i poden restar més temps al mateix lloc, com, per exemple, Rotylenchus robustus (prats i fruiterars de l’Alta Ribagorça i el Segrià), R. goodeyi (cítrics del País Valencià), o el conegut Helicotylenchus dihystera, molt freqüent, cosmopolita i polífag, trobat freqüentment en fruiterars i hortes de diverses comarques del Principat i del País Valencià; H. digonicus, Paratrofurus loofi, Tylenchorhynchus huesingi, T. goffarti i T. clarus i Merlinius brevidens són també espècies freqüents en conreus de diverses comarques catalanes. Els criconemàtids, són paràsits migradors àmpliament distribuïts. Hemicycliophora thienemanni i H. conida són freqüents en marges de rierols i torrenteres humides, del Segrià i el País Valencià, bé que de vegades es troben en presseguers i cítrics; Criconema annulifer i Ogma rhombosquamata es troben en alzinars subhumits del Principat; Crossonema multisquamata es troba en cítrics o fons de valls humides dels Prepirineus; Hemicriconemoides cocophilus i H. gaddi tenen una distribució meridional i es troben en cítrics; M. xenoplax és freqüent en vinyes i fruiters de pinyol, sovint associat a bacteris com Pseudomonas syringae; M. curvata és freqüent a la conca mediterrània i ataca el clavell, al qual produeix clorosi i retards en el creixement; M. antipolitana i M. sphaerocephala són freqüents en prats secs; i M. rustica ho és en els prats humits dels Pirineus.
Els nematodes ectoparàsits sedentaris són els que s’han adaptat més bé al parasitisme. El seu cicle inclou una fase juvenil d’ectoparàsit migrador i una altra d’ectoparàsit sedentari; el nematode deixa la meitat del cos dins la planta, on indueix la formació de cèl·lules nodridores que li proporcionaran l’aliment, i s’allunya així de les condicions externes. L’espècie més representativa d’aquest grup és Tylenchulus semipenetrans, freqüent a totes les àrees de conreu de cítrics. Anomenat nematode dels cítrics (bé que també ataca la vinya i l’olivera), només les femelles i els joves són paràsits, perquè el mascle madura al sòl, sense menjar; la larva preadulta és ectoparàsita migradora fins que arriba al quart estadi, i llavors canvia els seus hàbits i passa a ésser ectoparàsita sèssil.
Entre els ectoparàsits de fulles i tiges hi ha fonamentalment els afelènquides, també capaços de moure’s amb facilitat pels espais intercel·lulars, a les tiges, els bulbs i les fulles. Poden sobreviure diversos mesos en condicions de falta d’aigua, i també es poden cultivar amb fongs. Anguina tritici parasita inflorescències de gramínies, i Ditylenchus dipsaci parasita alls, cebes i bulbs de plantes ornamentals.
Els endoparàsits
Els nematodes endoparàsits es desenvolupen en l’interior de la planta, i generalment són adaptats a explotar un tipus determinat de teixit; generalment, els que ataquen fulles i tiges no ataquen rels.
Entre els endoparàsits de tiges i fulles hom troba tant tilènquides (anguínids i nototilènquids) com afelènquides, que alternen entre endoparasitisme i ectoparasitisme. Algunes espècies d’afelènquides són transportades per insectes. Aphelenchoides fragariae és paràsit de la maduixera, els teixits de la qual envaeix, si bé s’alimenta, a la superfície de les fulles, de les gemmes encara per obrir; A. ritaemabosi actua com a endoparàsit de la maduixera i com a ectoparàsit de les fulles del crisantem. Bursaphelenchus xylophilus és el nematode de la fusta del pi, capaç de matar ràpidament els arbres, que infecta tant per les fulles i les tiges com per les arrels; és transportat, en forma de larva dauer, entre les antenes i els èlitres d’un escarabat cerambícid (Monochamus alternatus); un sol escarabat pot transportar més de 15 000 larves. Rhadinaphelenchus cocophilus és el causant de la malaltia anomenada «anell vermell» del cocoter i la palma, i el seu cicle biològic inclou l’associació amb l’insecte Rhynchophorus palmatum; la larva del tercer estadi és la forma infectant i és la que es troba sobretot en l’anell roig; la larva de l’insecte es menja el tronc i durant el creixement ingereix una gran quantitat de larves del nematode, que passen al seu hemocel i s’hi estan fins que l’insecte es fa adult; els nematodes surten de l’insecte amb els ous que aquest diposita sobre la palma.
Entre els endoparàsits de rels hom distingeix dos grups, els migradors i els sedentaris. Els migradors poden desenvolupar tot el cicle complet a l’interior de la planta o bé tenir alguna fase lliure. Les espècies de Pratylenchus interactuen amb fongs, i llavors la seva acció patògena és molt superior. P. brachyurus, P. coffeae, P. penetrans, P. scribneri, P. vulnus i P. zeae, s’associen als cítrics (País Valencià); P. thornei i P. neglectus, als fruiters de pinyol, i Zygotylenchus guevarai, a la vinya (Penedès), els fruiters (Segrià) i els cítrics (País Valencià); Pratylenchoides crenicauda (Segrià) és molt menys comuna. Els endoparàsits sedentaris passen per la fase d’ou i de jove de segon estadi (anomenat larva L2) , i el mascle fa vida lliure al sòl. En els hetero dèrids, la larva L2 penetra la rel de l’hoste i hi indueix la formació d’unes cèl·lules grosses, de les quals es nodreix. El creixement de la femella fa que es trenquin els teixits de la planta i la regió posterior de l’animal queda exposada a l’exterior, mentre que la regió anterior, amb el cap, hi resta clavada; el mascle la fecunda i poc temps després la femella deixa la planta i cau a terra en forma de quist, que conté els ous en diferent estat de desenvolupament. Hom ha comprovat que hi ha diferents adaptacions de la planta per impedir la penetració de l’endoparàsit, i de l’endoparàsit per vèncer aquesta resistència, i s’han descrit diferents biotips d’ Heterodera, anomenats patotips, en diferents graus de virulència. Les espècies d’ Heterodera més àmpliament distribuïdes al nostre país són H. avenae (cereals) i H. rostochiensis (patata). Les espècies de Meloidogyne més esteses al Principat, M. incognita, M. javanica, M. hapla i M. arenaria, parasiten fruiters, plantes d’horta i plantes hornamentals; són molt esteses al Principat.
Els nematodes entomopatògens

Crisàlide de papallona parasitada per nematodes entomopatògens, els anomenats vulgarment «cucs grisos».
Fernando García del Pino.
D’entre els nematodes zooparàsits, mereixen una atenció especial els que parasiten insectes, ja que poden fer un paper molt important en el control biològic de les plagues. Pot tractar-se de paràsits facultatius o de paràsits obligats, que causen la debilitació, l’esterilitat o la mort d’un gran nombre d’insectes. Les qualitats més apreciades d’aquests nematodes, de cara al control de plagues, són la seva virulència i la rapidesa amb què maten l’insecte parasitat. Només algunes espècies de l’ordre dels mermítides i les famílies d’esteinernemàtids i heterorabdítids de l’ordre dels rabdítides presenten aquestes qualitats.
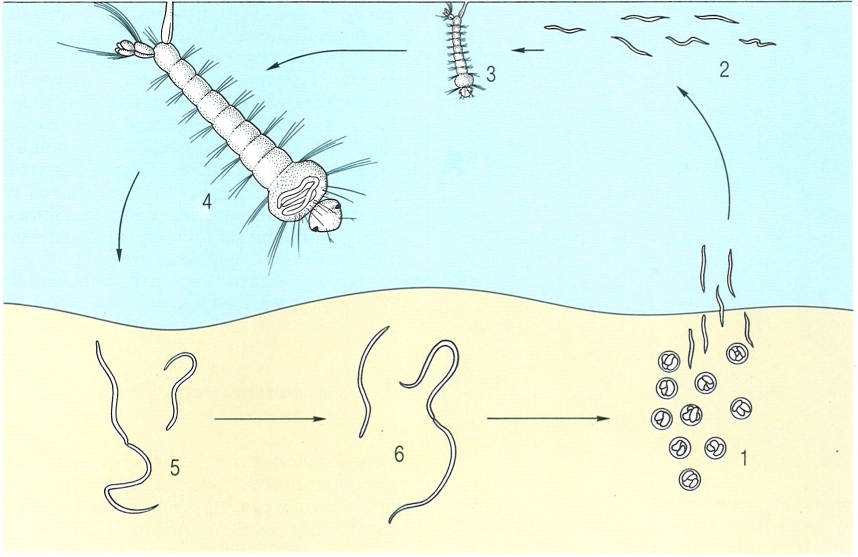
Cicle biològic del mermítid entomopatogen Romanomermis culicivorax. Hom hi ha indicat: 1 ous, 2 preparàsits, 3 fase de paràsit, 4 emergència i mort de l’hoste, 5 postparàsits, 6 adults.
Biopunt, a partir de fonts diverses
Els mermítides paràsits d’insectes actuen d’una manera anàloga a la dels parasitoides interns, ja que obtenen els nutrients dels hostes, que maten quan les formes juvenils completen el seu desenvolupament parasític i surten de l’hemocel de l’insecte. Apareixen tant en hàbitats terrestres com aquàtics, i utilitzen un ampli ventall d’insectes com a hostes. No obstant això, els que són més importants com a controladors biològics de plagues són els aquàtics, concretament els del gènere Romanomermis, que parasiten larves de mosquit. En el seu cicle biològic, els ous comencen a desenvolupar-se així que són deixats a l’exterior, i quan es desclouen, les formes juvenils preparàsites penetren dins l’insecte hoste a través de la cutícula. Alguns mermítids a penes es desenvolupen mentre l’insecte es troba en fase larval; així que aquest arriba a l’estat adult, els nematodes migren cap al seu abdomen on es desenvolupen considerablement, i moltes vegades li causen l’esterilitat i, posteriorment, la mort, que s’esdevé en el moment en què els nematodes surten de l’insecte adult. Altres espècies segueixen una estratègia diferent, com passa amb les del gènere Romanomermis, que es desenvolupen ràpidament durant la fase larval de l’insecte, amb la qual cosa no el deixen atènyer l’estadi de nimfa i el maten en el moment de sortir de l’insecte; llavors els nematodes postparàsits passen al medi (aigua o sòl), on acaben el seu desenvolupament i atenyen l’estadi adult, finalment alliberen els ous i s’inicia un nou cicle.
Els rabdítides entomopatògens es caracteritzen perquè mantenen una relació de mutualisme amb un bacteri (Xenorhabdus) que transporten en el tub digestiu; aquest bacteri és el que mata ràpidament l’insecte parasitat, i proporciona l’aliment necessari al nematode. Aquests nematodes es troben d’una manera natural en el sòl i en hàbitats molt humits, i per això generalment utilitzen com a hostes insectes també propis d’aquests hàbitats (escletxes, sòl, escorces d’arbre, etc.).
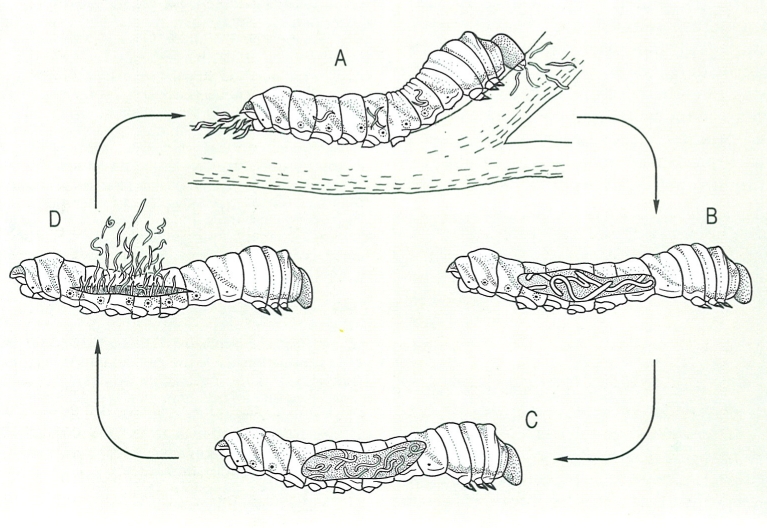
Cicle biològic dels nematodes entomopatògens de les famílies dels esteinermàtids i els heterorabdítids.
Biopunt, a partir de fonts diverses
Dins la família dels esteinernemàtids (Steinernematidae) s’inclou actualment un sol gènere, Steinernema, i set espècies: S. affinis, S. glaseri, S. carpocapsae, S. feltiae, S. intermedia, S. rara i S. anomali. En aquests nematodes, la infecció de l’insecte solament es pot iniciar en el tercer estadi larval infectiu (que correspon a la larva dauer dels nematodes lliures), en el qual l’animal té la capacitat de trobar l’hoste i penetrar-lo activament a través de les obertures naturals (boca, anus o espiracles) fins a atènyer l’hemocel. Així que hi arriben, els nematodes alliberen, a través del seu anus, els bacteris simbionts que duien a l’intestí, els quals es multipliquen i s’estenen per tot el cos de l’insecte fins a matar-lo; així, estableixen les condicions favorables perquè els nematodes esdevinguin adults. Llavors es produeix l’aparellament dels adults, per mitjà de la còpula. Les femelles comencen a alliberar els ous dins l’hemocel de l’insecte 24 hores després de la còpula; com que el desenvolupament dels ous és més ràpid que el procés de la posta, l’eclosió de molts ous es produeix dins l’úter matern, cosa que fa que la mare mori (endotòquia matricida). El desenvolupament dels petits nematodes acabats de sortir de l’ou pot seguir dos camins diferents, segons quina sigui la quantitat d’aliment disponible: si n’hi ha prou, completen el seu desenvolupament i donen lloc a la segona generació d’adults, que són de dimensions corporals més petites que els de la primera, i produeixen un nombre d’ous inferior; per contra, si no hi ha prou aliment, passen directament a formar estadis infectius que són els que surten del cadàver de l’insecte cap al medi extern (generalment el sòl), cercant un nou hoste; durant aquest temps, aquestes formes infectives no s’alimenten, i són resistents a condicions ambientals adverses i a certs productes químics.
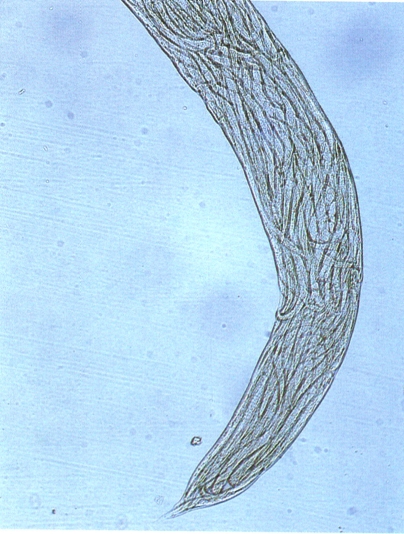
Detall del final del cos d’un heterorabdítid (Heterorhabditis) entomopatogen. La importància d’aquests nematodes ha crescut modernament, quan hom ha iniciat la recerca en el camp del control biològic de les plagues.
Fernando García del Pino.
La família dels heterorabdítids (Heterorhabditidae) inclou un únic gènere, amb quatre espècies: Heterorhabditis heliothidis, H. bacteriophora, H. hopta i H. hambetoni. Totes tenen un bacteri associat (Xenorhabdus luminescens), encarregat de matar l’insecte parasitat. El seu cicle biològic és heterogònic. La primera generació d’adults és composta únicament de femelles hermafrodites, que poden créixer fins a atènyer dimensions considerables (més de 6 mm de llargada); com en la família anterior, els ous són alliberats a l’hemolimfa de l’insecte, però molts eclosionen encara dins la femella que els produeix. Els nematodes que surten d’aquesta primera posta donen lloc a una segona generació, de mascles i femelles, els primers més petits. D’aquesta segona generació d’adults sorgiran les formes infectives, que sortiran del cadàver de l’insecte per a cercar un nou hoste.
El gran interès que han despertat els esteinernemàtids i els heterorabdítids en el camp del control biològic de plagues d’insectes es deu al fet que reuneixen una combinació de característiques intermèdies entre els depredadors o paràsits, els patògens i els insecticides químics. D’una banda tenen la mobilitat dels depredadors a l’hora de trobar el seu hoste i, d’altra banda, poden ésser produïts massivament, emmagatzemats i aplicats d’una manera semblant als patògens microbians. D’altres característiques, com el seu ampli espectre d’hostes i la seva virulència, recorden els insecticides químics, la qual cosa, a més del fet que no produeixen resistència en els insectes atacats i que són innocus per a l’ambient, fa que tinguin una bona acceptació per a ésser aplicats als programes de lluita integrada contra les plagues d’insectes. Els estudis fets fins ara demostren que els millors resultats s’han aconseguit amb la utilització del nematode Heterorhabditis heliothidis contra les plagues d’insectes que es desenvolupen al sòl, com el coleòpter Otiorynchus sulcatus. Igualment, s’han obtingut bons resultats amb la utilització del nematode Steinernema feltiae en el control del corc de la fusta Zeuzera pyrina, les larves del qual es desenvolupen en l’interior de galeries fetes als troncs dels arbres fruiters. Resultats similars (entre un 90 i un 99% de mortalitat de les larves) s’han obtingut en proves de camp fetes amb el nematode S. bibionis contra el sèssid Synanthedon tipuliformis, que ataca els grosellers.
Els nematodes zooparàsits
Principals tipus de cicle biològic en els nematodes zooparàsits. Cicles monoxens: 1 Trichostrongylus, 2 Ancylostoma i Necator. 3 Ascaris lumbricoides, 4 Syngamus (en el cicle del qual hi ha implicat un hoste paratènic facultatiu), 5 Oswaldocruzia, 6 Strongyloides (cicle heteroxè) i 7 Stephanurus. Cicles heteroxens. sense estadi larval lliure (excepte l’espècie 6): 8 Porrocaecum, 9 Toxocara (amb migració larval transplacentària), 10 Diplotriaenia, 11 Filària, 12 Onchocerca, 13 Gnathostoma spinigerum. Cicles heteroxens amb estadis larvals lliures: 14 Habronema muscae, 15 Protostrongylus, 16 Angiostrongylus (amb un hoste paratènic facultatiu), 17 Spiroxis contorta (amb un hoste paratènic facultatiu), 18 cuc de foc o de Medina (Dracunculus medinensis). A adult. O ou. L1 primer estadi de desenvolupament. L2 segon estadi de desenvolupament, L3 tercer estadi de desenvolupament. En moral s’indiquen els hostes intermediaris (P paratènics) i en gris els estadis encistats.
Biopunt, a partir Baer, 1971.
En els nematodes zooparàsits hom troba una gran complexitat de cicles vitals, des dels més simples, monoxens, fins a cicles heteroxens, amb un o més hostes intermediaris. Hom veurà, a continuació, un resum de les principals etapes dels seus cicles biològics, prenent d’exemple principalment els paràsits de l’home i els animals domèstics.
Els cicles vitals en els secernentis
Cicles directes amb estadis lliures en el medi extern
Entre les espècies que penetren en l’hoste per via cutània, destaca el cas de Strongyloides stercoralis (rabdítides), un paràsit de l’intestí humà, en el cicle del qual s’alternen generacions lliures i generacions paràsites. La forma parásita és feta solament de femelles partenogenètiques d’uns 2 o 3 mm de llargada, que viuen en la mucosa duodenal i alliberen els ous a l’intestí de l’hoste, dels quals surten larves rabditoides (de primer estadi) que surten a l’exterior amb els excrements de l’hoste. Aquestes larves, de menys de 300 µm de llargada, viuen al sòl humit i es nodreixen de detrits. Passen per quatre mudes successives i donen lloc a adults sexuats, els quals s’acoblen i engendren la segona generació. La femella lliure posa els ous, i d’aquests surten novament larves rabditoides, que, després de dues mudes, passen a ésser larves estrongiloides infestants (larves del tercer estadi). Aquestes són les que travessen la pell de l’home i passen a la circulació, per on arriben fins als pulmons, la tràquea, l’esòfag i finalment el duodè, on es transformen en femelles partenogenètiques. Pot haver-hi autoinfestació, en el qual cas les femelles partenogenètiques donen lloc directament a larves estrongiloides infestants, que no surten de l’hoste a l’exterior.
Entre els paràsits que poden penetrar en l’hoste per via cutània i per via bucal destaca el paràsit intestinal de l’home Ancylostoma duodenale (estrongílides), que viu fixat per la càpsula bucal a la mucosa del duodè i el jejú, i que s’alimenta de sang. La femella produeix ous amb solament 4 blastòmers, que continuen la segmentació al medi extern i alliberen una larva rabditoide (del primer estadi), que passa a estrongiloide (del segon estadi) i, finalment, a estrongiloide embeinada (del tercer estadi), molt resistent: arriba a viure fins deu mesos al sòl, i fins divuit mesos en l’aigua; aquesta és la que penetra per via cutània o bucal i migra fins als pulmons, la tràquea, l’esòfag i el duodè; passa dues mudes i es transforma en adult.
Hamonchus contortus, paràsit d’un remugant, sempre es transmet per via bucal. La larva del primer estadi que surt de l’ou, rabditoide, duu un aparell valvular esofàgic de tipus rabditiforme, i dona lloc a la larva del segon estadi; totes dues es nodreixen activament del medi extern. La larva del tercer estadi, estrongiloide i desproveïda d’aparell valvular, inclosa dins la muda de la larva del segon estadi, no es nodreix sinó que viu de reserves, i és la que penetra en l’hoste definitiu.
Els cosmocèrides tenen el cicle més senzill, possiblement més primitiu que el dels ascarídinis, però hom pensa que les dues primeres mudes (larves que no s’alimenten) són una primera etapa cap a les mudes de l’interior de l’ou o fins i tot cap als cicles amb hoste d’espera. El cicle d’Aplectana courdurieri recorda el dels tricostrongílides. Els dos primers estadis larvals són lliures i viuen a l’aigua a partir de les reserves vitel·lines; la forma infestant penetra en els amfibis solament per via oral. Hi ha el cas particular de Cosmocercoides dukae, que viu entre la conquilla i el mantell dels gasteròpodes terrestres, i que té la femella ovovivípara. El primer estadi larval (amb esòfag de tipus rabditoide) muda dues vegades i es converteix en una larva infectant (d’esòfag estrongiloide) que resta inclosa dins les dues mudes precedents; la primera muda, i de vegades la segona, té lloc dins l’úter matern. Tots els estadis evolutius poden trobar-se en el mol·lusc, i el mucus que aquest elimina conté tant adults com estadis larvals. La infestació és directa: les larves penetren entre la conquilla i el mantell o pel porus respiratori.
Cicle directe sense estadis lliures
Hom esmentarà els casos en què les mudes es fan a l’interior de l’ou. Per exemple, en Ostertagia marshalli, la larva muda una vegada en l’interior de l’ou i eclosiona al final del segon estadi; finalment s’enquista dos o tres dies després, sense haver-se alimentat. En Nematodirus filicollis, paràsit de l’ovella, la larva no eclosiona fins al tercer estadi enquistat; l’intestí de la larva infestant és format per una massa de 8 cèl·lules grans i és incapaç d’efectuar digestió. En el cas de l’oxiúrid Enterobius vermicularis, paràsit de l’home, el mascle i la femella viuen en la regió cega de l’intestí. Les femelles fecundades migren fins al recte i es fixen als marges de l’anus, on alliberen milers d’ous embrionats i, en descloure’s, es converteixen directament en formes infestants, ja que contenen larves del tercer estadi. Quan aquests ous són ingcrits, alliberen dins l’estómac de l’hoste una larva que es converteix en adult en l’il li terminal en tres o quatre setmanes. Un cas particular d’eclosió dins l’úter de la marc és el d’Ollulanus tricuspis, tricostrongílid paràsit del gat; els ous eclosionen dins l’úter matern i, encara sense sortir-ne, passen almenys una muda; les larves del segon i tercer estadis, que es formen dins l’estómac del gat, surten a l’exterior amb el vòmit, de manera que no continuen el seu desenvolupament si no són ingerides per un altre gat. Un altre cas semblant és el de Tachygometria vivípara, oxiür paràsit d’Uromastix, en el qual hi ha coexistència de dos tipus de femelles, que viuen conjuntament en el cec d’aquest hoste: una forma ovípara, que fa ous grossos de closca gruixuda, i una forma vivípara, a la qual els ous arribaran dins l’úter matern fins al tercer estadi, que sortirà per a convertir-se en adult dins el cec intestinal de l’hoste, sense passar al medi extern. El cicle dels cosmocèrids de la família dels atràctids és comparable al d’aquesta darrera espècie; sembla que els atràctids tenen la propietat de reproduir-se sense passar pel medi extern, i les femelles donen larves, aparentment en el tercer estadi, que es transformen en el quart estadi i, finalment, en adultes, sense sortir a l’exterior.
Hi ha casos en què la infestació és duta a terme per les larves del segon estadi, amb la intervenció facultativa d’un hoste de transport. L’espècie Heterakis gallinarum, paràsita de gal·liformes, té, com totes les del seu gènere, un bulb esofàgic que recorda el dels oxiúrides, però amb la bossa caudal ben diferenciada. El seu cicle comprèn una fase en el medi extern que va des de l’ou fins a l’estadi infestant (de dotze a quinze dies), que és el segon, com en els oxiúrides. La larva infestant resta dins l’ou. Sovint diverses espècies de eues de terra poden intervenir com a hostes facultatius de transport, com en una primera temptativa vers l’heteroxènia.
Cicles heteroxens amb transmissió per depredació
Cal distingir un primer grup de paràsits, en els quals la infestació de l’hoste vertebrat intermediari es fa en la fase de larva del segon estadi. En el cas d’Ascaris devosi, paràsit de la fura, la primera muda té lloc al voltant del dotzè dia, dins l’ou; quan un ratolí ingereix l’ou, la larva del segon estadi surt de l’ou, travessa la paret del tub digestiu i arriba fins als músculs i les vísceres, on passa a larva del tercer estadi. La fura s’infesta quan es menja el ratolí. Les dues darreres mudes s’efectuen dins l’intestí. Un cas particular és el d’Ascaris lumbricoides, una espècie en la qual el cicle directe deriva d’un cicle heteroxè. És un paràsit de l’home, cosmopolita i molt freqüent (es parla de 1200 milions de persones infectades al món). En aquesta espècie, els ous, alliberats dins l’intestí humà, són molt resistents i contenen una única cèl·lula. Quan surten a l’exterior, s’embrionen i al seu interior es forma la primera larva, la qual passa la primera muda i es converteix en larva del segon estadi, que és, doncs, la larva infectant. Quan l’ou és ingerit per l’home, es desclou dins l’estómac i la larva travessa la paret i se’n va fins al fetge i els pulmons; la segona muda s’efectua als alvèols pulmonars i, posteriorment, les larves del tercer estadi pugen fins a la tràquea, arriben a la faringe i són deglutides, de manera que cauen al tub digestiu, on fan les dues darreres mudes. L’home fa, així, alhora d’hoste intermediari (fins que la larva ateny el pulmó i passa la segona muda) i d’hoste definitiu. La infestadó de l’hoste intermediari en forma de larva del segon estadi també es produeix en els espirúrides més simples, com els fiselopteroides, en els quals el cicle es fa en el medi aquàtic. Dins la família dels gnatostòmids, aquest tipus d’infestació s’ha observat en els gèneres Spiroxys (una dotzena d’espècies paràsites de serps i tortugues aquàtics i batracis) i Gnathostoma. El cicle de Spiroxys contortus s’assembla al dels ascàrides; l’ou sembriona en el medi extern i és el segon estadi larval el que surt de l’ou i infesta el copèpode hoste intermediari. El cicle de Gnathostoma spinigerum s’assembla al dels ascàrides de tres hostes: la primera muda té lloc dins l’ou, el segon estadi infesta un copèpode i el tercer no es forma si no és dins un peix, que fa de segon hoste intermediari obligatori; la biologia d’aquesta espècie recorda molt la dels ascàrides marins, excepte pel fet que la larva del quart estadi segueix una migració dins el gat (hoste definitiu).
En un segon grup hom comentarà els paràsits que adquireixen l’heteroxènia en el primer estadi larval, cosa que es troba paral·lelament en diferents línies evolutives de nematodes, dins els ordres dels ascarídides, els estrongílides i els espirúrides.
En els ascarírides del grup dels cosmocercoïdeus hi ha una heteroxènia encara mal definida, amb infestació facultativa d’un oligoquet o un vertebrat: Megalobatrachonema compansae (família dels katlànids), paràsit de tritons, té larves que poden subsistir uns mesos dins oligoquets, però no creixen realment si no arriben a entrar dins el cos dels cap-grossos o dels tritonts adults, els quals s’infecten en ingerir els oligoquets, els cap-grossos o, directament, les larves.
En els estrongílides metastrongílids, l’hoste intermediari és un oligoquet o un mol·lusc. En el cicle de Metastrongylus apri, paràsit de l’intestí del porc, els ous madurs són eliminats amb les femtes i eclosionen solament si són ingerits per un cuc de terra (oligoquet); dins el cuc, les larves del primer estadi surten de l’ou, passen al sistema circulatori i arriben als òrgans propulsors; allà passen dues mudes, però la larva del tercer estadi manté la cutícula de la del segon estadi. El porc s’infecta ingerint els cucs de terra i llavors les larves migren pels seus vasos limfàtics fins als pulmons, on les femelles comencen a deixar ous al cap de 24 dies d’haver entrat en el porc. En el gènere Skrjabingylus els adults són paràsits de l’interior del nas de la fura; les larves del primer, segon i tercer estadis es desenvolupen en llimacs i cargols, els quals s’infesten per via digestiva o per larves que els travessen el tegument. Els cicles de Metastrongylus i Skrjabingylus es desenvolupen dins l’hoste intermediari (cucs de terra, mol·luscs), que viu al sòl.

Extrem caudal d’un mascle del nematode paràsit Gongylonema, per a mostrar les formacions auxiliars de la còpula, en aquest cas, espícules desiguals, amb la més llarga evaginada.
Santiago Mas-Coma.
En els ascarídides i els espirúrides heteroxens l’hoste intermediari és un artròpode, generalment un insecte. En aquest cas s’observa que el punt de localització del paràsit dins l’hoste definitiu (vertebrat) té relació amb les migracions larvals i el lloc que ocupaven les larves dins l’hoste intermediari. L’heteroxènia s’assoleix al final de la línia evolutiva en els ascarídides (en els subuluroïdeus), mentre que es converteix en fonamental en els espirúrides. Cal distingir tres grups diferents de cicles. En el primer grup hom troba espècies africanes, com el subuluroïdeu (ascarídides) Maupasina weissi, paràsit del cec intestinal d’un petit insectívor africà (un macroscèlid anomenat rata de trompa); i Subulura otolicni, del mateix grup, paràsit del cec intestinal de lemúrids africans (Galago senegalensis i G. demidovii), i espècies del nostre país, com Seuratum cadarachense (seuratoïdeu), paràsit intestinal de la rata cellarda, que presenta una larva del primer estadi dins l’ou, directament infectant, però que es manté uns quants dies dins l’intestí mitjà de l’insecte abans de travessar la paret del tub digestiu. En el segon grup hom inclou espirúrides com Pterygodermatites affinis (rictulàrid), que fa ous embrionats (larva del quart estadi), directament infestants; en aquest cas la larva eclosiona en l’insecte i s’encapsula en el seu intestí posterior; com Abbreviata caucasica (fisaloptèrid), paràsit de primats, que accidentalment pot parasitar l’home; i Ascarops strongylina, un paràsit cosmopolita molt freqüent en l’estómac i l’intestí prim del porc domèstic, la larva del qual infecta coleòpters copròfags. En el tercer grup s’inclouen espècies d’espirúrides com Mastophorus muris, paràsit de l’estómac de les rates comuna i grisa (Rattus rattus i R. norvegicus), els ous embrionats de la qual són directament infestants per a l’hoste intermediari (un blàtid, un coleòpter, un ortòpter, un sifonàpter, un dípter); la larva del primer estadi s’encapsula dins l’insecte; i les espècies del gènere Gongylonema, paràsites d’ocells i mamífers, que viuen incrustades en la seva mucosa estomacal, esofàgica i lingual, i a les quals la larva arriba a atènyer el tercer estadi dins de l’insecte.
Un cas particular és el de la filària ovípara Diplotriaena sokolowi, paràsit dels sacs aeris del blauet, a l’Àfrica. Les filàries ovípares són de bidogia primitiva, i els adults són paràsits dels sacs aeris de rèptils i ocells, i recorden els metastrongílides, per la seva biologia. Les larves del primer i tercer estadi evolucionen en el teixit adipós d’un insecte copròfag; la larva del tercer estadi, que és la infestant, travessa la paret intestinal de l’ocell, passa al circulatori i se situa primer als pulmons i després als sacs aeris, on es converteix en adult; els ous són regurgitats per l’ocell a través de la tràquea fins a la faringe i així arriben al tub digestiu.
Cicles heteroxens amb transmissió intermèdia
El pas d’una transmissió passiva a una transmissió activa augmenta considerablement l’eficàcia del cicle. Aquest pas es deu essencialment a una modificació del comportament dels hostes intermediaris, que en lloc d’ésser l’organisme consumit passen a ésser els consumidors. Cicles d’aquesta mena s’han observat en dos grups d’espirúrides: els habronemàtids i els telàzids.
Dins els habronemàtids hi ha, per exemple, Draschia megastoma, que viu dins la paret de l’estómac d’èquids, allotjat en la mucosa, on provoca la formació d’uns nòduls especials, proveïts d’un fi canal central a través del qual les femelles fecundades del paràsit ponen els ous. Aquests deixen anar a l’intestí unes larves del primer estadi que duen un estilet quitinós (larva aciculada), i surten a l’exterior amb les femtes del cavall. La continuació del cicle requereix la participació d’un insecte dípter, la mosca domèstica, la larva de la qual s’infecta ingerint, per depredació, la larva del primer estadi del nematode. Un cop dins la larva de mosca, la larva del nematode continua el seu desenvolupament, forada la paret del tub digestiu i li envaeix els tubs de Malphigi; quan la larva de mosca es converteix en nimfa, la larva del primer estadi passa una muda que la transforma en larva del segon estadi, la qual creix i provoca la formació de tumors, anomenats tilàcies, als tubs de Malphigi de la mosca. Quan la tilàcia s’obre allibera una larva del segon estadi a la cavitat general de la nimfa, i cap al vuitè dia muda i dona lloc a una larva del tercer estadi. En la mosca adulta, aquesta larva es manté dins la cavitat abdominal, i va avançant pel tòrax fins al cap, on es fica dins el llavi (cap al 13è dia). Quan la mosca es detura a la secreció labial d’un cavall, la larva surt del seu aparell bucal i cau a la boca del cavall, d’on posteriorment marxa cap a l’estómac, s’enfonsa a la submucosa i passa dues mudes abans de convertir-se en adult.
Pel que fa als telàzids, el gènere Oxyspirura conserva el cicle heteroxè dels espirúrides més simples i utilitza una panerola (blàtid) com a hoste intermediari. El gènere Thelazia, que comprèn més de trenta espècies paràsites del canal lacrimal de mamífers o de la membrana nictitant dels ocells, en canvi, té una biologia més complexa. L’hoste intermediari és un múscid, que és atret per les secrecions oculars. Les femelles vivípares de Thelazia alliberen unes larves a les secrecions lacrimals i són absorbides per l’insecte; un cop dins, passen de l’intestí a la cavitat abdominal i més tard als fol·licles ovàrics, on creixen i passen dues mudes. El tercer estadi larval abandona l’ovari de l’insecte i migra cap a les peces bucals per a ésser transmès directament a l’hoste definitiu. Les larves del tercer estadi surten activament de la trompa de la mosca quan aquesta es para sobre les parpelles de l’hoste.
Filàries vivípares
Les filàries són nematodes de cos filiforme molt llarg, que viuen en els vasos limfàtics. Les larves del primer estadi, en la majoria dels casos, són alliberades en la sang en forma de microfilàries.
Els dracunculoïdeus (espirúrides) s’han adaptat a la vida en els teixits de l’hoste definitiu. Deriven d’espirúrides primitius, del grup dels camal·lànids, paràsits intestinals de vertebrats aquàtics, el cicle dels quals inclou sempre la intervenció d’un copèpode (Cyclops). El gènere Dracunculus reuneix espècies paràsites de vertebrats (rèptils i marsupials) i inclou també l’anomenada filària de Medina (D. medinensis), paràsit de l’home i d’altres mamífers. El cicle d’aquesta filària manté un cert caràcter primitiu ja que s’inicia encara en el medi extern i la larva del tercer estadi penetra en l’hoste definitiu per via bucal. És, per tant, un cicle semblant al de les filàries ovípares, la biologia de les quals és del tipus dels espirúrides. Quan la femella és madura, es desplaça fins a situar-se sota la pell de l’hoste, generalment en els membres inferiors, i hi forma una petita úlcera cutània. En el moment que l’hoste introdueix aquesta úlcera en l’aigua, la femella allibera, a través de l’úlcera, uns quants milers de larves, que passen a fer vida lliure, nedadora, i no continuen creixent fins que no són ingerides per determinades espècies de copèpodes. En aquest cas, un cop dins el crustaci, passen dues mudes i donen lloc a larves del tercer estadi, que es converteixen en larves infestants entre quatre i vuit dies més tard. L’hoste definitiu s’infesta bevent aigua amb crustacis parasitats. El desenvolupament de la filària dins l’hoste definitiu és molt llarg. Els adults joves s’instal·len en el mesenteri, on s’estan fins al moment de la còpula. Després, el mascle, d’uns 2 cm de llargada, mor, i la femella, de vegades de més d’1 m, arriba fins a la pell al cap d’un any.
Les filàries humanes Onthocerca volvulus, Loa loa, Mansonella streptocerca, M. perstans i Wuchereria bancrofti, pertanyen totes a la família dels oncocèrids. Les larves són les anomenades microfilàries (larves del primer estadi), desproveïdes d’espines, que circulen per la sang i que només poden ésser transmeses per artròpodes hematòfags. Exceptuant alguns detalls, l’evolució de les larves en l’insecte és sempre molt semblant, i es pot resumir de la manera següent. L’insecte, un mosquit o un tàban, pica un home infestat per microfilàries i se les empassa; aquestes travessen ràpidament la paret de l’estómac i arriben al teixit adipós, on penetren als músculs toràcics i s’hi transformen en larves del segon estadi («larves salsitxa») i, més tard, en larves del tercer estadi infestants, llargues i primes. Les larves del tercer estadi migren fins a arribar al llavi inferior de l’insecte, que forma una beina al voltant de les peces bucals. Quan l’insecte pica un altre home, les larves penetren per la ferida en la sang, i al cap d’alguns mesos atenyen l’estat adult, i s’instal·len en un lloc diferent i característic de cada espècie. Analitzant aquests cicles, hom pot deduir els factors que fan que millorin les probabilitats de vida del paràsit en els cicles de les filàries : en primer lloc, l’atracció i la concentració de les microfilàries per la saliva del dípter; en segon lloc la cronobiologia de les microfilàries, que fa que migrin cap als vasos perifèrics durant les hores en què l’insecte s’alimenta.
Els cicles vitals en els adenoforis
L’evolució dels adenoforis difereix de la dels secernentis per dos elements: el primer estadi larval pot arribar a ésser infestant en l’hoste definitiu, de manera que aquí els cicles primitius són molt més simples, ja que les quatre mudes es fan en el mateix hoste; en segon lloc, els adenoforis mostren una tendència a abandonar el tracte digestiu per a colonitzar altres vísceres de l’hoste, i això dona lloc a cicles particulars, ja que els ous no poden ésser eliminats al medi extern sinó que la transmissió a un nou hoste es fa per canibalisme.
Entre els cicles monoxens més senzills, tenim el cas de Capillaria obsignata, que viu en estat adult en la mucosa intestinal d’ocells. Els ous s’eliminen amb la matèria fecal i passen la segmentació a l’exterior. El primer estadi infectant s’assoleix en sis dies. Quan l’ou és ingerit per un ocell, el primer estadi larval, proveït d’un arpó cefàlic, surt de l’ou i cau a l’intestí prim, on passa quatre mudes i arriba a l’estat adult. Aquest mateix cicle el comparteix Trichuris muris, que viu en el cec de rosegadors, i T. trichiura, en el de l’home.

El dimorfisme sexual és generalment molt accentuat entre els nematodes zooparàsits. En el cas de Trichosomoides crassicaudata (a dalt), paràsit de la rata comuna (Rattus norvegicus), es dona el cas, peculiar, que el mascle viu dins l’úter de la femella, com es veu en aquesta fotografia. A baix veiem una femella gràvida de Syphacia stroma, un oxiuroïdeu paràsit intestinal del ratolí de bosc (Apodemus sylvaticus), el mascle del qual és molt més petit.
Santiago Mas-Coma.
Entre els cicles monoxens complexos, amb organotropisme, hi ha el cas de Trichuris vulpis, en el qual la larva del primer estadi penetra a la mucosa intestinal i no retorna al lumen intestinal fins al cap de deu dies. En Capillaria aerophila, la larva, en lloc de retornar a l’intestí, migra per via sanguínia fins a un òrgan profund, en aquest cas els pulmons de la guineu, a partir dels quals són eliminades al medi extern. En Trichosomoides crassicauda la migració es fa fins a la bufeta de la rata. En Capillaria hepatica el cicle és més complex, ja que el paràsit migra fins al fetge; la mort de l’hoste és indispensable perquè s’alliberin els ous, encara no segmentats quan es troben en el fetge; la infestació d’un altre ratolí no pot obtenir-se si no és amb ous que han passat un temps al medi extern, després que el cadàver s’hagi destruït. En Muspicea borreli el cicle s’assembla al de la triquina; els embrions lliures dins el teixit del ratolí no creixen en el medi extern, i la infestació té lloc quan el ratolí llepa les larves emeses a la superfície de la pell o quan ingereix carn parasitada.
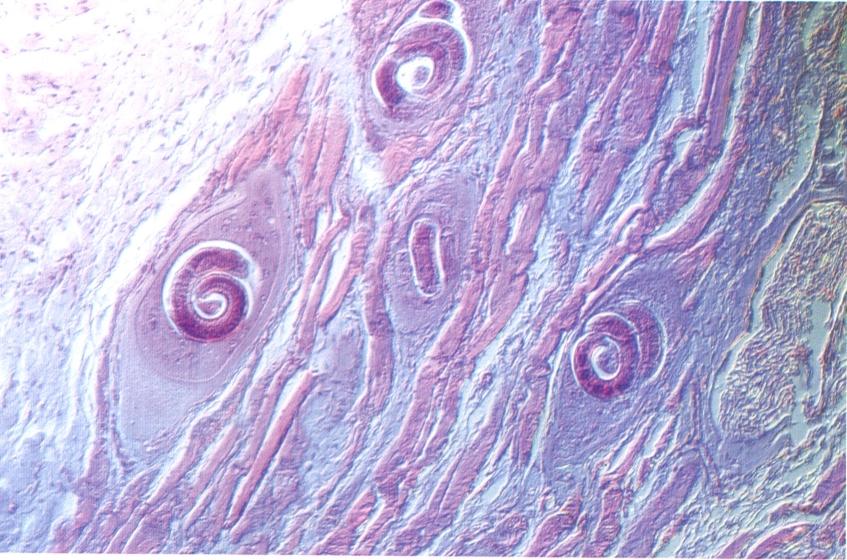
Teixit muscular de porc afectat de triquinosi. S’hi aprecien les larves encapsulades de Trichinella spiralis, el nematode causant de la malaltia.
Jacana.
El cicle de la triquina Trichinella spiralis és marcat per un desenvolupament molt ràpid de les larves dins l’intestí, les quals en tres dies hi atenyen l’estat adult; llavors, les femelles (vivípares) s’enfonsen en la mucosa i comencen a emetre larves, que passen a la circulació general i van a parar als músculs estriats, on s’encapsulen. La conservació de l’espècie ha estat possible gràcies al fenomen del canibalisme (cicle autoheteroxè).
L’heteroxènia s’estableix d’una manera progressiva en determinades espècies de Capillaria. En C. contorta la transmissió es pot fer bé directament pels ous o bé per mitjà d’un oligoquet. En C. plica, paràsit del tracte urinari d’ànecs, els ous es desclouen dins oligoquets; les larves penetren la paret de l’intestí i van a fixar-se en el teixit conjuntiu de la cavitat general del cos, on es fan infestants. Les larves de segon estadi es desenvolupen dins la paret de l’intestí prim de l’hoste definitiu, al cap d’uns vuit o deu dies. Les larves del tercer estadi es troben dins la bufeta als 30 dies aproximadament, on es transformen en quart estadi i posteriorment en adults. En el cas de Cystoopsis acipenseris, els adults viuen en còpula dins els quists subcutanis d’esturions joves; l’evaquació dels ous es fa a l’aigua, com en la filària de Medina, per ruptura del cos de la femella, i els ous són ingerits per un artròpode aquàtic (larva de simúlid o d’amfípode), dins el qual eclosionen; les larves es desenvolupen dins la cavitat general, seguint un cicle comparable al dels espirúrides. En Dioctophyme renale, paràsit renal del gos, el primer hoste intermediari és un oligoquet i el segon, un peix (però aquest no és indispensable); l’hoste definitiu (un gos o un altre mamífer) s’infesta ingerint el peix. La larva migra fins al ronyó, on arriba a atènyer unes dimensions considerables.